
Χέμινγουαιη Έρνεστ (Hemingway Ernest)-Για ποιον χτυπά η καμπάνα-Γράμματα (1992)

3685Journal of Cell Science 108, 3685-3694 (1995)Printed in Great Britain © The Company of Biologists Limited 1995JCS3270
Matrigel induces thymosin β4 gene in differentiating endothelial cells
D. S. Grant1,*, J. L. Kinsella2, M. C. Kibbey1, S. LaFlamme1, P. D. Burbelo1, A. L. Goldstein2
and H. K. Kleinman1
1Laboratory of Developmental Biology, National Institute of Dental Research, National Institutes of Health, Bldg 30, Rm 430,Bethesda, MD 20892, USA2Laboratory of Cardiovascular Science, Gerontology Reseach Center, NIA, Baltimore, MD, USA 3Department of Biochemistry and Molecular Biology, The George Washington University Medical School, Washington, DC, USA
*Author for correspondence
We performed differential cDNA hybridization using RNAfrom endothelial cells cultured for 4 hours on either plasticor basement membrane matrix (Matrigel), and identifiedearly genes induced during the morphological differen-tiation into capillary-like tubes. The mRNA for one clone,thymosin β4, was increased 5-fold. Immunostaininglocalized thymosin β4 in vivo in both growing and maturevessels as well as in other tissues. Endothelial cells trans-fected with thymosin β4 showed an increased rate of
attachment and spreading on matrix components, and anaccelerated rate of tube formation on Matrigel. Anantisense oligo to thymosin β4 inhibited tube formation onMatrigel. The results suggest that thymosin β4 is inducedand likely involved in differentiating endothelial cells.Thymosin β4 may play a role in vessel formation in vivo.
Key words: neovascularization, endothelium, matrix, differentiation,Matrigel, subtraction library, thymosin beta 4, basement membrane
SUMMARY
INTRODUCTION
The maintenance and integrity of the blood vessel wallprimarily depends on the stability of the endothelial cells andassociated vascular cells. Although both small and large bloodvessels are quiescent in the adult, they can readily form newvessels in response to changes such as injury or tumor growth(Folkman, 1992; Folkman and Hanahan, 1991; Folkman andIngber, 1992; Folkman and Shing, 1992; Paku and Paweletz,1991). The molecular events which occur in stable vessels orduring new blood vessel formation (angiogenesis) are stillunclear. Several in vivo and in vitro models have beendeveloped that mimic at least some aspects of the angiogenicprocess, and these models provide insight into the steps andinitiating factors involved in stimulating endothelial cells toform and maintain structurally intact vessels (Ingber et al.,1987; Jaffe et al., 1973; Kramer and Fuh, 1985; Madri andPratt, 1986; Nicosia et al., 1984). For example, Maciag andcoworkers (Maciag, 1990) have shown that when confluent,cultured endothelial cells are treated with phorbol estersapproximately 50% of the cells form tube networks. Using thismodel of endothelial cell differentiation with subtractivehybridization, a novel protein, EDG-1, was identified andshown to increase during tube formation (Hla and Maciag,1990a,b; Maciag, 1990).
Large and small blood vessels are composed of an innerendothelial cell layer, which is in direct contact with an asso-ciated basement membrane. A major factor regulating endo-thelial cell behavior is the basement membrane, whichprovides many signals and maintains the endothelium in a dif-
ferentiated state in vivo (Ingber and Folkman, 1989; Kramerand Fuh, 1985; Kubota et al., 1988; Madri et al., 1980) Abasement membrane substratum, Matrigel, has been used invitro with endothelial cells and found to induce rapid formationof a capillary-like tube network (Grant et al., 1992; Kubota etal., 1988). When the cells are seeded onto Matrigel, theyspread, attach and migrate across the surface over a 2 hourperiod, and then between 4-6 hours the cells align and formlateral associations which progress to the formation of tubesover the next 10 hours (Grant et al., 1990, 1991). During thismorphologic differentiation, cell division is greatly reduced.This process is blocked by cycloheximide suggesting thatprotein synthesis is important (Grant et al., 1991). While thismodel does not mimic all the events that occur during vesselformation, it provides a powerful tool for the examination ofcertain molecular and biochemical events that transpire duringendothelial cell morphogenesis and differentiation. In thisstudy, we have investigated the early genes which are inducedduring the endothelial cell differentiation on Matrigel.
Using a differential cDNA cloning technique performedwith the RNA from endothelial cells cultured for 4 hours oneither plastic or Matrigel, we have identified several geneswhich are induced by Matrigel. One of these genes encoded asmall polypeptide (4.9 kDa) first identified and isolated fromthe calf thymus, called thymosin β4 (Badamchian et al., 1988;Gomez-Marquez et al., 1989; Low and Goldstein, 1984, 1985;Low et al., 1990). Thymosin β4 contains 43 amino acids andexhibits important activities in the regulation and differen-tiation of thymus-dependent lymphocytes (Low and Goldstein,1984, 1985). For example, previous studies have demonstrated

3686 D. S. Grant and others
that thymosin β4 is effective in partially or fully reconstitutingimmune functions in thymic-deprived or immunodeprivedanimals (Low and Goldstein, 1981). The sequence of thisprotein is well conserved since it is over 95% homologous inhuman, mouse, rat and cow (Hall, 1991; Low and Goldstein,1985). In addition, this protein is ubiquitously expressed inmany cells and tissues (Condon and Hall, 1992; Lin andMorrison-Bogorad, 1990). Thymosin β4 is linked to the differ-entiation of not only hematopoietic cells but also to cells indeveloping human embryonic kidney (Hall, 1991). Thymosinβ4 is also increased in the serum of patients over 60, innewborns, and in patients suffering from AIDS, hepatitis, andinflammatory bowel disease (Naylor et al., 1986). This smallpolypeptide is differentially expressed in the developing braintissues during embryogenesis of the rat and is present in loweramounts in embryonic lung, kidney, spleen, adrenal glands,heart and liver (Lin and Morrison-Bogorad, 1990). Thymosinβ4 expression is also directly associated with cell cycle regu-lation (Schöbitz et al., 1991b). Nothing is known about theexpression or the regulation of thymosin β4 in endothelialcells. In this report, we provide evidence that this polypeptideis involved in the regulation of endothelial cell differentiation.
MATERIALS AND METHODS
CellsHuman umbilical vein endothelial cells (HUVEC) were isolated fromfreshly delivered cords (Grant et al., 1991). The cells were grown andpassaged (2 to 7 times) in Medium 199 (Gibco Inc.) containing 20%calf serum (Hyclone), 200 µg/ml ECGS (Collaborative Res. Inc.Bedford MA), 50 U/ml sodium heparin (Sigma), 100 µM penicillin-streptomycin (Gibco, BRL, Gaithersburg, MD), fungizone (2.7 µMamphotericin B), and 25 µg/ml gentamicin. Matrigel (at 15 mg/ml),collagen IV and laminin are routinely prepared in the laboratory(Timpl et al., 1979; Kleinman et al., 1986). Cells were grown to con-fluency on either plastic or glass coverslips for light microscopy. Lightmicrographs were taken with a Zeiss IM microscope equipped witheither Hoffman modulation contrast or phase contrast.
Preparation of RNA from HUVEC on Matrigel and plasticMatrigel at 15 mg/ml (Kleinman et al., 1986), was used to coat 40150-mm Nunc plates (approximately 1 ml of Matrigel per plate, athickness of approximately 0.3 mm) and was allowed to polymerizeat 37°C for a minimum of 30 minutes. One hundred and sixty controlplates lacking Matrigel were incubated at the same time at 37°C with5 ml of complete Medium 199. Trypsin-released passage fourHUVEC were plated at a density of 1 million cells per 150 mm plate(30% confluence) and incubated at 37°C, with 5% CO2/95% air for 4hours, at which time the cells on Matrigel were organizing intonetworks (see Fig. 1). The medium from all plates was aspirated andthen the plates were washed with Dulbecco’s phosphate-bufferedsaline (DPBS). The cells were lysed with 4 M guanidine isothio-cyanate, 25 mM sodium acetate, and 100 mM β-mercaptoethanol, andremoved after either gentle rocking (cells on Matrigel) or scraping(cells on plastic). Total cellular RNA was obtained by centrifugationover a CsCl2 cushion (Sambrook et al., 1989). Five milligrams of totalRNA were obtained from the cells grown on plastic for 4 hours. ThemRNA was isolated from total RNA of cells incubated on Matrigelby the use of an oligo dT Sepharose 4B column (Poly-Quick Kit,Stratagene, La Jolla, CA). A cDNA probe was generated from thisMatrigel-induced mRNA using Moloney murine leukemia (M-MuLV) reverse transcriptase (Boehringer Mannheim) for screeningthe library.
Library construction and identification of Matrigel inducedgenesA Matrigel-induced cDNA library was constructed using the Uni ZAPXR unidirectional cDNA synthesis Kit (Stratagene, La Jolla, CA)from the purified mRNA (as prepared above) and packaged (GigapackPlus 7, Stratagene, La Jolla, CA) as per the manufacturer’s pro-cedures. Following amplification, the cDNA library was screened bysubtractive hybridization using XL1 Blue cells (Stratagene, La Jolla,CA). Double lifts of the plaques were transferred to nitrocellulosemembranes (Schleicher and Schuell, Keene, NH) which were subse-quently denatured, neutralized, rinsed and crosslinked. For cDNAscreening, poly(A) RNA was isolated from HUVEC that werecultured on either plastic or Matrigel for 4 hours. Briefly, differentialplaque hybridization was performed using 32P-labeled probes synthe-sized by reverse transcriptase reaction from mRNA obtained fromcells cultured on Matrigel and plastic. Excess total RNA fromHUVEC grown on plastic was included with the Matrigel-inducedprobe during hybridization to compete out common genes. The dif-ferential hybridization was carried out using 5 mg of total RNA fromHUVEC on plastic with 0.5 µg of labeled Matrigel-induced cDNA,incubated overnight at 65°C in 5× Denhardt’s solution (0.1% (w/v)Ficoll, 1.0% (w/v) polyvinylpyrrolidone, 0.1% bovine serumalbumen), 6× SSC (0.9 M NaCl, 0.09 M sodium citrate), 0.5% SDS,and 15 µg/µl salmon sperm DNA. The filters were washed with 2×SSC + 0.1% SDS twice, then 0.5× SSC + 0.5% SDS three times at65°C. After exposure to Kodak XAR film, positive plaques were iden-tified and selected for further screening. Approximately 106 plaqueswere examined. Identified induced clones were rescreened threetimes. Several clones were found to be induced on Matrigel and theseinserts were rescued using R408 helper phage and XL1-Blue cells(Stratagene, La Jolla, CA).
Selected colonies were then streaked onto new LB/amp plates forfurther investigation and sequencing. Seventeen clones were identi-fied. Plasmid DNA was purified using a Qiagen (Chatsworth, CA)plasmid preparation kit. The double stranded cDNA inserts weresequenced from the T3 or T7 primer site in Bluescript with SequenaseVersion 1.0 T7 DNA polymerase kit (United States Biochemical,Cleveland, Ohio). In some cases, the sequencing reaction was doneusing Applied Biosystems Inc. AmpliTaqR DNA polymerase and a6% sequencing gel was then run on the automated sequencer. In eachcase, the sequence of the insert of a particular clone was compared(using the VAX, Fasta search; Pearson and Lipman, 1988) to knownsequences in the database.
Northern blot analysis HUVEC, at 70% confluence, were lifted and replated in fresh mediumon 150 mm Nunc tissue culture plates or on Matrigel-coated plates.The plates were incubated for 1, 2, 4, 6, or 20 hours at 37°C. TotalRNA was isolated as described above (Sambrook et al., 1989). A totalof 10 µg of RNA was loaded per lane in a formaldehyde-1% agarosegel, electrophoresed, and blotted onto Nytran (Schleicher and Schuell,Keene, NH). The integrity and equal loading of each sample wereconfirmed by staining the gel with ethidium bromide.
The cDNA inserts from the subtracted clones were gel purified afterelectrophoresis using a QIAEX kit for nucleic acid extraction fromgels (QIAGEN Inc., Chatsworth, CA), then 25-50 ng of the purifiedcDNA was labeled with [32P]dCTP using a random prime kit(Boehringer Mannheim, Indianapolis, IN) and used to probe thefilters. The Nytran filters with RNA from HUVEC were preincubatedin prehybridzation buffer (2× SSC, 2× Denhardt’s, 0.5% SDS, and 1mg/ml salmon sperm DNA) for 2 hours and then incubated with thedenatured labeled probe overnight. The following day, the filters werewashed with 2× SSC + 0.1% SDS, followed by 1× SSC + 0.1% SDS,and then 0.1× SSC + 0.1% SDS at 20°C or 37°C. The filters were thenwrapped in plastic and exposed to Kodak XAR film. The relative inte-grated density of RNA expression was done in the following way. Theimages on X-ray films were transferred to a Power Macintosh 7100/66
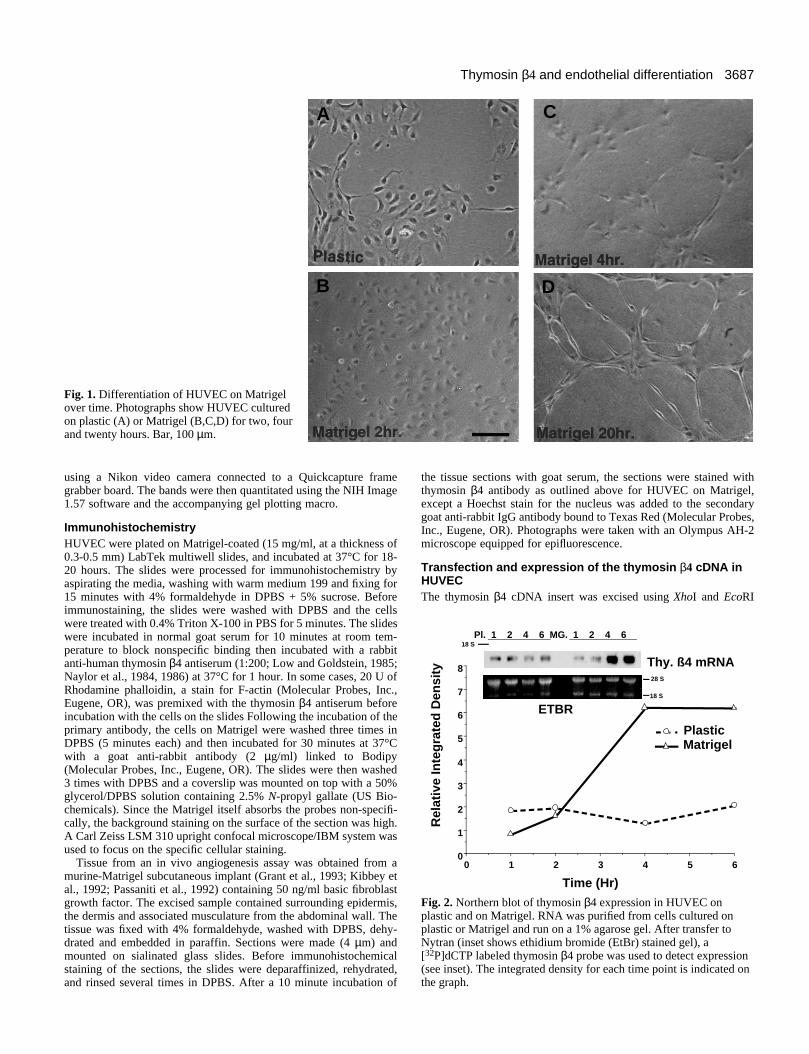
3687Thymosin β4 and endothelial differentiation
Fig. 1. Differentiation of HUVEC on Matrigelover time. Photographs show HUVEC culturedon plastic (A) or Matrigel (B,C,D) for two, fourand twenty hours. Bar, 100 µm.
A
B
C
D
65432100
1
2
3
4
5
6
7
8
PlasticMatrigel
Time (Hr)
Rel
ativ
e In
teg
rate
d D
ensi
ty
Thy. ß4 mRNA
Pl. 1 2 4 6 MG. 1 2 4 6
ETBR
18 S
18 S
28 S
Fig. 2. Northern blot of thymosin β4 expression in HUVEC onplastic and on Matrigel. RNA was purified from cells cultured onplastic or Matrigel and run on a 1% agarose gel. After transfer toNytran (inset shows ethidium bromide (EtBr) stained gel), a[32P]dCTP labeled thymosin β4 probe was used to detect expression(see inset). The integrated density for each time point is indicated onthe graph.
using a Nikon video camera connected to a Quickcapture framegrabber board. The bands were then quantitated using the NIH Image1.57 software and the accompanying gel plotting macro.
Immunohistochemistry HUVEC were plated on Matrigel-coated (15 mg/ml, at a thickness of0.3-0.5 mm) LabTek multiwell slides, and incubated at 37°C for 18-20 hours. The slides were processed for immunohistochemistry byaspirating the media, washing with warm medium 199 and fixing for15 minutes with 4% formaldehyde in DPBS + 5% sucrose. Beforeimmunostaining, the slides were washed with DPBS and the cellswere treated with 0.4% Triton X-100 in PBS for 5 minutes. The slideswere incubated in normal goat serum for 10 minutes at room tem-perature to block nonspecific binding then incubated with a rabbitanti-human thymosin β4 antiserum (1:200; Low and Goldstein, 1985;Naylor et al., 1984, 1986) at 37°C for 1 hour. In some cases, 20 U ofRhodamine phalloidin, a stain for F-actin (Molecular Probes, Inc.,Eugene, OR), was premixed with the thymosin β4 antiserum beforeincubation with the cells on the slides Following the incubation of theprimary antibody, the cells on Matrigel were washed three times inDPBS (5 minutes each) and then incubated for 30 minutes at 37°Cwith a goat anti-rabbit antibody (2 µg/ml) linked to Bodipy(Molecular Probes, Inc., Eugene, OR). The slides were then washed3 times with DPBS and a coverslip was mounted on top with a 50%glycerol/DPBS solution containing 2.5% N-propyl gallate (US Bio-chemicals). Since the Matrigel itself absorbs the probes non-specifi-cally, the background staining on the surface of the section was high.A Carl Zeiss LSM 310 upright confocal microscope/IBM system wasused to focus on the specific cellular staining.
Tissue from an in vivo angiogenesis assay was obtained from amurine-Matrigel subcutaneous implant (Grant et al., 1993; Kibbey etal., 1992; Passaniti et al., 1992) containing 50 ng/ml basic fibroblastgrowth factor. The excised sample contained surrounding epidermis,the dermis and associated musculature from the abdominal wall. Thetissue was fixed with 4% formaldehyde, washed with DPBS, dehy-drated and embedded in paraffin. Sections were made (4 µm) andmounted on sialinated glass slides. Before immunohistochemicalstaining of the sections, the slides were deparaffinized, rehydrated,and rinsed several times in DPBS. After a 10 minute incubation of
the tissue sections with goat serum, the sections were stained withthymosin β4 antibody as outlined above for HUVEC on Matrigel,except a Hoechst stain for the nucleus was added to the secondarygoat anti-rabbit IgG antibody bound to Texas Red (Molecular Probes,Inc., Eugene, OR). Photographs were taken with an Olympus AH-2microscope equipped for epifluorescence.
Transfection and expression of the thymosin β4 cDNA inHUVECThe thymosin β4 cDNA insert was excised using XhoI and EcoRI
3688 D. S. Grant and others
restriction enzymes, and subcloned by ligation into the pcDNA 1eukaryotic expression vector, containing a CMV promotor (InVitro-gen, San Diego, CA). After a plasmid MagicTMminiprep (PromegaCorp., Madison, WI), 10 µg of the plasmid, possessing the thymosinβ4 insert, was used to transiently transfect passage 4-6 HUVEC cells.Briefly, 1.1×106 cells were synchronized by the addition of 2 Mthymidine (Sigma; 174 µl per 10 ml of media) for 16-18 hours. Whenthe cells began to divide again, the transfection was done by electro-poration at 170 mV and a capacitance of 960 µF with a Gene Pulser(Bio-Rad Laboratories, Melville, NY) (Giordano et al., 1991;Goldstein et al., 1989). As a control, HUVEC were mock transfectedwith the pcDNA vector alone without the insert. Following the trans-fection, the cells were plated in normal HUVEC medium containing0.005 M sodium butyrate (Sigma Biochemicals, St Louis MO) for 14-16 hours. Butyrate treatment alone had no effect on either endogenousthymosin β4 expression or tube formation as evidenced by the mocktransfection control. Cells were incubated at 37°C for 24 and 48 hours
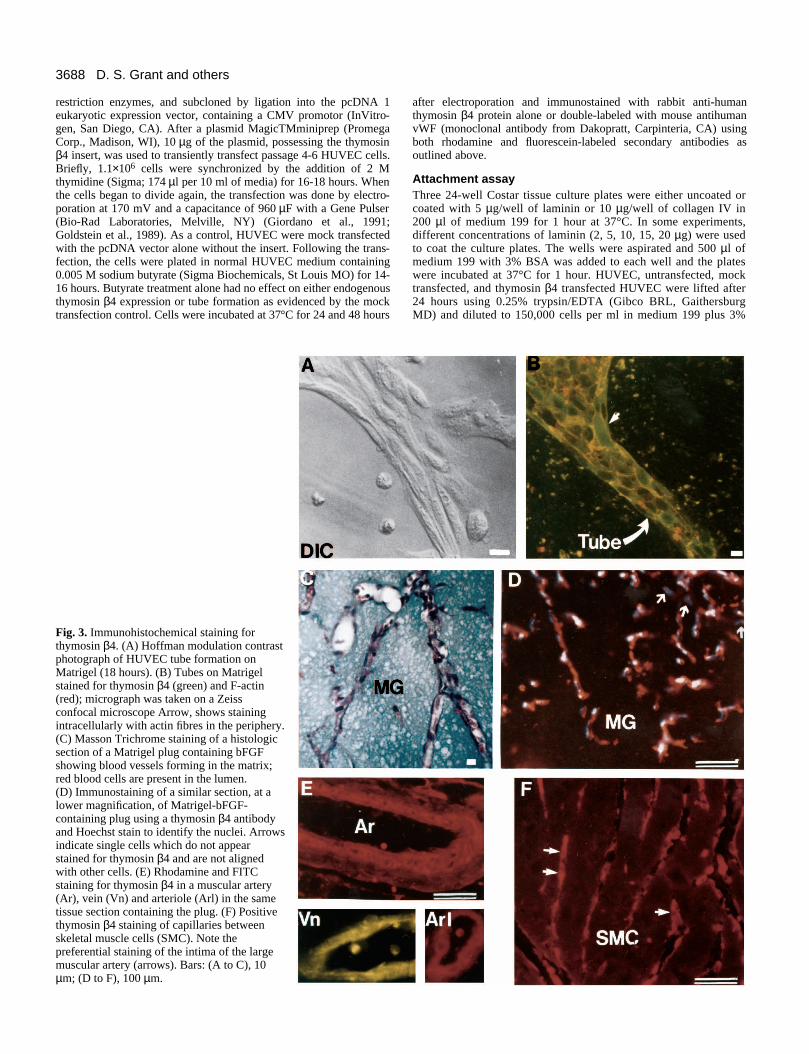
Fig. 3. Immunohistochemical staining forthymosin β4. (A) Hoffman modulation contrastphotograph of HUVEC tube formation onMatrigel (18 hours). (B) Tubes on Matrigelstained for thymosin β4 (green) and F-actin(red); micrograph was taken on a Zeissconfocal microscope Arrow, shows stainingintracellularly with actin fibres in the periphery.(C) Masson Trichrome staining of a histologicsection of a Matrigel plug containing bFGFshowing blood vessels forming in the matrix;red blood cells are present in the lumen.(D) Immunostaining of a similar section, at alower magnification, of Matrigel-bFGF-containing plug using a thymosin β4 antibodyand Hoechst stain to identify the nuclei. Arrowsindicate single cells which do not appearstained for thymosin β4 and are not alignedwith other cells. (E) Rhodamine and FITCstaining for thymosin β4 in a muscular artery(Ar), vein (Vn) and arteriole (Arl) in the sametissue section containing the plug. (F) Positivethymosin β4 staining of capillaries betweenskeletal muscle cells (SMC). Note thepreferential staining of the intima of the largemuscular artery (arrows). Bars: (A to C), 10µm; (D to F), 100 µm.
after electroporation and immunostained with rabbit anti-humanthymosin β4 protein alone or double-labeled with mouse antihumanvWF (monoclonal antibody from Dakopratt, Carpinteria, CA) usingboth rhodamine and fluorescein-labeled secondary antibodies asoutlined above.
Attachment assay Three 24-well Costar tissue culture plates were either uncoated orcoated with 5 µg/well of laminin or 10 µg/well of collagen IV in200 µl of medium 199 for 1 hour at 37°C. In some experiments,different concentrations of laminin (2, 5, 10, 15, 20 µg) were usedto coat the culture plates. The wells were aspirated and 500 µl ofmedium 199 with 3% BSA was added to each well and the plateswere incubated at 37°C for 1 hour. HUVEC, untransfected, mocktransfected, and thymosin β4 transfected HUVEC were lifted after24 hours using 0.25% trypsin/EDTA (Gibco BRL, GaithersburgMD) and diluted to 150,000 cells per ml in medium 199 plus 3%
3689Thymosin β4 and endothelial differentiation
BSA. In the attachment assay, the cells (75,000) were added to thewell and were incubated for 30 minutes at 37°C and then each wellwas washed once with DPBS. The attached cells were fixed andstained with the DiffQuick system (Baxter Scientific) and examinedwith an Olympus inverted microscope. For quantitation of cellattachment, the Optomax image analysis program attached to theOlympus microscope was used (Grant et al., 1992). Each conditionwas carried out in six replicates, ten random fields of view werequantitated per well, and the experiment was repeated at least threetimes for each condition.
Formation of capillary-like structures on MatrigelFour-well Nunc plates (16 mm well) were coated with 250 µl ofMatrigel and 40,000 mock transfected or thymosin β4 transfectedHUVEC were added to each well (Grant et al., 1990). The plates wereincubated for 24 hours, and, during this time, pictures were taken at2, 4 and 6 hours.
Effect of sense and antisense oligos on capillary-like tubeformation on MatrigelTwo 15mer phosphorothioated oligonucleotide sequences wereprepared by SyntheCell corporation (Rockville, MD): sense, 5′-TCTGACAAACCCGGGAT-3′, and antisense, 5′-ATCGG-GTTTGTCAGA-3′, which correspond to the first 15 nucleotides atthe 5′ initiation site of thymosin β4 mRNA. The oligos were purified(verified on an electrophoretic gel), dried, and rehydrated before use.The Matrigel-coated plates containing HUVEC were incubated at37°C for 18 hours in the presence of the oligos. The cells were fixedand stained (DiffQuick), and tube area was measured on theOptomax image analyzer. Data are expressed as the percentage tubearea in experimental wells compared to control.
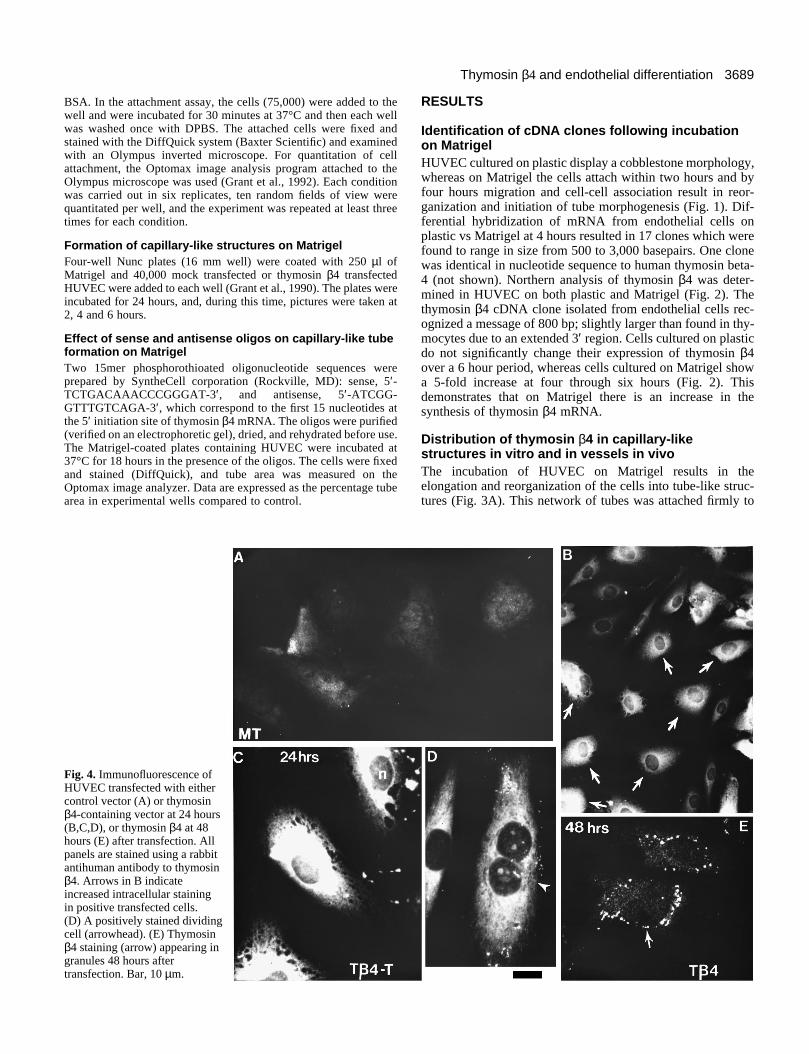
Fig. 4. Immunofluorescence ofHUVEC transfected with eithercontrol vector (A) or thymosinβ4-containing vector at 24 hours(B,C,D), or thymosin β4 at 48hours (E) after transfection. Allpanels are stained using a rabbitantihuman antibody to thymosinβ4. Arrows in B indicateincreased intracellular stainingin positive transfected cells.(D) A positively stained dividingcell (arrowhead). (E) Thymosinβ4 staining (arrow) appearing ingranules 48 hours aftertransfection. Bar, 10 µm.
RESULTS
Identification of cDNA clones following incubationon MatrigelHUVEC cultured on plastic display a cobblestone morphology,whereas on Matrigel the cells attach within two hours and byfour hours migration and cell-cell association result in reor-ganization and initiation of tube morphogenesis (Fig. 1). Dif-ferential hybridization of mRNA from endothelial cells onplastic vs Matrigel at 4 hours resulted in 17 clones which werefound to range in size from 500 to 3,000 basepairs. One clonewas identical in nucleotide sequence to human thymosin beta-4 (not shown). Northern analysis of thymosin β4 was deter-mined in HUVEC on both plastic and Matrigel (Fig. 2). Thethymosin β4 cDNA clone isolated from endothelial cells rec-ognized a message of 800 bp; slightly larger than found in thy-mocytes due to an extended 3′ region. Cells cultured on plasticdo not significantly change their expression of thymosin β4over a 6 hour period, whereas cells cultured on Matrigel showa 5-fold increase at four through six hours (Fig. 2). Thisdemonstrates that on Matrigel there is an increase in thesynthesis of thymosin β4 mRNA.
Distribution of thymosin β4 in capillary-likestructures in vitro and in vessels in vivoThe incubation of HUVEC on Matrigel results in theelongation and reorganization of the cells into tube-like struc-tures (Fig. 3A). This network of tubes was attached firmly to
3690 D. S. Grant and others
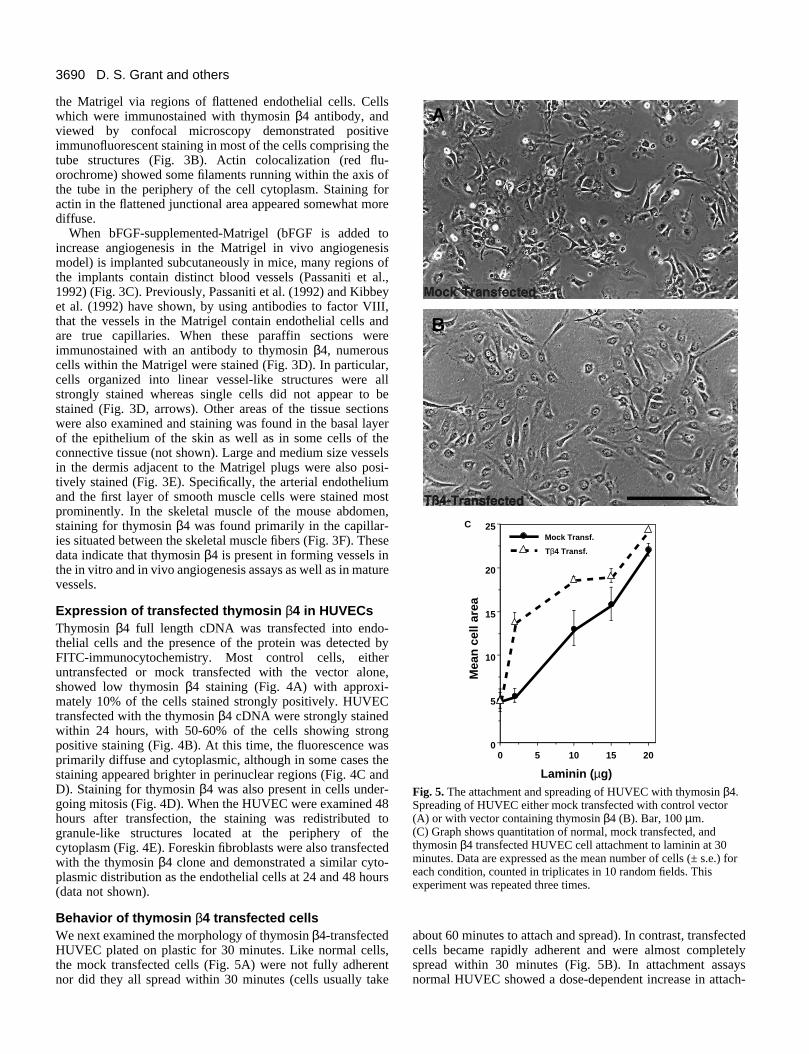
Fig. 5. The attachment and spreading of HUVEC with thymosin β4.Spreading of HUVEC either mock transfected with control vector(A) or with vector containing thymosin β4 (B). Bar, 100 µm.(C) Graph shows quantitation of normal, mock transfected, andthymosin β4 transfected HUVEC cell attachment to laminin at 30minutes. Data are expressed as the mean number of cells (± s.e.) foreach condition, counted in triplicates in 10 random fields. Thisexperiment was repeated three times.
A
B
201510500
5
10
15
20
25Mock Transf.
Tβ4 Transf.
Laminin (µg)
Mea
n c
ell a
rea
C
the Matrigel via regions of flattened endothelial cells. Cellswhich were immunostained with thymosin β4 antibody, andviewed by confocal microscopy demonstrated positiveimmunofluorescent staining in most of the cells comprising thetube structures (Fig. 3B). Actin colocalization (red flu-orochrome) showed some filaments running within the axis ofthe tube in the periphery of the cell cytoplasm. Staining foractin in the flattened junctional area appeared somewhat morediffuse.
When bFGF-supplemented-Matrigel (bFGF is added toincrease angiogenesis in the Matrigel in vivo angiogenesismodel) is implanted subcutaneously in mice, many regions ofthe implants contain distinct blood vessels (Passaniti et al.,1992) (Fig. 3C). Previously, Passaniti et al. (1992) and Kibbeyet al. (1992) have shown, by using antibodies to factor VIII,that the vessels in the Matrigel contain endothelial cells andare true capillaries. When these paraffin sections wereimmunostained with an antibody to thymosin β4, numerouscells within the Matrigel were stained (Fig. 3D). In particular,cells organized into linear vessel-like structures were allstrongly stained whereas single cells did not appear to bestained (Fig. 3D, arrows). Other areas of the tissue sectionswere also examined and staining was found in the basal layerof the epithelium of the skin as well as in some cells of theconnective tissue (not shown). Large and medium size vesselsin the dermis adjacent to the Matrigel plugs were also posi-tively stained (Fig. 3E). Specifically, the arterial endotheliumand the first layer of smooth muscle cells were stained mostprominently. In the skeletal muscle of the mouse abdomen,staining for thymosin β4 was found primarily in the capillar-ies situated between the skeletal muscle fibers (Fig. 3F). Thesedata indicate that thymosin β4 is present in forming vessels inthe in vitro and in vivo angiogenesis assays as well as in maturevessels.
Expression of transfected thymosin β4 in HUVECsThymosin β4 full length cDNA was transfected into endo-thelial cells and the presence of the protein was detected byFITC-immunocytochemistry. Most control cells, eitheruntransfected or mock transfected with the vector alone,showed low thymosin β4 staining (Fig. 4A) with approxi-mately 10% of the cells stained strongly positively. HUVECtransfected with the thymosin β4 cDNA were strongly stainedwithin 24 hours, with 50-60% of the cells showing strongpositive staining (Fig. 4B). At this time, the fluorescence wasprimarily diffuse and cytoplasmic, although in some cases thestaining appeared brighter in perinuclear regions (Fig. 4C andD). Staining for thymosin β4 was also present in cells under-going mitosis (Fig. 4D). When the HUVEC were examined 48hours after transfection, the staining was redistributed togranule-like structures located at the periphery of thecytoplasm (Fig. 4E). Foreskin fibroblasts were also transfectedwith the thymosin β4 clone and demonstrated a similar cyto-plasmic distribution as the endothelial cells at 24 and 48 hours(data not shown).
Behavior of thymosin β4 transfected cellsWe next examined the morphology of thymosin β4-transfectedHUVEC plated on plastic for 30 minutes. Like normal cells,the mock transfected cells (Fig. 5A) were not fully adherentnor did they all spread within 30 minutes (cells usually take
about 60 minutes to attach and spread). In contrast, transfectedcells became rapidly adherent and were almost completelyspread within 30 minutes (Fig. 5B). In attachment assaysnormal HUVEC showed a dose-dependent increase in attach-
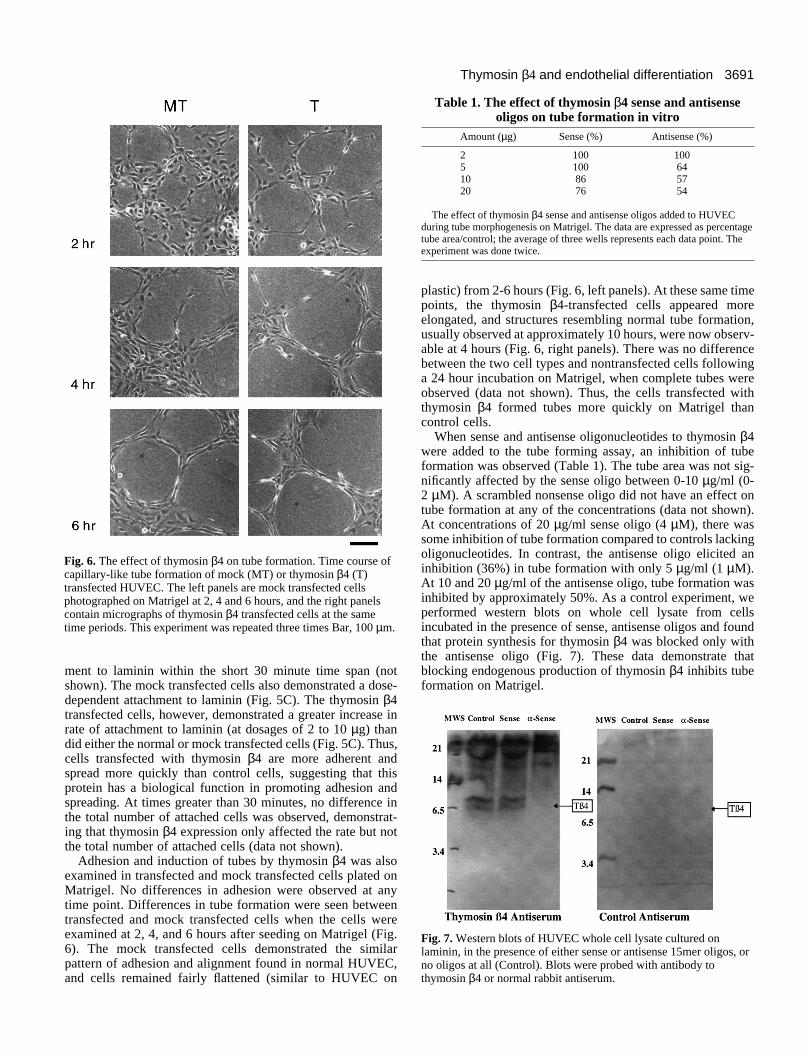
3691Thymosin β4 and endothelial differentiation
Fig. 6. The effect of thymosin β4 on tube formation. Time course ofcapillary-like tube formation of mock (MT) or thymosin β4 (T)transfected HUVEC. The left panels are mock transfected cellsphotographed on Matrigel at 2, 4 and 6 hours, and the right panelscontain micrographs of thymosin β4 transfected cells at the sametime periods. This experiment was repeated three times Bar, 100 µm.
Table 1. The effect of thymosin β4 sense and antisenseoligos on tube formation in vitro
Amount (µg) Sense (%) Antisense (%)
2 100 1005 100 6410 86 5720 76 54
The effect of thymosin β4 sense and antisense oligos added to HUVECduring tube morphogenesis on Matrigel. The data are expressed as percentagetube area/control; the average of three wells represents each data point. Theexperiment was done twice.
Fig. 7. Western blots of HUVEC whole cell lysate cultured onlaminin, in the presence of either sense or antisense 15mer oligos, orno oligos at all (Control). Blots were probed with antibody tothymosin β4 or normal rabbit antiserum.
ment to laminin within the short 30 minute time span (notshown). The mock transfected cells also demonstrated a dose-dependent attachment to laminin (Fig. 5C). The thymosin β4transfected cells, however, demonstrated a greater increase inrate of attachment to laminin (at dosages of 2 to 10 µg) thandid either the normal or mock transfected cells (Fig. 5C). Thus,cells transfected with thymosin β4 are more adherent andspread more quickly than control cells, suggesting that thisprotein has a biological function in promoting adhesion andspreading. At times greater than 30 minutes, no difference inthe total number of attached cells was observed, demonstrat-ing that thymosin β4 expression only affected the rate but notthe total number of attached cells (data not shown).
Adhesion and induction of tubes by thymosin β4 was alsoexamined in transfected and mock transfected cells plated onMatrigel. No differences in adhesion were observed at anytime point. Differences in tube formation were seen betweentransfected and mock transfected cells when the cells wereexamined at 2, 4, and 6 hours after seeding on Matrigel (Fig.6). The mock transfected cells demonstrated the similarpattern of adhesion and alignment found in normal HUVEC,and cells remained fairly flattened (similar to HUVEC on
plastic) from 2-6 hours (Fig. 6, left panels). At these same timepoints, the thymosin β4-transfected cells appeared moreelongated, and structures resembling normal tube formation,usually observed at approximately 10 hours, were now observ-able at 4 hours (Fig. 6, right panels). There was no differencebetween the two cell types and nontransfected cells followinga 24 hour incubation on Matrigel, when complete tubes wereobserved (data not shown). Thus, the cells transfected withthymosin β4 formed tubes more quickly on Matrigel thancontrol cells.
When sense and antisense oligonucleotides to thymosin β4were added to the tube forming assay, an inhibition of tubeformation was observed (Table 1). The tube area was not sig-nificantly affected by the sense oligo between 0-10 µg/ml (0-2 µM). A scrambled nonsense oligo did not have an effect ontube formation at any of the concentrations (data not shown).At concentrations of 20 µg/ml sense oligo (4 µM), there wassome inhibition of tube formation compared to controls lackingoligonucleotides. In contrast, the antisense oligo elicited aninhibition (36%) in tube formation with only 5 µg/ml (1 µM).At 10 and 20 µg/ml of the antisense oligo, tube formation wasinhibited by approximately 50%. As a control experiment, weperformed western blots on whole cell lysate from cellsincubated in the presence of sense, antisense oligos and foundthat protein synthesis for thymosin β4 was blocked only withthe antisense oligo (Fig. 7). These data demonstrate thatblocking endogenous production of thymosin β4 inhibits tubeformation on Matrigel.

3692 D. S. Grant and others
DISCUSSION
The formation of capillary-like tubes from human umbilicalvein endothelial cells cultured on Matrigel is a complexprocess which involves several steps and the biosynthesis ofnumerous gene products. We have shown previously (Grant etal., 1991) that the addition of cycloheximide or actinomycin Dto this assay system inhibits the differentiation of the cells intotubes. Here we have tried to further the understanding of thisprocess by determining the identity of some of the early genesby subtractive cDNA cloning. One of the clones, thymosin β4,previously not known to be associated with endothelial cells,nor known to be involved in their cell regulation, was foundby northern analysis to increase at early times (over a 6 hourperiod) in cells on Matrigel. Using an antibody to humanthymosin β4, we found this protein was also abundant in cellsforming tubes on Matrigel or in vessels in the in vivo murine-angiogenesis assay (Passaniti et al., 1992; Kibbey et al., 1992).In situ, thymosin β4 was found to be present in both large andsmall vessels in mouse tissue. Transfection of this gene and itssubsequent increased expression in cultured HUVEC demon-strated that the protein is associated with more rapid cellattachment, spreading, and formation of tubes on Matrigel. Allthese processes are essential for tube formation. Antisenseoligos to the thymosin β4 mRNA inhibited tube formation onMatrigel, again indicating its importance in the process.Although the cells attached more rapidly at very early timepoints, no effect on total cell adhesion to either laminin orMartigel was noted after 1 hour. Thus, thymosin β4 does notaffect vessel development by directly regulating adhesion.There are inherent toxicity problems with the antisense/senseprobes that make it difficult to clearly interpret the data. Thesedata, however, when examined in total, suggest that thymosinβ4 is expressed and is biologically active in differentiatingendothelial cells.
Although thymosin β4 was not previously shown to beinvolved in blood vessel formation and stabilization, it mayplay an important role in normal vessel development, differ-entiation, and maintenance. There is strong evidence thatthymosin β4, besides being a thymic hormone, is also a differ-entiation factor for many cell types (Low and Goldstein, 1984;Schöbitz et al., 1991b; Shimamura et al., 1990). Thymosin β4was first shown to be associated with the maturation of devel-oping T-lymphocytes in the thymus (Low et al., 1990; Schöbitzet al., 1991a; Talaev et al., 1991). More recently this moleculehas been shown to be associated with the development of B-lymphocytes, platelets, and macrophages, and thus appears beconsidered a marker for hematopoietic cell differentiation(Gomez-Marquez et al., 1989; Hall, 1991; Lin and Morrison-Bogorad, 1990). Thymosin β4 is also expressed in humantumors, the developing human kidney (Hall, 1991), and ispresent at high levels in the fetal cortex and cerebellum of thedeveloping rat brain (Lin and Morrison-Bogorad, 1990). Wefind that thymosin β4 may be associated with the morphogen-esis of HUVEC into tubes, and is present in vessels in vivo.Since this is the first study that illustrates that the expressionof thymosin β4 is involved in endothelial cell differentiation,further mechanistic investigations are needed to establish ifthymosin β4 induces differentiation in endothelial cells or is aproduct of the differentiation process.
It is an interesting fact that cultured endothelial cells plated
on Matrigel stop proliferating (Kubota et al., 1988), whereas,the proliferation of several tumor cell types is accelerated byMatrigel (Fridman et al., 1991). This difference in the two celltypes frequently correlates with the presence of thymosin β4.For example, in ROS osteosarcoma cells, thymosin β4 isexpressed more in cells possessing the slow growing, differ-entiated osteoblast-like phenotype (17/2.8 cells), than the moreundifferentiated cells (25/1 cells) (Atkinson et al., 1990)exhibiting the proliferative phenotype. An induction of thethymosin β4 mRNA has been shown during differentiation invarious phases of the cell cycle of lymphoid cell lines(Schöbitz et al., 1991a). In these studies, thymosin β4 mRNAcontent increased rapidly 1 hour after ConA stimulation oflymphocytes; thereafter, the mRNA levels continued toincrease, plateaued at about 48 hours, and then returned to theinitial values once the cells were quiescent again (Schöbitz etal., 1991b). It appears that thymosin β4 is modulated with cellproliferation changes, with the levels rising in rapidly prolif-erating cells. Thus, the stimulation of the lymphocyte prolifer-ation by ConA results in an increase in thymosin β4 and withsubsequent decrease the proliferative rate of the cells.Thymosin β4 appears to suppress cell proliferation.
HUVEC plated on Matrigel are also very sensitive tocytochalasin D (Grant et al., 1991), a compound that interruptsthe organization of the cytoskeleton. The reorganization ofactin filaments into fiber bundles is essential for tubeformation. Recent investigations have shown that thymosin β4is an actin monomer sequestering protein (Cassimeris et al.,1992; Safer, 1992; Safer et al., 1991; Sanders et al., 1992; Van-compernolle et al., 1992; Yu et al., 1993). Thymosin β4, likeother actin monomer sequestering proteins such as profilin,destrin, actinobindin, and cofilin, has the ability to inhibit thepolymerization of purified actin subunits, and has been recentlyshown to induce depolymerization of actin filaments incultured epithelial cells and in fibroblasts (Safer, 1992). Morerecently, the role of thymosin β4 in actin regulation has beenreevaluated. It now appears that thymosin β4 can also stimulatethe polymerization of actin monomers in the presence of lowamounts of profilin (Pantaloni and Carlier, 1993). The poly-merization and depolymerization of actin is important in celllocomotion and cell spreading (Safer et al., 1991; Stossel,1983), and now in tube formation. We showed that the over-expression of thymosin β4 could promote accelerated HUVECattachment, spreading, and tube formation. Based on indirectevidence, if thymosin can bind to monomeric actin in thisendothelial cell system, it may slow down migration of thecells across Matrigel and preferentially promote organizationand stabilization of the tube network on Matrigel. If profilin ispresent in endothelial cells during the over-expression ofthymosin β4, actin filament polymerization may increase andprovide additional filaments. Thus, thymosin β4’s actinsequestering ability may help to promote the formation ofvessels. Since cell-cell interactions are also important in tubeformation, and actin assembly is required for this process,thymosin β4 may also be enhancing cell cell organization. Wenow have preliminary results that indicate that the addition ofthymosin β4 protein to HUVEC on glass slides actually resultsin an increase as well as a rearrangement of the actin stressfibers (D. S. Grant, unpublished). Therefore, the exactmechanism of action of thymosin β4 on HUVEC cytoskeletonand the resulting biological response may be very complex.

3693Thymosin β4 and endothelial differentiation
The use of this subtractive cDNA cloning has providedinformation regarding the gene products induced during tubeformation on Matrigel. We have demonstrated that thymosinβ4 is biologically active in tube formation and possibly inangiogenesis in vivo. Thymosin β4 may not be acting alone invessel formation but is nevertheless essential. The mechanismof thymosin β4 action may be on cell cycle, and/or hormonal,and/or on actin regulation in the differentiating endothelialcells, but that has not yet been directly shown. Alternatively,thymosin β4 may have additional biological actions yet to bedefined.
The authors thank Mr Wesley Rose for preparing the western blotand carefully reviewing the manuscript.
REFERENCES
Atkinson, M. J., Freeman, M. W. and Kronenberg, H. M. (1990). Thymosinβ4 is expressed in ROS 17/2.8 osteosarcoma cells in a regulated manner.Mol. Endocrinol. 4, 69-74.
Badamchian, M., Strickler, M. P., Stone, M. J. and Goldstein, A. L. (1988).Rapid isolation of thymosin beta 4 from thymosin fraction 5 by preparativehigh-performance liquid chromatography. J. Chromatogr. 459, 291-300.
Cassimeris, L., Safer, D., Nachmias, V. T. and Zigmond, S. H. (1992).Thymosin beta 4 sequesters the majority of G-actin in resting humanpolymorphonuclear leukocytes. J. Cell Biol. 119, 1261-1270.
Condon, M. R. and Hall, A. K. (1992). Expression of thymosin beta-4 andrelated genes in developing human brain. J. Mol. Neurosci. 3, 165-170.
Folkman, J. and Hanahan, D. (1991). Switch to the angiogenic phenotypeduring tumorigenesis. Princess Takamatsu Symp. 22, 339-347.
Folkman, J. (1992). The role of angiogenesis in tumor growth. Semin. CancerBiol. 3, 65-71.
Folkman, J. and Ingber, D. (1992). Inhibition of angiogenesis. Semin. CancerBiol. 3, 89-96.
Folkman, J. and Shing, Y. (1992). Angiogenesis. J. Biol. Chem. 267, 10931-10934.
Fridman, R., Kibbey, M. C., Royce, L. S., Zain, M., Sweeney, M., Jicha, D.L., Yannelli, J. R., Martin, G. R. and Kleinman, H. K. (1991). Enhancedtumor growth of both primary and established human and murine tumor cellsin athymic mice after coinjection with Matrigel. J. Nat. Cancer Inst. 83, 769-774.
Giordano, T., Howard, T. H., Coleman, J., Sakamoto, K. and Howard, B.H. (1991). Isolation of a population of transiently transfected quiescent andsenescent cells by magnetic affinity cell sorting. Exp. Cell Res. 192, 193-197.
Goldstein, S., Fortdis, C. M. and Howard, B. H. (1989). Enhancedtransfection efficiency and improved cell survival after electroporation ofG2/M-synchronized cells and treatment with sodium butyrate. Nucl. AcidsRes. 17, 3959-3971.
Gomez-Marquez, J., Dosil, M., Segade, F., Bustelo, X., Pichel, J.,Dominguez, F. and Freire, M. (1989). Thymosin-β4 gene: preliminarycharacterisation and expression in tissues, thymic cells, and lymphocytes. J.Immunol. 143, 2740-2744.
Grant, D. S., Kleinman, H. K. and Martin, G. R. (1990). The role ofbasement membranes in vascular development. Ann. NY Acad. Sci. 588, 61-72.
Grant, D. S., Lelkes, P. I., Fukuda, K. and Kleinman, H. K. (1991).Intracellular mechanisms involved in basement membrane induced bloodvessel differentiation in vitro. In Vitro Cell Dev. Biol. 27a, 327-336.
Grant, D. S., Kinsella, J. L., Fridman, R., Auerbach, R., Piasecki, B. A.,Yamada, Y., Zain, M. and Kleinman, H. K. (1992). Interaction ofendothelial cells with a laminin A chain peptide (SIKVAV) in vitro andinduction of angiogenic behavior in vivo. J. Cell. Physiol. 153, 614-625.
Grant, D. S., Kleinman, H. K., Goldberg, I. D., Bhargava, M. M., Nickoloff,B. J., Kinsella, J. L., Polverini, P. and Rosen, E. M. (1993). Scatter factorinduces blood vessel formation in vivo. Proc. Nat. Acad. Sci. USA 90, 1937-1941.
Hall, A. (1991). Differential expression of thymosin genes in human tumorsand in the developing human kidney. Int. J. Cancer 48, 672-677.
Hla, T. and Maciag, T. (1990a). An abundant transcript induced in
differentiating human endothelial cells encodes a polypeptide with structuralsimilarities to G-protein-coupled receptors. J. Biol. Chem. 265, 9308-9313.
Hla, T. and Maciag, T. (1990b). Isolation of immediate-early differentiationmRNAs by enzymatic amplification of subtracted cDNA from humanendothelial cells. Biochem. Biophys. Res. Commun. 167, 637-643.
Ingber, D. E., Madri, J. A. and Folkman, J. (1987). Endothelial growthfactors and extracellular matrix regulate DNA synthesis through modulationof cell and nuclear expansion. In Vitro 23, 387-394.
Ingber, D. E. and Folkman, J. (1989). How does extracellular matrix controlcapillary morphogenesis? Cell 58, 803-805.
Jaffe, E. A., Nachman, R. L., Becker, C. G. and Minick, C. R. (1973).Culture of human endothelial cells derived from umbilical veins-identification by morphological and immunological criteria. J. Clin. Invest.52, 2745-2756.
Kibbey, M. C., Grant, D. S. and Kleinman, H. K. (1992). Role of theSIKVAV site of laminin in promotion of angiogenesis and tumor growth: anin vivo Matrigel model. J. Nat. Cancer Inst. 84, 1633-1638.
Kleinman, H. K., McGarvey, M. L., Liotta, L. A., Gehron-Robbey, P.,Tryggvasson, K. and Martin, G. R. (1986). Isolation and characterizationof type IV procollagen, laminin and heparan sulfate proteoglycan from theEHS sarcoma. Biochemistry 24, 6188.
Kramer, R. H. and Fuh, G. M. (1985). Type IV collagen synthesis by culturedhuman microvascular endothelial cells and its deposition in thesubendothelial basement membrane. Biochemistry 24, 7423-7430.
Kubota, Y., Kleinman, H. K., Martin, G. R. and Lawley, T. J. (1988). Roleof laminin and basement membrane in the morphological differentiation ofhuman endothelial cells into capillary-like structures. J. Cell Biol. 107, 1589-1598.
Lin, S. and Morrison-Bogorad, M. (1990). Developmental expression ofmRNAs encoding thymosins β4 and β10 in rat brain and other tissues. J. Mol.Neurosci. 2, 35-44.
Low, T. and Goldstein, A. (1984). Thymosins: structure, function andtherapeutic application. Thymus 6, 27-42.
Low, T. L. and Goldstein, A. L. (1985). Thymosin beta 4. Meth. Enzymol. 116,248-255.
Low, T. L., Lin, C. Y., Pan, T. L., Chiou, A. J. and Tsugita, A. (1990).Structure and immunological properties of thymosin beta 9 Met, a newanalog of thymosin beta 4 isolated from porcine thymus. Int. J. Pept. ProteinRes. 36, 481-488.
Low, T. L. K. and Goldstein, A. L. (1981). Chemical characterization ofthymosin β4. J. Biol. Chem. 257, 1000-1006.
Maciag, T. (1990). Molecular and cellular mechanisms of angiogenesis.Important Advan. Oncol. 85, 85-98.
Madri, J. A., Dryer, B., Pitlick, F. and Furthmayr, H. (1980). Thecollagenous components of the subendothelium: correlation of structure andfunction. Lab. Invest. 43, 303-315.
Madri, J. A. and Pratt, B. M. (1986). Endothelial cell-matrix interactions: invitro models of angiogenesis. J. Histochem. Cytochem. 34, 85-91.
Naylor, P. H., McClure, J. E., Spangelo, B. L., Low, T. L. K. and Goldstein,A. L. (1984). Immunochemical studies on thymosin: radioimmunoassay ofthymosin β4. Immunopharmacol. 7, 9-16.
Naylor, P. H., Friedman, K. A., Hersh, E., Erdos, M. and Goldstein, A. L.(1986). Thymosin alpha 1 and thymosin beta 4 in serum: comparison ofnormal, cord, homosexual and AIDS serum. Int. J. Immunopharmacol. 8,667-676.
Nicosia, R. F., McCormick, J. F. and Bielunas, J. (1984). The formation ofendothelial webs and channels in plasma clot culure. Scan. Elect. Microsc. 2,793-799.
Paku, S. and Paweletz, N. (1991). First steps of tumor-related angiogenesis.Lab. Invest. 65, 334-346.
Pantaloni, D. and Carlier, M.-F. (1993). How profilin promotes actin filamentassembly in the presence of thymosin β4. Cell 75, 1007-1014.
Passaniti, A., Taylor, R. M., Pili, R., Guo, Y., Long, P. V., Haney, J. A.,Pauly, R. R., Grant, D. S. and Martin, G. R. (1992). A simple, quantitativemethod for assessing angiogenesis and antiangiogenic agents usingreconstituted basement membrane, heparin, and fibroblast growth factor. LabInvest. 67, 519-528.
Pearson, W. R. and Lipman, D. J. (1988). Improved tools for biologicalsequence comparison. Proc. Nat. Acad. Sci. USA 85, 2444-2448.
Safer, D., Elzinga, M. and Nachmias, V. T. (1991). Thymosin beta 4 and Fx,an actin-sequestering peptide, are indistinguishable. J. Biol. Chem. 266,4029-4032.
Safer, D. (1992). The interaction of actin with thymosin beta 4 [news]. J.Muscle Res. Cell Motil. 13, 269-271.

3694 D. S. Grant and others
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning. ALaboratory Manual. Cold Spring Harbor, NY: Cold Spring HarborLaboratory Press.
Sanders, M. C., Goldstein, A. L. and Wang, Y. L. (1992). Thymosin beta 4(Fx peptide) is a potent regulator of actin polymerization in living cells. Proc.Nat. Acad. Sci. USA 89, 4678-4682.
Schöbitz, B., Hannappel, E. and Brand, K. (1991a). The early induction ofthe actin-sequestering peptide thymosin beta 4 in thymocytes depends on theproliferative stimulus. Biochim. Biophys. Acta 1095, 230-235.
Schöbitz, B., Netzker, R., Hannappel, E. and Brand, K. (1991b). Cell-cycle-regulated expression of thymosin beta 4 in thymocytes. Eur. J. Biochem. 199,257-262.
Shimamura, R., Kudo, J., Kondo, H., Dohmen, K., Gondo, H., Okamura,S., Ishibashi, H. and Niho, Y. (1990). Expression of the thymosin β4 geneduring differentiation of hematopoietic cells. Blood 76, 977-984.
Stossel, T. P. (1983). On the crawling of animal cells. Science 260, 1086-1094.
Talaev, V., Yarilin, A. A., Sharova, N. I., Shichkin, V. P. and Shuvaeva, T.M. (1991). Purification and characterization of a thymocyte growth factor. 2.Biological activity of the thymocyte growth factor. Biomed. Sci. 2, 515-519.
Timpl, R., Rohde, H., Gehron Robey, P., Rennard, S. I., Foidart, J. M. andMartin, G. R. (1979). Laminin-a glycoprotein from basement membranes. J.Biol. Chem. 254, 9933-9937.
Vancompernolle, K., Goethals, M., Huet, C., Louvard, D. andVandekerckhove, J. (1992). G- to F-actin modulation by a single amino acidsubstitution in the actin binding site of actobindin and thymosin beta 4.EMBO J. 11, 4739-4746.
Yu, F. X., Lin, S. C., Morrison, B. M., Atkinson, M. A. and Yin, H. L.(1993). Thymosin beta 10 and thymosin beta 4 are both actin monomersequestering proteins. J. Biol. Chem. 268, 502-509.
(Received 7 June 1995 - Accepted 11 September 1995)


















