
Kazi et al., J Autacoids 2012, 1:2 Autacoids · J Autacoids ISSN:2161-0479 Autacoids, an open...
6
Volume 1 • Issue 2 • 1000101 J Autacoids ISSN:2161-0479 Autacoids, an open access journal Open Access Research Article Autacoids Kazi et al., J Autacoids 2012, 1:2 http://dx.doi.org/10.4172/2161-0479.1000101 The role of Α 1b -Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake Raisa N Kazi 1 *, Munavvar A Sattar 2 , Nor A Abdullah 3 , Md A Hye Khan 4 and Edward J Johns 5 1 College of Applied Medical Science, Salman Bin Abdul-Aziz University, Saudi Arabia 2 School of Pharmaceutical Sciences, University Sains Malaysia, 11800, Penang, Malaysia 3 Department of Pharmacology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia 4 Department of Pharmacology and Toxicology, Medical College of Wisconsin, Wisconsin, USA 5 Department of Physiology, Aras Windle, University College Cork, College Road, Cork, Ireland Abstract Background: Renal α 1B -adrenoreceptors (α 1 -ARs) contributes to the pathogenesis of hypertension in spontaneously hypertensive rats (SHR). The present study examined the role of α 1B -ARs in renal hemodynamic and tubular functions in SHR subjected to high-salt diet (SHRHNa) for 6 weeks. Methods: Renal cortical vasoconstriction to noradrenaline (NA), phenylephrine (PE), and methoxamine (ME) in presence and absence of chloroethylclonidine (CEC) was measured in SHRHNa and SHR on normal sodium diet (SHRNNa). Renal tubular functional responses to PE and CEC were assessed as a measure of insulin clearance. Results: SHRHNa showed no signifcant change in the mean arterial pressure (MAP) as compared to SHRNNa. SHRHNa expressed enhanced renal cortical vascular sensitivity to NA, PE, and ME. Furthermore, renal vasoconstrictor response to NA, PE and ME was accentuated in the presence of CEC in SHRNNa. On the other hand in SHRHNa, renal cortical vasoconstriction to NA and ME was inhibited by CEC. However tubular response to PE was inhibited by CEC in SHRNNa and remains unaffected in SHRHNa. Conclusion: Thus it is concluded that, augmented α 1 -adrenergic response to adrenergic stimuli contribute to salt-related increase in renal vascular sensitivity in SHRHNa and these changes were independent of any further increase in MAP in SHRHNa. Irrespective of dietary sodium changes, α 1 -ARs are involved in mediation of tubular functions like antinatriuresis and antidiuresis of SHR. In addition, α 1B -ARs are the functional subtypes that mediate renal cortical vasoconstriction in SHRHNa and tubular function in SHRNNa, while high sodium in SHR did not infuence the functionality of α 1B -ARs in mediating tubular functions. Keywords: High salt; Renal α 1B AR Introduction Experimental evidences have implicated the role of a high-sodium in the pathogenesis of hypertension, particularly in salt-sensitive individuals [1,2] . In relation to this, high dietary sodium intake has been shown to infuence the expression of adrenoreceptors, possibly contributing to the pathogenesis of essential hypertension [3]. Renal α 1 -ARs play important role in the regulation of renal hemodynamic and tubular functions [4]. Pharmacological and cloning studies have documented, three subtypes of α -ARs: α , α , and α [5,6]. 1 1A 1B 1D Among these subtypes, α 1B -ARs have been found to be crucial in the regulation of renal vascular resistance in SHR and thus contribute to the increased vascular resistance in hypertension [7]. Furthermore, α 1B -ARs are also involved in the mediation of renal tubular sodium and water reabsorption [8]. α 1B -AR- mediated renal vasoconstriction has also been observed in some pathological conditions in rats, such as diabetes, hypertensive diabetes, heart failure, and renal failure [9,10]. Studies on the infuence of high salt load on renal α 1 -ARs in Wistar Kyoto rats have revealed signifcant involvement of α 1B- ARs in mediating the adrenergically-induced renal cortical vasoconstriction [11]. In addition, gene knockouts studies in mice have suggested the role of α 1D -AR in the development of high blood pressure in response to dietary salt load [12]. However, no information available regarding the α 1B -adrenoceptors functional contributions in the regulation of renal hemodynamic and renal tubular functions in SHR subjected to high sodium diet. Te present study investigated the infuence of a high-salt diet on the regulation of renal α 1 -ARs in hypertension, with particular attention to the functional contribution of α 1B -ARs in the regulation of renal hemodynamic and tubular functions in SHR on high sodium diet. Material and Methods Animals Male SHR with body weight ranging from 200 to 250 (g) were selected and maintained in the animal care facility, University Sains Malaysia, Penang, Malaysia. All procedures on animals were approved by the Animal Ethics Committee, University Sains Malaysia, Penang, Malaysia. Normal and high sodium diet feeding regime and experimental groups SHR rats were maintained for six weeks on a normal sodium and high sodium diet. SHR with normal sodium intake were given normal standard rat chow (Gold Coin Feed mills Sdn Bhd, Malaysia) and tap *Corresponding author: Raisa N Kazi, College of Applied Medical Science, Sal- man Bin Abdul-Aziz University, Saudi Arabia; E-mail: [email protected] Received March 26, 2012; Accepted June 11, 2012; Published June 14, 2012 Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α 1b -Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101 Copyright: © 2012 Kazi RN, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Transcript of Kazi et al., J Autacoids 2012, 1:2 Autacoids · J Autacoids ISSN:2161-0479 Autacoids, an open...

Volume 1 • Issue 2 • 1000101 J Autacoids ISSN:2161-0479 Autacoids, an open access journal
Open Access Research Article
Autacoids
Kazi et al., J Autacoids 2012, 1:2http://dx.doi.org/10.4172/2161-0479.1000101
The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake Raisa N Kazi1*, Munavvar A Sattar2, Nor A Abdullah3, Md A Hye Khan4 and Edward J Johns5
1 College of Applied Medical Science, Salman Bin Abdul-Aziz University, Saudi Arabia 2School of Pharmaceutical Sciences, University Sains Malaysia, 11800, Penang, Malaysia 3 Department of Pharmacology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia 4 Department of Pharmacology and Toxicology, Medical College of Wisconsin, Wisconsin, USA 5 Department of Physiology, Aras Windle, University College Cork, College Road, Cork, Ireland
Abstract
Background: Renal α1B-adrenoreceptors (α1-ARs) contributes to the pathogenesis of hypertension in spontaneously hypertensive rats (SHR). The present study examined the role of α1B-ARs in renal hemodynamic and tubular functions in SHR subjected to high-salt diet (SHRHNa) for 6 weeks.
Methods: Renal cortical vasoconstriction to noradrenaline (NA), phenylephrine (PE), and methoxamine (ME) in presence and absence of chloroethylclonidine (CEC) was measured in SHRHNa and SHR on normal sodium diet (SHRNNa). Renal tubular functional responses to PE and CEC were assessed as a measure of insulin clearance.
Results: SHRHNa showed no significant change in the mean arterial pressure (MAP) as compared to SHRNNa. SHRHNa expressed enhanced renal cortical vascular sensitivity to NA, PE, and ME. Furthermore, renal vasoconstrictor response to NA, PE and ME was accentuated in the presence of CEC in SHRNNa. On the other hand in SHRHNa, renal cortical vasoconstriction to NA and ME was inhibited by CEC. However tubular response to PE was inhibited by CEC in SHRNNa and remains unaffected in SHRHNa.
Conclusion: Thus it is concluded that, augmented α1-adrenergic response to adrenergic stimuli contribute to salt-related increase in renal vascular sensitivity in SHRHNa and these changes were independent of any further increase in MAP in SHRHNa. Irrespective of dietary sodium changes, α1-ARs are involved in mediation of tubular functions like antinatriuresis and antidiuresis of SHR. In addition, α1B-ARs are the functional subtypes that mediate renal cortical vasoconstriction in SHRHNa and tubular function in SHRNNa, while high sodium in SHR did not influence the functionality of α1B-ARs in mediating tubular functions.
Keywords: High salt; Renal α1B AR
Introduction Experimental evidences have implicated the role of a high-sodium
in the pathogenesis of hypertension, particularly in salt-sensitive individuals [1,2] . In relation to this, high dietary sodium intake has been shown to influence the expression of adrenoreceptors, possibly contributing to the pathogenesis of essential hypertension [3]. Renal α1-ARs play important role in the regulation of renal hemodynamic and tubular functions [4]. Pharmacological and cloning studies have documented, three subtypes of α -ARs: α , α , and α [5,6].1 1A 1B 1D Among these subtypes, α1B-ARs have been found to be crucial in the regulation of renal vascular resistance in SHR and thus contribute to the increased vascular resistance in hypertension [7]. Furthermore, α1B-ARs are also involved in the mediation of renal tubular sodium and water reabsorption [8]. α1B-AR- mediated renal vasoconstriction has also been observed in some pathological conditions in rats, such as diabetes, hypertensive diabetes, heart failure, and renal failure [9,10]. Studies on the influence of high salt load on renal α1-ARs in Wistar Kyoto rats have revealed significant involvement of α1B-ARs in mediating the adrenergically-induced renal cortical vasoconstriction [11]. In addition, gene knockouts studies in mice have suggested the role of α1D-AR in the development of high blood pressure in response to dietary salt load [12]. However, no information available regarding the α1B-adrenoceptors functional contributions in the regulation of renal hemodynamic and renal tubular functions in SHR subjected to high sodium diet.
The present study investigated the influence of a high-salt diet on the regulation of renal α1-ARs in hypertension, with particular
attention to the functional contribution of α1B-ARs in the regulation of renal hemodynamic and tubular functions in SHR on high sodium diet.
Material and Methods Animals
Male SHR with body weight ranging from 200 to 250 (g) were selected and maintained in the animal care facility, University Sains Malaysia, Penang, Malaysia. All procedures on animals were approved by the Animal Ethics Committee, University Sains Malaysia, Penang, Malaysia.
Normal and high sodium diet feeding regime and experimental groups
SHR rats were maintained for six weeks on a normal sodium and high sodium diet. SHR with normal sodium intake were given normal standard rat chow (Gold Coin Feed mills Sdn Bhd, Malaysia) and tap
*Corresponding author: Raisa N Kazi, College of Applied Medical Science, Salman Bin Abdul-Aziz University, Saudi Arabia; E-mail: [email protected]
Received March 26, 2012; Accepted June 11, 2012; Published June 14, 2012
Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Copyright: © 2012 Kazi RN, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Page 2 of 6
Volume 1 • Issue 2 • 1000101
water ad libitum. High sodium intake SHR were given normal standard rat chow and supplemented with (0.9%) isotonic saline to drink ad libitum. Once SHR had completed the high sodium and normal sodium diet feeding schedule of six weeks, they were subjected to acute renal vasoconstrictor and tubular functional studies. Four groups of rats were studied (each group n=6). Group 1 and 2 are SHRNNa and SHRHNa subjected to renal vasoconstrictor experiemnts. Group 3 and 4 are SHRNNa and SHRHNa subjected to renal tubular functional studies.
Renal vasoconstrictor study
Surgical preparation of animal: Overnight fasted (with water ad libitum) rats were anesthetized with 60 mg/kg (i.p.) sodium pentobarbitone (Nembutal®, CEVA Sante Animale, Liboure, France). After tracheostomy with endotracheal cannula (Portex, Kent, UK), the carotid artery was cannulated (PE 50, Portex, Kent, UK) and connected to a pressure transducer (P23 ID Gould, Statham Instrument, UK) coupled to a computerized data acquisition system (Power Lab, AD instrument, Australia) for continuous measurement of MAP. The left jugular vein was cannulated (PE 50, Portex, Kent, UK) to permit an intermittent administration of supplementary doses of anesthetic (10 mg/kg in 0.9% Saline). Following a midline abdominal incision, the left kidney was exposed. Then a cannula (PE 50, Portex, Kent, UK) was inserted via the iliac artery till the level of the renal artery of left kidney to enable exogenous administration of adrenergic agonists and antagonist in to the kidney. The urinary bladder was catheterized using PE 50 cannula (Protex Ltd. Kent, UK) to facilitate urination of the animal. Adrenal artery was located and the circulation was abolished by cutting the blood supply to the adrenal medulla to prevent the endogenous adrenaline and noradrenaline to enter the systemic circulation and reach the kidney. This was important to establish that the vasoactive responses obtained are induced by the exogenously administered adrenergic agonist only. Upon completion of surgery, 2 ml of saline were given slowly and intravenously through the jugular vein to the animal as a primer. Subsequently a continuous infusion (6 ml/hr) of saline started intrarenally through a PE 50 catheter inserted into the left iliac artery which was further connected to an infusion pump (Perfusor secura FT 50 ml, B, Braun). Renal cortical perfusion (RCP) was measured by Needle (implantable) Laser Doppler flow meter. The probe was connected to a Power lab AD instrument, Australia. An hour period was allowed for equilibrium and stabilization before commencing the acute renal cortical vasoconstrictor response study [13].
Experimental protocol for renal cortical vasoconstrictor response: Once the animal is stabilized, in each phase baseline MAP and RCP were recorded initially for 15 minutes in order to get a baseline values, followed by a three phase renal vasoconstrictor experiment. In the first phase saline was administered by continuous intra-renal infusion during which graded bolus doses of NA (25, 50, 100 & 200 ng), PE (0.25, 0.5, 1 & 2 µg) and ME (1, 2, 3 & 4 µg) were given in ascending and then descending orders through the iliac arterial cannula directly into the kidney. The renal vasoconstrictor responses to these adrenergic
agonists were expressed as % decrease in the RCP. During the second phase, low bolus dose of the CEC (5 µg/kg) was administered followed by a continuous infusion of CEC (1.25 µg/kg/hr) close renal arterially through the iliac arterial cannula, then 15 minute later, the second set of renal vasoconstrictor responses to NA, PE and ME was carried out. In the last phase, a high bolus dose of CEC (10 µg/kg) was injected followed by a continuous infusion of CEC (2.5 µg/kg/hr). Upon achieving the steady state, 15 minutes later, the same procedure used in the first and second phases was followed [13].
Renal tubular study
Surgical preparation for renal tubular studies: Surgical procedure for tubular functional studies was similar to those of the renal vasoconstriction studies, with an exception that the left ureter was cannulated for collection of urine. Two ml of insulin (10 mg/ml) in saline were given as a primer via the jugular vein cannula. The left iliac artery was isolated and catheterized (PE 50, Protex, Kent, UK), then the bevelled tip of the cannula was further inserted into the abdominal aorta up to the level of the entrance of the renal artery of the left kidney. This step was necessary to allow an exogenous administration of adrenergic agonist, antagonist and/or saline infusion (6 ml/hr) containing insulin (10 mg/ml) directly into the kidney. The left iliac artery cannula was further connected to a pressure transducer (P23 ID Gould, Statham Instrument, UK) coupled to a computerized data acquisition system for continuous measurement of renal arterial pressure (RAP). RCP was measured by Needle (implantable) Laser Doppler flow meter. The probe is connected to the Power lab (AD instrument, Australia). The MAP was recorded through the carotid artery cannula (PE 50. Protex, Kent, UK) which was connected to a pressure transducer (P23 ID Gould, Statham Instrument, UK) coupled to a computerized data acquisition system (Power Lab, AD instrument, Australia). In all the experiments 1 to 1.5 cm of the aorta was cleared and a screw-controlled snare was placed it above the renal arteries. The snare was slightly tightened to lower blood pressure distal to the construction site if systemic pressure was to rise as a result of adrenergic infusion. Later during experiment the snare could be released or tightened as needed and the mean blood pressure below the snare equivalent to RAP was maintained constant [14].
Experimental protocol for renal tubular studies: Once the animal is stabilized, three phase experiment was instigated and each part was continued for 2 hours, phase 1 was control (saline) phase. Phase 2 was PE-administration phase during which a dose of (100 µg/kg/hr) of PE was infused through the iliac arterial cannula directly into the kidney. Likewise phase 3 involved the administration of same dose of PE; however it was carried out in the presence of CEC (10 µg/kg/hr). RAP, RCP, and MAP was recorded initially to get a baseline values in all the three phases before the start of clearance. During each phase, three 15 minute urine clearance samples were collected to measure urine volume and subsequently calculate the urine flow rate (UFR). Blood samples were taken at the beginning and at the end of each phase for measurement of plasma sodium. Arterial blood samples (400 µl)
Adrenergic agonist SHRNNa SHRHNa
NA PE ME NA PE ME Phase1 Saline 50.1 ± 5.1 48.5 ± 5.4 29.9 ± 5.0 54.2 ± 4.5 61.3 ± 4.6 58.4 ± 4.8 Phase2 CEC (5µg/Kg) 51.7 ± 5.3 52.3 ± 6.2 47.1 ± 5.6* 62.3 ± 4.7* 64.6 ± 4.7 54.6 ± 4.5 Phase3 CEC (10µg/Kg) 60.4 ± 5.2# 69.4 ± 5.4# 50.7 ± 5.2# 49.3 ± 4.6§ 59.3 ± 4.7 64 ± 5.3#
Table1: Over all mean % change in RCP in response to NA, PE, ME in SHRNNa & SHRHNa rats in absence and presence of low(5µg/kg) and high dose of CEC (10µg/kg), Values are mean ± S.E.M *indicate p<0.05 between phase2 and phase1. # indicate p<0.05 between phase 3 and phase 1. § indicate p<0.05 between phase 2 and phase 3.
J AutacoidsISSN:2161-0479 Autacoids, an open access journal

Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Page 3 of 6
Volume 1 • Issue 2 • 1000101
were withdrawn from the carotid cannula into a precooled syringe, centrifuged for 2 min (6000 rpm) and the clear plasma was removed. The remaining packed blood cells were resuspended in an equal volume of saline and reinfused into the animal immediately through the jugular vein cannula [14]. Then the plasma and urine samples were assayed for insulin using the modified method [15]. Plasma and urine were also subjected to sodium measurements by flame emission photometry following this glomerular filtration rate (GFR), absolute sodium excretion (UNaV) and fractional sodium excretion (FENa) was calculated.
Drugs used: CEC in the present study is used to characterize the functional contribution of α1B-ARs in mediating adrenergically-induced renal cortical vasoconstriction and renal tubular functional responses. CEC is irreversible antagonists which alkylates and inactivates α1BAR there by decreases the renal cortical perfusion and function [16]. Noradrenaline is a mixed agonist and induces vasoconstriction in different vascular bed mediated by all the three subtypes of α1adrenoceptors and hence decreases the renal cortical perfusion [16]. PE is a nonselective synthetic agonist of α1-adrenoceptors with an ability to activate all three α , α and α -adrenoceptor subtypes 1A 1B 1D[16] that increase the renal vascular resistance and decrease the renal cortical perfusion when induced renal arterially. Methoxamine is a most powerful renal vasoconstrictor that leads to decrease in the renal cortical perfusion [16].
Statistical analysis
The RCP responses to NA, PE and ME in vasoconstrictor experiment were taken as the average percentage reductions from baseline values caused by each dose of adrenergic stimuli administered in ascending and descending orders. All data were expressed as mean % change ± S.E.M. of renal vasoconstrictor responses produced by all the doses (adrenergic agonists) and were compared between the phases (saline, low and high dose of antagonist treated phases). The data from the renal vasoconstriction experiments were subjected to two-way ANOVA, while the physiological data were analyzed by using one way ANOVA followed by the Bonferonni post-hoc test (Superanova, Abacus Inc., CA, USA). The differences between the means were considered significant at the 5% level.
The renal tubular functional responses in all the three phases were measured by taking the average value of the three clearances in each phase. All data were expressed as means ± S.E.M. and were compared between the phases (saline, agonist and antagonist treated phases). Statistical analysis was performed by one-way ANOVA on repeated
MAP (mmHg) RCP (BPU/min) Treatment SHRNNa SHRHNa SHRNNa SHRHNa
Phase 1 (saline) 136.4 ± 4.4 142.4 ± 4.5 216.0 ± 17.2 172.6 ± 20.6 Phase 2 (CEC 5µg/ kg) 121.0 ± 3.1 138.3 ± 3.2 201.8 ± 25.1 137.4 ± 12.4 Phase 3 (CEC 10µg /kg) 110.2 ± 2.3* 128.6 ± 6.1* 188.6 ± 20.6 158.6 ± 19.5
Table2: MAP and RCP (BPU) in SHRNNa and SHRHNa rats in the absence and presence of low and high dose of CEC in renal vasoconstrictor experiment. Data are presented as mean ± SEM * p<0.05 vs. Phase1.
measures (Superanova, Abacus Inc., CA, USA) followed by Bonferonni post-hoc test. Differences between the means were considered significant at the 5% level. The absolute and percentage changes quoted in the text represent the mean value calculated from individual rats.
Results Renal cortical vasoconstrictor response
Results show no significant change in the MAP between the SHRNNa and SHRHNa groups (SHRNNa-151.8 ± 3.3 vs. 154.2 ± 3.1mmHg). NA, PE and ME produced dose-dependent vasoconstrictions in the control phases1 in both SHRNNa and SHRHNa, which was evaluated in terms of % reduction in the RCP (Table1). In addition, NA, PE and ME-induced renal vasoconstrictions were significantly greater (p<0.05) in SHRHNa compared to SHRNNa when the measurements were performed during the control saline phase1 (in the absence of antagonist). In the later subsequent phases of the experiment, renal vasoconstrictor responses induced by NA, PE and ME were rather accentuated by both low dose of CEC (phase2) and more significantly (p<0.05) by high doses of CEC (phase3) in SHRNNa relative to their corresponding control phase 1. These effects collectively contributed to further significant reduction in the overall % change in the RCP (Table1).
In SHRHNa, NA induced renal vasoconstriction was significantly (p<0.05) accentuated by low dose of CEC in phase2, while high dose of CEC significantly (P<0.05) inhibited the NA induced renal vasoconstriction responses in phase3. Similarly PE induced renal cortical vasoconstriction were further enhanced by low dose (phase2) of CEC and attenuated by high dose of CEC in phase3. In spite of these observations neither of these two values attains statistical significance (Table1). On the contrary ME induced renal cortical vasoconstrictions in SHRHNa is attenuated by both low dose of CEC (phase2) and more significantly (p<0.05) by high doses of CEC (phase3) in SHRHNa (Table1). The administration of NA, PE, ME and CEC brought no effect on baseline measurements of RCP between all the three phases of in vivo acute renal vasoconstrictor experiment (Table2). However, we observed a significant (p<0.05) decrease in the MAP in the third phase of renal cortical vasoconstrictor experiment in both SHRNNa and SHRHNa (Table 2).
Renal tubular functional response
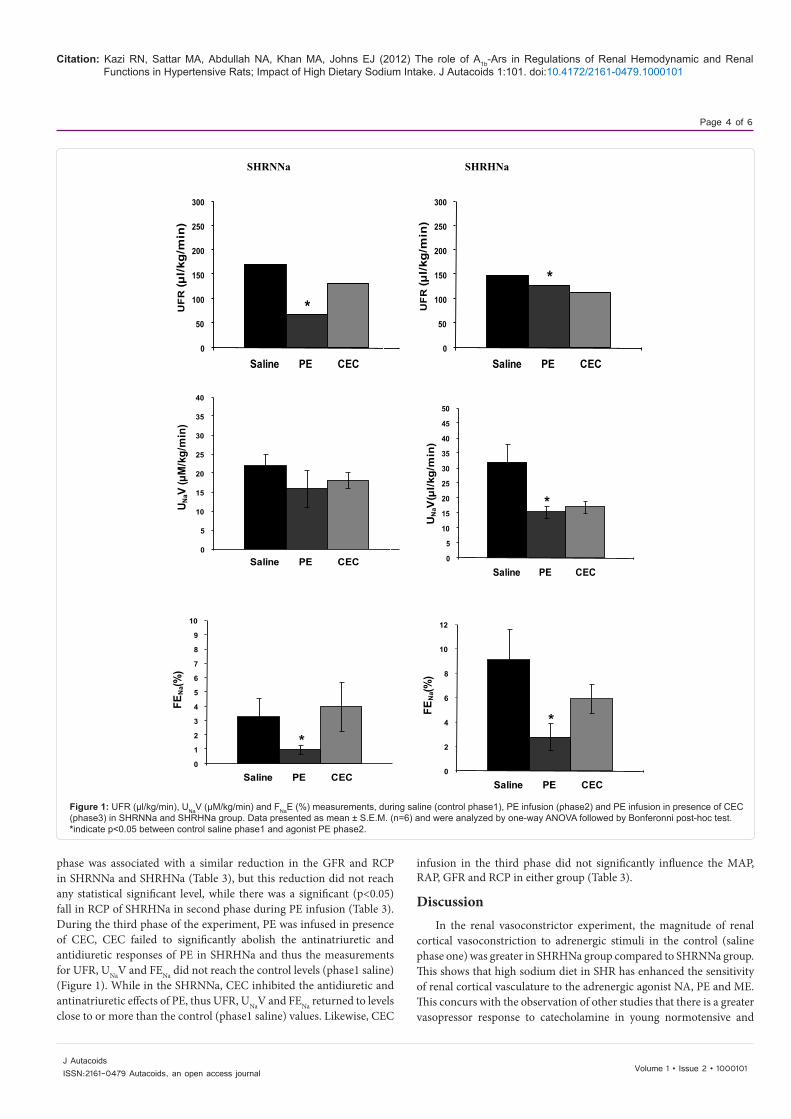
During the first phase, the tubular responses like UFR, UNaV and FENa were obtained while saline was infused (phase1: control saline phase). It can be observed that measurements of UNaV and FENa in control phase were significantly (p<0.05) greater in SHRHNa compared to the SHRNNa. In both SHRNNa and SHRHNa groups, during the second phase (Figure 1), PE infused close renal intra arterially caused a significant reduction in the UFR, UNaV and FENa compared to the corresponding control values of the first saline phase (Figure 1), except that the decrease in the UNaV in response to phenylephrine in the SHRNNa did not reach the statistically significant level. Close renal arterial infusion of PE brought no significant changes in MAP and RAP (Table 3). On the other hand, PE infusion in the second
MAP GFR RCP RAP Phase SHRNNa SHRHNa SHRNNa SHRHNa SHRNNa SHRHNa SHRNNa SHRHNa 1 139.2 ± 4.8 155.0 ± 5.6 3.5 ± 1.3 3.5 ± 1.1 206.1 ± 20.9 269.8 ± 14.9 141.6 ± 7.3 156.8 ± 4.3 2 137.4 ± 5.5 154.4 ± 5.9 1.2 ± 0.5 1.2 ± 0.3 185.2 ± 14.9 191.0 ± 14.8* 138.8 ± 8.7 153.6 ± 5.5 3 136.0 ± 5.1 149.8 ± 5.3 2.0 ± 0.5 1.4 ± 0.3 208.6 ± 11.4 223.2 ± 18.7 135.6 ± 10.2 149.0 ± 6.1
Table3: GFR (ml/kg/min), MAP (mmHg), RAP (mmHg) and RCP (BPU/min) in SHRNNa and SHRHNa rats in control (saline) phase1, phase2 (PE) and phase3 (PE in presence of CEC). Values are mean ± S.E.M *indicate p<0.05 between control and agonist PE
J AutacoidsISSN:2161-0479 Autacoids, an open access journal
Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Page 4 of 6
SHRNNa SHRHNa
300 300
0
50
1
UF
R (µ
l/kg
/min
)
Saline PE CEC
*
0
50
1
UF
R (µ
l/kg
/min
)
Saline PE CEC
*
30
35
40
U NaV
(µM
/kg/
min
)
40
45
50
UN
aV(µ
l/kg/
min
)
*
250 250
200 200
150 150
100 100
FEN
a(%)
FEN
a(%)
5 0
01Saline PE CEC 1Saline PE CEC
10 12 9
8 10
7
*
3525
3020
25 15
20
15
10
10
5
*
1
86
5
4
3
6
4 2
2
0
1
01Saline PE CEC
Saline PE CEC
Figure 1: UFR (µl/kg/min), UNaV (µM/kg/min) and FNaE (%) measurements, during saline (control phase1), PE infusion (phase2) and PE infusion in presence of CEC (phase3) in SHRNNa and SHRHNa group. Data presented as mean ± S.E.M. (n=6) and were analyzed by one-way ANOVA followed by Bonferonni post-hoc test. *indicate p<0.05 between control saline phase1 and agonist PE phase2.
phase was associated with a similar reduction in the GFR and RCP in SHRNNa and SHRHNa (Table 3), but this reduction did not reach any statistical significant level, while there was a significant (p<0.05) fall in RCP of SHRHNa in second phase during PE infusion (Table 3). During the third phase of the experiment, PE was infused in presence of CEC, CEC failed to significantly abolish the antinatriuretic and antidiuretic responses of PE in SHRHNa and thus the measurements for UFR, UNaV and FENa did not reach the control levels (phase1 saline) (Figure 1). While in the SHRNNa, CEC inhibited the antidiuretic and antinatriuretic effects of PE, thus UFR, UNaV and FENa returned to levels close to or more than the control (phase1 saline) values. Likewise, CEC
infusion in the third phase did not significantly influence the MAP, RAP, GFR and RCP in either group (Table 3).
Discussion In the renal vasoconstrictor experiment, the magnitude of renal
cortical vasoconstriction to adrenergic stimuli in the control (saline phase one) was greater in SHRHNa group compared to SHRNNa group. This shows that high sodium diet in SHR has enhanced the sensitivity of renal cortical vasculature to the adrenergic agonist NA, PE and ME. This concurs with the observation of other studies that there is a greater vasopressor response to catecholamine in young normotensive and
J Autacoids Volume 1 • Issue 2 • 1000101ISSN:2161-0479 Autacoids, an open access journal

Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Page 5 of 6
Volume 1 • Issue 2 • 1000101
hypertensive subjects with a high sodium intake [11,17]. Augmented vascular responsiveness because of high salt load results in a greater increase in vascular résistance, providing greater resistance to blood flow, and predisposing an individual to salt-induced blood pressure response [18].A variety of mechanisms appear to contribute to high-sodium intake-associated augmentation of vascular reactivity. Increases in the generation of reactive oxygen species that reduce endotheliumdependent vasodilation, [19] down-regulation of nitric oxide synthase expression in normotensive rats, [20] and abnormally high activation of intra-renal renin-angiotensin system [21] are among the possible mechanisms of high-salt-related alteration of vascular reactivity. Some reports have also suggested that in the aorta of SHR, excessive dietary salt directly affects the vascular smooth muscles and causes impairment of the relaxation response triggered by nitric oxide [22]. Studies using other salt-related hypertensive rat models have stated that the enhanced responsiveness of the mesenteric vascular bed to α1-adrenoceptor agonists could be due to a local alteration in the α1-adrenoceptor density [23].Results of the present study show that an elevated dietary sodium intake enhances the sensitivity of the renal cortical vasculature to α1-adrenergic agonists. Thus aside from the mechanisms already mentioned in relation to the high sodium associated change in the vascular reactivity, our study suggest that an augmented response of α1-adrenergic mechanism to adrenergic stimuli in the renal cortical vasculature could also contribute as one of the factor in the salt-related alterations of the renal vascular resistance in SHR.
Further, we observed that, NA, PE, and ME-induced renal cortical vasoconstriction were enhanced by both low and high dose of CEC in the second and third phase compared to the corresponding control (saline) in SHRNNa. The exact mechanism underlying this type of response is not completely understood, but, there is a possibility that the CEC antagonism of postsynaptic α1B-ARs interacts and enhances the sensitivity of other α1-adrenergic receptor subtypes to the exogenously administered NA, PE and ME, [16,24] resulting in a larger postsynaptic response in the renal cortical vasculature in SHRNNa. Similar CEC-mediated augmentation of renal vasoconstrictor response to adrenergic vasoconstrictor stimuli has also been reported in the pathophysiological state, such as in rats with renal impairment and in normotensive WKY rats subjected to high-sodium diet [11,16].
In contrast, the response to CEC of the SHRHNa group was comparatively different. Low dose of CEC significantly enhanced the renal cortical vasoconstrictor response to NA, and PE, the reason can be the above mentioned possibilty, while high dose of CEC attenuated this exaggerated renal cortical vasoconstrictor response to NA and PE. In addition CEC as a α1B-AR alkylating agent also caused significant attenuation of ME-induced renal cortical vasoconstriction in both phases of renal vasoconstrictor experiment in SHRHNa; these results suggests that there is a functional presence of α1B-AR in the mediation of adrenergically induced renal cortical vasoconstriction in SHRHNa. The involvement of α1B-ARs in renal vasoconstriction is concurrent with the results obtained from recent studies on WKY rats subjected to high-sodium diet and DOCA-salt hypertensive rats [11,25]. This has also been reported in other pathological conditions such as heart failure [10] ,diabetes, renal failure, and experimental early diabetic nephropathy [16,24]. This study is the first to acknowledge the involvement of α1Badrenoceptors in renal cortical vasoconstriction in SHR subjected to high sodium diet. Further it was also observed that both SHRNNa and SHRHNa showed significant fall in the baseline levels of MAP in third
phase, this could be due to spill over of some of the antagonist into the systemic circulation, however in these cases the baseline values of RCP was not altered in all the three phase in both experimental groups. Thus this fall in MAP has no effect on the renal vasoconstrictor response to the agonist.
A number of studies have indicated that the antidiuretic and antinatriuretic effects in response to low-frequency renal nerve stimulation are mediated by α1-ARs [26,27]. In the present study, PE produced marked reductions in UFR, UNaV and FENa in both SHRNNa and SHRHNa groups. These observations further strengthen some earlier findings which reports that α1-AR activation, either by renal nerve stimulation or exogenously administered α1-AR agonist, leads to antidiuresis and antinatriuresis [14,28] . Phenyephrine is adrenergic agonist used to stimulate the alpha 1 adrenoreceptors. The possible use of phenyephrine in the present study to stimulate α1B-ARs is due to the lack of a selective α1B-ARs agonist to directly evaluate the interaction of CEC with the α1B-ARs in vivo and is a limitation of the present study. Renal arterial infusion of PE did not bring any major changes in MAP and RAP in both SHRNNa and SHRHNa groups, indicating that the antidiuretic and antinatriuretic responses to PE was not due to alterations in systemic hemodynamics. In addition, PE infusion were associated with a decrease in GFR in SHRNNa and SHRHNa and a similar decrease in RCP only in SHRNNa but none of these values reach a statistically significant level. However there was a significant (P<0.05) reduction of RCP in SHRHNa because of PE infusion in the second phase. Giving that we did not measure the medullary blood flow this decrease may not have been responsible for the observed antidiuretic and antinatriuretic responses, as RCP did not represent the total renal blood flow. One could attribute the observed antinatriuresis and antidiuresis response to PE is induced by activation of renal α1ARs. However, CEC abolished the antinatriuretic and antidiuretic response to PE in SHRNNa, thus indicating that α1B-ARs are involved in the mediation of tubular antidiuresis and antinatriuresis of SHRNNa. On the other hand, CEC as a selective α1B-ARs antagonist, failed to significantly abolish the antidiuretic and antinatriuretic responses of PE in SHRHNa groups; accordingly the notion that, α1B-ARs could have been responsible for the observed antinatriuretic and antidiuretic responses is considered unlikely. However there is still a need to identify the roles of other α1-AR subtypes. Nonetheless, it can be suggested that high salt intake in SHR has no major influence on the functionality of α1B-ARs in the regulation of various renal tubular functions such as UFR, U V and FENa Na.
Conclusion Detrimental effect of high sodium diet cannot be attributed only to
rise in MAP, in the present study, independent of any further increase in MAP; a high-sodium diet in SHR has increases the sensitivity of renal cortical vascular smooth muscle α1-adrenergic responsiveness to adrenergic agonists. In addition irrespective of dietary sodium changes, α1-ARs are involved in the mediation of adrenergicallyinduced antinatriuresis, antidiuresis. Results also shows that, α1B-ARs are the functional subtypes involved in mediating the adrenergically induced renal cortical vasoconstriction in the SHRHNa and the tubular functions in SHRNNa, while they play a minimal role in mediating renal tubular functions in SHRHNa. Thus, high-sodium diet in SHRs did not influence the functionality of α1B-ARs in the regulation of tubular functions like UFR, UNaV and FENa, and there is a need to identify the role of other α1-AR subtypes.
J Autacoids ISSN:2161-0479 Autacoids, an open access journal

Citation: Kazi RN, Sattar MA, Abdullah NA, Khan MA, Johns EJ (2012) The role of Α1b-Ars in Regulations of Renal Hemodynamic and Renal Functions in Hypertensive Rats; Impact of High Dietary Sodium Intake. J Autacoids 1:101. doi:10.4172/2161-0479.1000101
Page 6 of 6
Volume 1 • Issue 2 • 1000101J Autacoids ISSN:2161-0479 Autacoids, an open access journal
Disclosures Conflict of interest statement
We have no financial, consultant, institutional and other relationships that might lead to bias or a conflict of interest.
Experimental investigation
No human subjects were used in the present research work.
Acknowledgement
Islamic development bank (IDB), Jeddah, Saudi Arabia is greatly acknowledged for providing merit scholarship programme for high technology. This work was supported by a research grant to Munavvar Zubaid Abdul Sattar from University Sains Malaysia, Penang, Malaysia is greatly acknowledged.
References
1. Houston MC (1986) Sodium and Hypertension: A Review. Arch Intern Med 146: 179-185.
2. Tobian L (1991) Salt and hypertension. Lessons from animal models that relate to human hypertension. hypertension 1: 152-158.
3. Michel MC, Insel PA, Brodde OE (1989) Renal alpha-adrenergic receptor alterations: a cause of essential hypertension? FASEB J 3:139-144.
4. DiBona GF, Kopp UC (1997) Neural control of renal function. Physiol Rev 77: 75-197.
5. García-Sáinz JA, Vázquez-Prado J, del Carmen Medina L (2000) Alpha1Adrenoceptors: function and phosphorylation. Eur J Pharmaco 389:1-12.
6. Salomonsson M, Brannstrom K, Arendshorst WJ (2000) Alpha (1)-adrenoceptor subtypes in rat renal resistance vessels: in vivo and in vitro studies. Am J Physiol Renal Physiol 278: 138-147.
7. Canessa LM, Piccio MM, Vachvanichsanong P, Sidhu A, Porter CC, et al. (1995) Alpha 1B-adrenergic receptors in rat renal microvessels. Kidney Int 48: 1412-1419.
8. Elhawary A, Pang CC (1994) Alpha 1b-adrenoceptors mediate renal tubular sodium and water re-absorption in the rat. Br J Pharmacol 111: 819-824.
9. Khan MA, Sattar MA, Abdullah NA, Johns EJ (2008) Alpha1B-adrenoceptors mediate adrenergically-induced renal vasoconstrictions in rats with renal impairment. Acta Pharmacol Sin 29: 193-203.
10. Abbas SA, Munavvar AS, Abdullah NA, Johns EJ (2006) Involvement of α1-adrenoceptor subtypes in the cardiac failure in spontaneously hypertensive rats. Journal of basic and applied science 2: 59-69.
11. Kazi RN, Munavvar AS, Abdullah NA, Khan AH, Johns EJ (2009) Influence of high dietary sodium intake on the functional subtypes of alpha-adrenoceptors in the renal cortical vasculature of Wistar–Kyoto rats. Auto Autacoid Pharmacol 29: 25-31.
12. Tanoue A, Koba M, Miyawaki S, Koshimizu TA, Hosoda C, et al. (2002) Role of the alpha1D-Adrenegric Receptor in the Development of Salt-Induced Hypertension. Hypertension 40:101-106.
13. Sattar MA, Johns EJ (1994) Evidence for an Alpha1-adrenoceptor Subtype Mediating Adrenergic Vasoconstriction in Wistar Normotensive and Stroke-Prone Spontaneously Hypertensive Rat Kidney. J Cardiovasc Pharmacol 23: 232-239.
14. Sattar MA, Johns EJ (1996) Alpha-1 adrenoceptor subtypes involved in mediating adrenergically induced antinatriuresis and antidiuresis in two kidney, one clip Goldblatt and deoxycorticosterone acetate-salt hypertensive rats. J Pharmacol Exp Ther 277: 245-252.
15. Bojesen E (1952) A method for determination of inulin in plasma and urine. Acta Med Scand Suppl 266: 275-282.
16. Mark AL, Lawton WJ, Abboud FM, Fitz AE, Connor WE, et al. (1975) Effects of high and low sodium intake on arterial pressure and forearm vasular resistance in borderline hypertension. A preliminary report. Circ Res 36: 194-198.
17. Egan ВМ (1992) Vascular reactivity, sympathetic tone, and stress. In Johnson EH, Gentry ED, Julius S (Eds.), Personality, elevated blood pressure, and essential hypertension.
18. Lenda DM, Sauls BA, Boegehold MA (2000) Reactive oxygen species may contribute to reduced endothelium-dependent dilation in rats fed high salt. Am J Physiol Heart Circ Physiol 279: H7-14.
19. Ni Z, Vaziri ND (2001) Effect of salt loading on nitric oxide synthase expression in normotensive rats. Am J Hypertens 14: 155-163.
20. Redgrave J, Rabinowe S, Hollenberg NK, Williams GH (1985) Correction of abnormal renal blood flow response to angiotensin II by converting enzyme inhibition in essential hypertensives. J Clin Invest 75: 1285-1290.
21. Kagota S, Tamashiro A, Yamaguchi Y, Nakamura K, Kunitomo M (2002) High Salt Intake Impairs Vascular Nitric Oxide/Cyclic Guanosine Monophosphate System in Spontaneously Hypertensive Rats. J Pharmacol Exp Ther 302: 344351.
22. Perry PA, Webb RC (1988) Sensitivity and adrenoceptor affinity in the mesenteric artery of the deoxycorticosterone acetate hypertensive rat. Can J Physiol Pharmacol 66: 1095-1099.
23. Armenia A, Munavvar AS, Abdullah NA, Helmi A, Johns EJ (2004) The contribution of adrenoceptor subtype(s) in the renal vasculature of diabetic spontaneously hypertensive rats. Br j pharmacol 142: 719-726.M
24. Pérez-Rivera AA, Fink GD, Galligan JJ (2005) Alpha-1B adrenoceptors mediate neurogenic constriction in mesenteric arteries of normotensive and DOCA-salt hypertensive mice. Auton neurosci 121: 64-73.
25. Chen C, Lokhandwala MF (1995) Identification of alpha1-adrenoceptor subtypes involved in the antinatriuretic response to intrarenal infusion of phenylephrine. Eur J Pharmacolo 287: 1-6.
26. Hesse IF, Johns EJ (1984) The effect of graded renal nerve stimulation on renal function in the anaesthetized rabbit. Comp Biochem Physiol A Comp Physiol 79: 409-414.
27. Akpogomeh BA, Johns EJ (1990) The Alphaadrenoceptor mediating the tubular actions of the renal nerves in spontaneously hypertensive and stroke-prone spontaneously hypertensive rats. J Auton Pharmacol 10: 201-212.
Submit your next manuscript and get advantages of OMICS Group submissions Unique features:
• User friendly/feasible website-translation of your paper to 50 world’s leading languages • Audio Version of published paper • Digital articles to share and explore
Special features:
• 100 Open Access Journals • 10,000 editorial team • 21 days rapid review process • Quality and quick editorial, review and publication processing • Indexing at PubMed (partial), Scopus, DOAJ, EBSCO, Index Copernicus and Google Scholar etc • Sharing Option: Social Networking Enabled • Authors, Reviewers and Editors rewarded with online Scientific Credits • Better discount for your subsequent articles
Submit your manuscript at: www.omicsonline.org/submission

















![PAPER OPEN ACCESS 3UHSDUDWLRQDQGFKDUDFWHUL ...spec-lab.ecnu.edu.cn/.../514d1fba-e0b4-43fa-95cd-35ba6a83b250.pdf · PAPER OPEN ACCESS 3UHSDUDWLRQDQGFKDUDFWHUL]DWLRQRIQDUURZEDQGJDSIHUURHOHFWULF](https://static.fdocument.org/doc/165x107/5e1b49bb1f7dfa13d250784c/paper-open-access-3uhsdudwlrqdqgfkdudfwhul-spec-labecnueducn514d1fba-e0b4-43fa-95cd-.jpg)
