
β-1,3-Glucanase, chitinase, and peroxidase activities in tobacco tissues resistant and susceptible...
8
Plrr+io/sgtealand Molecular PlantPatholstt ' 1991 39 . 43344f) 433 /3-1,3-Glucanase,chitinase,andperoxidaseactivitiesin tobaccotissuesresistantandsusceptibletobluemould asrelatedtoflowering,ageandsuckerdevelopment SARAHE .WYATT,SHENQ. PAN and JOSEPH Ki , (. Dc/sartnientofPlantPathology, University ofhentuckp,Letntgton .kentaekr40546. 1-SA . Irrupted/orpuhlicatimtSeptember19.91 Iobaccobecomesnaturallyresistanttobluemouldcausedby Peronospmatabacina asitages .This age-relatedresistanceiscorrelatedtothetimeofnormalfloraldevelopmentandthesenescence oflowerleaves ;however,itisdependentonneither ./3-1,3-Glucanase,chitinaseandperoxidase activitiesincreaseintobaccowithage .Theseincreasescorrelatewiththedevelopmentofage- relatedresistancetobluemouldandwereindependentoffloraldevelopment .Additionally,/1-1,3- glucanase .chitinase,andperoxidaseactivitieswerehigherinleaftissuefromthemainstalk resistanttobluemould)acomparedtoleaftissuefromsuckeringstemssusceptibletoblue mould'onthesameplant .Isozymepatternsof/3-1,3-glucanaseandchitinaseinallresistanttissues aretypicalofthoseoftissuessystemicallyprotectedbyeithersteminjectionwith Peronospora tabacina orfoliarinoculationwithTMV . INTRODUCTION Increasesinchitinase,f-1,3-glucanaseandperoxidaseactivitieshavebeenassociated with immunizationinseveralplant-pathogensystems [4,5,7,10,11,14,17 1 . Pathogenesis-related!PR)proteinshavealsobeenassociatedwithinducedresistance [8,19,20 ] . Tuzun etal . [17] demonstratedthepresenceofseveralPRproteinsinnon- inoculatedleavesoftobaccoplantssystemicallyimmunizedagainstbluemouldbystem injectionwith Peronosporatabacina . Theseproteinsappearedpriortochallenge,and increasedmorerapidlyafterchallengeinimmunizedplantsthanincontrols .Ye et al . [20]demonstratedthatasimilarpatternofPRproteinswaselicitedintobacco immunizedbyfoliarinoculationwithtobaccomosaicvirus ;TMV) .Theappearance andconcentrationoftheseproteinscoincidedwiththeappearanceandlevelof protectionagainstbluemould [20] . Severaloftheseproteinshavebeenidentifiedas chitinasesandf-1,3-glucanases [12,17] . However,Fraser [6] reportedthatPR proteinsaccumulatedinhealthytobaccoatthetimeofinflorescencedevelopmentand senescenceoflowerleaves .Lotan etal . [9]alsosuggestedthatPRproteinaccumulation wascorrelatedwiththenormaldevelopmentoffloweringintobacco . Resistancetobluemouldnaturallyoccursintobaccoasitages[15] . Thisage-related resistanceisassociatedwiththetimeofnormalfloraldevelopmentandsenescenceof Abbreviationsusedintext :PR,pathogenesis-related ;f''s .,fromseeding ;I'MV,tobaccomosaicvirus ; PACE,polyacrylamidegelelectrophoresis ;GlcNAc, .~'-acetyl-n-gLucosumiue . 088 .)-3765/91/120433+08$03 .00/0 tC 1991 AcademicPressLimited
Transcript of β-1,3-Glucanase, chitinase, and peroxidase activities in tobacco tissues resistant and susceptible...

Plrr+io/sgteal and Molecular Plant Patholst t ' 1991 39 . 433 44f)
433
/3-1,3-Glucanase, chitinase, and peroxidase activities intobacco tissues resistant and susceptible to blue mouldas related to flowering, age and sucker development
SARAH E . WYATT, SHEN Q. PAN and JOSEPH Ki , (.Dc/sartnient of Plant Pathology, University of hentuckp, Letntgton . kentaekr 40546. 1-SA.
Irrupted /or puhlicatimt September 19.91
I obacco becomes naturally resistant to blue mould caused by Peronospma tabacina as it ages . Thisage-related resistance is correlated to the time of normal floral development and the senescenceof lower leaves ; however, it is dependent on neither . /3-1,3-Glucanase, chitinase and peroxidaseactivities increase in tobacco with age. These increases correlate with the development of age-related resistance to blue mould and were independent of floral development . Additionally, /1-1,3-glucanase. chitinase, and peroxidase activities were higher in leaf tissue from the main stalkresistant to blue mould) a compared to leaf tissue from suckering stems susceptible to blue
mould' on the same plant . Isozyme patterns of/3-1,3-glucanase and chitinase in all resistant tissuesare typical of those of tissues systemically protected by either stem injection with Peronosporatabacina or foliar inoculation with TM V .
INTRODUCTION
Increases in chitinase, f-1,3-glucanase and peroxidase activities have been associatedwith immunization in several plant-pathogen systems [4, 5, 7, 10, 11, 14, 17 1 .Pathogenesis-related !PR) proteins have also been associated with induced resistance[8, 19, 20 ] . Tuzun et al . [17] demonstrated the presence of several PR proteins in non-inoculated leaves of tobacco plants systemically immunized against blue mould by steminjection with Peronospora tabacina . These proteins appeared prior to challenge, andincreased more rapidly after challenge in immunized plants than in controls . Ye et al .[20] demonstrated that a similar pattern of PR proteins was elicited in tobaccoimmunized by foliar inoculation with tobacco mosaic virus ;TMV) . The appearanceand concentration of these proteins coincided with the appearance and level ofprotection against blue mould [20] . Several of these proteins have been identified aschitinases and f-1,3-glucanases [12, 17] . However, Fraser [6] reported that PRproteins accumulated in healthy tobacco at the time of inflorescence development andsenescence of lower leaves . Lotan et al . [9] also suggested that PR protein accumulationwas correlated with the normal development of flowering in tobacco .
Resistance to blue mould naturally occurs in tobacco as it ages [15] . This age-relatedresistance is associated with the time of normal floral development and senescence of
Abbreviations used in text : PR, pathogenesis-related ; f' 's ., from seeding ; I'M V, tobacco mosaic virus ;PACE, polyacrylamide gel electrophoresis ; GlcNAc, .~'-acetyl-n-gLucosumiue .
088.)-3765/91/120433+08 $03 .00/0
tC 1991 Academic Press Limited

434
S . E. Wyatt et at.
lower leaycs ; however . i t is dependent ()It neither 'These reports 17rontpted us tut
examine f-I,3-glucanasc . chitinase and peroxidase activities iii premature!v ~utdnorrnall flowering tobacco to determine if these euzvtnr activ -itie, ssr •rc induted 1»floral development or correlated with age-related resistance .
MATERIALS AND METHODS
Tobacco plantsBurley tobacco (,,A'icotiana tabacum I . .1 cv. Ky 14 seeds were sown in Metro inix 2,50(W. R. Grace Co ., L ; .S ._1 .` in small trays . After 2 weeks, groups of' 12 seedlings weretransferred to trays in the same mix . The seedlings were fertilized daily soh Peter'sfertilizer 15-16-17 at half the recommended rate (1 . 5 gl t ! . .After another 2 weeks,individual seedlings were transplanted into 6-inch pots in Metro mix 260 . At 28--36days from seeding (Es . , plants were moved to Percival growth chambers . Half weresubjected to cool temperature, short day- conditions 18 °C, 8 h light, 16 It dark ; toinitiate premature flowering . The remainder were placed under warm long dayconditions (28 °C, 8 h light, 15 h dark with a I h night break at the midpoint ; . 'T)eseplants flowered normally at 11-12 weeks f .s . Plants remained under these conditions for21 days and were then returned to the greenhouse until challenge. Suckering wasinduced by removing the apical bud from 9-week-old plants grown in the greenhouseunder a 14 h photoperiod (daylight, supplemented with sodium lights) at 25--33 °C .
Pathogen culture and challenge inoculationPeronospora tabacina Adam isolate 79 . obtained in Kentucky in 1979, was maintained byweekly transfers of sporangiospores on young cy . KN 14 tobacco plants . Sporulatingleaves were frozen and stored at -20 °C between s-a et paper towels in scaled paperbags. Inoculation suspensions were made b y gently brushing sporangiospores from thefrozen leaves into distilled water . 'I'he resulting suspension was filtered through fourlavers of' cheesecloth, and the sporangiospores were washed three times with distilledwater, and were collected on 8 .0 pin membranes . Sporangiospores were thenresuspended in distilled water and the concentration adjusted to 1 x 10' spores ml ' .For challenge, plants were moved 24 h prior to challenge to a growth room illuminatedwith white fluorescent and incandescent light with a 14 h photoperiod at 23 ° C. Theupper surfaces of leaves were sprayed with a uniform mist of'P . tabacina sporangiosporesat I x 10 4 spores ml - ', and plants were covered with wet black plastic bags and kept inthe dark for 16 h .
Immunization of tobacco'The upper surfaces of three lower leaves of tobacco plants at 10 weeks [s . were dustedwith carborundum and were gently rubbed with 25 pg ml -' ofTMV in distilled water .At least one fully expanded leaf of each plant was left uninoculated . 'The leaves wereimmediately rinsed with water after inoculation . Plants were subsequently challengedwith P . tabacina at 14 days after inoculation .
Evaluation of disease resistanceResistance to blue mould ss as evaluated 7 days after challenge and based on a visualestimation of the percentage of the leaf area covered by chlorotic lesions 0 = no

Blue mould and tobacco enzymes and development
435
chlorosis evident, 1 = 1-25 °-o, 2 = 26-50" 0 „ 3 = 51-75 11 ,,, 4 = > 75 °,;, chlorosis) . Thefirst five expanded leaves from the top of the plant were assessed for disease resistance .Experiments were designed using a blind protocol and ratings were performed by animpartial researcher .
Extraction of proteinsLeaf tissues, with midribs removed, were collected at weekly intervals prior to and afterchallenge, and were immediately stored at -20 °C. Proteins were extracted fromfrozen tissues (1-4 g fresh weight) in 2 ml g - ' of 0 . 05 M sodium acetate buffer, pH 5 . 0,in a prechilled mortar and pestle at 4 °C . The homogenate was filtered through nylonsieve cloth and the filtrate was centrifuged at 12 500 g for 10 min . The supernatant wasdialysed against water at 4 °C for 24 h, then against 0 . 05 M sodium acetate buffer foranother 2 h. Extracts used for the determination ofisozyme patterns were concentratedby freeze-drying . Protein was determined by the method of Bradford [3] with y-globulin as standard . All samples were stored at -20 °C ; until needed .
En.cpine activity determinationsf-1,3-Glucanase activities were determined using the lamina rin-dinitrosalicylatecolorimetric assay of Abeles et al . [1] . The hydrolysis of colloidal chitin to V-acetylglucosamine, determined by the colorimetric assay of Boller & Mauch [2], was utilizedto determine chitinase activities . Chitinase and /-1,3-glucanase isozymes and proteinpatterns were detected after a single separation using native polyacrylamide gelelectrophoresis (PAGE) by the methods of Pan et al . [12] . Following electrophoreticseparation, PAGE gels attached to supporting glass plates were incubated in 0 . 1 Msodium acetate (pH 5 . 0) . They were then covered with a 7 . 5°,,, (0 . 75 mm thick lpolyacrylamide overlay gel containing 0 . 04"(, glycol chitin, synthesized according toTrudel & Asselin [16], in 0 . 1 M sodium acetate (pH 5 . 0 ; . The gels were incubated at40 ° C for 1 . 5 h under moist conditions . Overlay gels were then incubated in freshlyprepared 0 . 01 "-~, (w/v) fluorescent brightener 28 ;(Sigma, St Louis, Missouri, in500 mM tris-HCI (pH 8 .9) at room temperature for 5 min. Chitinase isozymes werevisualized as cleared zones, by placing the overlay gels on a UV transilluminator, andphotographed. To stain for fl-1,3-glucanases, the PAGE gels were washed, thenincubated at 40 ° C for 30 min in a mixture containing 75 ml of 0 .05 M sodium acetate(pH 5 . 0 and I g of laminarin (Sigma) dissolved in 75 ml of water . The gels were thenincubated in a mixture of methanol, water and acetic acid (5 :5 :2) for 5 min, washedwith water, and stained with 0 . 3 g of 2,3,5-triphenyltetrazolium chloride in 200 ml of1 . 0 M NaOH in a boiling water bath until red hands appeared (approx . 10 min' .Peroxidase activity was determined using guaicol as substrate .
RESULTS
Time-course studiesResistance to blue mould increases with age in Burley tobacco with significantresistance by 11-12 weeks [15, 1 .9] . fl-1,3-Glucanase, chitinase and peroxidase activitiesincreased with plant age and were correlated with the occurrence of age-relatedresistance (Fig . 1) . Neither resistance nor enzyme activities were increased by floral
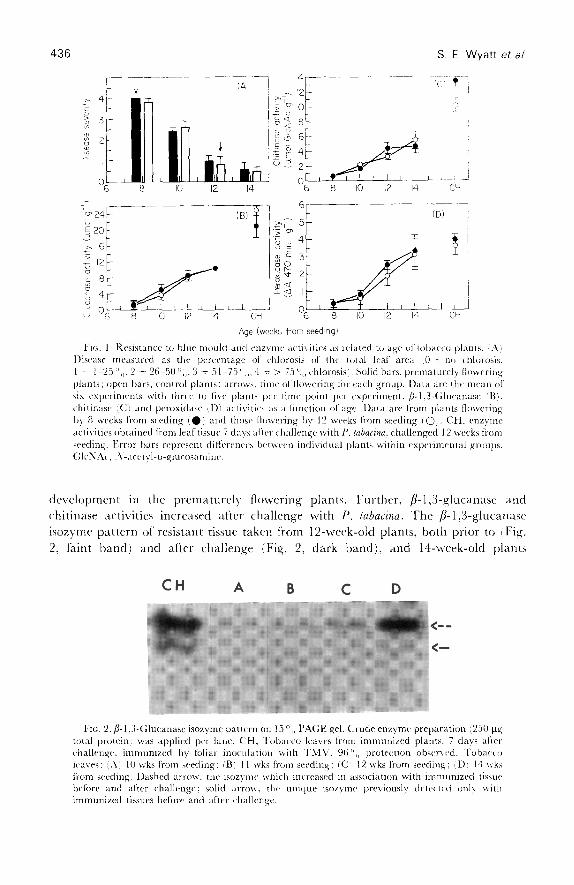
436
S . E. Wyatt et al.
CH A B C
CH
CT 24o20
>, 16
12U0
80
8
10
12
14
CH
6
8
10
12
14
CH
Age (weeks from seeding)I tc . 1. Resistance to blue mould and enzyme activities as related to age of tobacco plants . !Aj
Disease measured as the percentage of chlorosis of the total leaf area l0 = no chlorosis,1 = 1-25 °, ,, 2 = 26-50 3 = 51-75 °,,, 4 = > 75 °„ chlorosis) . Solid bars, prematurely floweringplants ; open bars, control plants ; arrows, time of flowering for each group . Data are the mean ofsix experiments with three to five plants per time point per experiment . /3-1 .3-Glucanase lBl .chitinase (C) and peroxidase ;Dl activities as a function of age . Data are from plants floweringby 8 weeks from seeding (S) and those flowering by 12 weeks from seeding 101 . CH, enzymeactivities obtained from leaf tissue 7 days after challenge with P . labacina, challenged 12 weeks fromseeding . Error bars represent diflcrences between individual plants within experimental groups .GIcNAc, :A`-acetyl-n-glucosamine .
development in the prematurely flowering plants . Further, fl-1,3-glucanase andchitinase activities increased after challenge with P . labacina . The fl-1,3-glucanaseisozyme pattern of resistant tissue taken from 12-week-old plants, both prior to (Fig .2, faint band) and after challenge (Fig. 2, dark band), and 14-week-old plants
Fte . 2 . f-1,3-Glen mase isozyme pattern on 15 PAGE gel . Crude enzyme preparation (250 pgtotal proteinn was applied per lane . CH, Tobacco leaves from immunized plants . 7 days afterchallenge, immunized by foliar inoculation with TMC. 96 protection observed . 'hobacco(caves : IA 1 10 wks from seeding, (B) 11 wks from seeding ; (Ci 12 wks from seeding ; ;Dl 14 wksfrom seeding . Dashed arrow, the isozyme which increased in association with immunized tissuebefore and after challenge ; solid arrow', the unique isozyme previously detected only withimmunized tissues before and after challenge .
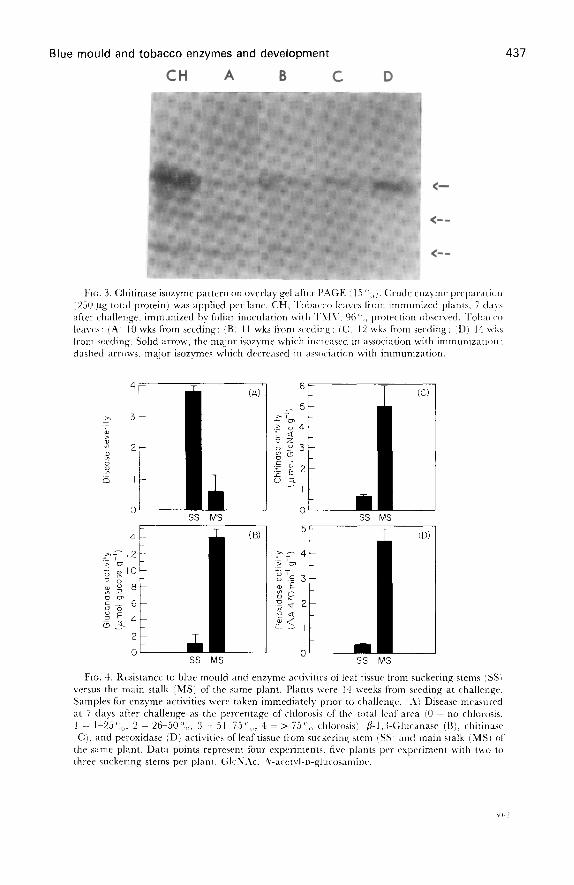
Blue mould and tobacco enzymes and development
437
CH
A B
a
0
2
0
SS MS
(A)
(B)
6
5
73
v 4cr•
z•
2 3o O0 o•
B 2i
i
0
5
C D
8 1
( C)
SS MS
(D)
SS MS
0
SS MS
<-
<--
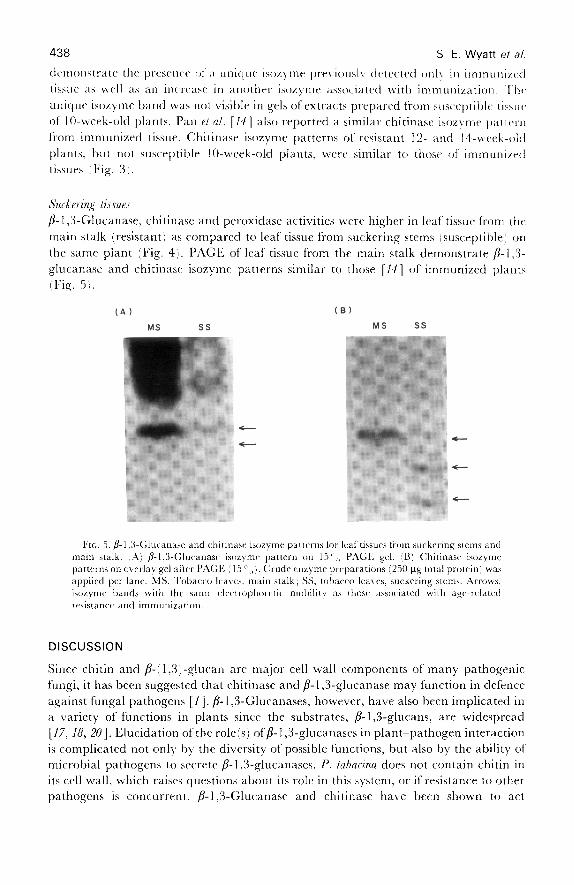
Ft(;. 3. Chitinase isozyme pattern on overlay gel after PAGE (15' . Crude enzyme preparation250 µg total protein) was applied per lane . CH, Toba( o leaves from immunized plants, 7 daysafter challenge, immunized by foliar inoculation with 'I \1\-, 96 protection observed . Tobaccoleaves : (A) 10 wks from seeding : ;B) I I wks from seeding : fC : 12 wks from seeding : ;1) 14 wksfrom seeding . Solid arrow, the major isozyme which increased in association with immunization ;dashed arrows, major isozymes which decreased in association with immunization .
Fio . 4 . Resistance to blue mould and enzyme activities of leaf tissue from suckering stems (SS 1versus the main stalk !MS) of the same plant. Plants were 14 weeks from seeding at challenge.Samples for enzyme activities were taken immediately prior to challenge . (A! Disease measuredat 7 days after challenge as the percentage of chlorosis of the total leaf area (0 = no chlorosis,I = 1-25'_ 2 = 26-501 , 3 = 51--75 °,,, 4 = > 75 °,, chlorosis! . /3-1 .3-Glucanase (B), chitinaseC), and peroxidase (D) activities of leaf tissue from suckering stem (SS) and main stalk (MS) ofthe same plant. Data points represent four experiments, five plants per experiment with two tothree suckering stems per plant . GlcNAc, .9-acetyl-D-glucosamine .
438
S . E. Wyatt et al .
demonstrate the presence of a unique isozyme previously detected only in immunizedtissue as well as an increase in another isozyme associated with immunization . Theunique isozyme band was not visible in gels of extracts prepared front susceptible tissueof 10-week-old plants . Pan et al . [14] also reported a similar chitinase isozyme patternfrom immunized tissue . Chitinase isozyme patterns of resistant 12- and 14-week-oldplants, but not susceptible 10-week-old plants. were similar to those of immunizedtissues ( Fig. 3 ; .
Suckering Tissuesfl- 1,3-Glucanase, chitinase and peroxidase activities were higher in leaf tissue from themain stalk (resistant) as compared to leaf tissue from suckering stems (susceptible) onthe same plant (Fig . 4) . PAGE of leaf tissue from the main stalk demonstrate /3-1,3-glucanase and chitinase isozyme patterns similar to those [14] of immunized plantsFig. 51 .
to 1
MS SScel
MS SS
Fic . 5 . /3-1,3-Glucanase and chitinase isozyme patterns for leaf tissues from suckering stems andmain stalk . iA,l f3-1,3-Glucanase isozyme pattern on 15"„ PAGE gel . (B) Chitinase isozymepatterns on overlay gel after PAGE (15 „i . Crude enzyme preparations (250 µg total protein] wasapplied per lane . VIS, Tobacco leaves, main stalk ; SS, tobacco leaves, suckering stems . Arrows,isozyme bands with the same electrophoretic mobility as those associated with age-relatedresistance and immunization .
DISCUSSION
Since chitin and /3-(1,3)-glucan are major cell wall components of many pathogenicfungi, it has been suggested that chitinase and /3-1,3-glucanase may function in defenceagainst fungal pathogens [1] . /3-1,3-Glucanases, however, have also been implicated ina variety of functions in plants since the substrates, /3-1,3-glucans, are widespread[17, 18, 20] . Elucidation of the role(s) of/3-1,3-glucanases in plant-pathogen interactionis complicated not only by the diversity of possible functions, but also by the ability ofmicrobial pathogens to secrete /3-1,3-glucanases . P. tahacina does not contain chitin inits cell wall, which raises questions about its role in this system, or if resistance to otherpathogens is concurrent

Blue mould and tobacco enzymes and development
439
synergistically in the partial degradation of isolated fungal cell walls containing chitin,and, in combination, strongly inhibit growth of several fungi containing chitinincluding those not inhibited by either enzyme alone [18] . Parallel increases may heimportant for their optimal function in plant defence .
Increased /4-1,3-glucanase, chitinase and peroxidase activities in the tobacco tissuestested are not dependent on floral development . Increases were associated with plantage and related to the development of resistance . These increases, both before and afterchallenge, are similar to those seen in tissues protected by immunization [13, 14] .
This suggests that immunization in tobacco may result from physiological changesand hormonal regulation related to ageing . Conventional assays of total /1-1,3-glucanase and chitinase activities using crude extracts have limited value for studyingthe roles of these enzymes . Such assays do not distinguish isozymes, although differentisozymes might be involved in the diverse roles of the enzymes . Using the PAGE systemdescribed, we were able to determine the specific /1-1,3-glucanase and chitinaseisozymes involved in the increases seen in resistant tissues . Pan eat al . [14] found thatelectrophoretic separation of intercellular fluids of uninfected leaves of immunizedplants revealed one dominant acidic intercellular isozyme of'#- 1,3-glucanase which wasassociated only with immunized tissues both prior to and after challenge . It was notfound in control tissues . Although a causal relationship has not yet been established,the increases in ,8-1,3-glucanases and chitinases after challenge and the occurrence ofthese enzymes in all resistant tissues tested further supports the supposition that theseenzymes are important in resistance mechanisms . The occurrence of fl- 1,3-glucanaseand chitinase isozyme patterns typical of resistant tissues immunized by priorinoculation with P. tabacina or TMV (induced systemic resistance) further supports thehypothesis that these enzymes are important in resistance . The role for peroxidase inresistance is not as clear . Peroxidase, although potentially important in resistantmechanisms as a generator of toxic free radicals and lignin, may be more closelyassociated with leaf senescence .
"The research reported in this manuscript was supported in part by grants from theR . J . Reynolds Tobacco Company and Cooperative Agreement 43YK-5-0030 of theUSDA/ARS. Journal paper 91-11-92 of the Kentucky Agricultural ExperimentStation, Lexington, Kentucky .
REFERENCES
1 . ABELES, F. B ., BOSSHART, R . P ., FORRENCE, L . E . & HABIG, W . H . ; 1971 . . Preparation and purificationof glucanase and chitinase from bean leaves . Plant Physiology 47, 129-134 .
2 . BOLLER, T . & MAUCH, F . (1988'i . Colorimetric assay for chitinase . Methods in Enzymology 161, 430-435 .3 . BRADFORD, M . M . (1976) . A rapid and sensitive method for the quantitation of microgram quantities
of protein utilizing the principle of protein-dye binding . Analytical Biochemistry 72, 248-254 .4 . CHAT, H . B . & DOKE, N . (1987) . Systemic activation of 0 2 -generating reaction, superoxide dismutase,
and peroxidase in potato plants in relation to induction of systemic resistance to Phvtophthora infestans .Annals of the Phytopathological Society of Japan 53, 585-590 .
5 . DALISAY, R . F. & Kcd, J . (1988) . Enhanced peroxidase and its persistence in cucumber plantsimmunized against anthracnose by foliar inoculation with Colletotrichum lagenarium . Ph_vtopathologv 79,1150 (abstr ; .
6 . FRASER, R. S . S . (1981) . Evidence for the occurrence of the "pathogenesis-related" proteins in leases ofhealthy tobacco plants during flowering . Physiological Plant Pathology 19, 69-76 .

440
S. E. Wyatt et al
7 . HAM3n;RSCHMInT, R., -Nu cm ts . E . & Ki
1982 . Association of cnhanccd pcroxtdasc 1(11\ It\ mil :nduccd so sicmic resistance hi ucumbcr to (,illrlotridunu /q,cnrrntnn . ll! iolngiizrl P/artt Pnthulnpr 20,71 82 .
8 . KASSANis, B . & t1-HI i . R . 1 . 1978_ l .fhCct of polyacr)-tic acid and b-proteins on I N1V' nuiltiplir nir,nin tobacco protoplasts . Pkv(apatholo,,is he `eitsdrifi 91, 269-272 .
9 . I .O'rA'o, T ., Out, A . & FIA HR . R . ; 1989' . Pathogenesis-related protcius are CIC\ clopmrntalls re 3ulatc(lill tobacco floc+ers . Plant (all 1, 881 887 .
10 . ME IRAt?x,,j . P . & Bott .e.R .'1' . H . 1986 . Local and svstcmic induction of chitinase Ill( ucumber plantsin response to viral, bacterial . and fun,t;al infections . Pltvnnlo,ical and aloleodar Plant Patholo8, 28,161 - 169 .
11 . b1SrRAOX, J . P ., S'rRETi, L . & S'rACH, I H . 1988! . A pathogenesis-related protein in cucumber is achitinase . Phvsiological and Molecular P/ant Patho/ogv 33, 1-9) .
12 . Pas, S . Q,., Ys, X . S . & Ki c,,J . 1989' . Direct detection of//-1 .3-glucanase isozymes oil polyacrylamideelectrophoresis and isoelectrofbcusing gels . dnnhli(ol Biodonnitrv 182, 136-140 .
13 . PAN, S . Q., YL, X . S . & Kt (", .J . i 1991 ; . Association off-1,3 glucanase activity, and isoform pattern withss-stemie resistance to blue mold in tobacco induced by stem injection with Perom)spora tabacina or lealinocultion with tobacco mosaic virus . Physiological and 3Alolecular Plant Pathology 39, 25- 39 .
14 . PA's, S . Q, YE, X . S . & Ktc,,J . (1991
Induction ofchitinases in tobacco plants systemically protectedagainst blue mold by Peronospool tabacina or tobacco mosaic eirus . Phvlopathologt !in press') .
15 . REUVENI, A-1-'1m it s, S ., COLE, J . S ., SiLOEt., M9 . R . & Kuu .,j . 19861 . The eflects of plant age and leafposition on the susceptibility of tobacco to him mold caused by Peronospora tabacina . Phvtapathology 76,477 458 .
16 . IRt DEL,,J . & AssELIN, A . 1989 Detection of chitinase O tis l's after polyacrolanude gel dectrophorests .Analytical Biachemislit 178, 362-366 .
17 . '1vzus . S ., RAO, &1 . N ., VOOErI, U., SmHARDt, C. L. & Kr( , J . ; 1988, . Early accumulation of chitinases,/3-1,3-glucanases and other b-proteins in tobacco immunized against blue mold . Ph_ytspathnlogt' 78,1556 ;abstr) .
18 . A'ocsii, C ., MEINS, F. & BOILER, B . ' 1988 . Co-ordinated regulation of chitinase and /3-1,3-glucanasein bean leaves . Planta 174, 364--372 .
19 . A1'V err, S . 1? . & Kung. , 1990 ; . hhe cflccts of age, Ibnsering, and senescence on the resistance of tobaccoto blue mold . Phytopathologv 80, 1000 shotri .
20 . YE. X . S ., Pas, S . Q. & Kuc, J . (1989x . Pathogenesis-related protc61s and systemic resistance to bluemold and tobacco mosaic virus induced by tobacco mosaic sires, Peronospora tahaeina and aspirin .Physiological and Molecular Plant Pathology 35, 161-175 .

![RESEARCH ARTICLE Open Access Loss of the interferon-γ...mania [36], and Trypanosoma [37]. Irgm1 −/ mice are also reported to be unusually susceptible to lipopolysac-charide injection](https://static.fdocument.org/doc/165x107/6092380d80c8922067614dc3/research-article-open-access-loss-of-the-interferon-mania-36-and-trypanosoma.jpg)










