
Journal of Negative Results in BioMedicine - BioMed Central
10
BioMed Central Page 1 of 10 (page number not for citation purposes) Journal of Negative Results in BioMedicine Open Access Research Wnt expression is not correlated with β-catenin dysregulation in Dupuytren's Disease David B O'Gorman 1,2 , Yan Wu 1 , Shannon Seney 1 , Rebecca D Zhu 1 and Bing Siang Gan* 1,2,3,4 Address: 1 Cell and Molecular Biology Laboratory, Hand and Upper Limb Centre, Lawson Health Research Institute, St. Joseph's Health Centre, London, Ontario, Canada, 2 Department of Surgery, University of Western Ontario, London, Ontario, Canada, 3 Department of Physiology and Pharmacology, University of Western Ontario, London, Ontario, Canada and 4 Department of Medical Biophysics, University of Western Ontario, London, Ontario, Canada Email: David B O'Gorman - [email protected]; Yan Wu - [email protected]; Shannon Seney - [email protected]; Rebecca D Zhu - [email protected]; Bing Siang Gan* - [email protected] * Corresponding author Abstract Background: Dupuytren's contracture or disease (DD) is a fibro-proliferative disease of the hand that results in finger flexion contractures. Increased cellular β-catenin levels have been identified as characteristic of this disease. As Wnts are the most widely recognized upstream regulators of cellular β-catenin accumulation, we have examined Wnt gene expression in surgical specimens and in DD-derived primary cell cultures grown in two-dimensional monolayer culture or in three- dimensional FPCL collagen lattice cultures. Results: The Wnt expression profile of patient-matched DD and unaffected control palmar fascia tissue was determined by a variety of complimentary methods; Affymetrix Microarray analysis, specific Wnt and degenerative primer-based Reverse Transcriptase (RT)-PCR, and Real Time PCR. Microarray analysis identified 13 Wnts associated with DD and control tissues. Degenerate Wnt RT-PCR analysis identified Wnts 10b and 11, and to a lesser extent 5a and 9a, as the major Wnt family members expressed in our patient samples. Competitive RT-PCR analysis identified significant differences between the levels of expression of Wnts 9a, 10b and 11 in tissue samples and in primary cell cultures grown as monolayer or in FPCL, where the mRNA levels in tissue > FPCL cultures > monolayer cultures. Real Time PCR data confirmed the down-regulation of Wnt 11 mRNA in DD while Wnt 10b, the most frequently isolated Wnt in DD and control palmar fascia, displayed widely variable expression between the methods of analysis. Conclusion: These data indicate that changes in Wnt expression per se are unlikely to be the cause of the observed dysregulation of β-catenin expression in DD. Background Dupuytren's contracture or disease (DD) is a benign fibro- proliferative disease of the hand that causes permanent finger flexion contractures [1,2]. Despite its long medical history and high prevalence among Caucasians of North- ern European ancestry, reportedly as high as 30–40% [3], the underlying genetic etiology of the disease remains unknown [4]. Numerous risk factors have been reported Published: 30 August 2006 Journal of Negative Results in BioMedicine 2006, 5:13 doi:10.1186/1477-5751-5-13 Received: 20 February 2006 Accepted: 30 August 2006 This article is available from: http://www.jnrbm.com/content/5/1/13 © 2006 O'Gorman et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Transcript of Journal of Negative Results in BioMedicine - BioMed Central

BioMed Central
Journal of Negative Results in BioMedicine
ss
Open AcceResearchWnt expression is not correlated with β-catenin dysregulation in Dupuytren's DiseaseDavid B O'Gorman1,2, Yan Wu1, Shannon Seney1, Rebecca D Zhu1 and Bing Siang Gan*1,2,3,4Address: 1Cell and Molecular Biology Laboratory, Hand and Upper Limb Centre, Lawson Health Research Institute, St. Joseph's Health Centre, London, Ontario, Canada, 2Department of Surgery, University of Western Ontario, London, Ontario, Canada, 3Department of Physiology and Pharmacology, University of Western Ontario, London, Ontario, Canada and 4Department of Medical Biophysics, University of Western Ontario, London, Ontario, Canada
Email: David B O'Gorman - [email protected]; Yan Wu - [email protected]; Shannon Seney - [email protected]; Rebecca D Zhu - [email protected]; Bing Siang Gan* - [email protected]
* Corresponding author
AbstractBackground: Dupuytren's contracture or disease (DD) is a fibro-proliferative disease of the handthat results in finger flexion contractures. Increased cellular β-catenin levels have been identified ascharacteristic of this disease. As Wnts are the most widely recognized upstream regulators ofcellular β-catenin accumulation, we have examined Wnt gene expression in surgical specimens andin DD-derived primary cell cultures grown in two-dimensional monolayer culture or in three-dimensional FPCL collagen lattice cultures.
Results: The Wnt expression profile of patient-matched DD and unaffected control palmar fasciatissue was determined by a variety of complimentary methods; Affymetrix Microarray analysis,specific Wnt and degenerative primer-based Reverse Transcriptase (RT)-PCR, and Real Time PCR.Microarray analysis identified 13 Wnts associated with DD and control tissues. Degenerate WntRT-PCR analysis identified Wnts 10b and 11, and to a lesser extent 5a and 9a, as the major Wntfamily members expressed in our patient samples. Competitive RT-PCR analysis identifiedsignificant differences between the levels of expression of Wnts 9a, 10b and 11 in tissue samplesand in primary cell cultures grown as monolayer or in FPCL, where the mRNA levels in tissue >FPCL cultures > monolayer cultures. Real Time PCR data confirmed the down-regulation of Wnt11 mRNA in DD while Wnt 10b, the most frequently isolated Wnt in DD and control palmar fascia,displayed widely variable expression between the methods of analysis.
Conclusion: These data indicate that changes in Wnt expression per se are unlikely to be the causeof the observed dysregulation of β-catenin expression in DD.
BackgroundDupuytren's contracture or disease (DD) is a benign fibro-proliferative disease of the hand that causes permanentfinger flexion contractures [1,2]. Despite its long medical
history and high prevalence among Caucasians of North-ern European ancestry, reportedly as high as 30–40% [3],the underlying genetic etiology of the disease remainsunknown [4]. Numerous risk factors have been reported
Published: 30 August 2006
Journal of Negative Results in BioMedicine 2006, 5:13 doi:10.1186/1477-5751-5-13
Received: 20 February 2006Accepted: 30 August 2006
This article is available from: http://www.jnrbm.com/content/5/1/13
© 2006 O'Gorman et al; licensee BioMed Central Ltd.This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Page 1 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
for DD, including alcoholism, trauma, diabetes, smoking,and epilepsy, but their exact role in the disease is not clear[5]. Epidemiological studies show an increased total mor-tality and cancer mortality rates among men with estab-lished DD [6], suggesting the pathophysiology of thisdisease may overlap with that of certain cancers.
β- catenin, the central component of the 'canonical' Wntsignalling pathway (herein referred to as Wnt/β-catenin)has been implicated in the pathogenesis of DD [7-9], andabnormal β-catenin levels in primary DD cell cultureshave been shown to vary with specific cell culture condi-tions [8,9]. β-Catenin plays both a structural role, as a cad-herin-binding protein in cell adhesion junctions [10,11],and a signalling role, as part of the Wnt/β-catenin path-way [12]. Wnts are a large family of lipid modified glyco-proteins [13] that regulate various cellular processesimportant to normal embryonic development [14]. Wntsact as paracrine factors, initiating cell signalling by bind-ing to Frizzled (Fz) receptors. The Wnt/Fz complex canthen activate one of three distinct signalling pathways thatcontrol either cell fate or differentiation (Wnt/β-cat-enin)[14], planar cell polarity (PCP)[15], or cell adhesion(Wnt/Ca+2/PKC)[16,17]. The co-receptor LRP5/6 (lipo-protein receptor-related proteins 5 or 6) is required forWnt/β-catenin pathway signalling [18-20]. Once acti-vated, the Wnt/Fz/LRP complex triggers a cascade of sig-nalling events that ultimately lead to the stabilization of a'cadherin-free' cytoplasmic pool of β-catenin. The cyto-plasmic accumulation of β-catenin results in its transloca-tion to the nucleus where it functions as a transcriptionalactivator for members of the lymphoid enhancer factor/T-cell factor (Lef/Tcf) family of DNA binding proteins[21,22].
The importance of the Wnt/β-catenin signalling is under-scored by its targeted disruption in human diseases. Forexample, several members of the Wnt/β-catenin pathwayare mutated in a variety of human malignancies [23-27].Normally, in the absence of a 'canonical' Wnt signal or anactivating mutational event, the cytoplasmic 'free' pool ofβ-catenin becomes serine/threonine phosphorylated,ubiquitinated (Ub) and degraded in the proteasome, viaan axin-based 'destruction' complex. Axin with the aid ofAPC (adenomatous polyposis coli) binds to β-catenin[28], which facilitates its phosphorylation [29] via a dualkinase mechanism involving CKI (casein kinase-1) andGSK-3β (glycogen synthase kinase-3β) [30-32]. CKI,which is recruited to the destruction complex by the axinbinding protein diversin [33], phosphorylates β-catenin atserine 45, an important priming step required by GSK-3βto mediate β-catenin phosphorylation at threonine 41,serine 37 and serine 33. This hyper-phosphorylated formof β-catenin is then recognized by the F-box containingprotein slimb/β-TrCP, a component of the E3 ubiquitin
(Ub) ligase complex, and β-catenin is targeted for degra-dation via the 26S proteasome [34-38]. Not surprisingly,the critical serine/threonine residues of β-catenin that arephosphorylated by GSK-3β are mutational 'hot spots' inmany cancers. We have previously shown that, unlike thesituation in tumors, this region (exon 3) of the β-cateningene derived from DD samples does not contain suchmutations [8]. Given the proposed role of Wnt/β-cateninsignalling in DD, in this paper, we set out to examine Wntexpression in DD.
Utilizing multiple approaches, we demonstrate here thatmultiple Wnts are expressed within patient lesions andcontrol normal palmar fascia (PF) tissue. The pattern ofWnt expression observed in tissue samples is altered by invitro culture method. Comparison of Wnt mRNA levels inDD and control tissues as well as examination of primarycultures of DD cells reveal that the level and type of Wntexpression is highly variable in this fibroproliferative dis-ease with the only consistent finding being down-regu-lated Wnt 11 mRNA expression in disease tissue. As Wnt-11 signalling is independent of β-catenin and no otherWnt family members display consistent alteration inexpression, this data suggests other, as yet unidentified,factors are dysregulating β-catenin processing in DD.
ResultsAffymetrix microarray analysisThe primary goal of this project was to determine theexpression status of all Wnts expressed in DD and controlpalmar fascia (PF). To achieve this, surgically resected DDand control patient samples were examined using Affyme-trix Microarray as described in the methods. As shown inFigure 1, 13 of the 19 Wnts (for a review of Wnt factors:http://www.stanford.edu/~rnusse/wntgenes/humanwnt.html) were identified as being expressed in DD. Dataindicated that the majority of Wnts detected in both DDand control were expressed at very similar levels with onlyWnts 5a and 11 displaying any variance in expressionbetween disease and control tissues.
Wnt expression profiling in vivo and in vitro using Wnt specific primersTo confirm the Microarray data and to assess the relativecontribution of each of these 13 Wnts in DD and controlPF, a Wnt expression profiling study was initiated utilizingWnt degenerate primer analysis. Prior to commencingthese experiments, however, it was necessary to determinethe optimal samples for analysis. We have previouslyreported that β-catenin levels are abnormal in DD tissuebut not in primary cell isolates grown in two-dimensionalmonolayer culture, whereas three-dimensional culture ofthe same primary cell lines in FPCL can recapitulateabnormal β-catenin expression [8,9]. The initial experi-ments involved the isolation of total RNA from surgically
Page 2 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
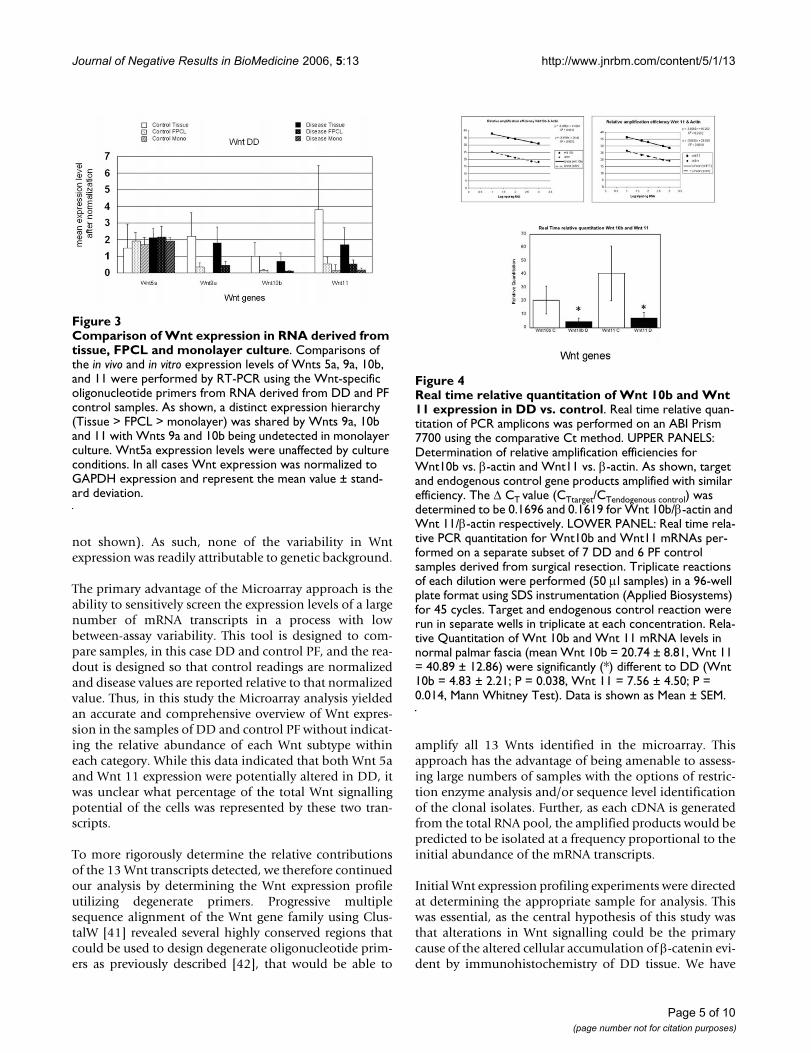
resected patient samples and primary cell cultures growneither in two-dimensional monolayer culture or in three-dimensional FPCL culture as described in the methods.Total RNA samples were then reverse transcribed and thecDNA templates were amplified by PCR using Wnt degen-erate oligonucleotide primers (Fig. 2). Clones of the PCRamplified products were isolated, and the cloned insertsfrom purified plasmid DNA digested with a series of diag-nostic restriction enzymes to identify the correspondingWnt subtype as described in the methods. Clones werecategorized based on the Wnt-specific DNA fragmentsizes, and several representative clones from each groupsequenced. The clones isolated from control and diseasePF tissue displayed a similar Wnt expression pattern, withWnt 5a, 9a, 10b, and 11 accounting for almost all of theidentifiable disease and control clones detected by thisapproach (as shown in Table 1). Two clones representingWnts 8b and 9b were also isolated. Comparisons of the invivo and in vitro expression levels of the major group ofWnts (5a, 9a, 10b, and 11) were performed by RT-PCRusing the Wnt-specific oligonucleotide primers asdescribed in the methods. As shown in Figure 3, three ofthe four Wnts (5a, 9a and 11) were readily detectablewithin FPCL cultures, while only two Wnts (5a and 11)were detected in confluent monolayer cultures. Wnt5aexpression levels were relatively high for both monolayerand FPCL cultures and quite similar to the levels seen invivo. However, comparisons between the in vivo and invitro expression levels of Wnt 9a, 10b and 11 showed sig-
nificant differences. Specifically, a distinct expressionhierarchy (in vivo > FPCL > confluent monolayer) wasshared by Wnts 9a, 10b and 11. Wnt10b expression wasvery low or undetectable in the majority of the FPCL cul-tures and largely undetectable in the confluent monolayercultures.
Based on this data, we concluded that these in vitro cul-ture conditions do not recapitulate in vivo Wnt expressionand that only RNA isolated directly from surgical speci-mens would be suitable to obtain an accurate representa-tion of Wnt expression in DD.
Wnt expression profiling in DD and control palmar fascia by reverse transcription and degenerate primer PCR analysisIn light of the results from the in vitro and in vivo studiesdescribed above, a comprehensive Wnt expression profil-ing study was performed on RNA isolated directly fromsurgical specimens utilizing reverse transcription and Wntdegenerate primer PCR analysis. PCR products (~400 bp)were subcloned into the pCR® 4-TOPO® sequencing vectorand clones were isolated as described in the methods. TheDNA restriction analysis was performed and a representa-tive subgroup of clones were sequenced to confirm Wntidentity. In total, 182 clones were isolated and identifiedby restriction enzyme analysis to yield a representativeoverview of Wnt expression in DD and control palmar fas-cia. As shown in Table 1, Wnts 5a, 9a, 10b, and 11accounted for all clonal isolates. Specifically, Wnts 5a, 9a,10b, and 11 represented 18%, 19%, 14% and 49% of thecontrol clones (n = 57), and 15%, 2%, 73% and 12% ofthe disease clones (n = 125), respectively (total 182).
Real Time PCR of Wnt 10b and Wnt 11 expression in DD and control palmar fasciaWnts 10b and 11 represented the majority of clonal iso-lates from DD and control tissues and their representationwithin each group appeared to correlate with the presenceof disease. To better compare the expression levels of theseWnts, we performed Real Time PCR of Wnt 10b and Wnt11 mRNA in surgical samples of DD and normal PF con-trol using the relative quantitation method. As shown inFigure 4, both Wnt 10b and Wnt 11 mRNA levels were sig-nificantly decreased in total RNA derived from DD cord(Mann-Whitney Test, Wnt 10b P = 0.038, Wnt 11 P =0.014). The expression levels of Wnt10b did not reflectthe representation in the clones derived from the degener-ate primer analysis or the Affymetrix Microarray analysis.Wnt 11 mRNA expression, by comparison, was lower inDD tissue than control palmar fascia by all of the tech-niques employed. With the exception of Wnt 11, directcomparison of the Wnt expression analysis reported hereindicated that the level and type of Wnt expression is notaltered between DD and control PF.
Affymetrix microarray of Wnt expressionFigure 1Affymetrix microarray of Wnt expression. 3 DD and 3 control samples were analyzed using the Human Genome U133 Plus 2.0 Array. The data generated represents the expression analysis from all samples. The data was sorted and normalized using Genespring software. The control value (mean of three samples relative to normalization of control #1 and corresponding disease value (mean of three samples) is shown beneath each Wnt designation on the X axis. The graph displays the mean values of each group ± standard deviation.
Page 3 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
DiscussionWnt expression is altered in a variety of cancers includingprostate and colon carcinoma [39,40], where β-catenin
accumulation and associated gene transcription arebelieved to contribute to the tumor growth. As we andothers have previously shown that abnormal cytoplasmicaccumulation of β-catenin is present in DD, we havehypothesized that β-catenin is a central molecular media-tor of its pathophysiology [7-9]. As Wnt factors are themost upstream regulating factors of β-catenin levels, wehave undertaken a comprehensive analysis of Wnt expres-sion in this disease.
Of the 19 Wnts identified to-date, 13 were detected byAffymetrix Microarray analysis in both DD and PF controltissue. Normalized expression analysis indicated that themajority of Wnts were expressed at equivalent levels inboth disease and control samples with only Wnt 11 andWnt 5a displaying any suggestion of altered expression. Alarge standard deviation was noted in the control samplesfor Wnt 11 (mean 1.29, SD 0.72) and the disease samplesfor Wnt5a (mean 2.62, SD 1.05). The Wnt expression ofthe control sample derived from a patient undergoing car-pal tunnel release was not significantly different from theWnt expression of the control samples derived from pal-mar fascia adjacent to disease cord in all cases includingWnt 11, where it was the sample closest to the mean (data
Table 1: Wnt expression profiling of surgical fascia tissue specimens.
Patient Fascia Type Wnt 5a Wnt 9a Wnt 10b Wnt 11
D2 Control 5/37 9/37 3/37 20/37Pooled Control 4/17 0/17 5/17 8/17W Control 1/3 2/3 0/3 0/3D Disease 0/28 1/28 27/28 0/28D2 Disease 1/4 0/4 2/4 1/4E Disease 0/16 0/16 16/16 0/16M Disease 0/9 0/9 9/9 0/9H2 Disease 0/16 0/16 16/16 0/16L Disease 0/2 1/2 1/2 0/2R Disease 2/9 0/9 7/9 0/9T Disease 5/22 1/22 10/22 6/22W Disease 11/19 0/19 3/19 8/19
In total, 9 disease, 7 control or normal fascia specimens (2 individual + 5 that were 'pooled' together) and were subjected to RT-PCR using degenerate Wnt primers. Four different Wnt species were identified; Wnt5a, 9a, 10b and 11 which were isolated from 18%, 19%, 14% and 49% of the control clones (n = 57), and 15%, 2%, 73% and 12% of the disease clones (n = 125), respectively (total 182)
Degenerate primer designFigure 2Degenerate primer design. Degenerative Wnt primers (designated Upper and Lower Primers) were designed using the ClustalW program with the aid of the BioEdit sequence alignment editor. Comparison of the published nucleic acid sequences for the 13 human Wnts detected by Affymetrix Microarray revealed two highly conserved regions that were used to design degenerate oligonucleotide primers for PCR amplification as shown.
Page 4 of 10(page number not for citation purposes)
Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
not shown). As such, none of the variability in Wntexpression was readily attributable to genetic background.
The primary advantage of the Microarray approach is theability to sensitively screen the expression levels of a largenumber of mRNA transcripts in a process with lowbetween-assay variability. This tool is designed to com-pare samples, in this case DD and control PF, and the rea-dout is designed so that control readings are normalizedand disease values are reported relative to that normalizedvalue. Thus, in this study the Microarray analysis yieldedan accurate and comprehensive overview of Wnt expres-sion in the samples of DD and control PF without indicat-ing the relative abundance of each Wnt subtype withineach category. While this data indicated that both Wnt 5aand Wnt 11 expression were potentially altered in DD, itwas unclear what percentage of the total Wnt signallingpotential of the cells was represented by these two tran-scripts.
To more rigorously determine the relative contributionsof the 13 Wnt transcripts detected, we therefore continuedour analysis by determining the Wnt expression profileutilizing degenerate primers. Progressive multiplesequence alignment of the Wnt gene family using Clus-talW [41] revealed several highly conserved regions thatcould be used to design degenerate oligonucleotide prim-ers as previously described [42], that would be able to
amplify all 13 Wnts identified in the microarray. Thisapproach has the advantage of being amenable to assess-ing large numbers of samples with the options of restric-tion enzyme analysis and/or sequence level identificationof the clonal isolates. Further, as each cDNA is generatedfrom the total RNA pool, the amplified products would bepredicted to be isolated at a frequency proportional to theinitial abundance of the mRNA transcripts.
Initial Wnt expression profiling experiments were directedat determining the appropriate sample for analysis. Thiswas essential, as the central hypothesis of this study wasthat alterations in Wnt signalling could be the primarycause of the altered cellular accumulation of β-catenin evi-dent by immunohistochemistry of DD tissue. We have
Real time relative quantitation of Wnt 10b and Wnt 11 expression in DD vsFigure 4Real time relative quantitation of Wnt 10b and Wnt 11 expression in DD vs. control. Real time relative quan-titation of PCR amplicons was performed on an ABI Prism 7700 using the comparative Ct method. UPPER PANELS: Determination of relative amplification efficiencies for Wnt10b vs. β-actin and Wnt11 vs. β-actin. As shown, target and endogenous control gene products amplified with similar efficiency. The Δ CT value (CTtarget/CTendogenous control) was determined to be 0.1696 and 0.1619 for Wnt 10b/β-actin and Wnt 11/β-actin respectively. LOWER PANEL: Real time rela-tive PCR quantitation for Wnt10b and Wnt11 mRNAs per-formed on a separate subset of 7 DD and 6 PF control samples derived from surgical resection. Triplicate reactions of each dilution were performed (50 μl samples) in a 96-well plate format using SDS instrumentation (Applied Biosystems) for 45 cycles. Target and endogenous control reaction were run in separate wells in triplicate at each concentration. Rela-tive Quantitation of Wnt 10b and Wnt 11 mRNA levels in normal palmar fascia (mean Wnt 10b = 20.74 ± 8.81, Wnt 11 = 40.89 ± 12.86) were significantly (*) different to DD (Wnt 10b = 4.83 ± 2.21; P = 0.038, Wnt 11 = 7.56 ± 4.50; P = 0.014, Mann Whitney Test). Data is shown as Mean ± SEM.
Comparison of Wnt expression in RNA derived from tissue, FPCL and monolayer cultureFigure 3Comparison of Wnt expression in RNA derived from tissue, FPCL and monolayer culture. Comparisons of the in vivo and in vitro expression levels of Wnts 5a, 9a, 10b, and 11 were performed by RT-PCR using the Wnt-specific oligonucleotide primers from RNA derived from DD and PF control samples. As shown, a distinct expression hierarchy (Tissue > FPCL > monolayer) was shared by Wnts 9a, 10b and 11 with Wnts 9a and 10b being undetected in monolayer culture. Wnt5a expression levels were unaffected by culture conditions. In all cases Wnt expression was normalized to GAPDH expression and represent the mean value ± stand-ard deviation.
Page 5 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
previously reported that this accumulation of β-catenin inDD was not detected by western blotting of primary cellisolates grown in two-dimensional monolayer culture [8].Three-dimensional cultures of the same primary cell linesin Fibroblast Populated Collagen Lattices (FPCL), how-ever, were able to qualitatively reflect in vivo β-cateninexpression due, at least in part, to the addition of appro-priate isometric tension [9]. As this study was inherentlyquantitative, it was of interest to determine the effect ofisometric tension and three-dimensional FPCL culture onthe level of Wnt expression and to determine how Wntexpression level in FPCL compared to Wnt expression invivo.
The analysis of clones by restriction analysis and sequenc-ing revealed that Wnts 5a, 9a, 10b, and 11 accounted forthe vast majority of the disease and control-derived clonesthat could be identified by this approach. Comparisons ofthe in vivo and in vitro expression levels of these Wnts (5a,9a, 10b, and 11) were performed by RT-PCR using theWnt-specific oligonucleotide primers. Three of the fourWnts (5a, 9a and 11) were readily detectable within FPCLcultures, while only two Wnts (5a and 11) were detectedin confluent monolayer cultures. Wnt 5a expression levelswere relatively high for both monolayer and FPCL cul-tures and quite similar to the levels seen in vivo. However,comparisons between the in vivo and in vitro expressionlevels of Wnt 9a, 10b and 11 showed significant differ-ences. Specifically, a distinct expression hierarchy (in vivo> FPCL > confluent monolayer) was shared by Wnts 9a,10b and 11. Wnt10b expression was very low or undetec-table in the majority of the FPCL cultures and largelyundetectable in the confluent monolayer cultures.
Based on these data, we concluded that these in vitro cul-ture conditions do not recapitulate in vivo Wnt expressionand that RNA isolated directly from surgical specimens isrequired to obtain an accurate representation of Wntexpression in DD. A comprehensive Wnt expression pro-file of 182 clones was generated utilizing reverse transcrip-tion and Wnt degenerate primer PCR analysis. As shownin Table 1, Wnt 5a was evident in 15% of DD- derivedclones and 18% of control derived clones, indicating thatthere was no difference in expression level between thesegroups. In contrast, the relative abundance of clones con-taining Wnts 10b and 11, and to a lesser extent 9b,sequences varied considerably between DD and controls.Wnt 11 was evident in 49% of control tissue – derivedclones but only 12% of those derived from disease tissue,whereas Wnt 10b displayed the opposite trend beingpresent in only 14% of control-derived clones comparedto 73% of DD-derived clones. Wnt10b is recognized toprimarily signal via the "canonical" pathway leading todownregulation of GSK-3β activity and cytoplasmic accu-mulation of β-catenin [43]. Wnt 11, while less well char-
acterized, has been shown to be associated with the "non-canonical" cell adhesion (Wnt/Ca+2/PKC) pathway [44].As this data could imply a shift to increased Wnt 10bexpression and decreased Wnt 11 expression in DD, con-sistent with an increase in signalling through the canoni-cal pathway, it was essential to independently confirmthese data.
To achieve this, Wnt 10b and Wnt 11 expression werequantitated by Real Time PCR on an additional subset of7 DD and 6 PF control samples derived directly from sur-gical resection. As shown in Figure 4, these results indicatethat Wnt 10b expression was significantly decreasedbetween disease and control samples. The lack of correla-tion between Wnt 10b expression in the Affymetrix Micro-array analysis, which indicated no change between DDand control samples, the degenerate primer analysis,which indicated an increased abundance of cDNAsderived from Wnt 10b mRNA, and the decreased Wnt 10bmRNA levels revealed by Real Time PCR data, was unex-pected. These contradictory findings dictate that an exclu-sion of changes in Wnt 10b mRNA expression in thepathophysiology of DD cannot be made at this time. Wnt11 mRNA expression was also variable, however in thiscase a consistent decrease in expression between diseaseand control samples was evident in all of the analyticaltechniques employed. Wnt 11 is reported to signalthrough a non-canonical pathway that does not affect β-catenin accumulation [44,45]. The absence of consistentchanges in the mRNA expression of any other Wnts iden-tified in DD that could lead to a shift to canonical Wnt sig-nalling indicates that, accounting for variability betweenindividual samples, there is no evidence of a consistentalteration in Wnt mRNA expression in DD that wouldalter β-catenin accumulation.
While Wnt expression is unchanged, it should be notedthat these data do not rule out alterations in Wnt signal-ling in the pathogenesis of this disease. Wnt signallingefficiency has been shown to be altered in a variety oftumors secondary to changes in the expression of secretedWnt antagonists, such as the Dickkopf family, Wnt inhib-itory factor-1 and secreted Frizzled-related protein (sFRP)family [46-48]. Our Affymetrix Microarray data indicatedthat there was a consistent reduction in sFRP-1 expressionin the 3 DD samples screened relative to PF control levels(data not shown). It is possible, therefore, that alterationsin total (rather than individual) Wnt signalling activitymay be affected by down-regulation of sFRP-1 in DD.Importantly, post-translational modification of Wnt/β-catenin signalling pathway components, such as alteredphosphorylation, acetylation, methylation, ubiquitina-tion, sumoylation, glycosylation or lipidation, could con-tribute to an altered responses of DD cells to upstreamsignalling molecules. In addition, while we have shown
Page 6 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
that exon 3 mutations of the β-catenin gene are not evi-dent in DD [8], we have not ruled out the possibility thatchanges to other regions of this or other genes encodingintegral components of this pathway such as APC may bealtering the sensitivity of DD cells to Wnt signalling. Weare presently assessing the activity of GSK-3β in DD todetermine if upstream signalling molecules are regulatingkinase activity and cytoplasmic stability of β-catenin inthis fibroproliferative disorder.
ConclusionWnt gene expression was shown to be affected by in vitroculture, both in routine monolayer culture and in threedimensional culture in FPCL, when assessed by RT-PCR.Affymetrix Microarray analysis, degenerate primer analy-sis and Real Time relative PCR quantitation were per-formed and, with the exception of Wnt 10b whereexpression was highly variable between analyses, the dataindicate that overall Wnt expression is unchangedbetween samples derived from surgical specimens of DDand those derived from normal PF. As such, alterations inthe expression levels of individual Wnt subtypes areunlikely to be contributing to the observed dysregulationof β-catenin expression in DD.
MethodsClinical specimen collectionDD patient specimens (normal and disease PF) were col-lected in compliance with the University's HumanResearch Ethics Committee. Disease cords and nodules(disease) and uninvolved normal fascia (control) werecollected from patients undergoing surgical resection ofDD lesions as following: All samples used in the currentstudy were from primary resections. In the operatingroom, the superficial surface of the affected palmar fasciaincluding a surrounding area of normal appearing fasciawas widely exposed. The diseased part was subsequentlyresected with a cuff of normal appearing palmar fascia. Onthe side table, the diseased cord was then removed fromthe resected specimen and immediately sent to the lab. Inaddition, an area of uninvolved normal appearing fascia(control) well-away from the resected specimen was har-vested. To incorporate a "true normal", palmar fascia frompatients who do not have any signs of Dupuytren's diseasewas harvested. Thus, one sample from a patient undergo-ing carpal tunnel release was also utilized in the Affyme-trix Microarray study. The majority of clinical specimenswere processed immediately for total RNA isolation asdescribed below, unless otherwise indicated, in whichcase samples were stored at -80°C prior to processing.
Affymetrix microarray analysisSurgical specimens were transferred to dry ice immedi-ately after clinical dissection. Approximately 100 mg tis-sue samples were minced into small pieces on dry ice and
then snap-frozen in Liquid Nitrogen. Tissue samples wereground in a mortar and pestle and 1 ml TRIzol reagent(Invitrogen Canada Inc., Burlington, Ontario) was addeduntil all the powder dissolved. Samples were transferredto 1.5 ml microcentrifuge tubes and total RNA was iso-lated using TRIzol procedure. Total RNA was purified andstabilized using RNeasy Minikit (Qiagen Inc., Missis-sauga, Ontario). Aliquots (3 μl) were screened using anAgilent 2100 Bioanalyzer (Agilent Technologies, Missis-sauga, Ontario) and high quality RNA samples were sub-mitted to the London Regional Genomic Center http://www.lrgc.ca for Microarray analysis on a Human GenomeU133 Plus 2.0 Array (Affymetrix, Santa Clara, CA). Thedata generated represents the expression analysis from allsamples (3 DD compared to 3 control consisting of 2 PFand 1 carpal tunnel release). The data was analyzed usingGenespring software (Agilent Technologies, Mississauga,Ontario).
Primary cell monolayer culturePrimary cell cultures were established as previouslydescribed [9,49]. Initially, primary cell cultures weregrown in starter media containing α-MEM (Gibco, Invit-rogen Corporation), 20% fetal bovine serum (FBS, Clon-tech Laboratories, Palo Alto, CA), penicillin G +streptomycin sulfate, and fungizone (Gibco, InvitrogenCorporation). Established primary cell cultures weremaintained in α-MEM + 10% FBS + antibiotics + fungi-zone at 37°C in a humidified chamber with 5% CO2.
Fibroblast populated collagen lattice (FPCL) culturesCollagen lattices were prepared as a modified version ofthe method described previously [8]. Briefly, phosphatebuffered solution (PBS) suspensions of primary cell cul-tures (passages 2 – 6) were mixed with a neutralized solu-tion of Vitrogen100 (Collagen Corp, Santa Clara, CA,USA) collagen type I matrix (8 parts Vitrogen100, 2.9 mg/ml + 1 part 10× α-MEM + 1 part HEPES buffer, pH 9.0).The cell-collagen concentrations were adjusted with PBSto attain a final collagen concentration of 2.0 mg/ml anda final cell concentration of 105 cells/ml of matrix. Thecell-collagen mixture was then dispensed into 24 well cul-ture dishes (0.5 ml/well) that were pre-treated with a PBSsolution containing 2% (w/v) bovine serum albumin(BSA). Following FPCL polymerization, 0.5 ml of growthmedium (α-MEM, 10% FBS) was added on top of each lat-tice. After 4 days of culture the attached FPCLs were har-vested for RNA extraction using the RNeasy® columnsaccording to the manufacturer's instructions (Qiagen,Mississauga, ON, Canada).
Degenerate Wnt primer designProgressive multiple sequence alignment of the Wnt genefamily was carried out using the ClustalW program withthe aid of the BioEdit sequence alignment editor [41].
Page 7 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
Comparison of the published nucleic acid sequences forthe 13 human Wnts detected by Affymetrix Microarrayrevealed several highly conserved regions that were usedto design degenerate oligonucleotide primers for PCRamplification as previously described [42].
RNA extraction and PCR amplification of Wnt genes using degenerate oligonucleotide primersRNA extraction from tissues was carried out as previouslydescribed. First-strand cDNA was reverse transcribed (RT)from 1 mg of total RNA using random hexamer primingand SuperScript™ II reverse transcriptase in a final volumeof 50 ml, as recommended by the manufacturer (Invitro-gen, Carlsbad, CA, USA). Following first strand synthesis,cDNA was amplified using the following degenerate oli-gonucleotide primers:
(UP) upper primer 5'-GGGGAATTCANCGAVTSYAART-GYCAY-3'
(LP) lower primer 5' – AAAAGATCTGCACCACCAVYG-SWM-3'
where V = ACG, N = ACGT, R = AG, M = AC, Y = CT, H =ACT, W = AT, S = CG, K = TG and B=CGT
Standard PCR amplification was performed in a PX2 Ther-mal Cycler (Thermo Electron Corporation) using 4 μl ofRT cDNA, 1 mM MgCl2, 200 μM of dNTPs (Invitrogen,Carlsbad, CA, USA), 1 μM of primers, and 1 unit of Plati-num® Taq DNA Polymerase (Invitrogen, Carlsbad,CA,.USA) in Taq PCR buffer (10 mM Tris pH8.3, 50 mMKCl). PCR cycling parameters were 94°C for 5 min, fol-lowed by 40 cycles of 94°C for 1 min., 59°C for 1.5 min.,72°C for 2 min., followed by a final extension at 72°C of10 min.
Cloning of PCR products and screening of transformantsPCR products (~400 bp) were directly subcloned into thepCR® 4-TOPO® sequencing vector using a TOPO TA clon-ing kit, as recommended by the manufacturer (Invitro-gen). Ligation products were then used to transformTOP10 chemically competent E. coli cells, and transform-ants selected for using standard ampicillin (100 μg/ml)supplemented LB agar plates. Plasmid DNA was purifiedfrom selected colonies using Qiagen mini-prep columns(Qiagen, Mississauga, ON, Canada). Plasmid DNA wasquantified by spectrophotometry (OD260), digested withEcoRI (pCR4-TOPO subcloned amplicons are flanked byEcoRI sites), and subjected to 2% agarose gel electrophore-sis. DNA bands were gel purified (Qiagen gel extractionkit) and then digested with a series of diagnostic restric-tion endonuclease (RE), including Eco RI, Eco RII and SacII. DNA fragments were then size-separated using native(non-denaturing) acrylamide (12%) gel electrophoresis.
Gels were stained with ethidium bromide, placed on a UVtransilluminator (FisherBiotech) and photographed usingPolaroid film (type 667) and a Polaroid DS-34 camera fit-ted with a UV filter (Polaroid, UK). The DNA fragmenta-tion patterns were analyzed (i.e. RE grouping), and severalclones from each RE group were sequenced to confirmtheir Wnt identity.
RT-PCR using Wnt-specific primersBriefly, 200 ng of total RNA from tissue or cells werereverse transcribed using random hexamers and Super-Script™ II RT as described above (Invitrogen, Carlsbad,CA, USA). Following first strand synthesis, Wnt specificPCR amplifications were carried out using 1 μl of cDNA,1.5 mM MgCl2, 200 μM of dNTP mix (Invitrogen,Carlsbad, CA, USA), 0.8 μM of primers (see below), and0.75 units of Platinum® Taq DNA Polymerase (Invitrogen,Carlsbad, CA., USA) in Taq PCR buffer (10 mM TrispH8.3, 50 mM KCl), in a final reaction volume of 25 μl.Sequences of the Wnt-specific primer were as following:
Wnt5a upper primer (Wnt5a-UP): 5' – aagaagtgcacgga-gatcgt – 3'
Wnt5a lower primer (Wnt5a-LP): 5' – tggaacctacccatcccata– 3'
Wnt9a upper primer (Wnt9a-UP): 5' – gcaagcatctgaag-cacaag – 3'
Wnt9a lower primer (Wnt9a-LP): 5' – tgctctcgcagttcttctca– 3'
Wnt10b upper primer (Wnt10b-UP): 5' – ctggtgctgctatgt-gctgt – 3'
Wnt10b lower primer (Wnt10b-LP): 5' – cccagccaaaaggag-tatga – 3'
Wnt11 upper primer (Wnt11-UP): 5' – tgacctcaagacccga-tacc – 3'
Wnt11 lower primer (Wnt11-LP): 5' – tgagggtccttgagca-gagt – 3'
GAPDH upper primer: 5' – gtcagtggtggacctgacct – 3'
GAPDH lower primer: 5' – aggggtctacatggcaactg – 3'
PCR cycling parameters were optimized for Wnt5a,Wnt9a, Wnt10b, Wnt11 and GAPDH to ensure log linearamplification of these products. For Wnt5a, Wnt9a,Wnt10b, Wnt11 were 94°C for 3 min, followed by 32cycles of 94°C for 30 sec., 62°C for 45 sec., 72°C for 45sec., followed by a final extension at 72°C of 7 min. PCR
Page 8 of 10(page number not for citation purposes)

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
conditions for GAPDH were the same as above except 25cycles were used. PCR samples were then subjected to 2%agarose gel electrophoresis, with the resulting ampliconsbeing visualized by ethidium bromide staining. Digitalimages of the gels were captured and analyzed using a geldocumentation workstation (Alpha Innotech Corp., SanLeandro, CA, USA).
Real Time PCRReal time PCR was performed on an ABI Prism 7700(Applied Biosystems, Foster City, CA. USA) using the rel-ative quantitation, or "comparative Ct" method. In brief,TRIzol reagent (Invitrogen) and RNeasy® Mini Kits (Qia-gen, Mississauga, ON) were used to isolate total RNA fromsurgically resected normal and diseased tissue frompatients with Dupuytren's Contracture. RNA quality wasdetermined on an Agilent 2100 Bioanalyzer and onlysamples with minimal degradation were used for analysis.10 μg of total RNA was reverse transcribed into cDNA firststrand using the High-Capacity cDNA Archive Kit(Applied Biosystems) in accordance with the manufac-turer's instructions. For validity, the comparative Ctmethod requires that the amplification efficiency of thetarget and endogenous control transcripts be equivalent.To assess this, dilutions of cDNA first strand correspond-ing to 1000 ng, 500 ng, 100 ng, 50 ng, 10 ng and 0 ng wereintroduced into the PCR reactions. Coupled with the tar-get genes Wnt 10b and Wnt 11, endogenous control geneproducts were amplified by gene specific probe sets con-taining TaqMan® MGB probes labelled with 6-FAM™(Applied Biosystems, Wnt 10b Assay Id Hs00559664_m1;Wnt 11 Assay ID Hs00182986_m1). Triplicate reactionsof each dilution were performed (50 μl samples) in a 96-well plate format using SDS instrumentation (AppliedBiosystems) for 45 cycles. Target and endogenous controlreaction were run in separate wells in triplicate at eachconcentration. To determine if the target and endogenouscontrol gene products amplified with the same efficiency,the Δ CT value (CT target/CT endogenous control) was plottedagainst the log input cDNA to create a semi-log regressionline. A "pass" value of <0.2 for the slope of Δ CT vs. loginput was used in this study. β-actin amplification passedvalidation with Δ CT vs. log input values of 0.1696 and0.1619 for Wnt 10b and Wnt 11 respectively, and this con-trol was subsequently used for all real time relative quan-titation in this study.
Competing interestsThe author(s) declare that they have no competing inter-ests.
Authors' contributionsYW, SS and RDZ carried out (RT-) PCR, DNA microarrayand cell culture, as well as first interpretation of the data.DBO and BSG are responsible for study conception and
design, coordinated the entire project, performed finalinterpretation of the data and completed the manuscript.All authors read and approved the final manuscript.
AcknowledgementsRebecca D. Zhu is the recipient of a Canadian Institutes of Health Research Summer Research Studentship. BSG is the recipient of a Canadian Institutes of Health Research Short-Term Clinician Investigator Grant. BSG also holds a Clinician Scientist Salary Award from the Dept. of Surgery at the University of Western Ontario and a Salary Award from the Dean's Fund at the Schulich School of Medicine and Dentistry at the University of West-ern Ontario. Financial support for this work was provided by the Canadian Institutes of Health Research, the Lawson Health Research Institute Inter-nal Research Fund, the US Plastic Surgery Educational Foundation, and the LHRI Advanced Surgical Technologies Research Group.
References1. Dupuytren G: Permanent retraction of the fingers, produced
by an affection of the palmar fascia. Lancet 1834, 2:222-225.2. Rayan GM: Clinical presentation and types of Dupuytren's dis-
ease. Hand Clin 1999, 15(1):87-96, vii.3. Gudmundsson KG, Arngrimsson R, Sigfusson N, Bjornsson A, Jons-
son T: Epidemiology of Dupuytren's disease: clinical, serolog-ical, and social assessment. The Reykjavik Study. J ClinEpidemiol 2000, 53(3):291-296.
4. Burge P: Genetics of Dupuytren's disease. Hand Clin 1999,15(1):63-71.
5. Ross DC: Epidemiology of Dupuytren's disease. Hand Clin 1999,15(1):53-62, vi.
6. Gudmundsson KG, Arngrimsson R, Sigfusson N, Jonsson T:Increased total mortality and cancer mortality in men withDupuytren's disease. A 15-year follow-up study. J Clin Epidemiol2002, 55(1):5-10.
7. Montgomery E, Lee JH, Abraham SC, Wu TT: Superficial fibroma-toses are genetically distinct from deep fibromatoses. ModPathol 2001, 14(7):695-701.
8. Varallo VM, Gan BS, Seney S, Ross DC, Roth JH, Richards RS, McFar-lane RM, Alman B, Howard JC: Beta-catenin expression inDupuytren's disease: potential role for cell-matrix interac-tions in modulating beta-catenin levels in vivo and in vitro.Oncogene 2003, 22(24):3680-3684.
9. Howard JC, Varallo VM, Ross DC, Roth JH, Faber KJ, Alman B, GanBS: Elevated levels of beta-catenin and fibronectin in three-dimensional collagen cultures of Dupuytren's disease cellsare regulated by tension in vitro. BMC Musculoskelet Disord 2003,4:16.
10. Ozawa M, Baribault H, Kemler R: The cytoplasmic domain of thecell adhesion molecule uvomorulin associates with threeindependent proteins structurally related in different spe-cies. Embo J 1989, 8(6):1711-1717.
11. Nagafuchi A, Takeichi M: Transmembrane control of cadherin-mediated cell adhesion: a 94 kDa protein functionally associ-ated with a specific region of the cytoplasmic domain of E-cadherin. Cell Regul 1989, 1(1):37-44.
12. Rubinfeld B, Albert I, Porfiri E, Fiol C, Munemitsu S, Polakis P: Bind-ing of GSK3beta to the APC-beta-catenin complex and reg-ulation of complex assembly. Science 1996,272(5264):1023-1026.
13. Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, ReyaT, Yates JR, Nusse R: Wnt proteins are lipid-modified and canact as stem cell growth factors. Nature 2003,423(6938):448-452.
14. Cadigan KM, Nusse R: Wnt signaling: a common theme in ani-mal development. Genes Dev 1997, 11(24):3286-3305.
15. Wallingford JB, Rowning BA, Vogeli KM, Rothbacher U, Fraser SE,Harland RM: Dishevelled controls cell polarity during Xenopusgastrulation. Nature 2000, 405(6782):81-85.
16. Slusarski DC, Corces VG, Moon RT: Interaction of Wnt and aFrizzled homologue triggers G-protein-linked phosphatidyli-nositol signalling. Nature 1997, 390(6658):410-413.
Page 9 of 10(page number not for citation purposes)
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2788574
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2788574
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2788574
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2519616
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2519616
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2519616
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8638126
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8638126
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8638126
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9407023
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9407023
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9389482

Journal of Negative Results in BioMedicine 2006, 5:13 http://www.jnrbm.com/content/5/1/13
Publish with BioMed Central and every scientist can read your work free of charge
"BioMed Central will be the most significant development for disseminating the results of biomedical research in our lifetime."
Sir Paul Nurse, Cancer Research UK
Your research papers will be:
available free of charge to the entire biomedical community
peer reviewed and published immediately upon acceptance
cited in PubMed and archived on PubMed Central
yours — you keep the copyright
Submit your manuscript here:http://www.biomedcentral.com/info/publishing_adv.asp
BioMedcentral
17. Winklbauer R, Medina A, Swain RK, Steinbeisser H: Frizzled-7 sig-nalling controls tissue separation during Xenopus gastrula-tion. Nature 2001, 413(6858):856-860.
18. Wehrli M, Dougan ST, Caldwell K, O'Keefe L, Schwartz S, Vaizel-Ohayon D, Schejter E, Tomlinson A, DiNardo S: arrow encodes anLDL-receptor-related protein essential for Wingless signal-ling. Nature 2000, 407(6803):527-530.
19. Tamai K, Semenov M, Kato Y, Spokony R, Liu C, Katsuyama Y, HessF, Saint-Jeannet JP, He X: LDL-receptor-related proteins in Wntsignal transduction. Nature 2000, 407(6803):530-535.
20. Pinson KI, Brennan J, Monkley S, Avery BJ, Skarnes WC: An LDL-receptor-related protein mediates Wnt signalling in mice.Nature 2000, 407(6803):535-538.
21. Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R,Birchmeier W: Functional interaction of beta-catenin with thetranscription factor LEF-1. Nature 1996, 382(6592):638-642.
22. van de Wetering M, Cavallo R, Dooijes D, van Beest M, van Es J,Loureiro J, Ypma A, Hursh D, Jones T, Bejsovec A, Peifer M, MortinM, Clevers H: Armadillo coactivates transcription driven bythe product of the Drosophila segment polarity gene dTCF.Cell 1997, 88(6):789-799.
23. Korinek V, Barker N, Morin PJ, van Wichen D, de Weger R, KinzlerKW, Vogelstein B, Clevers H: Constitutive transcriptional acti-vation by a beta-catenin-Tcf complex in APC-/- colon carci-noma [see comments]. Science 1997, 275(5307):1784-1787.
24. Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B,Kinzler KW: Activation of beta-catenin-Tcf signaling in coloncancer by mutations in beta-catenin or APC. Science 1997,275(5307):1787-1790.
25. Rubinfeld B, Robbins P, El-Gamil M, Albert I, Porfiri E, Polakis P: Sta-bilization of beta-catenin by genetic defects in melanomacell lines. Science 1997, 275(5307):1790-1792.
26. Polakis P: Wnt signaling and cancer. Genes Dev 2000,14(15):1837-1851.
27. Giles RH, van Es JH, Clevers H: Caught up in a Wnt storm: Wntsignaling in cancer. Biochim Biophys Acta 2003, 1653(1):1-24.
28. Behrens J, Jerchow BA, Wurtele M, Grimm J, Asbrand C, Wirtz R,Kuhl M, Wedlich D, Birchmeier W: Functional interaction of anaxin homolog, conductin, with beta- catenin, APC, andGSK3beta. Science 1998, 280(5363):596-599.
29. Yost C, Torres M, Miller JR, Huang E, Kimelman D, Moon RT: Theaxis-inducing activity, stability, and subcellular distributionof beta-catenin is regulated in Xenopus embryos by glycogensynthase kinase 3. Genes Dev 1996, 10(12):1443-1454.
30. Amit S, Hatzubai A, Birman Y, Andersen JS, Ben-Shushan E, Mann M,Ben-Neriah Y, Alkalay I: Axin-mediated CKI phosphorylation ofbeta-catenin at Ser 45: a molecular switch for the Wnt path-way. Genes Dev 2002, 16(9):1066-1076.
31. Liu T, Malbon CC, Wang HY: Activation of the b-catenin/Lef-Tcfpathway is obligate for formation of primitive endoderm bymouse F9 totipotent, teratocarcinoma cells in response toretinoic acid. J Biol Chem 2002, 12:12.
32. Yanagawa S, Matsuda Y, Lee JS, Matsubayashi H, Sese S, Kadowaki T,Ishimoto A: Casein kinase I phosphorylates the Armadillo pro-tein and induces its degradation in Drosophila. Embo J 2002,21(7):1733-1742.
33. Schwarz-Romond T, Asbrand C, Bakkers J, Kuhl M, Schaeffer HJ,Huelsken J, Behrens J, Hammerschmidt M, Birchmeier W: Theankyrin repeat protein Diversin recruits Casein kinase Iepsi-lon to the beta-catenin degradation complex and acts inboth canonical Wnt and Wnt/JNK signaling. Genes Dev 2002,16(16):2073-2084.
34. Aberle H, Bauer A, Stappert J, Kispert A, Kemler R: beta-catenin isa target for the ubiquitin-proteasome pathway. Embo J 1997,16(13):3797-3804.
35. Orford K, Crockett C, Jensen JP, Weissman AM, Byers SW: Serinephosphorylation-regulated ubiquitination and degradationof beta-catenin. J Biol Chem 1997, 272(40):24735-24738.
36. Hart M, Concordet JP, Lassot I, Albert I, del los Santos R, Durand H,Perret C, Rubinfeld B, Margottin F, Benarous R, Polakis P: The F-boxprotein beta-TrCP associates with phosphorylated beta-cat-enin and regulates its activity in the cell. Curr Biol 1999,9(4):207-210.
37. Latres E, Chiaur DS, Pagano M: The human F box protein beta-Trcp associates with the Cul1/Skp1 complex and regulatesthe stability of beta-catenin. Oncogene 1999, 18(4):849-854.
38. Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, HarperJW: The SCFbeta-TRCP-ubiquitin ligase complex associatesspecifically with phosphorylated destruction motifs in Ikap-paBalpha and beta-catenin and stimulates IkappaBalphaubiquitination in vitro. Genes Dev 1999, 13(3):270-283.
39. Zhu H, Mazor M, Kawano Y, Walker MM, Leung HY, Armstrong K,Waxman J, Kypta RM: Analysis of Wnt gene expression in pros-tate cancer: mutual inhibition by WNT11 and the androgenreceptor. Cancer Res 2004, 64(21):7918-7926.
40. Holcombe RF, Marsh JL, Waterman ML, Lin F, Milovanovic T, TruongT: Expression of Wnt ligands and Frizzled receptors incolonic mucosa and in colon carcinoma. Mol Pathol 2002,55(4):220-226.
41. Holmes I, Bruno WJ: Evolutionary HMMs: a Bayesian approachto multiple alignment. Bioinformatics 2001, 17(9):803-820.
42. Gavin BJ, McMahon JA, McMahon AP: Expression of multiplenovel Wnt-1/int-1-related genes during fetal and adultmouse development. Genes Dev 1990, 4(12B):2319-2332.
43. Bhatia N, Spiegelman VS: Activation of Wnt/beta-catenin/Tcfsignaling in mouse skin carcinogenesis. Mol Carcinog 2005,42(4):213-221.
44. Pandur P, Lasche M, Eisenberg LM, Kuhl M: Wnt-11 activation of anon-canonical Wnt signalling pathway is required for cardio-genesis. Nature 2002, 418(6898):636-641.
45. Ryu JH, Chun JS: Opposing Roles of WNT-5A and WNT-11 inInterleukin-1beta Regulation of Type II Collagen Expressionin Articular Chondrocytes. J Biol Chem 2006,281(31):22039-22047.
46. Suzuki H, Watkins DN, Jair KW, Schuebel KE, Markowitz SD, ChenWD, Pretlow TP, Yang B, Akiyama Y, Van Engeland M, Toyota M,Tokino T, Hinoda Y, Imai K, Herman JG, Baylin SB: Epigenetic inac-tivation of SFRP genes allows constitutive WNT signaling incolorectal cancer. Nat Genet 2004, 36(4):417-422.
47. Lee AY, He B, You L, Dadfarmay S, Xu Z, Mazieres J, Mikami I,McCormick F, Jablons DM: Expression of the secreted frizzled-related protein gene family is downregulated in human mes-othelioma. Oncogene 2004, 23(39):6672-6676.
48. Klopocki E, Kristiansen G, Wild PJ, Klaman I, Castanos-Velez E, SingerG, Stohr R, Simon R, Sauter G, Leibiger H, Essers L, Weber B, Her-mann K, Rosenthal A, Hartmann A, Dahl E: Loss of SFRP1 is asso-ciated with breast cancer progression and poor prognosis inearly stage tumors. Int J Oncol 2004, 25(3):641-649.
49. Howard JC, Varallo VM, Ross DC, Faber KJ, Roth JH, Seney S, GanBS: Wound healing-associated proteins Hsp47 and fibronec-tin are elevated in Dupuytren's contracture. J Surg Res 2004,117(2):232-238.
Page 10 of 10(page number not for citation purposes)
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8757136
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8757136
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9118222
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9118222
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065401
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065401
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065401
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065402
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065402
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065403
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065403
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9065403
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9554852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9554852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9554852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8666229
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8666229
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=8666229
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9233789
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9233789
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9312064
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9312064
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9312064
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9990852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9990852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=9990852
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2279700
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Abstract&list_uids=2279700


















