
Cytokine Receptor Common ² Chain as a Potential Activator of Cytokine Withdrawal-Induced
11
MOLECULAR AND CELLULAR BIOLOGY, 0270-7306/99/$04.0010 Nov. 1999, p. 7399–7409 Vol. 19, No. 11 Copyright © 1999, American Society for Microbiology. All Rights Reserved. Cytokine Receptor Common b Chain as a Potential Activator of Cytokine Withdrawal-Induced Apoptosis SHERN-FWU LEE, 1 HUEI-MEI HUANG, 1,2 JYH-RONG CHAO, 3 SHIRLEY LIN, 1,4 HSIN-FANG YANG-YEN, 3 AND JEFFREY J.-Y. YEN 1 * Institute of Biomedical Sciences 1 and Institute of Molecular Biology, 3 Academia Sinica, Graduate Institute of Life Sciences, National Defense Medical Center, 2 and Graduate Institute of Biology, Fu-Jen Catholic University, 4 Taipei, Taiwan Received 8 April 1999/Returned for modification 18 May 1999/Accepted 29 July 1999 Growth factors and cytokines play an important role in supporting cellular viability of various tissues during development due to their ability to suppress the default cell death program in each cell type. To date, neither the triggering molecule nor the transduction pathway of these default apoptosis programs is understood. In this study, we explored the possibility that cytokine receptors are involved in modulating cytokine withdrawal- induced apoptosis (CWIA) in hematopoietic cells. Expression of the exogenous cytokine receptor common b chain (bc), but not the a chains, accelerated CWIA in multiple cytokine-dependent cell lines. Reduction of the expression level of endogenous bc by antisense transcripts resulted in prolonged survival during cytokine deprivation, suggesting a critical role of bc in modulating CWIA. Fine mapping of the bc subunit revealed that a membrane-proximal cytoplasmic sequence, designated the death enhancement region (DER), was critical to the death acceleration effect of bc. Furthermore, DER accelerated cell death either as a chimeric membrane protein or as a cytosolic protein, suggesting that DER functions independently of the cytokine receptor and membrane anchorage. Cross-linking of the chimeric membrane-bound DER molecules by antibody or of the FK506-binding protein–DER fusion protein by a synthetic dimerizing agent, AP1510, did not abrogate the death acceleration effect. Transient transfection assays further indicated that DER promoted cell death in the absence of serum in the nonhematopoietic 293 cell line. In summary, our data suggest that bc plays an important role in modulating CWIA via an anchorage-independent and aggregation-insensitive mechanism. These findings may facilitate further studies on the signaling pathways of CWIA. During animal development, cells undergo vigorous selec- tion in most tissues where only some will survive. The number of cells that survive within a tissue is thought to be determined by the concentration of its survival factor(s). In a variety of growth factor-dependent cell types, supplementation with ex- ogenous survival growth factors usually results in hypertrophy of a given organ whereas withdrawal of survival factors induces programmed cell death, also called apoptosis (20). As the death-triggering molecule(s) is poorly defined, little is known about the signal transduction pathway of survival factor with- drawal-induced apoptosis. The hematopoietic system is one of the best-characterized examples where the population of each cell lineage is tightly regulated by cytokines (6). In response to an antigenic stimulus, the blood concentration of cytokines increases causing a rapid proliferation and accumulation of certain blood cells. When the antigen is later cleared, massive cell death occurs via apoptosis due to a lack of growth and survival cytokines. This prompt growth and death regulation of primary blood cells by cytokines is well preserved in many cytokine-dependent leukemic cell lines. Therefore, the cyto- kine-dependent hematopoietic cell line provides an excellent model for exploring the triggering mechanism of cytokine with- drawal-induced apoptosis (CWIA). Granulocyte-macrophage colony-stimulating factor (GM- CSF), interleukin 5 (IL-5), and IL-3 are potent hematopoietic growth and survival factors that not only promote proliferation but also support survival via their membrane receptor com- plexes on the cell surface. Functional GM-CSF, IL-5, and IL-3 receptors are composed of a ligand-recruiting a chain, which is specific for each cytokine, and a signal-transducing b subunit (bc), which is shared by all three cytokine receptors (8, 24, 46, 54). Humans possess only one b subunit (hbc) gene, whereas mice have two highly related genes for the signal transducing subunit mbc and mb IL-3 .mbc is 56% identical to hbc at the amino acid level and serves as the common subunit for murine IL-3 (mIL-3), GM-CSF, and IL-5 receptors. However, there is an extensive sequence homology between mbc and mb IL-3 (91% identity at the amino acid level). In addition, mb IL-3 forms a high-affinity receptor and transmits a proliferation signal with the mIL-3 receptor a subunit (mIL3Ra) but not with IL5Ra or the GM-CSF receptor a subunit (GMRa) (13, 15, 19). Upon ligand binding, bc becomes heavily tyrosine phosphorylated (46) and associates with many SH2-containing signaling proteins (45, 48). Although the antiapoptotic func- tion of the activated receptor is not clearly elucidated (5), several observations suggest that the receptor a chain and the phosphotyrosine-mediated signals play important roles in ac- tivating the antiapoptotic signals. The activation of tyrosine phosphorylation and proliferation function of bc were shown to depend on the presence of the cytoplasmic domain of the receptor a chain (38, 48), which associates with JAK2 kinase. The deletion mutant of bc which lacks a tyrosine residue in the cytoplasmic domain (b590) not only partially lost mitogenic activity but completely lost its antiapoptotic function (4). Fur- thermore, a cytoplasmic region required for activation of the Ras/Raf/mitogen-activated protein kinase pathway is essential for bc to transduce cytokine-dependent survival activity (21, 23). Expression of activated Ras protein in trans comple- mented the defect in apoptosis prevention of the mutant bc * Corresponding author. Mailing address: Institute of Biomedical Sciences, Academia Sinica, No. 128, Sec. 2, Yen-Jiou-Yuan Rd., Taipei 11529, Taiwan. Phone: 886-2-2652-3077. Fax: 886-2-2785-8847. E-mail: [email protected]. 7399 Downloaded from https://journals.asm.org/journal/mcb on 21 January 2022 by 222.117.231.222.
Transcript of Cytokine Receptor Common ² Chain as a Potential Activator of Cytokine Withdrawal-Induced

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/99/$04.0010
Nov. 1999, p. 7399–7409 Vol. 19, No. 11
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Cytokine Receptor Common b Chain as a Potential Activatorof Cytokine Withdrawal-Induced Apoptosis
SHERN-FWU LEE,1 HUEI-MEI HUANG,1,2 JYH-RONG CHAO,3 SHIRLEY LIN,1,4
HSIN-FANG YANG-YEN,3 AND JEFFREY J.-Y. YEN1*
Institute of Biomedical Sciences1 and Institute of Molecular Biology,3 Academia Sinica, Graduate Institute ofLife Sciences, National Defense Medical Center,2 and Graduate Institute of Biology,
Fu-Jen Catholic University,4 Taipei, Taiwan
Received 8 April 1999/Returned for modification 18 May 1999/Accepted 29 July 1999
Growth factors and cytokines play an important role in supporting cellular viability of various tissues duringdevelopment due to their ability to suppress the default cell death program in each cell type. To date, neitherthe triggering molecule nor the transduction pathway of these default apoptosis programs is understood. Inthis study, we explored the possibility that cytokine receptors are involved in modulating cytokine withdrawal-induced apoptosis (CWIA) in hematopoietic cells. Expression of the exogenous cytokine receptor common bchain (bc), but not the a chains, accelerated CWIA in multiple cytokine-dependent cell lines. Reduction of theexpression level of endogenous bc by antisense transcripts resulted in prolonged survival during cytokinedeprivation, suggesting a critical role of bc in modulating CWIA. Fine mapping of the bc subunit revealed thata membrane-proximal cytoplasmic sequence, designated the death enhancement region (DER), was critical tothe death acceleration effect of bc. Furthermore, DER accelerated cell death either as a chimeric membraneprotein or as a cytosolic protein, suggesting that DER functions independently of the cytokine receptor andmembrane anchorage. Cross-linking of the chimeric membrane-bound DER molecules by antibody or of theFK506-binding protein–DER fusion protein by a synthetic dimerizing agent, AP1510, did not abrogate thedeath acceleration effect. Transient transfection assays further indicated that DER promoted cell death in theabsence of serum in the nonhematopoietic 293 cell line. In summary, our data suggest that bc plays animportant role in modulating CWIA via an anchorage-independent and aggregation-insensitive mechanism.These findings may facilitate further studies on the signaling pathways of CWIA.
During animal development, cells undergo vigorous selec-tion in most tissues where only some will survive. The numberof cells that survive within a tissue is thought to be determinedby the concentration of its survival factor(s). In a variety ofgrowth factor-dependent cell types, supplementation with ex-ogenous survival growth factors usually results in hypertrophyof a given organ whereas withdrawal of survival factors inducesprogrammed cell death, also called apoptosis (20). As thedeath-triggering molecule(s) is poorly defined, little is knownabout the signal transduction pathway of survival factor with-drawal-induced apoptosis. The hematopoietic system is one ofthe best-characterized examples where the population of eachcell lineage is tightly regulated by cytokines (6). In response toan antigenic stimulus, the blood concentration of cytokinesincreases causing a rapid proliferation and accumulation ofcertain blood cells. When the antigen is later cleared, massivecell death occurs via apoptosis due to a lack of growth andsurvival cytokines. This prompt growth and death regulation ofprimary blood cells by cytokines is well preserved in manycytokine-dependent leukemic cell lines. Therefore, the cyto-kine-dependent hematopoietic cell line provides an excellentmodel for exploring the triggering mechanism of cytokine with-drawal-induced apoptosis (CWIA).
Granulocyte-macrophage colony-stimulating factor (GM-CSF), interleukin 5 (IL-5), and IL-3 are potent hematopoieticgrowth and survival factors that not only promote proliferationbut also support survival via their membrane receptor com-
plexes on the cell surface. Functional GM-CSF, IL-5, and IL-3receptors are composed of a ligand-recruiting a chain, which isspecific for each cytokine, and a signal-transducing b subunit(bc), which is shared by all three cytokine receptors (8, 24, 46,54). Humans possess only one b subunit (hbc) gene, whereasmice have two highly related genes for the signal transducingsubunit mbc and mbIL-3. mbc is 56% identical to hbc at theamino acid level and serves as the common subunit for murineIL-3 (mIL-3), GM-CSF, and IL-5 receptors. However, there isan extensive sequence homology between mbc and mbIL-3(91% identity at the amino acid level). In addition, mbIL-3forms a high-affinity receptor and transmits a proliferationsignal with the mIL-3 receptor a subunit (mIL3Ra) but notwith IL5Ra or the GM-CSF receptor a subunit (GMRa) (13,15, 19). Upon ligand binding, bc becomes heavily tyrosinephosphorylated (46) and associates with many SH2-containingsignaling proteins (45, 48). Although the antiapoptotic func-tion of the activated receptor is not clearly elucidated (5),several observations suggest that the receptor a chain and thephosphotyrosine-mediated signals play important roles in ac-tivating the antiapoptotic signals. The activation of tyrosinephosphorylation and proliferation function of bc were shownto depend on the presence of the cytoplasmic domain of thereceptor a chain (38, 48), which associates with JAK2 kinase.The deletion mutant of bc which lacks a tyrosine residue in thecytoplasmic domain (b590) not only partially lost mitogenicactivity but completely lost its antiapoptotic function (4). Fur-thermore, a cytoplasmic region required for activation of theRas/Raf/mitogen-activated protein kinase pathway is essentialfor bc to transduce cytokine-dependent survival activity (21,23). Expression of activated Ras protein in trans comple-mented the defect in apoptosis prevention of the mutant bc
* Corresponding author. Mailing address: Institute of BiomedicalSciences, Academia Sinica, No. 128, Sec. 2, Yen-Jiou-Yuan Rd., Taipei11529, Taiwan. Phone: 886-2-2652-3077. Fax: 886-2-2785-8847. E-mail:[email protected].
7399
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.

and supported long-term proliferation in association with GM-CSF (22). Additionally, another serine/threonine kinase, Akt/PKB, was suggested to be involved in the antiapoptotic func-tion of IL-3. Akt/PKB was activated by IL-3 in aphosphatidylinositol 3-kinase-dependent manner in the IL-3-dependent cell line Ba/F3. Active but not inactive forms ofAkt/PKB were found to phosphorylate BAD, a distinct mem-ber of the Bcl-2 family that promotes cell death, in vivo and invitro at the same residues that are phosphorylated in responseto IL-3 (7, 49).
Like most apoptotic programs, CWIA in the IL-3-dependentcell line requires activation of the caspase-3 like proteases andis sensitive to caspase inhibitors (39). However, to date neitherthe triggering molecule nor the transduction pathway of thisdefault apoptotic program is well understood. We therefore setout to determine whether the cytokine receptor itself was in-volved in CWIA. In this report, we show that the bc moleculeplays an important role in modulating CWIA. The bc moleculepromoted apoptosis via a cytoplasmic sequence, named thedeath enhancement region (DER), in a membrane anchorage-independent, aggregation-insensitive manner. The novel func-tion of bc in the modulation of apoptosis may shed light on themechanism of leukemogenesis of hematopoietic cells.
MATERIALS AND METHODS
Cell lines and culture conditions. Ba/F3, a murine IL-3-dependent pro-B-cellline (41), was a gift from Atsushi Miyajima, Institute of Molecular and CellularBiosciences, The University of Tokyo, Tokyo, Japan. Ba/F3 cells were main-tained in 10% fetal calf serum (FCS)-containing RPMI 1640 medium supple-mented with 55 mM b-mercaptoethanol and 10 U of mIL-3 per ml. HT-2, anmIL-2-dependent T-lymphocytic cell line (55) purchased from the AmericanType Culture Collection (Rockville, Md.), was maintained in a medium similarto that for Ba/F3 cells but supplemented with 10 U of mIL-2 per ml. TF-1, ahuman GM-CSF- or IL-3-dependent erythroleukemic cell line (25), was a giftfrom Toshio Kitamura, Department of Hematopoietic Growth Factor, Univer-sity of Tokyo. TF-1 cells were maintained in a medium similar to that for Ba/F3but supplemented with 20 U of human GM-CSF per ml. Teo4 is a pTet-Off (14)(Clontech, Palo Alto, Calif.)-transfected TF-1 cell line and was maintained inTF-1 culture medium plus 200 mg of G418 per ml. Antisense hbc transfectantsb(AS)1 and b(AS)2 were maintained in the same medium as that used for Teo4,with the inclusion of hygromycin B (200 mg/ml). The ectotropic package cell lineBOSC23 (43) and the human embryonic kidney cell line 293 were purchasedfrom the American Type Culture Collection and maintained in Dulbecco mod-ified Eagle medium (DMEM) supplemented with 10% FCS.
Plasmid DNA construction. Retroviral expression plasmids for wild-type hbc(pBabe- hbc) and hGMRa were constructed by inserting the coding sequencesfrom pKH97 (16) and pKH125 (47) (gifts from A. Miyajima) into the retroviralexpression vector pBabeHygro and pBabeNeo (32) (gifts from Hartmut Land,Imperial Cancer Research Fund, London, England), respectively, by standardmolecular cloning methods.
The antisense hbc plasmid was constructed by inserting the full-length hbc intothe tetracycline-regulatable vector pTRE (Clontech) in a reverse orientation.
Deletion mutants of hbc were created by inserting the Stop linker (top oligo-nucleotide, 59 CCCGGGTTAACTTAACTTAAGGATCC 39; bottom oligonu-cleotide, 59 GGATCCTTAAGTTAAGTTAACCCGGG 39) into the FspI, ApaI,and MunI sites of the hbc cDNA to generate b470, b590, and b772, respectively.To generate mutants b512, b590(D473-505), and b560(D473-505), the wild-typecytoplasmic domain of hbc was replaced with PCR-amplified DNA fragments atthe FspI site. Primers 59 CGCACGCGTAGAAAGTGGGAGGAGAAG 39 (59primer) and 59CGGGAATTCTCACCCCTGGTGTGGGGGACT 39 (39 primer)were used for the construction of b512, and primers 59 CGCACGCGTAGTCCCCCACACCAGGGG 39 (59 primer) and 59 CGCGAATTCTCAATCTGAGGCAGCTGGAGT 39 (39 primer) were used for the construction of b560(D473-505). The 59 primer of b560(D473-505) and the bottom primer of the Stop linkerwere used to generate b590(D473-505), with the mutant b590 used as the tem-plate DNA.
Plasmids pCD16/7/b471-590 and pCD16/7/Stop were both derived from plas-mid pCD16/7/Syk (30) (provided by Tadatsugu Taniguchi, Institute for Molec-ular and Cellular Biology, Osaka University, Osaka, Japan). The Stop linker(described above) was inserted into the blunted MluI site at the junction betweenCD7 and Syk cDNA to generate pCD16/7/Stop. For the construction of pCD16/7/b471-590, the PCR fragment encoding amino acids (aa) 471 to 590 of hbc(amplified by the 59 primer of b512 and the bottom primer of the Stop linker withb590 as the template DNA) was digested with XbaI and MluI and ligated with theXbaI-MluI DNA fragment containing the CD16/7 extracellular and transmem-
brane domain to generate the CD16/7/b471-590 chimeric molecule. The result-ing plasmid was cut at the XbaI site and blunted and then cut at the EcoRI siteto release the entire coding sequence. This fragment was subcloned into theSnaBI and EcoRI sites of pBabeHygro (32) to generate the retroviral expressionplasmid pCD16/7/b471-590.
The cDNA sequence encoding aa 471 to 590 of hbc was modified to be flankedby the MluI restriction enzyme site and the stop codon by using standard PCRtechnique. This PCR product was cloned into the EcoRI site of pcDNA3.1.His-A(Invitrogen) to generate plasmid pcDNA-3/ hbc(471-590). This plasmid encodesa cytoplasmic DER protein that fuses to the hexahistidine tag. An extra IRLRPpentapeptide sequence was created between the expression vector and the hbccoding sequence as a result of the plasmid construction strategy. The entirecoding sequence of this chimeric cDNA was then transferred to the retroviralexpression vector pBabeNeo by inserting the PmeI fragment into the SnaBI siteof pBabeNeo to generate pBabeHisbDER.
Plasmids pBabeF1E and pBabeF1bE were both derived from pCF1E (AriadPharmaceuticals, Inc., Cambridge, Mass.). The nucleotide sequence of bc en-coding aa 466 to 590 was amplified by PCR and subcloned into the SpeI site ofpCF1E to generate pCF1bE. The flanking restriction sites of the entire codingsequences of pCF1E and pCF1bE were modified and subcloned into the SnaBIand BamHI sites of pBabeNeo to generate plasmids pBabeF1E and pBabeF1bE.
Gene transfer by retroviral infection and lipofection. For retroviral infection,12 mg of plasmid DNA was transfected into 3 3 106 cells of the packaging cellline BOSC23 (42) with Lipofectamine (Life Science, Gaithersburg, Md.) accord-ing to the manufacturer’s protocol, and the 24-h conditioned media from thetransfected cells were used to infect mIL-3-dependent cell line Ba/F3, using thestandard protocol. The stable infectants were selected in medium containinghygromycin B (500 mg/ml) or G418 (800 mg/ml), and the expression of receptorsubunits was confirmed by flow cytometry and Western blot analysis.
TF-1 cells were transfected as previously described (4). Briefly, 2 mg of plasmidDNA was mixed with 2.5 ml of DMRIE-C (Life Science) in 0.5 ml of serum-freeDMEM and incubated at room temperature for 30 min; 1.2 3 106 TF-1 cellswere washed and resuspended in 50 ml of serum-free DMEM. Cells were mixedwith DMRIE-C-bound plasmid DNA and incubated at 37°C for 5 h. The trans-fected cells were washed twice and resuspended in fresh growth medium for 24to 48 h before G418 or hygromycin B was added to select for the stable clones.
For each transfection experiment, more than 20 independent subclones werefirst analyzed by flow cytometry (see below). All candidate clones were furtheranalyzed by Western blotting (see below). More than three positive transfectantswere subjected to functional analysis. Since the properties of all positive cloneswere similar, only the results from one representative clone, unless specificallyindicated, are presented and discussed.
Antibodies, immunoprecipitation, Western blot analysis, and antibody cross-linking. Antibodies were as follows; for immunoprecipitation and flow cytomet-ric analysis of hbc, S-16 (catalog no. sc-457; Santa Cruz Biotechnology, SantaCruz, Calif.); for Western blot analysis of the carboxyl terminus of hbc, C-20(catalog no. sc-675; Santa Cruz); for the amino terminus of hbc, N-20 (catalogno. sc-676; Santa Cruz); for immunoprecipitation, flow cytometric analysis, andWestern blot analysis of human GMRa (hGMRa), S-50 (catalog no. sc-456;Santa Cruz); for Western blot analysis of phosphotyrosine, PY20 (catalog no.p11120; Transduction Laboratories, Lexington, Ky.); for immunoprecipitationand Western blot analysis of STAT3, C-20 (catalog no. sc-482; Santa Cruz). Themonoclonal antibody for immunoprecipitation, flow cytometric analysis, andcross-linking of CD16 was LNK16 (catalog no. MCA1193XZ; Serotec, Oxford,England), and that for Western blot analysis of CD16 was 2H7 (catalog no.NCL-CD16; Novocastra, Newcastle upon Tyne, England).
Cells were lysed in radioimmunoprecipitation assay lysis buffer, and the celllysates were subjected to immunoprecipitation and Western blot analysis aspreviously described (4) except that the bands of interest were visualized with anenhanced chemiluminescence (ECL) detection system (Amersham, Little Chal-font, England). When reprobing the same blot with another antibody was nec-essary, membranes were treated in stripping buffer (100 mM 2-mercaptoethanol,2% sodium dodecyl sulfate, 62.5 mM Tris-HCl [pH 6.7]) at 55°C for 30 min priorto reprobing.
For antibody cross-linking experiments, cells were initially incubated with 2.5mg of anti-CD16 monoclonal antibody LNK16 per 107 cells in 0.5 ml of mediumat room temperature for 15 min. Cells were then cross-linked with 7.5 mg of goatanti-mouse immunoglobulin G1 antibody (Jackson ImmunoResearch Laborato-ries, Inc., West Grove, Pa.) at 37°C for 15 min before harvesting and analysis.
Cell viability, proliferation, and DNA fragmentation assay. Cell viability wasdetermined by trypan blue exclusion staining. Proliferation activity was measuredby a colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazoliumbromide (MTT) as a substrate as described by Mosmann et al. (33). For detec-tion of the DNA ladder during cell death, cells were treated as described in thetext and subjected to lysis and agarose gel electrophoresis as previously described(59).
Flow cytometric analysis of surface receptor expression. For analyzing surfacereceptor expression, 1 million cells were washed and resuspended in 1 ml ofstaining buffer (2% FCS and 0.1% sodium azide in phosphate-buffered saline).Cells were pelleted after 30 min on ice and mixed with 0.5 mg of primaryantibody. After incubation, cells were mixed with fluorescein isothiocyanate-
7400 LEE ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.
conjugated goat anti-mouse secondary antibody. Fluorescence intensity was an-alyzed by FACScan (Becton Dickinson, Mountain View, Calif.).
RT-PCR analysis. A single-tube reverse transcription (RT)-PCR format wasused to analyze the expression of the exogenous genes. The cytoplasmic RNAwas prepared from the target cells and subjected to RT-PCR analysis as specifiedfor the Access RT-PCR system (Promega, Madison, Wis.).
RESULTS
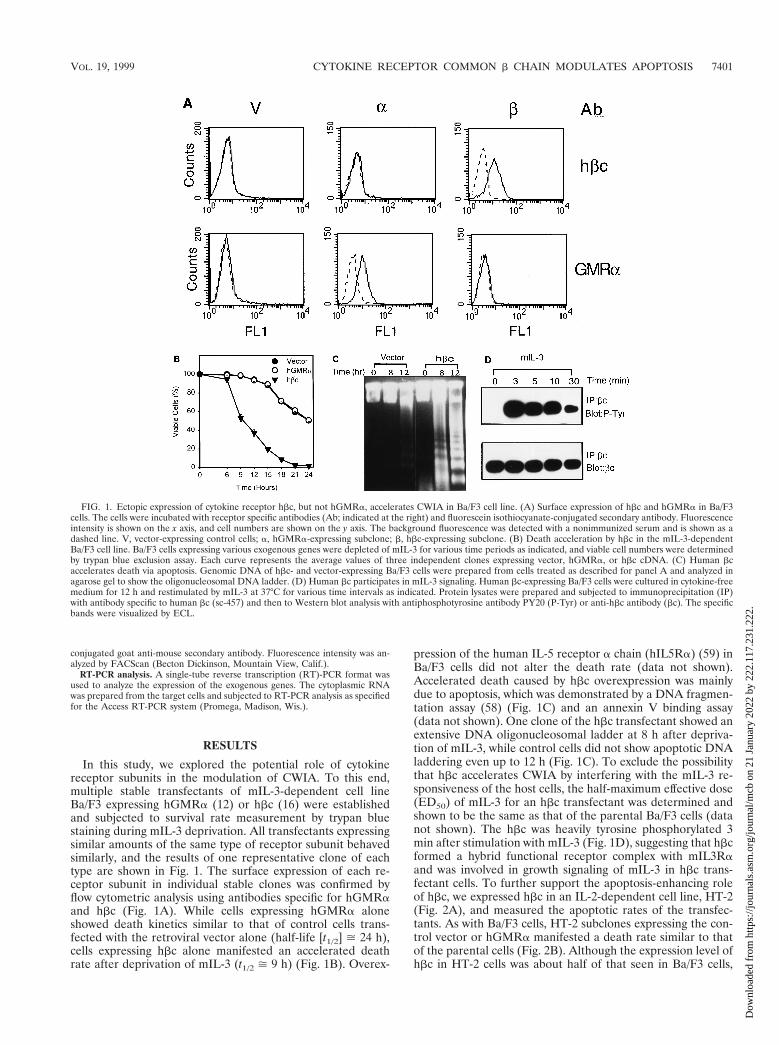
In this study, we explored the potential role of cytokinereceptor subunits in the modulation of CWIA. To this end,multiple stable transfectants of mIL-3-dependent cell lineBa/F3 expressing hGMRa (12) or hbc (16) were establishedand subjected to survival rate measurement by trypan bluestaining during mIL-3 deprivation. All transfectants expressingsimilar amounts of the same type of receptor subunit behavedsimilarly, and the results of one representative clone of eachtype are shown in Fig. 1. The surface expression of each re-ceptor subunit in individual stable clones was confirmed byflow cytometric analysis using antibodies specific for hGMRaand hbc (Fig. 1A). While cells expressing hGMRa aloneshowed death kinetics similar to that of control cells trans-fected with the retroviral vector alone (half-life [t1/2] > 24 h),cells expressing hbc alone manifested an accelerated deathrate after deprivation of mIL-3 (t1/2 > 9 h) (Fig. 1B). Overex-
pression of the human IL-5 receptor a chain (hIL5Ra) (59) inBa/F3 cells did not alter the death rate (data not shown).Accelerated death caused by hbc overexpression was mainlydue to apoptosis, which was demonstrated by a DNA fragmen-tation assay (58) (Fig. 1C) and an annexin V binding assay(data not shown). One clone of the hbc transfectant showed anextensive DNA oligonucleosomal ladder at 8 h after depriva-tion of mIL-3, while control cells did not show apoptotic DNAladdering even up to 12 h (Fig. 1C). To exclude the possibilitythat hbc accelerates CWIA by interfering with the mIL-3 re-sponsiveness of the host cells, the half-maximum effective dose(ED50) of mIL-3 for an hbc transfectant was determined andshown to be the same as that of the parental Ba/F3 cells (datanot shown). The hbc was heavily tyrosine phosphorylated 3min after stimulation with mIL-3 (Fig. 1D), suggesting that hbcformed a hybrid functional receptor complex with mIL3Raand was involved in growth signaling of mIL-3 in hbc trans-fectant cells. To further support the apoptosis-enhancing roleof hbc, we expressed hbc in an IL-2-dependent cell line, HT-2(Fig. 2A), and measured the apoptotic rates of the transfec-tants. As with Ba/F3 cells, HT-2 subclones expressing the con-trol vector or hGMRa manifested a death rate similar to thatof the parental cells (Fig. 2B). Although the expression level ofhbc in HT-2 cells was about half of that seen in Ba/F3 cells,
FIG. 1. Ectopic expression of cytokine receptor hbc, but not hGMRa, accelerates CWIA in Ba/F3 cell line. (A) Surface expression of hbc and hGMRa in Ba/F3cells. The cells were incubated with receptor specific antibodies (Ab; indicated at the right) and fluorescein isothiocyanate-conjugated secondary antibody. Fluorescenceintensity is shown on the x axis, and cell numbers are shown on the y axis. The background fluorescence was detected with a nonimmunized serum and is shown as adashed line. V, vector-expressing control cells; a, hGMRa-expressing subclone; b, hbc-expressing subclone. (B) Death acceleration by hbc in the mIL-3-dependentBa/F3 cell line. Ba/F3 cells expressing various exogenous genes were depleted of mIL-3 for various time periods as indicated, and viable cell numbers were determinedby trypan blue exclusion assay. Each curve represents the average values of three independent clones expressing vector, hGMRa, or hbc cDNA. (C) Human bcaccelerates death via apoptosis. Genomic DNA of hbc- and vector-expressing Ba/F3 cells were prepared from cells treated as described for panel A and analyzed inagarose gel to show the oligonucleosomal DNA ladder. (D) Human bc participates in mIL-3 signaling. Human bc-expressing Ba/F3 cells were cultured in cytokine-freemedium for 12 h and restimulated by mIL-3 at 37°C for various time intervals as indicated. Protein lysates were prepared and subjected to immunoprecipitation (IP)with antibody specific to human bc (sc-457) and then to Western blot analysis with antiphosphotyrosine antibody PY20 (P-Tyr) or anti-hbc antibody (bc). The specificbands were visualized by ECL.
VOL. 19, 1999 CYTOKINE RECEPTOR COMMON b CHAIN MODULATES APOPTOSIS 7401
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.
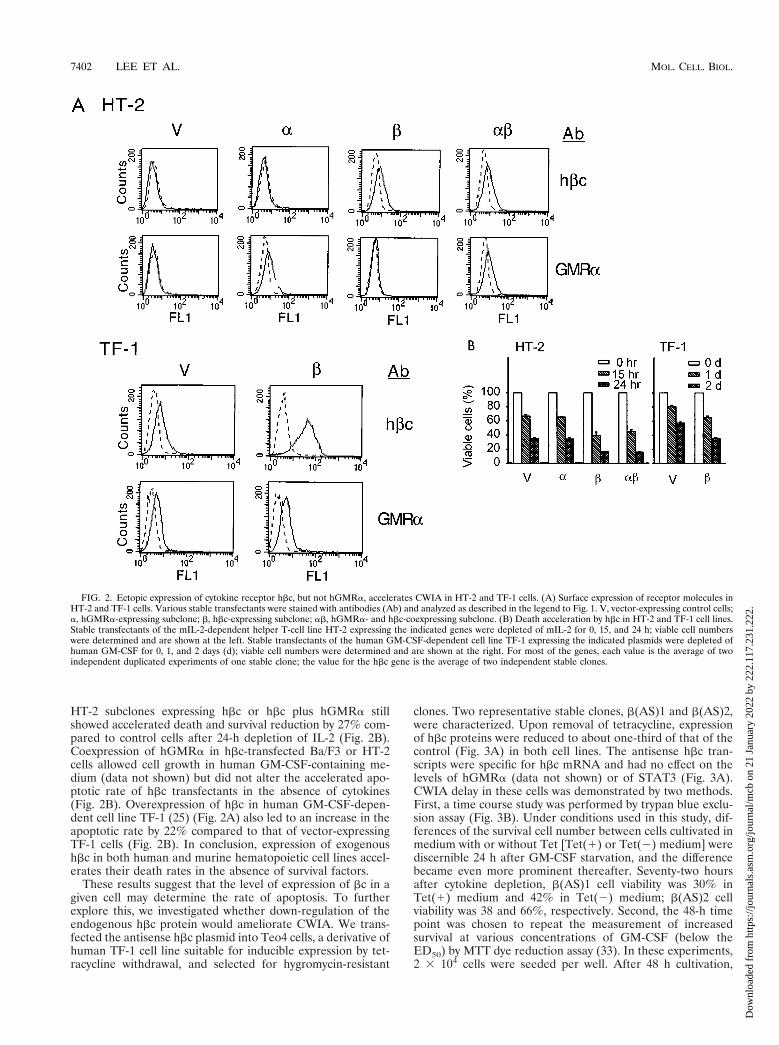
HT-2 subclones expressing hbc or hbc plus hGMRa stillshowed accelerated death and survival reduction by 27% com-pared to control cells after 24-h depletion of IL-2 (Fig. 2B).Coexpression of hGMRa in hbc-transfected Ba/F3 or HT-2cells allowed cell growth in human GM-CSF-containing me-dium (data not shown) but did not alter the accelerated apo-ptotic rate of hbc transfectants in the absence of cytokines(Fig. 2B). Overexpression of hbc in human GM-CSF-depen-dent cell line TF-1 (25) (Fig. 2A) also led to an increase in theapoptotic rate by 22% compared to that of vector-expressingTF-1 cells (Fig. 2B). In conclusion, expression of exogenoushbc in both human and murine hematopoietic cell lines accel-erates their death rates in the absence of survival factors.
These results suggest that the level of expression of bc in agiven cell may determine the rate of apoptosis. To furtherexplore this, we investigated whether down-regulation of theendogenous hbc protein would ameliorate CWIA. We trans-fected the antisense hbc plasmid into Teo4 cells, a derivative ofhuman TF-1 cell line suitable for inducible expression by tet-racycline withdrawal, and selected for hygromycin-resistant
clones. Two representative stable clones, b(AS)1 and b(AS)2,were characterized. Upon removal of tetracycline, expressionof hbc proteins were reduced to about one-third of that of thecontrol (Fig. 3A) in both cell lines. The antisense hbc tran-scripts were specific for hbc mRNA and had no effect on thelevels of hGMRa (data not shown) or of STAT3 (Fig. 3A).CWIA delay in these cells was demonstrated by two methods.First, a time course study was performed by trypan blue exclu-sion assay (Fig. 3B). Under conditions used in this study, dif-ferences of the survival cell number between cells cultivated inmedium with or without Tet [Tet(1) or Tet(2) medium] werediscernible 24 h after GM-CSF starvation, and the differencebecame even more prominent thereafter. Seventy-two hoursafter cytokine depletion, b(AS)1 cell viability was 30% inTet(1) medium and 42% in Tet(2) medium; b(AS)2 cellviability was 38 and 66%, respectively. Second, the 48-h timepoint was chosen to repeat the measurement of increasedsurvival at various concentrations of GM-CSF (below theED50) by MTT dye reduction assay (33). In these experiments,2 3 104 cells were seeded per well. After 48 h cultivation,
FIG. 2. Ectopic expression of cytokine receptor hbc, but not hGMRa, accelerates CWIA in HT-2 and TF-1 cells. (A) Surface expression of receptor molecules inHT-2 and TF-1 cells. Various stable transfectants were stained with antibodies (Ab) and analyzed as described in the legend to Fig. 1. V, vector-expressing control cells;a, hGMRa-expressing subclone; b, hbc-expressing subclone; ab, hGMRa- and hbc-coexpressing subclone. (B) Death acceleration by hbc in HT-2 and TF-1 cell lines.Stable transfectants of the mIL-2-dependent helper T-cell line HT-2 expressing the indicated genes were depleted of mIL-2 for 0, 15, and 24 h; viable cell numberswere determined and are shown at the left. Stable transfectants of the human GM-CSF-dependent cell line TF-1 expressing the indicated plasmids were depleted ofhuman GM-CSF for 0, 1, and 2 days (d); viable cell numbers were determined and are shown at the right. For most of the genes, each value is the average of twoindependent duplicated experiments of one stable clone; the value for the hbc gene is the average of two independent stable clones.
7402 LEE ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.

survival percentage was referred to the optical density readingof each sample compared to that of initiating cells. In theabsence of tetracyline, the survival cell numbers at all concen-trations of GM-CSF tested increased (Fig. 3C). The lower theconcentration of GM-CSF, the more profound the protectioneffects. In GM-CSF-free medium, survival cell numbers in-creased from 25% in Tet(1) medium to 60% in Tet(2) me-dium for b(AS)1 cells and from 35 to 70%, respectively, forb(AS)2 cells (Fig. 3C). The antisense construct did not com-pletely prevent CWIA in these experiments, possibly due to
incomplete ablation of hbc expression and/or because hbc maynot be the only cytokine receptor which can modulate CWIAin TF-1 cells (see below). Nevertheless, CWIA was dramati-cally inhibited when the expression level of endogenous hbcwas reduced. Therefore, not only exogenous hbc but also en-dogenous hbc regulates the death rate in a seemingly dose-dependent manner.
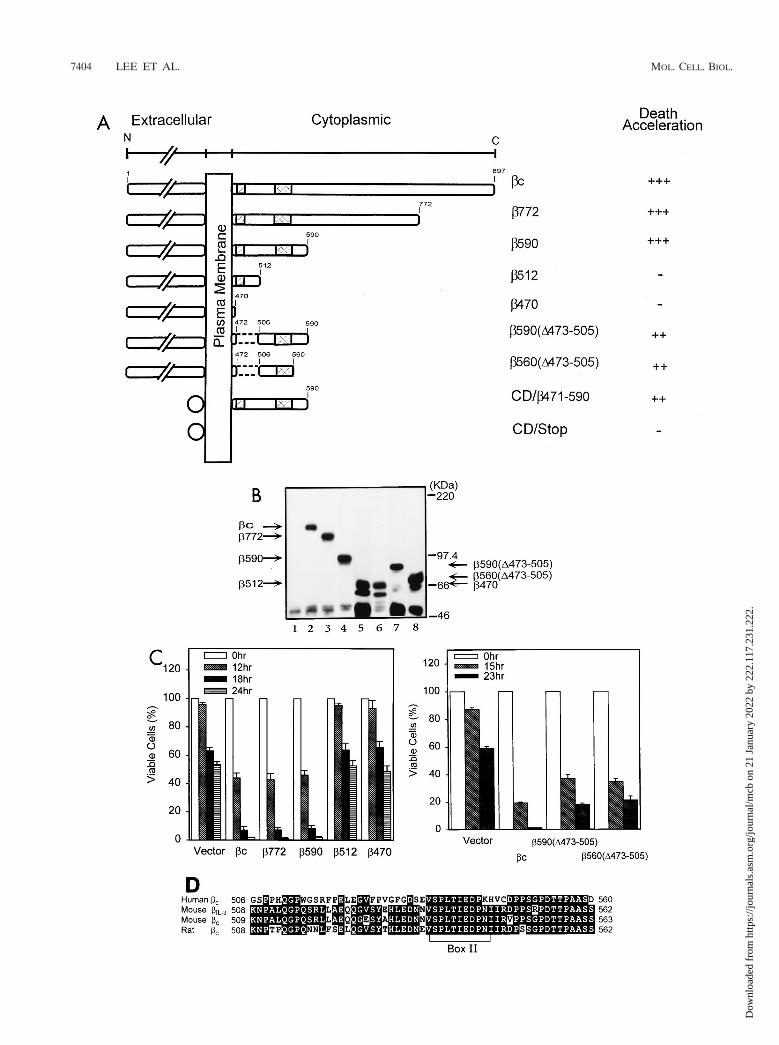
The structural domains essential for transducing prolifera-tion signals of the hbc molecule have been mapped (45, 47). Todetermine whether the domains needed for proliferation over-lap with that for apoptosis modulation, we constructed a seriesof deletion mutants to delineate sequence domains of hbccritical for the death acceleration effect (Fig. 4A). These con-structs were introduced into Ba/F3 cells by retroviral infection,and the hygromycin-resistant clones were analyzed and verifiedfor the expression of hbc mutants by flow cytometry (data notshown) and Western blot analysis (Fig. 4B). At least threeindependent clones for each mutation were subjected to sur-vival kinetics study after depletion of mIL-3. The death accel-eration ability of each mutation is summarized in Fig. 4A. Thehbc mutations with C-terminal deletions up to aa 773 (b772) or591 (b590) retained full death acceleration ability. However,with further deletions to aa 513 (b512) and 471 (b470), death-promoting activity was completely lost (Fig. 4C). Internal de-letion of the DNA sequence containing the conserved box I(31, 34, 40) from both b590 [i.e., b590(D473-505)] and b560[i.e., b560(D473-505)] did not abolish, but resulted in a slightreduction in, death acceleration ability (Fig. 4C). These datasuggest that the box I sequence is not essential for the apop-tosis-enhancing activity of hbc per se but may be required foroptimizing its death activity. The minimum sequence essentialfor accelerating apoptosis, designated the DER, is mapped bythis deletion analysis to a 54-aa peptide fragment from aa 506to 560 (Fig. 4D). In contrast to the dependence on the pres-ence of hGMRa for transducing proliferation signals of GM-CSF (24), coexpression of hGMRa with all hbc mutants testeddid not alter the survival kinetics in the absence of cytokines(data not shown).
These observations question whether DER can accelerateapoptosis via a mechanism independent of the cytokine recep-tor complex. To address this question, we fused the cDNAsequence encoding aa 471 to 590 of hbc, which contains DER,to a heterologous transmembrane protein, the chimeric CD16-CD7 molecule (30) (Fig. 4A, CD/b471-590), to test the abilityof this chimeric hbc molecule to affect CWIA. The constructCD16/7/Stop (Fig. 4A, CD/Stop), which contains no cytoplas-mic domain, was introduced to serve as a negative control.Surface expression of these chimeric CD16 antigens in alltransfectants was confirmed by flow cytometry (Fig. 5A) andWestern blot analysis (Fig. 5B). Ba/F3 cells expressing CD16/7/Stop did not show an altered rate of CWIA compared toparental cells (Fig. 5D, CD/S), whereas Ba/F3 cells expressingCD16/7/b471-590 showed moderately accelerated CWIA com-pared to parental cells, with the decrease of viable cells from 80to 50% at the 12-h time point (Fig. 5D, CD/b). These datasuggest that promotion of CWIA is independent of the GM-CSF receptor complex and is mediated via cis element(s) dis-tinct from those required for proliferation.
By taking advantage of the presence of the CD16 tag in thechimeric CD16/7/b471-590 protein, we further tested whetherdimerization or aggregation of the DER sequence would ab-rogate death acceleration activity. Since both box I and box IImotifs of hbc are present in the chimeric CD16/7/b471-590molecule and the JAK kinases were reported to be preassoci-ated with the box I sequence (38, 43), cross-linking of thismolecule is likely to activate the associated tyrosine kinases
FIG. 3. Antisense hbc transcript delays CWIA of GM-CSF-dependent cells.(A) Suppression of endogenous hbc protein expression by the antisense hbcplasmid in stable cell lines. Two antisense hbc-expressing TF-1 subclones[b(AS)1 and b(AS)2] and control cells (TRE) were cultured with (1) or without(2) tetracycline (2 mg/ml) for 48 h. Cell lysates were then prepared and subjectedto immunoprecipitation (IP) and Western blot analysis (Blot) with anti-hbcantibody (bc) and anti-STAT3 antibody (STAT3), respectively. (B) Time coursestudy of retarded CWIA of stable cell lines expressing hbc antisense RNA.Viable cell numbers of each cell line were determined by trypan blue staining andare relative to that of initial seeded cell number during cytokine depletion inTet(1) or Tet(2) medium. The values are averages of two independent exper-iments performed in duplicate. (C) Survival effect of antisense hbc in low-GM-CSF medium. Experiments were conducted as for panel B except that variouslow concentrations of human GM-CSF were used in the presence (open bar) orabsence (solid bar) of tetracycline and samples were analyzed only at the 48-htime point by MTT dye reduction assay. Percentage of survival is relative to thatof cells initially seeded. The values are averages of two independent experimentsperformed in triplicate.
VOL. 19, 1999 CYTOKINE RECEPTOR COMMON b CHAIN MODULATES APOPTOSIS 7403
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.
7404 LEE ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.

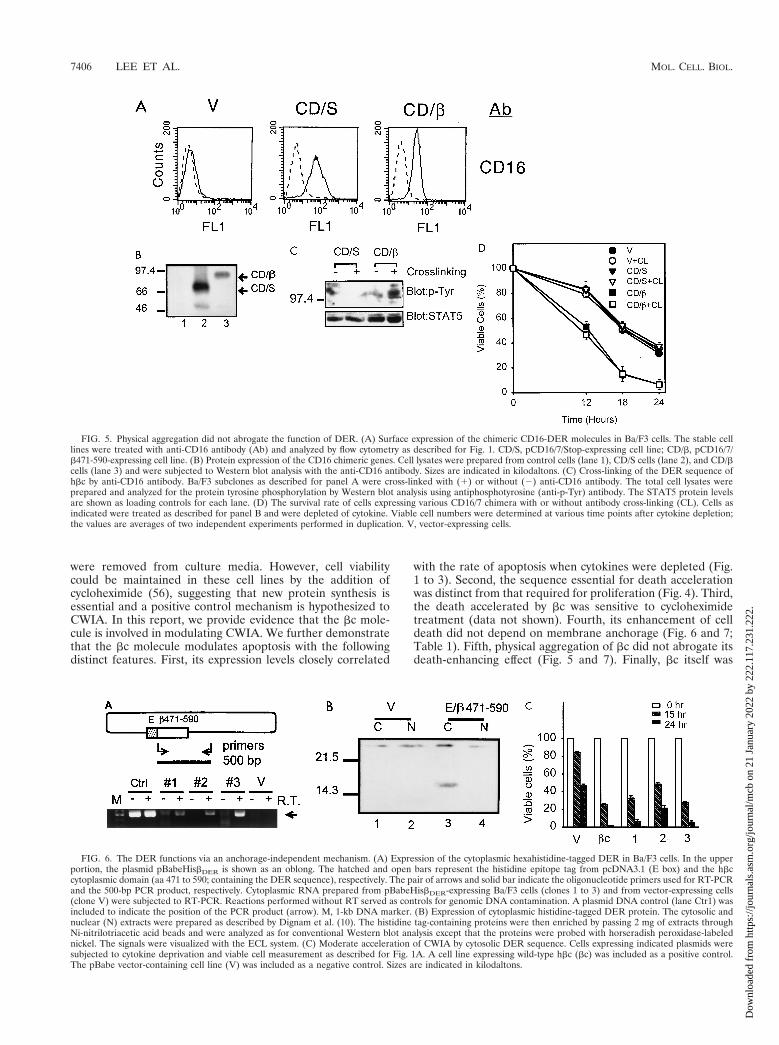
and result in tyrosine phosphorylation of certain cellular pro-teins. As indicated by the appearance of tyrosine-phosphory-lated protein signals in Western blot analysis (Fig. 5C, right-most lane), we successfully cross-linked the surface CD16/7/b471-590 molecules. Several tyrosine-phosphorylated cellularproteins (ranging from 97 to 150 kDa) were detectable inCD16/7/b471-590-expressing cells when the surface CD16 wascross-linked. Upon aggregation, the CD16/7/b471-590 mole-cules retained their ability to accelerate apoptosis (Fig. 5D,CD/b 1 CL). At 18 h, the surviving cells decreased from 55%for all control cells to 15% for CD16/7/b471-590-expressingcells regardless of cross-linking with the antibody. Our datastrongly suggest that physical aggregation caused by antibodycross-linking does not down-regulate the death accelerationactivity of the DER sequence. Given the fact that subclonesexpressing CD16/7/b471-590 grew well in mIL-3-containingmedium and that hbc-expressing HT-2 cells grew satisfactorilyin mIL-2, cytokines obviously abrogate in trans the apoptosisenhancing activity of DER.
Many membrane proteins function in an anchorage-depen-dent manner, due to the special membrane localization of theirsignaling components. To further understand the mechanismof death promotion by hbc and to explore whether this apo-ptosis acceleration activity is anchorage dependent, we con-structed the retroviral expression plasmid pBabeHisbDER, en-coding a hexahistidine-tagged cytoplasmic DER of hbc, andestablished several pBabeHisbDER-expressing cell lines by retro-viral infection. Although the protein product of pBabeHisbDERwas readily detectable by Western blot analysis in a transienttransfection assay with HeLa cells (data not shown), the ex-pression level of the cytoplasmic chimeric hbc protein in thesestable lines was very low and undetectable by a conventionalWestern blot analysis. Instead, we demonstrated the presenceof the RNA transcript of pBabeHisbDER by RT-PCR using aset of transgene specific oligonucleotide primers as depicted inFig. 6A. Three pBabeHisbDER-expressing clones, but not thecontrol vector transformant cells, showed a reverse transcrip-tase-dependent 500-bp specific PCR product (Fig. 6A, lanes 4,6, and 8 versus lane 10). Additionally, we enriched the histidinetag-containing proteins with Ni-nitrilotriacetic acid beads fromabout 2 mg of the cytoplasmic and nuclear proteins and ana-lyzed these protein samples as for conventional Western blot-ting except that the histidine tag-containing proteins wereprobed with horseradish peroxidase-labeled nickel (Kirke-gaard & Perry Laboratories, Gaithersburg, Md.). As shown inFig. 6B, a 14-kDa protein was detected in the cytoplasmicfraction of pBabeHisbDER-expressing cells (lane 3) but not inthe nuclear fraction or in vector-expressing cells (lanes 1, 2,and 4). In the death rate determination experiment (Fig. 6C),clones 1 and 3 showed death kinetics comparable to that of thewild-type hbc transformant (columns 1 and 3 versus columnV). Clone 2 showed a death rate slightly lower than the wild-type hbc control rate but still much higher than the control rate
(column 2). Therefore, the DER sequence is capable of accel-erating CWIA in an anchorage-independent manner.
To further elucidate the mode of DER’s function, we ex-pressed cytoplasmic DER as an FK506-binding protein(FKBP) fusion protein to explore the effect of dimerization onthe deleterious function of DER. Intriguingly, the cytoplasmicFKBP-DER fusion protein was again undetectable in stabletransfectants with anti-FKBP or hemagglutinin antigen anti-body even though it was highly expressed in a transient trans-fection experiment (data not shown). An RT-PCR analysis wascarried out instead to demonstrate the expression of the fusiongene. Using the pair of primers depicted in Fig. 7A, DNAfragments the size of ;500 and ;1,000 bp were specificallyamplified from the plasmids pBabeF1E and pBabeF1bE, re-spectively (Fig. 7A, lanes 1 to 4). The stable clone expressingpBabe vector showed no PCR product (lanes 5 and 6), andclones expressing pBabeF1E and pBabeF1bE showed a RT-dependent PCR product of predicted size (lanes 8, 10, 12, and14). Apoptotic kinetics was then studied in the presence of thedimerizing agent AP1510. Following the course of cytokinedeprivation, pBabe-F1bE-expressing cells manifested acceler-ated death faster than that of controls regardless of the pres-ence of AP1510 (Fig. 7B). These results further supported thenotion that DER-enhanced apoptosis is independent of mem-brane anchorage and insensitive to physical dimerization.
Finally, we examined whether hbc can promote death in anonhematopoietic cell line. This issue was explored in thehuman kidney cell line 293 by a transient assay measuring lossof green fluorescence protein (GFP) expression in apoptoticcells as previously described (4). As observed in the controlgroup, neither pBabe vector nor pcDNA3 vector caused anysignificant loss of GFP expression in the absence of serum(Table 1). However, the expression of wild-type hbc led to adecrease of 18.95% GFP1 cells upon serum starvation com-pared to cells in serum medium (Table 1). The expression ofcytosolic DER-containing sequence, i.e., pcDNA-hbc(471-590), also caused a decrease of 11.25% GFP1 cells upon serumstarvation (Table 1). These data confirm the anchorage-inde-pendent nature of the death acceleration activity of DER andfurther suggest that serum starvation sensitizes the 293 cells tothe death-promoting effect of hbc.
DISCUSSION
Hematopoietic cells differ from most other cell types in thattheir survival in vitro has an absolute requirement for specificgrowth factors. Growth factors are required continuouslythroughout the developmental program, and removal ofgrowth factors at any stage during differentiation into the ma-ture cells leads to apoptosis. Therefore, programmed celldeath was suggested to have a physiological significance inhematopoiesis (9). In many factor-dependent hematopoieticcell lines, apoptosis was observed when the relevant cytokines
FIG. 4. Mapping of the DER of hbc. (A) Schematic depiction of cytoplasmic deletion mutants of cytokine receptor hbc. Position 1 is the first methionine of theopen reading frame bc, full-length wild-type molecule of hbc; hatched and double-hatched boxes, conserved cytoplasmic box I and box II sequences, respectively; dashedlines, the internal deletion; circles, extracellular domains of the chimeric CD16/7/b471-590 and control CD16/7/Stop molecules. Death acceleration is indicated as 111(strong enhancement [decrease of 50 to 60% of survival cells compared to the control]), 11 (moderate enhancement [decrease of 20 to 40%]), and 2 (noenhancement). (B) Expression of the mutant hbc proteins in Ba/F3 cells. Stable Ba/F3 subclones infected with vector pBabeHygro (lane 1) or with hbc (lane 2), b772(lane 3), b590 (lane 4), b512 (lane 5), b470 (lane 6), b590(D473-505) (lane 7), and b560(D473-505) (lane 8) were lysed and subjected to Western blot analysis usinga polyclonal antibody specific to the extracellular domain of the hbc protein. The predicted position of each mutant hbc in the blot is indicated by arrows. (C) Survivalof cells expressing various hbc mutants during the course of cytokine deprivation. Viability of cells expressing various hbc mutants at various time points were measuredas described in legend to Fig. 1A. Each number is the average of two experiments of two to three independent subclones of each mutation. (D) Alignment of DERsequences of different species of bc. The amino acid sequence from 506 to 560 of human bc was compared to equivalent regions from mbIL-3, mbc, and rat bc. Identicalamino acids are shown as white letters; the conserved box II sequence is indicated by a bracket.
VOL. 19, 1999 CYTOKINE RECEPTOR COMMON b CHAIN MODULATES APOPTOSIS 7405
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.
were removed from culture media. However, cell viabilitycould be maintained in these cell lines by the addition ofcycloheximide (56), suggesting that new protein synthesis isessential and a positive control mechanism is hypothesized toCWIA. In this report, we provide evidence that the bc mole-cule is involved in modulating CWIA. We further demonstratethat the bc molecule modulates apoptosis with the followingdistinct features. First, its expression levels closely correlated
with the rate of apoptosis when cytokines were depleted (Fig.1 to 3). Second, the sequence essential for death accelerationwas distinct from that required for proliferation (Fig. 4). Third,the death accelerated by bc was sensitive to cycloheximidetreatment (data not shown). Fourth, its enhancement of celldeath did not depend on membrane anchorage (Fig. 6 and 7;Table 1). Fifth, physical aggregation of bc did not abrogate itsdeath-enhancing effect (Fig. 5 and 7). Finally, bc itself was
FIG. 5. Physical aggregation did not abrogate the function of DER. (A) Surface expression of the chimeric CD16-DER molecules in Ba/F3 cells. The stable celllines were treated with anti-CD16 antibody (Ab) and analyzed by flow cytometry as described for Fig. 1. CD/S, pCD16/7/Stop-expressing cell line; CD/b, pCD16/7/b471-590-expressing cell line. (B) Protein expression of the CD16 chimeric genes. Cell lysates were prepared from control cells (lane 1), CD/S cells (lane 2), and CD/bcells (lane 3) and were subjected to Western blot analysis with the anti-CD16 antibody. Sizes are indicated in kilodaltons. (C) Cross-linking of the DER sequence ofhbc by anti-CD16 antibody. Ba/F3 subclones as described for panel A were cross-linked with (1) or without (2) anti-CD16 antibody. The total cell lysates wereprepared and analyzed for the protein tyrosine phosphorylation by Western blot analysis using antiphosphotyrosine (anti-p-Tyr) antibody. The STAT5 protein levelsare shown as loading controls for each lane. (D) The survival rate of cells expressing various CD16/7 chimera with or without antibody cross-linking (CL). Cells asindicated were treated as described for panel B and were depleted of cytokine. Viable cell numbers were determined at various time points after cytokine depletion;the values are averages of two independent experiments performed in duplication. V, vector-expressing cells.
FIG. 6. The DER functions via an anchorage-independent mechanism. (A) Expression of the cytoplasmic hexahistidine-tagged DER in Ba/F3 cells. In the upperportion, the plasmid pBabeHisbDER is shown as an oblong. The hatched and open bars represent the histidine epitope tag from pcDNA3.1 (E box) and the hbccytoplasmic domain (aa 471 to 590; containing the DER sequence), respectively. The pair of arrows and solid bar indicate the oligonucleotide primers used for RT-PCRand the 500-bp PCR product, respectively. Cytoplasmic RNA prepared from pBabeHisbDER-expressing Ba/F3 cells (clones 1 to 3) and from vector-expressing cells(clone V) were subjected to RT-PCR. Reactions performed without RT served as controls for genomic DNA contamination. A plasmid DNA control (lane Ctr1) wasincluded to indicate the position of the PCR product (arrow). M, 1-kb DNA marker. (B) Expression of cytoplasmic histidine-tagged DER protein. The cytosolic andnuclear (N) extracts were prepared as described by Dignam et al. (10). The histidine tag-containing proteins were then enriched by passing 2 mg of extracts throughNi-nitrilotriacetic acid beads and were analyzed as for conventional Western blot analysis except that the proteins were probed with horseradish peroxidase-labelednickel. The signals were visualized with the ECL system. (C) Moderate acceleration of CWIA by cytosolic DER sequence. Cells expressing indicated plasmids weresubjected to cytokine deprivation and viable cell measurement as described for Fig. 1A. A cell line expressing wild-type hbc (bc) was included as a positive control.The pBabe vector-containing cell line (V) was included as a negative control. Sizes are indicated in kilodaltons.
7406 LEE ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.
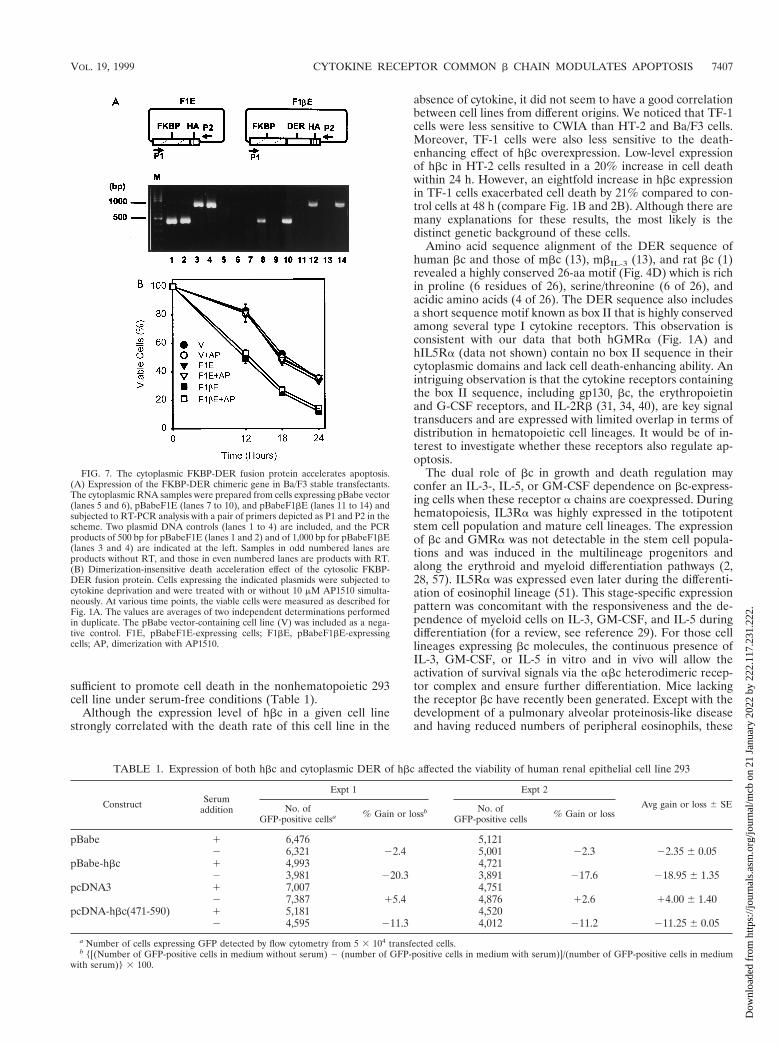
sufficient to promote cell death in the nonhematopoietic 293cell line under serum-free conditions (Table 1).
Although the expression level of hbc in a given cell linestrongly correlated with the death rate of this cell line in the
absence of cytokine, it did not seem to have a good correlationbetween cell lines from different origins. We noticed that TF-1cells were less sensitive to CWIA than HT-2 and Ba/F3 cells.Moreover, TF-1 cells were also less sensitive to the death-enhancing effect of hbc overexpression. Low-level expressionof hbc in HT-2 cells resulted in a 20% increase in cell deathwithin 24 h. However, an eightfold increase in hbc expressionin TF-1 cells exacerbated cell death by 21% compared to con-trol cells at 48 h (compare Fig. 1B and 2B). Although there aremany explanations for these results, the most likely is thedistinct genetic background of these cells.
Amino acid sequence alignment of the DER sequence ofhuman bc and those of mbc (13), mbIL-3 (13), and rat bc (1)revealed a highly conserved 26-aa motif (Fig. 4D) which is richin proline (6 residues of 26), serine/threonine (6 of 26), andacidic amino acids (4 of 26). The DER sequence also includesa short sequence motif known as box II that is highly conservedamong several type I cytokine receptors. This observation isconsistent with our data that both hGMRa (Fig. 1A) andhIL5Ra (data not shown) contain no box II sequence in theircytoplasmic domains and lack cell death-enhancing ability. Anintriguing observation is that the cytokine receptors containingthe box II sequence, including gp130, bc, the erythropoietinand G-CSF receptors, and IL-2Rb (31, 34, 40), are key signaltransducers and are expressed with limited overlap in terms ofdistribution in hematopoietic cell lineages. It would be of in-terest to investigate whether these receptors also regulate ap-optosis.
The dual role of bc in growth and death regulation mayconfer an IL-3-, IL-5, or GM-CSF dependence on bc-express-ing cells when these receptor a chains are coexpressed. Duringhematopoiesis, IL3Ra was highly expressed in the totipotentstem cell population and mature cell lineages. The expressionof bc and GMRa was not detectable in the stem cell popula-tions and was induced in the multilineage progenitors andalong the erythroid and myeloid differentiation pathways (2,28, 57). IL5Ra was expressed even later during the differenti-ation of eosinophil lineage (51). This stage-specific expressionpattern was concomitant with the responsiveness and the de-pendence of myeloid cells on IL-3, GM-CSF, and IL-5 duringdifferentiation (for a review, see reference 29). For those celllineages expressing bc molecules, the continuous presence ofIL-3, GM-CSF, or IL-5 in vitro and in vivo will allow theactivation of survival signals via the abc heterodimeric recep-tor complex and ensure further differentiation. Mice lackingthe receptor bc have recently been generated. Except with thedevelopment of a pulmonary alveolar proteinosis-like diseaseand having reduced numbers of peripheral eosinophils, these
FIG. 7. The cytoplasmic FKBP-DER fusion protein accelerates apoptosis.(A) Expression of the FKBP-DER chimeric gene in Ba/F3 stable transfectants.The cytoplasmic RNA samples were prepared from cells expressing pBabe vector(lanes 5 and 6), pBabeF1E (lanes 7 to 10), and pBabeF1bE (lanes 11 to 14) andsubjected to RT-PCR analysis with a pair of primers depicted as P1 and P2 in thescheme. Two plasmid DNA controls (lanes 1 to 4) are included, and the PCRproducts of 500 bp for pBabeF1E (lanes 1 and 2) and of 1,000 bp for pBabeF1bE(lanes 3 and 4) are indicated at the left. Samples in odd numbered lanes areproducts without RT, and those in even numbered lanes are products with RT.(B) Dimerization-insensitive death acceleration effect of the cytosolic FKBP-DER fusion protein. Cells expressing the indicated plasmids were subjected tocytokine deprivation and were treated with or without 10 mM AP1510 simulta-neously. At various time points, the viable cells were measured as described forFig. 1A. The values are averages of two independent determinations performedin duplicate. The pBabe vector-containing cell line (V) was included as a nega-tive control. F1E, pBabeF1E-expressing cells; F1bE, pBabeF1bE-expressingcells; AP, dimerization with AP1510.
TABLE 1. Expression of both hbc and cytoplasmic DER of hbc affected the viability of human renal epithelial cell line 293
Construct Serumaddition
Expt 1 Expt 2
Avg gain or loss 6 SENo. ofGFP-positive cellsa % Gain or lossb No. of
GFP-positive cells % Gain or loss
pBabe 1 6,476 5,1212 6,321 22.4 5,001 22.3 22.35 6 0.05
pBabe-hbc 1 4,993 4,7212 3,981 220.3 3,891 217.6 218.95 6 1.35
pcDNA3 1 7,007 4,7512 7,387 15.4 4,876 12.6 14.00 6 1.40
pcDNA-hbc(471-590) 1 5,181 4,5202 4,595 211.3 4,012 211.2 211.25 6 0.05
a Number of cells expressing GFP detected by flow cytometry from 5 3 104 transfected cells.b {[(Number of GFP-positive cells in medium without serum) 2 (number of GFP-positive cells in medium with serum)]/(number of GFP-positive cells in medium
with serum)} 3 100.
VOL. 19, 1999 CYTOKINE RECEPTOR COMMON b CHAIN MODULATES APOPTOSIS 7407
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.

mbc-null mice seem to have normal hematopoiesis (36, 37, 50).The expression of another highly homologous protein, mbIL-3,in these mbc-null mice was suggested to be responsible fortransducing appropriate proliferation and differentiation sig-nals during hematopoiesis. Similarly, since the DER sequenceof mbIL-3 is almost identical to that of mbc, it is possible thatin these mbc-null mice, mbIL-3 can substitute for mbc to reg-ulate apoptosis in a subset of hematopoietic cells. If this sce-nario is the case and given the fact that the receptor bc levelsaffect the death rate of a given cell type, it would then beinteresting to examine whether the expression level of mbIL-3in these mbc-null mice is elevated, a compensatory effect thatis frequently observed in mice lacking one member of an im-portant gene family (18, 26).
The functional and structural properties of DER are distinctfrom those of the death domains identified in the tumor ne-crosis factor receptor family and their adaptor signaling mol-ecules (53). These death domains can override the survivaleffects of growth factors and drive the target cell to undergoapoptosis. Functionally, DER is more closely related to theaddiction/dependence domains (ADD) in the nerve growthfactor receptor (NGFR) (44) and the androgen receptor (AR)(3) wherein the expression of these receptors establishes aligand-dependent cellular status. The ADD-containing recep-tors confer host cells with proliferation activity in the presenceof ligands and will drive cells to undergo apoptosis only whenligands are removed from the culture medium. However, thesethree functionally related peptide motifs are structurally verydistinct. DER of the receptor bc is a proline-rich peptide,ADD of NGFR is an a-helical peptide containing two criticalbasic residues (17), whereas ADD of AR is a stretch of glu-tamines that results from the trinucleotide repeats in the ARcoding sequence (3). It remains to be determined how thesestructurally distinct motifs can manifest similar biological func-tions.
Although our results strongly suggest that the receptor bc isinvolved in modulating growth factor withdrawal-induced ap-optosis, the underlying mechanism is still not clear. On thebasis of current knowledge on the signaling molecules involvedin growth and death control, we propose the following threelikely explanations. (i) Forced expression of bc might result insequestering some common signaling components which inter-act with the box II sequence, and consequently these cells diefaster upon deprivation of their dependent growth factors. (ii)The CIS/SOCS/SSI/Jab gene family could be induced by thereceptor bc through the JAK/STAT pathway (11, 27, 35, 52,60). Induction of these negative regulators may lead to prema-ture termination of the residual survival signal of the receptorafter cytokine removal, and the CWIA rate is subsequentlyenhanced. (iii) In the absence of cytokines, the receptor bc cantrigger a death cascade that eventually kill cells. While thesethree mechanisms are distinct from each other, they are notnecessarily mutually exclusive in the context of bc function.More experiments are required to unravel this issue.
ACKNOWLEDGMENTS
We thank Young-Sun Lin, Yu-Chung Yang, and Atsushi Miyajimafor critical comments on the manuscript, David Baltimore for theBOSC23 packaging cell line, Hartmut Land for retroviral expressionvectors, and Ariad Pharmaceuticals for plasmid pCF1E and the syn-thetic dimerizing agent AP1510. We are also grateful to Derek W.Gilroy for preparation of the manuscript.
This work was supported in part by Academia Sinica, Taiwan(J.J.-Y.Y.) and the National Science Council of Taiwan (J.J.-Y.Y.).S.-F.L. was supported by a postdoctoral fellowship from AcademiaSinica, Taiwan. H.-M.H. and J.-R.C. were supported by the postdoc-toral fellowships of the National Science Council of Taiwan.
REFERENCES
1. Appel, K., M. Buttini, A. Sauter, and P. J. Gebicke-Haerter. 1995. Cloning ofrat interleukin-3 receptor b-subunit from cultured microglia and its mRNAexpression in vivo. J. Neurosci. 15:5800–5809.
2. Ashihara, E., A. M. Vannucchi, G. Migliaccio, and A. R. Migliaccio. 1997.Growth factor receptor expression during in vitro differentiation of partiallypurified populations containing murine stem cells. J. Cell. Physiol. 171:343–356.
3. Bredesen, D., X. Ye, A. Tasinato, S. Sperandio, J. J. Wang, N. Assa-Munt,and S. Rabizadeh. 1998. p75NTR and the concept of cellular dependence:seeing how the other half die. Cell Death Differ. 5:365–371.
4. Chao, J. R., J. M. Wang, S. F. Lee, H. W. Peng, Y. H. Lin, C. H. Chou, J. C.Li, H. M. Huang, C. K. Chou, M. L. Kuo, J. J. Y. Yen, and H. F. Yang-Yen.1998. mcl-1 is an immediate-early gene activated by the granulocyte-mac-rophage colony-stimulating factor (GM-CSF) signaling pathway and is onecomponent of the GM-CSF viability response. Mol. Cell. Biol. 18:4883–4898.
5. Cleveland, J. L., M. Dean, N. Rosenberg, J. Y. Wang, and U. R. Rapp. 1989.Tyrosine kinase oncogenes abrogate interleukin-3 dependence of murinemyeloid cells through signaling pathways involving c-myc: conditional regu-lation of c-myc transcription by temperature-sensitive v-abl. Mol. Cell. Biol.9:5685–5695.
6. Cowling, G. J., and T. M. Dexter. 1994. Apoptosis in the haemopoieticsystem. Philos. Trans. R. Soc. Lond. Ser. B 345:257–263.
7. Datta, S. R., H. Dudek, X. Tao, S. Masters, H. Fu, Y. Gotoh, and M. E.Greenberg. 1997. Akt phosphorylation of BAD couples survival signals to thecell-intrinsic death machinery. Cell 91:231–241.
8. Devos, R., G. Plaetinck, J. Van-der-Heyden, S. Cornelis, J. Vandekerckhove,W. Fiers, and J. Tavernier. 1991. Molecular basis of a high affinity murineinterleukin-5 receptor. EMBO J. 10:2133–2137.
9. Dexter, T. M., A. D. Whetton, and C. M. Heyworth. 1986. The relevance ofprotein kinase C activation, glucose transport and ATP generation in theresponse of haemopoietic cells to growth factors, p. 163–169. In P. Kahn andT. Graf (ed.), Oncogenes and growth control. Springer-Verlag Press, Berlin,Germany.
10. Dignam, J., R. Lebovitz, and R. Roeder. 1983. Accurate transcription initi-ation by RNA polymerase II in a soluble extract from isolated mammaliannuclei. Nucleic Acids Res. 11:1475–1489.
11. Endo, T. A., M. Masuhara, M. Yokouchi, R. Suzuki, H. Sakamoto, K. Mitsui,A. Matsumoto, S. Tanimura, M. Ohtsubo, H. Misawa, T. Miyazaki, N.Leonor, T. Taniguchi, T. Fujita, Y. Kanajura, S. Komiya, and A. Yoshimura.1997. A new protein containing an SH2 domain that inhibits JAK kinases.Nature 387:921–924.
12. Gearing, D. P., J. A. King, N. M. Gough, and N. A. Nicola. 1989. Expressioncloning of a receptor for human granulocyte-macrophage colony-stimulatingfactor. EMBO J. 8:3667–3676.
13. Gorman, D. M., N. Itoh, T. Kitamura, J. Schreurs, S. Yonehara, I. Yahara,K. Arai, and A. Miyajima. 1990. Cloning and expression of a gene encodingan interleukin 3 receptor-like protein: identification of another member ofthe cytokine receptor gene family. Proc. Natl. Acad. Sci. USA 87:5459–5463.
14. Gossen, M., and H. Bujard. 1992. Tight control of gene expression in mam-malian cells by tetracycline responsive promoters. Proc. Natl. Acad. Sci. USA89:5547–5551.
15. Hara, T., and A. Miyajima. 1992. Two distinct functional high affinity recep-tors for mouse interleukin-3 (IL-3). EMBO J. 11:1875–1884.
16. Hayashida, K., T. Kitamura, D. M. Gorman, K. Arai, T. Yokota, and A.Miyajima. 1990. Molecular cloning of a second subunit of the receptor forhuman granulocyte-macrophage colony-stimulating factor (GM-CSF): re-constitution of a high-affinity GM-CSF receptor. Proc. Natl. Acad. Sci. USA87:9655–9659.
17. Hileman, M. R., B. S. Chapman, S. Rabizadeh, V. V. Krishnan, D. Bredesen,N. Assa-Munt, and L. A. Plesniak. 1997. A cytoplasmic peptide of theneurotrophin receptor p75NTR: induction of apoptosis and NMR deter-mined helical conformation. FEBS Lett. 415:145–154.
18. Hummler, E., T. J. Cole, J. A. Blendy, R. Ganss, A. Aguzzi, W. Schmid, F.Beermann, and G. Schutz. 1994. Targeted mutation of the CREB gene:compensation within the CREB/ATF family of transcription factors. Proc.Natl. Acad. Sci. USA 91:5647–5651.
19. Itoh, N., S. Yonehara, J. Schreurs, D. M. Gorman, K. Maruyama, A. Ishii, I.Yahara, K. Arai, and A. Miyajima. 1990. Cloning of an interleukin-3 recep-tor: a member of a distinct receptor gene family. Science 247:324–327.
20. Jacobson, M. D., M. Weil, and M. C. Raff. 1997. Programmed cell death inanimal development. Cell 88:347–354.
21. Kinoshita, T., M. Shirouzu, A. Kamiya, K. Hashimoto, S. Yokoyama, and A.Miyajima. 1997. Raf/MAPK and rapamycin-sensitive pathways mediate theanti-apoptotic function of p21Ras in IL-3-dependent hematopoietic cells.Oncogene 15:619–627.
22. Kinoshita, T., T. Yokota, K. Arai, and A. Miyajima. 1995. Regulation ofBcl-2 expression by oncogenic Ras protein in hematopoietic cells. Oncogene10:2207–2212.
23. Kinoshita, T., T. Yokota, K. Arai, and A. Miyajima. 1995. Suppression ofapoptosis death in hematopoietic cells by signalling through the IL-3/GM-CSF receptors. EMBO J. 14:266–275.
7408 LEE ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.

24. Kitamura, T., N. Sato, K. Arai, and A. Miyajima. 1991. Expression cloningof the human IL-3 receptor cDNA reveals a shared beta subunit for thehuman IL-3 and GM-CSF receptors. Cell 66:1165–1174.
25. Kitamura, T., T. Tange, T. Terasawa, S. Chiba, T. Kuwaki, K. Miyagawa,Y. F. Piao, K. Miyazono, A. Urabe, and F. Takaku. 1989. Establishment andcharacterization of a unique human cell line that proliferates dependently onGM-CSF, IL-3, or erythropoietin. J. Cell. Physiol. 140:323–334.
26. LeCouter, J. E., B. Kablar, W. R. Hardy, C. Ying, L. A. Megeney, L. L. May,and M. A. Rudnicki. 1998. Strain-dependent myeloid hyperplasia, growthdeficiency, and accelerated cell cycle in mice lacking the Rb-related p107gene. Mol. Cell. Biol. 18:7455–7465.
27. Matsumoto, A., M. Masuhara, K. Mitsui, M. Yokouchi, M. Ohtsubo, H.Misawa, A. Miyajima, and A. Yoshimura. 1997. CIS, a cytokine inducibleSH2 protein, is a target of the JAK-STAT5 pathway and modulates STAT5activation. Blood 89:3148–3154.
28. McKinstry, W. J., C. L. Li, J. E. Rasko, N. A. Nicola, G. R. Johnson, and D.Metcalf. 1997. Cytokine receptor expression on hematopoietic stem andprogenitor cells. Blood 89:65–71.
29. Metcalf, D. 1993. Hematopoietic regulators: redundancy or subtlety? Blood82:3515–3523.
30. Minami, Y., Y. Nakagawa, A. Kawahara, T. Miyazaki, K. Sada, H.Yamamura, and T. Taniguchi. 1995. Protein tyrosine kinase Syk is associatedwith and activated by the IL-2 receptor: possible link with the c-myc induc-tion pathway. Immunity 2:89–100.
31. Miura, O., J. L. Cleveland, and J. N. Ihle. 1993. Inactivation of erythropoi-etin receptor function by point mutations in a region having homology withother cytokine receptors. Mol. Cell. Biol. 13:1788–1795.
32. Morgensten, J. P., and H. Land. 1990. Advanced mammalian gene transfer:high titer retroviral vectors with multiple drug selection markers and acomplementary helper-free packaging cell line. Nucleic Acids Res. 18:3587–3596.
33. Mosmann, T. 1983. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J. Immunol. Methods65:55–63.
34. Murakami, M., M. Narazaki, M. Hibi, H. Yawata, K. Yasukawa, M.Hamaguchi, T. Taga, and T. Kishimoto. 1991. Critical cytoplasmic region ofthe interleukin 6 signal transducer gp130 is conserved in the cytokine recep-tor family. Proc. Natl. Acad. Sci. USA 88:11349–11353.
35. Naka, T., M. Narazaki, M. Hirata, T. Matsumoto, S. Minamoto, A. Aono, N.Nishimoto, T. Kajita, T. Taga, K. Yoshizaki, S. Akira, and T. Kishimoto.1997. Structure and function of a new STAT-induced STAT inhibitor. Na-ture 387:924–929.
36. Nishinakamura, R., N. Nakayama, Y. Hirabayashi, T. Inoue, D. Aud, T.McNeil, S. Azuma, S. Yoshida, Y. Toyoda, K. Arai, A. Miyajima, and R.Murray. 1995. Mice deficient for the IL-3/GM-CSF/IL-5 bc receptor exhibitlung pathology and impaired immune response, while bIL-3 receptor-defi-cient mice are normal. Immunity 2:211–222.
37. Nishinakamura, R., A. Miyajima, P. J. Mee, V. L. J. Tybulewixz, and R.Murray. 1996. Hematopoiesis in mice lacking the entire granulocyte-mac-rophage colony-stimulating factor/interleukin-5 functions. Blood 88:2458–2464.
38. Ogata, N., T. Kouro, A. Yamada, M. Koike, N. Hanai, T. Ishikawa, and K.Takatsu. 1998. JAK2 and JAK1 constitutively associate with an interleukin-5(IL-5) receptor a and bc subunit, respectively, and are activated upon IL-5stimulation. Blood 91:2264–2271.
39. Ohta, T., T. Kinoshita, M. Naito, T. Nozaki, M. Masutani, T. Tsuruo, and A.Miyajima. 1997. Requirement of the caspase-3/CPP32 protease cascade forapoptotic death following cytokine deprivation in hematopoietic cells.J. Biol. Chem. 272:23111–23116.
40. O’Neal, K. D., and L. Y. Yu-Lee. 1993. The proline-rich motif (PRM): a novelfeature of the cytokine/hematopoietin receptor superfamily. LymphokineCytotokine Res. 12:309–312.
41. Palacios, R., and M. Steinmetz. 1985. IL-3-dependent mouse clones thatexpress B-220 surface antigen, contain Ig genes in germ-line configuration,and generate B lymphocytes in vivo. Cell 41:727–734.
42. Pear, W. S., G. P. Nolan, M. L. Scott, and D. Baltimore. 1993. Production of
high-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad.Sci. USA 88:8392–8396.
43. Quelle, F. W., N. Sato, B. A. Witthuhn, R. C. Inhorn, M. Eder, A. Miyajima,J. D. Griffin, and J. N. Ihle. 1994. JAK2 associates with the bc chain of thereceptor for granulocyte-macrophage colony-stimulating factor, and its acti-vation requires the membrane-proximal region. Mol. Cell. Biol. 14:4335–4341.
44. Rabizadeh, S., J. Oh, L. T. Zhong, J. Yang, C. M. Bitler, L. L. Butcher, andD. E. Bredesen. 1993. Induction of Apoptosis by the low-affinity NGF re-ceptor. Science 261:345–348.
45. Rao, P., and R. A. Mufson. 1995. A membrane proximal domain of thehuman interleukin-3 receptor bc subunit that signals DNA synthesis in NIH3T3 cells specifically binds a complex of Src and Janus family tyrosine kinasesand phosphatidylinositol 3-kinase. J. Biol. Chem. 270:6886–6893.
46. Sakamaki, K., I. Miyajima, T. Kitamura, and A. Miyajima. 1992. Criticalcytoplasmic domains of the common b subunit of the human GM-CSF, IL-3and IL-5 receptors for growth signal transduction and tyrosine phosphory-lation. EMBO J. 11:3541–3549.
47. Sato, N., K. Sakamaki, N. Terada, K. Arai, and A. Miyajima. 1993. Signaltransduction by the high-affinity GM-CSF receptor: two distinct cytoplasmicregions of the common b subunit responsible for different signaling. EMBOJ. 12:4181–4189.
48. Sato, S., T. Katagiri, S. Takaki, Y. Kikuchi, Y. Hitoshi, S. Yonehara, S.Tsukada, D. Kitamura, T. Watanabe, O. Witte, and K. Takatsu. 1994. IL-5receptor-mediated tyrosine phosphorylation of SH2/SH3-containing pro-teins and activation of Bruton’s tyrosine and Janus 2 kinases. J. Exp. Med.180:2101–2111.
49. Scheid, M. P., and V. Duronio. 1998. Dissociation of cytokine-induced phos-phorylation of Bad and activation of PKB/akt: involvement of MEK up-stream of Bad phosphorylation. Proc. Natl. Acad. Sci. USA 95:7439–7444.
50. Scott, C. L., D. A. Hughes, D. Cary, N. A. Nicola, C. G. Begley, and L. Robb.1998. Functional analysis of mature hematopoietic cells from mice lackingthe bc chain of the granulocyte-macrophage colony-stimulating factor recep-tor. Blood 92:4119–4127.
51. Shalit, M., S. Sekhsaria, and H. L. Malech. 1995. Modulation of growth anddifferentiation of eosinophils from human peripheral blood CD341 cells byIL5 and other growth factors. Cell Immunol. 160:50–57.
52. Starr, R., T. A. Willson, E. M. Viney, L. J. L. Murray, J. R. Rayner, B. J.Jenkins, T. J. Gonda, W. S. Alexander, D. Metcalf, N. A. Nicola, and D. J.Hilton. 1997. A family of cytokine-inducible inhibitors of signalling. Nature387:917–921.
53. Tartaglia, L. A., T. M. Ayres, G. H. W. Wong, and D. V. Goeddel. 1993. Anovel domain within the 55 kd TNF receptor signals cell death. Cell 74:845–853.
54. Tavernier, J., R. Devos, S. Cornelis, T. Tuypens, J. Van-der-Heyden, W.Fiers, and G. Plaetinck. 1991. A human high affinity interleukin-5 receptor(IL5R) is composed of an IL5-specific a chain and a b chain shared with thereceptor for GM-CSF. Cell 66:1175–1184.
55. Watson, J. 1979. Continuous proliferation of murine antigen-specific helperT lymphocytes in culture. J. Exp. Med. 150:1510–1519.
56. Williams, G. T., C. A. Smith, E. Spooncer, T. M. Dexter, and D. R. Taylor.1990. Haemopoietic colony stimulating factors promote cell survival by sup-pressing apoptosis. Nature. 343:76–79.
57. Wognum, A. W., M. O. de-Jong, and G. Wagemaker. 1996. Differentialexpression of receptors for hemopoietic growth factors on subsets of CD341hemopoietic cells. Leuk. Lymphoma 24:11–25.
58. Wyllie, A. H. 1980. Glucocorticoid-induced thymocyte apoptosis is associatedwith endogenous endonuclease activation. Nature 284:555–556.
59. Yen, J. J. Y., Y. C. Hsieh, C. L. Yen, C. C. Chang, S. Lin, and H. F. Yang-Yen.1995. Restoring the apoptosis suppression response to IL-5 confers on eryth-roleukemic cells a phenotype of IL-5-dependent growth. J. Immunol. 154:2144–2152.
60. Yoshimura, A., T. Ohkubo, T. Kigushi, N. A. Jenkins, D. J. Gilbert, N. G.Copeland, T. Hara, and A. Miyajima. 1995. A novel cytokine-inducible geneCIS encodes an SH2-containing protein that binds to tyrosine-phosphory-lated interleukin 3 and erythropoietin receptors. EMBO J. 14:2816–2826.
VOL. 19, 1999 CYTOKINE RECEPTOR COMMON b CHAIN MODULATES APOPTOSIS 7409
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 21
Jan
uary
202
2 by
222
.117
.231
.222
.


















