
Improved protective effects of American Ginseng Berry ...
38
Accepted Manuscript Improved protective effects of American Ginseng Berry against Acetaminophen-induced liver toxicity through TNF-α-mediated caspase-3/-8/-9 signaling pathways Xing-Yue Xu , Zi Wang , Shen Ren , Jing Leng , Jun-nan Hu , Zhi Liu , Chen Chen , Wei Li PII: S0944-7113(18)30514-2 DOI: https://doi.org/10.1016/j.phymed.2018.09.234 Reference: PHYMED 52699 To appear in: Phytomedicine Received date: 10 February 2018 Revised date: 23 August 2018 Accepted date: 28 September 2018 Please cite this article as: Xing-Yue Xu , Zi Wang , Shen Ren , Jing Leng , Jun-nan Hu , Zhi Liu , Chen Chen , Wei Li , Improved protective effects of American Ginseng Berry against Acetaminophen- induced liver toxicity through TNF-α-mediated caspase-3/-8/-9 signaling pathways, Phytomedicine (2018), doi: https://doi.org/10.1016/j.phymed.2018.09.234 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Improved protective effects of American Ginseng Berry ...

Accepted Manuscript
Improved protective effects of American Ginseng Berry againstAcetaminophen-induced liver toxicity through TNF-α-mediatedcaspase-3/-8/-9 signaling pathways
Xing-Yue Xu , Zi Wang , Shen Ren , Jing Leng , Jun-nan Hu ,Zhi Liu , Chen Chen , Wei Li
PII: S0944-7113(18)30514-2DOI: https://doi.org/10.1016/j.phymed.2018.09.234Reference: PHYMED 52699
To appear in: Phytomedicine
Received date: 10 February 2018Revised date: 23 August 2018Accepted date: 28 September 2018
Please cite this article as: Xing-Yue Xu , Zi Wang , Shen Ren , Jing Leng , Jun-nan Hu , Zhi Liu ,Chen Chen , Wei Li , Improved protective effects of American Ginseng Berry against Acetaminophen-induced liver toxicity through TNF-α-mediated caspase-3/-8/-9 signaling pathways, Phytomedicine(2018), doi: https://doi.org/10.1016/j.phymed.2018.09.234
This is a PDF file of an unedited manuscript that has been accepted for publication. As a serviceto our customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, andall legal disclaimers that apply to the journal pertain.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
1
Improved protective effects of American Ginseng Berry against
Acetaminophen-induced liver toxicity through TNF-α-mediated
caspase-3/-8/-9 signaling pathways
Xing-Yue Xua, Zi Wang
a, Shen Ren
a, Jing Leng
a, Jun-nan Hu
a, Zhi Liu
a, Chen Chen
b, and Wei Li
a, c*
a College of Chinese Medicinal Materials, Jilin Agricultural University, Changchun 130118, China.
b School of Biomedical Sciences, University of Queensland, Brisbane, Queensland 4072, Australia;
c National & Local Joint Engineering Research Center for Ginseng Breeding and Development, Changchun
130118, China
*Correspondence
Professor Wei Li, College of Chinese Medicinal Materials, Jilin Agricultural University,
Changchun 130118, China. E-mail: [email protected], Tel. /Fax: +86-431-84533304.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
2
Abstract
Background: Similar to the leaves of P. Quinquefolius, American ginseng berry (AGB) is
another important part of P. Quinquefolius with alternative therapeutic potential. The liver
protection capabilities of the former have been demonstrated previously, however, the later
has not yet been evaluated.
Purpose: Based on our previous observation, the present work was designed to evaluate the
hepatic protective effects for novel mechanisms of AGB in acetaminophen (APAP)-induced
liver injury in vivo.
Study Design/Methods: All mice were divided into four groups as follows: normal group,
APAP group and APAP+AGB (150 mg/kg and 300 mg/kg) groups. AGB were orally
administered for one week before exposure to APAP (250 mg/kg). Severe liver injury was
observed and hepatotoxicity was evaluated after 24 h through evaluating the biochemical
markers, protein expressions levels and liver histopathology.
Results: Our study results clearly demonstrated that AGB pretreatment ameliorated
APAP-induced hepatic injury as evidenced by decreasing plasma alanine aminotransferase
(ALT), aspartate transaminase (AST), tumor necrosis factor α (TNF-α) and interleukin-1β
(IL-1β) compared to the APAP group. Western blotting analysis showed that pretreatment
with AGB decreased the expressions levels of TNF-α and nuclear transcription factor-κB
(NF-κB p65) in liver tissues. Meanwhile, the protein expression levels of caspases,
cytochrome c, and Bax were elevated by AGB treatment for seven days, while the protein
expression level of Bcl-2 was inhibited comparison with that in APAP group. Furthermore,
supplement of AGB resulted in increase of superoxide dismutase (SOD) and glutathione

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
3
(GSH), while decrease of malondialdehyde (MDA) content and the expression levels of
4-hydroxynonenal (4-HNE) and cytochrome P450 E1 (CYP2E1). The results of
histopathological staining demonstrated that AGB pretreatment inhibited APAP-induced
hepatocyte infiltration, congestion, and necrosis.
Conclusion: The present study demonstrated that AGB pretreatment protected liver cells
against APAP-induced hepatotoxicity through inhibition of oxidative stress, inflammation
responses via TNF-α-mediated caspase-3/-8/-9 signaling pathways.
Key words: American ginseng berry; APAP-induced liver injury; oxidative stress;
anti-inflammation; anti-apoptosis
Abbreviations:
APAP Acetaminophen SOD Superoxide dismutase
AGB American ginseng berry Caspase Cysteine aspartic acid specific protease
ALI Acute liver injury NF-κB Nuclear transcription factor-κB
AST Aspartate transaminase TNF-α Tumor necrosis factor-α
ALT Alanine aminotransferase IL-1β Interleukin-1β
GSH Glutathione CYP2E1 Cytochrome P450 E1
MDA Malondialdehyde 4-HNE 4-hydroxynonenal

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
4
1. Introduction
Acetaminophen (APAP) is commonly used as an anodyne and febrifuge, which is safe and
no side effects within therapeutic dosages. Nevertheless, the overdose APAP can induce
severe liver injuries from hepatic necrosis, fibrosis and even cirrhosis (Truong et al., 2016).
APAP-induced liver toxicity is triggered by massive accumulation of reactive metabolic
N-acetyl-p-benzoquinone imine (NAPQI), which is catalyzed by metabolism of hepatic
cytochromes P450, mainly CYP2E1 (Shanmugam et al., 2016). NAPQI is detoxified by
reacting with reduced glutathione (GSH) in the cells. Excessive NAPQI bindings to cellular
mitochondrial proteins also inhibit the mitochondrial oxidative phosphorylation, thus
generating reactive oxygen species (ROS) which leads to mitochondrial dysfunction, damage
and eventually the pathways of apoptosis are activated (Zhao et al., 2012). In addition, during
the metabolism processes, antioxidant enzymes, such as superoxide dismutase (SOD), play a
vital role in modulating the APAP-induced hepatotoxicity (Michael Brown et al., 2012).
Accumulated evidences have revealed that APAP overdose caused the transcriptional
activation of pro-inflammatory cytokines such as nuclear transcription factor-κB (NF-κB),
tumor necrosis factor α (TNF-α) and interleukin 1β (IL-1β) (Song et al., 2014). Bax, Bcl-2
and cytochrome-c were mitochondria-related apoptotic factors and playing important roles in
the cell apoptosis (Zhao et al., 2014). Caspases as members of protease family exert necessary
effects in cell apoptosis. Initiator caspases (containing 8, 9) were tightly coupled to
pro-apoptotic signals (Hsiao et al., 2014). Once activated, these caspases would activate the
downstream effector caspases-3 following by cleaving cell scaffolds, nucleoproteins and
finally induce apoptosis (Peng et al., 2016). Taken together, a continuing search for a

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
5
promising hepatoprotective medicine is warranted.
American ginseng berry (AGB) is the ripe fruit of Panax quinquefolius. Many reports have
proved that AGB exerts similar or same physiological activities as P. ginseng or P.
quinquefolius (Lin et al., 2008; Xie et al., 2004). So far, researchers have reported the
chemical constituents and biological activities of roots of American ginseng, while the studies
on its berry are scarce. In order to fully understand pharmaceutical value of American ginseng,
studies on the extracted saponin of American ginseng berry have been carried out (Sritularak
et al., 2009). A recent report indicated that AGB exerted protective effect on myocardial
ischemia by decreasing myocardial oxygen consumption and increasing coronary blood flow
in dogs (Lu et al., 2012). AGB also exerted anti-hyperglycemic effects in diabetic ob/ob mice
(Xie et al., 2002) and protective effect in chemotherapy of 5-FU on human colorectal cancer
cells (Li et al., 2009). We have showed that extracted saponins from the leaves of P.
quinquefolius (PQS) suppressed APAP-induced liver toxicity in mice (Xu et al., 2017).
Pharmacological studies have showed that AGB exerted anti-apoptotic (Wang et al., 2006b),
antioxidant (Lin et al., 2008) and anti-hyperglycemic (Xie et al., 2004) effects. Taking into
account the significant liver protection activity of American ginseng leaves, it is worth
exploring the hepatoprotective effects and mechanisms of AGB on APAP-induced liver injury.
In our study, molecular mechanisms of AGB action on APAP-induced liver injury have
been clarified, in which TNF-α-mediated caspase-3/-8/-9 signaling pathway was identified
clearly. To our knowledge, it is the first report of potential molecular mechanisms of AGB
action in protecting liver injuries against APAP-induced hepatotoxicity in mice.
2. Materials and methods

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
6
2.1. Materials and reagents
AGB extracts from the American ginseng berry was prepared and quantified by our own as
previous description (Liu et al., 2008; Wang et al., 2006a). In brief, AGB were eluted three
times with 70% ethanol, and then collecting from AB-8 resin column chromatogram. As
described in our previous work (Xu et al., 2017), the quantitative analysis of AGB was
implemented on a Hypersil ODS column using high-performance liquid chromatography
(Waters HPLC, Milford, MA, USA) at 203 nm. The contents of the crude saponins in the
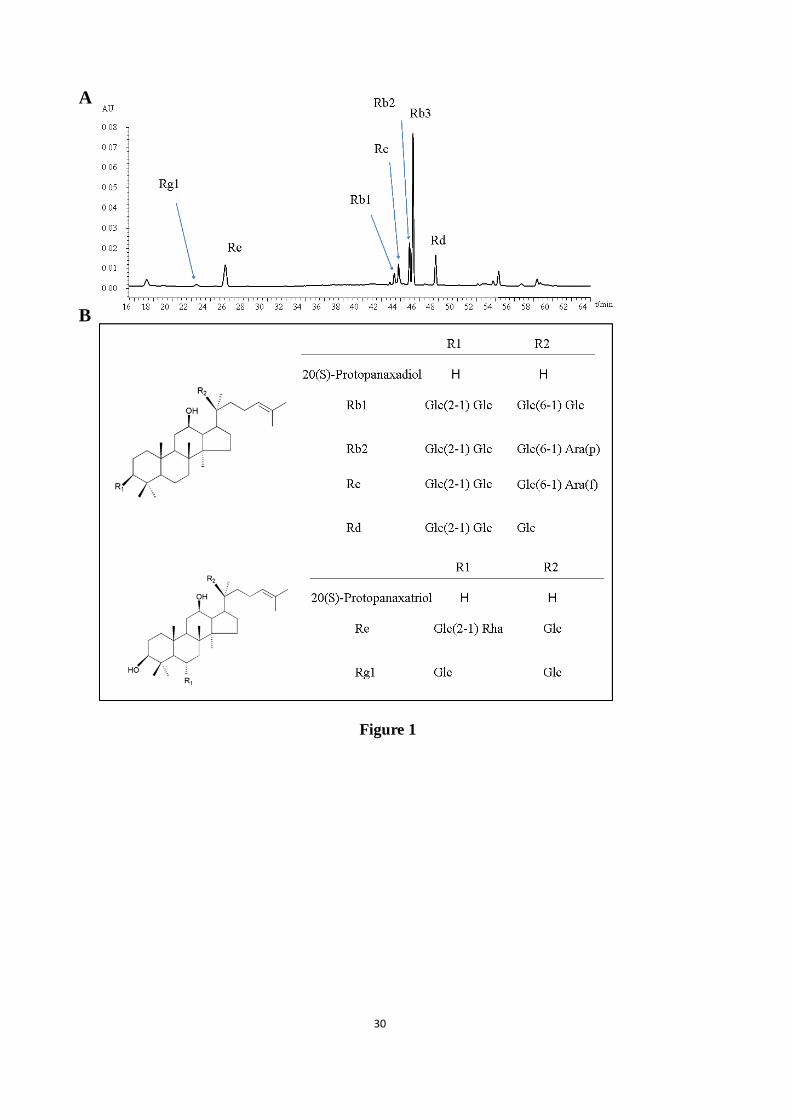
ABG were determined as follows: 0.336% Rg1, 9.107% Re, 0.504% Rb1, 8.805% Rb2,
29.523% Rb3, 3.171% Rc, and 6.022% Rd. the HPLC chromatogram and chemical structures
of seven saponins in AGB including protopanaxadiol ginsenosides (ginsenosides Rb1, Rb2,
Rb3, Rc, and Rd) and protopanaxatriol ginsenosides (ginsenosides Re and Rg1) as shown in
Fig. 1.
APAP was provided by Sigma-Aldrich (St. Louis, MO). Diagnostic kits applied to the
evaluation of ALT, AST, GSH, MDA, SOD and hematoxylin and eosin (H&E) dye kits were
obtained from Nanjing Jiancheng Institute of Biotech (Nanjing, China). Two-site sandwich
enzyme-linked immunosorbent assays (ELISA) for serum TNF-α and IL-1β were provided by
R&D systems (Minneapolis, MN, USA). Hoechst 33258 dye kit was bought from Beyotime
Biotech Co., Ltd. (Shanghai, China). The CPY2E1, 4-HNE, NF-κB (p65), phospho-p65,
TNF-α, Bax, Bcl-2, cytochrome-c, caspases-3/-8/-9, cleaved-caspases-3/-8/-9 and GAPDH
antibodies were purchased from Cell Signaling Technology, Inc (Danvers, MA, USA).
DyLight488-labeled second antibody was obtained from BOSTER Biological Technology Co.,
Ltd. (Wuhan, China). For the analysis of cell apoptosis, TUNEL cell apoptosis diagnostic kit

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
7
was obtained from Roche Applied Science (No. 11684817910). All the other chemical agents
during the study were of the highest grade commercially available.
2.2. Animals and experimental design
Adult male ICR mice (8 weeks old, 25~27g), SPF grade, were purchased from YISI
Experimental Animal Co., Ltd. with Certificate of Quality No. of SCXK (JI) 2017-0003
(Changchun, Jilin province, China). All mice were maintained 23~27°C and humidity of 50%
~70% with a 12-h light: 12-h dark cycle. All mice were continuously fed with standard chow
diet and libitum. All experimental protocols were strictly abided by the Committee of Animal
Care and Use. All animal protocols were in accordance with the Ethical Committee for
Laboratory Animals of Jilin Agricultural University (Permit No.: ECLA-JLAU-17626).
All animals were randomly divided into four groups (8, per group) as follows: normal
group, APAP-treated group, and APAP/AGB (150 mg/kg)-treated group and APAP/AGB (300
mg/kg)-treated group. AGB powder was dissolved in water. Mice were administered AGB
(150 and 300 mg/kg, respectively) by oral administration once daily for 7 days, the normal
and APAP-treated groups were managed with 0.9% normal saline only. On the 6th day, all
animals were intraperitoneally injected with 250 mg/kg APAP dissolved in warm saline
(65°C). The mice were administered saline in normal group. Subsequently, at 24 h post-APAP,
all mice were euthanized. Blood was collected and centrifuged at 3000 g, 4℃ for 15 min to
gain serum then stored at −80°C for biochemical analysis. Meanwhile, the fresh liver tissue
samples were harvested. A portion of the liver tissues immediately fixed in 10% buffered
formalin solution and embedded in paraffin for pathologic analysis. The remaining tissues
were maintained at −80°C for biochemical markers estimation.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
8
2.3. Biochemical marker assay
The contents of AST and ALT in serum were quantified using commercial available kits
(Nanjing Jiancheng Bioengineering Institute, Nanjing China) in accordance with the
manufactures’ protocols. The contents of TNF-α and IL-1β in serum were determined using
ELISA assay kits (R&D, Minneapolis, MN, USA). The absorbance was measured at 450 nm
in an ELISA reader (Bio-Rad, California, USA).
The hepatic levels of GSH, MDA and SOD were analyzed by commercial reagent kits
(Nanjing Jiancheng, Bioengineering Institute, Nanjing China) according to the manufacturer’s
protocols.
2.4. H&E and Hoechst 33258 Staining
For evaluation of hepatotoxicity, formalin-fixed liver tissues, which were embedded in
paraffin, and sliced into a thickness of 5-μm. Slices were stained with H&E according to a
commercial protocol, and subsequently performing an observation for histopathological
assessment used light microscope (Leica, DN750, Solms, Germany). These histopathological
changes would be evaluated including hepatocyte necrosis, congestion and inflammatory cell
infiltration.
As we known, Hoechst 33258 staining is commonly used for apoptosis detection which
was implemented as previously performed (Li et al., 2015). In brief, 5-μm paraffin slices were
stained by Hoechst 33258 (10 μg/ml). Stained nuclei were visualized use a fluorescent
microscope (Olympus BX-60, Tokyo, Japan). Image-Pro Plus 6.0 software (Media
Cybernetics, Maryland, USA) was applied to evaluate the degree of liver apoptosis through
quantifying the fragmented and condensed staining.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
9
2.5. TUNEL Staining
TUNEL analysis, in which apoptotic cells in the liver tissues would be precisely detected
by an situ cell death detection kit (Roche Applied Science, Germany), was implemented as
previously described (Li et al., 2016). In brief, the liver tissue slices were covered with 20
μg/mL of proteinase K for 10 min and then incubated in methanol including 3% H202 for 20
min. The slices were subsequently incubated with equilibrium buffer solution and terminal
deoxynucleotidyl transferase (TdT). Finally, the sections were incubated with anti-fluorescein
antibody. The activity of peroxidase (POD) in sections were revealed by applying
diaminobenzidine (DAB), every section was counterstained with hematoxylin. Subsequently
apoptotic cells assessment used light microscope (Leica, DN750, Solms, Germany).
2.6. Immunofluorescence Staining
Immunohistochemical analysis was executed on liver tissue sections as prior report
(Limsrichamrern et al., 2016). Specifically, the 5-μm paraffin slices were fixed to de-paraffin
and rehydrate with a series of xylene and different concentration alcohol solutions. After
antigen retrieval in citrate buffer solution (0.01 M, pH 6.0) for 8 min, the sections were
swashed with PBS and incubated with 1% BSA for 10 min. The blocking serum was casted
off, then incubated primary antibodies including CYP2E1 (1:200) and 4-HNE (1:100)
overnight at 4℃. The liver tissues were blotted with secondary antibody for 30 min at 37℃.
Then liver tissues slides were exposed to the DyLight 488-labeled (BOSTER, Wuhan, China).
Nuclear staining was implemented with DAPI. The sections were visualized using a Leica
microscope (Leica TCS SP8, Solms, Germany). The degree of liver expression was analyzed
by Image-Pro Plus 6.0 software analyzed (Media Cybernetics, Maryland, USA)

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
10
2.7. Western Blotting Analysis
Western blotting analysis was implemented as previously depicted (Noh et al., 2015).
Briefly, liver tissues were homogenized with cold RIPA buffer. After centrifuging 3 times for
10 min, the protein concentrations were analyzed using the BCA protein Trizol Reagent
(Beyotime Biotechnology, China). Equivalent protein extracts from samples were subjected to
12% SDS-PAGE and then electrophoretically transferred onto a PVDF membrane.
Phosphorylated proteins were blocked with 5% bovine serum albumin (BSA) in Tris-buffered
saline (TBS) with 0.1% Tween-20, and other proteins with 5% skimmed milk in TBS with
0.1% Tween-20 as least 1 h, and then incubated overnight at 4℃ with primary antibodies
including NF-κB p65 (1:1000), p-p65 (1:1000), TNF-α (1:1000), Bcl-2 (1:1000), Bax
(1:1000), cytochrome-c (1:1000) and caspases family (caspase-3/-8/-9 1:1000). Followed by,
the secondary antibody (1:1000) incubated for 1 h. Signals were captured with Emitter
Coupled Logic (ECL) substrate (Pierce Chemical Co., Rockford, IL, USA). Protein band
intensities were assessed with Quantity One software (Bio-Rad Laboratories, Hercules, CA,
USA).
2.8. Statistical Analysis
All of the experimental data are expressed as the means ± SD. SPSS version 17.0 for
Windows (SPSS Inc., Chicago, IL, USA) was used for the least significance difference (LDS)
multiple-comparison tests. Statistically significant differences between four groups were
evaluated by using one-way analysis of variance (ANOVA) followed by Tukey-Kramer tests.
p <0.05 or <0.01 were considered dramatically. Statistical graphs were presented by GraphPad
Prism 6.0.4. (GraphPad Software, La Jolla, CA, USA).

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
11
3. Results
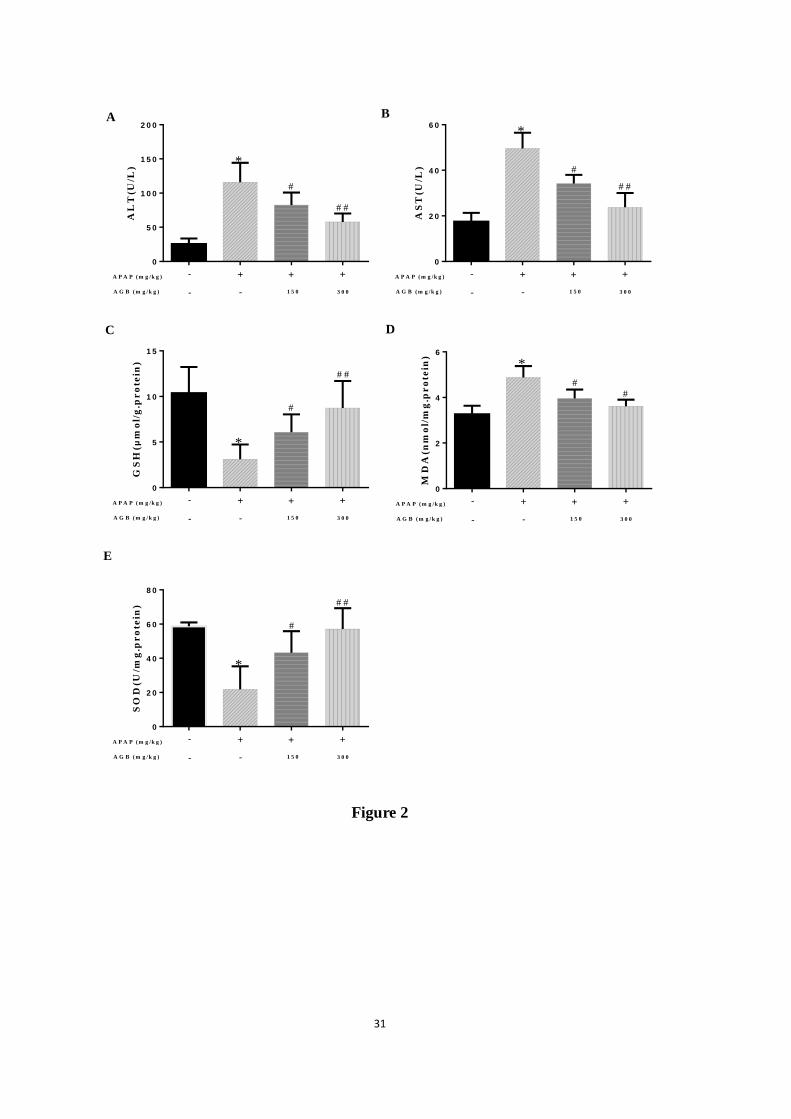
3.1. AGB pretreatment reduces AST and ALT levels after APAP treatment
To evaluate the function of AGB on liver-protection from APAP-induced hepatotoxicity, we
measured the serum levels of ALT and AST, the markers of acute liver injury in all mice. As
shown in Fig 2A and B, compared to the normal group, a single injection of 250 mg/kg APAP
significantly increased the levels of ALT and AST (p < 0.05 or p < 0.01), reflecting a severe
liver injury. However, pretreatment with AGB once daily for seven days (150 and 300 mg/kg)
reduced APAP-induced ALT and AST activities. These results indicated that AGB may exert a
protective effect in APAP-induced liver injury.
3.2. AGB pretreatment inhibits oxidative stress after APAP treatment
It has been reported that oxidative stress damage was caused by the APAP-induced
molecular mechanisms linking to acute liver failure (Nagi et al., 2010). As shown in Fig 2C,
D and E, the results indicated that GSH content and SOD activity were dramatically reduced
accompanied by an increase in lipid peroxidation product MDA by APAP treatment (p < 0.05
or p < 0.01). Interestingly, oral administration with 150 mg/kg and 300 mg/kg AGB for seven
days dramatically decreased the level of MDA and restored the controlled oxidant status as
verified by reinforce of SOD activity and suppression of GSH depletion (p < 0.05 or p < 0.01).
These results demonstrated that AGB had a remarkable antioxidant capacity against
APAP-induced live injury via increasing antioxidant enzyme activity.
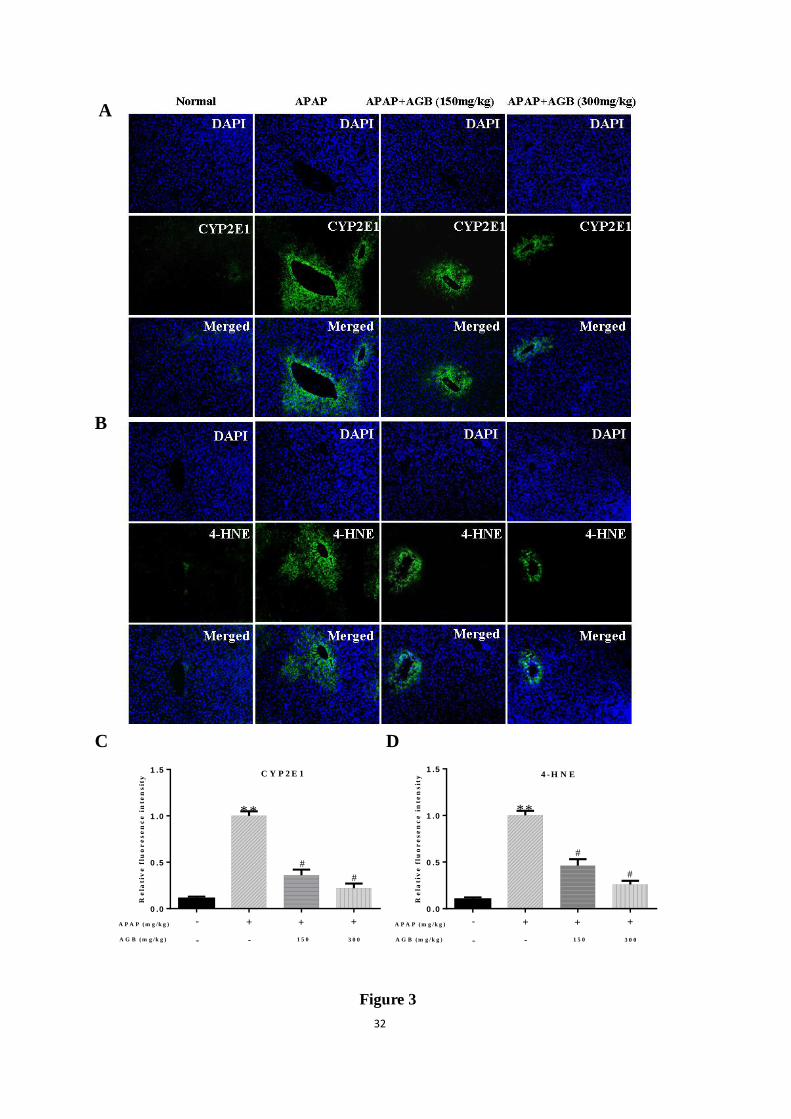
In order to further demonstrate whether oxidative stress was relevant with development of
APAP-induced hepatotoxicity in mice, lipid peroxidation was verified using 4-HNE staining.
Strong fluorescence intensities of 4-HNE were shown in the central veins of the liver tissues,

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
12
after administration APAP alone for 24 h. Nevertheless, Pretreatment AGB for one week
dramatically reduced 4-HNE positive areas, especially in the group of dosage 300 mg/kg. The
sites of lipid peroxidation were closely correlated to the necrotic areas in the liver. According
to the lipid peroxidation product MDA analysis, results showed that pretreatment with AGB
significantly reduced the increase in lipid peroxidation, as shown in Fig 3A.
APAP toxicity is triggered through its metabolic activation, which is catalyzed by
cytochrome P450 enzymes, mainly CYP2E1 (Du et al., 2015). In this study, the protein
expression level of CYP2E1 in liver tissues was determined after 24 h APAP treatment. The
expression of CYP2E1 was elevated after APAP treatment. However, pretreatment with AGB
remarkably reduced APAP-induced the expression of CYP2E1 (Fig 3B). These results
demonstrated that administration of AGB may exert a protective effect on oxidative stress
caused by APAP.
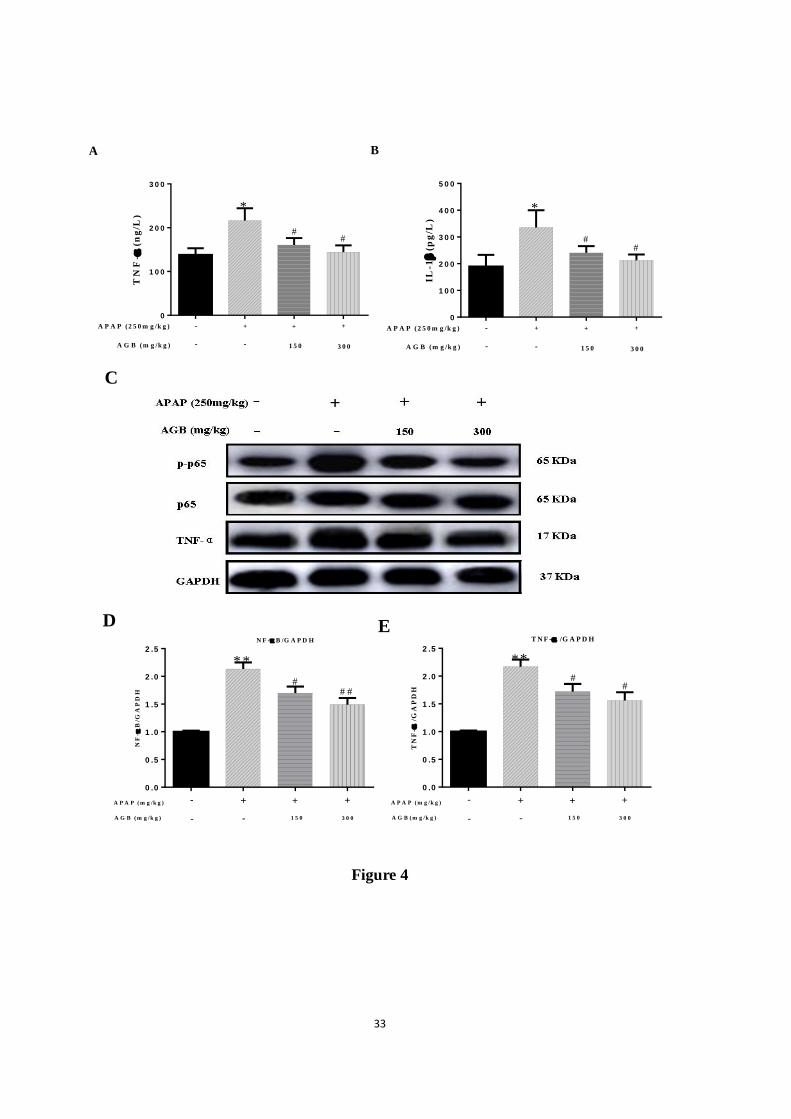
3.3. AGB pretreatment inhibits Inflammation response after APAP treatment
Prior studies reported that oxidative stress increased pro-inflammatory cytokine during the
progression of APAP-induced hepatotoxicity (Barman et al., 2016). As described in Fig 4A
and B, TNF-α and IL-1β contents were markedly increased compared with normal group after
24 h APAP injection alone (p < 0.05 or p < 0.01). In contrast, significantly increased TNF-α
and IL-1β levels following exposure to APAP were diminished by AGB pretreatment for 7
days (p < 0.05 or p < 0.01).
To gain an insight into the anti-inflammatory effects of AGB with APAP-induced liver
damage, we assessed the protein expression levels of NF-κB p65, p-p65 and TNF-α in all
groups by western blotting analysis. As depicted in Fig 4C, the results clearly showed that

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
13
exposure to APAP caused significant overproduction of pro-inflammatory cytokines
(phosphorylation of p65 and TNF-α) in the liver tissues. However, AGB pretreatment result
reduced the expressions of these pro-inflammatory proteins in the liver.
3.4. AGB pretreatment inhibits liver Histological alteration after APAP treatment
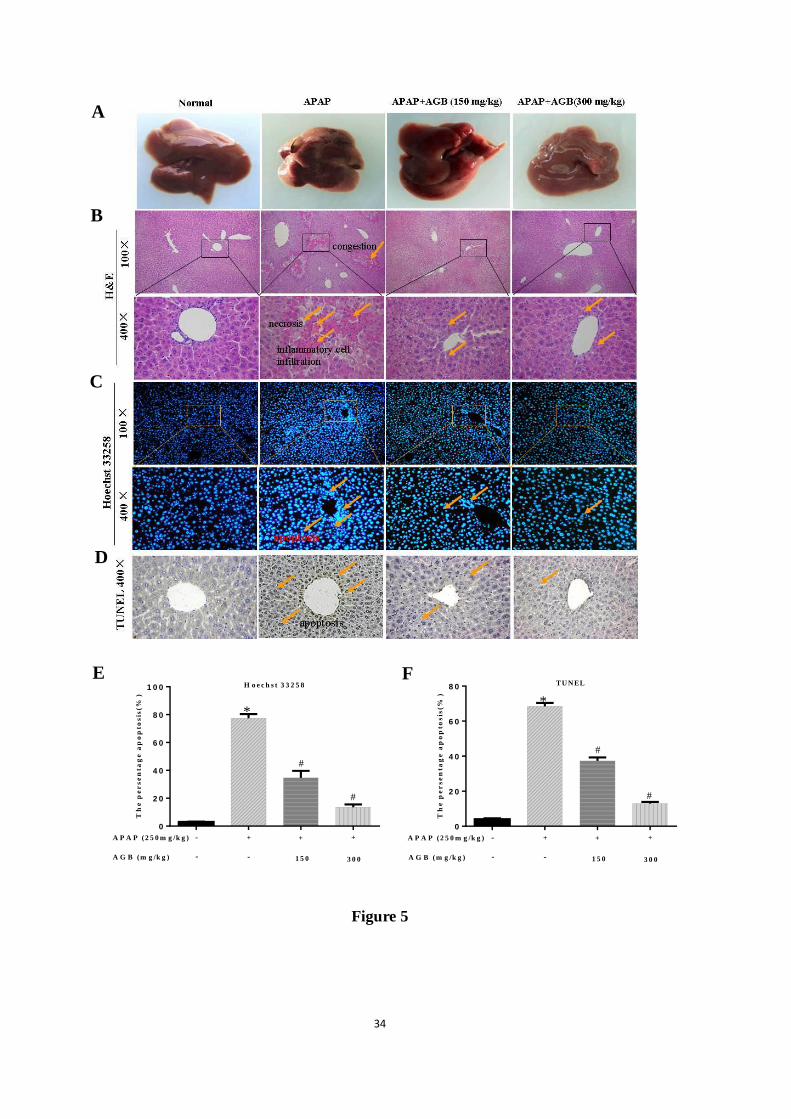
In liver tissues from 4 groups (Fig. 5A), we found that APAP treatment aroused more liver
hemorrhage than that in the normal control mice. Besides, AGB pretreatment ameliorated the
hemorrhage injury by APAP. H&E staining revealed that the livers with APAP treatment
presented more necrotic regions, congestion and inflammatory cell infiltration than that in
normal control group. Pretreatment with AGB reversed these APAP-induced hepatic lesions.
Light microscopic methods for histopathological analysis were performed on liver tissues
from all groups. As shown in Fig 5B, liver tissues in APAP treated mice showed necrosis,
congestion, and inflammatory infiltration. After treatment with low dose of AGB (150 mg/kg),
there was dramatic decrease in cytoplasm damage. Interestingly, the group treated with high
dosage AGB (300 mg/kg) after exposure to APAP markedly attenuated the hepatocyte
necrosis, inflammatory infiltration to the similar level in the normal group.
3.5. AGB pretreatment inhibits cell apoptosis after APAP treatment
To investigate whether AGB pretreatment decreased hepatocyte apoptosis induced by
APAP, Hoechst 33258 staining assay was performed to evaluate apoptosis of cells. As shown
in Fig 5C, significant nuclear fragmentation and condensation of liver cell nucleus were
observed to indicate apoptosis of liver cells in the APAP group. With AGB pretreatment, most
cell nucleus exhibited the regular homogeneous fluorescent density and normal cell structures.
To further evaluate whether there is any coexistence of apoptosis and by APAP, the liver cell

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
14
apoptosis was verified and quantified by TUNEL staining. As described in Fig 5D, compared
to normal group, TUNEL staining showed abundant cell apoptosis in APAP group. However,
AGB pretreatment (300 mg/kg) markedly reduced the number of TUNEL positive cells.
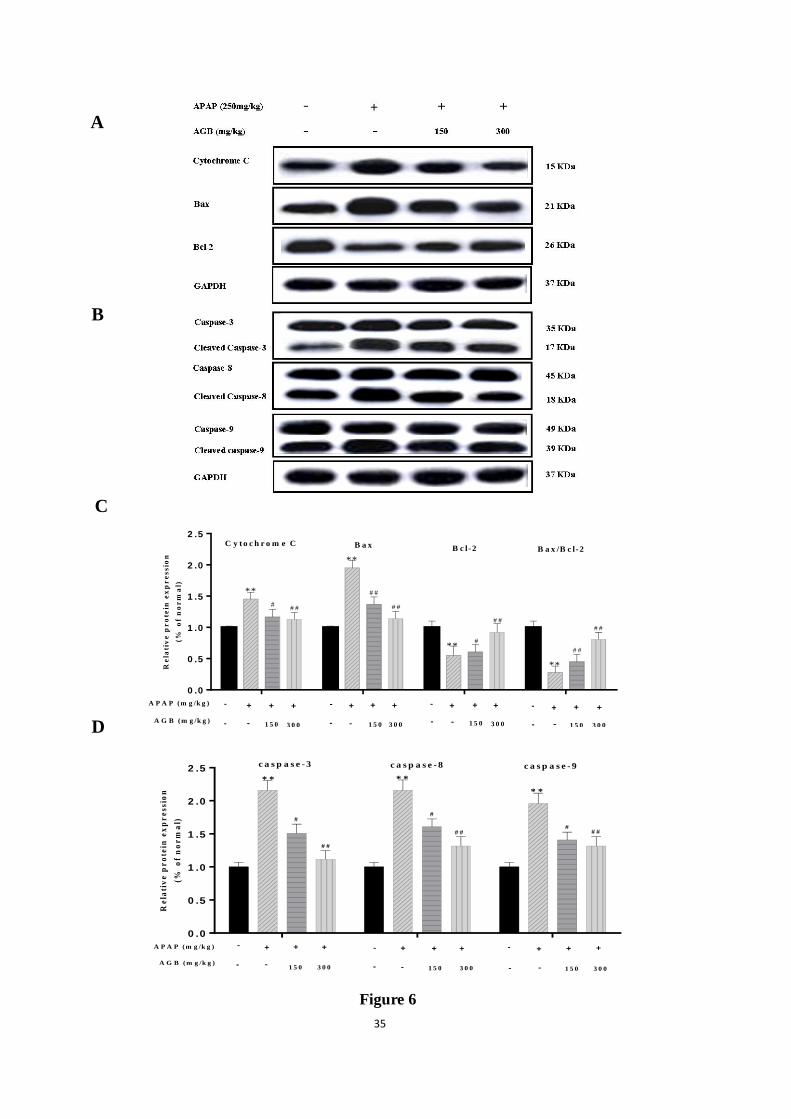
To clarify the signaling systems in this hepatocyte apoptosis, expressions of pro-apoptotic
factor Bax, cytochrome-c, the members of caspase family (including 3, 8, 9), and
anti-apoptotic factor Bcl-2 were determined in all experimental groups. As showed in Fig 6,
The results also demonstrated that pretreatment with AGB 150 mg/kg and 300 mg/kg
significantly reduced the protein expression of Bax, cytochrome-c and cleaved caspase-3/-8/-9,
and increased the protein expression of Bcl-2 compared with that in APAP alone group.
4. Discussion
APAP was commonly used as an anodyne and febrifuge agent in clinical application when
administered at therapeutic doses. However, accidental or intentional APAP overdose could
trigger acute hepatic injury and even acute liver cirrhosis (Holubek et al., 2006). To date,
N-acetylcysteine (NAC) has considered to be the first clinical antidote against APAP
hepatotoxicity. Although NAC was very effective partially for preventing APAP-induced liver
toxicity to avoid morbidity and mortality, some patients still arouse severe toxicity with
nausea, anaphylactic reaction and headaches (Saito et al., 2010). Therefore, it was necessary
to develop therapeutic options and approaches for APAP-induced liver toxicity.
It has considered that many natural extracts and bioactive compounds exert potential
protective effects against APAP-caused liver toxicity (Yuan et al., 2010; Zhang et al., 2017).
American ginseng (P. quinquefolius), a famous medicinal plant, was originated in virgin forest
of North America (Jiao et al., 2015). In the last century, American ginseng was introduced to

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
15
China for cultivation and breeding. So far, numerous scientists have made a great effort to
identify the chemical compositions and pharmacological effects of American ginseng (Yu et
al., 2014). Although many published reports have focused on its roots, there were little
systematic studies or reports on the active components and the utilization value of American
ginseng berry (Wang et al., 2006b). Like the roots, American ginseng berry also contains
similar ginsenosides. These ginsenosides mainly include protopanaxadiol-type saponins (PDS,
e.g. Rb1, Rb2, Rb3, Rc, and Rd) and a few protopanaxatriol-type saponins (PTS, e.g. Rg1 and
Re), of which PDS accounts for more than 80% of all saponins (Wang et al., 2006a).
A recent report indicated that AGB exerted protective effect on myocardial ischemia by
decreasing myocardial oxygen consumption and increasing coronary blood flow in dogs (Lu
et al., 2012). Several studies have found AGB exerted anti-hyperglycemic effects in diabetic
ob/ob mice (Xie et al., 2002) and enhanced chemopreventive effect of 5-FU on human
colorectal cancer cells (Li et al., 2009). AGB produced cardiovascular effects because of its
bioactive ingredients (Wang and Shi, 2011). Accordingly, Korean red ginseng extract mainly
including ginsenosides Rg3, prevented APAP-induced hepatotoxicity via metabolic enzyme
regulation (Gum and Cho, 2013), and panaxatriol saponin ameliorated APAP-induced liver
injury by restoring thioredoxin-1 and pro-caspase-12 (Wang et al., 2014). Importantly, our
recent results showed that saponins from the leaves of Panax quinquefolius attenuated
APAP-induced liver toxicity in mice (Xu et al., 2017). There were many reports about
experimental liver injury models, including APAP-induced (Xu et al., 2017), alcohol-induced
(Liu et al., 2016), CCl-4-induced (Xie et al., 2013) and LPS/D-GaLN-induced (Yan et al.,
2016). The hepatoprotective effects of AGB and the underlying mechanisms against

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
16
APAP-induced liver toxicity have not been elucidated yet.
This study clearly showed that AGB attenuated APAP-induced toxicity via attenuating liver
cells damage, ameliorating oxidative stress, inhibiting the expressions of apoptosis protein
and pro-inflammatory factors. The serum levels of ALT and AST were commonly referred as
liver enzymes that enter the bloodstream after hepatocyte structural integrity damage, and
were generally regarded as useful quantitative markers to reflect hepatic diseases (Yuan et al.,
2010). Our research demonstrated that the serum levels of AST and ALT remarkably increased
in mice exposed to APAP, but AGB pretreatment inhibited APAP-induced increase in AST
and ALT. Meanwhile, histological analysis demonstrated that AGB attenuated liver pathologic
changes including inflammatory infiltration and cellular necrosis and apoptosis. These results
indicated that the hepatocellular damage was reduced as a direct consequence of AGB oral
administration.
Oxidative stress played an important role in APAP-induced hepatotoxicity (Nagi et al.,
2010). APAP extensively affected hepatic metabolism and then promoted the formation of
reactive metabolite NAPQI, which induced the APAP’s toxicity, as it reacted rapidly with
GSH with depleting GSH. In APAP overdose situation, GSH became exhausted, and excess
NAPQI irreversibly adhered to mitochondrial proteins (Smith et al., 2016). As a result,
accumulation of NAPQI induced hepatocellular damage by aggravating oxidative stress in
conjunction with mitochondrial dysfunction (Kang et al., 2017). Our findings clearly
displayed the depletion of hepatic GSH in the APAP-intoxicated mice compared to the normal
group. However, pretreatment with AGB (150 and 300 mg/kg) reversed the depletion of liver
GSH. APAP metabolic activation and formation of NAPQI were mediated by cytochrome

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
17
P450-mediated enzymes, especially CYP2E1. We found overexpression of CYP2E1 in liver
tissues of APAP-treated mice reported in the previous reports. (Na et al., 2017). However,
AGB inhibited the overexpression of APAP-induced CYP2E1 expression evidenced by IHC
analyses. Furthermore, oxidative stress acted an important contribution to liver injury, which
was characterized by LPO and the accumulation of ROS (Du et al., 2016). Oxidative stress
induced by APAP was normally detoxified by the enzymatic antioxidant defense system.
Antioxidant enzymes, such as SOD, were massively depleted with APAP exposure (Wang et
al., 2017). The increase in SOD was reversed after AGB administration (150 and 300 mg/kg),
demonstrating the protective effect of AGB on APAP toxicity. The hepatic level of MDA was
generally used as a reliable biomarker of free radical mediated LPO injury (Noh et al., 2015).
The results in this investigation demonstrated that the level of hepatic MDA was remarkably
increased after the APAP challenge and these alterations were effectively alleviated by AGB
pretreatment for seven days. A study conducted by Guo et al also supported our findings (Guo
et al., 2016). In supporting MDA data, 4-HNE staining also showed an overexpression in liver
slices after exposure to APAP, and the overexpression was remarkably reduced in the groups
of AGB pretreatment.
Many pro-inflammatory mediators, like TNF-α and IL-1β, are involved in the development
of inflammatory responses. They were released due to activation of Kupffer cells by the APAP
toxic metabolite NAPQI (Legert et al., 2015). Besides, NF-κB translocation to the nucleus
was needed for the induction of cytokines that were associated with APAP-induced liver
injury (Wang et al., 2016). Inflammatory cytokines TNF-α and IL-1β were activated by the
ubiquitous NF-κB pathway and their receptors engaged in activating NF-κB (Sullivan et al.,

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
18
2014). It was reported that APAP activated the NF-κB pathway through expression of TNF-α,
IL-1β and others in a recent study (Truong et al., 2016), where TNF-α mediated the death
receptor pathway and apoptosis. In this study, the levels of TNF-α and IL-1β, and the protein
expressions of phosphorylated NF-κB and TNF-α were highly increased in the APAP-injured
mice and these increases were significantly attenuated by AGB treatment (150 and 300
mg/kg). These results may suggest that AGB exerted the protective effect against
APAP-induced liver inflammation.
Accumulating evidences demonstrated that APAP-induced liver injury was relevant to
apoptosis (Xu et al., 2017). Apoptosis acted as an important cell death mode in APAP
hepatotoxicity (Yuan et al., 2010). Mitochondria-mediated apoptosis was often accurately
regulated by the Bcl-2 family members. Bcl-2 as a prominent pro-survival protein was
binding to the mitochondrial external membrane to antagonize Bax homodimers mediated
pro-apoptosis (Joshi et al., 2011). The Bcl-2/Bax ratio was regarded as a vital index to cause
hepatocyte apoptosis (Guo et al., 2017). Our study showed that pretreatment AGB decreased
Bax and increased Bcl-2 protein expression levels as well as the Bcl-2/Bax ratio was clearly
decreased compared to that in only APAP group. Those results demonstrated that the balance
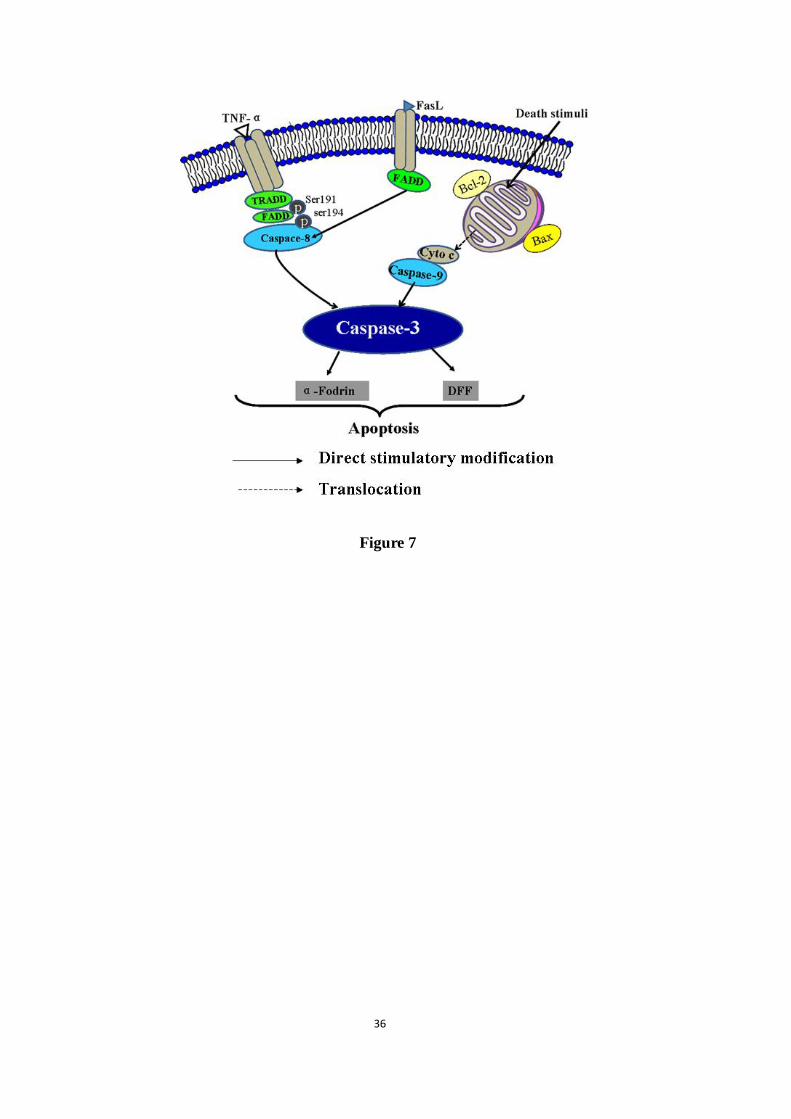
between Bcl-2 and Bax were broken. TNF-α, a significant pro-inflammatory cytokine,
induced hepatocyte apoptosis or necrosis by activating caspase-3 (Fan et al., 2014). In this
study, we demonstrated TNF-α-mediated caspase-3/-8/-9 signaling pathway. TNF-α mediated
apoptosis by recruiting death domain-containing adaptor proteins FADD and TRADD to their
receptor, which in turn leads to the activation of the initiator caspase-8. Caspase-8 cleaved and
activated cytosolic protein caspase-3 in cell apoptosis. In fact, caspase-3 functions as a central

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
19
effector to cleave various cellular substrates and trigger cell apoptosis eventually (Nagase et
al., 2002). Caspase-9 directly linked to the pathway of mitochondrial apoptosis, and it can be
activated by death stress (Peng et al., 2016). Caspase-3 was activated by caspase-9 and
involved in the mitochondrial apoptosis pathway (Zhu et al., 2012). Alternatively, caspase-3
can be directly triggered apoptosis via activation of the initiator caspase-8/-9 (Hayakawa et al.,
2008). Western blotting analyses in this report clearly revealed that APAP increased
caspase-3/8/9 in hepatocytes, which were similar to the results by Yuan et al (Yuan et al.,
2010). However, the protein expression of caspase-3/-8/-9 was dramatically suppressed after
AGB administration, indicating that AGB exerted anti-apoptotic properties in the presence of
APAP hepatotoxicity. These data indicated that AGB remarkably reduced apoptosis of
hepatocytes in APAP-induced hepatotoxicity. Overall, AGB ameliorated activation of
TNF-α-mediated caspase pathway by reducing the expression of cleaved caspase-3/-8/-9,
cytochrome-c and the Bcl-2/Bax signaling pathway, eventually suppressing hepatocyte
apoptosis (Fig 7).
5. Conclusions
In summary, AGB pretreatment possessed a great positive effect in ameliorating
antioxidant mechanisms, inflammation responses, and TNF-α-mediated caspase-3/-8/-9
hepatocyte apoptosis on APAP-induced hepatotoxicity. AGB exhibited anti-inflammatory
ability through the inhibition of pro-inflammatory cytokines including NF-κB, TNF-α and
IL-1β in APAP-induced acute hepatotoxicity. Furthermore, the APAP toxicity might be
reduced by preventing the production of lipid peroxidation to alleviate oxidative stress level
as well as hepatic antioxidant status. Likewise, AGB attenuated activation of TNF-α-mediated

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
20
caspase pathway and decreased the Bcl-2/Bax ratio and the expression of cytochrome-c,
which eventually prevented hepatocyte apoptosis and necrosis. The findings from the present
work clearly revealed that AGB may be serves as a hepatoprotective agent in near future.
Acknowledgements
This work was supported by the grants of National Natural Science Foundation of China (No.
81403052 & 314707418), the Scientific Research Foundation for the Returned Overseas
Chinese Scholars (Jilin Province, 2015), Jilin Science & Technology Development Plan (No.
20160209008YY), and the Program for the Young Top-notch and Innovative Talents of Jilin
Agricultural University (2016).
Conflicts of Interest
The authors declare no conflict of interest.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
21
References
Barman, P.K., Mukherjee, R., Prusty, B.K., Suklabaidya, S., Senapati, S., Ravindran, B., 2016. Chitohexaose
protects against acetaminophen-induced hepatotoxicity in mice. Cell Death Dis 7, e2224.
Du, K., McGill, M.R., Xie, Y., Jaeschke, H., 2015. Benzyl alcohol protects against acetaminophen hepatotoxicity
by inhibiting cytochrome P450 enzymes but causes mitochondrial dysfunction and cell death at higher
doses. Food Chem Toxicol 86, 253-261.
Du, K., Ramachandran, A., Jaeschke, H., 2016. Oxidative stress during acetaminophen hepatotoxicity: Sources,
pathophysiological role and therapeutic potential. Redox Biol 10, 148-156.
Fan, J.H., Feng, G.G., Huang, L., Tang, G.D., Jiang, H.X., Xu, J., 2014. Naofen promotes TNF-alpha-mediated
apoptosis of hepatocytes by activating caspase-3 in lipopolysaccharide-treated rats. World J Gastroenterol
20, 4963-4971.
Gum, S.I., Cho, M.K., 2013. Korean red ginseng extract prevents APAP-induced hepatotoxicity through
metabolic enzyme regulation: the role of ginsenoside Rg3, a protopanaxadiol. Liver Int 33, 1071-1084.
Guo, Q., Shen, Z., Yu, H., Lu, G., Yu, Y., Liu, X., Zheng, P., 2016. Carnosic acid protects against
acetaminophen-induced hepatotoxicity by potentiating Nrf2-mediated antioxidant capacity in mice. Korean
J Physiol Pharmacol 20, 15-23.
Guo, Y.L., Jiang, W.D., Wu, P., Liu, Y., Zhou, X.Q., Kuang, S.Y., Tang, L., Tang, W.N., Zhang, Y.A., Feng, L.,
2017. The decreased growth performance and impaired immune function and structural integrity by dietary
iron deficiency or excess are associated with TOR, NF-kappaB, p38MAPK, Nrf2 and MLCK signaling in
head kidney, spleen and skin of grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol 65, 145-168.
Hayakawa, A., Kawamoto, Y., Nakajima, H., Sakai, J., Takasawa, R., Nakashima, I., Magae, J., Tanuma, S., 2008.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
22
Bid truncation mediated by caspases-3 and -9 in vinorelbine-induced apoptosis. Apoptosis 13, 523-530.
Holubek, W.J., Kalman, S., Hoffman, R.S., 2006. Acetaminophen-induced acute liver failure: results of a United
States multicenter, prospective study. Hepatology 43, 880-882.
Hsiao, P.C., Lee, W.J., Yang, S.F., Tan, P., Chen, H.Y., Lee, L.M., Chang, J.L., Lai, G.M., Chow, J.M., Chien,
M.H., 2014. Nobiletin suppresses the proliferation and induces apoptosis involving MAPKs and
caspase-8/-9/-3 signals in human acute myeloid leukemia cells. Tumour Biol 35, 11903-11911.
Jiao, X.L., Bi, X.B., Zhang, X.S., Gao, W.W., 2015. Autotoxic effect of ginsenoside extrats on growth of
American ginseng in different medium. China Journal of Chinese Materia Medica 40, 1433-1438.
Joshi, S., Braithwaite, A.W., Robinson, P.J., Chircop, M., 2011. Dynamin inhibitors induce caspase-mediated
apoptosis following cytokinesis failure in human cancer cells and this is blocked by Bcl-2 overexpression.
Mol Cancer 10, 78.
Kang, E.S., Lee, J., Homma, T., Kurahashi, T., Kobayashi, S., Nabeshima, A., Yamada, S., Seo, H.G., Miyata, S.,
Sato, H., Fujii, J., 2017. xCT deficiency aggravates acetaminophen-induced hepatotoxicity under inhibition
of the transsulfuration pathway. Free Radic Res 51, 80-90.
Legert, K.G., Tsilingaridis, G., Remberger, M., Ringden, O., Heimdahl, A., Yucel-Lindberg, T., Dahllof, G., 2015.
The relationship between oral mucositis and levels of pro-inflammatory cytokines in serum and in gingival
crevicular fluid in allogeneic stem cell recipients. Support Care Cancer 23, 1749-1757.
Li, W., Su, X.-m., Han, Y., Xu, Q., Zhang, J., Wang, Z., Wang, Y.-p., 2015. Maltol, a Maillard reaction product,
exerts anti-tumor efficacy in H22 tumor-bearing mice via improving immune function and inducing
apoptosis. RSC Advances 5, 101850-101859.
Li, W., Yan, M.H., Liu, Y., Liu, Z., Wang, Z., Chen, C., Zhang, J., Sun, Y.S., 2016. Ginsenoside Rg5 Ameliorates
Cisplatin-Induced Nephrotoxicity in Mice through Inhibition of Inflammation, Oxidative Stress, and

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
23
Apoptosis. Nutrients 8, 566.
Li, X.L., Wang, C.Z., Sun, S., Mehendale, S.R., Du, W., He, T.C., Yuan, C.S., 2009. American ginseng berry
enhances chemopreventive effect of 5-FU on human colorectal cancer cells. Oncol Rep 22, 943-952.
Limsrichamrern, S., Chanapul, C., Mahawithitwong, P., Sirivatanauksorn, Y., Kositamongkol, P., Asavakarn, S.,
Tovikkai, C., Dumronggittigule, W., 2016. Correlation of Hematocrit and Tacrolimus Level in Liver
Transplant Recipients. Transplant Proc 48, 1176-1178.
Lin, E., Wang, Y., Mehendale, S., Sun, S., Wang, C.Z., Xie, J.T., Aung, H.H., Yuan, C.S., 2008. Antioxidant
protection by American ginseng in pancreatic beta-cells. Am J Chin Med 36, 981-988.
Liu, W., Zheng, Y., Han, L., Wang, H., Saito, M., Ling, M., Kimura, Y., Feng, Y., 2008. Saponins (Ginsenosides)
from stems and leaves of Panax quinquefolium prevented high-fat diet-induced obesity in mice.
Phytomedicine 15, 1140-1145.
Liu, X., Wang, T., Cai, L., Qi, J., Zhang, P., Li, Y., 2016. Biochanin A protects
lipopolysaccharide/D-galactosamine-induced acute liver injury in mice by activating the Nrf2 pathway and
inhibiting NLRP3 inflammasome activation. Int Immunopharmacol 38, 324-331.
Lu, D., Li, P., Liu, J., 2012. Quinquenoside F(6), a new triterpenoid saponin from the fruits of Panax
quinquefolium L. Nat Prod Res 26, 1395-1401.
Michael Brown, J., Ball, J.G., Wright, M.S., Van Meter, S., Valentovic, M.A., 2012. Novel protective
mechanisms for S-adenosyl-L-methionine against acetaminophen hepatotoxicity: improvement of key
antioxidant enzymatic function. Toxicol Lett 212, 320-328.
Na, S., Li, J., Zhang, H., Li, Y., Yang, Z., Zhong, Y., Dong, G., Yang, J., Yue, J., 2017. The induction of
cytochrome P450 2E1 by ethanol leads to the loss of synaptic proteins via PPARalpha down-regulation.
Toxicology 15,18-27.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
24
Nagase, M., Shiota, T., Tsushima, A., Murshedul Alam, M., Fukuoka, S., Yoshizawa, T., Sakato, N., 2002.
Molecular mechanism of satratoxin-induced apoptosis in HL-60 cells: activation of caspase-8 and caspase-9
is involved in activation of caspase-3. Immunol Lett 84, 23-27.
Nagi, M.N., Almakki, H.A., Sayed-Ahmed, M.M., Al-Bekairi, A.M., 2010. Thymoquinone supplementation
reverses acetaminophen-induced oxidative stress, nitric oxide production and energy decline in mice liver.
Food Chem Toxicol 48, 2361-2365.
Noh, J.R., Kim, Y.H., Hwang, J.H., Choi, D.H., Kim, K.S., Oh, W.K., Lee, C.H., 2015. Sulforaphane protects
against acetaminophen-induced hepatotoxicity. Food Chem Toxicol 80, 193-200.
Peng, X., Gan, J., Wang, Q., Shi, Z., Xia, X., 2016. 3-Monochloro-1,2-propanediol (3-MCPD) induces apoptosis
via mitochondrial oxidative phosphorylation system impairment and the caspase cascade pathway.
Toxicology 372, 1-11.
Saito, C., Zwingmann, C., Jaeschke, H., 2010. Novel mechanisms of protection against acetaminophen
hepatotoxicity in mice by glutathione and N-acetylcysteine. Hepatology 51, 246-254.
Shanmugam, S., Thangaraj, P., Lima, B.D., Chandran, R., de Souza Araujo, A.A., Narain, N., Serafini, M.R.,
Junior, L.J., 2016. Effects of luteolin and quercetin 3-beta-d-glucoside identified from Passiflora subpeltata
leaves against acetaminophen induced hepatotoxicity in rats. Biomed Pharmacother 83, 1278-1285.
Smith, A.K., Petersen, B.K., Ropella, G.E., Kennedy, R.C., Kaplowitz, N., Ookhtens, M., Hunt, C.A., 2016.
Competing Mechanistic Hypotheses of Acetaminophen-Induced Hepatotoxicity Challenged by Virtual
Experiments. PLoS Comput Biol 12, e1005253.
Song, E., Fu, J., Xia, X., Su, C., Song, Y., 2014. Bazhen decoction protects against acetaminophen induced acute
liver injury by inhibiting oxidative stress, inflammation and apoptosis in mice. PLoS One 9, e107405.
Sritularak, B., Morinaga, O., Yuan, C.S., Shoyama, Y., Tanaka, H., 2009. Quantitative analysis of ginsenosides

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
25
Rb1, Rg1, and Re in American ginseng berry and flower samples by ELISA using monoclonal antibodies. J
Nat Med 63, 360-363.
Sullivan, C.B., Porter, R.M., Evans, C.H., Ritter, T., Shaw, G., Barry, F., Murphy, J.M., 2014. TNFalpha and
IL-1beta influence the differentiation and migration of murine MSCs independently of the NF-kappaB
pathway. Stem Cell Res Ther 5, 104.
Truong, V.L., Ko, S.Y., Jun, M., Jeong, W.S., 2016. Quercitrin from Toona sinensis (Juss.) M.Roem. Attenuates
Acetaminophen-Induced Acute Liver Toxicity in HepG2 Cells and Mice through Induction of Antioxidant
Machinery and Inhibition of Inflammation. Nutrients 8, 431.
Wang, C., Shi, D.Z., 2011. [Research of cardiovascular effects and mechanism of Panax quinquefolius saponin].
Chin J Iteg Trad Wes Med 31, 825-831.
Wang, C.Z., Wu, J.A., McEntee, E., Yuan, C.S., 2006a. Saponins composition in American ginseng leaf and
berry assayed by high-performance liquid chromatography. J Agric Food Chem 54, 2261-2266.
Wang, C.Z., Zhang, B., Song, W.X., Wang, A., Ni, M., Luo, X., Aung, H.H., Xie, J.T., Tong, R., He, T.C., Yuan,
C.S., 2006b. Steamed American ginseng berry: ginsenoside analyses and anticancer activities. J Agric Food
Chem 54, 9936-9942.
Wang, H., Zhang, R., Bridle, K.R., Jayachandran, A., Thomas, J.A., Zhang, W., Yuan, J., Xu, Z.P., Crawford,
D.H., Liang, X., Liu, X., Roberts, M.S., 2017. Two-photon dual imaging platform for in vivo monitoring
cellular oxidative stress in liver injury. Sci Rep 7, 45374.
Wang, H., Zhu, Y., Xu, X., Wang, X., Hou, Q., Xu, Q., Sun, Z., Mi, Y., Hu, C., 2016. Ctenopharyngodon idella
NF-kappaB subunit p65 modulates the transcription of IkappaBalpha in CIK cells. Fish Shellfish Immunol
54, 564-572.
Wang, S., Wang, X., Luo, F., Tang, X., Li, K., Hu, X., Bai, J., 2014. Panaxatriol saponin ameliorated liver injury

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
26
by acetaminophen via restoring thioredoxin-1 and pro-caspase-12. Liver Int 34, 1068-1073.
Xie, J., Wan, J., Jiang, R., Lu, H., Peng, X., Zhang, L., 2013. Upregulation of Sirt1 in
carbon-tetrachloride-induced acute liver injury. Drug Chem Toxicol 36, 277-283.
Xie, J.T., Aung, H.H., Wu, J.A., Attel, A.S., Yuan, C.S., 2002. Effects of American ginseng berry extract on
blood glucose levels in ob/ob mice. Am J Chin Med 30, 187-194.
Xie, J.T., Wu, J.A., Mehendale, S., Aung, H.H., Yuan, C.S., 2004. Anti-hyperglycemic effect of the
polysaccharides fraction from American ginseng berry extract in ob/ob mice. Phytomedicine 11, 182-187.
Xu, X.Y., Hu, J.N., Liu, Z., Zhang, R., He, Y.F., Hou, W., Wang, Z.Q., Yang, G., Li, W., 2017. Saponins
(Ginsenosides) from the Leaves of Panax quinquefolius Ameliorated Acetaminophen-Induced
Hepatotoxicity in Mice. J Agric Food Chem 65, 3684-3692.
Yan, D., Liu, H.L., Yu, Z.J., Huang, Y.H., Gao, D., Hao, H., Liao, S.S., Xu, F.Y., Zhou, X.Y., 2016. BML-111
Protected LPS/D-GalN-Induced Acute Liver Injury in Rats. Int J Mol Sci 17, 453-74.
Yu, C., Wang, C.Z., Zhou, C.J., Wang, B., Han, L., Zhang, C.F., Wu, X.H., Yuan, C.S., 2014. Adulteration and
cultivation region identification of American ginseng using HPLC coupled with multivariate analysis. J
Pharm Biomed Anal 99, 8-15.
Yuan, H.D., Jin, G.Z., Piao, G.C., 2010. Hepatoprotective effects of an active part from Artemisia sacrorum
Ledeb. against acetaminophen-induced toxicity in mice. J Ethnopharmacol 127, 528-533.
Zhang, J., Zhang, S., Bi, J., Gu, J., Deng, Y., Liu, C., 2017. Astaxanthin pretreatment attenuates
acetaminophen-induced liver injury in mice. Int Immunopharmacol 45, 26-33.
Zhao, G., Zhu, Y., Eno, C.O., Liu, Y., Deleeuw, L., Burlison, J.A., Chaires, J.B., Trent, J.O., Li, C., 2014.
Activation of the proapoptotic Bcl-2 protein Bax by a small molecule induces tumor cell apoptosis. Mol
Cell Biol 34, 1198-1207.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
27
Zhao, X., Cong, X., Zheng, L., Xu, L., Yin, L., Peng, J., 2012. Dioscin, a natural steroid saponin, shows
remarkable protective effect against acetaminophen-induced liver damage in vitro and in vivo. Toxicol Lett
214, 69-80.
Zhu, L., Yuan, H., Guo, C., Lu, Y., Deng, S., Yang, Y., Wei, Q., Wen, L., He, Z., 2012. Zearalenone induces
apoptosis and necrosis in porcine granulosa cells via a caspase-3- and caspase-9-dependent mitochondrial
signaling pathway. J Cell Physiol 227, 1814-1820.

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
28
Figure legends
Figure 1. HPLC chromatogram and chemical structures of seven saponins in the AGB. These
saponins includes four protopanaxadiol ginsenosides (e.g. Rb1, Rb2, Rb3, Rc, and Rd) and
two protopanaxatriol ginsenosides (e.g. Re and Rg1).
Figure 2. Pretreatment with AGB protected against APAP-induced liver injury: The levels of
ALT (A), AST (B), GSH (C), MDA (D) and SOD (E) were measured in APAP-induced
hepatotoxicity. All data were expressed as mean ± S.D., n = 8. * p < 0.05, ** p < 0.01 vs.
normal group; #
p < 0.05, ##
p < 0.01 vs. APAP group. GSH = glutathione; MDA
=malondialdehyde; SOD = superoxide dismutase.
Figure 3. Pretreatment with AGB protected against APAP-induced liver injury. 4-HNE (A);
CYP2E1 (B), the 4-HNE of relative fluorescence intensity (C), the CYP2E1 of relative
fluorescence intensity (D). The expression level of CYP2E1 and 4-HNE (Green) in tissue
section isolated from different groups was assessed by immunofluorescence. Representative
immunofluorescence images were taken at 400×4’ 6-Diamidino-2-phenylindole (DAPI) (Blue)
was used as a nuclear counterstain.
Figure 4. Pretreatment with AGB protected against APAP-induced liver injury. The levels of:
TNF-α (A) and IL-1β (B) in liver tissues were determined by ELISA kits. The protein
expression was examined by western blotting analysis in liver tissues. p-p65 and TNF-α (C),
the relative protein expression of p-p65 (D), the relative protein expression of TNF-α (E). All
data were expressed as mean ± S.D., n = 8. * p < 0.05, ** p < 0.01 vs. normal group; # p <
0.05, ##
p < 0.01 vs. APAP group.
Figure 5. Histological examination of morphological changes in liver tissues. Livers of four

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
29
groups were observed (A), Liver tissues stained with H&E (100×, 400×) (B), Hoechst 33258
(100×, 400×) (C), and TUNEL (400×) (D), the Hoechst 33258 percentage apoptosis (%) (E),
the TUNEL percentage apoptosis (%) (F). Arrows show hepatocyte, necrosis, congestion,
inflammatory cell infiltration. All data were expressed as mean ± S.D., n = 8. * p < 0.05, ** p
< 0.01 vs. normal group; # p < 0.05,
## p < 0.01 vs. APAP group.
Figure 6. Effects of AGB on the protein expression of Cytochrome C, Bax, Bcl-2 (A),
cspase-3, caspase-8, caspase-9 (B), relative protein expression of Cytochrome C, Bax, Bcl-2,
Bcl-2/Bax (C); caspase-3 caspase-8 caspase-9 (D) band intensities were analyzed. The protein
expression was examined by western blotting analysis in liver tissues from normal, APAP,
APAP + AGB (150 mg/kg), and APAP + AGB (300 mg/kg) groups. All data were expressed
as mean ± S.D., n = 8. * p < 0.05, ** p < 0.01 vs. normal group; # p < 0.05,
## p < 0.01 vs.
APAP group.
Figure 7. Schematic diagram of signaling pathway involved in the hepatoprotective effects of
AGB on APAP-induced hepatotoxicity. AGB exerts the liver protection effects against
APAP-induced hepatotoxicity mainly through TNF-α-mediated caspase-3/-8/-9 signaling
pathways.
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
30
Figure 1
A
B
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
31
C
A B
0
2
4
6
MD
A(n
mo
l/m
g.p
ro
tein
)
*
##
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
0
2 0
4 0
6 0
8 0
SO
D(U
/mg
.pr
ote
in)
*
#
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
# #
0
5
1 0
1 5
GS
H(μ
mo
l/g
.pr
ote
in)
*
#
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
# #
0
2 0
4 0
6 0
AS
T(U
/L)
*
#
# #
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
0
5 0
1 0 0
1 5 0
2 0 0A
LT
(U/L
) *
#
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
# #
D
E
Figure 2
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
32
0 .0
0 .5
1 .0
1 .5
Re
lativ
e f
luo
re
se
nc
e i
nte
ns
ity
C Y P 2 E 1
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
* *
#
#
0 .0
0 .5
1 .0
1 .5
Re
lativ
e f
luo
re
se
nc
e i
nte
ns
ity 4 - H N E
**
#
#
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
Figure 3
A
B
C D
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
33
A B
0
1 0 0
2 0 0
3 0 0
TN
F-
(ng
/L)
*
##
A P A P (2 5 0 m g /k g )
A G B (m g /k g )
- +
- -
+ +
1 5 0 3 0 0
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
IL
-1
(pg
/L)
*
##
A P A P (2 5 0 m g /k g )
A G B (m g /k g )
- +
- -
+ +
1 5 0 3 0 0
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
TN
F-
/GA
PD
H
**
#
T N F - /G A P D H
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
#
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
NF
- B
/GA
PD
H
* *
## #
N F - B /G A P D H
A P A P ( m g /k g )
A G B ( m g /k g )
- +
- -
+ +
1 5 0 3 0 0
Figure 4
C
D E
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
34
0
2 0
4 0
6 0
8 0
1 0 0
Th
e p
er
se
nta
ge
ap
op
to
sis
(%
)
H o e c h s t 3 3 2 5 8
*
#
#
A P A P (2 5 0 m g /k g )
A G B (m g /k g )
- +
- -
+ +
1 5 0 3 0 0
0
2 0
4 0
6 0
8 0
Th
e p
er
se
nta
ge
ap
op
to
sis
(%
)
TU N EL
*
#
#
A P A P (2 5 0 m g /k g )
A G B (m g /k g )
- +
- -
+ +
1 5 0 3 0 0
Figure 5
E F
C
B
A
D
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
35
1 2 3 4
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Re
lati
ve
pr
ote
in e
xp
re
ss
ion
(% o
f n
or
ma
l)
C y t o c h r o m e C B a x B c l-2 B a x /B c l-2
**
A P A P (m g /k g )
A G B (m g /k g ) -
+-
-
+ +
1 5 0 3 0 0 -
+-
-
+ +
1 5 0 3 0 0
- +
--
+ +
1 5 0 3 0 0
- +
- -
+ +
1 5 0 3 0 0
## #
**
# #
# #
**#
# #
**
# #
# #
1 2 3
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Re
lati
ve
pr
ote
in e
xp
re
ss
ion
(% o
f n
or
ma
l)
c a s p a s e - 3 c a s p a s e - 8 c a s p a s e - 9
* *
#
# #
* *
#
# #
* *
## #
A P A P (m g /k g )
A G B (m g /k g ) -
+-
-
+ +
1 5 0 3 0 0 -
+-
-
+ +
1 5 0 3 0 0
- +
- -
+ +
1 5 0 3 0 0
Figure 6
A
B
D
C
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
36
Figure 7
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIP
T
37
Graphical Abstract


















