
Motivation for Top-Up: A beamline perspective David Paterson Top-Up Workshop.
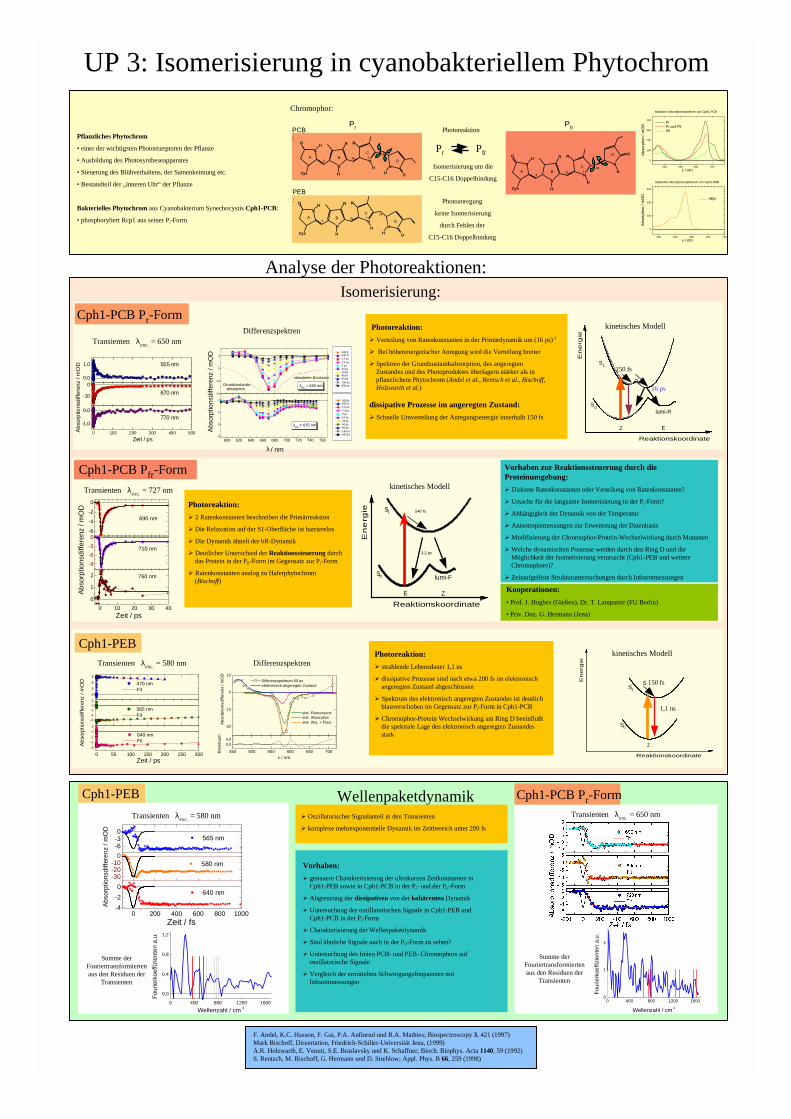
UP 3: Isomerisierung in cyanobakteriellem Phytochrom
400 500 600 700
0
100
200
300
400
statische Absorptionsspektren von Cph1-PCB
Pr Pr und Pfr Pfr
Abso
rptio
n / m
OD
λ / nm
500 550 600 650 700
0
100
200
300
statisches Absorptionsspektrum von Cph1-PEB
PEB
Abso
rptio
n / m
OD
λ / nm
0 100 200 300 400 500
-1,0
0,0
0,0
1,0
-10
0
770 nm
Zeit / ps
555 nm
670 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
Pflanzliches Phytochrom
• einer der wichtigsten Photorezeptoren der Pflanze
• Ausbildung des Photosyntheseapparates
• Steuerung des Blühverhaltens, der Samenkeimung etc.
• Bestandteil der „inneren Uhr“ der Pflanze
Bakterielles Phytochrom aus Cyanobakterium Synechocystis Cph1-PCB:
• phosphoryliert Rcp1 aus seiner Pr-Form
Photoanregung
keine Isomerisierung
durch Fehlen der
C15-C16 Doppelbindung
Cph1-PEB
Cph1-PCB Pr-Form
Isomerisierung:
Wellenpaketdynamik:
Cph1-PCB Pfr-Form
Pr Pfr
Isomerisierung um die
C15-C16 Doppelbindung
Photoreaktion
Analyse der Photoreaktionen:
0 10 20 30 400
1
2
-9-6-30
-6
-4
-2
0
760 nm
Zeit / ps
710 nm
690 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
Transienten λexc. = 727 nm
ReaktionskoordinateE Z
lumi-F
En
erg
ie
S0
S1 540 fs
3,2 ps
kinetisches Modell
Transienten λexc. = 650 nmDifferenzspektren
-6-4-202
0 50 100 150 200 250 300-6-4-202
0369
565 nm Fit
Abso
rptio
nsdi
ffere
nz /
mO
D
640 nm Fit
Zeit / ps
470 nm Fit
Transienten λexc. = 580 nm
450 500 550 600 650 700
0,04,0
-20
-10
0
10
Abso
rptio
nsdi
ffere
nz /
mO
D
Res
iduu
m
λ / nm
Differenzspektrum 60 ps elektronisch angeregter Zustand
stat. Fluoreszenz stat. Absorption stat. Abs. + Fluor.
Differenzspektren
Reaktionskoordinate
EZ
lumi-R
En
erg
ie
S0
S1
kinetisches Modell
150 fs
16 ps
Reaktionskoordinate
Z
Energ
ie
S0
S1
1,1 ns
≤ 150 fs
kinetisches Modell
Transienten λexc. = 580 nm Transienten λexc. = 650 nm
Cph1-PEB Cph1-PCB Pr-Form
Vorhaben zur Reaktionssteuerung durch dieProteinumgebung:� Diskrete Ratenkonstanten oder Verteilung von Ratenkonstanten?
� Ursache für die langsame Isomerisierung in der Pr-Form?
� Abhängigkeit der Dynamik von der Temperatur
� Anisotropiemessungen zur Erweiterung der Datenbasis
� Modifizierung der Chromophor-Protein-Wechselwirkung durch Mutanten
� Welche dynamischen Prozesse werden durch den Ring D und die Möglichkeit der Isomerisierung verursacht (Cph1-PEB und weitere Chromophore)?
� Zeitaufgelöste Strukturuntersuchungen durch Infrarotmessungen
Vorhaben:� genauere Charakterisierung der ultrakurzen Zeitkonstanten in Cph1-PEB sowie in Cph1-PCB in der Pr- und der Pfr-Form
� Abgrenzung der dissipativen von der kohärenten Dynamik
� Untersuchung der oszillatorischen Signale in Cph1-PEB und Cph1-PCB in der Pr-Form
� Charakterisierung der Wellenpaketdynamik
� Sind ähnliche Signale auch in der Pfr-Form zu sehen?
� Untersuchung des freien PCB- und PEB- Chromophors auf oszillatorische Signale
� Vergleich der ermittelten Schwingungsfrequenzen mit Infrarotmessungen
Pfr
Chromophor:
N
H
RN
O
Cys
H
NH
R
NH O
+A BC
D
Pr
510
15
PCB:
PEB:
1510
5 D
CBA +
N
H
RN
O
Cys
H
NH
R
OHN
N
H
R
N
O
Cys
H
N
H
R
N
H
O
+A BC
D
510
15
Photoreaktion:� Verteilung von Ratenkonstanten in der Primärdynamik um (16 ps)-1
� Bei höherenergetischer Anregung wird die Verteilung breiter
� Spektren der Grundzustandsabsorption, des angeregten Zustandes und des Photoproduktes überlagern stärker als in pflanzlichem Phytochrom (Andel et al., Rentsch et al., Bischoff, Holzwarth et al.)
dissipative Prozesse im angeregten Zustand:� Schnelle Umverteilung der Anregungsenergie innerhalb 150 fs
Photoreaktion:� 2 Ratenkonstanten beschreiben die Primärreaktion
� Die Relaxation auf der S1-Oberfläche ist barrierelos
� Die Dynamik ähnelt der bR-Dynamik
� Deutlicher Unterschied der Reaktionssteuerung durch das Protein in der Pfr-Form im Gegensatz zur Pr-Form
� Ratenkonstanten analog zu Haferphytochrom (Bischoff)
Kooperationen:• Prof. J. Hughes (Gießen), Dr. T. Lamparter (FU Berlin)
• Priv. Doz. G. Hermann (Jena)
Photoreaktion:� strahlende Lebensdauer 1,1 ns
� dissipative Prozesse sind nach etwa 200 fs im elektronisch angeregten Zustand abgeschlossen
� Spektrum des elektronisch angeregten Zustandes ist deutlich blauverschoben im Gegensatz zur Pr-Form in Cph1-PCB
� Chromophor-Protein Wechselwirkung am Ring D beeinflußt die spektrale Lage des elektronisch angeregten Zustandes stark
0 400 800 1200 1600
0,0
0,4
0,8
1,2
Four
ierk
oeffi
zien
ten
a.u.
Wellenzahl / cm-1
Summe derFouriertransformiertenaus den Residuen der
Transienten
Summe derFouriertransformiertenaus den Residuen der
Transienten
� Oszillatorischer Signalanteil in den Transienten
� komplexe mehrexponentielle Dynamik im Zeitbereich unter 200 fs
F. Andel, K.C. Hasson, F. Gai, P.A. Anfinrud und R.A. Mathies; Biospectroscopy 3, 421 (1997)Mark Bischoff, Dissertation, Friedrich-Schiller-Universität Jena, (1999)A.R. Holzwarth, E. Venuti, S.E. Braslavsky und K. Schaffner; Bioch. Biophys. Acta 1140, 59 (1992)S. Rentsch, M. Bischoff, G. Hermann und D. Strehlow; Appl. Phys. B 66, 259 (1998)
600 620 640 660 680 700 720 740 760-15
-10
-5
0
-15
-10
-5
0
λanr. = 615 nm
Abso
rptio
nsdi
ffere
nz /
mO
D
λ / nm
300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps
Grundzustands- absorption
stimulierte Emission
λanr. = 650 nm
300 fs 600 fs 1,2 ps 2,4 ps 5 ps 10 ps 20 ps 40 ps 80 ps 150 ps 300 ps
0 400 800 1200 16000
1
2
Four
ierk
oeffi
zien
ten
a.u.
Wellenzahl / cm-1
-30-20-10
0-6-30
0 200 400 600 800 1000-4-20
Ab
sorp
tions
diffe
renz
/ m
OD
580 nm
565 nm
Zeit / fs
640 nm


















