
Bioquímica - nutrifaciplac.files.wordpress.com · provenientes das reações acima passam pela...
42
Bioquímica Profª. Ana Elisa Matias Fundamentos da Bioquímica Β-Oxidação
Transcript of Bioquímica - nutrifaciplac.files.wordpress.com · provenientes das reações acima passam pela...
Bioquímica
Profª. Ana Elisa Matias
Fundamentos da Bioquímica
Β-Oxidação

• Os lipídeos ingeridos são constituídos principalmente por:• triacilgliceróis (90% do total),
• glicerofosfolipídeos (em menor grau),
• colesterol,
• ésteres de colesteril e
• ácidos graxos livres.
• No trato gastrointestinal, os lipídeos são emulsificados, digeridos porenzimas hidrolíticas e absorvidos pelas células da mucosa intestinal.
• Em razão da pouca solubilidade em meio aquoso, os lipídeos seagregam em grandes complexos dificultando a hidrólise enzimática ea absorção intestinal.

• Esses obstáculos são contornados pelo emprego de agentesemulsificantes que aumentam a interface lipídio-água permitindo a açãodas enzimas intestinais hidrossolúveis, também como a “solubilização”dos produtos de hidrólise.
• Os lipídeos da dieta são emulsificados no duodeno pela ação detergente dos sais biliares.
• Os sais biliares são moléculas anfipáticas sintetizadas pelo fígado a partir do colesterol e temporariamente armazenados na vesícula biliar e liberados no intestino delgado após ingestão de gorduras.
• A emulsificação é possível pela natureza anfipática dos sais biliares.

• A porção polar das moléculas de sais biliares, interage com a água, enquanto o grupo não-polar interage com os lipídeos hidrofóbicos.
• Desse modo, os lipídeos são finamente dispersos no meio aquoso.
• Três enzimas hidrolíticas são encontradas no suco pancreático secretado no duodeno:
• lipase−pancreática,
• colesterol−esterase e
• fosfolipase A2.
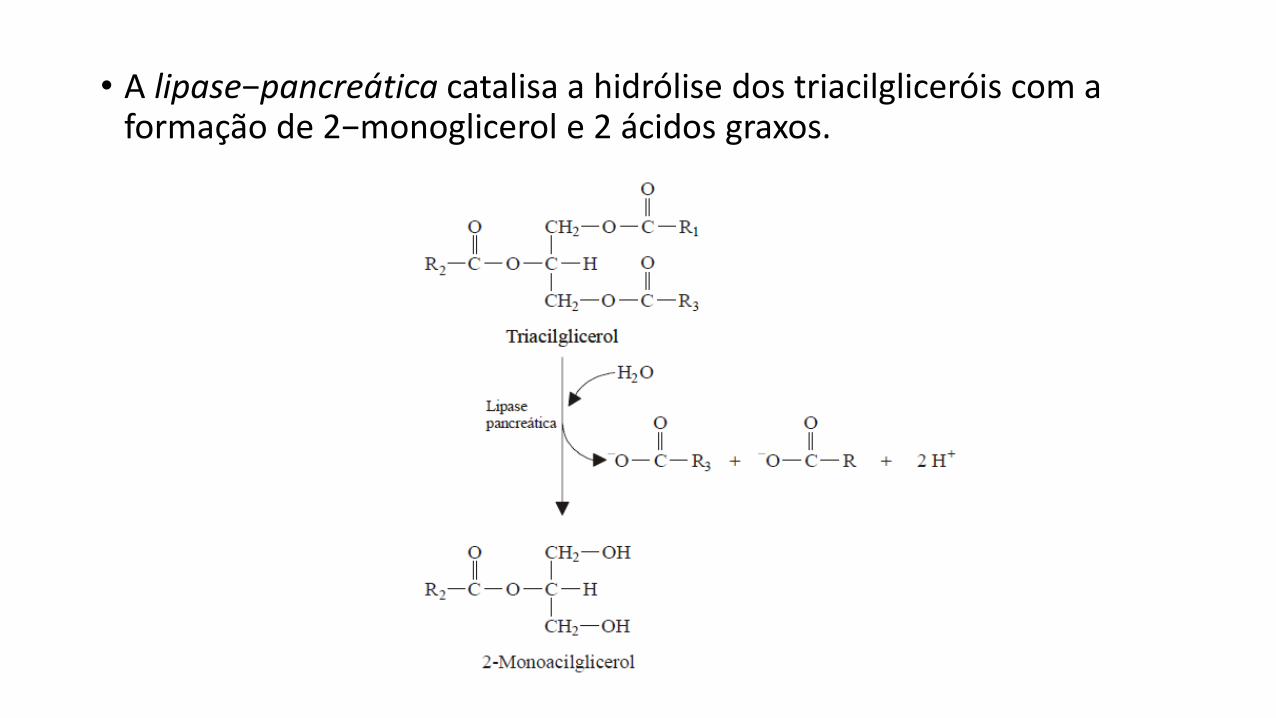
• A lipase−pancreática catalisa a hidrólise dos triacilgliceróis com a formação de 2−monoglicerol e 2 ácidos graxos.

• Os ácidos graxos de fontes alimentares e sintetizados no organismo, sãoesterificados a triacilgliceróis, transportados via corrente circulatória earmazenados como gotículas líquidas no citoplasma das células do tecidoadiposo.
• Os triacilgliceróis constituem a fonte mais concentrada de energia químicado corpo.
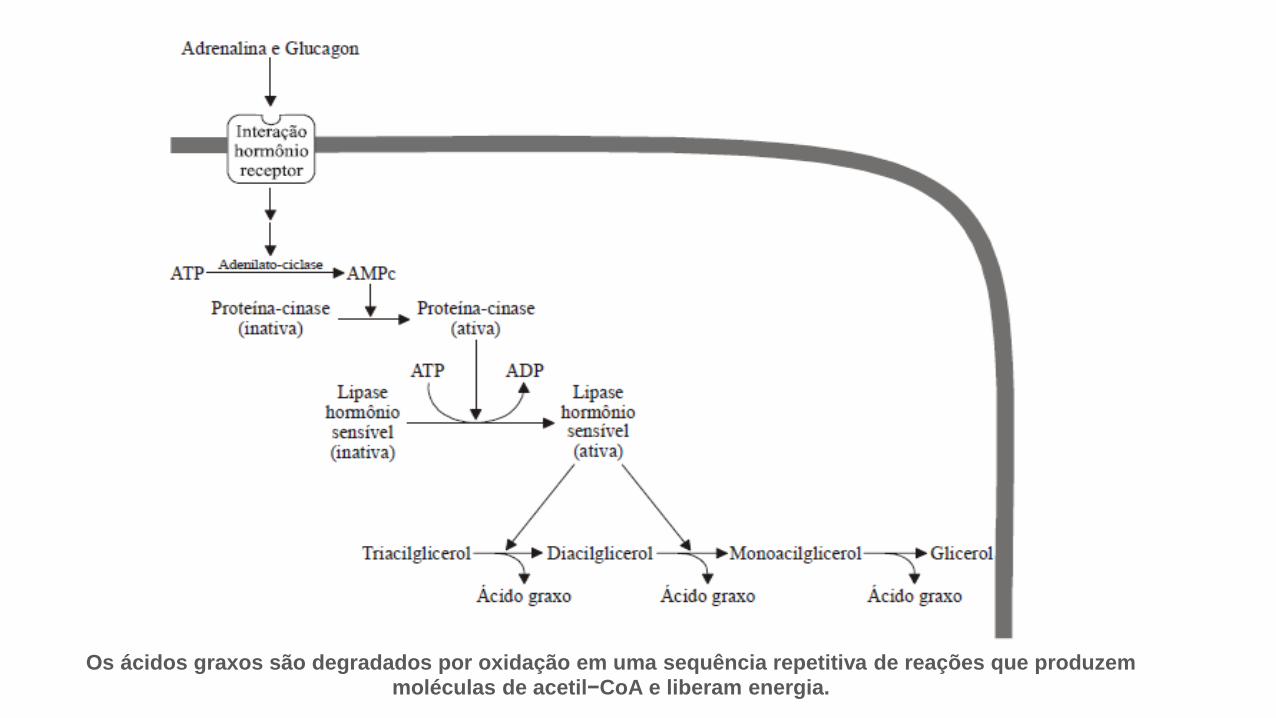
• Durante o jejum, exercício vigoroso e em resposta ao estresse, ostriacilgliceróis armazenados nos adipócitos são hidrolisados em ácidosgraxos e glicerol pela ação da lipase hormônio-sensível.
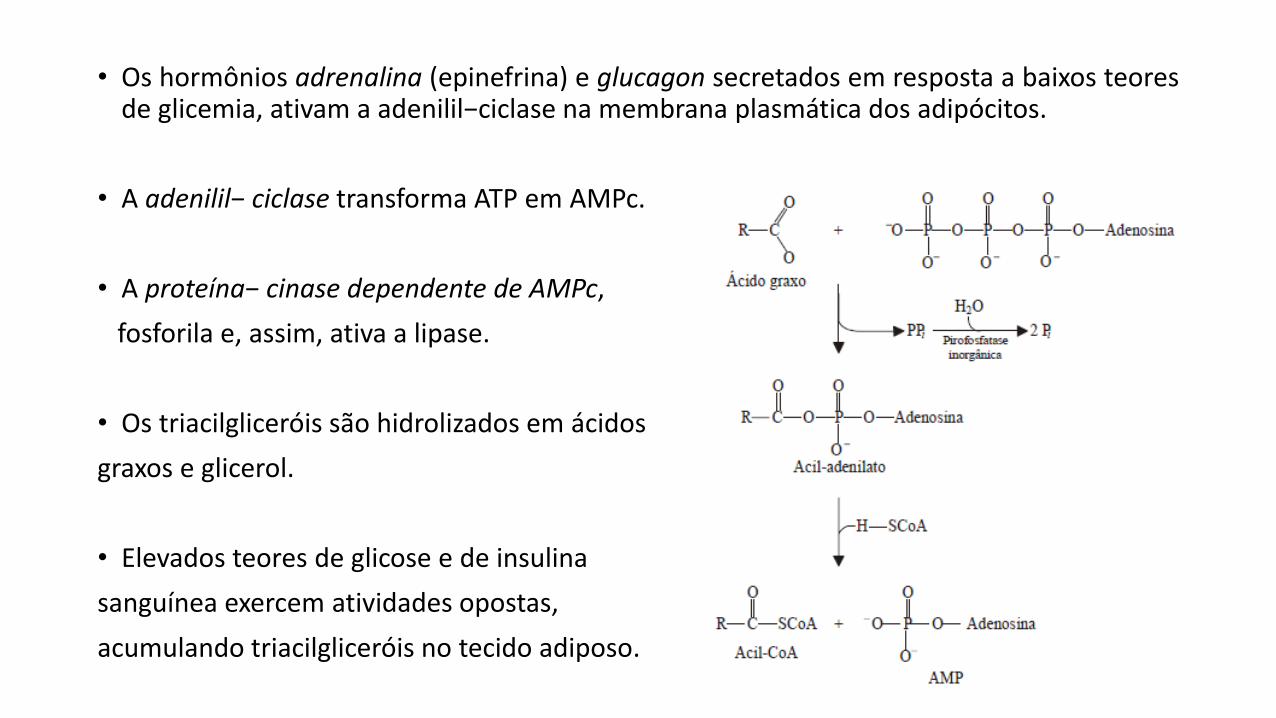
• Os hormônios adrenalina (epinefrina) e glucagon secretados em resposta a baixos teoresde glicemia, ativam a adenilil−ciclase na membrana plasmática dos adipócitos.
• A adenilil− ciclase transforma ATP em AMPc.
• A proteína− cinase dependente de AMPc,
fosforila e, assim, ativa a lipase.
• Os triacilgliceróis são hidrolizados em ácidos
graxos e glicerol.
• Elevados teores de glicose e de insulina
sanguínea exercem atividades opostas,
acumulando triacilgliceróis no tecido adiposo.
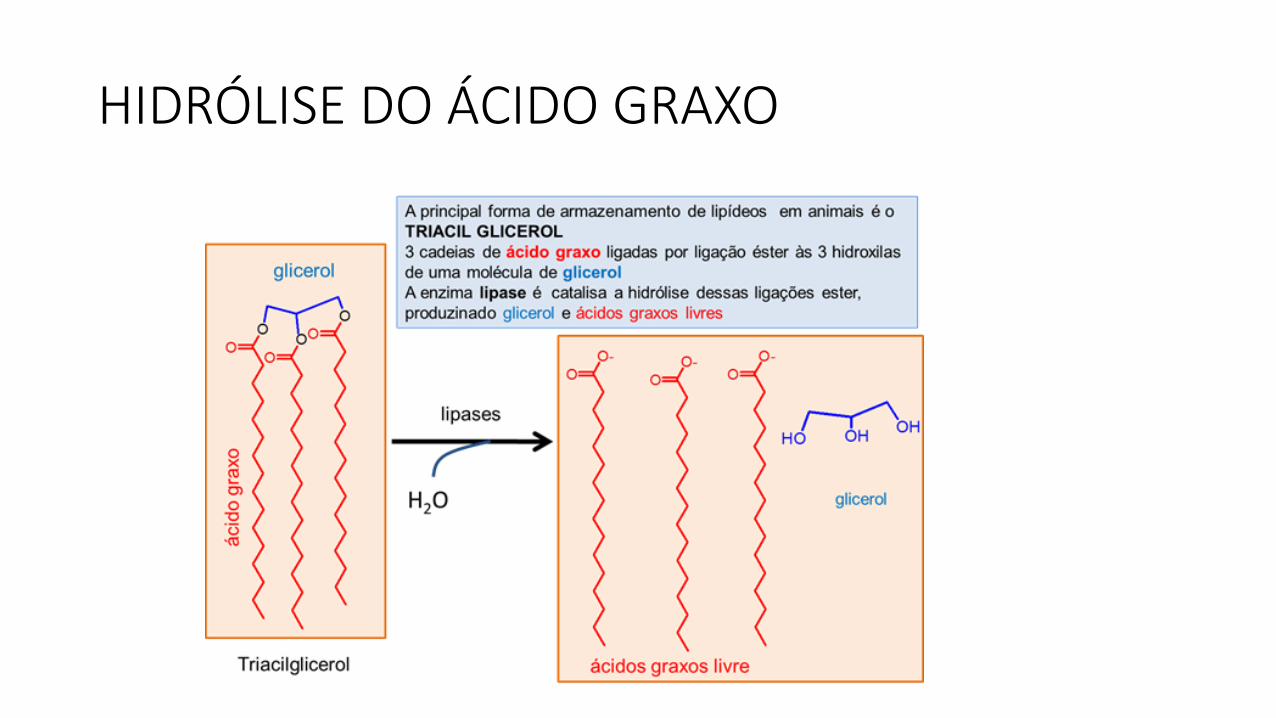
HIDRÓLISE DO ÁCIDO GRAXO

Os ácidos graxos são degradados por oxidação em uma sequência repetitiva de reações que produzem
moléculas de acetil−CoA e liberam energia.

A beta-oxidação é a via de oxidação dos ácidos graxos.
Via catabólica de degradação de ácidos graxos para produção de energia.
Ocorre na matriz mitocondrial, após a ativação e a entrada dos ácidos graxos namitocôndria.
Sua função é produzir acetil-coa que será utilizado no ciclo de krebs, da mesmaforma que o acetil-coa gerado pela glicólise.
13
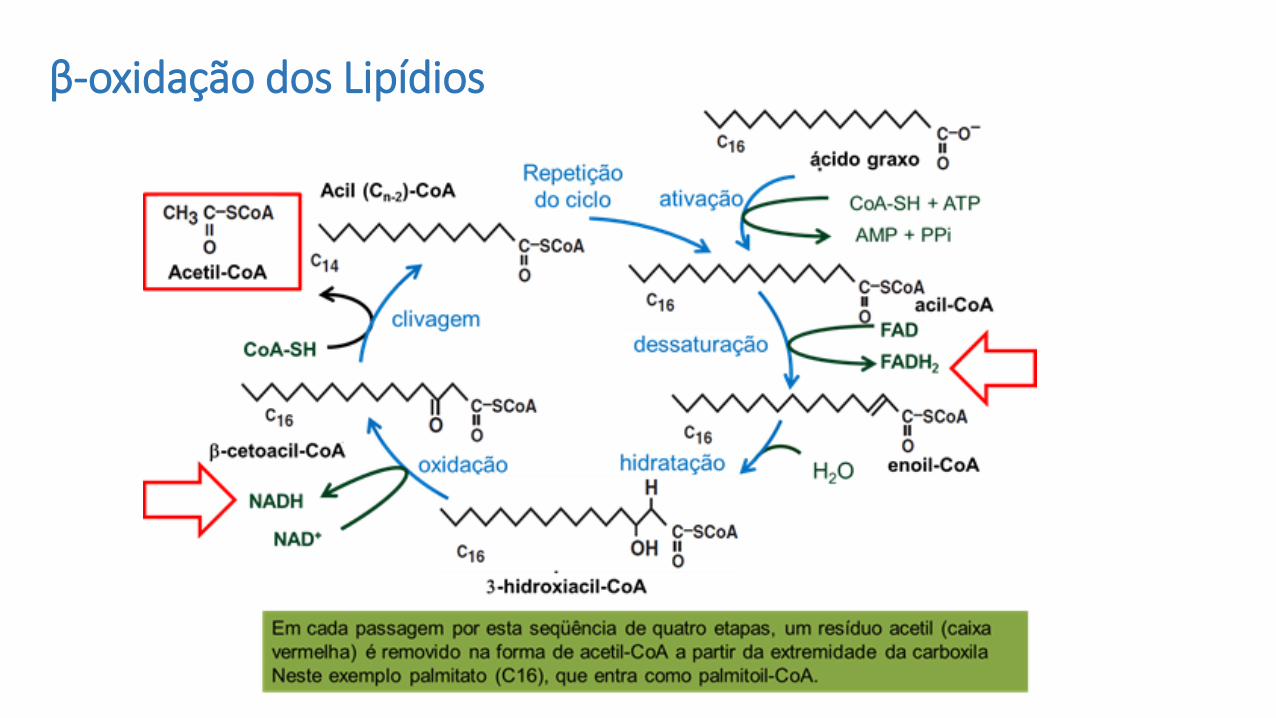
β-oxidação dos Lipídios

Esta via funciona degradando a cadeia carbonada do ácido graxo em ciclos, ecada ciclo é formado por uma sequência de quatro reações que no final gera umamolécula de acetil-coa. Portanto a cada ciclo é gerada uma molécula de acetil-coa.
A quantidade de acetil-coa gerado vai depender do tamanho da cadeia decarbonos, pois é isso que determina a quantidade de ciclos.
14
β-oxidação dos Lipídios

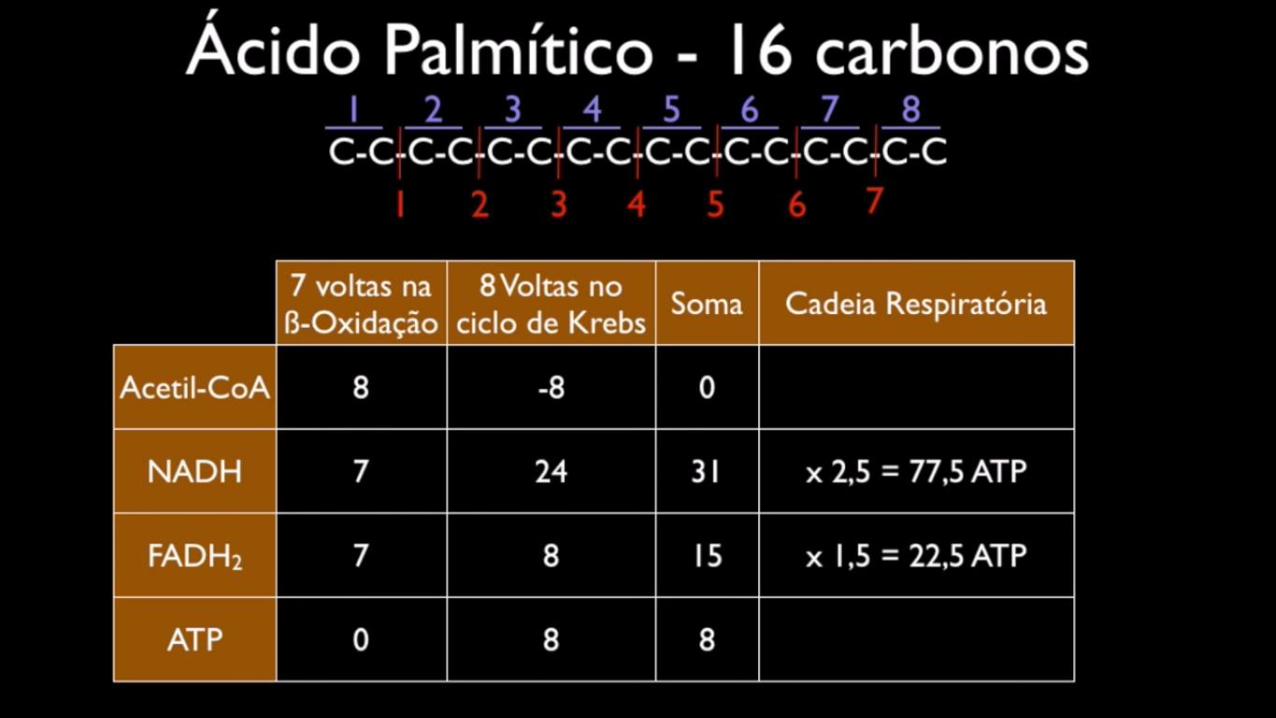
• Em um ácido graxo de 16C há a formação de 8 Acetil-CoA
• 7FADH
• 7NAD
Ciclo de Krebs

Em cada ciclo são retirados dois carbonos da cadeia carbonada do ácido graxopara a formação do acetil-coa.
Com isso é fácil pensar que a quantidade de ciclos seria a metade do valor dototal de carbonos da cadeia carbonada do ácido graxo. Portanto, para uma cadeiade 16 carbonos deveriam ocorrer 8 ciclos.
Entretanto este pensamento é errado, pois o ultimo ciclo gera 2 moléculas deacetil-coa, portanto precisando de um ciclo a menos do que no raciocínioanterior.
16
β-oxidação dos Lipídios

A quantidade correta de ciclos que deveriam acontecer em uma cadeia de 16 carbonos éde sete ciclos.
Isso é fácil entender, pois após ocorrerem 6 ciclos, com cada um retirando 2 carbonos,sobram 4 carbonos.
Logo no sétimo ciclos os 4 carbonos serão divididos em 2 grupos de 2 carbonos, gerandoportanto 2 moléculas de acetil-coa e por isso não sendo necessário a utilização de um cicloextra da beta-oxidação.
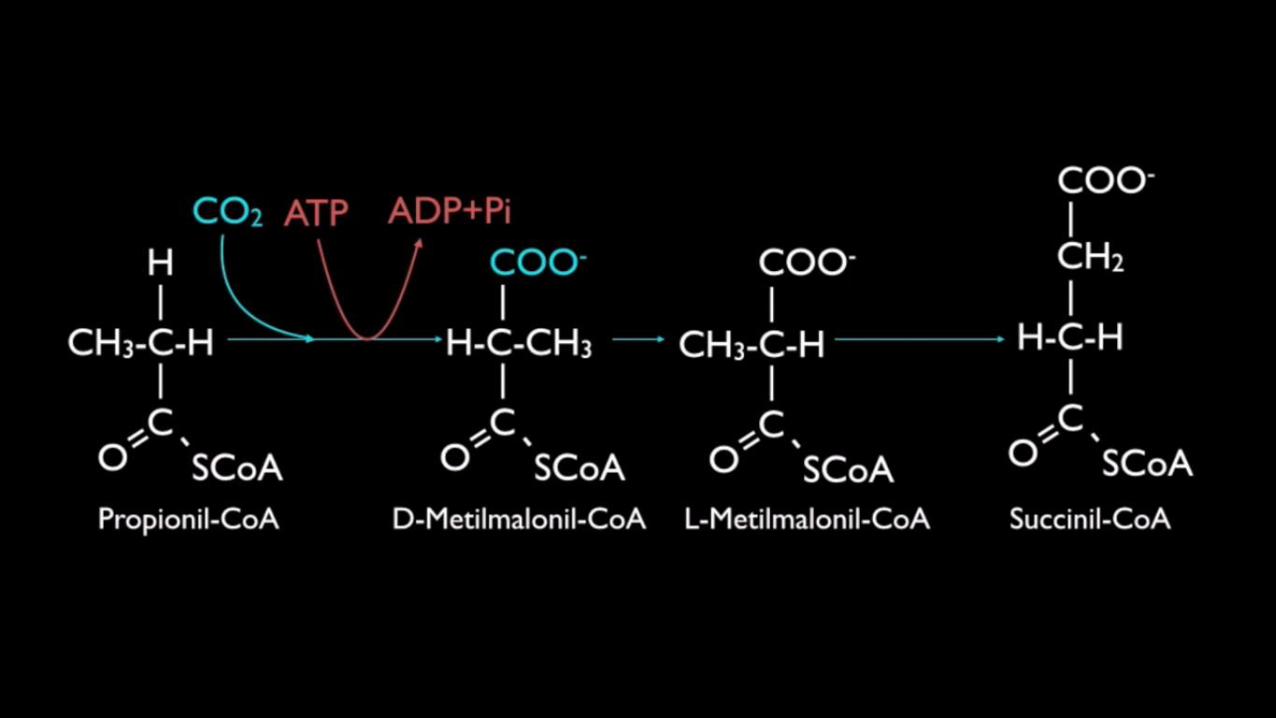
Quando se trata de cadeias ímpares, o ultimo ciclo gera uma molécula de acetil-coa (2carbonos) e uma de 3 carbonos chamada de propionil-coa. Esta ultima moléculaposteriormente é oxidada em succinil-coa a qual é utilizada no ciclo de krebs.
17
β-oxidação dos Lipídios
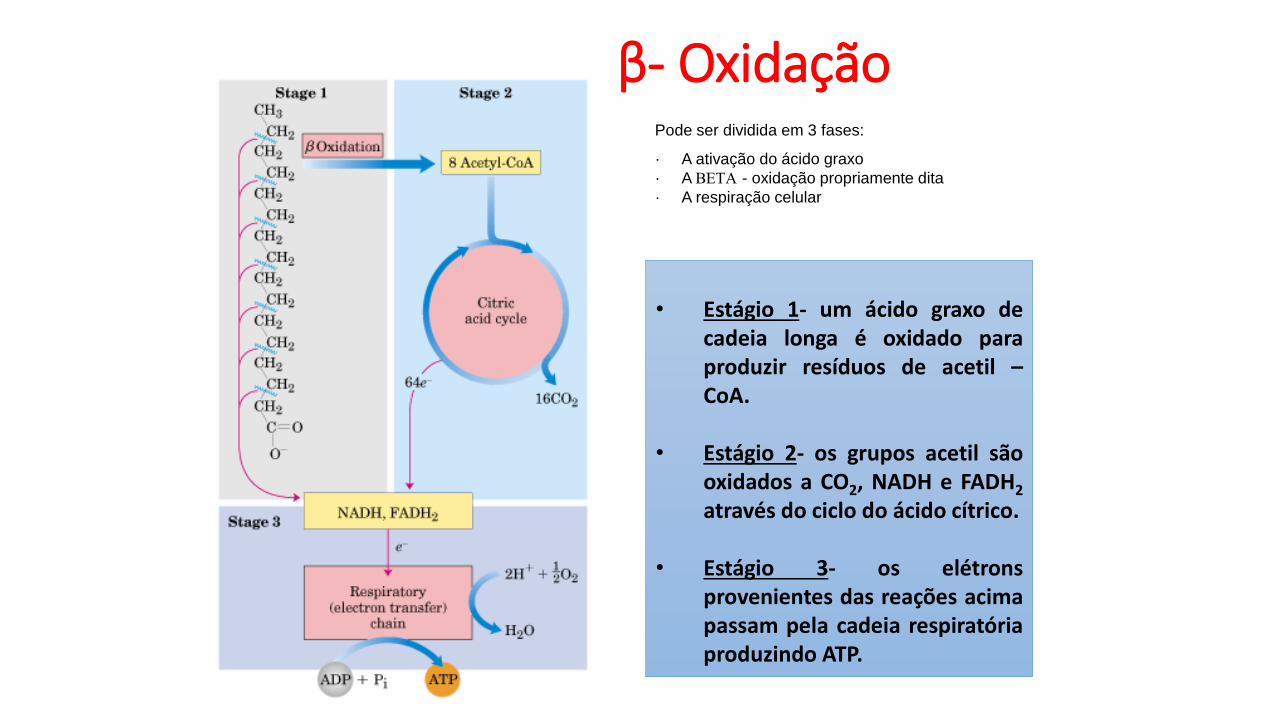
β- Oxidação
• Estágio 1- um ácido graxo decadeia longa é oxidado paraproduzir resíduos de acetil –CoA.
• Estágio 2- os grupos acetil sãooxidados a CO2, NADH e FADH2
através do ciclo do ácido cítrico.
• Estágio 3- os elétronsprovenientes das reações acimapassam pela cadeia respiratóriaproduzindo ATP.
Pode ser dividida em 3 fases:
· A ativação do ácido graxo
· A BETA - oxidação propriamente dita
· A respiração celular

19
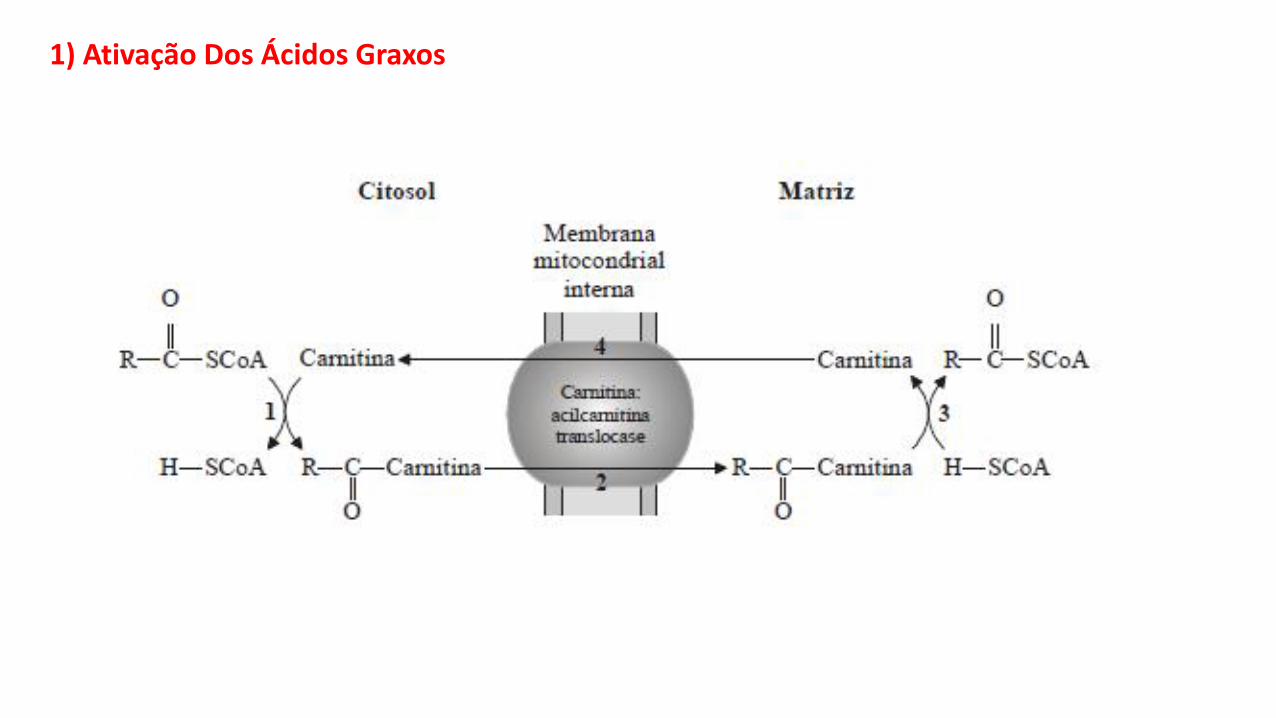
1) Ativação Dos Ácidos Graxos
A ativação dos ácidos graxos consiste na entrada destes na mitocôndria, na forma deACIL-CoA.
O processo depende:
• Da ligação do ácido graxo com a Coenzima A, formando o Acil-CoA no citosol. Areação é catalisada pela enzima Acil-CoA Sintetase, localizada na membranamitocondrial externa:
CH3-(CH2)n-COOH + ATP + CoA-SH CH3-(CH2)n-CO-S-CoA + AMP + PPi

20
1) Ativação Dos Ácidos Graxos
• Do transporte do radical acila através da MMI, do citosol para a matriz, mediado pelocarreador específico carnitina. A transferência do radical acila da CoA para a carnitinaé catalisada pela enzima carnitina-Acil-Transferase I:
Acil-S-CoA + Carnitina Acil-Carnitina + CoA-SH
• Do lado da matriz mitocondrial, a carnitina doa novamente o radical acila para a CoA,regenerando o Acil-CoA no interior da mitocôndria. A reação é catalisada pelaarnitina-Acil-Transferase II, localizada na face interna da MMI, e é exatamente oinverso da descrita acima.
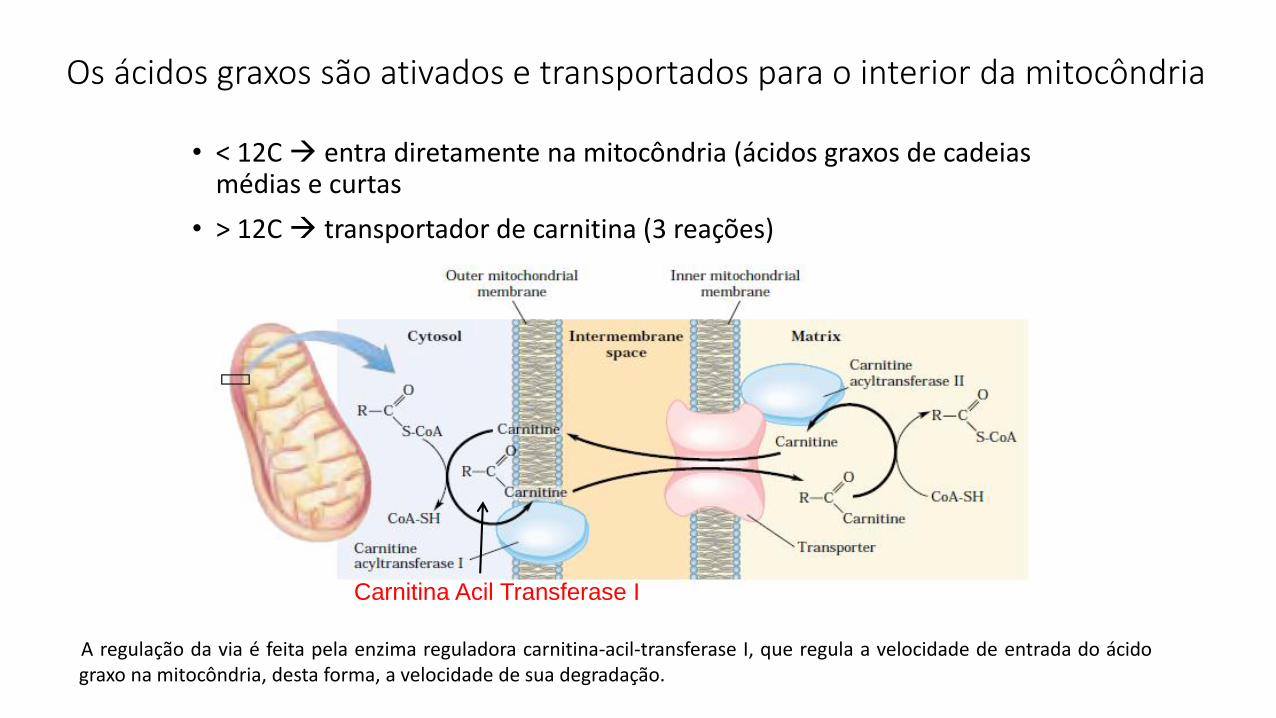
Os ácidos graxos são ativados e transportados para o interior da mitocôndria
• < 12C entra diretamente na mitocôndria (ácidos graxos de cadeias médias e curtas
• > 12C transportador de carnitina (3 reações)
Carnitina Acil Transferase I
A regulação da via é feita pela enzima reguladora carnitina-acil-transferase I, que regula a velocidade de entrada do ácidograxo na mitocôndria, desta forma, a velocidade de sua degradação.
1) Ativação Dos Ácidos Graxos

1. OXIDAÇÃO (FAD)
2. HIDRATAÇÃO (NAD+)
3. RE-OXIDAÇÃO
4. CLIVAGEM
1, 2, 3 e 4 se repetem “n” vezes, de acordo com o tamanho da cadeia de
carbono do ácido graxo
ATIVAÇÃO
ETAPAS da -OXIDAÇÃO

Oxidação
• Consiste na quebra por oxidação do ácido graxo sempre em seu carbono β , convertendo-o na nova carbonila de ácido graxo agora 2 carbonos mais curto.
• O processo é repetitivo, e libera à cada quebra:• 1 NADH+H+
• 1 FADH2
• 1 Acetil CoA
• São 4 as enzimas envolvidas em cada etapa de oxidação da via.
24

• Exemplo:
25
CH3-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CO-S-CoA + CoA-SH
CH3-CH2-CH2-CH2-CH2-CH2-CH2-CO-S-CoA + Acetil-CoA
CH3-CH2-CH2-CH2-CH2-CO-S-CoA + Acetil-CoA
CH3-CH2-CH2-CO-S-CoA + Acetil-CoA
Acetil-CoA + Acetil-CoA

1a OXIDAÇÃO

Oxidação de Ácidos Graxos Insaturados:
27
Se o ácido graxo a ser oxidado for insaturado, o processo tem dois passosenzimáticos adicionais:
A conversão do isômero "cis" em "trans“.
A saturação da dupla ligação pela adição de água.
Uma vez o ácido graxo saturado, ele pode seguir com o processo normal deoxidação.
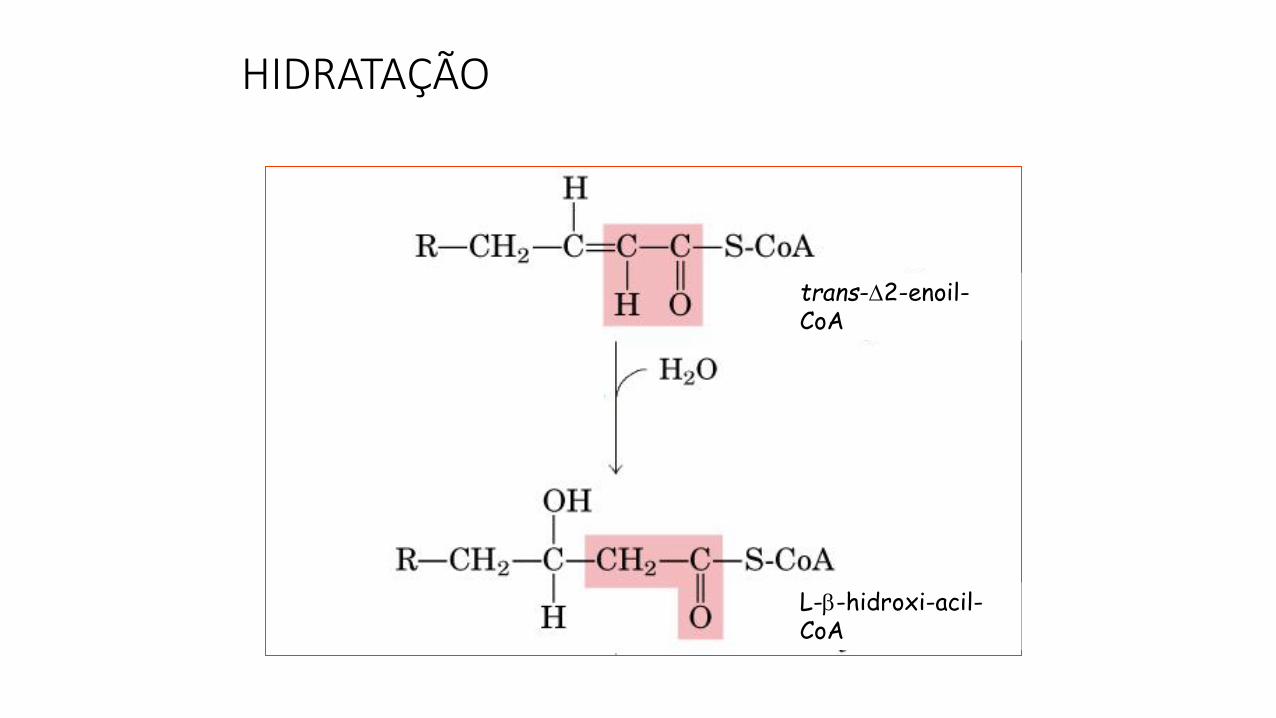
HIDRATAÇÃO
trans-2-enoil-CoA
L--hidroxi-acil-CoA
enoil-CoA hidratase
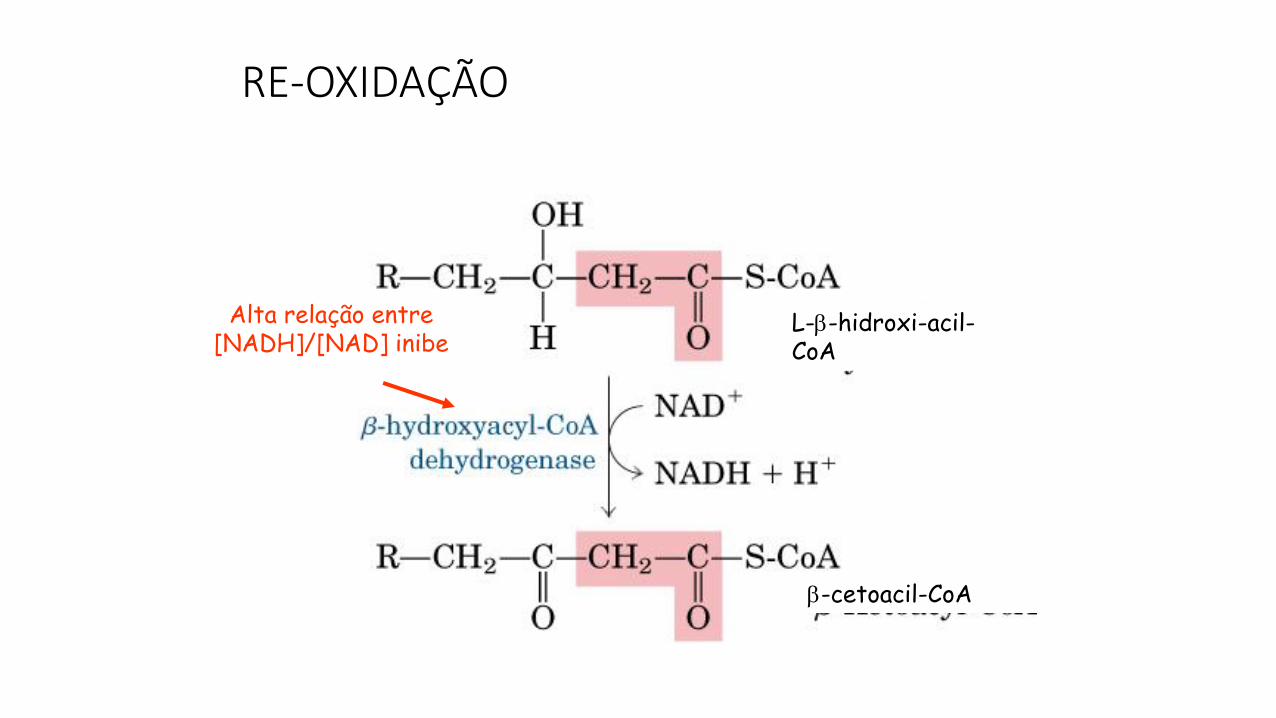
RE-OXIDAÇÃO
L--hidroxi-acil-CoA
-cetoacil-CoA
Alta relação entre [NADH]/[NAD] inibe

CLIVAGEM
-cetoacil-CoA
(C14) acil-CoA
(miristoil-CoA)
acetil-CoA
Alta concentração de acetil-CoA inibe
β-oxidação dos Lipídios
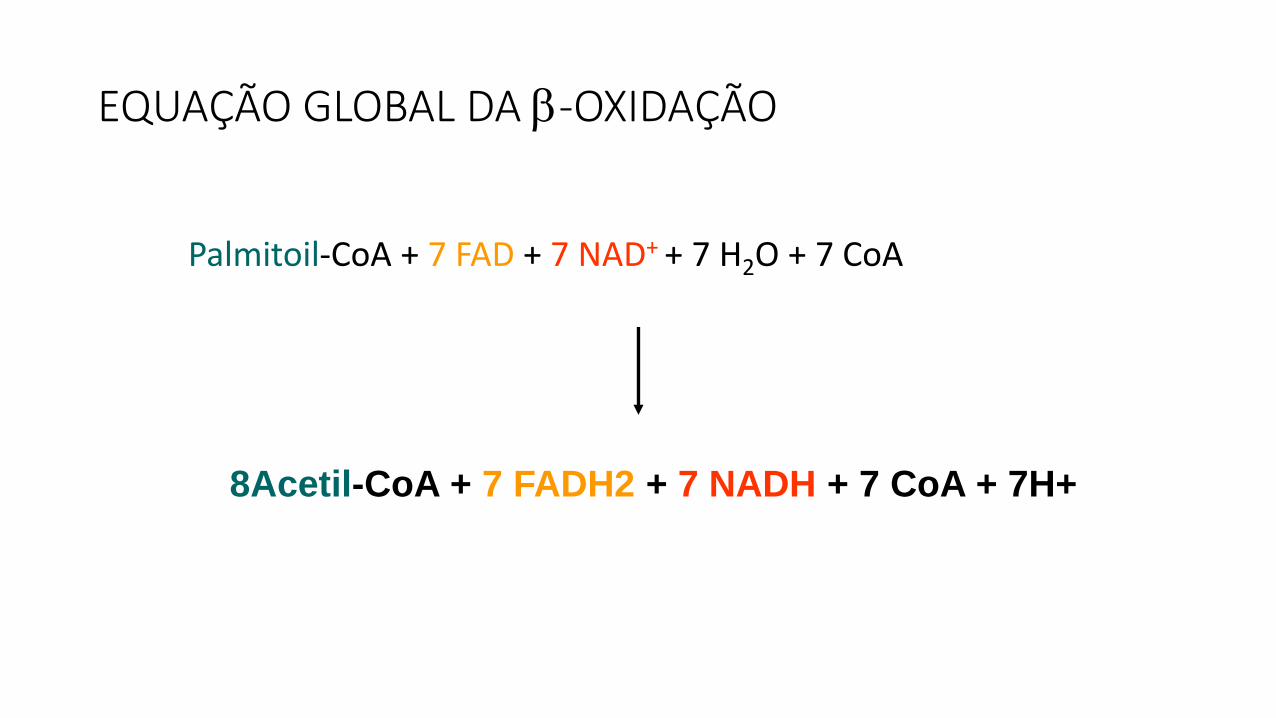
Palmitoil-CoA + 7 FAD + 7 NAD+ + 7 H2O + 7 CoA
8Acetil-CoA + 7 FADH2 + 7 NADH + 7 CoA + 7H+
EQUAÇÃO GLOBAL DA -OXIDAÇÃO


36
Após a β-oxidação, os resíduos acetila do acetil-CoA são oxidados atéchegarem a CO2, o que ocorre no ciclo do ácido cítrico.
Os acetil-coa vindos da oxidação vão entrar nessa via junto com os acetil-coAprovenientes da desidrogenação e descarboxilação do piruvato pelo complexoenzimático da piruvato desidrogenase.
Nessa etapa haverá produção de NADH e FADH2 para suprir de elétrons acadeia respiratória da mitocôndria, que os levará ao oxigênio. Junto a essefluxo de está a fosforilação do ADP em ATP. Com isso a energia gerada naoxidação de ácidos graxos vai ser conservada na forma de ATP.
β-oxidação dos Lipídios

37
Em situações de baixa concentração de glicose no sangue (como jejumprolongado) a β-oxidação é uma alternativa para a produção de energia (poislibera FADH2 e NADH).
Consequentemente, há muita produção de acetil-CoA.
O Ciclo de Krebs não consegue absorver todo esse substrato, estandoprejudicado, uma vez que seus intermediários estão envolvidosna gliconeogênese.
β-oxidação dos Lipídios

38
Essas moléculas de acetil-CoA se condensam, formando Corpos cetônicos, essacondensação acaba liberando Coenzima A, o que é essencial para que hajacontinuidade no Ciclo de Krebs.
Essa produção ocorre principalmente no fígado, que por sua vez não possui acapacidade de degradar corpos cetônicos.
Os corpos cetônicos podem ser usados como fonte de energia no cérebro em casosde desnutrição, nos quais a disponibilidade de glicose é mínima.
β-oxidação dos Lipídios

39
Em situações de baixa concentração de glicose no sangue (como jejum prolongado)a β-oxidação é uma alternativa para a produção de energia (pois libera FADH2 eNADH).
Consequentemente, há muita produção de acetil-CoA.
O Ciclo de Krebs não consegue absorver todo esse substrato, estando prejudicado,uma vez que seus intermediários estão envolvidos na gliconeogênese.
β-oxidação dos Lipídios

40
Essas moléculas de acetil-CoA se condensam, formando Corpos cetônicos, essacondensação acaba liberando Coenzima A, o que é essencial para que hajacontinuidade no Ciclo de Krebs.
Essa produção ocorre principalmente no fígado, que por sua vez não possui acapacidade de degradar corpos cetônicos.
Os corpos cetônicos podem ser usados como fonte de energia no cérebro emcasos de desnutrição, nos quais a a disponibilidade de glicose é mínima.

41
É importante notar que ácidos graxos em sua maioria não são precursores deglicose no corpo humano por não termos enzimas que permitam tal tipo de reação.
O que a beta-oxidação promove para a gliconeogênese é energia para a realizaçãoda gliconeogênese, e não um substrato para a formação de glicose.
No fígado, a energia liberada pela β-oxidação é empregada para dirigir a gliconeogênese.
β-oxidação dos Lipídios

42
Isso acontece porque a maioria dos ácidos graxos de nossa dieta são pares, queapenas geram acetil-coa com sua oxidação como foi visto acima, e não conseguimossintetizar glicose a partir do acetil-coa.
Somente conseguimos gerar glicose a partir de ácidos graxos de cadeia de carbonosímpares, por estas produzirem no final o succinil-coa que é um intermediário dociclo de krebs.
No entanto a quantidade de ácidos graxos de cadeia ímpar é muito pequena emnossa dieta, portanto quase não influenciando no processo da gliconeogênese.
β-oxidação dos Lipídios
![Metabolismo de los lipidos [Modo de compatibilidad] · PDF fileLos restos de acetil-CoA provenientes de la β-oxidación y de la degradación de glucosa o de las cadenas carbonadas](https://static.fdocument.org/doc/165x107/5a943a8b7f8b9aba4a8bd427/metabolismo-de-los-lipidos-modo-de-compatibilidad-restos-de-acetil-coa-provenientes.jpg)





