
Sodium Valproate Sensitizes Non-Small Lung Cancer A549 Cells to γδ T-Cell-Mediated Killing through...
7
J BIOCHEM MOLECULAR TOXICOLOGY Volume 27, Number 11, 2013 Sodium Valproate Sensitizes Non-Small Lung Cancer A549 Cells to γδ T-Cell-Mediated Killing through Upregulating the Expression of MICA Xian-zhi Du, 1 Qiong-ya Li, 1 Fa-wang Du, 2 Zheng-guang He, 2 and Juan Wang 1 1 Department of Respiratory Medicine, The Second Affiliated Hospital, Chongqing Medical University, Chongqing 400010, People’s Republic of China; E-mail: [email protected] 2 Department of Respiratory Medicine, Suining Central Hospital, Suining 629000, People’s Republic of China Received 25 June 2013; revised 25 June 2013; accepted 12 July 2013 ABSTRACT: Major histocompatibility complex (MHC) class I chain-related protein A (MICA) is in- volved in γδ T-cell recognition of target tumor cells. The aim of this study was to investigate the feasi- bility of utilization of sodium valproate (VPA), a hi- stone deacetylase inhibitor, to sensitize non-small cell lung cancer A549 cells to γδ T-cell-mediated killing. VPA induced a dose-dependent increase in the mRNA and protein expression of MICA in A549 cells. γδ T cells showed cytotoxicity to A549 cells, which was increased by about 50% in the presence of VPA. The concomitant addition of MICA antibody significantly attenuated the VPA-mediated sensitization to γδ T- cell killing. VPA enhanced the cleavage of caspase-3 and caspase-9 in A549 cells cocultured with γδ T cells, and such enhancement was reversed by the MICA an- tibody. In conclusion, VPA sensitizes tumor cells to γδ T-cell-mediated cytotoxicity through the upregulation of MICA and may thus have benefits in improving γδ T-cell-based cancer immunotherapy. C 2013 Wiley Pe- riodicals, Inc. J Biochem Mol Toxicol 27:492–498, 2013; View this article online at wileyonlinelibrary.com. DOI 10.1002/jbt.21513 KEYWORDS: Apoptosis; MICA; Non-Small Lung Cancer; Sodium Valproate; Immunotherapy INTRODUCTION T lymphocytes bearing γδ T-cell receptor (TCR) typically account for 1% to 5% of the total T cells in Correspondence to: Xian-zhi Du. Contract Grant Sponsor: National Natural Science Foundation of China. Contract Grant Number: 30872261. C 2013 Wiley Periodicals, Inc. the peripheral blood, with a predominant phenotype of CD3 + CD4 − CD8 − [1, 2]. The majority of γδ T cells in human peripheral blood are the Vγ 9Vδ2 subset that expresses the Vδ2 chain associated with Vγ 9. In con- trast to conventional αβ T cells, Vγ 9Vδ2 T cells do not recognize antigenic peptides presented by MHC class I or class II molecules, but rather respond to nonpro- cessed and nonpeptidic phosphoantigens in a major histocompatibility complex (MHC)-unrestricted fash- ion [3, 4]. Vγ 9Vδ2 T cells can be stimulated in a va- riety of infectious diseases and play a critical role in the immune defense against invaded pathogens [5]. Interestingly, γδ T cells exhibit the capacity to kill a broad range of solid tumor and leukemia/lymphoma cells [6, 7]. It has been documented that activated hu- man γδ T cells produce large amounts of cytokines, including interferon-γ , consequently inducing antitu- mor immune responses [8]. Natural killer (NK) group 2, member D (NKG2D) is a C-type lectin-like recep- tor expressed on NK cells and γδ T cells [9]. This receptor plays an important role in cancer immuno- surveillance. The ligands for NKG2D include the MHC class I chain-related molecules A and B (MICA and MICB), which are primarily expressed by tumor ep- ithelial cells [10, 11]. The MIC–NKG2D engagement is believed to activate cytolytic responses of γδ T cells against epithelial tumor cells [9, 12]. Release of MICA and MICB from cancer cells represents an immune- escape mechanism and contributes to tumor progres- sion [13, 14]. Therefore, stimulation of the expression and prevention of the shedding of MIC molecules is of importance in triggering effective antitumor immunity. Histone acetylation-mediated chromatin remod- eling is an important mechanism for transcriptional control of genes [15], and is involved in the down- regulation of important components of the “tumor 492
Transcript of Sodium Valproate Sensitizes Non-Small Lung Cancer A549 Cells to γδ T-Cell-Mediated Killing through...

J BIOCHEM MOLECULAR TOXICOLOGYVolume 27, Number 11, 2013
Sodium Valproate Sensitizes Non-Small Lung CancerA549 Cells to γ δ T-Cell-Mediated Killing throughUpregulating the Expression of MICAXian-zhi Du,1 Qiong-ya Li,1 Fa-wang Du,2 Zheng-guang He,2 and Juan Wang1
1 Department of Respiratory Medicine, The Second Affiliated Hospital, Chongqing Medical University, Chongqing 400010, People’s Republicof China; E-mail: [email protected] Department of Respiratory Medicine, Suining Central Hospital, Suining 629000, People’s Republic of China
Received 25 June 2013; revised 25 June 2013; accepted 12 July 2013
ABSTRACT: Major histocompatibility complex(MHC) class I chain-related protein A (MICA) is in-volved in γ δ T-cell recognition of target tumor cells.The aim of this study was to investigate the feasi-bility of utilization of sodium valproate (VPA), a hi-stone deacetylase inhibitor, to sensitize non-small celllung cancer A549 cells to γ δ T-cell-mediated killing.VPA induced a dose-dependent increase in the mRNAand protein expression of MICA in A549 cells. γ δ
T cells showed cytotoxicity to A549 cells, which wasincreased by about 50% in the presence of VPA. Theconcomitant addition of MICA antibody significantlyattenuated the VPA-mediated sensitization to γ δ T-cell killing. VPA enhanced the cleavage of caspase-3and caspase-9 in A549 cells cocultured with γ δ T cells,and such enhancement was reversed by the MICA an-tibody. In conclusion, VPA sensitizes tumor cells to γ δ
T-cell-mediated cytotoxicity through the upregulationof MICA and may thus have benefits in improving γ δ
T-cell-based cancer immunotherapy. C© 2013 Wiley Pe-riodicals, Inc. J Biochem Mol Toxicol 27:492–498, 2013;View this article online at wileyonlinelibrary.com. DOI10.1002/jbt.21513
KEYWORDS: Apoptosis; MICA; Non-Small Lung Cancer;Sodium Valproate; Immunotherapy
INTRODUCTION
T lymphocytes bearing γ δ T-cell receptor (TCR)typically account for 1% to 5% of the total T cells in
Correspondence to: Xian-zhi Du.Contract Grant Sponsor: National Natural Science Foundation
of China.Contract Grant Number: 30872261.
C© 2013 Wiley Periodicals, Inc.
the peripheral blood, with a predominant phenotypeof CD3+CD4−CD8− [1, 2]. The majority of γ δ T cellsin human peripheral blood are the Vγ 9Vδ2 subset thatexpresses the Vδ2 chain associated with Vγ 9. In con-trast to conventional αβ T cells, Vγ 9Vδ2 T cells do notrecognize antigenic peptides presented by MHC classI or class II molecules, but rather respond to nonpro-cessed and nonpeptidic phosphoantigens in a majorhistocompatibility complex (MHC)-unrestricted fash-ion [3, 4]. Vγ 9Vδ2 T cells can be stimulated in a va-riety of infectious diseases and play a critical role inthe immune defense against invaded pathogens [5].Interestingly, γ δ T cells exhibit the capacity to kill abroad range of solid tumor and leukemia/lymphomacells [6, 7]. It has been documented that activated hu-man γ δ T cells produce large amounts of cytokines,including interferon-γ , consequently inducing antitu-mor immune responses [8]. Natural killer (NK) group2, member D (NKG2D) is a C-type lectin-like recep-tor expressed on NK cells and γ δ T cells [9]. Thisreceptor plays an important role in cancer immuno-surveillance. The ligands for NKG2D include the MHCclass I chain-related molecules A and B (MICA andMICB), which are primarily expressed by tumor ep-ithelial cells [10, 11]. The MIC–NKG2D engagement isbelieved to activate cytolytic responses of γ δ T cellsagainst epithelial tumor cells [9, 12]. Release of MICAand MICB from cancer cells represents an immune-escape mechanism and contributes to tumor progres-sion [13, 14]. Therefore, stimulation of the expressionand prevention of the shedding of MIC moleculesis of importance in triggering effective antitumorimmunity.
Histone acetylation-mediated chromatin remod-eling is an important mechanism for transcriptionalcontrol of genes [15], and is involved in the down-regulation of important components of the “tumor
492

Volume 27, Number 11, 2013 SENSITIZING A549 CELLS TO γ δ T CELLS 493
recognition complex” in various tumor cells [16].The expression of NKG2D ligands has been found to beaffected by chromatin remodeling. Armeanu et al. [17]reported that treatment with the histone deacetylaseinhibitor (HDACi) sodium valproate (VPA) inducestranscription of MICA and MICB in hepatocellular car-cinoma cells, leading to increased susceptibility to NKcell-mediated cytotoxicity. Similarly, VPA is able to sen-sitize human osteosarcoma cells to cytotoxicity of NKcells through the upregulation of the surface expressionof MICA and MICB [18].
Non-small cell lung cancer (NSCLC) is one of theleading causes of cancer-related mortality in the world[19]. Despite significant progress in therapy, the prog-nosis of patients with advanced NSCLC is still poor,with less than 5% long-term survival at 5 years [20].A phase I clinical study has demonstrated that adop-tive γ δ T-cell immunotherapy is safe and beneficialfor patients with recurrent NSCLC [21]. In the presentstudy, we investigated the feasibility of utilization ofVPA to enhance the susceptibility of human NSCLCA549 cells to the cytotoxicity of ex vivo expanded γ δ
T cells.
MATERIALS AND METHODS
Antibodies and Reagents
Antihuman γ δ TCR and anti-NKG2D antibodieswere purchased from eBioscience (San Diego, CA),and anti-CD80, anti-CD83, and anti-HLA-DR werepurchased from Beijing 4A Biotech Company, Ltd.(Beijing, People’s Republic of China). Anti-MICA, anti-cleaved caspase-3, and anti-cleaved caspase-9 antibod-ies were purchased from Abcam (Cambridge, MA).Zoledronic acid, VPA, and Lactate Dehydrogenase(LDH) Assay Kit were purchased from Sigma (St. Louis,MO). Interleulin-2 (IL-2) was purchased from Pepro-tech (Rocky Hill, NJ).
Isolation of Peripheral Blood MononuclearCells (PBMCs)
Peripheral blood samples were harvested fromhealthy donors and isolation of PBMCs was done us-ing Ficoll–Hypaque (Shanghai Institutes for Biologi-cal Sciences, Shanghai, People’s Republic of China)density-gradient centrifugation. Informed consent wasobtained from each subject and the protocol of thisstudy was approved by the Medical Ethics Commit-tee of Chongqing Medical University (Chongqing, Peo-ple’s Republic of China).
Preparation of γ δ T Cells
γ δ T cells were sorted from isolated PBMCsby flow cytometry using fluorescein isothiocyanate(FITC)-conjugated antihuman γ δ TCR antibody. Cellswere cultured in RPMI 1640 medium (Gibco, Invitro-gen, Gaithersburg, MD) supplemented with 10% fetalbovine serum (FBS), streptomycin (100 μg/mL), andpenicillin (100 U/mL). To expand γ δ T cells, PBMCswere plated on plastic culture dishes in RPMI 1640medium supplemented with 10% FBS, zoledronic acid(300 pg/mL), and IL-2 (200 IU/mL). Cells were thencultured for 9–12 days. The purity and phenotypiccharacteristics of γ δ T cells were determined by flowcytometry using antibodies against γ δ TCR, NKG2D,HLA-DR, CD80, and CD83. γ δ T cells were employedas effector cells to kill target A549 cells at differenteffector–target ratios (i.e., 5:1, 10:1, 20:1, and 40:1).
A549 Cells and VPA Treatment
A549 cells were purchased from the AmericanType Culture Collection and maintained in RPMI 1640medium supplemented with 10% FBS, streptomycin(100 μg/mL), and penicillin (100 U/mL) at 37◦C in 5%CO2.
A549 cells were seeded onto six-well plates and ex-posed to different concentrations of VPA ranging from0.25–2.0 mM. After incubation for 24 h, the cells wereharvested and tested for the MICA expression.
Lactate Dehydrogenase Assay
A549 cells were cocultured with γ δ T cells for 24 hin the presence or absence of VPA (1.0 mM) and anti-MICA monoclonal antibody (10 μg/mL). The cytotox-icity of γ δ T cells against A549 cells was assessed bythe LDH Assay Kit according to the manufacturer’sinstructions.
Reverse Transcription Polymerase ChainReaction (RT-PCR)
Total RNA was extracted from A549 cells us-ing TRIzol reagent (TaKaRa Biotechnology CompanyLtd., Dalian, People’s Republic of China) accordingto the manufacturer’s protocol. Complementary DNA(cDNA) was synthesized from total RNA with theFirst-Strand cDNA Synthesis Kit (Roche, Mannheim,Germany). PCR amplification was achieved usingTaq DNA Polymerase (TaKaRa Biotechnology Com-pany Ltd.) at the following conditions: initial de-naturation at 94◦C for 3 min, 35 cycles of denat-uration at 94◦C for 30 s, annealing at 58◦C for
J Biochem Molecular Toxicology DOI 10.1002/jbt
494 DU ET AL. Volume 27, Number 11, 2013
30 s, and elongation at 72◦C for 30–50 s, and fi-nal elongation at 72◦C for 5 min. Human MICAcDNA was amplified with the following primers:forward 5′-TCTCCCAAAACCTGGAGACTAA-3′ andreverse 5′-CCAGCTCAGTGTGATATTCCAG-3′ (prod-uct size of 296 bp). Beta-actin, used as an inter-nal control, was amplified with the primers: forward5′-CTGGGACGACATGGAGAAA-3′ and reverse 5′-AAGGAAGGCTGGAAGAGTGC-3′ (production sizeof 564 bp). PCR products were separated by elec-trophoresis on 1% agarose gels and detected byethidium bromide staining. Densitometric analysis ofRT-PCR products was done using the QuantityOnesoftware (Bio-Rad, Hercules, CA).
Western Blot Analysis
After treatments, A549 cells were lysed in a 10 mMTris buffer (pH 7.4) containing 1% sodium dode-cyl sulfate (SDS) and complete protease inhibitors(Roche). Samples of the lysates were separated bySDS-polyacrylamide gel electrophoresis, transferredto polyvinylidene fluoride membranes, and incu-bated with the primary antibodies against MICA,cleaved caspase-3, and cleaved caspase-9 overnightat 4◦C. After incubation with horseradish peroxidase-conjugated secondary antibodies, blots were visu-alized with enhanced chemiluminescence reagents(Amersham Biosciences, Piscataway, NJ). The in-tensity of each band was measured by densit-ometric analysis using the QuantityOne software(Bio-Rad).
Enzyme-Linked Immunosorbent Assay(ELISA)
The supernatant of the cell culture was harvestedafter 24 h of incubation and tested for the release ofsoluble MICA (sMICA) using a commercial ELISA kit.The ELISA experiment was done according to the man-ufacturer’s recommendations. Optical densities weremeasured at 450 nm using a microplate ELISA reader.
Statistical Analysis
All statistical calculations were carried out usingthe Statistical Product and Service Solutions (SPSS)software, version 9.0 (SPSS, Chicago, IL). Data are ex-pressed as means ± standard deviation (SD). The dif-ference among the means of multiple groups was an-alyzed by one-way analysis of variance followed bythe Tukey’s multiple comparison test. A difference wasdefined as significant at p < 0.05.
RESULTS
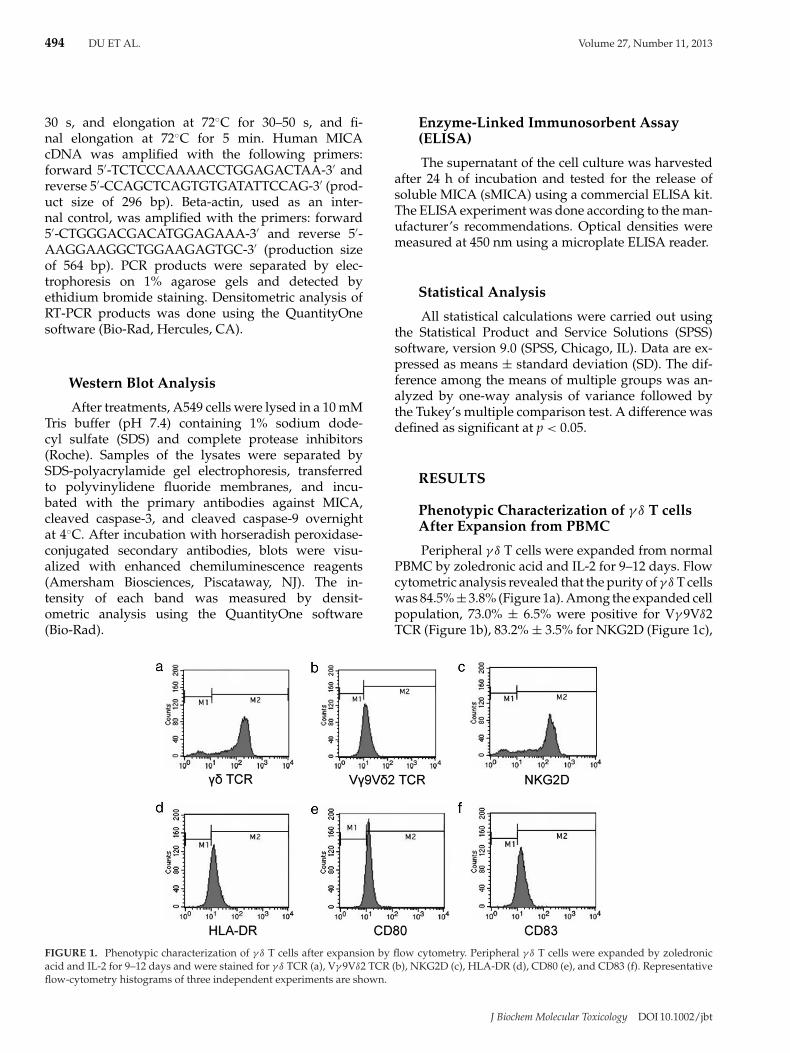
Phenotypic Characterization of γ δ T cellsAfter Expansion from PBMC
Peripheral γ δ T cells were expanded from normalPBMC by zoledronic acid and IL-2 for 9–12 days. Flowcytometric analysis revealed that the purity of γ δ T cellswas 84.5% ± 3.8% (Figure 1a). Among the expanded cellpopulation, 73.0% ± 6.5% were positive for Vγ 9Vδ2TCR (Figure 1b), 83.2% ± 3.5% for NKG2D (Figure 1c),
FIGURE 1. Phenotypic characterization of γ δ T cells after expansion by flow cytometry. Peripheral γ δ T cells were expanded by zoledronicacid and IL-2 for 9–12 days and were stained for γ δ TCR (a), Vγ 9Vδ2 TCR (b), NKG2D (c), HLA-DR (d), CD80 (e), and CD83 (f). Representativeflow-cytometry histograms of three independent experiments are shown.
J Biochem Molecular Toxicology DOI 10.1002/jbt
Volume 27, Number 11, 2013 SENSITIZING A549 CELLS TO γ δ T CELLS 495
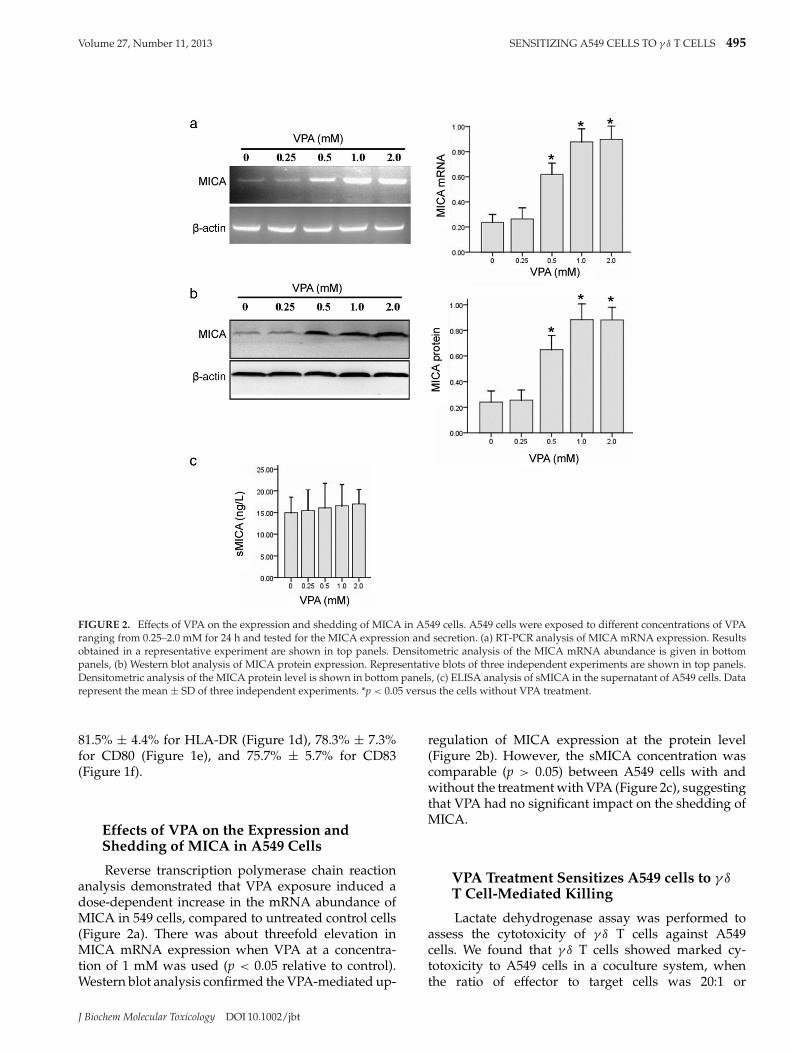
FIGURE 2. Effects of VPA on the expression and shedding of MICA in A549 cells. A549 cells were exposed to different concentrations of VPAranging from 0.25–2.0 mM for 24 h and tested for the MICA expression and secretion. (a) RT-PCR analysis of MICA mRNA expression. Resultsobtained in a representative experiment are shown in top panels. Densitometric analysis of the MICA mRNA abundance is given in bottompanels, (b) Western blot analysis of MICA protein expression. Representative blots of three independent experiments are shown in top panels.Densitometric analysis of the MICA protein level is shown in bottom panels, (c) ELISA analysis of sMICA in the supernatant of A549 cells. Datarepresent the mean ± SD of three independent experiments. *p < 0.05 versus the cells without VPA treatment.
81.5% ± 4.4% for HLA-DR (Figure 1d), 78.3% ± 7.3%for CD80 (Figure 1e), and 75.7% ± 5.7% for CD83(Figure 1f).
Effects of VPA on the Expression andShedding of MICA in A549 Cells
Reverse transcription polymerase chain reactionanalysis demonstrated that VPA exposure induced adose-dependent increase in the mRNA abundance ofMICA in 549 cells, compared to untreated control cells(Figure 2a). There was about threefold elevation inMICA mRNA expression when VPA at a concentra-tion of 1 mM was used (p < 0.05 relative to control).Western blot analysis confirmed the VPA-mediated up-
regulation of MICA expression at the protein level(Figure 2b). However, the sMICA concentration wascomparable (p > 0.05) between A549 cells with andwithout the treatment with VPA (Figure 2c), suggestingthat VPA had no significant impact on the shedding ofMICA.
VPA Treatment Sensitizes A549 cells to γ δT Cell-Mediated Killing
Lactate dehydrogenase assay was performed toassess the cytotoxicity of γ δ T cells against A549cells. We found that γ δ T cells showed marked cy-totoxicity to A549 cells in a coculture system, whenthe ratio of effector to target cells was 20:1 or
J Biochem Molecular Toxicology DOI 10.1002/jbt
496 DU ET AL. Volume 27, Number 11, 2013
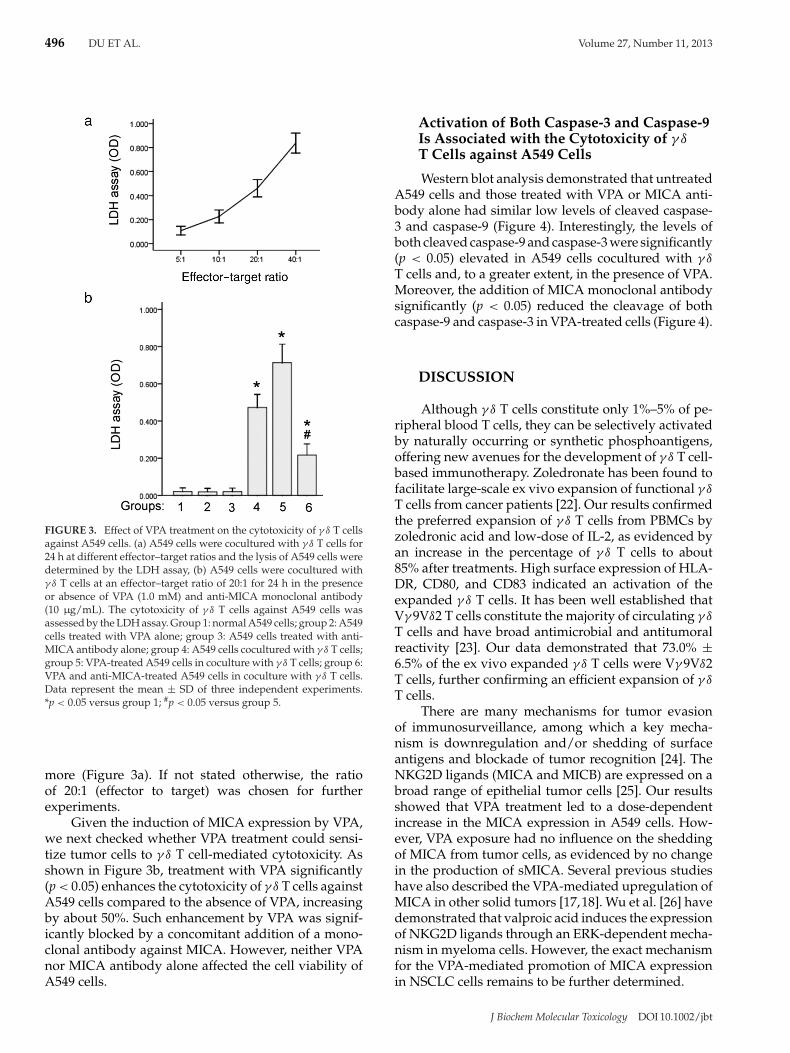
FIGURE 3. Effect of VPA treatment on the cytotoxicity of γ δ T cellsagainst A549 cells. (a) A549 cells were cocultured with γ δ T cells for24 h at different effector–target ratios and the lysis of A549 cells weredetermined by the LDH assay, (b) A549 cells were cocultured withγ δ T cells at an effector–target ratio of 20:1 for 24 h in the presenceor absence of VPA (1.0 mM) and anti-MICA monoclonal antibody(10 μg/mL). The cytotoxicity of γ δ T cells against A549 cells wasassessed by the LDH assay. Group 1: normal A549 cells; group 2: A549cells treated with VPA alone; group 3: A549 cells treated with anti-MICA antibody alone; group 4: A549 cells cocultured with γ δ T cells;group 5: VPA-treated A549 cells in coculture with γ δ T cells; group 6:VPA and anti-MICA-treated A549 cells in coculture with γ δ T cells.Data represent the mean ± SD of three independent experiments.*p < 0.05 versus group 1; #p < 0.05 versus group 5.
more (Figure 3a). If not stated otherwise, the ratioof 20:1 (effector to target) was chosen for furtherexperiments.
Given the induction of MICA expression by VPA,we next checked whether VPA treatment could sensi-tize tumor cells to γ δ T cell-mediated cytotoxicity. Asshown in Figure 3b, treatment with VPA significantly(p < 0.05) enhances the cytotoxicity of γ δ T cells againstA549 cells compared to the absence of VPA, increasingby about 50%. Such enhancement by VPA was signif-icantly blocked by a concomitant addition of a mono-clonal antibody against MICA. However, neither VPAnor MICA antibody alone affected the cell viability ofA549 cells.
Activation of Both Caspase-3 and Caspase-9Is Associated with the Cytotoxicity of γ δT Cells against A549 Cells
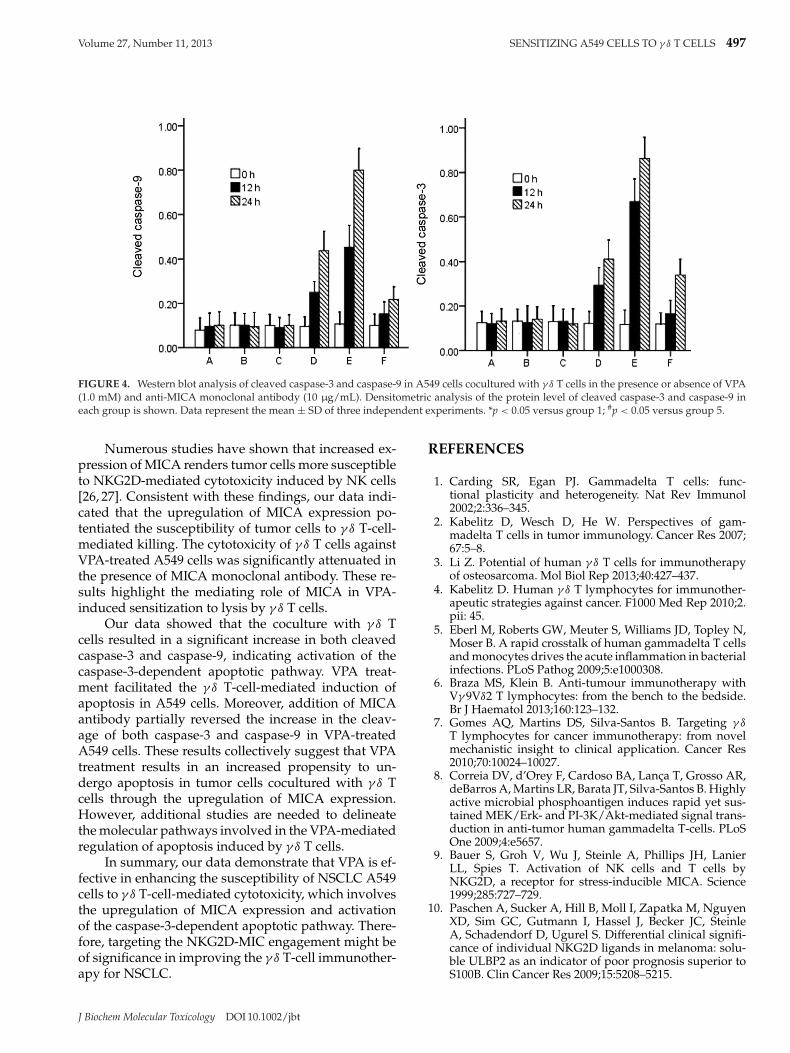
Western blot analysis demonstrated that untreatedA549 cells and those treated with VPA or MICA anti-body alone had similar low levels of cleaved caspase-3 and caspase-9 (Figure 4). Interestingly, the levels ofboth cleaved caspase-9 and caspase-3 were significantly(p < 0.05) elevated in A549 cells cocultured with γ δ
T cells and, to a greater extent, in the presence of VPA.Moreover, the addition of MICA monoclonal antibodysignificantly (p < 0.05) reduced the cleavage of bothcaspase-9 and caspase-3 in VPA-treated cells (Figure 4).
DISCUSSION
Although γ δ T cells constitute only 1%–5% of pe-ripheral blood T cells, they can be selectively activatedby naturally occurring or synthetic phosphoantigens,offering new avenues for the development of γ δ T cell-based immunotherapy. Zoledronate has been found tofacilitate large-scale ex vivo expansion of functional γ δ
T cells from cancer patients [22]. Our results confirmedthe preferred expansion of γ δ T cells from PBMCs byzoledronic acid and low-dose of IL-2, as evidenced byan increase in the percentage of γ δ T cells to about85% after treatments. High surface expression of HLA-DR, CD80, and CD83 indicated an activation of theexpanded γ δ T cells. It has been well established thatVγ 9Vδ2 T cells constitute the majority of circulating γ δ
T cells and have broad antimicrobial and antitumoralreactivity [23]. Our data demonstrated that 73.0% ±6.5% of the ex vivo expanded γ δ T cells were Vγ 9Vδ2T cells, further confirming an efficient expansion of γ δ
T cells.There are many mechanisms for tumor evasion
of immunosurveillance, among which a key mecha-nism is downregulation and/or shedding of surfaceantigens and blockade of tumor recognition [24]. TheNKG2D ligands (MICA and MICB) are expressed on abroad range of epithelial tumor cells [25]. Our resultsshowed that VPA treatment led to a dose-dependentincrease in the MICA expression in A549 cells. How-ever, VPA exposure had no influence on the sheddingof MICA from tumor cells, as evidenced by no changein the production of sMICA. Several previous studieshave also described the VPA-mediated upregulation ofMICA in other solid tumors [17,18]. Wu et al. [26] havedemonstrated that valproic acid induces the expressionof NKG2D ligands through an ERK-dependent mecha-nism in myeloma cells. However, the exact mechanismfor the VPA-mediated promotion of MICA expressionin NSCLC cells remains to be further determined.
J Biochem Molecular Toxicology DOI 10.1002/jbt
Volume 27, Number 11, 2013 SENSITIZING A549 CELLS TO γ δ T CELLS 497
FIGURE 4. Western blot analysis of cleaved caspase-3 and caspase-9 in A549 cells cocultured with γ δ T cells in the presence or absence of VPA(1.0 mM) and anti-MICA monoclonal antibody (10 μg/mL). Densitometric analysis of the protein level of cleaved caspase-3 and caspase-9 ineach group is shown. Data represent the mean ± SD of three independent experiments. *p < 0.05 versus group 1; #p < 0.05 versus group 5.
Numerous studies have shown that increased ex-pression of MICA renders tumor cells more susceptibleto NKG2D-mediated cytotoxicity induced by NK cells[26, 27]. Consistent with these findings, our data indi-cated that the upregulation of MICA expression po-tentiated the susceptibility of tumor cells to γ δ T-cell-mediated killing. The cytotoxicity of γ δ T cells againstVPA-treated A549 cells was significantly attenuated inthe presence of MICA monoclonal antibody. These re-sults highlight the mediating role of MICA in VPA-induced sensitization to lysis by γ δ T cells.
Our data showed that the coculture with γ δ Tcells resulted in a significant increase in both cleavedcaspase-3 and caspase-9, indicating activation of thecaspase-3-dependent apoptotic pathway. VPA treat-ment facilitated the γ δ T-cell-mediated induction ofapoptosis in A549 cells. Moreover, addition of MICAantibody partially reversed the increase in the cleav-age of both caspase-3 and caspase-9 in VPA-treatedA549 cells. These results collectively suggest that VPAtreatment results in an increased propensity to un-dergo apoptosis in tumor cells cocultured with γ δ Tcells through the upregulation of MICA expression.However, additional studies are needed to delineatethe molecular pathways involved in the VPA-mediatedregulation of apoptosis induced by γ δ T cells.
In summary, our data demonstrate that VPA is ef-fective in enhancing the susceptibility of NSCLC A549cells to γ δ T-cell-mediated cytotoxicity, which involvesthe upregulation of MICA expression and activationof the caspase-3-dependent apoptotic pathway. There-fore, targeting the NKG2D-MIC engagement might beof significance in improving the γ δ T-cell immunother-apy for NSCLC.
REFERENCES
1. Carding SR, Egan PJ. Gammadelta T cells: func-tional plasticity and heterogeneity. Nat Rev Immunol2002;2:336–345.
2. Kabelitz D, Wesch D, He W. Perspectives of gam-madelta T cells in tumor immunology. Cancer Res 2007;67:5–8.
3. Li Z. Potential of human γ δ T cells for immunotherapyof osteosarcoma. Mol Biol Rep 2013;40:427–437.
4. Kabelitz D. Human γ δ T lymphocytes for immunother-apeutic strategies against cancer. F1000 Med Rep 2010;2.pii: 45.
5. Eberl M, Roberts GW, Meuter S, Williams JD, Topley N,Moser B. A rapid crosstalk of human gammadelta T cellsand monocytes drives the acute inflammation in bacterialinfections. PLoS Pathog 2009;5:e1000308.
6. Braza MS, Klein B. Anti-tumour immunotherapy withVγ 9Vδ2 T lymphocytes: from the bench to the bedside.Br J Haematol 2013;160:123–132.
7. Gomes AQ, Martins DS, Silva-Santos B. Targeting γ δT lymphocytes for cancer immunotherapy: from novelmechanistic insight to clinical application. Cancer Res2010;70:10024–10027.
8. Correia DV, d’Orey F, Cardoso BA, Lanca T, Grosso AR,deBarros A, Martins LR, Barata JT, Silva-Santos B. Highlyactive microbial phosphoantigen induces rapid yet sus-tained MEK/Erk- and PI-3K/Akt-mediated signal trans-duction in anti-tumor human gammadelta T-cells. PLoSOne 2009;4:e5657.
9. Bauer S, Groh V, Wu J, Steinle A, Phillips JH, LanierLL, Spies T. Activation of NK cells and T cells byNKG2D, a receptor for stress-inducible MICA. Science1999;285:727–729.
10. Paschen A, Sucker A, Hill B, Moll I, Zapatka M, NguyenXD, Sim GC, Gutmann I, Hassel J, Becker JC, SteinleA, Schadendorf D, Ugurel S. Differential clinical signifi-cance of individual NKG2D ligands in melanoma: solu-ble ULBP2 as an indicator of poor prognosis superior toS100B. Clin Cancer Res 2009;15:5208–5215.
J Biochem Molecular Toxicology DOI 10.1002/jbt

498 DU ET AL. Volume 27, Number 11, 2013
11. Unni AM, Bondar T, Medzhitov R. Intrinsic sensor ofoncogenic transformation induces a signal for innateimmunosurveillance. Proc Natl Acad Sci U S A 2008;105:1686–1691.
12. Groh V, Rhinehart R, Secrist H, Bauer S, Grabstein KH,Spies T. Broad tumor-associated expression and recog-nition by tumor-derived gamma delta T cells of MICAand MICB. Proc Natl Acad Sci U S A 1999;96:6879–6884.
13. Holdenrieder S, Stieber P, Peterfi A, Nagel D, Steinle A,Salih HR. Soluble MICA in malignant diseases. Int J Can-cer 2006;118:684–687.
14. Salih HR, Goehlsdorf D, Steinle A. Release of MICBmolecules by tumor cells: mechanism and soluble MICBin sera of cancer patients. Hum Immunol 2006;67:188–195.
15. Kurdistani SK, Tavazoie S, Grunstein M. Mapping globalhistone acetylation patterns to gene expression. Cell2004;117:721–733.
16. Sigalotti L, Coral S, Fratta E, Lamaj E, Danielli R, Di Gi-acomo AM, Altomonte M, Maio M. Epigenetic modu-lation of solid tumors as a novel approach for cancerimmunotherapy. Semin Oncol 2005;32:473–478.
17. Armeanu S, Bitzer M, Lauer UM, Venturelli S, Pathil A,Krusch M, Kaiser S, Jobst J, Smirnow I, Wagner A, SteinleA, Salih HR. Natural killer cell-mediated lysis of hep-atoma cells via specific induction of NKG2D ligands bythe histone deacetylase inhibitor sodium valproate. Can-cer Res 2005;65:6321–6329.
18. Yamanegi K, Yamane J, Kobayashi K, Kato-Kogoe N,Ohyama H, Nakasho K, Yamada N, Hata M, Nish-ioka T, Fukunaga S, Futani H, Okamura H, Terada N.Sodium valproate, a histone deacetylase inhibitor, aug-ments the expression of cell-surface NKG2D ligands,MICA/B, without increasing their soluble forms to en-hance susceptibility of human osteosarcoma cells toNK cell-mediated cytotoxicity. Oncol Rep 2010;24:1621–1627.
19. Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statis-tics, 2002. CA Cancer J Clin 2005;55:74–108.
20. Schiller JH, Harrington D, Belani CP, Langer C, SandlerA, Krook J, Zhu J, Johnson DH; Eastern Cooperative On-cology Group. Comparison of four chemotherapy regi-mens for advanced non-small-cell lung cancer. N Engl JMed 2002;346:92–98.
21. Nakajima J, Murakawa T, Fukami T, Goto S, KanekoT, Yoshida Y, Takamoto S, Kakimi K. A phase I studyof adoptive immunotherapy for recurrent non-small-celllung cancer patients with autologous gammadelta T cells.Eur J Cardiothorac Surg 2010;37:1191–1197.
22. Kondo M, Sakuta K, Noguchi A, Ariyoshi N, Sato K,Sato S, Sato K, Hosoi A, Nakajima J, Yoshida Y, ShiraishiK, Nakagawa K, Kakimi K. Zoledronate facilitates large-scale ex vivo expansion of functional gammadelta T cellsfrom cancer patients for use in adoptive immunotherapy.Cytotherapy 2008;10:842–856.
23. Bonneville M, Scotet E. Human Vgamma9Vdelta2 T cells:promising new leads for immunotherapy of infectionsand tumors. Curr Opin Immunol 2006;18:539–546.
24. Seliger B. Strategies of tumor immune evasion. BioDrugs2005;19:347–354.
25. Groh V, Rhinehart R, Secrist H, Bauer S, Grabstein KH,Spies T. Broad tumor-associated expression and recogni-tion by tumor-derived gamma delta T cells of MICA andMICB. Proc Natl Acad Sci U S A 1999;96:6879–6884.
26. Wu X, Tao Y, Hou J, Meng X, Shi J. Valproic acid up-regulates NKG2D ligand expression through an ERK-dependent mechanism and potentially enhances NKcell-mediated lysis of myeloma. Neoplasia 2012;14:1178–1189.
27. Armeanu S, Bitzer M, Lauer UM, Venturelli S, Pathil A,Krusch M, Kaiser S, Jobst J, Smirnow I, Wagner A, SteinleA, Salih HR. Natural killer cell-mediated lysis of hep-atoma cells via specific induction of NKG2D ligands bythe histone deacetylase inhibitor sodium valproate. Can-cer Res 2005;65:6321–6329.
J Biochem Molecular Toxicology DOI 10.1002/jbt




![· Web view22]. AP-2α expression sensitized cancer cells to chemotherapy drugs and enhanced tumor killing, while AP-2α deletion led to drug resistance 23-25], suggesting the](https://static.fdocument.org/doc/165x107/5f073db07e708231d41c0297/web-view-22-ap-2-expression-sensitized-cancer-cells-to-chemotherapy-drugs-and.jpg)


![Una (mica tanto) breve introduzione a LATEX2 · 2017. 4. 3. · Prefazione LATEX [1] e un sistema di composizione di testo particolarmente indicato per la produzione di documenti](https://static.fdocument.org/doc/165x107/6080c8eb2f68053aeb0439a1/una-mica-tanto-breve-introduzione-a-2017-4-3-prefazione-latex-1-e-un-sistema.jpg)


![PFA(S)[S Spaces arXiv:1104.3471v1 [math.GN] 18 Apr 2011[45], [47], and [46] dealing with characterizing paracompactness and killing Dowker spaces in locally compact normal spaces,](https://static.fdocument.org/doc/165x107/60a0563f2ce08335df0bff54/pfass-spaces-arxiv11043471v1-mathgn-18-apr-2011-45-47-and-46-dealing.jpg)







