
Role for the endometrial epithelial protein MFG-E8 and its receptor integrin αvβ3 in human...
9
Role for the endometrial epithelial protein MFG-E8 and its receptor integrin avb3 in human implantation: results of an in vitro trophoblast attachment study using established human cell lines Carla Schmitz, M.D., a Liang Yu, Ph.D., a Silvina Bocca, M.D., Ph.D., a Sandra Anderson, B.S., a Jo~ ao Sabino Cunha-Filho, M.D., a Bhaskara S. Rhavi, Ph.D., b and Sergio Oehninger, M.D., Ph.D. a a Department of Obstetrics and Gynecology, Eastern Virginia Medical School, The Jones Institute for Reproductive Medicine, Norfolk, Virginia; and b Department of Mathematics and Statistics, Old Dominion University, Norfolk, Virginia Objective: To investigate the role of MFG-E8 and its receptor integrin avb3 in the attachment of trophoblast cells to the endometrial epithelium. Design: Experimental in vitro study. Setting: Academic center. Patient(s): None. Intervention(s): By using a well-differentiated endometrial adenocarcinoma cell line (Ishikawa cells) and choriocarcinoma human trophoblast cells (Jar cells), an in vitro assay mimicking human implantation was established. To investigate the impact of blocking MFG-E8 and integrin avb3, we pretreated the cell lines with antibodies against those proteins at different concentrations before the attachment assay. Main Outcome Measure(s): Attachment rate of Jar spheroids to the epithelial cell monolayer. Result(s): Pretreatment of Ishikawa cells with anti-MFG-E8 antibody caused a dose-dependent and significant inhibition of attachment. On the other hand, pretreatment of Jar spheroids did not result in a significant effect on the attachment rate. Pretreatment of Ishikawa cells as well as Jar spheroids with anti-integrin avb3 antibodies resulted in a dose-dependent, significant inhibition of attachment. Conclusion(s): This study showed that blocking MFG-E8 and its receptor integrin avb3 in Ishikawa cells diminishes Jar spheroid attachment. Moreover, blocking integrin avb3 in the trophoblastic cells also diminished their attachment to the Ishikawa monolayer. (Fertil Steril Ò 2014;101:874–82. Ó2014 by American Society for Reproductive Medicine.) Key Words: Attachment, integrin avb3, implantation, human endometrium, Jar spheroids, MFG-E8 Discuss: You can discuss this article with its authors and with other ASRM members at http:// fertstertforum.com/schmitzc-human-endometrium-mfg-e8-integrin-implantation/ Use your smartphone to scan this QR code and connect to the discussion forum for this article now.* * Download a free QR code scanner by searching for “QR scanner” in your smartphone’s app store or app marketplace. I t is well known that implantation requires a receptive endometrium, a normal and functional embryo at the blastocyst stage of development, and a synchronized dialogue between them (1). Nevertheless, there are many unknown factors that play an important role in this process (2, 3). Because human implantation studies in vivo are very limited (mainly owing to ethical reasons), have poor reproducibility, and Received March 29, 2013; revised November 29, 2013; accepted December 5, 2013; published online January 11, 2014. C.S. has nothing to disclose. L.Y. has nothing to disclose. S.B. has nothing to disclose. S.A. has nothing to disclose. J.S.C.-F. has nothing to disclose. B.S.R. has nothing to disclose. S.O. has nothing to disclose. C.S.'s present address is: Postgraduate Program, Medical Clinics of the Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil. J.S.C.-F.'s present address is: Department of Obstetrics and Gynecology, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil. C.S. and L.Y. contributed equally to this work. Reprint requests: Sergio Oehninger, M.D., Ph.D., The Jones Institute for Reproductive Medicine, 601 Colley Ave., Norfolk, Virginia 23507 (E-mail: [email protected]). Fertility and Sterility® Vol. 101, No. 3, March 2014 0015-0282/$36.00 Copyright ©2014 American Society for Reproductive Medicine, Published by Elsevier Inc. http://dx.doi.org/10.1016/j.fertnstert.2013.12.015 874 VOL. 101 NO. 3 / MARCH 2014
Transcript of Role for the endometrial epithelial protein MFG-E8 and its receptor integrin αvβ3 in human...

Role for the endometrial epithelialprotein MFG-E8 and its receptorintegrin avb3 in human implantation:results of an in vitro trophoblastattachment study using establishedhuman cell lines
Carla Schmitz, M.D.,a Liang Yu, Ph.D.,a Silvina Bocca, M.D., Ph.D.,a Sandra Anderson, B.S.,aJo~ao Sabino Cunha-Filho, M.D.,a Bhaskara S. Rhavi, Ph.D.,b and Sergio Oehninger, M.D., Ph.D.a
a Department of Obstetrics and Gynecology, Eastern Virginia Medical School, The Jones Institute for ReproductiveMedicine, Norfolk, Virginia; and b Department of Mathematics and Statistics, Old Dominion University, Norfolk, Virginia
Objective: To investigate the role of MFG-E8 and its receptor integrin avb3 in the attachment of trophoblast cells to the endometrialepithelium.Design: Experimental in vitro study.Setting: Academic center.Patient(s): None.Intervention(s): By using a well-differentiated endometrial adenocarcinoma cell line (Ishikawa cells) and choriocarcinoma humantrophoblast cells (Jar cells), an in vitro assay mimicking human implantation was established. To investigate the impact of blockingMFG-E8 and integrin avb3, we pretreated the cell lines with antibodies against those proteins at different concentrations before theattachment assay.Main Outcome Measure(s): Attachment rate of Jar spheroids to the epithelial cell monolayer.Result(s): Pretreatment of Ishikawa cells with anti-MFG-E8 antibody caused a dose-dependent and significant inhibition ofattachment. On the other hand, pretreatment of Jar spheroids did not result in a significant effect on the attachment rate.Pretreatment of Ishikawa cells as well as Jar spheroids with anti-integrin avb3 antibodies resulted in a dose-dependent, significantinhibition of attachment.Conclusion(s): This study showed that blocking MFG-E8 and its receptor integrin avb3 in Ishikawa cells diminishes Jar spheroid
Use your smartphone
attachment. Moreover, blocking integrin avb3 in the trophoblastic cells also diminished theirattachment to the Ishikawa monolayer. (Fertil Steril� 2014;101:874–82. �2014 by AmericanSociety for Reproductive Medicine.)Key Words: Attachment, integrin avb3, implantation, human endometrium, Jar spheroids,MFG-E8
Discuss: You can discuss this article with its authors and with other ASRM members at http://fertstertforum.com/schmitzc-human-endometrium-mfg-e8-integrin-implantation/
to scan this QR codeand connect to thediscussion forum forthis article now.*
* Download a free QR code scanner by searching for “QRscanner” in your smartphone’s app store or app marketplace.
Received March 29, 2013; revised November 29, 2013; accepted December 5, 2013; published onlineJanuary 11, 2014.
C.S. has nothing to disclose. L.Y. has nothing to disclose. S.B. has nothing to disclose. S.A. hasnothing to disclose. J.S.C.-F. has nothing to disclose. B.S.R. has nothing to disclose. S.O. hasnothing to disclose.
C.S.'s present address is: Postgraduate Program, Medical Clinics of the Universidade Federal do RioGrande do Sul, Porto Alegre, Brazil.
J.S.C.-F.'s present address is: Department of Obstetrics and Gynecology, Universidade Federal do RioGrande do Sul, Porto Alegre, Brazil.
C.S. and L.Y. contributed equally to this work.Reprint requests: Sergio Oehninger, M.D., Ph.D., The Jones Institute for Reproductive Medicine, 601
Colley Ave., Norfolk, Virginia 23507 (E-mail: [email protected]).
Fertility and Sterility® Vol. 101, No. 3, March 2014 0015-0282/$36.00Copyright ©2014 American Society for Reproductive Medicine, Published by Elsevier Inc.http://dx.doi.org/10.1016/j.fertnstert.2013.12.015
874
I t is well known that implantationrequires a receptive endometrium, anormal and functional embryo at
the blastocyst stage of development,and a synchronized dialogue betweenthem (1). Nevertheless, there are manyunknown factors that play an importantrole in this process (2, 3). Because humanimplantation studies in vivo are verylimited (mainly owing to ethicalreasons), have poor reproducibility, and
VOL. 101 NO. 3 / MARCH 2014

Fertility and Sterility®
lack capacity formanipulation/intervention, in vitro studies arethe most common ones performed (4–6).
Integrins are a diverse family of glycoprotein receptorsexpressed on the cell membrane as heterodimeric a and b sub-units involved in both cell–cell and cell–substratum adhesion(7, 8). One particular integrin, the avb3, has been studied inmany physiologic/pathologic processes, and it has beensuggested to be involved in implantation across manymammalian species, including mouse, rat, pig, rabbit, andhuman (8–12). In women, an adequate integrin avb3 receptorexpression seems to be necessary for a receptive endometrium(7, 13–15). Integrin's hallmark is the ability of individualfamily members to recognize multiple extracellular matrixligands, such as bone matrix proteins, collagens, fibronectins,fibrinogen, laminins, thrombospondins, vitronectin, and vonWillebrand factor (16, 17). Despite the structural diversity oftheir ligands, integrins recognize a core ligand motif centeredon an acidic residue (18).
Recently, our group reported for the first time on a novelendometrial protein, milk fat globule-epidermal growthfactor (MFG-E8). Initially it was found that MFG-E8 was up-regulated in the human endometrium during the window ofimplantation in a microarray study comparing the geneexpression profile during the mid- vs. early luteal phase (19).We showed that MFG-E8 protein is mostly expressed in theendometrial epithelial cells and during the window of implan-tation in healthy ovulatory women, both in luminal and glan-dular epithelial cells, and with intense staining at both apicaland basal cellular compartments (20). The MFG-E8 receptor,integrin anb3, was present in the epithelial and stromal com-partments, with cycle-dependent and coincident peak expres-sions at mid-secretory phase, as previously described (20, 21).
The secreted glycoprotein MFG-E8 (also known as lactad-herin [SED1] and breast antigen 46 [BA46]) was initiallydescribed as a component of the milk fat globule membrane,which is mainly secreted in micro-vesicles (22). It has alsobeen suggested as a tumor marker in breast carcinomas (23,24). Human MFG-E8 is a 46-kDa glycoprotein peripherallyassociated with the cell membrane having two N-terminalepidermal growth factor-like repeats, one of which includesan arginine-glycine-aspartic acid motif that serves as theligand to its anb3 integrin receptor, facilitating cell adhesionand signal transduction. The two C-terminal discoidin C1–C2domains are homologous to coagulation factors V and VIIIand are responsible for binding to cell membrane phospho-lipids or cell surface carbohydrate moieties as well as to theextracellular matrix (25, 26).
MFG-E8 has a variety of functions in many extrauterinetissues related to apoptosis, cell adhesion and remodeling,neovascularization, and immunomodulation. In particular,it promotes phagocytosis of apoptotic cells by anb3/b5integrin-expressing phagocytes (27) as well as endothelialand epithelial cells (28, 29). In systemic endothelial cells,MFG-E8 binding to anb3/b5 integrins promotes vascularendothelial growth factor–induced survival and proliferation,leading to angiogenesis (30). It stimulates cell–cell andcell–extracellular matrix adhesion during sperm–oocyteinteraction (25, 31). In intestinal, mammary gland, andepididyimal epithelium, MFG-E8 regulates migration of
VOL. 101 NO. 3 / MARCH 2014
epithelial cells (25, 32, 33). It also protects againstsymptomatic rotavirus infection (34).
On the basis of recent work from our laboratories, wepostulated MFG-E8 as a novel glycoprotein with key roles inthe regulation of endometrial function (20). We showed thatMFG-E8 protein was up-regulated by prolactin in primaryendometrial epithelial cell cultures. This finding supports amodulatory role for prolactin as a stromal–epithelial paracrinefactor controlling MFG-E8 production. We also demonstratedthat in epithelial Ishikawa cells, MFG-E8 protein secretion isup-regulated by hCG and secreted via microparticles (35).We also confirmed that MFG-E8 has proapoptotic activity inhuman endometrial stromal cells, suggesting that this mole-cule acts as a paracrine factor (36). Furthermore, we reportedthat MFG-E8 protein is highly expressed in human chorionicvilli at all trimesters of gestation (in both cytotrophoblasts andsyncytiotrophoblasts) and in murine implantation sites (37).Finally, we presented new evidence that recombinantMFG-E8 modulates endometrial endothelial cell proliferationand adhesion under in vitro conditions (38).
In the present experiments we investigated the role ofMFG-E8 and integrin avb3 in the attachment of trophoblastcells to the endometrial epithelium, to further define its func-tional roles during the early implantation events. We used anin vitro model with Jar spheroids and Ishikawa cells. TheIshikawa cell line is a human, well-differentiated endometrialadenocarcinoma cell line recognized as a well-characterizedmodel for studying endometrial function as a surrogate forepithelial endometrial cells (39, 40, 41). Jar cells are derivedfrom a human choriocarcinoma, tend to grow as multicellularspheroids when cultured under appropriate conditions, andare largely used in blastocyst/trophoblast attachment studies(1, 42–45). Ishikawa cells/Jar spheroids interactions havebeen demonstrated to represent an adequate model forstudying early implantation events (1, 40, 41).
MATERIALS AND METHODSCell Lines and Culture Conditions
Current work in our laboratories using endometrial tissueand cells has been approved by the Eastern Virginia MedicalSchool institutional review board (#04-10-FB-0279 and#09-07-EX-0153, active projects). In this particular studyconsent was not required because the use of established cellslines is considered exempt. The human endometrial adeno-carcinoma cell line Ishikawa was generously provided byC. Lockwood, M.D. from Yale University. Cells used in theseexperiments were from passage 10 and were characterized byimmunostaining as cytokeratin-18þ and vimentin�, andwere also estrogen receptor aþ and bþ, as previously re-ported by us (data not shown) (4, 35). Ishikawa cells werecultured in Dulbecco's modified essential medium/F12containing 100 U/mL penicillin and 100 mg/mLstreptomycin (Gibco) and supplemented with 5% fetalbovine serum (Atlanta Biological) in 75-mL flasks at 37�Cin a humid atmosphere with 5% CO2. The cells were thenplated in four- well plates (Thermo Fisher Scientific) untilreaching 100% confluence and used in subsequentexperiments.
875
ORIGINAL ARTICLE: REPRODUCTIVE SCIENCE
The human choriocarcinoma Jar cell line was purchasedfrom ATCC. Cells used in these experiments were frompassage 6 and were characterized by immunostaining ascytokeratin-7þ and hCGþ, as previously reported by us (datanot shown) (4, 46). The Jar cells were cultured in RoswellPark Memorial Institute 1640 medium (Gibco) with 5% fetalbovine serum (Atlanta Biological) and 100 U/mL penicillinand 100 mg/mL streptomycin (Gibco). After reaching 80%confluence, Jar cells were detached with 0.05% trypsin(Gibco). A single-cell suspension was transferred into anagarose-coated Petri dish and cultured for 2 days to formspheroids. All cultures were conducted in an incubator with5% CO2 at 37�C. The diameter of Jar spheroids as measuredby inverted microscopy was between 150 and 200 mm (4, 46).
Immunocytochemistry
The Dako EnVisionþ System-HRP kit (DAB, Dako) was usedto perform immunocytochemistry. Ishikawa cells weregrown on glass coverslips and fixed with methanol at�20�C for 10 minutes. After washing with phosphate-buffered saline, endogenous peroxidase activity wasremoved with 3% hydrogen peroxide for 10 minutes.Nonspecific binding sites were blocked with 2% normalgoat serum for 30 minutes at room temperature. Cells wereincubated overnight at 4�C with the following mouse mono-clonal primary antibodies prepared in 1% normal goat
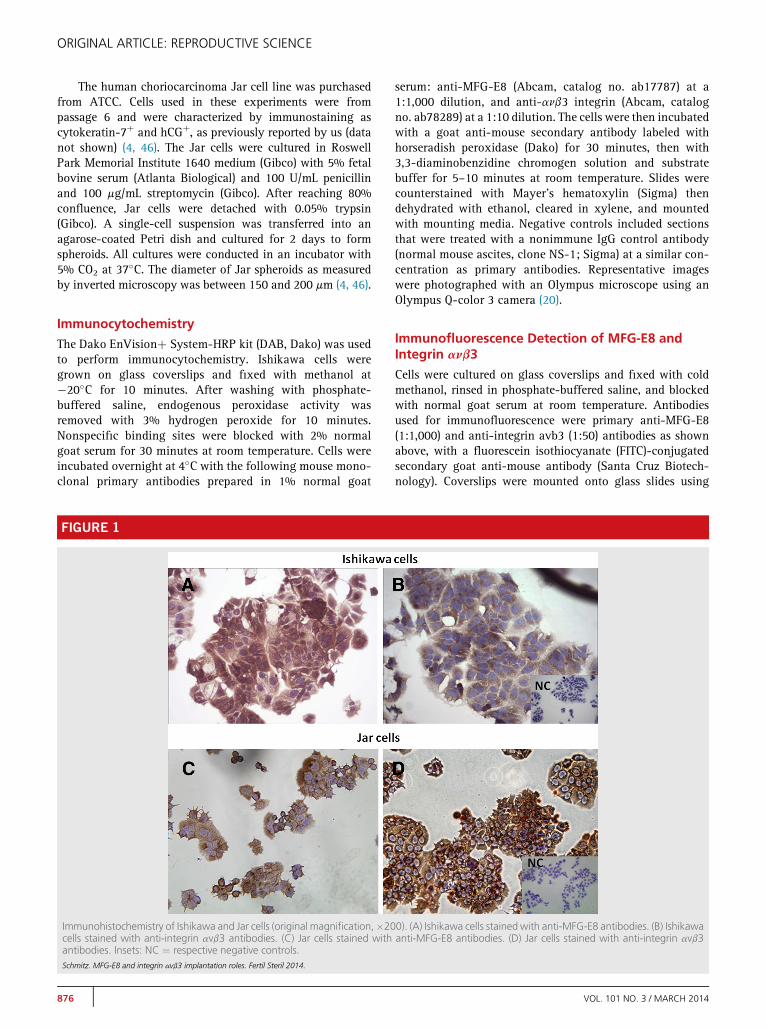
FIGURE 1
Immunohistochemistry of Ishikawa and Jar cells (original magnification,�20cells stained with anti-integrin avb3 antibodies. (C) Jar cells stained withantibodies. Insets: NC ¼ respective negative controls.Schmitz. MFG-E8 and integrin avb3 implantation roles. Fertil Steril 2014.
876
serum: anti-MFG-E8 (Abcam, catalog no. ab17787) at a1:1,000 dilution, and anti-anb3 integrin (Abcam, catalogno. ab78289) at a 1:10 dilution. The cells were then incubatedwith a goat anti-mouse secondary antibody labeled withhorseradish peroxidase (Dako) for 30 minutes, then with3,3-diaminobenzidine chromogen solution and substratebuffer for 5–10 minutes at room temperature. Slides werecounterstained with Mayer's hematoxylin (Sigma) thendehydrated with ethanol, cleared in xylene, and mountedwith mounting media. Negative controls included sectionsthat were treated with a nonimmune IgG control antibody(normal mouse ascites, clone NS-1; Sigma) at a similar con-centration as primary antibodies. Representative imageswere photographed with an Olympus microscope using anOlympus Q-color 3 camera (20).
Immunofluorescence Detection of MFG-E8 andIntegrin anb3
Cells were cultured on glass coverslips and fixed with coldmethanol, rinsed in phosphate-buffered saline, and blockedwith normal goat serum at room temperature. Antibodiesused for immunofluorescence were primary anti-MFG-E8(1:1,000) and anti-integrin avb3 (1:50) antibodies as shownabove, with a fluorescein isothiocyanate (FITC)-conjugatedsecondary goat anti-mouse antibody (Santa Cruz Biotech-nology). Coverslips were mounted onto glass slides using
0). (A) Ishikawa cells stained with anti-MFG-E8 antibodies. (B) Ishikawaanti-MFG-E8 antibodies. (D) Jar cells stained with anti-integrin avb3
VOL. 101 NO. 3 / MARCH 2014
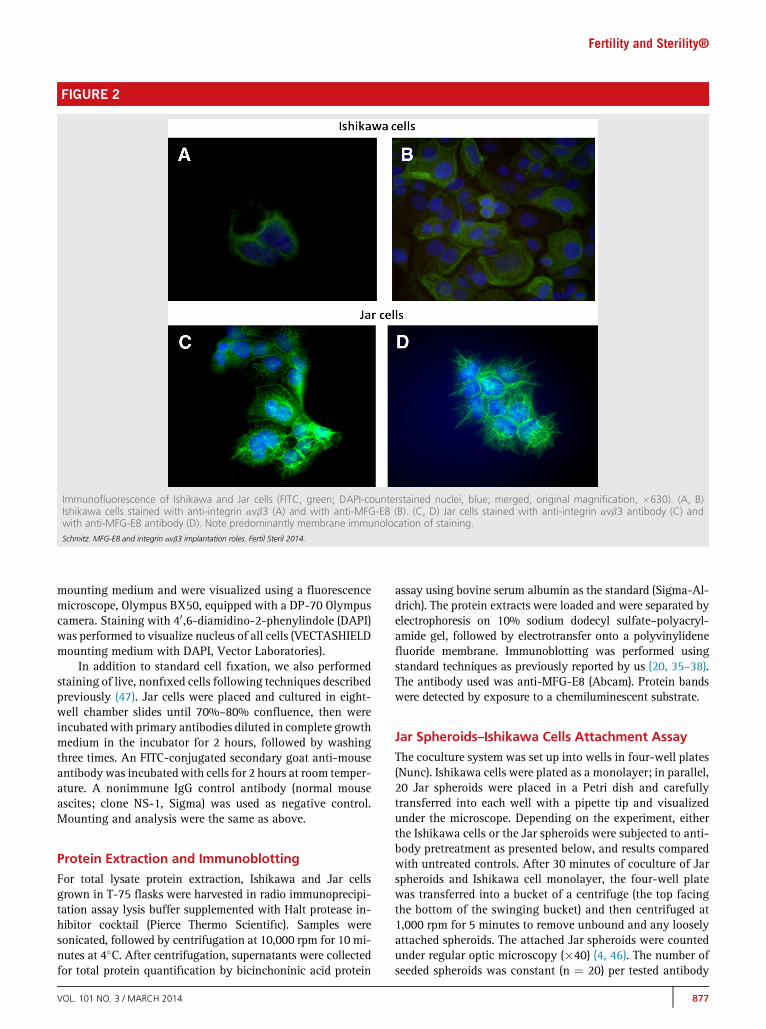
FIGURE 2
Immunofluorescence of Ishikawa and Jar cells (FITC, green; DAPI-counterstained nuclei, blue; merged, original magnification, �630). (A, B)Ishikawa cells stained with anti-integrin avb3 (A) and with anti-MFG-E8 (B). (C, D) Jar cells stained with anti-integrin avb3 antibody (C) andwith anti-MFG-E8 antibody (D). Note predominantly membrane immunolocation of staining.Schmitz. MFG-E8 and integrin avb3 implantation roles. Fertil Steril 2014.
Fertility and Sterility®
mounting medium and were visualized using a fluorescencemicroscope, Olympus BX50, equipped with a DP-70 Olympuscamera. Staining with 40,6-diamidino-2-phenylindole (DAPI)was performed to visualize nucleus of all cells (VECTASHIELDmounting medium with DAPI, Vector Laboratories).
In addition to standard cell fixation, we also performedstaining of live, nonfixed cells following techniques describedpreviously (47). Jar cells were placed and cultured in eight-well chamber slides until 70%–80% confluence, then wereincubated with primary antibodies diluted in complete growthmedium in the incubator for 2 hours, followed by washingthree times. An FITC-conjugated secondary goat anti-mouseantibody was incubated with cells for 2 hours at room temper-ature. A nonimmune IgG control antibody (normal mouseascites; clone NS-1, Sigma) was used as negative control.Mounting and analysis were the same as above.
Protein Extraction and Immunoblotting
For total lysate protein extraction, Ishikawa and Jar cellsgrown in T-75 flasks were harvested in radio immunoprecipi-tation assay lysis buffer supplemented with Halt protease in-hibitor cocktail (Pierce Thermo Scientific). Samples weresonicated, followed by centrifugation at 10,000 rpm for 10 mi-nutes at 4�C. After centrifugation, supernatants were collectedfor total protein quantification by bicinchoninic acid protein
VOL. 101 NO. 3 / MARCH 2014
assay using bovine serum albumin as the standard (Sigma-Al-drich). The protein extracts were loaded and were separated byelectrophoresis on 10% sodium dodecyl sulfate–polyacryl-amide gel, followed by electrotransfer onto a polyvinylidenefluoride membrane. Immunoblotting was performed usingstandard techniques as previously reported by us (20, 35–38).The antibody used was anti-MFG-E8 (Abcam). Protein bandswere detected by exposure to a chemiluminescent substrate.
Jar Spheroids–Ishikawa Cells Attachment Assay
The coculture system was set up into wells in four-well plates(Nunc). Ishikawa cells were plated as a monolayer; in parallel,20 Jar spheroids were placed in a Petri dish and carefullytransferred into each well with a pipette tip and visualizedunder the microscope. Depending on the experiment, eitherthe Ishikawa cells or the Jar spheroids were subjected to anti-body pretreatment as presented below, and results comparedwith untreated controls. After 30 minutes of coculture of Jarspheroids and Ishikawa cell monolayer, the four-well platewas transferred into a bucket of a centrifuge (the top facingthe bottom of the swinging bucket) and then centrifuged at1,000 rpm for 5 minutes to remove unbound and any looselyattached spheroids. The attached Jar spheroids were countedunder regular optic microscopy (�40) (4, 46). The number ofseeded spheroids was constant (n ¼ 20) per tested antibody
877
ORIGINAL ARTICLE: REPRODUCTIVE SCIENCE
dose (and control). There were 10 replicates per investigatedcondition (antibody dose and control) that were performedon three different experimental days. Additionalexperiments were performed in a similar fashion using anegative control with pretreatment with a nonimmune IgGcontrol antibody (normal mouse ascites; clone NS-1 Sigma)at equal concentration as primary antibodies and comparedwith nontreated (non-IgG) conditions.
Treatment Conditions
First, Ishikawa cells in the four-well plates were pretreatedwith the anti-MFG-E8 antibodies at concentrations of 1 mg/mL and 2 mg/mL or with the anti-integrin avb3 antibodiesat concentrations of 1 mg/mL, 10 mg/mL, and 50 mg/ml for2 hours, after which Jar spheroids were seeded on top of themonolayer for the attachment assay. When Ishikawa cellswere treated, no treatment was added to Jar spheroids for30 minutes.
Second, Jar spheroids were pretreated with either anti-MFG-E8 antibodies at concentrations of 1 mg/mL and 2 mg/mL or with anti-integrin avb3 antibodies at a concentrationof 50 mg/mL, for 15 minutes, after which Jar spheroids wereseeded on top of the (untreated) Ishikawa cell monolayer forthe attachment assay.
The anti-MFG-E8 and anti-integrin avb3 antibody dosesused for treatment were chosen on the basis of an in vitro hu-man endometrial endothelial cell adhesion and proliferation
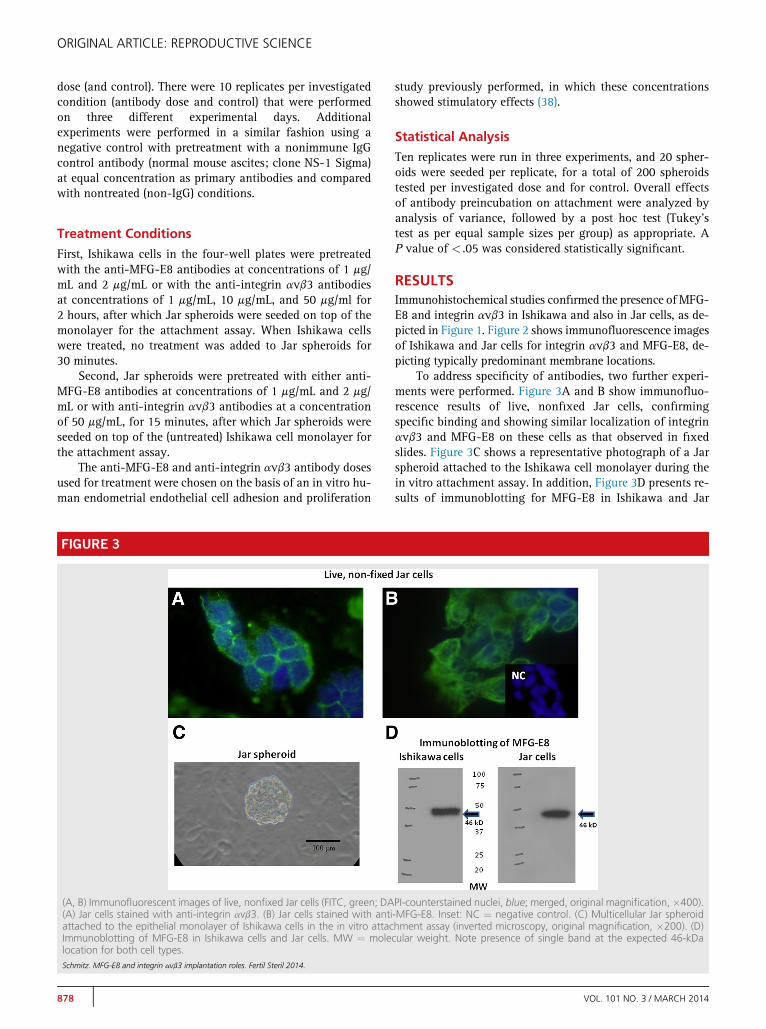
FIGURE 3
(A, B) Immunofluorescent images of live, nonfixed Jar cells (FITC, green; DA(A) Jar cells stained with anti-integrin avb3. (B) Jar cells stained with antiattached to the epithelial monolayer of Ishikawa cells in the in vitro attacImmunoblotting of MFG-E8 in Ishikawa cells and Jar cells. MW ¼ molelocation for both cell types.Schmitz. MFG-E8 and integrin avb3 implantation roles. Fertil Steril 2014.
878
study previously performed, in which these concentrationsshowed stimulatory effects (38).
Statistical Analysis
Ten replicates were run in three experiments, and 20 spher-oids were seeded per replicate, for a total of 200 spheroidstested per investigated dose and for control. Overall effectsof antibody preincubation on attachment were analyzed byanalysis of variance, followed by a post hoc test (Tukey'stest as per equal sample sizes per group) as appropriate. AP value of < .05 was considered statistically significant.
RESULTSImmunohistochemical studies confirmed the presence of MFG-E8 and integrin avb3 in Ishikawa and also in Jar cells, as de-picted in Figure 1. Figure 2 shows immunofluorescence imagesof Ishikawa and Jar cells for integrin avb3 and MFG-E8, de-picting typically predominant membrane locations.
To address specificity of antibodies, two further experi-ments were performed. Figure 3A and B show immunofluo-rescence results of live, nonfixed Jar cells, confirmingspecific binding and showing similar localization of integrinavb3 and MFG-E8 on these cells as that observed in fixedslides. Figure 3C shows a representative photograph of a Jarspheroid attached to the Ishikawa cell monolayer during thein vitro attachment assay. In addition, Figure 3D presents re-sults of immunoblotting for MFG-E8 in Ishikawa and Jar
PI-counterstained nuclei, blue; merged, original magnification, �400).-MFG-E8. Inset: NC ¼ negative control. (C) Multicellular Jar spheroidhment assay (inverted microscopy, original magnification, �200). (D)cular weight. Note presence of single band at the expected 46-kDa
VOL. 101 NO. 3 / MARCH 2014
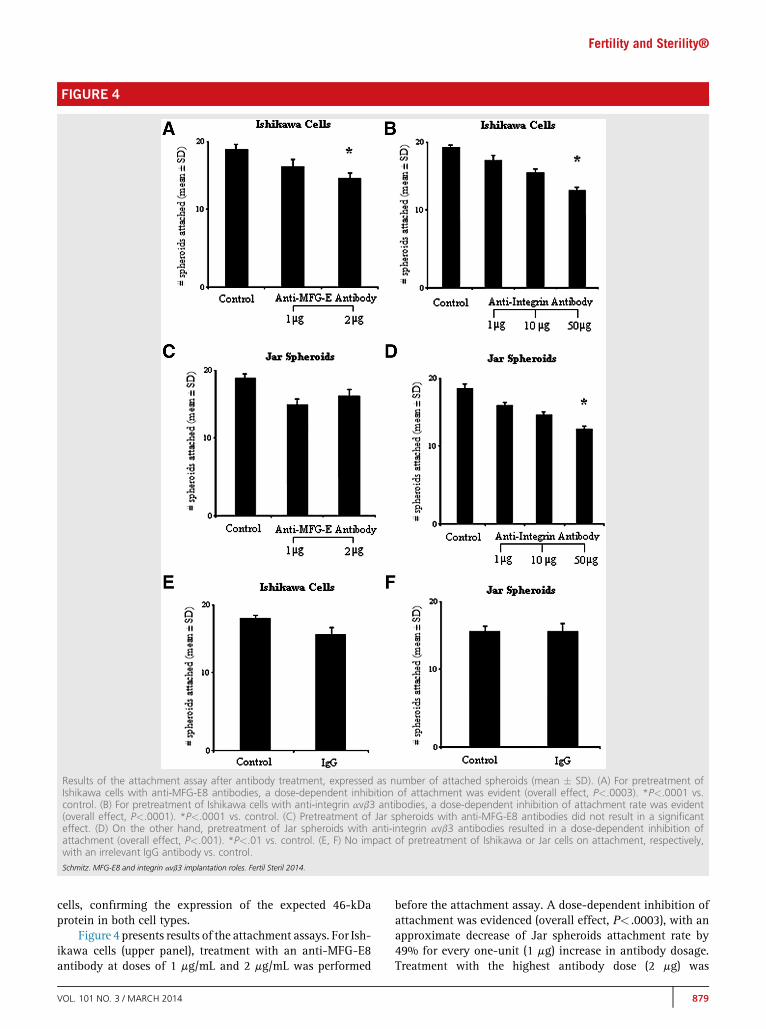
FIGURE 4
Results of the attachment assay after antibody treatment, expressed as number of attached spheroids (mean � SD). (A) For pretreatment ofIshikawa cells with anti-MFG-E8 antibodies, a dose-dependent inhibition of attachment was evident (overall effect, P<.0003). *P<.0001 vs.control. (B) For pretreatment of Ishikawa cells with anti-integrin avb3 antibodies, a dose-dependent inhibition of attachment rate was evident(overall effect, P<.0001). *P<.0001 vs. control. (C) Pretreatment of Jar spheroids with anti-MFG-E8 antibodies did not result in a significanteffect. (D) On the other hand, pretreatment of Jar spheroids with anti-integrin avb3 antibodies resulted in a dose-dependent inhibition ofattachment (overall effect, P<.001). *P<.01 vs. control. (E, F) No impact of pretreatment of Ishikawa or Jar cells on attachment, respectively,with an irrelevant IgG antibody vs. control.Schmitz. MFG-E8 and integrin avb3 implantation roles. Fertil Steril 2014.
Fertility and Sterility®
cells, confirming the expression of the expected 46-kDaprotein in both cell types.
Figure 4 presents results of the attachment assays. For Ish-ikawa cells (upper panel), treatment with an anti-MFG-E8antibody at doses of 1 mg/mL and 2 mg/mL was performed
VOL. 101 NO. 3 / MARCH 2014
before the attachment assay. A dose-dependent inhibition ofattachment was evidenced (overall effect, P< .0003), with anapproximate decrease of Jar spheroids attachment rate by49% for every one-unit (1 mg) increase in antibody dosage.Treatment with the highest antibody dose (2 mg) was
879

ORIGINAL ARTICLE: REPRODUCTIVE SCIENCE
significantly lower than controls (P< .0001), with a reductionof attachment by 76% (Fig. 4A). Treatment with anti-integrinavb3 antibodies at doses of 1 mg/mL, 10 mg/mL, and 50 mg/mLvs. controls was carried out before the attachment assay. Adose-dependent inhibition of attachment rate was observed(overall effect, P< .0001), and the attachment rate decreasedby approximately 2% per every one-unit (1 mg) increase inantibody dosage. The treatment with 4 mg was significantlylower than controls (P< .0001), with a reduction of attachmentby 81% (Fig. 4B).
For Jar spheroids (Fig. 4, middle panel), pretreatment ofJar spheroids with anti-MFG-E8 antibody did not result in asignificant effect on attachment rate (Fig. 4C). Conversely,pretreatment of Jar spheroids with anti-integrin avb3antibodies showed a dose-dependent inhibition of attachment(overall effect, P< .001), and treatment with the highestantibody dose (2 mg) was significantly lower than controls(P< .01), with a reduction of attachment by 44% (Fig. 4D).
Pretreatment with IgG antibody (negative control)showed no difference of attachment compared with control,untreated conditions (P>.7) in either Ishikawa (Fig. 4E) orJar (Fig. 4F) cells.
DISCUSSIONPrevious studies have shown that integrin avb3 and MFG-E8are expressed in primary human epithelial cells, more inten-sively during the window of implantation (7, 19, 20). Ourstudy confirmed that both of them are also expressed inIshikawa cells (20, 35, 48). Furthermore, we demonstratedthat both proteins are present in trophoblast Jar cells byimmunochemistry and immunofluorescence. Integrin avb3has been previously found to be expressed in mouse andhuman embryos (49, 50), and MFG-E8 has been shown tobe expressed in human placental trophoblasts in all threetrimesters of pregnancy (36). Here we show for the first timethat Jar cells express MFG-E8. After these characterizationswe were able to proceed with our experiments to test ourhypothesis that blocking MFG-E8 and integrin avb3 by spe-cific antibodies might impair the attachment of Jar spheroids(mimicking the trophoblast/embryo) to the Ishikawa cellmonolayer (simulating the endometrial epithelium).
Integrins divalent-cation-dependent binding to theirligands regulates many cellular processes, such as remodelingof bone, homeostasis, fertilization, and implantation (15, 18).Integrin avb3 and its ligands' roles in implantation have beensubject to extensive investigation in the last few years. It wasshown that blocking integrin avb3 and Lewis Y, adifucosylated oligosaccharide carried by this integrin, inepithelial RL95-2 cells diminishes Jar spheroids attachment(5). Another study performed in Ishikawa cells knocked downintegrin avb3 and demonstrated diminished attachment of ratblastocysts to those cells (51). In agreement with these results,we found diminished attachment of Jar's spheroids to Ishikawacells when this integrin was blocked. Additionally, we alsoshowed that blocking integrin avb3 present in the trophoblastcells decreases attachment, an effect not reported before.
MFG-E8 is a glycoprotein that contains two epidermalgrowth factor domains, a proline/threonine-rich domain,
880
and two factor-VIII-homologous domains (25, 26). It has anessential role in an adequate functioning immune system,promoting the engulfment of apoptotic cells bymacrophages and dendritic cells (52). MFG-E8 dysfunctionseems to be implicated in some pathologic processes, suchas tumor progression and systemic lupus erythematosus (53,54). Furthermore, MFG-E8 promotes angiogenesis and is acrucial mediator of vascular endothelial growth factor proan-giogenic effect, and its therapeutic potential has been pointedout for future disease treatment, such as sepsis and certaintypes of cancers (55–57).
Although its function in mediating phagocytic engulf-ment seems to be the most investigated one, MFG-E8's rolein the implantation process has also received some attentionrecently. It has been shown that there is an increase in itsendometrial expression during the window of implantationin human (19, 20). Here we demonstrated that blockingMFG-E8 in Ishikawa cells significantly diminishes Jarspheroid attachment, although blocking the protein in theJar spheroids did not have the same effect. To the best ofour knowledge this is the first evidence of MFG-E8 adhesionproperties in a trophoblast/endometrial epithelial cell modelthat simulates human implantation.
In conclusion, results demonstrated that independentblockage ofMFG-E8and its receptor integrinavb3 in Ishikawacells significantly diminished Jar spheroid attachment. More-over, we showed that blockage of integrin avb3 in the tropho-blastic tissue (Jar cells) also diminished their attachment to theIshikawa monolayer. Further characterization of MFG-E8 andintegrin avb3 intracellular pathways involved in this processis required for a better understanding of the roles of theseproteins in normal and pathologic human implantation.
REFERENCES1. Evron A, Goldman S, Shalev E. Effect of primary human endometrial stromal
cells on epithelial cell receptivity and protein expression is dependent onmenstrual cycle stage. Hum Reprod 2011;26:176–90.
2. Diedrich K, Fauser BC, Devroey P, Griesinger G, Evian Annual Reproduction(EVAR) Workshop Group. The role of the endometrium and embryo in hu-man implantation. Hum Reprod Update 2007;13:365–77.
3. Oehninger S. Revealing the enigmas of implantation: what is the true impactof ovarian hyperstimulation? Fertil Steril 2008;89:27–30.
4. Wang H, Pilla F, Anderson S, Martínez-Escribano S, Herrer I, Moreno-Moya JM, et al. A novel model of human implantation: 3D endometrium-like culture system to study attachment of human trophoblast (Jar) cellspheroids. Mol Hum Reprod 2012;18:33–43.
5. Zhang D, Wei J, Wang J, Liu S, Wang X, Yan Q. Difucosylated oligosaccha-ride Lewis Y is contained within integrin avb3 on RL95-2 cells and requiredfor endometrial receptivity. Fertil Steril 2011;95:1446–51.
6. AboussahoudW, Bruce C, Elliott S, Fazeli A. Activation of Toll-like receptor 5decreases the attachment of human trophoblast cells to endometrial cellsin vitro. Hum Reprod 2010;25:2217–28.
7. Lessey BA, Damjanovich L, Coutifaris C, Castelbaum A, Albelda SM,Buck CA. Integrin adhesion molecules in the human endometrium. Corre-lation with the normal and abnormal menstrual cycle. J Clin Invest 1992;90:188–95.
8. Illera MJ, Cullinan E, Gui Y, Yuan L, Beyler SA, Lessey BA. Blockade of thealpha(v)beta(3) integrin adversely affects implantation in the mouse. Biol Re-prod 2000;62:1285–90.
9. Srinivasan KR, Blesson CS, Fatima I, Kitchlu S, Jain SK, Mehrotra PK, et al.Expression of alphaVbeta3 integrin in rat endometrial epithelial cells and itsfunctional roleduring implantation.GenCompEndocrinol2009;160:124–33.
VOL. 101 NO. 3 / MARCH 2014

Fertility and Sterility®
10. Lin H, Wang X, Liu G, Fu J, Wang A. Expression of alphaV and beta3 integrinsubunits during implantation in pig. Mol Reprod Dev 2007;74:1379–85.
11. Illera MJ, Lorenzo PL, Gui YT, Beyler SA, Apparao KB, Lessey BA. A role foralphavbeta3 integrin during implantation in the rabbit model. Biol Reprod2003;68:766–71.
12. LesseyBA. Twopathwaysof progesteroneaction in thehumanendometrium:implications for implantation and contraception. Steroids 2003;68:809–15.
13. Surrey ES, Minjarez DA, Schoolcraft WB. The incidence of aberrant endome-trial alphavbeta(3) vitronectin expression in a high risk infertility population:could prolonged GnRH agonist therapy play a role? J Assist Reprod Genet2007;24:553–6.
14. Lessey BA, Castelbaum AJ, Buck CA, Lei Y, Yowell CW, Sun J. Further char-acterization of endometrial integrins during the menstrual cycle and in preg-nancy. Fertil Steril 1994;62:497–506.
15. Lindhard A, Bentin-Ley U, Ravn V, Islin H, Hviid T, Rex S, et al. Biochemicalevaluation of endometrial function at the time of implantation. Fertil Steril2002;78:221–33.
16. Plow EF, Haas TA, Zhang L, Loftus J, Smith JW. Ligand binding to integrins. JBiol Chem 2000;275:21785–8.
17. Aziz M, Jacob A, Matsuda A, Wang P. Review: milk fat globule-EGF factor 8expression, function and plausible signal transduction in resolving inflamma-tion. Apoptosis 2011;16:1077–86.
18. Arnaout MA. Integrin structure: new twists and turns in dynamic cell adhe-sion. Immunol Rev 2002;186:125–40.
19. Mirkin S, Arslan M, Churikov D, Corica A, Diaz JI, Williams S, et al. In searchof candidate genes critically expressed in the human endometrium duringthe window of implantation. Hum Reprod 2005;20:2104–17.
20. Franchi A, Bocca S, Anderson S, Riggs R, Oehninger S. Expression of milk fatglobule EGF-factor 8 (MFG-E8) mRNA and protein in the human endome-trium and its regulation by prolactin. Mol Hum Reprod 2011;17:360–71.
21. Apparao KB, Murray MJ, Fritz MA, Meyer WR, Chambers AF, Truong PR,et al. Osteopontin and its receptor alphavbeta(3) integrin are coexpressedin the human endometrium during the menstrual cycle but regulated differ-entially. J Clin Endocrinol Metab 2001;86:4991–5000.
22. Stubbs JD, Lekutis C, Singer KL, Bui A, Yuzuki D, Srinivasan U, et al. cDNAcloning of a mouse mammary epithelial cell surface protein reveals the exis-tence of epidermal growth factor-like domains linked to factor VIII-like se-quences. Proc Natl Acad Sci U S A 1990;87:8417–21.
23. Peterson JA, Zava DT, Duwe AK, Blank EW, Battifora H, Ceriani RL. Biochem-ical and histological characterization of antigens preferentially expressed onthe surface and cytoplasm of breast carcinoma cells identified by mono-clonal antibodies against the human milk fat globule. Hybridoma 1990;9:221–35.
24. Taylor MR, Couto JR, Scallan CD, Ceriani RL, Peterson JA. Lactadherin(formerly BA46), a membrane-associated glycoprotein expressed in humanmilk and breast carcinomas, promotes Arg-Gly-Asp (RGD)-dependent celladhesion. DNA Cell Biol 1997;16:861–9.
25. Ensslin MA, Shur BD. The EGF repeat and discoidin domain protein,SED1/MFG-E8, is required for mammary gland branching morphogenesis.Proc Natl Acad Sci U S A 2007;104:2715–20.
26. Raymond A, Ensslin MA, Shur BD. SED1/MFG-E8: a bi-motif protein that or-chestrates diverse cellular interactions. J Cell Biochem 2009;106:957–66.
27. Hanayama R, TanakaM, Miwa K, Shinohara A, Iwamatsu A, Nagata S. Iden-tification of a factor that links apoptotic cells to phagocytes. Nature 2002;417:182–7.
28. Nandrot EF, Anand M, Almeida D, Atabai K, Sheppard D, Finnemann SC.Essential role for MFG-E8 as ligand for alphavbeta5 integrin in diurnal retinalphagocytosis. Proc Natl Acad Sci USA 2007;104:12005–10.
29. FensMH,Mastrobattista E, de Graaff AM, Flesch FM, Ultee A, Rasmussen JT,et al. Angiogenic endothelium shows lactadherin-dependent phagocytosisof aged erythrocytes and apoptotic cells. Blood 2008;111:4542–50.
30. Silvestre JS, Th�ery C, Hamard G, Boddaert J, Aguilar B, Delcayre A, et al. Lac-tadherin promotes VEGF-dependent neovascularization. Nat Med 2005;11:499–506.
31. Ensslin MA, Shur BD. Identification of mouse sperm SED1, a bimotif EGFrepeat and discoidin-domain protein involved in sperm-egg binding. Cell2003;114:405–17.
VOL. 101 NO. 3 / MARCH 2014
32. Bu HF, Zuo XL, Wang X, Ensslin MA, Koti V, HsuehW, et al. Milk fat globule-EGF factor 8/lactadherin plays a crucial role in maintenance and repair ofmurine intestinal epithelium. J Clin Invest 2007;117:3673–83.
33. Raymond AS, Elder B, Ensslin M, Shur BD. Loss of SED1/MFG-E8 results inaltered luminal physiology in the epididymis. Mol Reprod Dev 2010;77:550–63.
34. Newburg DS, Peterson JA, Ruiz-Palacios GM, Matson DO, Morrow AL,Shults J, et al. Role of human-milk lactadherin in protection against symp-tomatic rotavirus infection. Lancet 1998;351:1160–4.
35. Sarhan A, Bocca S, Anderson S, Jacot T, Burch T, Nyalwidhe J, et al. Hu-man endometrial milk fat globule-epidermal growth factor 8 (MFGE8) isup regulated by estradiol at the transcriptional level, and its secretionvia microvesicles is stimulated by human chorionic gonadotropin (hCG).Cell Signalling Trafficking 2013;1:1–10. http://dx.doi.org/10.7243/2054-1481-1-1.
36. Riggs RM, Bocca S, Anderson S, Franchi A, Rhavi BS, Oehninger S. Epithelialcell protein milk fat globule-epidermal growth factor 8 and human chorionicgonadotropin regulate stromal cell apoptosis in the human endometrium.Fertil Steril 2012;98:1549–56.
37. Bocca SM, Anderson S, Amaker B, Swanson RJ, Franchi A, Lattanzio F, et al.Milk fat globule epidermal growth factor 8 (MFG-E8): a novel protein in themammalian endometrium with putative roles in implantation and placenta-tion. Placenta 2012;33:795–802.
38. Bocca S, Anderson S, Oehninger S. Milk fat globule epithelial growth factor8 (MFG-E8) regulates human endometrial endothelial cell adhesion and pro-liferation. Fertil Steril 2010;94:S215–6.
39. Nishida M, Kasahara K, KanekoM, Iwasaki H, Hayashi K. [Establishment of anew human endometrial adenocarcinoma cell line, Ishikawa cells, contain-ing estrogen and progesterone receptors]. Nihon Sanka Fujinka Gakkai Zas-shi 1985;37:1103–11.
40. Hannan NJ, Paiva P, Dimitriadis E, Salamonsen LA. Models for study of humanembryo implantation: choice of cell lines? Biol Reprod 2010;82:235–45.
41. Singh H, Nardo L, Kimber SJ, Aplin JD. Early stages of implantation as re-vealed by an in vitro model. Reproduction 2010;139:905–14.
42. Harduf H, Goldman S, Shalev E. Human uterine epithelial RL95-2 andHEC-1A cell-line adhesiveness: the role of plexin B1. Fertil Steril 2007;87:1419–27.
43. Kodithuwakku SP, Ng PY, Liu Y, Ng EH, Yeung WS, Ho PC, et al. Hormonalregulation of endometrial olfactomedin expression and its suppressive effecton spheroid attachment onto endometrial epithelial cells. Hum Reprod2011;26:167–75.
44. Heneweer C, Kruse LH, Kindh€auser F, Schmidt M, Jakobs KH, Denker HW,et al. Adhesiveness of human uterine epithelial RL95-2 cells to trophoblast:rho protein regulation. Mol Hum Reprod 2002;8:1014–22.
45. Heneweer C, Schmidt M, Denker HW, Thie M. Molecular mechanismsin uterine epithelium during trophoblast binding: the role of smallGTPase RhoA in human uterine Ishikawa cells. J Exp Clin Assist Reprod2005;2:4.
46. Wang H, Bocca S, Anderson S, Yu L, Rhavi BS, Horcajadas J, et al. Sex ste-roids regulate epithelial-stromal cell cross talk and trophoblast attachmentand invasion in a three dimensional human endometrial culture system. Tis-sue Eng Part C Methods 2013;19:676–87.
47. Manos PD, Ratanasirintrawoot S, Loewer S, Daley GQ, Schlaeger TM. Live-cell immunofluorescence staining of human pluripotent stem cells. Curr Pro-toc Stem Cell Biol 2011. Chapter 1:Unit 1C.12.
48. Castelbaum AJ, Ying L, Somkuti SG, Sun J, Ilesanmi AO, Lessey BA. Charac-terization of integrin expression in a well differentiated endometrial adeno-carcinoma cell line (Ishikawa). J Clin Endocrinol Metab 1997;82:136–42.
49. Sutherland AE, Calarco PG, Damsky CH. Developmental regulation of integ-rin expression at the time of implantation in the mouse embryo. Develop-ment 1993;119:1175–86.
50. Campbell S, Swann HR, Seif MW, Kimber SJ, Aplin JD. Cell adhesion mole-cules on the oocyte and preimplantation human embryo. Hum Reprod1995;10:1571–8.
51. Kaneko Y, Day ML, Murphy CR. Integrin b3 in rat blastocysts and epithelialcells is essential for implantation in vitro: studies with Ishikawa cells andsmall interfering RNA transfection. Hum Reprod 2011;26:1665–74.
881

ORIGINAL ARTICLE: REPRODUCTIVE SCIENCE
52. Jinushi M, Nakazaki Y, Dougan M, Carrasco DR, Mihm M, Dranoff G. MFG-E8-mediated uptake of apoptotic cells by APCs links the pro- and antiinflam-matory activities of GM-CSF. J Clin Invest 2007;117:1902–13.
53. Oba J, Moroi Y, Nakahara T, Abe T, Hagihara A, Furue M. Expression of milkfat globule epidermal growth factor-VIII may be an indicator of poor prog-nosis in malignant melanoma. Br J Dermatol 2011;165:506–12.
54. Hu CY, Wu CS, Tsai HF, Chang SK, Tsai WI, Hsu PN. Genetic polymorphismin milk fat globule-EGF factor 8 (MFG-E8) is associated with systemic lupuserythematosus in human. Lupus 2009;18:676–81.
882
55. Shah KG, Wu R, Jacob A, Molmenti EP, Nicastro J, Coppa GF, et al. Recom-binant human milk fat globule-EGF factor 8 produces dose-dependent ben-efits in sepsis. Intensive Care Med 2012;38:128–36.
56. Frierson HF Jr, El-Naggar AK, Welsh JB, Sapinoso LM, Su AI, Cheng J,et al. Large scale molecular analysis identifies genes with alteredexpression in salivary adenoid cystic carcinoma. Am J Pathol 2002;161:1315–23.
57. Chen X, Cheung ST, So S, Fan ST, Barry C, Higgins J, et al. Gene expressionpatterns in human liver cancers. Mol Biol Cell 2002;13:1929–39.
VOL. 101 NO. 3 / MARCH 2014




![The Ising model of a ferromagnet from 1920 to 2020smirnov/slides/slides-ising-model.pdfMuch fascinating mathematics, expect more: • [Zamolodchikov, JETP 1987]: E8 symmetry in 2D](https://static.fdocument.org/doc/165x107/5fdbf69f1ab2af4dc43ecbfe/the-ising-model-of-a-ferromagnet-from-1920-to-2020-smirnovslidesslides-ising-modelpdf.jpg)













