Cloning of the cDNA for Canine Interferon-γ
4
Click here to load reader
Transcript of Cloning of the cDNA for Canine Interferon-γ

JOURNAL OF INTERFERON RESEARCH 12:191-194 (1992)Mary Ann Liebert, Inc., Publishers SHORT COMMUNICATION
Cloning of the cDNA for Canine Interferon-7KEITH ZUCKER, PING LU, VIOLET ESQUENAZI, and JOSHUA MILLER
ABSTRACT
We have taken advantage of conserved regions of cDNA sequences for interferon-7 (IFN-7) from other speciesto design polymerase chain reaction (PCR) primers capable of amplifying the protein-coding region of thecanine mRNA. We report here the cDNA cloning of this region from dog lymphocytes and the cDNA sequence.The predicted amino acid sequence is also reported and compared to the known sequences of these otherspecies. The molecular clone for the canine IFN-7 will allow direct study of this important cytokine's role ingraft rejection in the widely used experimental canine transplant model.
THE CELLULAR CYTOKINE interferon-7 (IFN-7) is known tobe a key regulator of cell-mediated immune responses.'"
IFN-7 production increases as a result of antigen- or mitogen-induced T-cell activation, and this, in turn, effects the produc-tion of other proteins, including class II histocompatibility anti-gens, other cytokines, and even IFN-7 itself.'2-4' Evidencefrom kidney transplant studies suggests that IFN-7 may also bea key molecule in the rejection process, with at least one actionbeing to up-regulate class II-MHC proteins in donor-derivedkidney cells.'571 Despite the apparent importance of IFN-7 inthese processes, studies on the regulation of this cytokine at themolecular level still lag behind those of other cytokines such as
interleukin-2 (IL-2).Transplantations of vascularized organs and bone marrow in
dogs are widely used preclinical experimental models. In spiteof this, very little is known about canine cytokines or theirrespective genes. The present brief communication reports theuse of polymerase chain reaction (PCR) methods for the cloningof the protein-coding region of the canine IFN-7 cDNA withprimers designed from conserved regions described in otherspecies, with the cDNA and predicted amino acid sequencescompared to that of other known IFN-7 molecules. The avail-ability of a clone for the canine IFN-7 cDNA will allow formore detailed studies on the regulation of this important cytok-ine in dogs at the molecular level. It will also help to elucidatethe role of IFN-7 in the canine transplant model.
Sense and anti-sense degenerate PCR primers were designedto regions +52 to +71 (with the + 1 position corresponding tothe initiation codon of the signal peptide) and +507 to +531 ofthe human IFN-7 gene.'81 The sense primer was a 20-mer
consisting of the sequence 5-TCTCTTGGCTGTTACTGCCA-3'. The anti-sense primer was a 24-mer consisting of the se-
quence 5'-AAAATTCAAATATGTGCATGGCAGGAA-3'. To-tal RNA was isolated from beagle lymph node lymphocytesstimulated for 24 h with phorbol myristate acetate(PMA) + A23187'7' using the method of Chirgwin et a/.'9)cDNA was synthesized from 1.0 p.g of RNA using a RNA-PCRkit (Cetus Corp., Emeryville, CA) in a final volume of 20 u.1,according to the manufacturer's instructions using randomprimers. The amplification reaction contained 0.3 p.M of eachsense and antisense primer with the remaining components ofthe PCR reaction. Amplification conditions consisted of thefirst five cycles at 97°C for 2 min, 37°C for 2 min, 72°C for 3min, followed by 35 cycles at 94°C for 2 min, 55°C for 2 min,72°C for 3 min. The entire PCR reaction was then loaded onto a
1.0% low-melting-temperature agarose gel and separated for 2h at 60 volts. A specific 480-bp DNA band was excised fromthe gel after visualization with ethidium bromide and the agar-ose was melted at 65°C. Fifteen microliters of this DNA suspen-sion was added to a second PCR reaction identical to the firstand an additional 35 cycles of 94°C for 2 min, 55°C for 2 min,72°C for 3 min was performed. The entire PCR product was
again separated on a 1% regular agarose gel and the specific480-bp band was excised. The DNA was eluted out of the gelusing an IBI electroelution apparatus and precipitated withEtOH overnight at
—
70°C. The resuspended DNA was quanti-tated and 75 ng was added to 25 ng of the plasmid pCR2000(Invitrogen, San Diego, CA). Ligation of the vector and PCRproduct was performed according to the manufacturer's instruc-tions. Several transformed bacterial colonies were grown up
Division of Transplantation. Department of Surgery. Miami Veterans Administration Medical Center and the University of Miami School ofMedicine, Miami, FL 33101.
191

192 ZUCKER ET AL.
and the plasmids were recovered.'"" Presence of the IFN-7insert was confirmed using PCR. Inserts from several individ-ual clones were exised with Hind III and Eco RI restrictionenzymes and ligated directly into the same sites of the sequenc-ing vector M13mpl8. Sequencing was done on the isolatedsingle-stranded phage DNA by the method of Sanger et al.al)Several individual recombinant clones containing inserts inboth orientation were sequenced using the M13 universalprimer so that the complete sequence of both strands of DNAcould be ascertained.
In addition to the full-length 480-bp band, many other lower-molecular-weight bands were also seen on the agarose-gel (datanot shown). After excision and cloning of the full-length PCRproduct, inserts from several isolated recombinant clones were
sequenced. The resulting cDNA sequence is shown in Fig. 1.The sequence of the PCR primers is underlined. Using theknown reading frame of other IFN-7 genes, the predicted aminoacid sequence was deduced and is also shown in Fig. 1. Thecanine cDNA sequence shows a sequence homology of 83%,83%, 81%, 78%, 66%, and 68% to the known sequences ofbovine, ovine, porcine, human, rat, and mouse cDNAs, respec-tively. To insure that the sequence reported here for the caninecDNA is not the result of alteration during the PCR amplifica-tion by the relatively low fidelity of the Taq polymerase, ampli-fications were also performed from the cDNA samples using
Pfu polymerase (Stratagene, La Jolla, CA). This enzyme pos-sesses a 3' to 5' exonuclease activity resulting in a 12-foldincrease in fidelity of DNA synthesis over Taq polymerase. Thesequences for all clones obtained using the Pfu enzyme were
identical to those of the clones obtained using the Taq poly-merase.
Comparison of the predicted amino acid sequence of themature canine IFN-7 protein to that of other known species isshown in Fig. 2. The first six amino acids of the canine se-
quence could not be determined accurately, because they corre-
spond to the PCR primer sequence. Highly conserved aminoacids are indicated on the figure as well as the percent homol-ogy of each sequence to that of the human IFN-7. The predictedcanine protein shows an 80%, 80%, 73%, 66%, 45%, and 46%sequence homology to bovine, ovine, porcine, human, rat, andmouse proteins, respectively.
High sequence conservation at the cDNA and protein levelsconfirm that this is indeed an authentic clone. Figure 2 showsthat every highly conserved amino acid from mouse to human isalso conserved in the canine sequence. As would be expected,the predicted dog protein is most closely related to the bovineand ovine sequences. Interestingly, the canine amino acid se-
quence is slightly closer in homology to the human IFN-7 thanis the bovine sequence. Several amino acid positions can beseen in which there has been a change between the bovine and
1 10TCT CTT GGC TGT TAC TGC CAG GCC ATG TTT TTT AAA GAA ATA GAA AACSLGCYCQAMFFKEIEN
48
20CTA AAG GAA TAT TTT AAT GCA AGT AAT CCA GAT GTA TCG GAC GGT GGGLKEYFNASNPDVSDGG
96
30 40TCT CTT TTC GTA GAT ATT TTG AAG AAA TGG AGA GAG GAG AGT GAC AAASLFVDILKKWREESDK
144
50 60ACA ATC ATT CAG AGC CAA ATT GTC TCT TTC TAC TTG AAA CTG TTT GAC 192TIIQSQIVSFYLKLFD
70AAC TTT AAA GAT AAC CAG ATC ATT CAA AGG AGC ATG GAT ACC ATC AAG 240NFKDNQI IQRMSDT IK
80 90GAA GAC ATG CTT GGC AAG TTC TTA AAT AGC AGC ACC AGT AAG AGG GAG 288E'DMLGKFLNSSTSKRE
100GAC TTC CTT AAG CTG ATT CAA ATT CCT GTG AAC GAT CTG CAG GTC CAGDFLKLIQIPVNDLQVQ
336
110 120CGC AAG GCG ATA AAT GAA CTC ATC AAA GTG ATG AAT GAT CTC TCA CCARKAINELIKVMNDLSP
384
130 140AGA TCC AAC CTA AGG AAG CGG AAA AGG AGT CAG AAT CTG TTT CGA GGCRSNLRKRKRSQNLFRG
432
CGC AGA GCA TCG AAA TAA TGG TCA TCC TGC CAG CAC TAT TTG AAT TTT 480R R A S K
FIG. 1. cDNA and predicted amino acid sequence of canine IFN-7. The sequences of the PCR primers are underlined. Numbersoverhead refer to the amino acid positions of the protein after signal peptide cleavage.

CLONING OF THE cDNA FOR CANINE IFN-7 193
HumanCanineOvineBovinePorcineHurineRat
SI Sll 1 10 20 30MKY1SYILAF QiCIVLGSLG CY.CQDPYVK EAENLKKY_EN AGHSDVADNG TLFLGJLKNÜ
--AMFF- -I-E--SNP--S-G- S--VD-K-I-E--SNP-KG- P--SE-I-E--SSP-KG- P--SD-ITI--D- -STS--PNG- P-E-
SSGI--.EEK S-D-WR---SA-RRV-VL -LMAL-.- --GTLIE SL-S--N- SSSM-AMEGK S-L-D-WR--
.-SF--F--S--T-F--NA-HC--
L L--VL--FS- L L--GL--FS-•--VT-CFS- L --FLMAV-.-
S-G-G-FF-S-G-GQFFRS-APFF--HGTVIE SL-S-NN---GTLIE SL-S--N--
HumanCanineOvineBovinePorcineHurineRat
40 50 60 70 80 90KEESDRK1MQ SOIVSFYFKL FKNFKDDOSI QKSVETIKED MNVKEFNSNK K£RDD£EKLTR-KT-I--K--I--D-K--I--K--I-QKDG-M--L- -1-LR-
-D----E-L--E-L--EI---EVL-
-N-I--N-V--N-V--N-A--N-A-
-R-MD---R-MDI--R-MDI--R-MDV-SNNISV-
-Q--Q--Q-
-LG---FQ---FQ---FQR-
-STGSSGSSGSS
ESH LITT--SNS-
S--E-E-LE-E-LE-G-LN---A-K-A-MSIA
-L--I-KR-I-K--I
QKDGNT--LE -I-LR- -EVL--N-A- SNNISV-ESH LITN--SNS- A-K-A-MSIA
HumanCanineOvineBovinePorcineMurineRat
100 110 120 130 140NYSYTDLNVQ RKAIHEilQy. MAELS£AAKT GKRKRSOMLF RGRRASOQIP-N--Q--QIP-D--QI-QIP-D--QI-KIP-DN-QI-KFE-NNPQ--KFE-NNPQI-
-N--N--N--S--Q-FN-H--VN-
-K--K--K--K--R--R-
-ND----ND----ND----ND—
RSNL R-KSNL R-KSNL R-RSNL R-
VHQ-L-ESSLIHQ-ESSL
--N----N----N----TM--RC-RC
-K-M-MQ-Q---K
% match toHuman
100656262594239
Reference8
1714181516
FIG. 2. Comparison of the aminoacid sequences of known mammalian IFNs-7. Amino acids S1-S20 refer to the putative signalpeptide. Highly conserved amino acids are underlined. The dashed lines indicate amino acids that are identical to those of thehuman sequence.
canine sequences that has been conserved between human andcanine proteins. Unfortunately, as mentioned, the cDNA se-
quence encoding the signal peptide and first three amino acidsof the mature protein could not be obtained because of theposition of the PCR sense primer. Efforts to obtain this se-
quence information using the rapid amplification of cDNA ends(RACE) technique"21 has not yet been successful.
The cDNA clone obtained here has been used also now as a
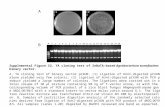
molecular probe in Northern blot studies on dog lymphocytesresponding to either mitogenic or allogenic stimulus in vitro.Figure 3 shows the results of a Northern blot study of IFN-7mRNA levels after in vitro PMA + A23187 induction of puri-fied dog lymph node lymphocytes. Details of the experimentare found in the figure legend. The results show a markedincrease in IFN-7 mRNA after mitogen stimulation, whichpeaked at 6 h. After this time, the mRNA level decreased. Theexpression of this gene shows the same pattern of upregulationthat is observed for other species,"1' adding support to theclaim that this is an authentic canine cDNA clone.
Lack of availability of canine-specific cytokines or their re-
spective molecular clones has made it difficult to study theirrole in transplantation using the canine model system. A purifi-cation protocol for the dog IFN-7 protein has only recently beendescribed.I7) Furthermore, lack of cross-species reactivity haslimited the use of commercially available native or recombinantIFN-7 proteins or the respective antisera to these proteins. Onlythe polyclonal anti-human IFN-7 has been observed to cross-
react with the canine IFN-7 protein.'71 Native or recombinanthuman, rat, or mouse IFN-7 proteins have not been observed towork on canine lymphocytes. The availability of this cDNAclone should allow for the production of a recombinant IFN-7protein that can be used in transplant studies. Efforts to engineer
Time(hr) 0 3 6 14 24
28S-
18S-
FIG. 3. Upregulation of IFN-7 mRNA in lymphocytes.Lymph node lymphocytes were isolated and stimulated in cul-ture for various times with mitogen as described.'7' Total RNAwas prepared as described in the text. Ten micrograms of totalRNA from each sample was separated on a 1.2% agarose gel for5 h at 60 volts and transferred to a hydrophobic membrane(Immobilon-N, Millipore Corp., Bedford MA) using a vac-uum-assisted transfer apparatus (Pharmacia-LKB, Piscataway,NJ). The membrane was sequentially probed with the canineIFN-7 cDNA clone and the full-length mouse ß-actin cDNAclone labeled with ",2P using the random primer labeling kit(Boehringer-Mannheim, Indianapolis, IN). Prehybridizationand hybridizations were performed using rapid hybridizationbuffer (Amersham, Arlington Heights, IL) according to themanufacturer's specifications. The final wash conditions wereat 65°C in 0.1 x SSC, 0.1% SDS for 20 min. Exposure toX-ray film (X-omat AR, Kodak, Rochester, NY) was for 16 hat —70°C using one intensifying screen. Positions of the 18Sand 28S RNA markers are indicated on the figure.

194
bacteria genetically to produce this protein are currently under-way, as are studies on the expression of this gene in dog trans-
plantation, to ascertain the role of this important cytokine ingraft rejection.
ACKNOWLEDGMENTS
This work was supported by the Miami Veteran's Adminis-tration Hospital Research Support, National Institute of Healthgrant no. ROI DK25243-11, and a grant from the South FloridaKidney Foundation.
REFERENCES
1. ARAL K., LEE, F., MIXAJIMA, A., MIXATAKE, S., ARAI,N., and YOKOTA, T. (1990). Cytokines: Coordinates of theimmune and inflammatory responses. Annu. Rev. Biochem. 59,783-836.
2. GAUCHAT, JG., GAUCHAT, D., QIU, G., MANDALLAZ,M., and STADLER, B.M. (1991). Detection of cytokine mRNAin polyclonally-, antigen-, or allergen-stimulated mononuclearcells. Immunol. Rev. 119, 147-161.
3. BENOIST, C, and MATHIS, D. (1990). Regulation of majorhistocompatibility complex class II genes. Annu. Rev. Immunol.8,681-715.
4. IJZERMANS, J.N.M. and MARQUET, R.L. (1989). Interferon-gamma: A review. Immunobiology 179, 456-473.
5. ROTH, D., FULLER, L., ESQUENAZI, V., KYRIAKIDES,G., PARDO, V., and MILLER, J. (1985). The biologic signifi-cance of the mixed lymphocyte kidney culture in humans. Trans-plantation 40, 376-383.
6. ESQUENAZI, V., FULLER, L., PARDO, V., ROTH, D., MIL-GROM, M., and MILLER, J. (1987). In vivo and in vitro induc-tion of class II molecules in canine renal cells and their effect on
the mixed lymphocyte kidney cell culture. Transplantation 44,680-692.
7. FULLER, L., FERNANDEZ, J., ZHENG. S., CARRENO, M.,ESQUENAZI, V., YANG, W-C, and MILLER, J. (1992). Im-muno and biochemical characterization of purified canine inter-feron-7. Production of a monoclonal antibody, affinity purifica-tion and its effect on MLC and MLKC reaction. Transplantation(in press).
8. GRAY, P.W. and GOEDDEL, D.V. (1982). Structure of thehuman immune interferon gene. Nature 298, 859-863.
ZUCKER ET AL.
9. CHIRGWIN, M.M., PRZYLOYLA, A.E., MACDONALD,R.J., and RUTTER, W.J. (1979). Isolation of biologically activeribonucleic acid from sources enriched in ribonuclease. Biochem-istry 18, 5294-5299.
10. AUSUBEL, FM.. BRENT, R., KINGSTON, R.E., MOORE,DP., SEIDMAN, J.G., SMITH, JA., and STRUHL, K. (eds.).(1989). Purification of Plasmid DNA, in: Short Protocols in Mo-lecular Biology. New York: John Wiley & Sons, 387 pp.
11. SANGER, F., NICKLEN, S., and COULSON, A.R. (1977).DNA sequencing with chain-terminating inhibitors. Proc. Nati.Acad. Sei. USA 74, 5463-5467.
12. FROHMAN, MA. (1990). Rapid Amplification of cDNA ends(RACE): User-friendly cDNA cloning. Amplifications 5, 11-15.
13. GRANELLI-PIPERNO, A.. ANDRUS, L., and STEINMAN,R.M. (1986). Lymphokine and non-lymphokine mRNA levels instimulated human T cells: Kinetics, mitogen requirements, andeffects of cyclosporin A. J. Exp. Med. 163, 922-937.
14. CERRETTI. D.P.. MCKEREGRAN, K., LARSEN, A., COS-MAN, D., GILLIS, S., and BAKER, P.E. (1986). Cloning, se-
quence, and expression of bovine ¡nterferon-gamma. J. Immunol.136,4561-4564.
15. GRAY, P.W.. and GOEDDEL, D.V. (1983). Cloning and ex-
pression of murine immune interferon cDNA. Proc. Nati. Acad.Sei. USA 80, 5842-5846.
16. DIJKEMA, R., VAN DER MEIDE, P.H., POUWELS, PH.,CASPERS, M.. DUBBELD, M., and SCHELLEKENS, H.(1985). Cloning and expression of the chromosomal immune in-terferon gene of the rat. EMBO J. 4, 761-767.
17. MCINNES. C.J., LOGAN, M., REDMOND. J.. ENTRICAN,G., and BAIRD, G.D. (1990). The molecular cloning of the ovinegamma-interferon cDNA using the polymerase chain reaction.Nucleic Acids Res. 18, 4012.
18. DIJKMANS, R., VANDENBROECK, K., BEUKEN, E., andBILLIAU, A. (1990). Sequence of the porcine ¡nterferon-gamma(IFN-7) gene. Nucleic Acids Res. 18, 4259.
Address reprint requests to:Dr. Keith Zucker
Department of Surgery—R310Division of Transplantation
University of Miami School of MedicineP.O. Box 016310
Miami, Florida 33101
Received 8 November 1991/Accepted 21 February 1992