Flow Cytometric Analysis of Hydroxyurea Effects on Fetal Hemoglobin Production in Cultures of...
11
ORIGINAL ARTICLE Flow Cytometric Analysis of Hydroxyurea Effects on Fetal Hemoglobin Production in Cultures of b-Thalassemia Erythroid Precursors Ilana Amoyal, Ada Goldfarb, and Eitan Fibach * Department of Hematology, Hadassah University Hospital, Jerusalem, Israel ABSTRACT An increase in fetal hemoglobin (Hb F) ameliorates the clinical symptoms of the underlying disease in the b hemoglobinopathies—sickle cell anemia and b-thalassemia (thal). Hydroxyurea (HU) can elevate Hb F production in erythroid cells and is the agent currently in clinical use for patients with sickle cell anemia; it is presently being tested in clinical trials for thalassemia. We have developed a two-phase liquid culture system that mimics the in vivo hematological changes that are observed in patients treated with HU. Adding HU during the second phase of the culture increases the proportion of Hb F, increases the levels of total hemoglobin (Hb) content per cell and increases cell size, but it decreases the numbers of cells and the total amount of Hb produced. In the present study we developed and utilized a double labeling procedure for flow cytometric analysis of the cellular distribution of Hb F. Cells exposed to various concentrations of HU on day 6 of the second phase of the culture were harvested on day 12, and stained simultaneously with fluorochrome-conjugated monoclonal antibodies specific for human glycophorin A, an erythroid specific marker, and human Hb F. Both the percentage of the Hb F-containing cells and their intensity of fluorescence were recorded. The latter value gives a semi-quantitative estimation of the mean cellular Hb F content. The results indicated that cultures derived from different b-thalassemic patients differ in their response to HU. In most patients, low doses of HU decreased the percentage of Hb F-cells as well as their Hb F content. At high doses, * Correspondence: Dr. Eitan Fibach, Department of Hematology, Hadassah University Hospital, Ein- Kerem, P.O. Box 12,000, Jerusalem 91120, Israel; Fax: 972-2-6423067; E-mail: fi[email protected]. HEMOGLOBIN Vol. 27, No. 2, pp. 77–87, 2003 DOI: 10.1081=HEM-120021539 0363-0269 (Print); 1532-432X (Online) Copyright # 2003 by Marcel Dekker, Inc. www.dekker.com 77 Hemoglobin Downloaded from informahealthcare.com by CDL-UC Berkeley on 10/28/14 For personal use only.
Transcript of Flow Cytometric Analysis of Hydroxyurea Effects on Fetal Hemoglobin Production in Cultures of...

ORIGINAL ARTICLE
Flow Cytometric Analysis of Hydroxyurea Effectson Fetal Hemoglobin Production in Cultures of
b-Thalassemia Erythroid Precursors
Ilana Amoyal, Ada Goldfarb, and Eitan Fibach*
Department of Hematology, Hadassah University Hospital, Jerusalem, Israel
ABSTRACT
An increase in fetal hemoglobin (Hb F) ameliorates the clinical symptoms of the
underlying disease in the b hemoglobinopathies—sickle cell anemia and b-thalassemia
(thal). Hydroxyurea (HU) can elevate Hb F production in erythroid cells and is the
agent currently in clinical use for patients with sickle cell anemia; it is presently being
tested in clinical trials for thalassemia. We have developed a two-phase liquid culture
system that mimics the in vivo hematological changes that are observed in patients
treated with HU. Adding HU during the second phase of the culture increases the
proportion of Hb F, increases the levels of total hemoglobin (Hb) content per cell and
increases cell size, but it decreases the numbers of cells and the total amount of
Hb produced. In the present study we developed and utilized a double labeling
procedure for flow cytometric analysis of the cellular distribution of Hb F. Cells
exposed to various concentrations of HU on day 6 of the second phase of the culture
were harvested on day 12, and stained simultaneously with fluorochrome-conjugated
monoclonal antibodies specific for human glycophorin A, an erythroid specific marker,
and human Hb F. Both the percentage of the Hb F-containing cells and their intensity of
fluorescence were recorded. The latter value gives a semi-quantitative estimation of the
mean cellular Hb F content. The results indicated that cultures derived from different
b-thalassemic patients differ in their response to HU. In most patients, low doses of HU
decreased the percentage of Hb F-cells as well as their Hb F content. At high doses,
*Correspondence: Dr. Eitan Fibach, Department of Hematology, Hadassah University Hospital, Ein-
Kerem, P.O. Box 12,000, Jerusalem 91120, Israel; Fax: 972-2-6423067; E-mail: [email protected].
HEMOGLOBIN
Vol. 27, No. 2, pp. 77–87, 2003
DOI: 10.1081=HEM-120021539 0363-0269 (Print); 1532-432X (Online)
Copyright # 2003 by Marcel Dekker, Inc. www.dekker.com
77
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

some patients showed an increase in both parameters, while others showed an increase
in the percentage of Hb F-cells with minimal increase in their mean Hb F content, while
still other patients showed little effect at all. In all patients, high doses of HU caused
a decrease in cell numbers. These results suggest that HU has mixed effects on
erythroid precursors. Both the two-phase liquid culture and the flow cytometric
analysis procedures described herein provide the experimental tools for screening of
Hb F-inducing drugs and for evaluating patients’ cell response prior to treatment.
Key Words: Flow cytrometric analysis; Hydroxyurea (HU); Fetal hemoglobin (Hb F);
Thalassemia (thal); Erythroid precursors.
INTRODUCTION
Fetal hemoglobin (Hb F) is frequently increased in the b hemoglobinopathies—sickle
cell anemia and b-thalassemia (thal) (1). Epidemiological studies have indicated that an
increase in Hb F ameliorates the clinical symptoms of the underlying disease (2,3). In
sickle cell anemia not only do Hb F-containing cells have a lower concentration of sickle
hemoglobin (Hb), but Hb F inhibits polymerization of sickle Hb directly, accounting for
the lower propensity of such cells to undergo sickling (4–6). In b-thal, elevated Hb F
should compensate partially for the deficiency in b-globin chains.
Several pharmacological agents have been used to stimulate Hb F synthesis in vitro
and in vivo. The initial studies involved treatment of baboons (7) or patients with either
b-thal (8) or sickle cell anemia (9), with 5-azacytidine. The treatment resulted in a
considerable stimulation of g chain synthesis. Since this drug has been implicated as a
carcinogen (10), hydroxyurea (HU) is the agent currently in clinical use for patients with
sickle cell anemia and is being tested in clinical trials for thalassemia (11–14). The
response to HU treatment is variable; about 30% of the patients do not respond at all, and
the patients who do respond do it to various degrees.
Various cell culture systems have been used to better characterize the molecular
mechanisms by which HU mediates these effects (15). We developed a two-phase liquid
culture procedure that mimics the hematological changes that are observed in patients
treated with HU (16). In phase I, peripheral blood mononuclear cells are cultured in the
presence of a combination of growth factors, but in the absence of erythropoietin (Epo).
During this stage, early erythroid committed progenitors (BFU-E) proliferate and differ-
entiate into erythroid colony-forming unit (CFU-E)-like progenitors. In phase II, the latter
cells, in response to Epo, continue to proliferate and mature into orthochromatic
normoblasts (17,18). This system yields large and relatively pure populations of erythroid
cells that are semi-synchronous in maturity (19). Cells exposed to high concentrations
of HU during the second part of phase II were found to have an increased proportion of
Hb F (%Hb F), increased levels of total Hb content per cell (MCH), and increased cell
size (MCV), but the numbers of cells and the total amount of Hb produced were
decreased (16,20).
The analysis of the Hb content of these cultures was carried out by high performance
liquid chromatography (HPLC) (16) that measures the percentage of Hb F produced, and
by reverse transcriptase polymerase chain reaction (RT–PCR) that measures the content of
mRNA for the various globin genes (21). However, the results obtained by these
78 Amoyal, Goldfarb, and Fibach
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

techniques provide an average data of the entire cell population studied, and do not allow
analysis at the single cell basis. Such an analysis is possible by flow cytometry.
In the present study we developed and utilized a double labeling procedure for a flow
cytometric analysis. Cells were stained simultaneously with monoclonal antibodies
specific for human glycophorin A (GPA) and human Hb F. Glycophorin A is an erythroid
specific marker that appears on the surface of erythroid precursors in parallel with the
intracellular accumulation Hb. The GPA positive cells were gated and analyzed for their
distribution with respect to Hb F. Both the percentage of Hb F-containing cells (%Hb
F-cells) and their intensity of fluorescence, expressed as the mean fluorescence channel
(MFC), were recorded. The MFC provides a comparative, semi-quantitative estimation of
the cellular Hb F content. The results indicated that cultures derived from different
b-thalassemic patients differ in their response to HU. In most patients, low doses of HU
resulted in a decrease in both the %Hb F-cells as well as in their Hb F content. At high
doses, some patients showed an increase in both parameters; other patients showed an
increase in the %Hb F-cells with no increase in Hb F content; while still other patients
showed no effect at all. High doses of HU caused a decrease in cell numbers in all patients.
MATERIALS AND METHODS
Patients
Twelve b-thal patients were studied. Table 1A summarizes some clinical data of
these patients.
Culture of b-Thalassemic Erythroid Cells
After informed consent was obtained, 10 mL blood was withdrawn into a heparin-
containing tube. In the case of transfused patients, blood was obtained just prior to
transfusion. Mononuclear cells were isolated by centrifugation over Ficoll-Hypaque
(1.077 g=mL; Sigma, St. Louis, MO, USA). The cells were cultured as previously
described (17,18,22). Briefly, the cells were first cultured in a-minimal essential medium
containing 10% pre-selected fetal bovine serum (FBS) (both from Biological Industries,
Beit-HaEmek, Israel), 1 mg=mL of cyclosporine A (Sandoz, Basel, Switzerland), and 10%
conditioned medium collected from cultures of a human bladder carcinoma 5637 cell line
[American Type Culture Collection (ATCC), Manassas, VA, USA]. After 5–7 day
incubation in this phase I culture, non adherent cells were harvested, washed and
resuspended in phase II medium composed of a medium, 30% FBS, 1% deionized bovine
serum albumin (BSA; Sigma) supplemented with 10�6 mol=L dexamethasone (Elkin-Sinn,
Cherry Hill, NJ, USA), 10�5 mol=L b-mercaptoethanol (Sigma), 0.3 mg=mL of human
holo-transferrin (Sigma), and 1 U=mL of human recombinant Epo (Ortho Pharmaceuticals,
Raritan, NJ, USA). After exposure to Epo, erythroid progenitors proliferated and matured
after 12 days into Hb-containing orthochromatic normoblasts. Various concentrations of
HU (Sigma) were added to the cultures on day 6 of phase II. On day 12, all cultures were
terminated, and the cells were harvested and analyzed. The Hb-containing cells were
Flow Cytometry Analysis of HU Effect on Hb F 79
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

Ta
ble
1.
Th
ep
atie
nt
po
pu
lati
on
stu
die
d:
som
ecl
inic
ald
ata
(A)
and
resp
on
seo
fcu
ltu
red
cell
sto
hy
dro
xy
ure
a(B
).
(A)
(B)
Hb
F(m
ax.
fold
incr
ease
)a
No
.S
ex-a
ge
b-T
hal
asse
mia
Mu
tati
on
Tra
nsf
usi
on
req
uir
emen
t
Des
ferr
al
trea
tmen
t
%H
bF
cell
sbM
FC
c
1M
-37
Inte
rmed
iaIV
S-I
-6(T
!C
)=IV
S-I
-6(T
!C
)O
ccas
ion
ally
No
ne
1.7
61
.53
2F
-41
Inte
rmed
iaIV
S-I
-6(T
!C
)=IV
S-I
-1(G
!T
)�
1=m
on
thY
es4
.38
3.6
4
3F
-38
Inte
rmed
iaIV
S-I
-6(T
!C
)=IV
S-I
-6(T
!C
)O
ccas
ion
ally
No
ne
1.7
71
.90
4M
-43
Inte
rmed
iaIV
S-I
-6(T
!C
)=IV
S-I
-1(G
!T
)�
1=m
on
thY
es1
.44
2.7
2
5M
-51
Inte
rmed
ia�
28
(A!
C)=�
28
(A!
C)
Occ
asio
nal
lyN
on
e1
.16
1.6
1
6M
-49
Inte
rmed
ia�
28
(A!
C)=�
28
(A!
C)
Occ
asio
nal
lyN
on
e1
.26
1.5
6
7F
-27
Inte
rmed
iaIV
S-I
-6(T
!C
)=IV
S-I
-6(T
!C
)N
on
eN
on
e1
.17
1.1
3
8M
-45
Inte
rmed
ia�
28
(A!
C)=�
28
(A!
C)
No
ne
No
ne
1.3
12
.03
9M
-37
Maj
or
IVS
-I-1
10
(G!
A)=
IVS
-I-1
10
(G!
A)
�1=m
on
thY
es4
.50
2.9
4
10
M-4
4M
ajo
rF
SC
44
(–C
)=P
oly
A(A
AT
AA
A!
AA
TA
AG
)�
1=m
on
thY
es1
.60
2.8
1
11
F-3
9M
ajo
rF
SC
44
(–C
)=P
oly
A(A
AT
AA
A!
AA
TA
AG
)�
1=m
on
thY
es2
.72
1.3
8
12
M-2
3M
ajo
rF
SC
44
(–C
)=F
SC
44
(–C
)�
1=m
on
thY
es1
.16
1.1
2
aC
ells
wer
ecu
ltu
red
inth
ep
rese
nce
of
dif
fere
nt
con
cen
trat
ion
so
fH
Uan
dan
aly
zed
by
flow
cyto
met
ryfo
llow
ing
do
uble
stai
nin
gw
ith
anti
hu
man
Hb
Fan
d
anti
hu
man
GPA
anti
bo
die
s.bT
he
pro
po
rtio
no
fH
bF
-co
nta
inin
gce
lls
ou
to
fth
eto
tal
po
pu
lati
on
of
ery
thro
idce
lls.
cT
he
MF
Co
fth
eH
bF
-co
nta
inin
ger
yth
roid
cell
s.F
or
each
pat
ien
t,th
era
tio
of
%H
bF
cell
san
dM
FC
of
cell
str
eate
dw
ith
op
tim
alco
nce
ntr
atio
no
fH
Up
er
un
trea
ted
cell
sw
asca
lcu
late
d.
Th
eo
pti
mal
con
cen
trat
ion
of
HU
var
ied
amo
ng
the
pat
ien
ts.
80 Amoyal, Goldfarb, and Fibach
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

counted following staining with a mixture of benzidine dihydrochloride and hydrogen
peroxide, as described previously (18).
Flow Cytometry
For immuno-phenotyping, cells were harvested and washed with phosphate buffered
saline containing 1% BSA. The cells were fixed in 1 mL of 4% paraformaldehyde (Sigma)
for 15 min at room temperature. The cells were then centrifuged and the pellet permea-
bilized by resuspending the cells in 1 mL mixture of ethanol : acetone (1 : 4, v=v) for 1 min
at room temperature. The cells were washed twice, the pellet was resuspended in 0.1 mL
buffer and the cells stained by incubation (at room temperature for 60 min) with
fluorescein isothiocyanate (FITC)-conjugated anti-human GPA and=or phycoerythrin
(PE)-conjugated anti-human Hb F (both from Immunoquality Products, Groningen,
The Netherlands). The cells were then washed with the same buffer and analyzed, using
a FACS-Calibur1 flow cytometer (Becton-Dickinson, Immunofluorometry Systems,
Mountain View, CA, USA). Cells were passed at a rate of about 1000 cells=second,
using saline as the sheath fluid. A 488 nm argon laser beam served as the light source for
excitation. Emission of 10,000 cells was measured using logarithmic amplification for
fluorescence and linear amplification for forward light scattering and side scatter. FL-1
measured FITC-derived fluorescence and FL-2 measured PE-derived fluorescence. In both
cases geometrical amplification was used. Background noise was determined using isotype
control stained cells. The percentage of Hb F-containing cells (%Hb F-cells) out of the
total population or out of the GPA-positive (erythroid) population was calculated using the
CELLQuest1 (Becton-Dickinson) software. The geometrical MFC of the entire cell
population or of the Hb F-cells, as indicated in the text, were also calculated.
RESULTS
Peripheral blood mononuclear cells were cultured according to the two-phase liquid
culture protocol. On day 6 of phase II, cells were exposed to different concentrations of
HU. On day 12, the cultures were harvested; an aliquot was counted for Hb-containing
cells by benzidine staining, and another aliquot was double labeled for GPA and Hb F and
analyzed by flow cytometry. Figure 1 shows the data as dot plots of a representative
experiment with cultures derived from a thalassemic patient. Figure 1(A), (B) show the
controls: cells stained with PE-conjugated anti-Hb F antibodies had minimal (0.44%) FL-1
fluorescence [Fig. 1(A)], and cells stained with FITC-conjugated anti-GPA antibodies
had minimal (0.73%) FL-2 fluorescence [Fig. 1(B)]. These results indicate that under the
flow conditions used, there was no spillover of fluorescence of cells stained for FL-2 (Hb F)
into the FL-1 detector, and likewise, no FL-2 fluorescence by cells stained for FL-1 (GPA).
Figure 1(C) shows the FL-1 and FL-2 fluorescence of cells double stained for both
GPA and Hb F. The results of this particular experiment show that 8.0% of the entire
population had no fluorescence at all; these (GPA�Hb F�) cells were probably non
erythroid cells (lymphocytes, myeloid precursors, and some macrophages) or very early
erythroid precursors prior to accumulation of either Hb or GPA; 77.2% of the cells were
GPAþHb F�, erythroid cells not containing appreciable amount of Hb F; 12.6% of the cells
Flow Cytometry Analysis of HU Effect on Hb F 81
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

were GPAþHb Fþ, Hb F-containing erythroid cells; and 2.2% GPA�Hb Fþ cells, probably
very immature erythroid precursors that do not yet contain GPA but already contain Hb F.
Fetal hemoglobin has been shown to start accumulating at early stages of the erythroid
maturation (23), probably prior to GPA expression.
A gate was set on the GPAþ cells, and cells within this gate were analyzed for Hb F.
The results show 14.5% Hb Fþ cells within this population. The MFC was calculated to be
36. Figure 1(D) shows the FL-1=FL-2 dot plot of cells from the same patient treated with
150 mM HU. The results show that within the total population 24% were GPAþHb Fþ.
Within the GPAþ cells 29.4% were Hb Fþ with an MFC of 50.3. Thus, in this particular
experiment, HU increased the percentage of erythroid cells with Hb F from 14.5 to 29.4%
(a 2.0-fold increase) and the MFC from 36 to 50.3 (a 1.4-fold increase).
We next studied cultures derived from 12 b-thalassemic patients treated with different
concentrations of HU. The maximal fold increase by HU of the %Hb F cells and their MFCs
are summarized in Table 1 (column B). The results show a variable degree of response. Some
patients (patients 2 and 9) showed increase in both the %Hb F cells and their MFCs. Other
Figure 1. Flow cytometric analyses of Hb F content in thalassemic erythroid precursors. Cultures
derived from a b-thal patient, untreated (A–C) or treated with 150mM HU (D), were stained with
PE-conjugated antibodies for Hb F (A), with FITC-conjugated antibodies for GPA (B), or both
(C–D). The percentages of cells out of the entire population in each quadrant are shown.
82 Amoyal, Goldfarb, and Fibach
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

patients (e.g., patient 11) showed and increase in the %Hb F cells with minimal increase in
MFC, while still other patients (e.g., patients 7 and 12) showed little effect at all. In all
patients, low doses of HU resulted in a decrease in both the %Hb F-cells as well as their MFC,
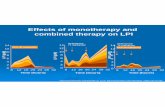
whereas in all patients high doses of HU caused a decrease in cell numbers. Figure 2 depicts
the results of the %Hb F cells and MFC obtained in cultures derived from three
representative patients. Patient 9 [see Table 1 (column B)] showed an HU dose-response
increase of either parameter, patient 11 showed an increase in %Hb F cells but no change
in MFC, and patient 5 showed no change in either parameter.
Figure 2. Effect of different doses of HU on Hb F in cultured thalassemic cells. Cells derived from
three representative patients were cultured from day 6 of phase II in the presence of the indicated
doses of HU. On day 12, the cells were harvested and stained for Hb F and GPA. The variations in
%Hb F cells (A) and MFC (B) are shown.
Flow Cytometry Analysis of HU Effect on Hb F 83
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

DISCUSSION
The two-phase liquid culture procedure is a valuable tool for screening of new Hb
F-stimulating drugs and for studying their mechanism of action. Two broad hypotheses
have been explored for the drug effect. One is based on drug-induced modifications of
DNA, such as hypo-methylation (9) or activation of responsive regions of the globin
promoter (24). The other mechanism involves modification of the rate of differentiation or
maturation of erythroid progenitors, such as rapid recruitment of erythroblasts directly
from progenitors with higher Hb F programs due to the cytoreduction effect of drugs such
as HU (25), or direct induction of down-stream maturation of progenitor cells (26,27).
Observations of elevated Hb F following accelerated erythropoiesis strongly suggest that
the rate of erythroid maturation is involved in the modulation of Hb F production (28).
This was supported by our experiments with hemin (29) and Epo (30).
In order to study the mechanism of action of HU, we analyzed the cultured cell
distribution with respect to Hb F content. In normal adults, most Hb F is restricted to a few
‘‘F cells.’’ Both the number and the Hb F content of these F cells may be increased in
various acquired and genetic conditions (31,32). In the present study, two parameters were
studied: the percentage of Hb F-containing cells and their Hb F content. For this purpose,
we developed a double staining flow cytometric procedure. In order to exclude from
analysis some non erythroid cells, such as lymphocytes, myeloid precursors, and macro-
phages (33) or mature red blood cells (RBC) derived from the original blood that may
contaminate the culture, the cells were labeled for GPA. Glycophorin A is a specific
erythroid marker that is present on intermediate erythroid precursors, but not on non
erythroid cells or mature RBC. By gating on GPAþ cells, only erythroid precursors were
included in the analysis. The cellular Hb F content was inferred from the MFC. Although a
direct relationship between MFC and Hb F content is not available, the MFC provides a
comparative, semi-quantitative estimation of Hb F content. Thus, MFC of cord blood RBC
is 6-fold higher than thalassemic Hb F-containing RBC (F-cells) and 10-fold higher than
normal F-cells. Similar results were obtained when cultured erythroid precursors from cord
blood, thalassemic blood, and normal blood were compared (data not shown).
Increase in %Hb F-cells without a change in Hb F content would suggest that HU
works through its cytoreduction effect; e.g., selective survival of Hb F-containing cells or a
selective maturation of Hb F-committed progenitors. In contrast, increased cellular Hb F
content would suggest that HU induces the potential of cells to produce Hb F, probably
through direct action at the DNA level. Molecular studies reported that HU affects the
proportion of Hb F, g-globin chain, and g-globin mRNA produced (21). But these effects
fail to suggest a mode of action; both selection or induction mechanism would cause an
elevation of these parameters.
The results of flow cytometry analyses indicated that cultures derived from different
b-thalassemic patients differ in their response to HU. In most patients, low doses of HU
resulted in a decrease in both the %Hb F-cells as well as their Hb F content. These results
are in agreement with a recent report showing that HU exerts a bimodal, dose-dependent,
effect on erythropoiesis in culture; low doses increase the cellularity and total Hb content,
but not the Hb F level. High doses, decreased the cellularity and total Hb content, but
increased the proportion of Hb F (20). The present study substantiates these results with
thalassemic patients, and further demonstrates that while some patients respond by an
increase in both the %Hb F-cells and Hb F content, others show an increase in %Hb F-cells
84 Amoyal, Goldfarb, and Fibach
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

with minimal increase in Hb F content, while still other patients show little effect at all.
These results, albeit based on a relatively small group of patients, indicated no correlation
between the pattern of response to HU and the genetic (the mutations involved) and
phenotype (clinical manifestations and mode of treatment, e.g., blood transfusion, chelation
therapy by desferral or deferiprone, etc.) of the patients. These results suggest that HU has
mixed effects on erythroid precursors. The relationship of changes in these two parameters
of Hb F to the clinical outcome of the treatment is a subject of further study.
Most of the patients have been treated by regular or occasional blood transfusions.
However, in the culture procedure used, the transfused blood did not affect the Hb
produced, since the transfused normal donor progenitors do not develop in culture (data
not shown). This is due to: (a) thalassemic blood contains elevated number of erythroid
progenitors compared to normal blood; (b) progenitors are removed from the donor blood
in the process of separation of the RBC fraction (by removal of the ‘‘buffy-coat’’)
following blood collection in the blood bank; (c) progenitors do not survive the storing
conditions of the blood bank.
The unresponsiveness of a significant proportion of patients to HU suggests that in vitro
predictive assay prior to treatment is highly desirable. Both the two-phase liquid culture and
the flow cytometric analysis described herein provide the experimental tools for such an
assay. In vivo studies have shown that continuous infusion of a therapeutic dose of HU at
20 mg=kg will maintain a serum concentration at 100 mM (34). The HU doses used in the
two-phase culture closely resembled the in vivo pharmacokinetics of HU treatment. In
addition, these cultures have been demonstrated to recapitulate many aspects of the in vivo
hematological changes following treatment with HU [reviewed in Ref. (15)]. By using the
flow cytometric method less than one million cells are required for analysis; a cell number
that can be obtained by culturing mononuclear cells derived from 1 mL of peripheral blood.
Flow cytometers are standard equipment in most hematological laboratories (for diagnostic
immunophenotyping of immunological and hematological diseases), thus, the procedure
provides an affordable, objective, accurate, and quantitative assay for determining Hb F
production in culture. However, further studies are required to conclude that results
obtained in culture could be extrapolated to patient’s response to HU and other drugs.
ACKNOWLEDGMENTS
We wish to thank Ms. A. Sinberger, Ms. A. Treves, and Dr. E. Prus (Department of
Hematology, Hadassah University Hospital, Jerusalem, Israel) for their help throughout the
study.
REFERENCES
1. Bunn HF, Forget BG. Hemoglobins A2, F, and A1c and other human hemoglobin
components. In: Hemoglobin: Molecular, Genetic and Clinical Aspects. Philadelphia:
W.B. Saunders Company, 1986:73–74.
2. Al-Awamy BH, Niazi GA, el-Mouzan MI, Altorki MT, Naeem MA. Relationship of
Haemoglobin F and a thalassaemia to severity of sickle-cell anaemia in the Eastern
Province of Saudi Arabia. Ann Trop Paediatr 1986; 6:261–265.
Flow Cytometry Analysis of HU Effect on Hb F 85
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

3. Haghshenass M, Ismail-Beigi F, Clegg JB, Weatherall DJ. Mild sickle-cell anaemia in
Iran associated with high levels of fetal haemoglobin. J Med Genet 1977; 14:168–171.
4. Benesch RE, Edalji R, Benesch R, Kwong S. Solubilization of Hemoglobin S by other
hemoglobins. Proc Natl Acad Sci USA 1980; 77:5130–5134.
5. Noguchi CT, Rodgers GP, Serjeant G, Schechter AN. Levels of fetal hemoglobin
necessary for treatment of sickle cell disease. N Engl J Med 1988; 318:96–99.
6. Eaton WA, Hofrichter J. The biophysics of sickle cell hydroxyurea therapy. Science
1995; 268:1142–1143.
7. DeSimone J, Heller P, Hall L, Zwiers D. 5-Azacytidine stimulates fetal hemoglobin
synthesis in anemic baboons. Proc Natl Acad Sci USA 1982; 79:4428–4431.
8. Ley TJ, DeSimone J, Anagnou NP, Keller GH, Humphries RK, Turner PH, Young NS,
Keller P, Nienhuis AW. 5-Azacytidine selectively increases g-globin synthesis in a
patient with bþ thalassemia. N Engl J Med 1982; 307:1469–1475.
9. Charache S, Dover G, Smith K, Talbot CC Jr, Moyer M, Boyer S. Treatment of
sickle cell anemia with 5-azacytidine results in increased fetal hemoglobin production
and is associated with nonrandom hypomethylation of DNA around the g-d-b-globin
gene complex. Proc Natl Acad Sci USA 1983; 80:4842–4846.
10. Carr BI, Reilly JG, Smith SS, Winberg C, Riggs A. The tumorigenicity of
5-azacytidine in the male Fischer rat. Carcinogenesis 1984; 5:1583–1590.
11. Rodgers GP, Dover GJ, Noguchi CT, Schechter AN, Nienhuis AW. Hematologic
responses of patients with sickle cell disease to treatment with hydroxyurea. N Engl J
Med 1990; 322:1037–1045.
12. Charache S, Terrin ML, Moore RD, Dover GJ, Barton FB, Eckert SV, McMahon RP,
Bonds DR. Effect of hydroxyurea on the frequency of painful crises in sickle cell
anemia. Investigators of the multicenter study of hydroxyurea in sickle cell anemia.
N Engl J Med 1995; 332:1317–1322.
13. Rodgers GP, Rachmilewitz EA. Novel treatment options in the severe b-globin
disorders. Br J Haematol 1995; 91:263–268.
14. Olivieri NF, Rees DC, Ginder GD, Thein SL, Waye JS, Chang L, Brittenham GM,
Weatherall DJ. Elimination of transfusions through induction of fetal hemoglobin
synthesis in Cooley’s anemia. Ann NY Acad Sci 1998; 850:100–109.
15. Fibach E. Cell culture and animal models to screen for promising fetal hemoglobin-
stimulating compounds. Semin Hematol 2001; 38:374–381.
16. Fibach E, Burke LP, Schechter AN, Noguchi CT, Rodgers GP. Hydroxyurea increases
fetal hemoglobin in cultured erythroid cells derived from normal individuals and
patients with sickle cell anemia or b-thalassemia. Blood 1993; 81:1630–1635.
17. Fibach E, Rachmilewitz EA. Proliferation and differentiation of erythroid progenitors
in liquid culture: analysis of progenitors derived from patients with polycythemia vera.
Am J Hematol 1990; 35:151–156.
18. Fibach E. Techniques for studying stimulation of fetal hemoglobin production in
human erythroid cultures. Hemoglobin 1998; 22(5&6):445–458.
19. Fibach E, Rachmilewitz EA. The two-step liquid culture: a novel procedure for
studying maturation of human normal and pathological erythroid precursors. Stem
Cells 1993; 11(suppl 1):36–41.
20. Wang M, Tang DC, Liu W, Chin K, Zhu J, Fibach E, Rodgers GP. Hydroxyurea exerts
bi-modal dose-dependent effects on erythropoiesis in human cultured erythroid cells.
Br J Haematol 2002; 119:1–8.
86 Amoyal, Goldfarb, and Fibach
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.

21. Pope SH, Fibach E, Sun J, Chin K, Rodgers GP. Two-phase liquid culture system
models normal human adult erythropoiesis at the molecular level. Eur J Haematol
2000; 64:292–303.
22. Fibach E, Manor D, Treves A, Rachmilewitz EA. Growth of human normal erythroid
progenitors in liquid culture: a comparison with colony growth in semisolid culture.
Int J Cell Cloning 1991; 9:57–64.
23. Dalyot N, Fibach E, Rachmilewitz EA, Oppenheim A. Adult and neonatal patterns of
human globin gene expression are recapitulated in liquid cultures. Exp Hematol 1992;
20:1141–1145.
24. Hudgins WR, Fibach E, Safaya S, Rieder RF, Miller AC, Samid D. Transcriptional
upregulation of g-globin by phenylbutyrate and analogous aromatic fatty acids.
Biochem Pharmacol 1996; 52:1227–1233.
25. Letvin NL, Linch DC, Beardsley GP, McIntyre KW, Miller BA, Nathan DG. Influence
of cell cycle phase-specific agents on simian fetal hemoglobin synthesis. J Clin Invest
1985; 75:1999–2005.
26. Torrealba-de Ron AT, Papayannopoulou Th, Knapp MS, Fu MF, Knitter G,
Stamatoyannopoulos G. Perturbations in the erythroid marrow progenitor cell
pools may play a role in the augmentation of Hb F by 5-azacytidine. Blood 1984;
63:201–210.
27. Humphries RK, Dover G, Young NS, Moore JG, Charache S, Ley T, Nienhuis AW.
5-Azacytidine acts directly on both erythroid precursors and progenitors to increase
production of fetal hemoglobin. J Clin Invest 1985; 75:547–557.
28. Alter BP. Fetal erythropoiesis in stress hematopoiesis. Exp Hematol 1979; 7:200–209.
29. Fibach E, Kollia P, Schechter AN, Noguchi CT, Rodgers GP. Hemin-induced
acceleration of hemoglobin production in immature cultured erythroid cells:
preferential enhancement of fetal hemoglobin. Blood 1995; 85:2967–2974.
30. Fibach E, Schechter AN, Noguchi CT, Rodgers GP. Reducing erythropoietin in
cultures of human erythroid precursors elevates the proportion of fetal haemoglobin.
Br J Haematol 1994; 88:39–45.
31. Dover GJ, Boyer SH, Pembrey ME. F-cell production in sickle cell anemia: regulation
by genes linked to b-hemoglobin locus. Science 1981; 211:1441–1444.
32. Garner C, Tatu T, Reittie JE, Littlewood T, Darley J, Cervino S, Farrall M, Kelly P,
Spector TD, Thein SL. Genetic influences on F cells and other hematologic variables:
a twin heritability study. Blood 2000; 95:342–346.
33. Miller JL, Njoroge JM, Gubin AN, Rodgers GP. Prospective identification of erythroid
elements in cultured peripheral blood. Exp Hematol 1999; 27:624–629.
34. Fabricius E, Rajewsky F. Determination of hydroxyurea in mammalian tissues and
blood. Rev Eur Etud Clin Biol 1971; 16:679–683.
Received January 6, 2003
Accepted January 28, 2003
Flow Cytometry Analysis of HU Effect on Hb F 87
Hem
oglo
bin
Dow
nloa
ded
from
info
rmah
ealth
care
.com
by
CD
L-U
C B
erke
ley
on 1
0/28
/14
For
pers
onal
use
onl
y.





![-Thalassemia:HiJAKingIneffectiveErythropoiesisand IronOverloaddownloads.hindawi.com/journals/ah/2010/938640.pdf · 2019-07-31 · the Jak2-Stat5 pathway in erythroid cells [35]. Since](https://static.fdocument.org/doc/165x107/5e61a711f943864ec2353be9/thalassemiahijakingineffectiveerythropoiesisand-i-2019-07-31-the-jak2-stat5-pathway.jpg)

![Electronic Supplementary Information Magnesium β ... · 1 Electronic Supplementary Information Magnesium β-Ketoiminates as CVD Precursors for MgO Formation Elaheh Pousaneh[a], Tobias](https://static.fdocument.org/doc/165x107/60651f68f5d4f347af3c4c60/electronic-supplementary-information-magnesium-1-electronic-supplementary.jpg)