![New -Halo–lactones and -Hydroxy–lactones with Strong ...€¦ · Table1. Only two of these compounds have been described thus far in the literature [26]. Figure 1. A four-step](https://static.fdocument.org/doc/165x107/60bc7b780cebbb784b0fd7cc/new-haloalactones-and-hydroxyalactones-with-strong-table1-only-two-of.jpg)
Anticancer Activity of New Synthetic α-Methylene-δ-Lactones on Two Breast Cancer Cell Lines
10
Anticancer Activity of New Synthetic a-Methylene-d-Lactones on Two Breast Cancer Cell Lines Anna Wyre z bska 1 , Katarzyna Gach 1 , Urszula Lewandowska 2 , Karolina Szewczyk 2 , El _ zbieta Hrabec 2 , Jakub Modranka 3 , Rafal Jakubowski 3 , Tomasz Janecki 3 , Jacek Szyma nski 4 and Anna Janecka 1 1 Department of Biomolecular, Chemistry Medical University of Lodz, Lodz, Poland, 2 Department of Enzymology, Medical University of Lodz, Lodz, Poland, 3 Institute of Organic Chemistry, Technical University of Lodz, Lodz, Poland and 4 Central Laboratory, Medical University of Lodz, Lodz, Poland (Received 14 January 2013; Accepted 18 July 2013) Abstract: Natural products are important leads in drug discovery. The search for effective plant-derived anticancer agents or their synthetic analogues has continued to be of interest to biologists and chemists for a long time. In this report, cytotoxicity and anti- cancer activity of new synthetic a-methylene-d-lactones was tested against two breast cancer cell lines, invasive, hormone-inde- pendent MDA-MB-231 and hormone-dependent MCF-7. Cytotoxicity was examined using MTT assay. The ability to induce apoptosis and changes in mitochondrial membrane potential was studied by flow cytometry. The expression levels of pro- and anti-apoptotic genes were determined by quantitative real-time PCR. Cancer cell migration and invasion were assessed by wound healing and Matrigel assays. Additionally, secretion of proteins associated with invasiveness, metalloproteinase-9 (MMP-9) and urokinase plasminogen activator (uPA) was investigated using commercial ELISA kits and MMP-9 activity by gelatin zymogra- phy. A natural sesquiterpene lactone, parthenolide, was used as a positive control. Screening results showed all four analogues to be highly cytotoxic. The most potent compound of the series, 1-isopropyl-2-methylene-1,2-dihydrobenzochromen-3-one, desig- nated DL-3, which reduced the number of viable MDA-MB-231 and MCF-7 cells with the IC 50 values of 5.3 lM and 3.54 lM, respectively, was selected for further research. DL-3 activated the intrinsic pathway of apoptosis, associated with the loss of mitochondrial membrane potential and changes in Bax/Bcl-2 ratio. DL-3 also inhibited the movement of both types of breast cancer cells. Suppression of cell migration and invasion was the result of the decreased secretion of enzymes responsible for the degradation of the extracellular matrix, MMP-9 and uPA. These findings show that the synthetic a-methylene-d-lactone, DL-3, displays potential to be further explored in the development of new anticancer agents. Throughout history, plants have always been an excellent source of pharmaceutical agents used in traditional medicine. More recently, a big number of important drugs have been obtained from plants, either directly, by extracting an active component or, more often, by structural modifications of natu- ral compounds or by the synthesis of their analogues with improved pharmacological properties. Many of the compounds used for cancer chemotherapy, such as the Vinca alkaloids, paclitaxel, camtothecin and etoposide, were originally derived from plants [1], and plants continue to be viewed as major sources for the development of new anticancer drugs. A vast number of biologically significant natural products are characterized by the a-methylene-c-lactone structural motif. Such products are abundant in plants of the Compositae family and possess a broad spectrum of biological activities, ranging from anti-inflammatory, phytotoxic, antibacterial and antifungal to cytotoxic/anticancer [2]. First a-methylene-c-lac- tones 1 (fig. 1) were isolated from plants of the Compositae family over 100 years ago, but real interest in their synthesis and biological activities came in the last decade. The unsatu- rated lactone functionality can react as the Michael-type acceptor with mercapto groups in cysteine residues of enzymes, other proteins and free intracellular glutathione, which leads to the formation of covalent adducts. Such alkyl- ation of cellular thiols is an important reaction resulting in the disruption of some major processes in the cells. Various groups of cytotoxic agents that can alkylate cellular thiols are used or tested as anticancer agents [3]. In contrast to a-methylene-c-lactones, a-methylene-d- lactones 2 (fig. 1) are much less abundant in nature. First a-methylene-d-lactones, vernolepin and vernomenin isolated from Vernonia hymenolepis [4] and crassin from Pseudoplexa- ura porosa [5] showed some cytotoxicity against human pha- ryngeal carcinoma (KB) cells. However, the data on the cytotoxic activity of natural a-methylene-d-lactones are scarce. From some time now, we and others have been developing general methodologies for the introduction of a methylene group onto the c- or d-lactone ring, using the Horner–Wads- worth–Emmons olefination reaction. Recently, we have Author for correspondence: Anna Janecka, Laboratory of Biomolecular Chemistry, Medical University of Lodz, Mazowiecka 6/8, 92-215 Lodz, Poland (fax: +48 42 42 272 56 94, e-mail anna.janecka@umed. lodz.pl). O O O O R R 1 2 Fig. 1. General structure of a-methylene-c-lactones 1 and a-methy- lene-d-lactones 2. © 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd Basic & Clinical Pharmacology & Toxicology Doi: 10.1111/bcpt.12120
Transcript of Anticancer Activity of New Synthetic α-Methylene-δ-Lactones on Two Breast Cancer Cell Lines

Anticancer Activity of New Synthetic a-Methylene-d-Lactones onTwo Breast Cancer Cell Lines
Anna Wyrezbska1, Katarzyna Gach1, Urszula Lewandowska2, Karolina Szewczyk2, El _zbieta Hrabec2, Jakub Modranka3, RafałJakubowski3, Tomasz Janecki3, Jacek Szyma�nski4 and Anna Janecka1
1Department of Biomolecular, Chemistry Medical University of Lodz, Lodz, Poland, 2Department of Enzymology, Medical University of Lodz,Lodz, Poland, 3Institute of Organic Chemistry, Technical University of Lodz, Lodz, Poland and 4Central Laboratory, Medical University of Lodz,
Lodz, Poland
(Received 14 January 2013; Accepted 18 July 2013)
Abstract: Natural products are important leads in drug discovery. The search for effective plant-derived anticancer agents or theirsynthetic analogues has continued to be of interest to biologists and chemists for a long time. In this report, cytotoxicity and anti-cancer activity of new synthetic a-methylene-d-lactones was tested against two breast cancer cell lines, invasive, hormone-inde-pendent MDA-MB-231 and hormone-dependent MCF-7. Cytotoxicity was examined using MTT assay. The ability to induceapoptosis and changes in mitochondrial membrane potential was studied by flow cytometry. The expression levels of pro- andanti-apoptotic genes were determined by quantitative real-time PCR. Cancer cell migration and invasion were assessed by woundhealing and Matrigel assays. Additionally, secretion of proteins associated with invasiveness, metalloproteinase-9 (MMP-9) andurokinase plasminogen activator (uPA) was investigated using commercial ELISA kits and MMP-9 activity by gelatin zymogra-phy. A natural sesquiterpene lactone, parthenolide, was used as a positive control. Screening results showed all four analogues tobe highly cytotoxic. The most potent compound of the series, 1-isopropyl-2-methylene-1,2-dihydrobenzochromen-3-one, desig-nated DL-3, which reduced the number of viable MDA-MB-231 and MCF-7 cells with the IC50 values of 5.3 lM and 3.54 lM,respectively, was selected for further research. DL-3 activated the intrinsic pathway of apoptosis, associated with the loss ofmitochondrial membrane potential and changes in Bax/Bcl-2 ratio. DL-3 also inhibited the movement of both types of breastcancer cells. Suppression of cell migration and invasion was the result of the decreased secretion of enzymes responsible for thedegradation of the extracellular matrix, MMP-9 and uPA. These findings show that the synthetic a-methylene-d-lactone, DL-3,displays potential to be further explored in the development of new anticancer agents.
Throughout history, plants have always been an excellentsource of pharmaceutical agents used in traditional medicine.More recently, a big number of important drugs have beenobtained from plants, either directly, by extracting an activecomponent or, more often, by structural modifications of natu-ral compounds or by the synthesis of their analogues withimproved pharmacological properties. Many of the compoundsused for cancer chemotherapy, such as the Vinca alkaloids,paclitaxel, camtothecin and etoposide, were originally derivedfrom plants [1], and plants continue to be viewed as majorsources for the development of new anticancer drugs.A vast number of biologically significant natural products
are characterized by the a-methylene-c-lactone structuralmotif. Such products are abundant in plants of the Compositaefamily and possess a broad spectrum of biological activities,ranging from anti-inflammatory, phytotoxic, antibacterial andantifungal to cytotoxic/anticancer [2]. First a-methylene-c-lac-tones 1 (fig. 1) were isolated from plants of the Compositaefamily over 100 years ago, but real interest in their synthesisand biological activities came in the last decade. The unsatu-rated lactone functionality can react as the Michael-type
acceptor with mercapto groups in cysteine residues ofenzymes, other proteins and free intracellular glutathione,which leads to the formation of covalent adducts. Such alkyl-ation of cellular thiols is an important reaction resulting in thedisruption of some major processes in the cells. Variousgroups of cytotoxic agents that can alkylate cellular thiols areused or tested as anticancer agents [3].In contrast to a-methylene-c-lactones, a-methylene-d-
lactones 2 (fig. 1) are much less abundant in nature. Firsta-methylene-d-lactones, vernolepin and vernomenin isolatedfrom Vernonia hymenolepis [4] and crassin from Pseudoplexa-ura porosa [5] showed some cytotoxicity against human pha-ryngeal carcinoma (KB) cells. However, the data on thecytotoxic activity of natural a-methylene-d-lactones are scarce.From some time now, we and others have been developing
general methodologies for the introduction of a methylenegroup onto the c- or d-lactone ring, using the Horner–Wads-worth–Emmons olefination reaction. Recently, we have
Author for correspondence: Anna Janecka, Laboratory of BiomolecularChemistry, Medical University of Lodz, Mazowiecka 6/8, 92-215Lodz, Poland (fax: +48 42 42 272 56 94, e-mail [email protected]).
O O O ORR
1 2
Fig. 1. General structure of a-methylene-c-lactones 1 and a-methy-lene-d-lactones 2.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
Basic & Clinical Pharmacology & Toxicology Doi: 10.1111/bcpt.12120

prepared a series of twenty a-methylene-d-lactones all ofwhich, when evaluated by the MTT test, showed high-to-mod-erate cytotoxicity against several cancer cell lines (HL-60 andNALM-6 leukaemias and MCF-7 and HT-29 adenocarcino-mas) [6]. Here, we have chosen four of these d-lactones thatcould be classified as 2-methylene-1,2-dihydrobenzochromen-3-ones 3 (designated DL-1 to DL-4) (fig. 2), for further evalu-ation of their anticancer potential in two breast cancer celllines, the oestrogen receptor (ER)-negative MDA-MB-231 andhormone-independent MCF-7. Natural sesquiterpene lactone,parthenolide (PTL), was used as a positive control.
Materials and Methods
Materials and general procedures. 2-Methylene-1,2-dihydroben-zochromen-3-ones (DL-1 to DL-4) were synthesized as describedearlier [6]. Parthenolide (PTL) used as a positive control incytotoxicity assays was purchased from Tocris Bioscience (Bristol,UK). In all experiments, parthenolide and the tested compounds weredissolved in DMSO and further diluted in the culture medium toobtain 0.1% DMSO concentration. In each experiment, controlswithout and with 0.1% DMSO were performed. 0.1% DMSO had noeffect on the observed parameters.
Cell culture. The MCF-7 and MDA-MB-231 cell lines werepurchased from the European Collection of Cell Cultures (ECACC).The cells were cultured in high-glucose Dulbecco’s modified Eagle’smedium (DMEM, BioWhittaker, Lonza, Basel, Switzerland)supplemented with glutamine (2 mM) (Sigma-Aldrich, St. Louis, MO,USA) gentamicin (5 lg/mL) and 10% heat-inactivated foetal bovineserum (FBS) (both from Biological Industries, Beit-Haemek, Israel).Cells were maintained at 37°C in a 5% CO2 atmosphere and weregrown until 80% confluent.Human aortic smooth muscle cells (HASMC) (a gift from prof.
Janusz Szemraj, Medical University of Lodz) were cultured in thesame medium as above.
MTT assay. The MTT assay was performed according to a knownprocedure [7]. The cells were grown to subconfluent levels in theculture medium and then plated onto 24-well plates (104 cells/well) in
the final volume of 1 ml of the culture medium. After 24 hr, variousconcentrations of the tested lactones were added, and the plates wereincubated for 24 hr. Then, the cells were incubated for 2 hr at 37°Cwith 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide[MTT, 5 mg/ml in phosphate-buffered saline (PBS, GIBCO,Invitrogen, Carlsbad, CA, USA)]. The absorbance of the blueformazan product was measured at 540 nm using an automated platereader (iMark Bio-Rad, Hercules, CA, USA) and compared withcontrol (untreated cells). All experiments were performed in triplicate.
Quantitative real-time PCR. The cells were seeded in 25-ml cellculture flasks in 10 ml of standard growth medium. After 24 hr, thegrowth medium was replaced by a fresh growth mediumsupplemented with DL-3 in the desired concentration and incubatedfor 24 hr (apoptosis-related genes) and 48 hr (invasiveness-relatedgenes). Then, the cells were washed twice with PBS to remove theadded compound and harvested by trypsinization. The cells werefrozen and kept at �80°C, awaiting further experiments.Total RNA was extracted using Total RNA Mini Kit (A&A Bio-
technology, Poland) [8]. cDNA was synthesized using EnhancedAvian HS RT-PCR Kit and oligo(dT)12-18 primers (Sigma-Aldrich, St.Louis, MO, USA). Quantitative real-time PCR was performed with theMx3005P QPCR Systems (Agilent Technologies, Inc. Santa Clara,CA, USA) according to the manufacturer’s instructions for the Bril-liant II SYBR Green QPCR Master Mix (Agilent Technologies, Inc.Santa Clara, CA, USA). Primers used are listed in table 1. Real-timePCRs were run in triplicate using the following thermal cycling pro-file: 95°C for 10 min., followed by 40 steps of 95°C for 30 sec. and58°C for 1 min. and 72°C for 1 min. After 40 cycles, samples wererun for the dissociation protocol (i.e. melting curve analysis). Theexpression levels of the tested genes were determined by the 2�ΔΔCT
method [9]. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)was used as a reference gene. All results were presented asmean � S.D.
Analysis of apoptosis by flow cytometry. Apoptotic cell death wasdetermined using the FITC Annexin V Apoptosis Detection Kit (BDBioscience), according to the manufacturer guidelines. Briefly, cellswere treated for 24 hr with the indicated concentrations of DL-3.Then, cells were harvested, stained with annexin V and propidiumiodide (PI) and kept for 15 min. at room temperature in the dark.Apoptosis of cells was analysed by the flow cytometry (BectonDickinson Canto II). Early and late apoptosis was visualized andquantified by constructing a dot plot using the BD FACSDivasoftware.
Assessment of mitochondrial membrane potential. The integrity ofmitochondrial membrane was assessed by JC-10 staining and flowcytometry. This dye can selectively enter into mitochondrial matrix,switching colour from red to green when DΨm decreases. The healthynon-apoptotic cells maintain a high DΨm, while unhealthy apoptoticcells have a low DΨm, which is the result of the formation of pores inthe mitochondria by dimerized Bax or activated Bid, Bak, or Badproteins.
O O
3
RDL-1, R = MeDL-2, R = BuDL-3, R = i-PrDL-4, R = vinyl
Fig. 2. Structure of new synthetic 2-methylene-1,2-dihydrobenzochro-men-3-ones 3.
Table 1.Primers used for quantitative real-time PCR.
Target gene Forward primers Reverse primers
Bax 5′ ACCCGGTGCCTCAGGATGCGT 3′ 5′ GGCAAAGTAGAAAAGGGCGAC 3′Bcl-2 5′ CATGCTGGGGCCGTACAG 3′ 5′ GAACCGGCACCTGCACAC 3′MMP-9 5′ GACCAATCTCACCGACAGG 3′ 5′ GCCACCCGAGTGTAACCATA 3′uPA 5′ GACCCCCTCGTCTGTTCCCTCCAAG 3′ 5′ CTCTTCCTTGGTGTGACTGCGG 3′GAPDH 5′ GTCGCTGTTGAAGTCAGAGGAG 3′ 5′ CGTGTCAGT GGTGGACCTGAC 3′
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
2 ANNA WYREzBSKA ET AL.

To monitor changes in mitochondrial membrane potential (DΨm),cells treated with a tested compound for 24 hr were loaded with JC-10 (Cell MeterTM JC-10 Mitochondrial Membrane Potential Assay Kit,AAT, Bioquest Inc., Sunnyvale, CA, USA) and analysed by flowcytometry (Becton Dickinson Canto II). Carbonyl cyanide-4-(trifluoro-methoxy)phenylhydrazone (FCCP), provided by the producer, wasused as a positive control. Changes in DΨm were visualized and quan-tified by constructing a dot plot using the BD FACSDiva software.
Wound-healing assay. The assay was performed according to aknown procedure [10]. The cells were seeded in 6-well plates. Whenthey achieved 90% confluence, a wound was made in the centre ofthe cell monolayer with a sterile 200 lm disposable tip. The cellswere washed with PBS, and the fresh culture medium with DL-3 wasadded. The scratched area was photographed at the reference pointswith a phase-contrast microscope (OLYMPUS CKX41, Olympus,London, UK) for the first image, and the following images (after 0, 6,12, and 24 hr). Assays were performed in triplicate and repeated threetimes. The distances between two edges of the scratch were measuredat the reference points and analysed statistically.
Invasion assay. The invasion assay was performed as previouslydescribed [11]. Briefly, the cells were plated on the upper
compartment of a Transwell chamber and invaded through a Matrigel-coated membrane. After 48 hr, the cells at the upper side of themembrane were removed and the invaded cells were fixed, stainedand counted.
Metalloproteinase-9 (MMP-9) and urokinase plasminogen activator
(uPA) secretion. The supernatant collected after treatment of the cellswith DL-3 was subsequently analysed for MMP-9 and uPA proteinlevels using MMP-9 ELISA Kit (RayBiotech, Norcross, GA, USA)and AssayMax Human Urokinase (uPA) ELISA Kit (AssayPro, St.Charles, MO, USA), respectively, according to the manufacturer’sinstructions.
Zymography. The cells were seeded into 96-well plates at a density104 cells/well in the final volume of 200 ll of the culture medium.After 24 hr, the growth medium was replaced by a fresh serum-freemedium supplemented with DL-3 in the desired concentration. Thecells were additionally incubated with tumour necrosis factor-alpha(TNF-a, 25 ng/ml). After 48-hr incubation, the culture supernatantwas collected.Gelatin zymography was performed as described previously [12].
Briefly, the same volume of conditioned media (20 ll) was dissolvedin electrophoresis sample buffer containing sodium dodecyl sulphate
Table 2.Growth inhibitory effects of a-methylene-d-lactones in human breast cancer cell lines MDA-MB-231 and MCF-7 and in human aortic smooth mus-cle cells (HASMC).
Compound Structure
IC50 (lM)
MDA-MB-231 MCF-7 HASMC
DL-1
O O
11.4 � 2.10 8.17 � 0.58 –
DL-2
O O
15.1 � 1.82 12.67 � 0.29 –
DL-3
O O
5.3 � 0.69 3.54 � 0.76 6.5 � 0.85
DL-4
O O
n-Bu 7.9 � 0.99 4.75 � 1.09 –
PTL
OO O
14.5 � 2.10 9.5 � 0.70 17.5 � 1.5
All values are expressed as mean � S.E.M. of three independent experiments performed in triplicate.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
ANTICANCER ACTIVITY OF NEW SYNTHETIC a-METHYLENE-d-LACTONES 3
(SDS) (Applichem Darmstadt, Germany) and subjected to electropho-resis in 10% polyacrylamide gel embedded with 1.5 mg/ml gelatin inthe absence of b-mercaptoethanol. After electrophoresis, the enzymeswere renatured by incubation with 2.5% Triton X-100 and the enzy-matic reaction was allowed to proceed at 37°C for 21 hr. Thereafter,the gels were stained for 1.5 hr with 0.0125% Amido Black (POCH,Gliwice, Poland) in 7% acetic acid and 20% ethanol, and photo-graphed using Olympus camera. The densitometry of the gels was car-ried out using the Quantity One 4.4 software (Bio-Rad Laboratories,Hercules, CA, USA).
Statistical analysis. Statistical analyses were performed using Prism4.0 (GraphPad Software Inc., San Diego, CA, USA). The data wereexpressed as means � S.D. Statistical comparisons were assessed by aone-way ANOVA followed by a post hoc multiple comparison Student–Newman–Keuls test. A probability level of 0.05 or lower wasconsidered statistically significant.
Results
Cytotoxicity assay. Cytotoxicity of the synthetic a-methylene-d-lactones was examined using the MTT assay. The survivalof MDA-MB-231 or MCF-7 cells, after incubation for 24 hr
with the tested compounds in increasing concentrations, wasassessed (table 2). All synthetic lactones showed highcytotoxic activity, with analogue DL-3 being the most potent(about 2 times lower IC50 value than for PTL in both celllines). DL-3, as the most cytotoxic analogue, was selected forfurther evaluation as a potential anticancer agent.Cytotoxicity of DL-3 and PTL was also tested against
HASMC cells, to evaluate the influence of these compoundson normal cells. No significant differences in IC50 valuesbetween DL-3- or PTL-treated cancer and HASMC cells wereobserved.
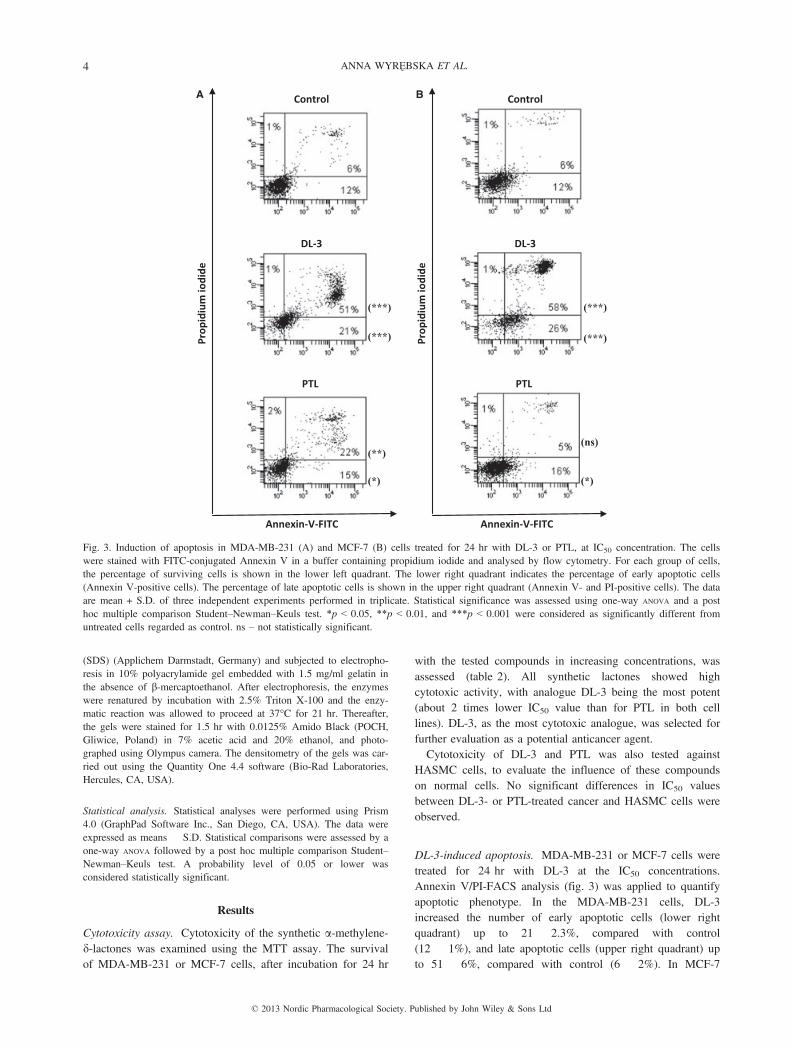
DL-3-induced apoptosis. MDA-MB-231 or MCF-7 cells weretreated for 24 hr with DL-3 at the IC50 concentrations.Annexin V/PI-FACS analysis (fig. 3) was applied to quantifyapoptotic phenotype. In the MDA-MB-231 cells, DL-3increased the number of early apoptotic cells (lower rightquadrant) up to 21 � 2.3%, compared with control(12 � 1%), and late apoptotic cells (upper right quadrant) upto 51 � 6%, compared with control (6 � 2%). In MCF-7
PTL PTL
Prop
idiu
m io
dide
Prop
idiu
m io
dide
Control Control
DL-3 DL-3
Annexin-V-FITC Annexin-V-FITC
(***)(***)
(***)
(**)
(*)
(***)
(ns)
(*)
A B
Fig. 3. Induction of apoptosis in MDA-MB-231 (A) and MCF-7 (B) cells treated for 24 hr with DL-3 or PTL, at IC50 concentration. The cellswere stained with FITC-conjugated Annexin V in a buffer containing propidium iodide and analysed by flow cytometry. For each group of cells,the percentage of surviving cells is shown in the lower left quadrant. The lower right quadrant indicates the percentage of early apoptotic cells(Annexin V-positive cells). The percentage of late apoptotic cells is shown in the upper right quadrant (Annexin V- and PI-positive cells). The dataare mean + S.D. of three independent experiments performed in triplicate. Statistical significance was assessed using one-way ANOVA and a posthoc multiple comparison Student–Newman–Keuls test. *p < 0.05, **p < 0.01, and ***p < 0.001 were considered as significantly different fromuntreated cells regarded as control. ns – not statistically significant.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
4 ANNA WYREzBSKA ET AL.
cells, the number of early and late apoptotic cells was26 � 2% and 58 � 6%, respectively, compared with control(12 � 2% and 6 � 2%). For PTL used as a positive control,the number of early and late apoptotic cells was 15 � 0.5%and 22 � 1.5%, respectively, for MDA-MB-231 cells, whilethe same values for MCF-7 cells were 16 � 1.8% and5 � 1%, respectively.It can be concluded that DL-3 was a much stronger apopto-
tic inducer than PTL, causing about 2 and 10 times increasein the late apoptotic cell population in MDA-MB-231 andMCF-7 cells, respectively.
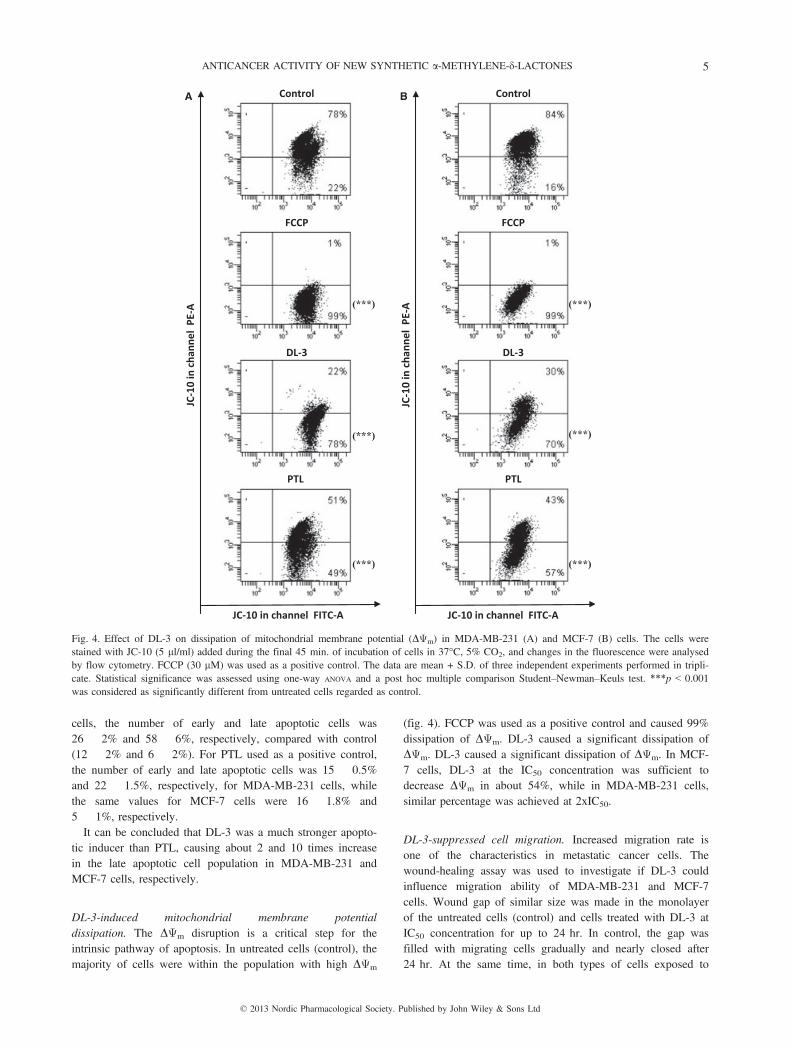
DL-3-induced mitochondrial membrane potentialdissipation. The DΨm disruption is a critical step for theintrinsic pathway of apoptosis. In untreated cells (control), themajority of cells were within the population with high DΨm
(fig. 4). FCCP was used as a positive control and caused 99%dissipation of DΨm. DL-3 caused a significant dissipation ofDΨm. DL-3 caused a significant dissipation of DΨm. In MCF-7 cells, DL-3 at the IC50 concentration was sufficient todecrease DΨm in about 54%, while in MDA-MB-231 cells,similar percentage was achieved at 2xIC50.
DL-3-suppressed cell migration. Increased migration rate isone of the characteristics in metastatic cancer cells. Thewound-healing assay was used to investigate if DL-3 couldinfluence migration ability of MDA-MB-231 and MCF-7cells. Wound gap of similar size was made in the monolayerof the untreated cells (control) and cells treated with DL-3 atIC50 concentration for up to 24 hr. In control, the gap wasfilled with migrating cells gradually and nearly closed after24 hr. At the same time, in both types of cells exposed to
DL-3
Control
PTL
FCCP
JC-1
0 in
cha
nnel
PE-
A
JC-10 in channel FITC-A
DL-3
Control
PTL
FCCP
JC-1
0 in
cha
nnel
PE-
A
JC-10 in channel FITC-A
(***)
(***)
(***)
(***)
(***)
(***)
A B
Fig. 4. Effect of DL-3 on dissipation of mitochondrial membrane potential (DΨm) in MDA-MB-231 (A) and MCF-7 (B) cells. The cells werestained with JC-10 (5 ll/ml) added during the final 45 min. of incubation of cells in 37°C, 5% CO2, and changes in the fluorescence were analysedby flow cytometry. FCCP (30 lM) was used as a positive control. The data are mean + S.D. of three independent experiments performed in tripli-cate. Statistical significance was assessed using one-way ANOVA and a post hoc multiple comparison Student–Newman–Keuls test. ***p < 0.001was considered as significantly different from untreated cells regarded as control.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
ANTICANCER ACTIVITY OF NEW SYNTHETIC a-METHYLENE-d-LACTONES 5
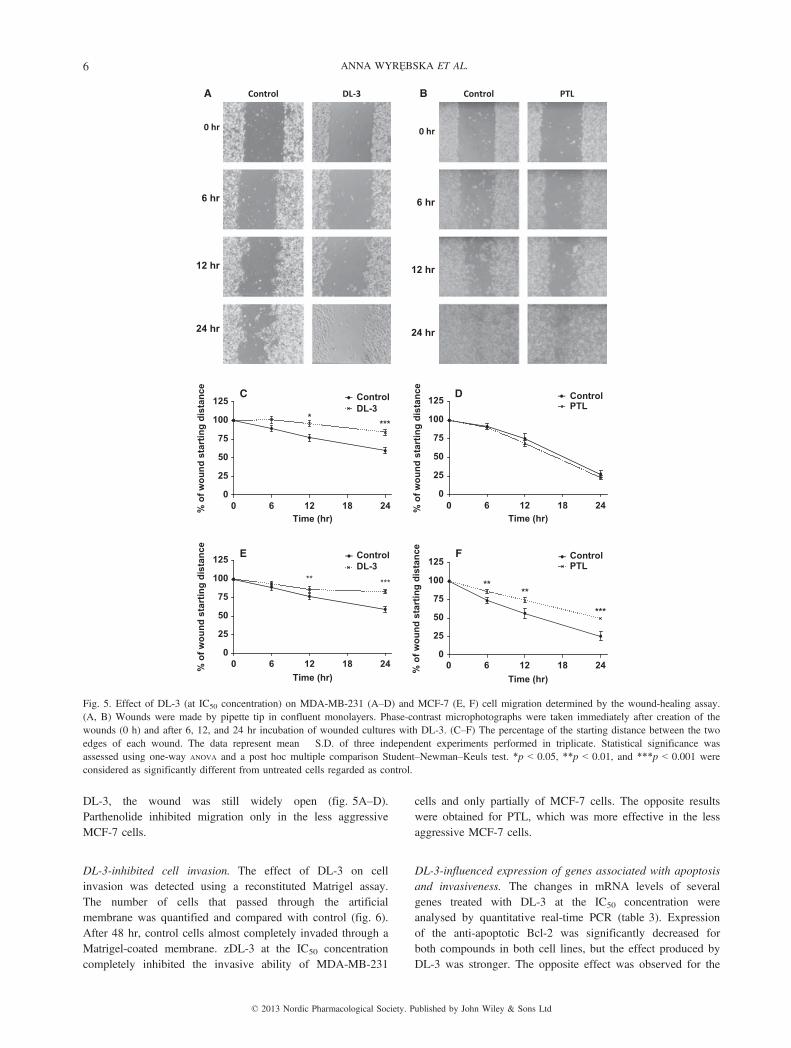
DL-3, the wound was still widely open (fig. 5A–D).Parthenolide inhibited migration only in the less aggressiveMCF-7 cells.
DL-3-inhibited cell invasion. The effect of DL-3 on cellinvasion was detected using a reconstituted Matrigel assay.The number of cells that passed through the artificialmembrane was quantified and compared with control (fig. 6).After 48 hr, control cells almost completely invaded through aMatrigel-coated membrane. zDL-3 at the IC50 concentrationcompletely inhibited the invasive ability of MDA-MB-231
cells and only partially of MCF-7 cells. The opposite resultswere obtained for PTL, which was more effective in the lessaggressive MCF-7 cells.
DL-3-influenced expression of genes associated with apoptosisand invasiveness. The changes in mRNA levels of severalgenes treated with DL-3 at the IC50 concentration wereanalysed by quantitative real-time PCR (table 3). Expressionof the anti-apoptotic Bcl-2 was significantly decreased forboth compounds in both cell lines, but the effect produced byDL-3 was stronger. The opposite effect was observed for the
Control
0 hr
6 hr
12 hr
24 hr
DL-3 Control
0 hr
6 hr
12 hr
24 hr
PTLA B
0 6 12 18 240
25
50
75
100
125DL-3Control
****
Time (hr)
% o
f wou
nd s
tart
ing
dist
ance
0 6 12 18 240
25
50
75
100
125DL-3Control
** ***
Time (hr)
% o
f wou
nd s
tart
ing
dist
ance
C
E
0 6 12 18 240
25
50
75
100
125 PTLControl
Time (hr)
% o
f wou
nd s
tart
ing
dist
ance
0 6 12 18 240
25
50
75
100
125 PTLControl
****
***
Time (hr)
% o
f wou
nd s
tart
ing
dist
ance
D
F
Fig. 5. Effect of DL-3 (at IC50 concentration) on MDA-MB-231 (A–D) and MCF-7 (E, F) cell migration determined by the wound-healing assay.(A, B) Wounds were made by pipette tip in confluent monolayers. Phase-contrast microphotographs were taken immediately after creation of thewounds (0 h) and after 6, 12, and 24 hr incubation of wounded cultures with DL-3. (C–F) The percentage of the starting distance between the twoedges of each wound. The data represent mean � S.D. of three independent experiments performed in triplicate. Statistical significance wasassessed using one-way ANOVA and a post hoc multiple comparison Student–Newman–Keuls test. *p < 0.05, **p < 0.01, and ***p < 0.001 wereconsidered as significantly different from untreated cells regarded as control.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
6 ANNA WYREzBSKA ET AL.
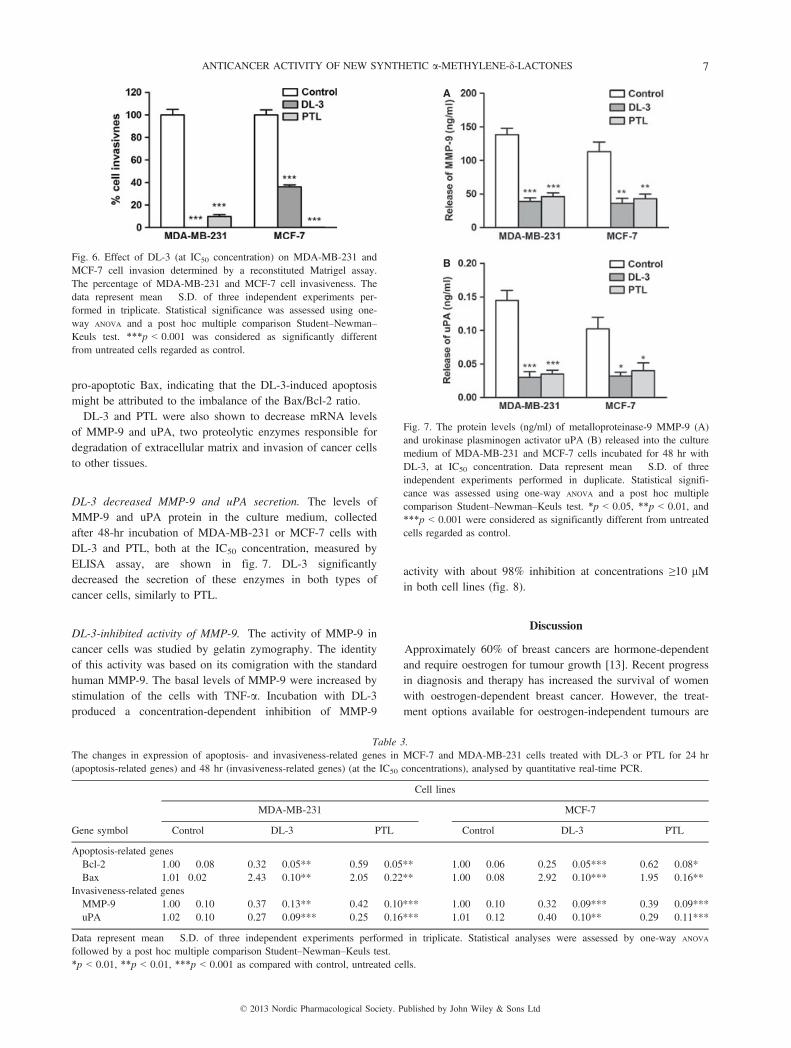
pro-apoptotic Bax, indicating that the DL-3-induced apoptosismight be attributed to the imbalance of the Bax/Bcl-2 ratio.DL-3 and PTL were also shown to decrease mRNA levels
of MMP-9 and uPA, two proteolytic enzymes responsible fordegradation of extracellular matrix and invasion of cancer cellsto other tissues.
DL-3 decreased MMP-9 and uPA secretion. The levels ofMMP-9 and uPA protein in the culture medium, collectedafter 48-hr incubation of MDA-MB-231 or MCF-7 cells withDL-3 and PTL, both at the IC50 concentration, measured byELISA assay, are shown in fig. 7. DL-3 significantlydecreased the secretion of these enzymes in both types ofcancer cells, similarly to PTL.
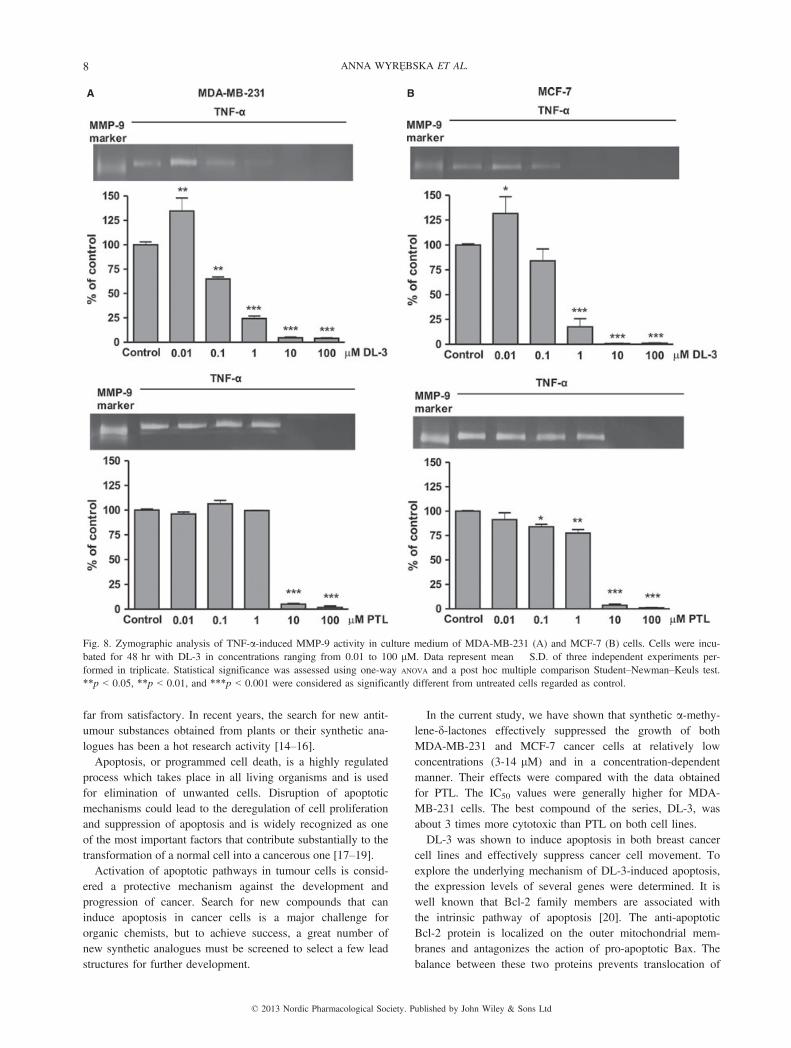
DL-3-inhibited activity of MMP-9. The activity of MMP-9 incancer cells was studied by gelatin zymography. The identityof this activity was based on its comigration with the standardhuman MMP-9. The basal levels of MMP-9 were increased bystimulation of the cells with TNF-a. Incubation with DL-3produced a concentration-dependent inhibition of MMP-9
activity with about 98% inhibition at concentrations ≥10 lMin both cell lines (fig. 8).
Discussion
Approximately 60% of breast cancers are hormone-dependentand require oestrogen for tumour growth [13]. Recent progressin diagnosis and therapy has increased the survival of womenwith oestrogen-dependent breast cancer. However, the treat-ment options available for oestrogen-independent tumours are
Fig. 6. Effect of DL-3 (at IC50 concentration) on MDA-MB-231 andMCF-7 cell invasion determined by a reconstituted Matrigel assay.The percentage of MDA-MB-231 and MCF-7 cell invasiveness. Thedata represent mean � S.D. of three independent experiments per-formed in triplicate. Statistical significance was assessed using one-way ANOVA and a post hoc multiple comparison Student–Newman–Keuls test. ***p < 0.001 was considered as significantly differentfrom untreated cells regarded as control.
A
B
Fig. 7. The protein levels (ng/ml) of metalloproteinase-9 MMP-9 (A)and urokinase plasminogen activator uPA (B) released into the culturemedium of MDA-MB-231 and MCF-7 cells incubated for 48 hr withDL-3, at IC50 concentration. Data represent mean � S.D. of threeindependent experiments performed in duplicate. Statistical signifi-cance was assessed using one-way ANOVA and a post hoc multiplecomparison Student–Newman–Keuls test. *p < 0.05, **p < 0.01, and***p < 0.001 were considered as significantly different from untreatedcells regarded as control.
Table 3.The changes in expression of apoptosis- and invasiveness-related genes in MCF-7 and MDA-MB-231 cells treated with DL-3 or PTL for 24 hr(apoptosis-related genes) and 48 hr (invasiveness-related genes) (at the IC50 concentrations), analysed by quantitative real-time PCR.
Gene symbol
Cell lines
MDA-MB-231 MCF-7
Control DL-3 PTL Control DL-3 PTL
Apoptosis-related genesBcl-2 1.00 � 0.08 0.32 � 0.05** 0.59 � 0.05** 1.00 � 0.06 0.25 � 0.05*** 0.62 � 0.08*Bax 1.01�0.02 2.43 � 0.10** 2.05 � 0.22** 1.00 � 0.08 2.92 � 0.10*** 1.95 � 0.16**
Invasiveness-related genesMMP-9 1.00 � 0.10 0.37 � 0.13** 0.42 � 0.10*** 1.00 � 0.10 0.32 � 0.09*** 0.39 � 0.09***uPA 1.02 � 0.10 0.27 � 0.09*** 0.25 � 0.16*** 1.01 � 0.12 0.40 � 0.10** 0.29 � 0.11***
Data represent mean � S.D. of three independent experiments performed in triplicate. Statistical analyses were assessed by one-way ANOVA
followed by a post hoc multiple comparison Student–Newman–Keuls test.*p < 0.01, **p < 0.01, ***p < 0.001 as compared with control, untreated cells.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
ANTICANCER ACTIVITY OF NEW SYNTHETIC a-METHYLENE-d-LACTONES 7
far from satisfactory. In recent years, the search for new antit-umour substances obtained from plants or their synthetic ana-logues has been a hot research activity [14–16].Apoptosis, or programmed cell death, is a highly regulated
process which takes place in all living organisms and is usedfor elimination of unwanted cells. Disruption of apoptoticmechanisms could lead to the deregulation of cell proliferationand suppression of apoptosis and is widely recognized as oneof the most important factors that contribute substantially to thetransformation of a normal cell into a cancerous one [17–19].Activation of apoptotic pathways in tumour cells is consid-
ered a protective mechanism against the development andprogression of cancer. Search for new compounds that caninduce apoptosis in cancer cells is a major challenge fororganic chemists, but to achieve success, a great number ofnew synthetic analogues must be screened to select a few leadstructures for further development.
In the current study, we have shown that synthetic a-methy-lene-d-lactones effectively suppressed the growth of bothMDA-MB-231 and MCF-7 cancer cells at relatively lowconcentrations (3-14 lM) and in a concentration-dependentmanner. Their effects were compared with the data obtainedfor PTL. The IC50 values were generally higher for MDA-MB-231 cells. The best compound of the series, DL-3, wasabout 3 times more cytotoxic than PTL on both cell lines.DL-3 was shown to induce apoptosis in both breast cancer
cell lines and effectively suppress cancer cell movement. Toexplore the underlying mechanism of DL-3-induced apoptosis,the expression levels of several genes were determined. It iswell known that Bcl-2 family members are associated withthe intrinsic pathway of apoptosis [20]. The anti-apoptoticBcl-2 protein is localized on the outer mitochondrial mem-branes and antagonizes the action of pro-apoptotic Bax. Thebalance between these two proteins prevents translocation of
A B
Fig. 8. Zymographic analysis of TNF-a-induced MMP-9 activity in culture medium of MDA-MB-231 (A) and MCF-7 (B) cells. Cells were incu-bated for 48 hr with DL-3 in concentrations ranging from 0.01 to 100 lM. Data represent mean � S.D. of three independent experiments per-formed in triplicate. Statistical significance was assessed using one-way ANOVA and a post hoc multiple comparison Student–Newman–Keuls test.**p < 0.05, **p < 0.01, and ***p < 0.001 were considered as significantly different from untreated cells regarded as control.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
8 ANNA WYREzBSKA ET AL.

cytochrome c from the mitochondria to cytoplasm and deter-mines the apoptosis resistance. Suppression of Bcl-2 and/orup-regulation of Bax breaks this balance and leads to apopto-sis. The Bax/Bcl-2 ratio in cells treated with DL-3 increaseddrastically, indicating that apoptosis induced in cancer cellsmight be mediated by the mitochondrial pathway. This path-way is associated with changes of DΨm as well as changes inexpression of Bcl-2 family of proteins. DL-3 caused a signifi-cant increase of Bax and decrease of Bcl-2 mRNA and pro-tein levels, as well as dissipation of DΨm in both cell lines.Therefore, our results indicate that intrinsic pathway wasinvolved in DL-3-induced apoptosis in MDA-MB-231 andMCF-7 cells.The critical step of cancer dissemination leading to metas-
tasis is migration of cancer cells through the extracellularmatrix (ECM) [21,22]. Indispensable in this process is theactivation of proteolytic enzymes capable of degrading theECM. The key proteases involved in the ECM degradationare uPA [23] and matrix metalloproteinases (MMPs), espe-cially MMP-2 (gelatinase A) and MMP-9 (gelatinase B) [24].Expression and activity of all these enzymes changes inmany pathological conditions, including cancer. A simplerelationship has been established between overproduction ofMMPs or uPA in tumour cells and cancer progression. Toexplore the molecular mechanism underlying the DL-3-inducedinhibition of migration and invasive potential of MDA-MB-231 and MCF-7 cells, we investigated expression and secre-tion levels of MMP-9 and uPA. Significant down-regulationof these enzymes, both on mRNA and protein level, wasobserved, indicating that DL-3 is a potent inhibitor of theseenzymes.A series drawback of DL-3 anticancer action in breast
cancer cells is the fact that this compound is also highlycytotoxic for normal cells and therefore cannot be consid-ered as a potential therapeutic agent. However, simple andefficient preparation of a-methylene-d-lactones and theirpotential for further functionalization may enable selectivemodulation of their reactivity with biological nucleophiles,thus allowing for more specific interactions and better selec-tivity as drug candidates. DL-3 can be used as a lead struc-ture for further development of synthetic anticancer drugcandidates.
Conclusions
To the best of our knowledge, this is the first report demon-strating the broad range of anticancer activities produced bysimple synthetic compounds with an a-methylene-d-lactonemotif. We have found that 1-isopropyl-2-methylene-1,2-di-hydrobenzochromen-3-one, DL-3, can induce pro-apoptoticand antimetastatic effects in two breast cancer cell lines, hor-mone-independent MDA-MB-231 and oestrogen-dependentMCF-7. Cytotoxic action of DL-3 was related to the intrinsicpathway of apoptosis and was caused by breaking the balancebetween Bax and Bcl-2 and dissipation of mitochondrialmembrane potential. The underlying mechanism of suppressedcell migration and invasion involved down-regulation of genes
responsible for the degradation of the extracellular matrix.The results presented here indicate that simple synthetica-methylene-d-lactones, in particular DL-3, have great poten-tial as lead compounds for further structure–activity relation-ship studies in the search for new chemotherapy agentsagainst breast cancer.
AcknowledgementsThis work was supported by a grant from the Medical Uni-
versity of Lodz (No 503/1-156-02/503-01).
References
1 Cragg GM, Newman DJ. Plants as a source of anti-cancer agents.J Ethnopharmacol 2005;100:72–9.
2 Zhang S, Won YK, Ong CN, Shen HM. Anti-cancer potential ofsesquiterpene lactones: bioactivity and molecular mechanisms. CurrMed Chem Anticancer Agents 2005;5:239–49.
3 Pati HN, Das U, Sharma RK, Dimmock JR. Cytotoxic thiol alkyla-tors. Mini Rev Med Chem 2007;7:131–9.
4 Kupchan SM, Hemingway RJ, Werner D, Karim A, McPhail AT,Sim GA. Vernolepin, a novel elemanolide dilactone tumor inhibi-tor from Vernonia hymenolepis. J Am Chem Soc 1968;90:3596–7.
5 Weinheimer AJ, Chang CWJ, Matson JA. Naturally occurringcembranes. Fortschr Chem Org Naturst 1979;36:285–387.
6 Modranka J, Albrecht A, Jakubowski R, Krawczyk H, R�o _zalski M,Krajewska U et al. Synthesis and Biological Evaluation of a-Methylidene-d-lactones with 3,4-Dihydrocoumarin Skeleton. Bio-org Med Chem 2012;20:5017–26.
7 Mosmann T. Rapid colorimetric assay for cellular growth and sur-vival: application to proliferation and cytotoxicity assays. J Immu-nol Methods 1983;65:55–63.
8 Chomczynski P, Sacchi N. Single-step method of RNA isolationby acid guanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem 1987;162:156–9.
9 Winer J, Jung CK, Shackel I, Williams PM. Development and vali-dation of real-time quantitative reverse transcriptase-polymerasechain reaction for monitoring gene expression in cardiac myocytesin vitro. Anal Biochem 1999;270:41–9.
10 Rodriguez LG, Wu X, Guan JL. Wound-healing assay. MethodsMol Biol 2005;294:23–9.
11 Lewandowska U, Szewczyk K, Owczarek K, Hrabec Z, PodsezdekA, Koziołkiewicz M et al. Flavanols from Japanese quince (chae-nomeles japonica) fruit inhibit human prostate and breast cancercell line invasiveness and cause favorable changes in bax/bcl-2mRNA ratio. Nutr Cancer 2013;65:273–85.
12 Hrabec E, Strek M, Nowak D, Greger J, Suwalski M, Hrabec Z.Activity of type IV collagenases (MMP-2 and MMP-9) in primarypulmonary carcinomas: a quantitative analysis. J Cancer Res ClinOncol 2002;128:197–204.
13 Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011:the impact of eliminating socioeconomic and racial disparities onpremature cancer deaths. CA Cancer J Clin 2011;61:212–36.
14 Brooke DG, Shelley EJ, Roberts CG, Denny WA, Sutherland RL,Butt AJ. Synthesis and in vitro evaluation of analogues of avo-cado-produced toxin (+)-(R)-persin in human breast cancer cells.Bioorg Med Chem 2011;19:7033–43.
15 Ren Y, Acu~na UM, Jim�enez F, Garc�õa R, Mej�õa M, Chai H et al.Cytotoxic and NF-jB inhibitory sesquiterpene lactones from Pipto-coma rufescens. Tetrahedron 2012;68:2671–8.
16 Telang NT, Li G, Sepkovic DW, Bradlow HL, Wong GY. Anti-proliferative effects of Chinese herb Cornus officinalis in a cellculture model for estrogen receptor-positive clinical breast cancer.Mol Med Rep 2012;5:22–8.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
ANTICANCER ACTIVITY OF NEW SYNTHETIC a-METHYLENE-d-LACTONES 9

17 Hu W, Kavanagh JJ. Anticancer therapy targeting the apoptoticpathway. Lancet Oncol 2003;4:721–9.
18 Qi F, Li A, Zhao L, Xu H, Inagaki Y, Wang D et al. Cinobufacini,an aqueous extract from Bufo bufo gargarizans Cantor, inducesapoptosis through a mitochondria-mediated pathway in human hepa-tocellular carcinoma cells. J Ethnopharmacol 2010;128:654–61.
19 Reed JC. Apoptosis-targeted therapies for cancer. Cancer Cell2003;3:17–22.
20 Adams JM, Cory S. The Bcl-2 protein family: arbiters of cell sur-vival. Science 1998;281:1322–6.
21 Steeg PS, Theodorescu D. Metastasis: a therapeutic target for can-cer. Nat Clin Pract Oncol 2008;5:206–19.
22 Geiger TR, Peeper DS. Metastasis mechanisms. Biochim BiophysActa 2009;1796:293–308.
23 Duffy MJ. The urokinase plasminogen activator system: role inmalignancy. Curr Pharm Des 2004;10:39–49.
24 Das S, Banerji A, Frei E, Chatterjee A. Rapid expression and acti-vation of MMP-2 and MMP-9 upon exposure of human breast can-cer cells (MCF-7) to fibronectin in serum free medium. Life Sci2007;82:467–76.
© 2013 Nordic Pharmacological Society. Published by John Wiley & Sons Ltd
10 ANNA WYREzBSKA ET AL.



![Targeting macrophage checkpoint inhibitor SIRPα for anticancer … · 2020. 6. 18. · lymphocyte–associated protein 4 [CTLA-4] and programmed death 1 [PD-1]), or their ligands](https://static.fdocument.org/doc/165x107/5fd9a9e449b9f25d9f5898e6/targeting-macrophage-checkpoint-inhibitor-sirp-for-anticancer-2020-6-18-lymphocyteaassociated.jpg)



![Highly Branched Poly(α-Methylene-γ-Butyrolactone) from …file.scirp.org/pdf/OJPChem_2017112914172525.pdf · 2017-12-01 · ... (3.00 g, 0.013 mol), and L-valinol [(S)-(+)-2-Amino-3-methyl-1-butanol]](https://static.fdocument.org/doc/165x107/5b1be3007f8b9a28258f0d54/highly-branched-poly-methylene-butyrolactone-from-filescirporgpdfojpchem.jpg)










