
Subsensitivity to β-adrenoceptor agonists in right atria isolated from footshock-stressed rats
5
Gen. Pharmac. Vol. 18, No. 5, pp. 473-477, 1987 0306-3623/87 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1987 Pergamon Journals Ltd SUBSENSITIVITY TO fl-ADRENOCEPTOR AGONISTS IN RIGHT ATRIA ISOLATED FROM FOOTSHOCK-STRESSED RATS ROSANA ALMADA BASSANIand SERGIODE MORAES* Department of Pharmacology, Faculty of Medical Sciences, Universidade Estadual de Campinas (UNICAMP), Campinas, S~o Paulo, Brazil (Received 24 November 1986) Abstract--1. The effects of three daily sessions of inescapable footshock on the sensitivity of rat isolated right atria to the chronotropic effect of norepinephrine, tyramine and soterenol were studied. 2. Inescapable footshock induces subsensitivity to norepinephrine and tyramine. The maximum response to the partial agonist soterenol was reduced. 3. In vitro denervation and addition of cocaine prevented the demonstration of inescapable footshock- induced subsentivity to norepinephrine. 4. It is concluded that repeated inescapable footshock stress reduces the number of atrial fll-adrenoceptors and increases the efficiency of the neuronal reuptake process. INTRODUCTION It is generally accepted that repeated stressful situ- ations cause the development of a persistent sub- sensitivity of rat brain tissue to norepinephrine (Stone and McCarty, 1983). The mechanisms by which stressor agents produce subsensitivity have not been ascertained yet but it is likely that a common factor is the great stress-induced release of catecholamines (Wolfe et al., 1978). Chronic footshock decreases the sensitivity of rat brain slices norepinephrine-cyclic AMP generating system (Stone, 1978, 1979a,b). Con- sidering that fl-adrenoceptors partially mediate this norepinephfine effect, a reduction of density and/or affinity of fl-adrenoceptors is a possible mechanism for subsensitivity of rat brain slices seen after re- peated stress. Accordingly, several reports have shown decreases in the density of rat brain fl-adrenoceptors after repeated stress (U'Prichard and Kvetnansk~, 1980; Torda et al., 1981; Stone, 1983; Stone and Platt, 1982). Contrasting with brain slice studies few attempts have been made to study the sensitivity to norepinephrine in fl-adrenoceptors containing tissues isolated from stressed rats. Harriet al. (1974) observed that right atria isolated from cold-stressed rats showed subsensitivity to the chronotropic effect of norepinephrine. Recently, it has been reported that cold stress induces a con- formational alteration of atrial fl~-adrenoceptor population which is responsible for the observed atrial subsensitivity to the neurotransmitter (CaUia and De Moraes, 1984). Taken together, these studies suggest that fl-adrenoceptor subsensitivity is a mechanism related to adaptation to repeated stress (Stone, 1979b) both in brain and peripheral tissues. The present experiments were undertaken in order to investigate this question by studying the effects of *To whom all correspondence should be addressed. another type of stress (inescapable footshock) on the sensitivity of the rat isolated right atrium to fl-adrenoceptor agonists. Although footshock and cold exposure induce similar physiological and endocrinological stress reactions (Kvetnansk~ and Mikulaj, 1970) and predictably both should induce subsensitivity to norepinephrine, two features differentiate footshock from cold exposure. Foot- shock is an intermittent repeated stress and secondly, unlike some stressogenic conditions, footshock has a sensory stimulation component which induces pain and strong emotional distress. MATERIALSAND METHODS Animals Male Wistar rats weighing 200-300 g at the beginning of the experiments were used. The animals were housed four per hanging solid-bottom cage, with pine shavings as bedding and with tap water and laboratory chow freely available. Streys protocol Rats were individuallysubmitted to three daily sessionsof unsignalled randomized inescapable footshock. Footshock stress was applied by placing the animal into a Plexiglass chamber (26cm long by 21 cm wide by 26cm high) with a grid floor consisting of stainless steel rods, 0.3 cm dia and spaced 1.0 cm. Shocks were delivered by a constant-current d.c. source controlled by a microprocessor and were scrambled according to Hoffman and Fleshler (1967). Each session lasted 30 min. Current intensity was 1.0 mA and duration 1.0scc. Each rat received 120 footshocks at random intervals of 5-25 see with a mean interval of 15 sec. After the ending of the sessionwhich was always carried out between 10:00 and 11:00 a.m., the animals were returned to their home-cage. Isolated pacemaker Immediately after the ending of the third daily footshock sessions rats were killed by a sharp blow on the head and bleeding. Right atria were isolated, trimmed of excess tissue 473
Transcript of Subsensitivity to β-adrenoceptor agonists in right atria isolated from footshock-stressed rats

Gen. Pharmac. Vol. 18, No. 5, pp. 473-477, 1987 0306-3623/87 $3.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1987 Pergamon Journals Ltd
SUBSENSITIVITY TO fl-ADRENOCEPTOR AGONISTS IN RIGHT ATRIA ISOLATED FROM
FOOTSHOCK-STRESSED RATS
ROSANA ALMADA BASSANI and SERGIO DE MORAES* Department of Pharmacology, Faculty of Medical Sciences,
Universidade Estadual de Campinas (UNICAMP), Campinas, S~o Paulo, Brazil
(Received 24 November 1986)
Abstract--1. The effects of three daily sessions of inescapable footshock on the sensitivity of rat isolated right atria to the chronotropic effect of norepinephrine, tyramine and soterenol were studied.
2. Inescapable footshock induces subsensitivity to norepinephrine and tyramine. The maximum response to the partial agonist soterenol was reduced.
3. In vitro denervation and addition of cocaine prevented the demonstration of inescapable footshock- induced subsentivity to norepinephrine.
4. It is concluded that repeated inescapable footshock stress reduces the number of atrial fll-adrenoceptors and increases the efficiency of the neuronal reuptake process.
INTRODUCTION
It is generally accepted that repeated stressful situ- ations cause the development of a persistent sub- sensitivity of rat brain tissue to norepinephrine (Stone and McCarty, 1983). The mechanisms by which stressor agents produce subsensitivity have not been ascertained yet but it is likely that a common factor is the great stress-induced release of catecholamines (Wolfe et al., 1978). Chronic footshock decreases the sensitivity of rat brain slices norepinephrine-cyclic AMP generating system (Stone, 1978, 1979a,b). Con- sidering that fl-adrenoceptors partially mediate this norepinephfine effect, a reduction of density and/or affinity of fl-adrenoceptors is a possible mechanism for subsensitivity of rat brain slices seen after re- peated stress. Accordingly, several reports have shown decreases in the density of rat brain fl-adrenoceptors after repeated stress (U'Prichard and Kvetnansk~, 1980; Torda et al., 1981; Stone, 1983; Stone and Platt, 1982). Contrasting with brain slice studies few attempts have been made to study the sensitivity to norepinephrine in fl-adrenoceptors containing tissues isolated from stressed rats. Harriet al. (1974) observed that right atria isolated from cold-stressed rats showed subsensitivity to the chronotropic effect of norepinephrine. Recently, it has been reported that cold stress induces a con- formational alteration of atrial fl~-adrenoceptor population which is responsible for the observed atrial subsensitivity to the neurotransmitter (CaUia and De Moraes, 1984). Taken together, these studies suggest that fl-adrenoceptor subsensitivity is a mechanism related to adaptation to repeated stress (Stone, 1979b) both in brain and peripheral tissues. The present experiments were undertaken in order to investigate this question by studying the effects of
*To whom all correspondence should be addressed.
another type of stress (inescapable footshock) on the sensitivity of the rat isolated right atrium to fl-adrenoceptor agonists. Although footshock and cold exposure induce similar physiological and endocrinological stress reactions (Kvetnansk~ and Mikulaj, 1970) and predictably both should induce subsensitivity to norepinephrine, two features differentiate footshock from cold exposure. Foot- shock is an intermittent repeated stress and secondly, unlike some stressogenic conditions, footshock has a sensory stimulation component which induces pain and strong emotional distress.
MATERIALS AND METHODS
Animals Male Wistar rats weighing 200-300 g at the beginning of
the experiments were used. The animals were housed four per hanging solid-bottom cage, with pine shavings as bedding and with tap water and laboratory chow freely available.
Streys protocol
Rats were individually submitted to three daily sessions of unsignalled randomized inescapable footshock. Footshock stress was applied by placing the animal into a Plexiglass chamber (26cm long by 21 cm wide by 26cm high) with a grid floor consisting of stainless steel rods, 0.3 cm dia and spaced 1.0 cm. Shocks were delivered by a constant-current d.c. source controlled by a microprocessor and were scrambled according to Hoffman and Fleshler (1967). Each session lasted 30 min. Current intensity was 1.0 mA and duration 1.0scc. Each rat received 120 footshocks at random intervals of 5-25 see with a mean interval of 15 sec. After the ending of the session which was always carried out between 10:00 and 11:00 a.m., the animals were returned to their home-cage.
Isolated pacemaker
Immediately after the ending of the third daily footshock sessions rats were killed by a sharp blow on the head and bleeding. Right atria were isolated, trimmed of excess tissue
473

474 ROSANA ALMADA BASSANI
and set up for isometric recording of spontaneous beating in 20 ml tissue baths. The bathing medium was Krebs- Henseleit solution with the following mM concentrations: NaCI, 115.0; KC1, 4.6; CaCI2.2H20, 2.5; MgSO4.7H20 2.5; KH2PO4, 1.2; NaHCO3, 25.0; glucose I 1.0 and ascorbic acid 0.1. The bathing medium was kept at 36.5°C and continuously gassed with 95% 02-5% CO 2 (pH 7.2-7.4). Diastolic tension applied to each atrium was the maximum needed to obtain a pen deflection of about 0.5 cm/beat at the highest preamplifier sensitivity without recording background noise (Biickner et al., 1978). Bathing medium was changed every 15 min until the preparation reached a stable basal frequency. This took about 45min. Full cumulative concentration-effect curves to norepinephrine, tyramine or soterenol were obtained by a stepwise increase in the agonist concentration (0.5 log unit). Maximum response was considered to have been obtained when a 0.5 log unit increase in the agonist concentration did not induce an additional response. Only one concentration-effect curve for the chronotropic effect of the agonists was obtained from each atrium. Changes in sensitivity to the agonists were evaluated by determining the concentration which produces a response which was 50% of the maximum response (ECs0). The mean negative logarithm of ECs0 (POE) and SEM were calculated from values obtained in individual experiments.
In vitro pretreatment
To reduce the influence of neuronal dissipating mech- anisms on the sensitivity of the isolated pacemaker to norepinephrine, spontaneously beating atria were in vitro denervated using 6-hydroxydopamine (6-OHDA) according to Aprigliano and Hermsmeyer (1976). Briefly, NaHCO 3 and KH2PO 4 were omitted from the bathing medium and the antioxidant agent glutathione (20#M) was added. 6-OHDA in the concentration of 12 #M was applied for a 10-min period and then thoroughly washed out during 45min at 15-min intervals. Previous experiments have shown that this procedure causes a 90% reduction in the atrial norepinephrine endogenous content [Callia and De Moraes 0984)]. After 6-OHDA pretreatment atria were exposed to 3/zM cocaine. The neuronal uptake inhibitor remained in the tissue baths during determination of the norepinephrine concentration-effect curve.
Statistical analyses
Data were analysed using the Student's t-test for unpaired samples (Snedecor and Cochran, 1967).
RESULTS
During the three daily sessions of footshock the animals showed unequivocal signs of intense emo- tional distress such as uneasiness, piloerection and
and SERGIO DE MORAES
defaecation. Some animals also vocalize during the sessions. Right atria isolated from footshock-stressed rats showed resting rates which did not differ from those observed in right atria isolated from the control group. However, the concentration--effect curve for the chronotropic effect of norepinephrine was shifted to the right (2.34-fold at the pD 2 level; P < 0.05) without a significant alteration in the maximum response to the neurotransmitter. In vitro denervation using 6 -OHDA and addition of cocaine shift to the left the concentration-effect curves to norepinephrine in right atria isolated from the control group (6.60-fold) and from the stressed group (16.59-fold). As can be seen in Table 1, the effect of impairment of the neuronal uptake was significantly different (P < 0.05) in atria from stressed rats as compared to atria from the control group. Figure 1 also illustrates that 6 -OHDA and cocaine prevented the demonstra- tion of footshock-induced atrial subsensitivity to the chronotropic effect of norepinephrine.
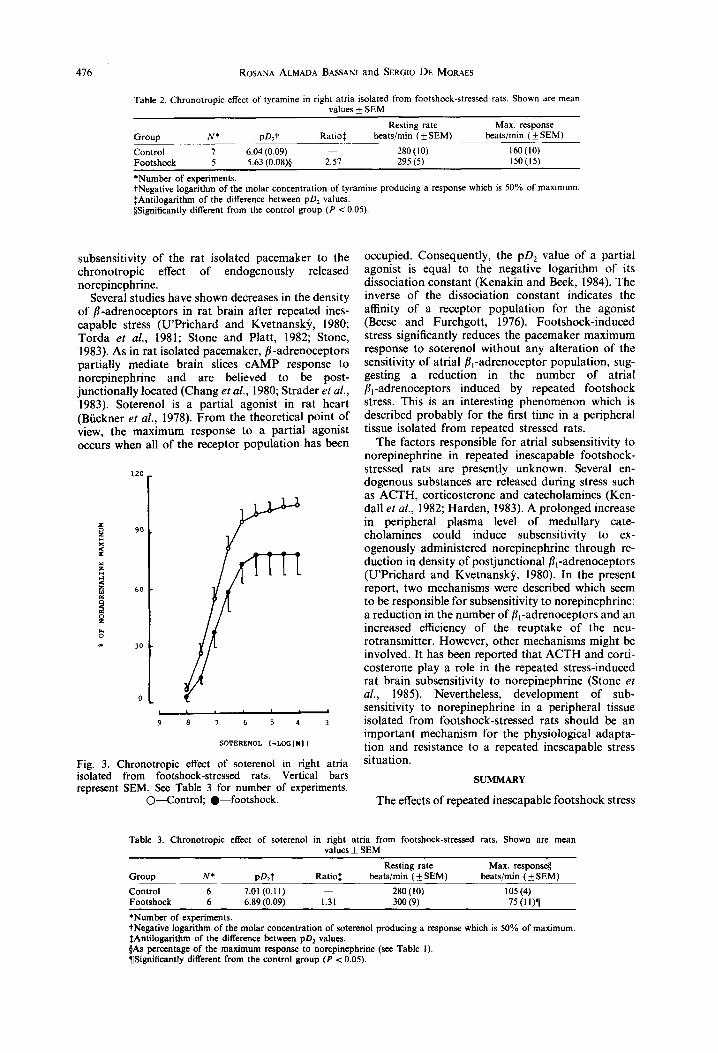
Right atria isolated from footshock-stressed rats also were subsensitive to the chronotropic effect of tyramine, an indirectly-acting fl~-adrenoceptor agonist (Fig. 2). Interestingly, the rightward displacement of the concentration-effect curve to tyramine has the same magnitude as that observed for norepinephrine (2.57-fold at the pD2 level). The maximum response to tyramine remains unaltered (see Table 2). Concentrat ion-effect curves to t h e partial fll-adrenoceptor agonist soterenol are shown in Fig. 3. It can be seen that repeated footshock stress induces a statistically significant diminution of the atrial maximum response to soterenol, when the concentration-effect curve is plotted in relation to the maximum response to norepinephrine (P < 0.05). However, the atrial sensitivity to soterenol was unaffected by repeated footshock (Table 3).
DISCUSSION
The development of rat brain subsensitivity to adrenoceptor agonists is thought to be related to the general mechanisms of adaptat ion to chronic stress (Stone, 1979b). Moreover , it has been suggested that the effect of repeated stressful situations on brain noradrenergic transmission is dependent of subtle psychological variables, such as the inability of the animal to cope with the stressor (Weiss et al., 1977). In the present report, three daily sessions of ines- capable footshock induces atrial subsensitivity to the
Table 1, Chronotropic effect of norepinephrine in rat atria isolated from footshock-stressed rats. Shown are mean values + SEM
Resting rate Max. response Group Nf pD2:~ Ratio§ beats/min (_SEM) beats/min (+SEM) Control 6 7.42 (0.06) - - 305 (6) 120 (12) Footshock 6 7.05 (0. l 0)¶l 2.34 305 (2) 150 (8) Control* 5 8,24 (0.08)11 6.60 320 (7) 110 (19) Footshock 13 8.27 (0.07)** 16.60 300 (6) 105 (5)
*In vitro pretreatment with 6-OHDA and cocaine (3 ,aM). fNumber of experiments. :[:Negative logarithm of the molar concentration of norepinephrine producing a response which is 50% of
maximum. §Antilogarithm of the difference between pD 2 values. ¶]Significantly different from the control/control* groups (P < 0.05). [ISignificantly different from control (P < 0.05). **Significantly different from footshock (P < 0.05).

Footshock-stressed rats 475
(n z 0
u)
x
10¢ A
• , | * *
11 i0 9 8 7 6 5 4
N O R E P I N E P H R I N E ( - L O G [ M ] )
ILl U) Z O a . U) W
:E 5O
X
1 0 0 B
0 i I , J . • •
11 10 9 S 7 6 5
N O R E P I N E P H R I N E - L O G £M~] )
Fig. 1. Chronotropic effect of norepinephrine in right atria isolated from footshock-stressed rats (A). In (B) the experiments were carried out after in vitro denervation using 6-hydroxydopamine and addition of 3 #M cocaine. Vertical bars represent SEM. See Table 1 for number of experiments. O--Control;
O--footshock.
chronotropic effect of norepinephrine, which seems to be caused by an increased efficiency of the. neuronal uptake process and a reduction in the number of atrial fl~-adrenoceptors.
Atrial subsensitivity to the chronotropic effect of norepinephrine could be produced by modifications in postjunctionally located fl]-adrenoceptors and/or alterations in the mechanisms which modulate the concentration of norepinephrine at the flradreno- ceptors. The latter mechanisms are represented by the neuronal reuptake process and by the intraneuronal metabolism of the catecholamine. Pharmacologically, a discreet reduction in the density of postjunctional fll-adrenoceptors would not be expected to shift to
Z o
x
o
I00
50
/ I I I !
7 6 5 4
TYRAMINE (-LOG[M])
Fig. 2. Chronotropic effect of tyramine in right atria isolated from footshock-stressed rats. Vertical bars represent SEM. See Table 2 for number of experiments. O--Control;
l--footshock.
the right the concentration-effect curve to norepine- phrine since, probably, there is a large amount of spare receptors for the neurotransmitter. Biickner et al. 0978) showed that the fractional occupation of rat atrial fl-adrenoceptors, needed to produce 50% of the maximum response (PD2) to the chronotropic effect of isoprenaline corresponds to fewer than 5% of the fl-adrenoceptor population. On the other hand, an increase in neuronal reuptake process and/or intraneuronal metabolism of norepinephrine should shift the curve to the right without any significant alteration in the maximum response to the neurotransmitter.
In the present report, three daily sessions of ines- capable footshock induce a parallel shift to the right of the concentration-effect curve to norepinephrine. Addition of cocaine, which selectively inhibits the neuronal reuptake process, prevented the demonstra- tion of atrial subsensitivity to the neurotransmitter. This is a strong, although indirect, evidence that inescapable footshock-induced stress increases the activity of the neuronal reuptake process. In order to investigate the effect of footshock stress on the atrial sensitivity to endogenously released norepinephrine, the indirectly-acting sympathomimetic agonist tyr- amine was used. There was subsensitivity to the chronotropic effect of tyramine. The norepinephrine- releasing effect of tyramine is calcium-independent and non-exocytotic (B6nisch et al., 1986). Tyramine mobilizes norepinephrine from storage vesicles and is transported together with Na ÷, which increases the affinity of free axoplasmic norepinephrine to neuronal membrane carriers, thus amplifying the outward transport of the neurotransmitter (B6nisch et al., 1986). Hence, the main neuronal mechanism for dissipation of tyramine-released norepinephrine is the reuptake of the neurotransmitter. Increased atrial reuptake of the neurotransmitter will induce sub- sensitivity to the chronotropic effect of tyramine. Taken together, these results clearly indicate, for the first time, that repeated inescapable footshock causes
O . P . 1 8 / ~ ' m B
476 ROSANA ALMADA BASSANI and SERGIO DE MORAES
Table 2. Chronotropic effect of tyramine in right atria isolated from footshock-stressed rats. Shown are mean values _+ SEM
Resting rate Max. response Group N* pD2t Ratio~: beats/min ( _+ S E M ) beats/min ( + SEM)
Control 7 6.04(0.09) - - 280(10) 160(10) Footshock 5 5.63 (0.08)§ 2.57 295 (5) 150 (I 5)
*Number of experiments. tNegative logarithm of the molar concentration of tyramine producing a response which is 50% of maximum. :~Antilogarithm of the difference between pD 2 values. §Significantly different from the control group (P < 0.05).
subsensitivity of the rat isolated pacemaker to the chronotropic effect of endogenously released norepinephrine.
Several studies have shown decreases in the density of fl-adrenoceptors in rat brain after repeated ines- capable stress (U'Pr ichard and Kvetnansk~, 1980; Torda et al., 1981; Stone and Platt, 1982; Stone, 1983). As in rat isolated pacemaker, fl-adrenoceptors partially mediate brain slices c A M P response to norepinephrine and are believed to be post- junctionally located (Chang et al., 1980; Strader et al., 1983). Soterenol is a partial agonist in rat heart (Biickner et al., 1978). F rom the theoretical point of view, the maximum response to a partial agonist occurs when all o f the receptor populat ion has been
120
9o
op 3 0
0
I I I I I I I
9 8 7 6 5 4 3
SOTERENOL (-LOGIM])
Fig. 3. Chronotropic effect of soterenol in right atria isolated from footshock-stressed rats. Vertical bars represent SEM. See Table 3 for number of experiments.
O--Control; O--footshock.
occupied. Consequently, the pD 2 value of a partial agonist is equal to the negative logarithm of its dissociation constant (Kenakin and Beck, 1984). The inverse of the dissociation constant indicates the affinity of a receptor populat ion for the agonist (Beese and Furchgott , 1976). Footshock-induced stress significantly reduces the pacemaker maximum response to soterenol without any alteration of the sensitivity of atrial fl ,-adrenoceptor population, sug- gesting a reduction in the number of atrial fll-adrenoceptors induced by repeated footshock stress. This is an interesting phenomenon which is described probably for the first time in a peripheral tissue isolated from repeated stressed rats.
The factors responsible for atrial subsensitivity to norepinephrine in repeated inescapable footshock- stressed rats are presently unknown. Several en- dogenous substances are released during stress such as A C T H , corticosterone and catecholamines (Ken- dall et al., 1982; Harden, 1983). A prolonged increase in peripheral plasma level of medullary cate- cholamines could induce subsensitivity to ex- ogenously administered norepinephrine through re- duction in density of postjunctional fll-adrenoceptors (U'Prichard and Kvetnansk~, 1980). In the present report, two mechanisms were described which seem to be responsible for subsensitivity to norepinephrine: a reduction in the number of fl~-adrenoceptors and an increased efficiency of the reuptake of the neu- rotransmitter. However, other mechanisms might be involved. It has been reported that A C T H and corti- costerone play a role in the repeated stress-induced rat brain subsensitivity to norepinephrine (Stone et al., 1985). Nevertheless, development of sub- sensitivity to norepinephrine in a peripheral tissue isolated from footshock-stressed rats should be an important mechanism for the physiological adapta- tion and resistance to a repeated inescapable stress situation.
SUMMARY
The effects of repeated inescapable footshock stress
Table 3. Chronotropic effect of soterenol in right atria from footshock-stressed rats. Shown are mean values ± SEM
Resting rate Max. response§ Group N* pD2t Ratio:~ beats/rain (±SEM) beats/rain (±SEM) Control 6 7.01 (0.11 ) - - 280 (10) 105 (4) Footshock 6 6.89 (0.09) 1.31 300 (9) 75 (1 l)¶ *Number of experiments. tNegative logarithm of the molar concentration of soterenol producing a response which is 50% of maximum. :[:Antilogarithm of the difference between pD 2 values. §As percentage of the maximum response to norepinephrine (see Table I). ¶Significantly different from the control group (P < 0.05).

Footshock-stressed rats 477
on the sensitivity of the rat isolated right atria to the chronotropic effect of norepinephrine, tyramine and soterenol were studied. Inescapable footshock shifts to the right the concentration-effect curve to nor- epinephrine (2.34-fold at the pD 2 level; P < 0.05) and tyramine (2.57-fold at the pD 2 level, P <0.05). Footshock stress reduces the atrial maximum re- sponse to the partial agonist soterenol when its concentration-effect curve is plotted in relation to the maximum response to norepinephrine (P < 0.05). In vitro denervation and addition of cocaine prevented the demonstrat ion of subsensitivity to nor- epinephrine. It is concluded that, as observed for rat brain slices, repeated inescapable footshock stress decreases the number of atrial postjunctional f l :adrenoceptors . However, subsensitivity to ex- ogenous or endogenously-released norepinephrine is caused by an increased efficiency of the neuronal reuptake process.
Acknowledgements--The authors wish to thank Mrs Solange Ap. dos Santos Basso for typing the manuscript. R.A.B. is the recipient of a pre-doctoral fellowship from FAPESP (Grant No. 84/2018-0).
REFERENCES
Aprighano O. and Hermsmeyer K. (1976) In vitro de- nervation of the portal vein and caudal artery of the rat. J. Pharmac. exp. Ther. 198, 568-577.
Besse J. C. and Furchgott R. F. (1976) Dissociation con- stants and relative efficacies of agonists acting on alpha- adrenergic receptors in rabbit aorta. J. Pharmac. exp. Ther. 197, 66-78.
B6nisch H., Fuchs G. and Graefe K. H. (1986) Sodium- dependence of the saturability of carrier-mediated nor- adrenaline effiux from noradrenergic neurones in the rat vas deferens. Naunyn-Schmiedebergs Arch. Pharmac. 332, 131-134.
Bfickner C. K., Torphy T. and Costa D. J. (1978) Studies on fl-adrenoceptors mediating changes in mechanical events and adenosine 3'-5' monophosphate levels in rat atria. Fur. J. Pharmac. 47, 259-271.
Callia M. L. and De Moraes S. (1984) Heterogeneity of beta adrenoceptors in right atria isolated from cold-exposed rats. J. Pharmac. exp. Ther. 230, 450-454.
Chang R. L. S., Tran V. T. and Snyder S. H. (1980) Neurotransmitter receptor localization: brain lesion- induced alteration in benzodiazepine, GABA, beta- adrenergic and histamine H:receptor binding. Brain Res. 190, 95-110.
Harden T. K. (1983) Agonist-induced desensitization of /~-adrenergic receptor-linked adenylate cyclase. Pharmac. Rev. 35, 5-32.
Hard M. N. E., Melender L. and Tirri R. (1974) Changed chronotropic sensitivity to sympathomimetic amines in isolated atria from rats following cold acclimation. Ex- perientia 30, 1041-1043.
Hoffman H. S. and Fleshler M. (1967) A relay sequencing
device for scrambling grid shock. J. exp. Anim. Behav. 5, 329-330.
Kenakin T. P. and Beek D. (1984) Relative efficacy of prenalterol and pirbuterol: measurement of agonist affinity by alteration of receptor number. J. Pharmac. exp. Ther. 299, 340-345.
Kendall D. A., Duman R., Slopes J. and Enna S. J. (1982) Influence of adrenocorticotropic hormone and yohimbine on antidepressant-induced declines in rat brain neuro- transmitter receptor binding and function. J. Pharmac. exp. Ther. 222, 566-571.
Kvetnanak~, R. and Mikulaj L. (1970) Adrenal and urinary catecholamines in rats during adaptation to repeated immobilization stress. Endocrinology 87, 738-743.
Snedecor G. W. and Cochran W. G. (1967) Statistical Methods. Iowa State University Press, Ames.
Stone E. A. (1978) Effect of stress on norepinephrine- stimulated accumulation of cyclic AMP in rat brain slices. Pharmac. Biochem. Behav. 8, 583-591.
Stone E. A. (1979a) Reduction by stress of norepinephrine- stimulated accumulation of cyclic AMP in rat cerebral cortex. J. Neurochem. 32, 1335-1337.
Stone E. A. (1979b) Subsensitivity to norepinephrine as a link between adaptation to stress and antidepressant therapy: an hypothesis. Res. Commun. Psychol. Psychiat. Behav. 4, 241-255.
Stone E. A. (1983) Adaptation to stress and brain adre- nergic receptors. Neurosci. Biobehav. Rev. 7, 503-509.
Stone E. A. and McCarty R. (1983) Adaptation to stress: tyrosine hydroxylase activity and catecholamine release. Neurosci. Biobehav. Rev. 7, 29-34.
Stone E. A. and Platt J. E. (1982) Brain adrenergic receptors and resistance to stress. Brain Res. 237, 405-514.
Stone E. A., Slucky A. V., Platt J. E. and Trullas R. (1985) Reduction of the cyclic adenosine 3'-5' monophosphate response to catecholamines in rat brain slices after re- peated restraint stress. J. Pharmac. exp. Ther. 233, 382-388.
Strader C. D., Pickel V. M., Joh T. H., Strohsacker M. W., Shore R., Lefkowitz R. F. and Caron M. G. (1983) Antibodies to the fl-adrenergic receptor: attenuation of catecholamine-sensitive adenylate cyclase and demonstra- tion of postsynaptic receptor localization in brain. Proc. natn. Acad. Sci. U.S.A. 80, 1840-1844.
Torda T., Yamaguchi I., Hirata F., Kopin I. J. and Axelrod J. (1981) Mepacrine treatment prevents immobilization- induced desensitization of beta-adrenergic receptors in rat hypothalamus and brain stem. Brain Res. 205, 441-444.
U'Prichard D. C. and Kvetnansk~, R. (1980) Central and peripheral adrenergic receptor in acute and repeated immobilization stress. In Catecholamines and Stress: Re- cent Advances (Edited by Usden E., Kvetnansk~, R. and Kopin I. J.), pp. 299-232. Elsevier North/Holland, New York.
Weiss J. M., Glazer H. I., Pohrecky L., Bailey W. H. and Schneider L. H. (1977) Coping behaviour and stress- induced behavioral depression: studies of the role of brain catecholamines. In The Psychobiology of Depressive Disorders: Implications for the Effects of Stress (Edited by Depul R.). Academic Press, New York.
Wolfe B. B., Harden T. K., Sporn J. R. and Molinoff P. B. • (1978) Presynaptic modulation of beta adrenergic recep-
tors in rat cerebral cortex after treatment with anti- depressants. J. Pharmac. exp. Ther. 207, 446~,57.


















