Rapid identification of genomic interferon genes by sequencing on phage λ DNA templates
6
Rapid Identification of Genomic Interferon Genes by Sequencing on Phage h DNA Templates XIN-YUAN LIU, SHUICHIRO MAEDA, KENNETH COLLIER, and SIDNEY PESTKA A method for rapid screening and identification of a gene or closely related genes from a genomic library by directly sequencing h phage DNA without laborious in- termediate steps and subcloning is presented. The Sanger dideoxy sequencing technique with a synthetic eicosadeoxynucleotide (20-mer) primer and double- stranded DNA template was performed at 50°C rather than at room temperature to avoid the formation of nonspecific bands. Detailed steps of the procedure are described. The technique permitted the rapid and con- venient sequencing and identification of multiple human leukocyte interferon genes that had been isolated from a human genomic library in phage h. The dideoxy sequencing technique has been widely applied to determine the sequences of DNA (1-3) with single-stranded DNA (M13) as template. If large double-stranded DNA (dsDNA) can be used as template for the dideoxy se- quencing technique, a specific gene in the genomic library can be rapidly screened without further la- borious steps involved in subcloning of the ge- nomic DNA. In 1979, Smith et al. (4) described the use of short synthetic oligonucleotides as primers to sequence directly DNA segments Gene Anal Techn 2:83-88 1985, f _.- cloned in the dsDNA plasmid vector pBR322. In ---~'¢ 1981, Wallace et al. (5) synthesized oligodeoxyn- ~-~ ucleotides complementary to plasmid pBR322 at positions adjacent to five of the unique restriction endonuclease cleavage sites and used these oli- godeoxynucleotides as primers to sequence the DNA adjacent to the respective sites. Hong (6) extended the method for sequencing on large dsDNA (h DNA, 48,502 bp) templates with the dideoxy sequencing technique. In Zagursky et al. (7, this issue), a similar technique with the use of reverse transcriptase for obtaining DNA sequence directly from phage h is described. Here we apply and modify the above methods for rapid screening of interferon DNA sequences from a human ge- nomic library cloned in h Charon 4A. The primer used is a single-stranded oligodeoxynucleotide (20-mer) common to all the known Hu-IFN-a se- quences. Materials and Methods T4-polynucleotide kinase, deoxynucleotide tri- phosphates, and dideoxynucleotide triphosphates were purchased from P-L Chemicals, Ltd. The Klenow fragment of DNA polymerase I was from Boehringer Mannheim; b/-32p]ATP (specific ac- tivity > 5000 Ci/mmol, 10 mCi/ml) was from Amersham International, Ltd; proteinase K was from Bethesda Research Laboratories, Inc. The concentrated Buffer K (5 x) for the kinase reac- tion consisted of the following components: 0.5 M Tris. HC1, pH 7.5, 0.1 mM MgCI 2, and 0.02 M dithiothreitol (DTT). Sequencing Buffer S con- sisted of 0.1 M Tris. HCI, pH 8.3, 0.1 M MgCI 2, 0.02 M DTT. The dideoxy terminator mixtures were comprised of the following components: 1 ddG: 240 txM ddGTP ddA: 200 txM ddATP ddT: 540 txM ddTTP ddC: 100 txM ddCTP 11.25 IxM dGTP 200 txM dATP 150 txM dTTP 150 ~M dCTP 150 I~M dGTP 15 IxM dATP 150 IxM dTTP 150 ~M dCTP 150 IxM dGTP 200 jxM dATP 11 ~M dTTP 150 ~M dCTP 150 IxM dGTP 200/~M dATP 150 txM dTTP 11 ~M dCTP . From the Roche Institute of Molecular Biology, Roche Re- search Center, Nutley, New Jersey (S.P.), the Department of Biochemistry, Kumamoto University School of Medicine, Ku- mamoto, Japan (S.M.), and the Department of Molecular Ge- netics, Hoffmann-La Roche Inc., Nutley, New Jersey (K.C.). Present address of X.-Y.L.: Shanghai Institute of Biochem- istry, Academia Sinica, Shanghai, China. Address reprint requests to: Dr. Sidney Pestka, Roche In- stitute of Molecular Biology, Roche Research Center, Nutley, NJ 07110. Received June 14, 1985. The primer was an eicosadeoxynucleotide (20- i Abbreviations: ddGTP, dideoxyguanidine triphosphate; ddATE dideoxyadenosine triphosphate; ddTTE dideoxythy- midine triphosphate; ddCTP, dideoxycytidine triphosphate; dGTP, deoxyguanidine triphosphate; dATE deoxyadenosine triphosphate; dTTE thymidine triphosphate; dCTE deoxycy- tidine triphosphate; ddA, ddC, ddG, and ddT, dideoxynucleo- side mixtures for termination at A, C, G, and T residues, re- spectively. © 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York, NY 10017 0735-0651/85/$03.30
Transcript of Rapid identification of genomic interferon genes by sequencing on phage λ DNA templates

Rapid Identification of Genomic Interferon Genes by Sequencing on Phage h DNA Templates
X I N - Y U A N LIU,
S H U I C H I R O M A E D A ,
K E N N E T H C O L L I E R , and
S I D N E Y P E S T K A
A method for rapid screening and identification of a gene or closely related genes from a genomic library by directly sequencing h phage DNA without laborious in- termediate steps and subcloning is presented. The Sanger dideoxy sequencing technique with a synthetic eicosadeoxynucleotide (20-mer) primer and double- stranded DNA template was performed at 50°C rather than at room temperature to avoid the formation of nonspecific bands. Detailed steps of the procedure are described. The technique permitted the rapid and con- venient sequencing and identification of multiple human leukocyte interferon genes that had been isolated from a human genomic library in phage h.
The dideoxy sequencing technique has been widely applied to determine the sequences of DNA (1-3) with single-stranded DNA (M13) as template. If large double-stranded DNA (dsDNA) can be used as template for the dideoxy se- quencing technique, a specific gene in the genomic library can be rapidly screened without further la- borious steps involved in subcloning of the ge- nomic DNA. In 1979, Smith et al. (4) described the use of short synthetic oligonucleotides as primers to sequence directly DNA segments
Gene Anal Techn 2:83-88 1985,
f _.- cloned in the dsDNA plasmid vector pBR322. In ---~'¢ 1981, Wallace et al. (5) synthesized oligodeoxyn- ~ - ~ ucleotides complementary to plasmid pBR322 at positions adjacent to five of the unique restriction endonuclease cleavage sites and used these oli- godeoxynucleotides as primers to sequence the DNA adjacent to the respective sites. Hong (6) extended the method for sequencing on large dsDNA (h DNA, 48,502 bp) templates with the dideoxy sequencing technique. In Zagursky et al. (7, this issue), a similar technique with the use of reverse transcriptase for obtaining DNA sequence directly from phage h is described. Here we apply and modify the above methods for rapid screening of interferon DNA sequences from a human ge- nomic library cloned in h Charon 4A. The primer used is a single-stranded oligodeoxynucleotide (20-mer) common to all the known Hu-IFN-a se- quences.
Materials and Methods T4-polynucleotide kinase, deoxynucleotide tri- phosphates, and dideoxynucleotide triphosphates were purchased from P-L Chemicals, Ltd. The Klenow fragment of DNA polymerase I was from Boehringer Mannheim; b/-32p]ATP (specific ac- tivity > 5000 Ci/mmol, 10 mCi/ml) was from Amersham International, Ltd; proteinase K was from Bethesda Research Laboratories, Inc. The concentrated Buffer K (5 x) for the kinase reac- tion consisted of the following components: 0.5 M Tris. HC1, pH 7.5, 0.1 mM MgCI 2, and 0.02 M dithiothreitol (DTT). Sequencing Buffer S con- sisted of 0.1 M Tris. HCI, pH 8.3, 0.1 M MgCI 2, 0.02 M DTT. The dideoxy terminator mixtures were comprised of the following components: 1
ddG: 240 txM ddGTP ddA: 200 txM ddATP ddT: 540 txM ddTTP ddC: 100 txM ddCTP
11.25 IxM dGTP 200 txM dATP 150 txM dTTP 150 ~M dCTP 150 I~M dGTP 15 IxM dATP 150 IxM dTTP 150 ~M dCTP 150 IxM dGTP 200 jxM dATP 11 ~M dTTP 150 ~M dCTP 150 IxM dGTP 200/~M dATP 150 txM dTTP 11 ~M dCTP .
From the Roche Institute of Molecular Biology, Roche Re- search Center, Nutley, New Jersey (S.P.), the Department of Biochemistry, Kumamoto University School of Medicine, Ku- mamoto, Japan (S.M.), and the Department of Molecular Ge- netics, Hoffmann-La Roche Inc., Nutley, New Jersey (K.C.). Present address of X.-Y.L.: Shanghai Institute of Biochem- istry, Academia Sinica, Shanghai, China.
Address reprint requests to: Dr. Sidney Pestka, Roche In- stitute of Molecular Biology, Roche Research Center, Nutley, NJ 07110.
Received June 14, 1985.
The primer was an eicosadeoxynucleotide (20-
i Abbreviations: ddGTP, dideoxyguanidine triphosphate; ddATE dideoxyadenosine triphosphate; ddTTE dideoxythy- midine triphosphate; ddCTP, dideoxycytidine triphosphate; dGTP, deoxyguanidine triphosphate; dATE deoxyadenosine triphosphate; dTTE thymidine triphosphate; dCTE deoxycy- tidine triphosphate; ddA, ddC, ddG, and ddT, dideoxynucleo- side mixtures for termination at A, C, G, and T residues, re- spectively.
© 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York, NY 10017
0735-0651/85/$03.30

84 - j ----- 1985, G e n e Ana l Techn 2 : 8 3 - 8 8
__--i met), 5' CTCATGATTTCTGCTCTGAC 3', lo- cated in the conserved region of Hu-IFN-c~ cor- responding to amino acids 144-150. It was pre- pared by the phosphoramidite method (8,9) and was purified by high-performance liquid chroma- tography (HPLC).
Preparation of 5'-End Labeled Primer To prepare the labeled primer, 40 ~1 of [y-32p]ATP was dried in a 0.5-ml Eppendorf tube and dis- solved with 4 ~1 of Buffer K; then 14 ~1 of primer (28 pmol) and 2 ~i (10 units) of polynucleotide kinase were added. The mixture was incubated at 37°C for 15 min. Then, an additional 2 ~1 of polynucleotide kinase were added and incubated for a further 15 rain. With the use of small vol- umes and by the addition of kinase twice, more efficient labeling of primer was obtained. The reaction was stopped by addition of 80 ~xl of Buffer K, 50 pA of phenol saturated with the same buffer, and shaking. After centrifugation, the upper aqueous layer was collected and extracted twice with chloroform. To precipitate the DNA, 0.1 volume of 5 M NaCI and 3 volumes of 95% ethanol were added and kept at -20°C for 1-2 hr (6). The pellet was collected by centrifugation and dis- solved in 52 Isl water (0.4-0.5 pmol primer/~tl).
Preparation of DNA Template Bacteriophage h clones were prepared as de- scribed (10) by amplification of the phage from a single plaque representing one leukocyte inter- feron gene from a human genomic library in phage
(11) and then purified by CsCI centrifugation. The pure bacteriophage band was collected and dialyzed for 1 hr twice against 1000 volumes of Solution A (0.05 M Tris • C1, pH 8.0, 0.01 M NaCI, 0.01 M MgCI2). The bacteriophage was then di- gested with proteinase K (final concentration 50 ~g/ml) in 20 mM EDTA/0.5% SDS solution at 65°C for 30 rain. An additional equivalent amount of proteinase K was added to the above incubation mixture for a further 30 rain digestion at 65°C. The reaction was stopped by addition of an equal volume of phenol. DNA was extracted with phenol/chloroform and precipitated after addition of 0.1 volume of 0.3 M sodium acetate (pH 5.2) and 2 volumes of 95% ethanol.
Dideoxy Sequencing Procedure Annealing of primer and template. In Eppendorf
tubes (0.5 ml) a solution containing 0.8-1.2 pmol 32p-primer and 0.8-1.6 ~g h DNA from a single recombinant in a volume of 18 ~1 of H20 was heated to denature the DNA at 95°C (Dri-Block DB-3, Techne, Inc.) for 3 rain and immediately transferred to dry ice/ethanol. The use of poly- propylene Eppendorf tubes instead of glass cap- illaries is more convenient and omits the sealing step of the two previous reports (5,6). Two mi- croliters of sequencing Buffer S was then added to the solution, and the resulting mixture was in- cubated in a 50°C water bath for 30 min to anneal the primer and template. Then, 18 ~1 of 15% glyc- erol and 2 ILl (10 units) of Klenow fragment of DNA polymerase l was added, kept in a 50°C bath, and used in the next step within 4 rain.
Sequencing. Two microliters of each termi- nator mixture was dispensed into four separate tubes and kept in an ice bath before use. Imme- diately before use the first terminator mixture (for example, ddC) was placed into a 50°C bath for 1 min, then the second one into 50°C bath tbr 1 rain, etc. To each tube, 8 p,l of the above 32P-labeled p r ime r - t emp la t e -K lenow mixture was added successively at l-rain intervals and incubated for an additional 30 min at 50°C.
The reaction was stopped by adding 4 ~1 form- amide dye solution (0.1% xylene cyanol FF/0.1% bromophenol blue/10 mM EDTA-Na 2 in 95% deionized formamide), incubating at 95°C for 3 min, and cooling in dry ice; then 5-6 pA was loaded onto a 0.6-cm thick 40-cm long 8% poly- acrylamide sequencing gel. Electrophoresis was performed for 3-5 hr (usually 3 hr) at 1500-1800 v with the plate surface temperature kept at about 50°C by monitoring with an electronic thermom- eter. The gel was fixed for 15 rain in 10% acetic acid and 10% methanol, then dried (Slab Gel Drier Model SE1140, Hoeffer Scientific Instruments) and exposed to Kodak X-OMAT AR film without an intensifier screen at -20°C for 1-2 days or with a screen overnight.
R e s u l t s a n d D i s c u s s i o n
Importance of Temperature for Sequencing Reaction Wallace et al. (5) used room temperature for se- quencing on pBR322 dsDNA templates. Hong (6) found that 100 nucleotides from the primer site were unreadable when the sequencing reaction was carried out at room temperature, and he
© 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York, NY 1(1017
93 pER322 ! ]
a,r ,rc
f _ 1985, Gene Anal Techn 2:83-88 ~ . ~ _
f J
26 27
G A T C G A T C
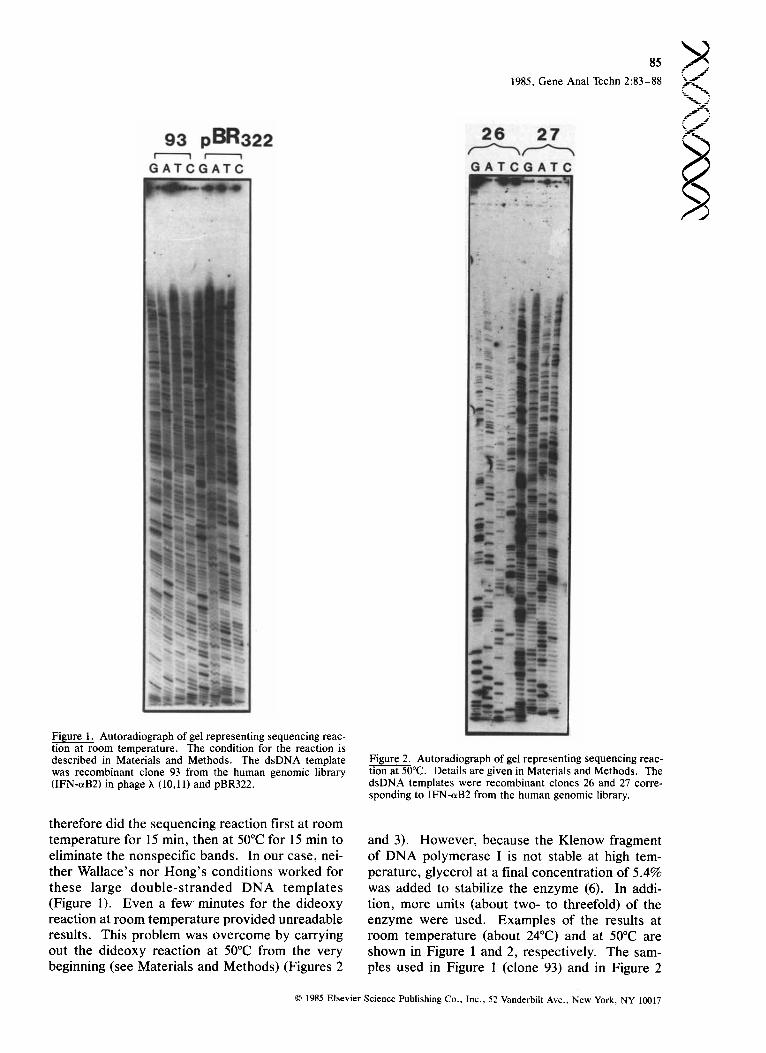
Figure 1. Autoradiograph of gel representing sequencing reac- tion at room temperature. The condition for the reaction is described in Materials and Methods. The dsDNA template was recombinant clone 93 from the human genomic library (IFN-~B2) in phage h (10,11) and pBR322.
Figure 2. Autoradiograph of gel representing sequencing reac- tion at 50°C. Details are given in Materials and Methods. The dsDNA templates were recombinant clones 26 and 27 corre- sponding to IFN-aB2 from the human genomic library.
therefore did the sequencing reaction first at room temperature for 15 rain, then at 50°C for 15 min to eliminate the nonspecific bands. In our case, nei- ther Wallace's nor Hong's conditions worked for these large double-stranded DNA templates (Figure 1). Even a few minutes for the dideoxy reaction at room temperature provided unreadable results. This problem was overcome by carrying out the dideoxy reaction at 50°C from the very beginning (see Materials and Methods) (Figures 2
and 3). However, because the Klenow fragment of DNA polymerase I is not stable at high tem- perature, glycerol at a final concentration of 5.4% was added to stabilize the enzyme (6). In addi- tion, more units (about two- to threefold) of the enzyme were used. Examples of the results at room temperature (about 24°C) and at 50°C are shown in Figure 1 and 2, respectively. The sam- ples used in Figure 1 (clone 93) and in Figure 2
© 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York, NY 10017
86 f J - _ _ j
j -
1985, Gene Anal Techn 2:83-88
H I
G A T C G A T C
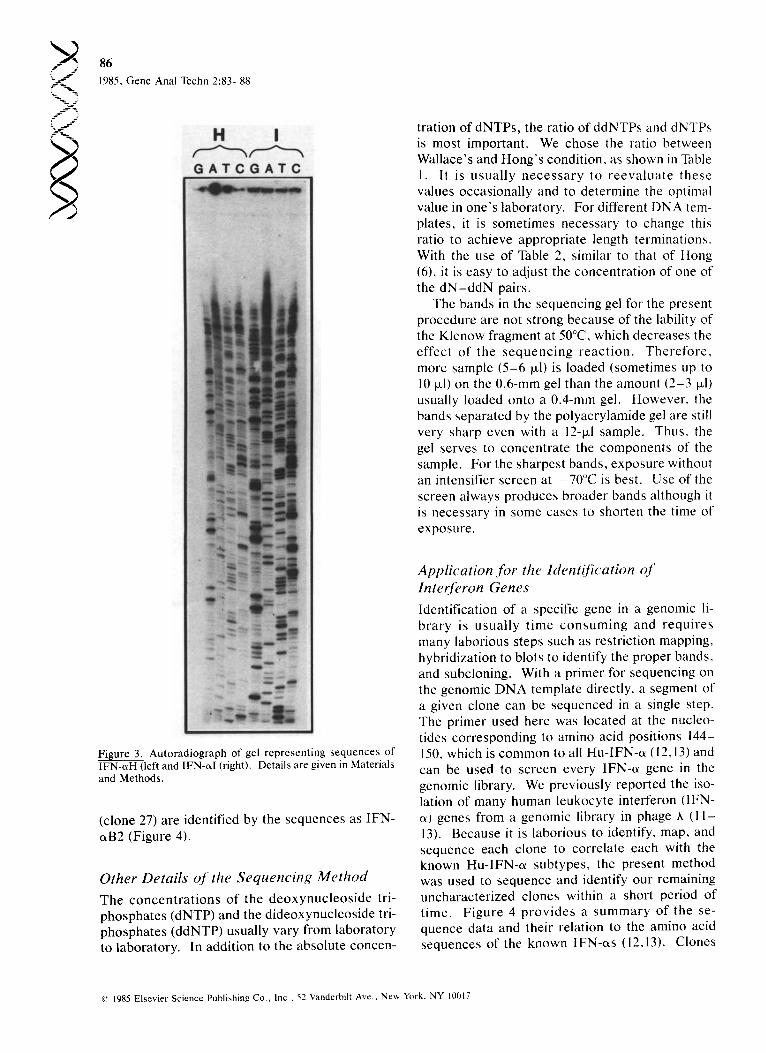
Figure 3. Au to r ad i og raph of gel r ep resen t ing s e q u e n c e s of IFN-~xH (left and IFN-~I (right). Details are given in Materials and Methods .
(clone 27) are identified by the sequences as IFN- eLB2 (Figure 4).
Other Details of the Sequencing Method The concen t ra t ions of the deoxynuc leos ide tri- phosphates (dNTP) and the dideoxynucleoside tri- phosphates (ddNTP) usually vary from laboratory to laboratory. In addition to the absolute concen-
tration of dNTPs, the ratio of ddNTP8 and dNTPs is most important. We chose the ratio between Wallace's and Hong 's condition, as shown in Table 1. It is usual ly n e c e s s a r y to reeva lua te these values occasionally and to determine the optimal value in one 's laboratory. For different DNA tem- plates, it is sometimes necessary to change this ratio to achieve appropriate length terminations. With the use of Table 2, similar to that of Hong (6), it is easy to adjust the concentrat ion of one of the d N - d d N pairs.
The bands in the sequencing gel for the present procedure are not strong because of the lability of the Klenow fragment at 50°C, which decreases the ef fec t of the sequenc ing reac t ion . Th e re fo r e , more sample (5 -6 ~1) is loaded (sometimes up to 10 p.l) on the 0.6-ram gel than the amount (2-3 ~l) usually loaded onto a 0.4-ram gel. However , the bands separated by the polyacrylamide gel are still very sharp even with a 12-~1 sample. Thus, the gel serves to concentrate the components of the sample. For the sharpest bands, exposure without an intensifier screen at -70°C is best. Use of the screen always produces broader bands although it is necessary in some cases to shorten the time of exposure.
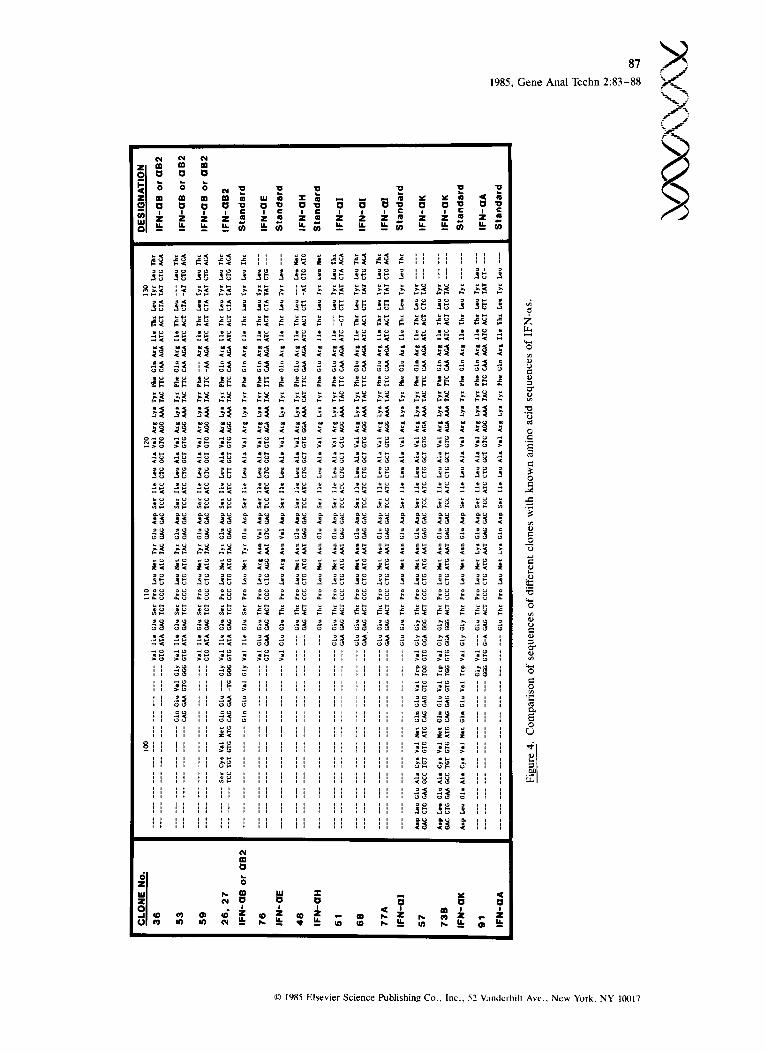
Application for the Identification of Interferon Genes Identification of a specific gene in a genomic li- b ra ry is usual ly t ime consuming and requi res many laborious steps such as restriction mapping, hybridization to blots to identify the proper bands, and subcloning. With a primer for sequencing on the genomic DNA template directly, a segment of a given clone can be sequenced in a single step. The primer used here was located at the nucleo- tides corresponding to amino acid positions 144- 150, which is common to all Hu-IFN-o~ (12,13) and can be used to screen every IFN-~ gene in the genomic library. We previously reported the iso- lation of many human leukocyte interferon (IFN- cx) genes from a genomic library in phage ;~ (11- 13). Because it is laborious to identify, map, and sequence each clone to correlate each with the known Hu-IFN-~ subtypes, the present method was used to sequence and identify our remaining uncharacterized clones within a short period of t ime. F igure 4 p rov ides a su m m ary of the se- quence data and their relation to the amino acid sequences of the known IFN-cxs (12,13). Clones
© 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York. NY 10017
1985, G e n e A n a l T e c h n 2 : 8 3 - 8 8
I_~[ " . ~ , ° ° .
" " 'i i , o , , , ~ ~ , _ .
I~1 ~ ~ - ~
ta i ~ ,
o . . . . . ~, . . . . . . . . ~ o o ~ . . . . . . . . ~ o o ~ o
i i ~ i i i i i i i , . . . . . . , , , , , o ~ s
I ' ; ~ I I I
~ , ~ , I I I I 1 I I I I I , , , , , , I 1 1 1 I I I I I
' ' ' ' ' I g 1 i i ~ I I I ' 1
m
Z
._= E
O e~
e .
0
e -
g
E 0
(..)
-%_ -
f ! J
© 1985 Elsevier Science Publishing Co., Inc., 52 Vandcrbil! Ave.. New York, NY 101117
- ._-
-5- __7 :.-_j
88
1985, Gene Anal Techn 2:83-88
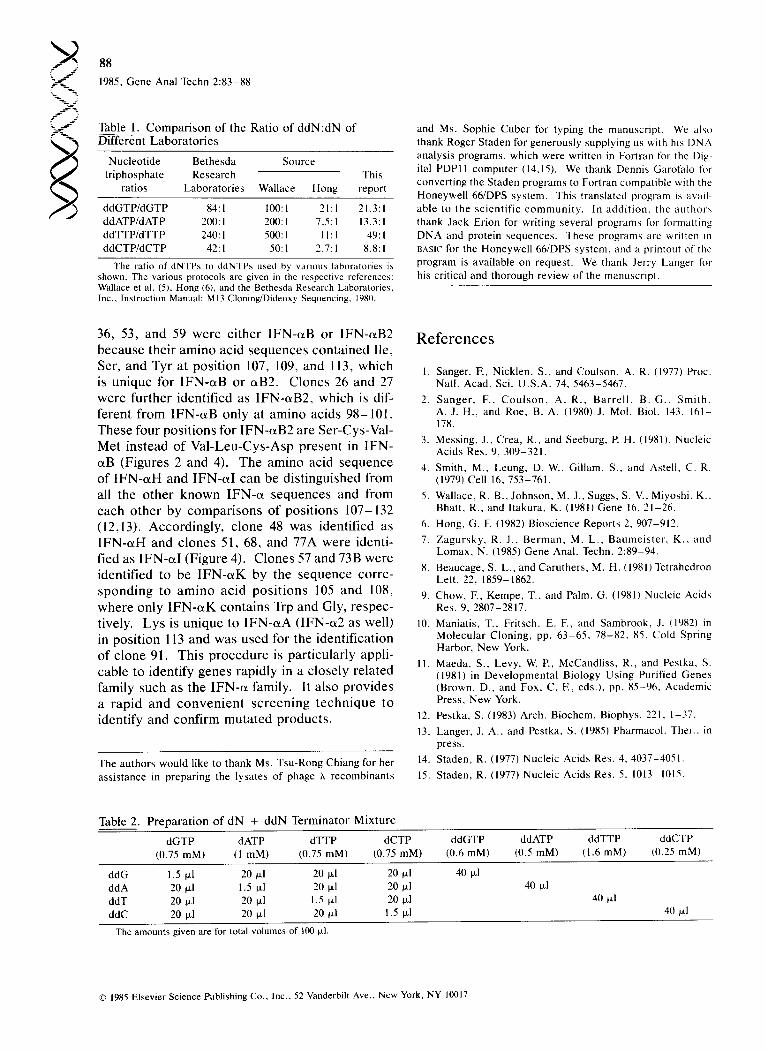
Table 1. Comparison of the Ratio of ddN:dN of ~ n t Laboratories
Nucleotide Bethesda Source triphosphate Research This
ratios Laboratories Wallace Hong report
ddGTP/dGTP 84:1 100:1 21:1 21.3:1 ddATP/dATP 200:1 200:1 7.5:1 13.3:1 ddTTP/dTTP 240: I 500: I 11 : 1 49: I ddCTP/dCTP 42:1 50:1 2.7:1 8.8: I
The ratio of dNTPs to ddNTPs used by various laboratories is shown. The various protocols are given in the respective references: Wallace et al. (5), Hong (6), and the Bethesda Research Laboratories, Inc., Instruction Manual: M13 Cloning/Dideoxy Sequencing, 1980.
and Ms. Sophie Cuber for typing the manuscript. We also thank Roger Staden for generously supplying us with his DNA analysis programs, which were written in Fortran for the Dig- ital PDPll computer (14,15). We thank Dennis Garofalo for converting the Staden programs to Fortran compatible with the Honeywell 66/DPS system. This translated program is avail- able to the scientific community, lnaddi t ion , theau thors thank Jack Erion for writing several programs for formatting DNA and protein sequences. These programs are written in UASJC for the Honeywell 66/DPS system, and a printout of the program is available on request. We thank Jerry Langer for his critical and thorough review of the manuscript.
36, 53, and 59 were either IFN-aB or IFN-aB2 because their amino acid sequences contained Ile, Ser, and Tyr at position 107, 109, and 113, which is unique for IFN-aB or c~B2. Clones 26 and 27 were further identified as IFN-aB2, which is dif- ferent from IFN-aB only at amino acids 98-101. These four positions for IFN-aB2 are Ser-Cys-Val- Met instead of Val-Leu-Cys-Asp present in IFN- aB (Figures 2 and 4). The amino acid sequence of IFN-aH and IFN-M can be distinguished from all the other known IFN-a sequences and from each other by comparisons of positions 107-132 (12,13). Accordingly, clone 48 was identified as IFN-oLH and clones 51, 68, and 77A were identi- fied as IFN-aI (Figure 4). Clones 57 and 73B were identified to be IFN-aK by the sequence corre- sponding to amino acid positions 105 and 108, where only IFN-o~K contains Trp and Gly, respec- tively. Lys is unique to IFN-aA (IFN-a2 as well) in position 113 and was used for the identification of clone 91. This procedure is particularly appli- cable to identify genes rapidly in a closely related family such as the IFN-a family. It also provides a rapid and convenient screening technique to identify and confirm mutated products.
The authors would like to thank Ms. Tsu-Rong Chiang for her assistance in preparing the lysates of phage h recombinants
References
1. Sanger, E, Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. Acad. Sci. U.S.A. 74, 5463-5467.
2. Sanger, F., Coulson, A. R., Barrell, B. G., Smith, A. J. H., and Roe, B. A. (1980) J. Mol. Biol. 143, 161- 178.
3. Messing, J., Crea, R., and Seeburg, P. H. (1981). Nucleic Acids Res. 9, 309-321.
4. Smith, M., Leung, D. W., Gillam, S.. and Astelt, C. R. (1979) Cell 16, 753-761.
5. Wallace, R. B., Johnson, M. J., Suggs, S. V., Miyoshi, K., Bhatt, R., and Itakura, K. (1981) Gene 16, 21-26.
6. Hong, G. E (1982) Bioscience Reports 2, 907-912.
7. Zagursky, R. J., Berman, M. L., Baumeister, K., and Lomax, N. (1985) Gene Anal. Techn. 2:89-94.
8. Beaucage, S. L., and Caruthers, M. H. (1981) Tetrahedron Len. 22, 1859-1862.
9. Chow, E, Kempe, T., and Palm, G. (1981) Nucleic Acids Res. 9, 2807-2817.
10. Maniatis, T., Fritsch, E. E, and Sambrook, J. (1982) in Molecular Cloning, pp. 63-65, 78-82, 85, Cold Spring Harbor, New York.
11. Maeda, S., Levy, W. P., McCandliss, R., and Pestka, S. (1981) in Developmental Biology Using Purified Genes (Brown, D., and Fox, C. E, eds.), pp. 85-96, Academic Press, New York.
12. Pestka, S. (1983) Arch. Biochem. Biophys. 221, 1-37.
13. Langer, J. A., and Pestka. S. (1985) Pharmacol. Ther., in press.
14. Staden, R. (1977) Nucleic Acids Res. 4, 4037-4051.
15. Staden, R. (1977) Nucleic Acids Res. 5, 1013-1015.
Table 2. Preparation of dN + ddN Terminator Mixture
dGTP dATP dTTP dCTP (0.75 mM) (1 mM) (0.75 mM) (0.75 mM)
ddGTP ddATP ddTTP ddCTP (0.6 mM) (0.5 mM) (1.6 mM) (0.25 mM)
ddG 1.5 ~1 20 ~1 20 ~1 20 gl ddA 20 ~1 1.5 ~1 20 ~1 20 gl ddT 20 ~1 20 ~1 1.5 ~1 20 ~1 ddC 20 ~1 20 ~1 20 ~1 1.5 ~1
40 ~1 40 p,l
40 Ixl 40 ~1
The amounts given are for total volumes of 100 ixl.
© 1985 Elsevier Science Publishing Co., Inc., 52 Vanderbilt Ave., New York, NY 10017