
Di (2-ethylhexyl) phthalate inhibits growth of mouse ovarian antral follicles through an oxidative...
8
Di (2-ethylhexyl) phthalate inhibits growth of mouse ovarian antral follicles through an oxidative stress pathway Wei Wang, Zelieann R. Craig, Mallikarjuna S. Basavarajappa, Rupesh K. Gupta, Jodi A. Flaws ⁎ Department of Comparative Biosciences, University of Illinois, 2001 S. Lincoln Ave, Urbana, IL, USA, 61802 abstract article info Article history: Received 8 September 2011 Revised 24 October 2011 Accepted 10 November 2011 Available online 23 November 2011 Keywords: Di (2-ethylhexyl) phthalate Antral follicles Oxidative stress Ovary Di (2-ethylhexyl) phthalate (DEHP) is a plasticizer that has been shown to inhibit growth of mouse antral fol- licles, however, little is known about the mechanisms by which DEHP does so. Oxidative stress has been linked to follicle growth inhibition as well as phthalate-induced toxicity in non-ovarian tissues. Thus, we hy- pothesized that DEHP causes oxidative stress and that this leads to inhibition of the growth of antral follicles. To test this hypothesis, antral follicles isolated from CD-1 mice (age 31–35 days) were cultured with vehicle control (dimethylsulfoxide [DMSO]) or DEHP (1–100 μg/ml) ± N-acetyl cysteine (NAC, an antioxidant at 0.25–1 mM). During culture, follicles were measured daily. At the end of culture, follicles were collected and processed for in vitro reactive oxygen species (ROS) assays to measure the presence of free radicals or for measurement of the expression and activity of various key antioxidant enzymes: Cu/Zn superoxide dis- mutase (SOD1), glutathione peroxidase (GPX) and catalase (CAT). The results indicate that DEHP inhibits the growth of follicles compared to DMSO control and that NAC (0.25–1 mM) blocks the ability of DEHP to inhibit follicle growth. Furthermore, DEHP (10 μg/ml) significantly increases ROS levels and reduces the ex- pression and activity of SOD1 compared to DMSO controls, whereas NAC (0.5 mM) rescues the effects of DEHP on ROS levels and SOD1. However, the expression and activity of GPX and CAT were not affected by DEHP treatment. Collectively, these data suggest that DEHP inhibits follicle growth by inducing production of ROS and by decreasing the expression and activity of SOD1. © 2011 Elsevier Inc. All rights reserved. Introduction Phthalates or phthalate esters (PE) are synthetic plasticizers that impart flexibility to polyvinylchloride products. They are present in a wide variety of products, including building materials, food packaging, toys, cosmetics, clothing and medical devices. More than 18 billion pounds of phthalates are used worldwide each year (Crinnion, 2010). Humans are exposed to phthalates through inhalation, ingestion and dermal absorption on a daily basis (Halden, 2010; Heudorf et al., 2007). Di (2-ethylhexyl) phthalate (DEHP) is one of the most common- ly used phthalate plasticizers. Since DEHP is not covalently bound to plastic matrix, it can leach out of products and contaminate the external environment. DEHP exposure is predominantly via food and appears to be close to the tolerable daily intake of 2 mg/day in the general popula- tion (Lyche et al., 2009). However, individuals undergoing certain medical procedures may be exposed to even higher levels of DEHP via plastic medical devices (Kamrin, 2009). DEHP is considered to be an endocrine disrupting chemical (EDC) based on its adverse effects on reproductive organs. Animal studies have shown that postnatal exposure to DEHP causes testicular atrophy and that in utero exposure to DEHP causes a number of abnormalities in male reproductive tract development in rodents (Christiansen et al., 2010; Clark and Cochrum, 2007; Martino-Andrade and Chahoud, 2009). In female rats, high dose acute exposure to DEHP reduces estra- diol production and causes anovulation (Lovekamp-Swan and Davis, 2003; Lyche et al., 2009; Martino-Andrade and Chahoud, 2009). Numer- ous human epidemiologic studies have reported associations between chronic exposure to DEHP and various adverse reproductive outcomes, including altered male reproductive development and function (Halden, 2010; Main et al., 2006; Swan, 2008), endometriosis (Cobellis et al., 2003; Kim et al., 2010a), and increased risk of premature birth and various pregnancy complications (Latini et al., 2006; Martino- Andrade and Chahoud, 2009; Whyatt et al., 2009). However, the mech- anisms by which DEHP affects the reproductive system are not fully elucidated. One potential mechanism by which DEHP causes reproductive abnormalities may be via oxidative stress. Oxidative stress is caused by an imbalance between the production and elimination of reactive oxygen species (ROS) in the system, leading to damage to DNA, lipid peroxidation of membranes and oxidative modification of proteins (Agarwal et al., 2005). ROS are a two-edged sword: they serve as key signal molecules in various physiological processes, but also play roles in pathological processes in female reproductive organs (Agarwal et Toxicology and Applied Pharmacology 258 (2012) 288–295 ⁎ Corresponding author. Fax: + 1 217 244 1652. E-mail addresses: [email protected] (W. Wang), [email protected] (Z.R. Craig), [email protected] (M.S. Basavarajappa), [email protected] (R.K. Gupta), jfl[email protected] (J.A. Flaws). 0041-008X/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2011.11.008 Contents lists available at SciVerse ScienceDirect Toxicology and Applied Pharmacology journal homepage: www.elsevier.com/locate/ytaap
Transcript of Di (2-ethylhexyl) phthalate inhibits growth of mouse ovarian antral follicles through an oxidative...

Toxicology and Applied Pharmacology 258 (2012) 288–295
Contents lists available at SciVerse ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /ytaap
Di (2-ethylhexyl) phthalate inhibits growth of mouse ovarian antral follicles throughan oxidative stress pathway
Wei Wang, Zelieann R. Craig, Mallikarjuna S. Basavarajappa, Rupesh K. Gupta, Jodi A. Flaws ⁎Department of Comparative Biosciences, University of Illinois, 2001 S. Lincoln Ave, Urbana, IL, USA, 61802
⁎ Corresponding author. Fax: +1 217 244 1652.E-mail addresses: [email protected] (W. Wang
(Z.R. Craig), [email protected] (M.S. Basavarajappa(R.K. Gupta), [email protected] (J.A. Flaws).
0041-008X/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.taap.2011.11.008
a b s t r a c t
a r t i c l e i n f oArticle history:Received 8 September 2011Revised 24 October 2011Accepted 10 November 2011Available online 23 November 2011
Keywords:Di (2-ethylhexyl) phthalateAntral folliclesOxidative stressOvary
Di (2-ethylhexyl) phthalate (DEHP) is a plasticizer that has been shown to inhibit growth of mouse antral fol-licles, however, little is known about the mechanisms by which DEHP does so. Oxidative stress has beenlinked to follicle growth inhibition as well as phthalate-induced toxicity in non-ovarian tissues. Thus, we hy-pothesized that DEHP causes oxidative stress and that this leads to inhibition of the growth of antral follicles.To test this hypothesis, antral follicles isolated from CD-1 mice (age 31–35 days) were cultured with vehiclecontrol (dimethylsulfoxide [DMSO]) or DEHP (1–100 μg/ml)±N-acetyl cysteine (NAC, an antioxidant at0.25–1 mM). During culture, follicles were measured daily. At the end of culture, follicles were collectedand processed for in vitro reactive oxygen species (ROS) assays to measure the presence of free radicals orfor measurement of the expression and activity of various key antioxidant enzymes: Cu/Zn superoxide dis-mutase (SOD1), glutathione peroxidase (GPX) and catalase (CAT). The results indicate that DEHP inhibitsthe growth of follicles compared to DMSO control and that NAC (0.25–1 mM) blocks the ability of DEHP toinhibit follicle growth. Furthermore, DEHP (10 μg/ml) significantly increases ROS levels and reduces the ex-pression and activity of SOD1 compared to DMSO controls, whereas NAC (0.5 mM) rescues the effects ofDEHP on ROS levels and SOD1. However, the expression and activity of GPX and CAT were not affected byDEHP treatment. Collectively, these data suggest that DEHP inhibits follicle growth by inducing productionof ROS and by decreasing the expression and activity of SOD1.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Phthalates or phthalate esters (PE) are synthetic plasticizers thatimpart flexibility to polyvinylchloride products. They are present in awide variety of products, including building materials, food packaging,toys, cosmetics, clothing and medical devices. More than 18 billionpounds of phthalates are used worldwide each year (Crinnion, 2010).Humans are exposed to phthalates through inhalation, ingestion anddermal absorption on a daily basis (Halden, 2010; Heudorf et al.,2007). Di (2-ethylhexyl) phthalate (DEHP) is one of themost common-ly used phthalate plasticizers. Since DEHP is not covalently bound toplasticmatrix, it can leach out of products and contaminate the externalenvironment. DEHP exposure is predominantly via food and appears tobe close to the tolerable daily intake of 2 mg/day in the general popula-tion (Lyche et al., 2009). However, individuals undergoing certainmedical procedures may be exposed to even higher levels of DEHP viaplastic medical devices (Kamrin, 2009).
DEHP is considered to be an endocrine disrupting chemical (EDC)based on its adverse effects on reproductive organs. Animal studies
), [email protected]), [email protected]
rights reserved.
have shown that postnatal exposure to DEHP causes testicular atrophyand that in utero exposure to DEHP causes a number of abnormalities inmale reproductive tract development in rodents (Christiansen et al.,2010; Clark and Cochrum, 2007; Martino-Andrade and Chahoud,2009). In female rats, high dose acute exposure to DEHP reduces estra-diol production and causes anovulation (Lovekamp-Swan and Davis,2003; Lyche et al., 2009;Martino-Andrade and Chahoud, 2009). Numer-ous human epidemiologic studies have reported associations betweenchronic exposure to DEHP and various adverse reproductive outcomes,including altered male reproductive development and function(Halden, 2010; Main et al., 2006; Swan, 2008), endometriosis(Cobellis et al., 2003; Kim et al., 2010a), and increased risk of prematurebirth and various pregnancy complications (Latini et al., 2006;Martino-Andrade and Chahoud, 2009; Whyatt et al., 2009). However, the mech-anisms by which DEHP affects the reproductive system are not fullyelucidated.
One potential mechanism by which DEHP causes reproductiveabnormalities may be via oxidative stress. Oxidative stress is causedby an imbalance between the production and elimination of reactiveoxygen species (ROS) in the system, leading to damage to DNA, lipidperoxidation of membranes and oxidative modification of proteins(Agarwal et al., 2005). ROS are a two-edged sword: they serve as keysignal molecules in various physiological processes, but also play rolesin pathological processes in female reproductive organs (Agarwal et

289W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
al., 2005). Studies have shown that ROS modulate multiple physiologi-cal processes from oocyte maturation to fertilization, embryo develop-ment and pregnancy (Al-Gubory et al., 2010; Dennery, 2010; Zhang etal., 2011). However, extra accumulation of ROS leads to ovarian apopto-sis, the process by which follicles undergo atresia and corpora luteaundergo regression (Agarwal et al., 2008; Lim and Luderer, 2010;Zhang et al., 2006). Excess ROS also affect angiogenesis, which is criticalfor follicular growth and corpus luteum formation (Agarwal et al.,2005). Epidemiology studies also have shown that oxidative stress isassociated with the age-related decline in fertility, endometriosis,pre-term labor, and unexplained infertility (Agarwal et al., 2005;Al-Gubory et al., 2010; Ruder et al., 2009).
Although studies have shown that DEHP is hydrolyzed into anactive metabolite mono(2-ethylhexyl) phthalate (MEHP) by lipasesand esterases in the intestine (Frederiksen et al., 2007), we electedto focus on revealing the effect of DEHP on mouse antral follicles forthe following reasons. First, after DEHP exposure, detectable amountsof DEHP remain in the plasma and peritoneal fluid and thus, can affectthe functions of reproductive organs (Cobellis et al., 2003). Second,high plasma levels of DEHP have been correlated with various femalereproductive disorders. For example, several studies have reported anassociation between high plasma levels of DEHP and endometriosis, in-dicating a possible role of DEHP in the pathogenesis of endometriosis(Cobellis et al., 2003; Kim et al., 2010a; Reddy et al., 2006a, 2006b). Astudy by Durmaz et al. also reported that high plasma levels of DEHPare correlated with pubertal gynecomastia (Durmaz et al., 2009).Third, DEHP has been used in different in vitro systems and has beenshown to exert toxic effects on endometrial cells, granulocytes andoocytes (Kim et al., 2010b; Mlynarcikova et al., 2009; Palleschi et al.,2009).
While there is limited information on whether DEHP causes oxi-dative stress in the female reproductive system, oxidative stress waspreviously suggested to represent a common mechanism in endo-crine disruptor-mediated dysfunction in reproduction (Gupta et al.,2006a, 2006b; Latchoumycandane and Mathur, 2002). Recent animalstudies have shown that DEHP causes oxidative stress in male repro-ductive tissues, specifically in Leydig cells, Sertoli cells and germ cellsby disrupting antioxidant defenses and increasing ROS (Botelho et al.,2009; Erkekoglu et al., 2010; Liu et al., 2005). Previous studies havealso shown that DEHP inhibits follicle growth of antral follicles inthe ovary (Gupta et al., 2010). However, the mechanisms bywhich DEHP inhibits growth are not fully understood. Thus, thisstudy tested the hypothesis that DEHP inhibits antral folliclegrowth through an oxidative stress pathway. To test this hypothe-sis, we determined whether DEHP reduces ROS in the ovary andtested whether N-acetyl-cysteine (NAC, an antioxidant) rescuesfollicles from the toxic effects of DEHP on antral follicles. Further-more, we investigated the mechanism by which DEHP induces ox-idative stress by examining the effect of DEHP on key antioxidantenzymes: Cu/Zn superoxide dismutase (SOD1), glutathione perox-idase (GPX) and catalase (CAT).
Materials and methods
Chemicals. DEHP was purchased from Sigma (St. Louis, MO). Stock so-lutions of DEHP were prepared using dimethylsulfoxide (DMSO)(Sigma, St. Louis, MO) as the solvent in various concentrations (133,
Table 1Sequences of primer sets used for gene expression analysis.
Gene name Abbreviation Forwa
Superoxide dismutase 1 Sod1 5′-AAGlutathione peroxidase Gpx 5′-CCCatalase Cat 5′-GCActin, beta Actb 5′-GG
13.3, and 1.33 mg/ml) that allowed an equal volume to be added toculture wells for each treatment group to control for solvent concen-tration. Final concentrations in culture were 1, 10, and 100 μg/ml ofDEHP, which are equivalent to approximately 2.56, 25.6, and256 μM. We chose these doses based on previous studies on the effectsof DEHP on cultured cells or follicles and based on their clinical rele-vance (Gillum et al., 2009; Lenie and Smitz, 2009). We also chosethese doses based on our preliminary dose–response experiment. N-acetyl-L-cysteine (NAC) was purchased from Sigma (St. Louis, MO).We chose NAC as opposed to other antioxidants as it is a known antiox-idant, free-radical scavenger, glutathione precursor, and increases theactivity of SOD and GPX enzymes (Tilly and Tilly, 1995). NAC has alsobeen shown to protect from oxidative stress in other tissues and celltypes (Gupta et al., 2006a; Hecht et al., 2002). For culture, a 100 mMstock solution of NACwas prepared usingα-MEM and the final concen-trations of NAC in each well of the culture were 0.25, 0.5, 1 and 2 mM.
Animals. CD-1 mice that were 31–35 days old were used for all experi-ments. The mice were housed at the University of Illinois at Urbana-Champaign, Veterinary Medicine Animal Facility under a 12:12 dark:light cycle. Food and water were provided ad libitum. All animal proce-dures were approved by the University of Illinois Institutional AnimalCare and Use Committee.
Follicle culture. Female CD-1 mice were euthanized and ovaries wereremoved. Based on relative size (250–350 μm) and appearance, antralfollicles were isolated mechanically from the ovaries and interstitialtissue was removed using fine watchmaker forceps (Gupta et al.,2006a). About 3–4 mice were used per experiment, providing ap-proximately 25–35 follicles per mouse. The isolated follicles wererandomly divided into different treatment groups (10–16 folliclesper group). Doses of vehicle control (DMSO), DEHP (1, 10, and100 μg/ml), DEHP (10 μg/ml)+NAC (0.5, 1, and 2 mM) were individu-ally prepared in supplemented α-MEM as described previously (Guptaet al., 2006a). The final solvent concentration was 0.075%. Antral folli-cles were cultured for 24–96 h at 37 °C in 95% air and 5% CO2. Non-treated controls (supplemented media only) were used in each experi-ment as a control for culture conditions. At the end of culture, follicleswere collected, snap frozen, and stored at -80 °C for later use.
Analysis of follicle growth. Follicle sizewas assessed at 24 h intervals bymeasuring follicle diameter on perpendicular axes using an invertedmicroscope equipped with a calibrated ocular micrometer. Follicle di-ameter measurements were averaged among treatment groups andplotted to compare the effects of chemical treatments on growth overtime. Data were presented as percent change over time. The statisticalanalyses were performed based on percentage change. At least threeseparate experiments were performed for each treatment group.
Gene expression analysis. Total RNA was extracted from follicles usingthe RNeasy Micro Kit (Qiagen, Inc., Valencia, CA) according to themanufacturer's protocol. To remove any possible genomic DNA con-tamination, RNA was further treated with DNAse. Messenger RNA(mRNA; 200 ng) was reverse transcribed to cDNA using an iScriptcDNA synthesis kit (Bio-Rad, Hercules, CA) following the man-ufacturer's instructions. The cDNA was diluted 1:4 with nuclease freewater. Quantitative real-time (qPCR) was conducted using a CFX96
rd Reverse
GGCCGTGTGCGTGCTGAA-3′ 5′-CAGGTCTCCAACATGCCTCT-3′TCAAGTACGTCCGACCTG-3′ 5′-CAATGTCGTTGCGGCACACC-3′AGATACCTGTGAACTGTC-3′ 5′-GTAGAATGTCCGCACCTGAG-3′GCACAGTGTGGGTGAC-3′ 5′-CTGGCACCACACCTTCTAC-3′
100
110
120
130
140
150
0h 24h 48h 72h 96h
Per
cen
t C
han
ge
NTDMSODEHP 1µg/mlDEHP 10µg/mlDEHP 100µg/ml
**
***
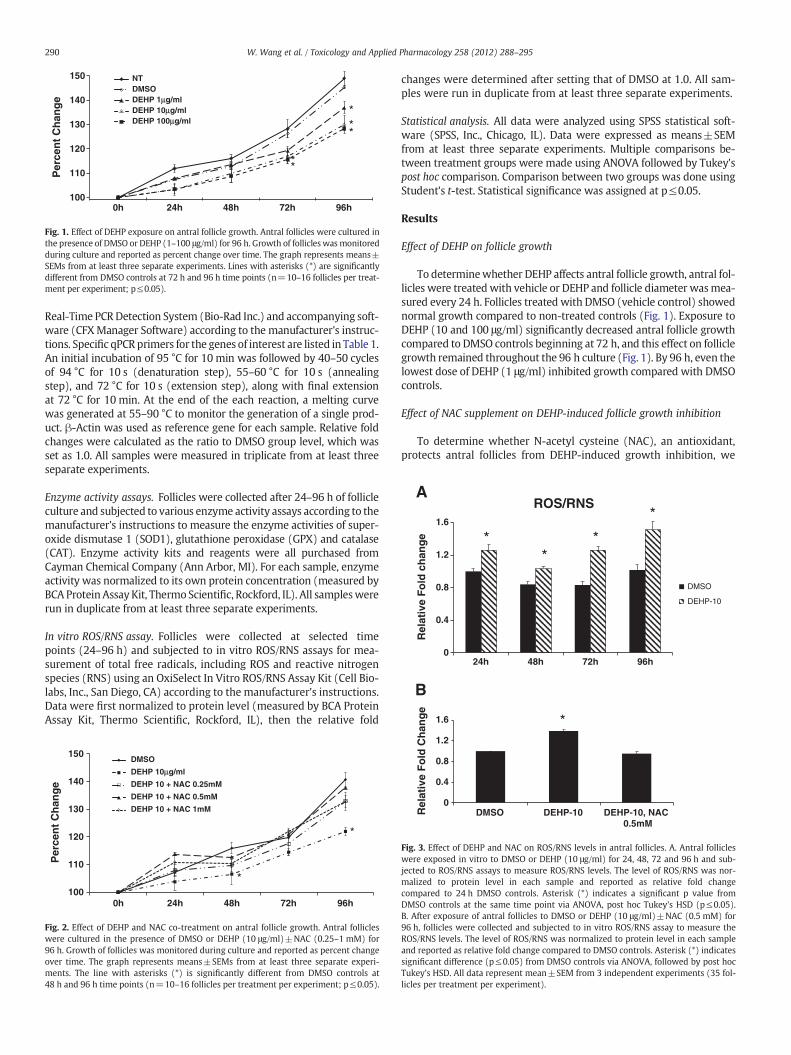
Fig. 1. Effect of DEHP exposure on antral follicle growth. Antral follicles were cultured inthe presence of DMSO or DEHP (1–100 μg/ml) for 96 h. Growth of follicles wasmonitoredduring culture and reported as percent change over time. The graph represents means±SEMs from at least three separate experiments. Lines with asterisks (*) are significantlydifferent from DMSO controls at 72 h and 96 h time points (n=10–16 follicles per treat-ment per experiment; p≤0.05).
1.6ng
e
*
0
0.4
0.8
1.2
1.6
24h 48h 72h 96h
Rel
ativ
e F
old
ch
ang
e
ROS/RNS
DMSO
DEHP-10
*
**
A
B
*
290 W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
Real-Time PCR Detection System (Bio-Rad Inc.) and accompanying soft-ware (CFXManager Software) according to the manufacturer's instruc-tions. Specific qPCR primers for the genes of interest are listed in Table 1.An initial incubation of 95 °C for 10 min was followed by 40–50 cyclesof 94 °C for 10 s (denaturation step), 55–60 °C for 10 s (annealingstep), and 72 °C for 10 s (extension step), along with final extensionat 72 °C for 10 min. At the end of the each reaction, a melting curvewas generated at 55–90 °C to monitor the generation of a single prod-uct. β-Actin was used as reference gene for each sample. Relative foldchanges were calculated as the ratio to DMSO group level, which wasset as 1.0. All samples were measured in triplicate from at least threeseparate experiments.
Enzyme activity assays. Follicles were collected after 24–96 h of follicleculture and subjected to various enzyme activity assays according to themanufacturer's instructions to measure the enzyme activities of super-oxide dismutase 1 (SOD1), glutathione peroxidase (GPX) and catalase(CAT). Enzyme activity kits and reagents were all purchased fromCayman Chemical Company (Ann Arbor, MI). For each sample, enzymeactivity was normalized to its own protein concentration (measured byBCA Protein Assay Kit, Thermo Scientific, Rockford, IL). All sampleswererun in duplicate from at least three separate experiments.
In vitro ROS/RNS assay. Follicles were collected at selected timepoints (24–96 h) and subjected to in vitro ROS/RNS assays for mea-surement of total free radicals, including ROS and reactive nitrogenspecies (RNS) using an OxiSelect In Vitro ROS/RNS Assay Kit (Cell Bio-labs, Inc., San Diego, CA) according to the manufacturer's instructions.Data were first normalized to protein level (measured by BCA ProteinAssay Kit, Thermo Scientific, Rockford, IL), then the relative fold
100
110
120
130
140
150
0h 24h 48h 72h 96h
Per
cen
t C
han
ge
DMSO
DEHP 10µg/ml
DEHP 10 + NAC 0.25mM
DEHP 10 + NAC 0.5mM
DEHP 10 + NAC 1mM
*
*
Fig. 2. Effect of DEHP and NAC co-treatment on antral follicle growth. Antral follicleswere cultured in the presence of DMSO or DEHP (10 μg/ml)±NAC (0.25–1 mM) for96 h. Growth of follicles was monitored during culture and reported as percent changeover time. The graph represents means±SEMs from at least three separate experi-ments. The line with asterisks (*) is significantly different from DMSO controls at48 h and 96 h time points (n=10–16 follicles per treatment per experiment; p≤0.05).
changes were determined after setting that of DMSO at 1.0. All sam-ples were run in duplicate from at least three separate experiments.
Statistical analysis. All data were analyzed using SPSS statistical soft-ware (SPSS, Inc., Chicago, IL). Data were expressed as means±SEMfrom at least three separate experiments. Multiple comparisons be-tween treatment groups were made using ANOVA followed by Tukey'spost hoc comparison. Comparison between two groups was done usingStudent's t-test. Statistical significance was assigned at p≤0.05.
Results
Effect of DEHP on follicle growth
To determinewhether DEHP affects antral follicle growth, antral fol-licles were treated with vehicle or DEHP and follicle diameter wasmea-sured every 24 h. Follicles treated with DMSO (vehicle control) showednormal growth compared to non-treated controls (Fig. 1). Exposure toDEHP (10 and 100 μg/ml) significantly decreased antral follicle growthcompared to DMSO controls beginning at 72 h, and this effect on folliclegrowth remained throughout the 96 h culture (Fig. 1). By 96 h, even thelowest dose of DEHP (1 μg/ml) inhibited growth compared with DMSOcontrols.
Effect of NAC supplement on DEHP-induced follicle growth inhibition
To determine whether N-acetyl cysteine (NAC), an antioxidant,protects antral follicles from DEHP-induced growth inhibition, we
0
0.4
0.8
1.2
DMSO DEHP-10 DEHP-10, NAC 0.5mM
Rel
ativ
e F
old
Ch
a
Fig. 3. Effect of DEHP and NAC on ROS/RNS levels in antral follicles. A. Antral follicleswere exposed in vitro to DMSO or DEHP (10 μg/ml) for 24, 48, 72 and 96 h and sub-jected to ROS/RNS assays to measure ROS/RNS levels. The level of ROS/RNS was nor-malized to protein level in each sample and reported as relative fold changecompared to 24 h DMSO controls. Asterisk (*) indicates a significant p value fromDMSO controls at the same time point via ANOVA, post hoc Tukey's HSD (p≤0.05).B. After exposure of antral follicles to DMSO or DEHP (10 μg/ml)±NAC (0.5 mM) for96 h, follicles were collected and subjected to in vitro ROS/RNS assay to measure theROS/RNS levels. The level of ROS/RNS was normalized to protein level in each sampleand reported as relative fold change compared to DMSO controls. Asterisk (*) indicatessignificant difference (p≤0.05) from DMSO controls via ANOVA, followed by post hocTukey's HSD. All data represent mean±SEM from 3 independent experiments (35 fol-licles per treatment per experiment).
291W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
conducted preliminary experiments to select a nontoxic level of NACfor the studies. Using the in vitro follicle culture system, the effect ofNAC on follicle growth was evaluated for 96 h. No significant folliclegrowth differences were observed in the NT, DMSO and NAC (0.25–2 mM) groups. However, follicles treated with NAC (5–10 mM) didnot grow (data not shown). Thus, NAC at 0.25–1 mM was used inall subsequent experiments.
Inhibition of follicle growth was observed with DEHP (10 μg/ml)compared to DMSO controls (Fig. 2). In contrast, NAC (0.25–1 mM)blocked the effect of DEHP-induced growth inhibition. Specifically,follicles co-treated with DEHP (10 μg/ml) and NAC (0.25–1 mM)had similar growth over time to DMSO controls (Fig. 2).
1.4Sod1 Expression
DMSO
A
Effect of DEHP on oxidative stress levels in antral follicles in vitro
Weobserved that the DEHP-induced follicle growth inhibition startsas early as 48 h (Fig. 2) and 72 h (Fig. 1), which suggests that DEHPmight induce oxidative stress even before 48 h or 72 h. To address thisquestion, we compared the levels of ROS/RNS in cultured follicles trea-ted with vehicle or DEHP (10 μg/ml) for 24, 48, 72 and 96 h. The results
0
0.2
0.4
0.6
0.8
1
1.2
DMSO DEHP-1 DEHP-10 DEHP-100
Rel
ativ
e F
old
Ch
ang
e
Sod1 Expression
*
0
0.2
0.4
0.6
0.8
1
1.2
DMSO DEHP-1 DEHP-10 DEHP-100
Rel
ativ
e F
old
Ch
ang
e
Gpx Expression
0
0.2
0.4
0.6
0.8
1
1.2
DMSO DEHP-1 DEHP-10 DEHP-100
Rel
ativ
e F
old
Ch
ang
e
Cat Expression
A
B
C
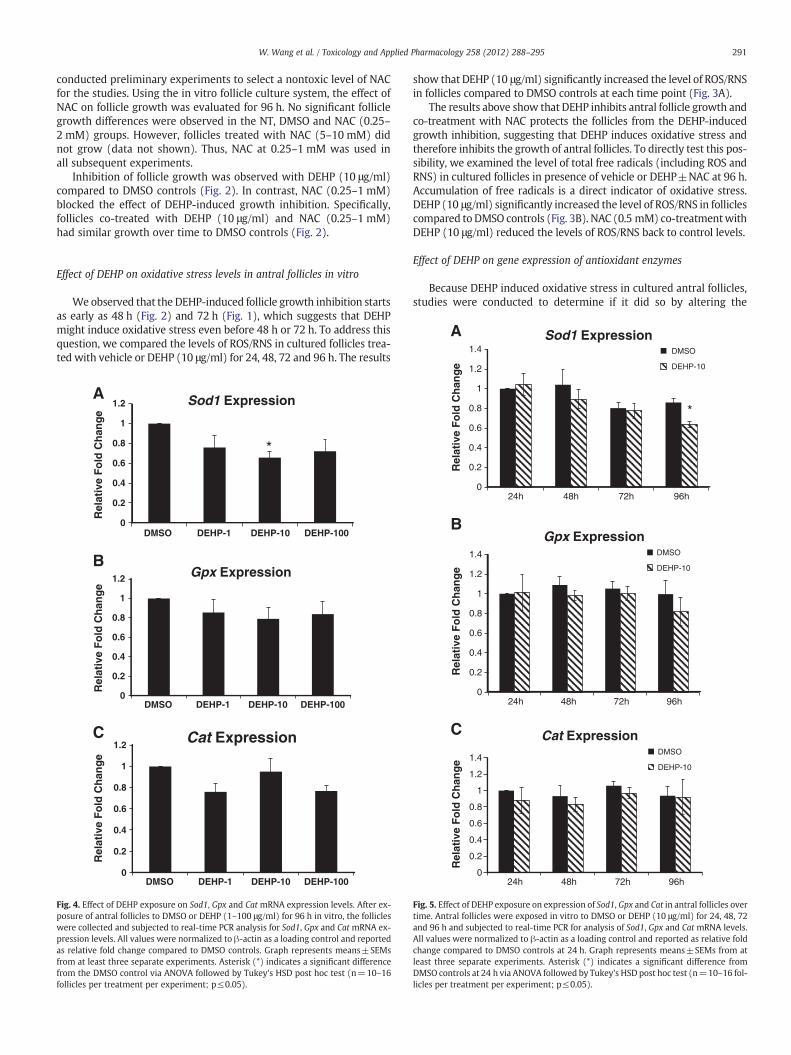
Fig. 4. Effect of DEHP exposure on Sod1, Gpx and Cat mRNA expression levels. After ex-posure of antral follicles to DMSO or DEHP (1–100 μg/ml) for 96 h in vitro, the follicleswere collected and subjected to real-time PCR analysis for Sod1, Gpx and Cat mRNA ex-pression levels. All values were normalized to β-actin as a loading control and reportedas relative fold change compared to DMSO controls. Graph represents means±SEMsfrom at least three separate experiments. Asterisk (*) indicates a significant differencefrom the DMSO control via ANOVA followed by Tukey's HSD post hoc test (n=10–16follicles per treatment per experiment; p≤0.05).
show that DEHP (10 μg/ml) significantly increased the level of ROS/RNSin follicles compared to DMSO controls at each time point (Fig. 3A).
The results above show that DEHP inhibits antral follicle growth andco-treatment with NAC protects the follicles from the DEHP-inducedgrowth inhibition, suggesting that DEHP induces oxidative stress andtherefore inhibits the growth of antral follicles. To directly test this pos-sibility, we examined the level of total free radicals (including ROS andRNS) in cultured follicles in presence of vehicle or DEHP±NAC at 96 h.Accumulation of free radicals is a direct indicator of oxidative stress.DEHP (10 μg/ml) significantly increased the level of ROS/RNS in folliclescompared to DMSO controls (Fig. 3B). NAC (0.5 mM) co-treatmentwithDEHP (10 μg/ml) reduced the levels of ROS/RNS back to control levels.
Effect of DEHP on gene expression of antioxidant enzymes
Because DEHP induced oxidative stress in cultured antral follicles,studies were conducted to determine if it did so by altering the
0
0.2
0.4
0.6
0.8
1
1.2
24h 48h 72h 96h
Rel
ativ
e F
old
Ch
ang
e DEHP-10
*
0
0.2
0.4
0.6
0.8
1
1.2
1.4
24h 48h 72h 96h
Rel
ativ
e F
old
Ch
ang
e
Gpx ExpressionDMSO
DEHP-10
0
0.2
0.4
0.6
0.8
1
1.2
1.4
24h 48h 72h 96h
Rel
ativ
e F
old
Ch
ang
e
Cat ExpressionDMSO
DEHP-10
B
C
Fig. 5. Effect of DEHP exposure on expression of Sod1, Gpx and Cat in antral follicles overtime. Antral follicles were exposed in vitro to DMSO or DEHP (10 μg/ml) for 24, 48, 72and 96 h and subjected to real-time PCR for analysis of Sod1, Gpx and Cat mRNA levels.All values were normalized to β-actin as a loading control and reported as relative foldchange compared to DMSO controls at 24 h. Graph represents means±SEMs from atleast three separate experiments. Asterisk (*) indicates a significant difference fromDMSO controls at 24 h via ANOVA followed by Tukey's HSD post hoc test (n=10–16 fol-licles per treatment per experiment; p≤0.05).

292 W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
expression of antioxidant enzymes required to detoxify the ROS/RNSin the system. Specifically, the expression levels of endogenous anti-oxidant enzymes Sod1, Gpx and Cat were compared in DMSO controlDEHP-treated follicles. At 96 h, only DEHP (10 μg/ml) significantly de-creased the expression of Sod1 compared to DMSO controls. OtherDEHP doses (1 and 100 μg/ml) did not significantly affect SOD1 ex-pression compared to controls (Fig. 4A). DEHP (1–100 μg/ml) didnot affect Gpx and Cat expression compared to DMSO controls(Figs. 4B–C).
To further elucidate whether DEHP affects gene expression atearly time points during the culture, the expression profiles of Sod1,Gpx and Cat were compared in DMSO and DEHP (10 μg/ml)-treatedfollicles at 24 h, 48 h, 72 h and 96 h. All the antioxidant enzymes(Sod1, Gpx and Cat) were consistently expressed throughout the cul-ture (Figs. 5A–C). Only DEHP (10 μg/ml) reduced the expression ofSod1 at 96 h (Fig. 5A). No significant changes were observed in anyof the selected enzymes at 0–72 h compared to DMSO controls(Figs. 5A–C).
0
0.2
0.4
0.6
0.8
1
1.2
DMSO DEHP-1 DEHP-10 DEHP-100
Rel
ativ
e F
old
Ch
ang
e
SOD1 Activity
*
0
0.2
0.4
0.6
0.8
1
1.2
DMSO DEHP-1 DEHP-10 DEHP-100
DMSO DEHP-1 DEHP-10 DEHP-100
Rel
ativ
e F
old
Ch
ang
e
GPX Activity
0
0.5
1
1.5
2
Rel
ativ
e F
old
Ch
ang
e
CAT Activity
A
B
C
Fig. 6. Effect of DEHP exposure on SOD1, GPX and CAT activities in antral follicles. An-tral follicles were exposed in vitro to DMSO or DEHP (1–100 μg/ml) for 96 h and thencollected and subjected to specific assays to measure the enzyme activities of SOD1,GPX and CAT. All values were normalized to protein level as a loading control andreported as relative fold change compared to DMSO controls. Graph representsmeans±SEMs from at least three separate experiments. Asterisk (*) indicates a signif-icant difference from DMSO controls via ANOVA followed by Tukey's HSD post hoc test(n=24 follicles per treatment per experiment; p≤0.05).
Effect of DEHP on activities of antioxidant enzymes
Since DEHP only inhibited expression of Sod1 and not Gpx or Catexpression, we then determined whether DEHP affects the activitiesof SOD1, GPX and CAT. Compared to DMSO controls, only DEHP(10 μg/ml) significantly inhibited the activity of SOD1, the key antiox-idant enzyme that catalyzes the dismutation of superoxide into oxy-gen and hydrogen peroxide (Fig. 6A). DEHP treatments (1–100 μg/ml) did not significantly affect the activities of GPX and CAT, the en-zymes that catalyze the decomposition of hydrogen peroxide(Figs. 6B–C).
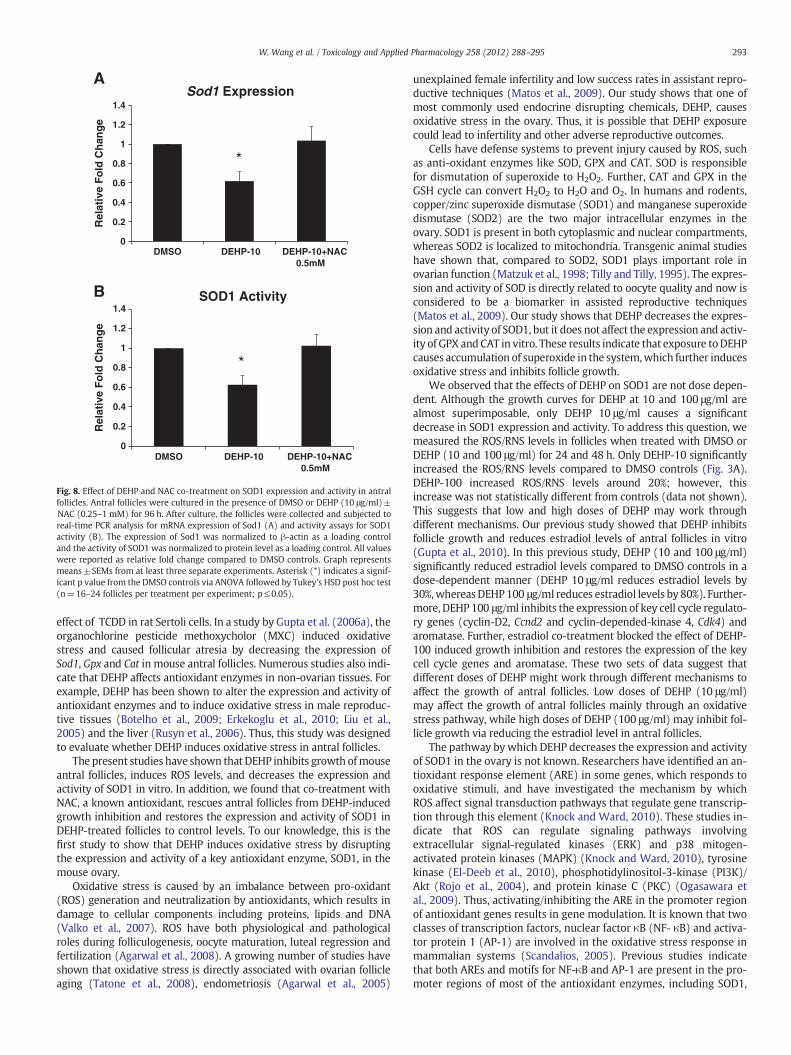
To further elucidate whether DEHP affects SOD1 activity at earlytime points during culture, the activity profiles of SOD1 were com-pared in DMSO and DEHP (10 μg/ml)-treated follicles at 24 h, 48 h,72 h and 96 h. The results show that DEHP (10 μg/ml) exposure sig-nificantly increases SOD1 activity at 24 h, but it significantly de-creases SOD activity at 72 and 96 h (Fig. 7).
Effect of NAC co-treatment on SOD1 expression and activity
DEHP-induced oxidative stress and inhibition of follicle growthcould be the result of decreased SOD1 expression and activity in thefollicles since SOD1 is the key antioxidant enzyme required for elim-ination of accumulated ROS/RNS in the system. Because addition ofNAC protects follicle growth from the DEHP-induced inhibition byreducing the oxidative stress in the system, studies were conductedto investigate whether NAC co-treatment protects the follicle fromthe DEHP-induced oxidative stress by rescuing the expression andactivity of SOD1. Compared to DMSO controls, DEHP (10 μg/ml) sig-nificantly reduced the expression and inhibited the activity of SOD1(Figs. 8A–B). Addition of NAC (0.5 mM) to the supplemented mediarestored the expression and activity of SOD1 to DMSO control level(Figs. 8A–B).
Discussion
Numerous studies have shown that EDCs induce oxidative stressin biological systems. In a study by Jin et al. (2008), TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin) induced oxidative stress by decreasingthe activity of antioxidant enzymes (SOD, GPX, CAT and glutathionereductase) in mouse testes. Aly and Khafagy (2011) reported a similar
0
0.4
0.8
1.2
1.6
24h 48h 72h 96h
Rel
ativ
e F
old
Ch
ang
e
SOD1 Activity
DMSO
DEHP-10
*
*
*
Fig. 7. Effect of DEHP exposure on SOD1 activity profile in antral follicles. Antral folli-cles were exposed in vitro to DMSO or DEHP (10 μg/ml) for 24, 48, 72 and 96 h andsubjected to SOD1 activity assays for analysis of SOD1 activity over time. All valueswere normalized to protein level as a loading control and shown as relative fold changecompared to 24 h DMSO controls. Graph represents means±SEMs from at least 3 sep-arate experiments. Asterisk (*) indicates a significant p value from the DMSO controlsat same time point via ANOVA, post hoc Tukey's HSD (n=16 follicles per treatment perexperiment; p≤0.05).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
DMSO DEHP-10 DEHP-10+NAC 0.5mM
Rel
ativ
e F
old
Ch
ang
eSod1 Expression
A
0
0.2
0.4
0.6
0.8
1
1.2
1.4
DMSO DEHP-10 DEHP-10+NAC 0.5mM
Rel
ativ
e F
old
Ch
ang
e
SOD1 Activity
*
*
B
Fig. 8. Effect of DEHP and NAC co-treatment on SOD1 expression and activity in antralfollicles. Antral follicles were cultured in the presence of DMSO or DEHP (10 μg/ml)±NAC (0.25–1 mM) for 96 h. After culture, the follicles were collected and subjected toreal-time PCR analysis for mRNA expression of Sod1 (A) and activity assays for SOD1activity (B). The expression of Sod1 was normalized to β-actin as a loading controland the activity of SOD1 was normalized to protein level as a loading control. All valueswere reported as relative fold change compared to DMSO controls. Graph representsmeans±SEMs from at least three separate experiments. Asterisk (*) indicates a signif-icant p value from the DMSO controls via ANOVA followed by Tukey's HSD post hoc test(n=16–24 follicles per treatment per experiment; p≤0.05).
293W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
effect of TCDD in rat Sertoli cells. In a study by Gupta et al. (2006a), theorganochlorine pesticide methoxycholor (MXC) induced oxidativestress and caused follicular atresia by decreasing the expression ofSod1, Gpx and Cat in mouse antral follicles. Numerous studies also indi-cate that DEHP affects antioxidant enzymes in non-ovarian tissues. Forexample, DEHP has been shown to alter the expression and activity ofantioxidant enzymes and to induce oxidative stress in male reproduc-tive tissues (Botelho et al., 2009; Erkekoglu et al., 2010; Liu et al.,2005) and the liver (Rusyn et al., 2006). Thus, this study was designedto evaluate whether DEHP induces oxidative stress in antral follicles.
The present studies have shown that DEHP inhibits growth ofmouseantral follicles, induces ROS levels, and decreases the expression andactivity of SOD1 in vitro. In addition, we found that co-treatment withNAC, a known antioxidant, rescues antral follicles from DEHP-inducedgrowth inhibition and restores the expression and activity of SOD1 inDEHP-treated follicles to control levels. To our knowledge, this is thefirst study to show that DEHP induces oxidative stress by disruptingthe expression and activity of a key antioxidant enzyme, SOD1, in themouse ovary.
Oxidative stress is caused by an imbalance between pro-oxidant(ROS) generation and neutralization by antioxidants, which results indamage to cellular components including proteins, lipids and DNA(Valko et al., 2007). ROS have both physiological and pathologicalroles during folliculogenesis, oocyte maturation, luteal regression andfertilization (Agarwal et al., 2008). A growing number of studies haveshown that oxidative stress is directly associated with ovarian follicleaging (Tatone et al., 2008), endometriosis (Agarwal et al., 2005)
unexplained female infertility and low success rates in assistant repro-ductive techniques (Matos et al., 2009). Our study shows that one ofmost commonly used endocrine disrupting chemicals, DEHP, causesoxidative stress in the ovary. Thus, it is possible that DEHP exposurecould lead to infertility and other adverse reproductive outcomes.
Cells have defense systems to prevent injury caused by ROS, suchas anti-oxidant enzymes like SOD, GPX and CAT. SOD is responsiblefor dismutation of superoxide to H2O2. Further, CAT and GPX in theGSH cycle can convert H2O2 to H2O and O2. In humans and rodents,copper/zinc superoxide dismutase (SOD1) and manganese superoxidedismutase (SOD2) are the two major intracellular enzymes in theovary. SOD1 is present in both cytoplasmic and nuclear compartments,whereas SOD2 is localized to mitochondria. Transgenic animal studieshave shown that, compared to SOD2, SOD1 plays important role inovarian function (Matzuk et al., 1998; Tilly and Tilly, 1995). The expres-sion and activity of SOD is directly related to oocyte quality and now isconsidered to be a biomarker in assisted reproductive techniques(Matos et al., 2009). Our study shows that DEHP decreases the expres-sion and activity of SOD1, but it does not affect the expression and activ-ity of GPX and CAT in vitro. These results indicate that exposure toDEHPcauses accumulation of superoxide in the system,which further inducesoxidative stress and inhibits follicle growth.
We observed that the effects of DEHP on SOD1 are not dose depen-dent. Although the growth curves for DEHP at 10 and 100 μg/ml arealmost superimposable, only DEHP 10 μg/ml causes a significantdecrease in SOD1 expression and activity. To address this question, wemeasured the ROS/RNS levels in follicles when treated with DMSO orDEHP (10 and 100 μg/ml) for 24 and 48 h. Only DEHP-10 significantlyincreased the ROS/RNS levels compared to DMSO controls (Fig. 3A).DEHP-100 increased ROS/RNS levels around 20%; however, thisincrease was not statistically different from controls (data not shown).This suggests that low and high doses of DEHP may work throughdifferent mechanisms. Our previous study showed that DEHP inhibitsfollicle growth and reduces estradiol levels of antral follicles in vitro(Gupta et al., 2010). In this previous study, DEHP (10 and 100 μg/ml)significantly reduced estradiol levels compared to DMSO controls in adose-dependent manner (DEHP 10 μg/ml reduces estradiol levels by30%,whereas DEHP100 μg/ml reduces estradiol levels by 80%). Further-more, DEHP 100 μg/ml inhibits the expression of key cell cycle regulato-ry genes (cyclin-D2, Ccnd2 and cyclin-depended-kinase 4, Cdk4) andaromatase. Further, estradiol co-treatment blocked the effect of DEHP-100 induced growth inhibition and restores the expression of the keycell cycle genes and aromatase. These two sets of data suggest thatdifferent doses of DEHP might work through different mechanisms toaffect the growth of antral follicles. Low doses of DEHP (10 μg/ml)may affect the growth of antral follicles mainly through an oxidativestress pathway, while high doses of DEHP (100 μg/ml) may inhibit fol-licle growth via reducing the estradiol level in antral follicles.
The pathway by which DEHP decreases the expression and activityof SOD1 in the ovary is not known. Researchers have identified an an-tioxidant response element (ARE) in some genes, which responds tooxidative stimuli, and have investigated the mechanism by whichROS affect signal transduction pathways that regulate gene transcrip-tion through this element (Knock and Ward, 2010). These studies in-dicate that ROS can regulate signaling pathways involvingextracellular signal-regulated kinases (ERK) and p38 mitogen-activated protein kinases (MAPK) (Knock and Ward, 2010), tyrosinekinase (El-Deeb et al., 2010), phosphotidylinositol-3-kinase (PI3K)/Akt (Rojo et al., 2004), and protein kinase C (PKC) (Ogasawara etal., 2009). Thus, activating/inhibiting the ARE in the promoter regionof antioxidant genes results in gene modulation. It is known that twoclasses of transcription factors, nuclear factor κB (NF- κB) and activa-tor protein 1 (AP-1) are involved in the oxidative stress response inmammalian systems (Scandalios, 2005). Previous studies indicatethat both AREs and motifs for NF-κB and AP-1 are present in the pro-moter regions of most of the antioxidant enzymes, including SOD1,

294 W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
SOD2, CAT and GPX (Banning et al., 2005; Scandalios, 2005). Interest-ingly, numerous studies have shown that DEHP induces oxidativestress and causes apoptosis in hepatocytes by activating ERK/MAPKand p38/MAPK, therefore activating several transcription factors, in-cluding NF-κB, AP-1 c-jun and c-fos (Ghosh et al., 2010; Lee andLim, 2011a). In mouse allergic effector cells, DEHP increases AP-1transcriptional activation via p38 MAPK phosphorylation (Oh andLim, 2010). Further, mono- (2-ethylhexyl) phthalate (MEHP), the ac-tive metabolite of DEHP, also activates NF-κB signaling in the rat testisand induces germ cell apoptosis (Rasoulpour and Boekelheide, 2005;Rogers et al., 2008). Thus, it is quite possible that DEHP alters the ex-pression of SOD1 through any of these pathways.
NAC is a thiol antioxidant and has beneficial clinical implicationsin HIV infection and cancer, as well as in heart, kidney and liver dis-ease (Kelly, 1998). It acts as a free radical scavenger and glutathioneprecursor in reproductive and non-reproductive tissues and protectsthem from oxidative stress induced apoptosis (Erkkila et al., 1998;Otala et al., 2002; Priya et al., 2011). Numerous studies have shownthat NAC inhibits apoptosis in cultured porcine ovarian primordialgerm cells (Lee et al., 2010), rat ovarian follicles (Tilly and Tilly,1995), bovine corpora luteal cells (Lohrke et al., 2010) and in culturedhuman ovarian cortex (Otala et al., 2002). NAC was able to restore thefertility of mice lacking γ glutamyl transpeptidase, an enzyme impor-tant for glutathione synthesis (Kumar et al., 2000). Our previous stud-ies also show that NAC protects mouse antral follicles from MXC-induced growth inhibition and atresia (Gupta et al., 2010). Thus,NAC seems to be a survival factor in reproductive organs via its effectsof redox control. Our current study supports and expands thesefindings.
The mechanism by which NAC acts to prevent DEHP-induced tox-icity is unknown. Recent studies show that DEHP induces oxidativestress via the MAPK–NF-κB pathway (Lee and Lim, 2011a, 2011b)and that NAC decreases the phosphorylation of various kinases (p38MAPK and ERK) and inhibits the activation of NF-κ (Lee and Lim,2011b; Sigala et al., 2011). Further, Rojo et al. (2004) reported thatmotifs for NF-κB are located in the promoter regions of SOD. Wehave shown that DEHP treatment decreases the expression and activ-ity of SOD1 and that this effect of DEHP is blocked by NAC. Thus, it ispossible that DEHP-induced ROS alters the NF-κB pathway to furtherregulate the antioxidant expression causing oxidative stress and thatNAC prevents these processes. In addition, a recent study shows thatNAC pre-treatment upregulates SOD2 and CAT in cadmium-inducedoxidative stress (Doi et al., 2010). Thus, it is possible that NAC co-treatment induces SOD2; therefore, decreasing the ROS level and re-storing the expression and activity of SOD1.
In conclusion, these studies show that DEHP causes oxidativestress in antral follicles, which leads to growth inhibition of these fol-licles. Future studies should examine if DEHP-induced oxidativestress leads to infertility in animal models and humans and if so,whether NAC can protect from DEHP-induced infertility.
Conflict of interest statement
The authors have no conflicts to disclose.
Acknowledgments
The authors thank Liying Gao for her outstanding technical help.This work was supported by the National Institutes of Health (NIH)R01ES019178 (JAF), a Billie Field Fellowship in Reproductive Biology(WW and ZRC) and an Environmental Toxicology Scholarship (MSB).
References
Agarwal, A., Gupta, S., Sharma, R.K., 2005. Role of oxidative stress in female reproduc-tion. Reprod. Biol. Endocrinol. 3, 28.
Agarwal, A., Gupta, S., Sekhon, L., Shah, R., 2008. Redox considerations in female repro-ductive function and assisted reproduction: from molecular mechanisms to healthimplications. Antioxid. Redox Signal. 10, 1375–1403.
Al-Gubory, K.H., Fowler, P.A., Garrel, C., 2010. The roles of cellular reactive oxygen species,oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 42,1634–1650.
Aly, H.A., Khafagy, R.M., 2011. 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-inducedcytotoxicity accompanied by oxidative stress in rat Sertoli cells: possible role ofmitochondrial fractions of Sertoli cells. Toxicol. Appl. Pharmacol. 252, 273–280.
Banning, A., Deubel, S., Kluth, D., Zhou, Z., Brigelius-Flohe, R., 2005. The GI-GPx gene is atarget for Nrf2. Mol. Cell. Biol. 25, 4914–4923.
Botelho, G.G., Bufalo, A.C., Boareto, A.C., Muller, J.C., Morais, R.N., Martino-Andrade, A.J.,Lemos, K.R., Dalsenter, P.R., 2009. Vitamin C and resveratrol supplementation to ratdams treated with di(2-ethylhexyl)phthalate: impact on reproductive and oxidativestress end points in male offspring. Arch. Environ. Contam. Toxicol. 57, 785–793.
Christiansen, S., Boberg, J., Axelstad, M., Dalgaard, M., Vinggaard, A.M., Metzdorff, S.B.,Hass, U., 2010. Low-dose perinatal exposure to di(2-ethylhexyl) phthalate inducesanti-androgenic effects in male rats. Reprod. Toxicol. 30, 313–321.
Clark, B.J., Cochrum, R.K., 2007. The steroidogenic acute regulatory protein as a target ofendocrine disruption in male reproduction. Drug Metab. Rev. 39, 353–370.
Cobellis, L., Latini, G., De Felice, C., Razzi, S., Paris, I., Ruggieri, F., Mazzeo, P., Petraglia, F.,2003. High plasma concentrations of di-(2-ethylhexyl)-phthalate in women withendometriosis. Hum. Reprod. 18, 1512–1515.
Crinnion, W.J., 2010. Toxic effects of the easily avoidable phthalates and parabens.Altern. Med. Rev. 15, 190–196.
Dennery, P.A., 2010. Oxidative stress in development: nature or nurture? Free Radic.Biol. Med. 49, 1147–1151.
Doi, T., Puri, P., Bannigan, J., Thompson, J., 2010. Pre-treatment with N-acetylcysteineupregulates superoxide dismutase 2 and catalase genes in cadmium-induced oxi-dative stress in the chick omphalocele model. Pediatr. Surg. Int. 27, 131–136.
Durmaz, E., Ozmert, E.N., Erkekoglu, P., Giray, B., Derman, O., Hincal, F., Yurdakok, K.,2009. Plasma phthalate levels in pubertal gynecomastia. Pediatrics 125, e122–129.
El-Deeb, I.M., Yoo, K.H., Lee, S.H., 2010. ROS receptor tyrosine kinase: a new potentialtarget for anticancer drugs. Med. Res. Rev. 31, 794–818.
Erkekoglu, P., Rachidi, W., Yuzugullu, O.G., Giray, B., Favier, A., Ozturk, M., Hincal, F.,2010. Evaluation of cytotoxicity and oxidative DNA damaging effects of di(2-ethyl-hexyl)-phthalate (DEHP) and mono(2-ethylhexyl)-phthalate (MEHP) on MA-10Leydig cells and protection by selenium. Toxicol. Appl. Pharmacol. 248, 52–62.
Erkkila, K., Hirvonen, V., Wuokko, E., Parvinen, M., Dunkel, L., 1998. N-acetyl-L-cysteineinhibits apoptosis in human male germ cells in vitro. J. Clin. Endocrinol. Metab. 83,2523–2531.
Frederiksen, H., Skakkebaek, N.E., Andersson, A.M., 2007. Metabolism of phthalates inhumans. Mol. Nutr. Food Res. 51, 899–911.
Ghosh, J., Das, J., Manna, P., Sil, P.C., 2010. Hepatotoxicity of di-(2-ethylhexyl)phthalateis attributed to calcium aggravation, ROS-mediated mitochondrial depolarization,and ERK/NF-kappaB pathway activation. Free Radic. Biol. Med. 49, 1779–1791.
Gillum, N., Karabekian, Z., Swift, L.M., Brown, R.P., Kay, M.W., Sarvazyan, N., 2009. Clin-ically relevant concentrations of di (2-ethylhexyl) phthalate (DEHP) uncouple car-diac syncytium. Toxicol. Appl. Pharmacol. 236, 25–38.
Gupta, R.K., Miller, K.P., Babus, J.K., Flaws, J.A., 2006a. Methoxychlor inhibits growthand induces atresia of antral follicles through an oxidative stress pathway. Toxicol.Sci. 93, 382–389.
Gupta, R.K., Schuh, R.A., Fiskum, G., Flaws, J.A., 2006b. Methoxychlor causes mitochon-drial dysfunction and oxidative damage in the mouse ovary. Toxicol. Appl. Pharma-col. 216, 436–445.
Gupta, R.K., Singh, J.M., Leslie, T.C.,Meachum, S., Flaws, J.A., Yao, H.H., 2010. Di-(2-ethylhexyl)phthalate and mono-(2-ethylhexyl) phthalate inhibit growth and reduce estradiollevels of antral follicles in vitro. Toxicol. Appl. Pharmacol. 242, 224–230.
Halden, R.U., 2010. Plastics and health risks. Annu. Rev. Public Health 31, 179–194.Hecht, S.S., Upadhyaya, P.,Wang,M., Bliss, R.L., McIntee, E.J., Kenney, P.M., 2002. Inhibition
of lung tumorigenesis in A/J mice by N-acetyl-S-(N-2-phenethylthiocarbamoyl)-L-cysteine and myo-inositol, individually and in combination. Carcinogenesis 23,1455–1461.
Heudorf, U., Mersch-Sundermann, V., Angerer, J., 2007. Phthalates: toxicology and ex-posure. Int. J. Hyg. Environ. Health 210, 623–634.
Jin, M.H., Hong, C.H., Lee, H.Y., Kang, H.J., Han, S.W., 2008. Toxic effects of lactational ex-posure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on development of male re-productive system: involvement of antioxidants, oxidants, and p53 protein.Environ. Toxicol. 25, 1–8.
Kamrin, M.A., 2009. Phthalate risks, phthalate regulation, and public health: a review. J.Toxicol. Environ. Health B Crit. Rev. 12, 157–174.
Kelly, G.S., 1998. Clinical applications of N-acetylcysteine. Altern. Med. Rev. 3, 114–127.Kim, S.H., Chun, S., Jang, J.Y., Chae, H.D., Kim, C.H., Kang, B.M., 2010a. Increased plasma
levels of phthalate esters in women with advanced-stage endometriosis: a pro-spective case–control study. Fertil. Steril. 95, 357–359.
Kim, Y.H., Kim, S.H., Lee, H.W., Chae, H.D., Kim, C.H., Kang, B.M., 2010b. Increased via-bility of endometrial cells by in vitro treatment with di-(2-ethylhexyl) phthalate.Fertil. Steril. 94, 2413–2416.
Knock, G.A., Ward, J.P., 2010. Redox regulation of protein kinases as a modulator of vas-cular function. Antioxid. Redox. Signal 15, 1531–1547.
Kumar, T.R., Wiseman, A.L., Kala, G., Kala, S.V., Matzuk, M.M., Lieberman, M.W., 2000.Reproductive defects in gamma-glutamyl transpeptidase-deficient mice. Endocri-nology 141, 4270–4277.
Latchoumycandane, C., Mathur, P.P., 2002. Induction of oxidative stress in the rat testisafter short-term exposure to the organochlorine pesticide methoxychlor. Arch.Toxicol. 76, 692–698.

295W. Wang et al. / Toxicology and Applied Pharmacology 258 (2012) 288–295
Latini, G., Del Vecchio, A., Massaro, M., Verrotti, A., C. D. E. F., 2006. In utero exposure tophthalates and fetal development. Curr. Med. Chem. 13, 2527–2534.
Lee, J., Lim, K.T., 2011a. Plant-originated glycoprotein (24 kDa) has an inhibitory effecton proliferation of BNL CL.2 cells in response to di(2-ethylhexyl)phthalate. CellBiochem. Funct. 29, 496–505.
Lee, J., Lim, K.T., 2011b. Plant-originated glycoprotein (24 kDa) has an inhibitory effecton proliferation of BNL CL.2 cells in response to di(2-ethylhexyl)phthalate. CellBiochem. Funct. 29, 496–505.
Lee, Y.J., Lee, D.M., Lee, C.H., Heo, S.H., Won, S.Y., Im, J.H., Cho, M.K., Nam, H.S., Lee, S.H.,2010. Suppression of human prostate cancer PC-3 cell growth by N-acetylcysteineinvolves over-expression of Cyr61. Toxicol. In Vitro 25, 199–205.
Lenie, S., Smitz, J., 2009. Steroidogenesis-disrupting compounds can be effectively stud-ied for major fertility-related endpoints using in vitro cultured mouse follicles.Toxicol. Lett. 185, 143–152.
Lim, J., Luderer, U., 2010. Oxidative damage increases and antioxidant gene expressiondecreases with aging in the mouse ovary. Biol. Reprod. 84, 775–782.
Liu, K., Lehmann, K.P., Sar, M., Young, S.S., Gaido, K.W., 2005. Gene expression profilingfollowing in utero exposure to phthalate esters reveals new gene targets in the eti-ology of testicular dysgenesis. Biol. Reprod. 73, 180–192.
Lohrke, B., Xu, J., Weitzel, J.M., Kruger, B., Goldammer, T., Viergutz, T., 2010. N-acetyl-cysteine impairs survival of luteal cells through mitochondrial dysfunction. Cyto-metry A 77, 310–320.
Lovekamp-Swan, T., Davis, B.J., 2003. Mechanisms of phthalate ester toxicity in the fe-male reproductive system. Environ. Health Perspect. 111, 139–145.
Lyche, J.L., Gutleb, A.C., Bergman, A., Eriksen, G.S., Murk, A.J., Ropstad, E., Saunders, M.,Skaare, J.U., 2009. Reproductive and developmental toxicity of phthalates. J. Toxi-col. Environ. Health B Crit. Rev. 12, 225–249.
Main, K.M., Mortensen, G.K., Kaleva, M.M., Boisen, K.A., Damgaard, I.N., Chellakooty, M.,Schmidt, I.M., Suomi, A.M., Virtanen, H.E., Petersen, D.V., Andersson, A.M., Toppari,J., Skakkebaek, N.E., 2006. Human breast milk contamination with phthalates andalterations of endogenous reproductive hormones in infants three months of age.Environ. Health Perspect. 114, 270–276.
Martino-Andrade, A.J., Chahoud, I., 2009. Reproductive toxicity of phthalate esters. Mol.Nutr. Food Res. 54, 148–157.
Matos, L., Stevenson, D., Gomes, F., Silva-Carvalho, J.L., Almeida, H., 2009. Superoxide dis-mutase expression in human cumulus oophorus cells.Mol. Hum. Reprod. 15, 411–419.
Matzuk, M.M., Dionne, L., Guo, Q., Kumar, T.R., Lebovitz, R.M., 1998. Ovarian function insuperoxide dismutase 1 and 2 knockout mice. Endocrinology 139, 4008–4011.
Mlynarcikova, A., Nagyova, E., Fickova, M., Scsukova, S., 2009. Effects of selected endo-crine disruptors on meiotic maturation, cumulus expansion, synthesis of hyaluro-nan and progesterone by porcine oocyte–cumulus complexes. Toxicol. In Vitro23, 371–377.
Ogasawara, N., Oguro, T., Sakabe, T., Matsushima, M., Takikawa, O., Isobe, K., Nagase, F.,2009. Hemoglobin induces the expression of indoleamine 2,3-dioxygenase in den-dritic cells through the activation of PI3K, PKC, and NF-kappaB and the generationof reactive oxygen species. J. Cell. Biochem. 108, 716–725.
Oh, P.S., Lim, K.T., 2010. IgE, COX-2, and IL-4 are ExpressedbyDEHP through p38MAPKandSuppressed by Plant Glycoprotein (75 kDa) in ICR Mice. Inflammation 34, 326–334.
Otala, M., Erkkila, K., Tuuri, T., Sjoberg, J., Suomalainen, L., Suikkari, A.M., Pentikainen,V., Dunkel, L., 2002. Cell death and its suppression in human ovarian tissue culture.Mol. Hum. Reprod. 8, 228–236.
Palleschi, S., Rossi, B., Diana, L., Silvestroni, L., 2009. Di(2-ethylhexyl)phthalate stimulatesCa(2+) entry, chemotaxis and ROS production in human granulocytes. Toxicol. Lett.187, 52–57.
Priya, S., Vijayalakshmi, P., Vivekanandan, P., Karthikeyan, S., 2011. Influence of N-acetylcysteine against dimethylnitrosamine induced hepatotoxicity in rats. Toxi-col. Ind. Health 27, 914–922.
Rasoulpour, R.J., Boekelheide, K., 2005. NF-kappaB is activated in the rat testis follow-ing exposure to mono-(2-ethylhexyl) phthalate. Biol. Reprod. 72, 479–486.
Reddy, B.S., Rozati, R., Reddy, B.V., Raman, N.V., 2006a. Association of phthalate esterswith endometriosis in Indian women. BJOG 113, 515–520.
Reddy, B.S., Rozati, R., Reddy, S., Kodampur, S., Reddy, P., Reddy, R., 2006b. High plasmaconcentrations of polychlorinated biphenyls and phthalate esters in women withendometriosis: a prospective case control study. Fertil. Steril. 85, 775–779.
Rogers, R., Ouellet, G., Brown, C., Moyer, B., Rasoulpour, T., Hixon, M., 2008. Cross-talkbetween the Akt and NF-kappaB signaling pathways inhibits MEHP-induced germcell apoptosis. Toxicol. Sci. 106, 497–508.
Rojo, A.I., Salinas, M., Martin, D., Perona, R., Cuadrado, A., 2004. Regulation of Cu/Zn-superoxide dismutase expression via the phosphatidylinositol 3 kinase/Akt path-way and nuclear factor-kappaB. J. Neurosci. 24, 7324–7334.
Ruder, E.H., Hartman, T.J., Goldman, M.B., 2009. Impact of oxidative stress on femalefertility. Curr. Opin. Obstet. Gynecol. 21, 219–222.
Rusyn, I., Peters, J.M., Cunningham, M.L., 2006. Modes of action and species-specificeffects of di-(2-ethylhexyl)phthalate in the liver. Crit. Rev. Toxicol. 36, 459–479.
Scandalios, J.G., 2005. Oxidative stress: molecular perception and transductionof signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 38,995–1014.
Sigala, I., Zacharatos, P., Toumpanakis, D., Michailidou, T., Noussia, O., Theocharis, S.,Roussos, C., Papapetropoulos, A., Vassilakopoulos, T., 2011. MAPKs and NF-kappaB differentially regulate cytokine expression in the diaphragm in responseto resistive breathing: the role of oxidative stress. Am. J. Physiol. Regul. Integr.Comp. Physiol. 300, R1152–R1162.
Swan, S.H., 2008. Environmental phthalate exposure in relation to reproductive out-comes and other health endpoints in humans. Environ. Res. 108, 177–184.
Tatone, C., Amicarelli, F., Carbone, M.C., Monteleone, P., Caserta, D., Marci, R., Artini, P.G., Piomboni, P., Focarelli, R., 2008. Cellular and molecular aspects of ovarian folli-cle ageing. Hum. Reprod. Update 14, 131–142.
Tilly, J.L., Tilly, K.I., 1995. Inhibitors of oxidative stress mimic the ability of follicle-stimulating hormone to suppress apoptosis in cultured rat ovarian follicles. Endo-crinology 136, 242–252.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M.T., Mazur, M., Telser, J., 2007. Free radicalsand antioxidants in normal physiological functions and human disease. Int. J. Bio-chem. Cell Biol. 39, 44–84.
Whyatt, R.M., Adibi, J.J., Calafat, A.M., Camann, D.E., Rauh, V., Bhat, H.K., Perera, F.P., An-drews, H., Just, A.C., Hoepner, L., Tang, D., Hauser, R., 2009. Prenatal di(2-ethyl-hexyl)phthalate exposure and length of gestation among an inner-city cohort.Pediatrics 124, e1213–e1220.
Zhang, X., Li, X.H., Ma, X., Wang, Z.H., Lu, S., Guo, Y.L., 2006. Redox-induced apoptosis ofhuman oocytes in resting follicles in vitro. J. Soc. Gynecol. Investig. 13, 451–458.
Zhang, W., Liu, Y., An, Z., Huang, D., Qi, Y., Zhang, Y., 2011. Mediating effect of ROS onmtDNA damage and low ATP content induced by arsenic trioxide in mouse oo-cytes. Toxicol. In Vitro 25, 979–984.


















