
1.1 Introduction - Shodhganga : a reservoir of Indian...
46
Chapter 1 General Introduction ………… - 2 - 1.1 Introduction One of the greatest challenges for society in the 21 st century is to meet the increasing demand of fuel energy for transportation and industrial processes in a sustainable way. Global energy consumption continues to increase while fossil-fuel reserves are diminishing. To prevent a looming energy crisis, the development of renewable energy sources is becoming a priority. Bioethanol is the most widely used biofuel for transportation worldwide. Bioethanol from lignocellulosic biomass including agricultural, forestry residues and woody crops appears promising, as the raw material are ubiquitous, abundant; their non-competitiveness with food crops. Lignocellulosic materials consist mainly of cellulose (35-50%), hemicellulose (20-35%) and lignin (10-25%) (Sun and Cheng, 2002). The chemical properties of components of lignocellulose make them a substrate of enormous biotechnological value. Thus, extensive research has been carried out on ethanol production from lignocellulosic materials because it is a cleaner fuel in addition to being a renewable alternative to petroleum (Cadoche et al., 1989; Yang and Wyman, 2008; Binod et al., 2010). Currently, it is a hotspot in the bioenergy research field. 1.2 Cellulose occurrence, structure and composition Cellulose is the most abundant and common renewable organic biopolymer on earth, representing about 1.5 x 10 12 tons (in the tropics) of the total annual biomass production through photosynthesis. It is considered to be an inexhaustible source of raw material for different products. Cellulose is a crystalline biopolymer, where cellulosic chains in the crystals are stiffened by inter- and intra- hydrogen bonds and the adjacent sheets are held together by weak Van-der Waals forces. An important feature of this crystalline array is the relative impermeability of large molecules such as water. In this polymeric structure there are both crystalline and amorphous regions in addition to several types of surface irregularities. This heterogeneity makes the fibers capable of swelling when partially hydrated, resulting in the enlargement of micropores and cavities that allow the penetration of large molecules including enzymes.
Transcript of 1.1 Introduction - Shodhganga : a reservoir of Indian...

Chapter 1 General Introduction …………
- 2 -
1.1 Introduction One of the greatest challenges for society in the 21st century is to meet the increasing
demand of fuel energy for transportation and industrial processes in a sustainable
way. Global energy consumption continues to increase while fossil-fuel reserves are
diminishing. To prevent a looming energy crisis, the development of renewable
energy sources is becoming a priority. Bioethanol is the most widely used biofuel for
transportation worldwide. Bioethanol from lignocellulosic biomass including
agricultural, forestry residues and woody crops appears promising, as the raw material
are ubiquitous, abundant; their non-competitiveness with food crops. Lignocellulosic
materials consist mainly of cellulose (35-50%), hemicellulose (20-35%) and lignin
(10-25%) (Sun and Cheng, 2002). The chemical properties of components of
lignocellulose make them a substrate of enormous biotechnological value. Thus,
extensive research has been carried out on ethanol production from lignocellulosic
materials because it is a cleaner fuel in addition to being a renewable alternative to
petroleum (Cadoche et al., 1989; Yang and Wyman, 2008; Binod et al., 2010).
Currently, it is a hotspot in the bioenergy research field.
1.2 Cellulose occurrence, structure and composition
Cellulose is the most abundant and common renewable organic biopolymer on earth,
representing about 1.5 x 1012 tons (in the tropics) of the total annual biomass
production through photosynthesis. It is considered to be an inexhaustible source of
raw material for different products. Cellulose is a crystalline biopolymer, where
cellulosic chains in the crystals are stiffened by inter- and intra- hydrogen bonds and
the adjacent sheets are held together by weak Van-der Waals forces. An important
feature of this crystalline array is the relative impermeability of large molecules such
as water. In this polymeric structure there are both crystalline and amorphous regions
in addition to several types of surface irregularities. This heterogeneity makes the
fibers capable of swelling when partially hydrated, resulting in the enlargement of
micropores and cavities that allow the penetration of large molecules including
enzymes.

Chapter 1 General Introduction …………
- 3 -
The cellulose is embedded in a matrix of hemicellulose, pectin and lignin (Eriksson
and Bermek, 2009). Lignin and hemicellulose are found in the spaces between
cellulose micro fibrils in primary and secondary cell walls, as well as the middle
lamellae (Eriksson and Bermek, 2009). Fig 1.1 demonstrates a simplified model to
illustrate the cross linking of cellulose microfibril and hemicellulose in lignocellulosic
biomass.
Fig. 1.1 A simplified model to illustrate the cross linking of cellulose microfibril and hemicellulose in lignocellulosic biomass
At the molecular level, cellulose is a linear polymer of glucose composed of
anhydrous units coupled to each other by β-1-4 glycosidic bonds. The number of
glucose units in the cellulose molecules varies and degree of polymerization ranges
from 250 to well over 10,000 units depending on the biomass. Fig 1.2 shows the
schematic representation of a cellulosic chain.
Fig. 1.2 Schematic representation of a cellulosic chain
Chapter 1 General Introduction …………
- 4 -
Hemicellulose is more varied in structure and composition than cellulose and includes
xylan, mannan, galactan and arabinan polymers (Beg et al., 2001). The most abundant
hemicellulose in nature is xylan, containing mainly β-D-xylopyranosyl residues
linked by β-1,4-glycosidic bonds (Koukiekolo et al., 2005). In plants, the xylan forms
an overlying layer through hydrogen bonding with the cellulose, while covalently
linked with lignin which forms an outside sheath to protect the plant. Xylan forms an
important part of the plant cell walls, forming 30–35% of total dry weight, although
the exact abundance of the xylan may differ between plants (Beg et al., 2001).
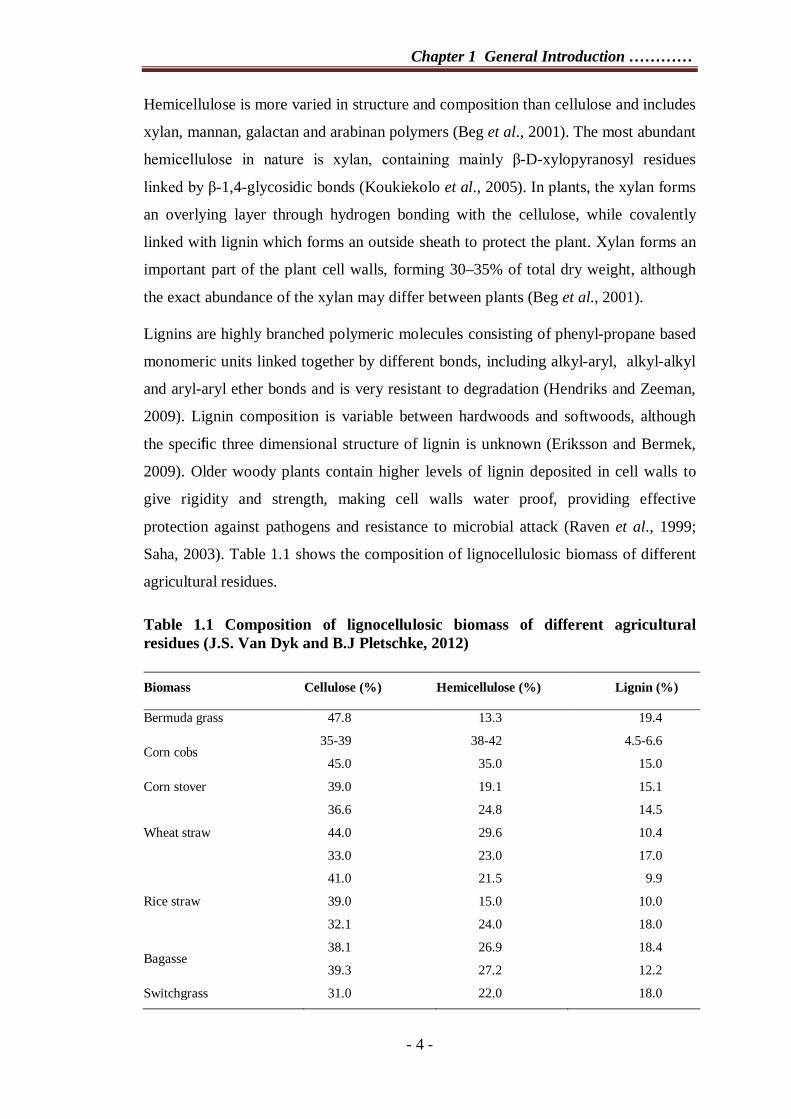
Lignins are highly branched polymeric molecules consisting of phenyl-propane based
monomeric units linked together by different bonds, including alkyl-aryl, alkyl-alkyl
and aryl-aryl ether bonds and is very resistant to degradation (Hendriks and Zeeman,
2009). Lignin composition is variable between hardwoods and softwoods, although
the specific three dimensional structure of lignin is unknown (Eriksson and Bermek,
2009). Older woody plants contain higher levels of lignin deposited in cell walls to
give rigidity and strength, making cell walls water proof, providing effective
protection against pathogens and resistance to microbial attack (Raven et al., 1999;
Saha, 2003). Table 1.1 shows the composition of lignocellulosic biomass of different
agricultural residues.
Table 1.1 Composition of lignocellulosic biomass of different agricultural residues (J.S. Van Dyk and B.J Pletschke, 2012) Biomass Cellulose (%) Hemicellulose (%) Lignin (%)
Bermuda grass 47.8 13.3 19.4
Corn cobs 35-39 38-42 4.5-6.6
45.0 35.0 15.0
Corn stover 39.0 19.1 15.1
Wheat straw
36.6 24.8 14.5
44.0 29.6 10.4
33.0 23.0 17.0
Rice straw
41.0 21.5 9.9
39.0 15.0 10.0
32.1 24.0 18.0
Bagasse 38.1 26.9 18.4
39.3 27.2 12.2
Switchgrass 31.0 22.0 18.0

Chapter 1 General Introduction …………
- 5 -
1.2.1 Cellulase(s) system
Cellulases belonging to a large family of glycosyl hydrolases (GHs) and are classified
into 132 glycoside hydrolase families (http://afmb.cnrs-mrs.fr/CAZY/index.html).
Cellulases mostly have a small independently folded carbohydrate binding module
(CBM) which is connected to the catalytic domain by a flexible linker (Fig 1.3). The
CBMs are responsible for binding the substrate for enzyme activity (Bayer et al.,
1998). Based on the presence and absence of CBMs, the cellulase systems of
microbes can be generally regarded as complexed or non-complexed.
Fig. 1.3 Schematic representation of a CBM with linker
Utilization of insoluble cellulose requires a cellulolytic enzyme system consisting of
either secreted or cell associated cellulases belonging to different classes which are
categorized based on their mode of action and structural properties. The three major
type of cellulase activities recognized are: i) endoglucanase / 1-4-β-D-
gluconohydrolases /EG-(EC3.2.1.4); ii) exoglucanases/1-4-β-D-
glucanglucanohydrolases/cellobiohydrolases/CBH-(EC 3.2.1.91); and iii) β-
glucosidase/BG/BGL/β-glucosdisase glucohydrolases-(EC 3.2.1.21).
Endoglucanases (EGs) are also referred to as carboxymethylcellulases (CMCase),
named after the artificial substrate, carboxymethyl-cellulose (CMC), which is
generally used to measure the enzyme activity. Endoglucanases cut randomly at
internal amorphous sites in the cellulose chain generating oligosaccharides and new
chain ends. This has shown by the effect of the enzyme on CMC and amorphous

Chapter 1 General Introduction …………
- 6 -
cellulose. Fungal EGs are generally monomers with no or low glycosylation and have
an open binding cleft. There are several reports on the existence of multiple EGs in
fungi. In Trichoderma reesei strain at least 5 EGs are present (EG1/Cel7B,
EGII/Cel5A, EGIII/Cel12A, EGIV/Cel61A and EGV/Cel45A). Among these EGI,
EGII, EGIV and EGV possess CBM domain while EGIII does not contain the CBM
(Sandgren et al., 2005). Cellobiohydrolases (CBHs) act on both the reducing and
non reducing ends of the cellulose chains liberating glucose, cellobiose or
cellooligosaccharides as major products, this in turn increases the synergy between
opposite acting enzymes. For example, Trichoderma reesei has two CBHs acting
from non reducing (CBHII/Cel6A) and reducing (CBHI/Cel7A) ends, which results in
a more efficient cellulolytic degrader. Moreover, both CBHI and CBHII of
Trichoderma reesei have CBM at the carboxy terminus or at the amino terminus of
the catalytic module. Similar to EGs, CBHs are monomers with no or low
glycosylation. Cellobiose, the end product of CBHs, act as a competitive inhibitor,
which can limit the ability of the enzymes to degrade all of cellulose molecules in a
system (Divne et al., 1998; Sandgren et al., 2005). β-glucosidases (BGLs) hydrolyze
soluble cellodextrins and cellobiose to glucose. BGLs have been placed mainly in
families 1 and 3 of GH based on their amino acid sequence (Henrissat et al., 1991).
Family 3 includes BGLs from fungi, bacteria and plants, whereas family 1 BGLs are
from bacterial, plant and mammalian origin with galactosidase activity in addition to
β-glucosidase activity. In Trichoderma reesei, two β-glucosidases (BGL1/Cel3A and
BGLII/Cel1A) were isolated from culture supernatant, but the enzymes were found to
be primarly bound to the cell wall (Kubicek et al., 1981). In brief, endoglucanase
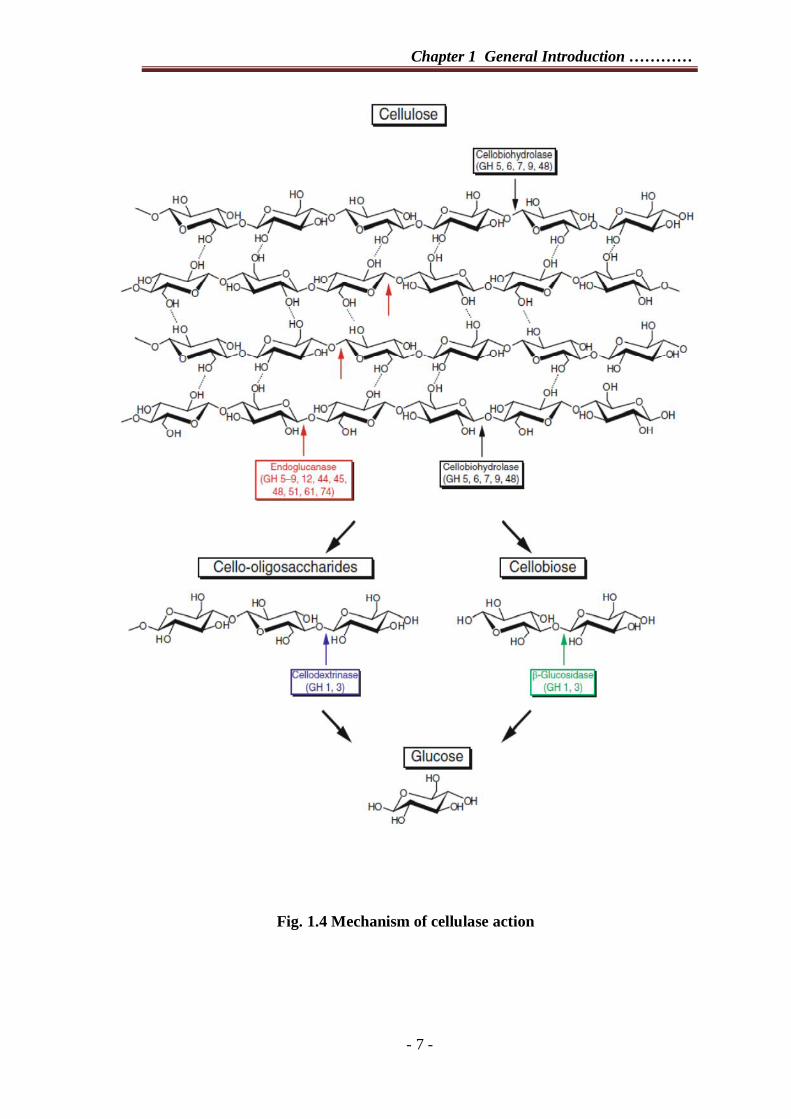
enzymes randomly cleave β-1-4 glucosidic linkages within the backbone of cellulose.
Cellobiohydrolase enzymes cleave from either the reducing or non reducing end of
cellulose chains in a processive manner. Oligosaccharides released as a result of these
activities are converted to glucose by the action of cellodextrinases, whereas the
cellobiose released mainly by the action of cellobiohydrolases is converted to glucose
by β-glucosidases (Fig 1.4).
Chapter 1 General Introduction …………
- 7 -
Fig. 1.4 Mechanism of cellulase action

Chapter 1 General Introduction …………
- 8 -
1.2.2 Synergy among cellulases
Synergy between cellulases have been reviewed and identified between either
different cellobiohydrolases; endo and/or exo-glucanases; endo-glucanases and β-
glucosidases (Walker and Wilson; 1991; Nidetzky et al., 1994; Boisset et al., 2000;
Schwarz, 2001; Zhang and Lynd, 2004).
The degree of synergy or synergism is defined as the ratio of the rate or yield of
product released by enzymes when used together to the sum of the rate or yield of
these products when the enzymes are used separately in the same amount as they were
employed in the mixture (Kumar and Wyman, 2009). Synergy depends on the ratio of
the enzymes involved (Nidetzky et al., 1994), as well as the specific characteristics of
the enzymes and the characteristics of the substrate. The ratios of various cellulases
used in different studies to obtain maximum synergy displayed large variations.
Bothwell et al. (1993) used a ratio of endoglucanase to cellobiohydrolase of 3:1,
while other studies used higher ratios of cellobiohydrolase to endoglucanase. Berger
et al. (2007) used 17:1 cellobiohydrolase to endoglucanase and Jung et al. (2008)
used a ratio of 10:1 cellobiohydrolase to endoglucanase. Hoshino et al. (1997), on
the other hand, found optimal synergy with a combination of 1:1 cellobiohydrolase to
endoglucanase. Boisset et al. (2001) used 98.75% cellobiohydrolase to 1.25%
endoglucanase. It is not clear why such varied ratios have been used, but this may be
as a result of enzyme characteristics, assay conditions or variations in substrate
characteristics, particularly resulting from different pretreatment methods.
1.2.3 Thermostable cellulases
Thermostability is defined as the capacity of an enzyme to retain its active structural
confirmation at a selected temperature for a long period of time. Thermostable
enzymes in the hydrolysis of lignocellulosic materials have several potential
advantages: higher specific activity and higher stability which allow longer life-time,
improved recyclability and increased flexibility for the process configurations. The
two first characteristics would expectedly improve the overall performance of the
enzymatic hydrolysis even at the range of presently used temperatures. Thus, the use
of thermophilic enzymes would ultimately lead to improved performances i.e.
decreased enzyme dosage and reduced hydrolysis time and consequently decreased

Chapter 1 General Introduction …………
- 9 -
hydrolysis costs. Thermostable enzymes would also expectedly allow hydrolysis at
higher consistency due to lower viscosity at elevated temperatures and thus allow more
flexibility in the process configurations.
Several mesophilic or moderately thermophilic fungal strains are known to produce
thermostable enzymes. These enzymes are stable and active at temperatures that can
be clearly higher than the optimum temperatures for the growth of the microorganism.
Filamentous fungi reported to produce cellulases that retain relatively high cellulose
degrading activity at elevated temperatures include particularly those from the species
Talaromyces emersonii, Thermoascus aurantiacus, Chaetomium thermophilum,
Myceliophthora thermophilla and Thielavia terrestris as reviewed by Vikari and
coworkers (Vikari et al., 2007). Thermophilic β-glucosidases have been characterized
from e.g. Aureobasidium pullulans, Clostridium thermphillum, Talaromyces
emersonii, Thermoascus aurantiacus and Thermomyces lanuginose. Although
thermostability is one of the most studied and engineered protein properties, no
generally applicable rules have been established so far. With the availability of
thermostable enzymes a number of new applications in the future are likely. Although,
believed to provide tremendous economical benefits, production of the enzymes to the
required level by the industries has remained a challenge.
1.2.4 Molecular expression and regulation of cellulases
Research on cellulase gene(s) expression is increasing exponentially with a potential to
use the tremendous resources of cellulosic waste for different applications. Owing to
the progress in heterologous expression techniques, recombinant enzyme production
systems are now promising platforms for efficient industrial cellulase production with
many areas where they may enhance the productivity of biomass to biofuel processing
(Alper et al., 2009). Bacterial expression systems for cellulase production have been
modified to efficiently screen heterogenous cellulase genes and organisms like
Zymomonas mobilis (Linger et al., 2010) and Bacillus subtilis (Liu et al., 2012) are
being investigated for roles in consolidated bioprocessing (CBP) (Pottkamper et al.,
2009). Recombinant production of cellulases in bacteria often leads to an increase in
enzyme yield, as compared to the original host, thought to be a result of changes to the
expression systems involving gene copy number, promoters, and inducer types (Afzal
et al., 2010). Yeast expression systems for cellulase production are becoming

Chapter 1 General Introduction …………
- 10 -
increasingly popular even on an individual scale (Mattanovich et al., 2012). Several
yeast expression systems such as Saccharomyces cerevisiae, Pichia pastoris and
Kluyveromyces marxianus have been developed for use in the biofuel industry (Yanase
et al., 2010; Akcapinar et al., 2011). Although important progress has been made
towards industrial, heterologous cellulase production in yeasts, the yield and activity of
enzymes remains substantially lower than Trichoderma reesei that are in currently
industrial use (Elkins et al., 2010). Plant expression systems are potentially an ideal
system for cost effective cellulase production because of their ability to be
economically produced on an industrial scale (Shen et al., 2012; Jung et al., 2012).
Fungal expression systems are the mainstay of cellulase production in current
industrial scale, particularly Trichoderma reesei, Trichoderma longibrachiatum and
Aspergillus niger (Chandel et al., 2012). However, significant efforts have also been
made towards increasing productivity using synthetic biological tools. Although
specific expression strategies in various species of bacteria, yeasts, plants and fungi
show distinct potential, no ideal system has yet been established (Elkins et al., 2010;
Chandel et al., 2012). Thus, an amalgamation of genetically modified plant biomass
and the most promising enzymes, whether identified from natural sources or
synthetically enhanced, incorporated within surface cellulase displaying, ethanol
tolerant yeast or cellulosome displaying bacteria, may be required in combination for
the final crucial breakthrough in this ever expanding field.
Regulation of cellulase production is finely controlled by activation repression
mechanisms (Sukumaran et al., 2008) and genes are found to be coordinately regulated
(Ilmen et al., 1997). Trichoderma reesei cellulases are inducible enzymes and several
inducers were reported in Trichoderma cellulase system where sophorose is one of the
major inducer (Vaheri et al., 1979; Kubicek et al., 1998). Cellobiose, δ-cellobiose-1-5
lactone and other oxidized products of cellulose hydrolysis were also found to act as
inducers. Lactose, another known inducer of cellulase is utilized in commercial
production of enzyme due to its economic reasons. Though mechanism of lactose
induction is not fully understood, it is now known that lack of galactose mutarotase
activity is crucial for cellulase induction in fungus.
Analyses of promoters of cellobiohydrolase I and II has shown binding sites of at least
three transcription activators (ACE I, ACE II and HAP 2/3/5) and one carbon

Chapter 1 General Introduction …………
- 11 -
catabolite repressor (CRE I). Molecular mechanism of gene induction in presence of
cellulose is still unclear. However, it is reported that expression of cellobiohydrolase
and at least two endoglucanases (EG1 and EG2) are believed to be controlled by ACE
II binding to their promoters (Aro et al., 2001; Wurleitner et al., 2003). HAP 2/3/4
binding to cellobiohydrolase I promoter is evidenced by the presence of its binding
sequence in the promoter region (Ilmen et al., 1996a). Glucose repression of cellulase
is supposed to be mediated through carbon catabolite repressor protein CRE I (Ilmen et
al., 1996a) and promoter regions of cbh 1, cbh 2, eg 1 and eg 2 genes have CRE I
binding sites indicating fine control of these genes by carbon catabolite repression
(Kubicek et al., 1998). Glucose repression of cellulase system overrides its induction
(Ilmen et al., 1997) and de-repression is believed to occur by an induction mechanism
mediated by trans-glycosylation of glucose (Sternberg et al., 1979; Fritscher et al.,
1990).
1.2.5 Production of cellulases
1.2.5.1 Microorganisms producing cellulases
Cellulolytic microbes are carbohydrate degraders that primarily utilize cellulose as
energy source for metabolic growth. However, bacteria belonging to Cellulomonas sp.
Cytophaga sp. and fungi utilize a variety of other carbohydrates in addition to cellulose.
Whereas, the anaerobic cellulolytic species have a restricted carbohydrate range,
limited to cellulose and or its hydrolytic products. Most commonly studied cellulolytic
organisms include: Fungal species-Trichoderma, Humicola, Penicillium and
Aspergillus; Bacteria-Bacillus, Psuedomonas, Cellulomonas; and Actinomycetes-
Streptomycetes, Actinomucor and Streptomyces. One of the most extensively studied
fungi is Trichoderma reesei, which converts native cellulose and its derivatives to
glucose by the action of cellulolytic enzymes namely cellulases. Besides Trichoderma
reesei, several fungi like Humicola, Penicillium and Aspergillus can metabolize
cellulose as an energy source, but only few strains are capable of secreting a complex
of cellulase enzymes, which could have potential application in the enzymatic
hydrolysis of cellulose.

Chapter 1 General Introduction …………
- 12 -
Cellulase production is a major research area and with the rejuvenated interest created
due their applications in lignocellulose conversion, several investigations worldwide
are working on these aspects of cellulase (Sukumaran et al., 2005). Production of low
titres of cellulase has always been major concern and researchers are trying to improve
the production titres by adopting multifaceted approaches, which include the use of
better bioprocess technologies and use of cheaper or crude raw materials as substrate
for enzyme production (Sukumaran et al., 2005). Majority of the reports on microbial
production of cellulases utilizes submerged fermentation (SmF) technology. However,
in nature, the growth and cellulose utilization of cellulase producing aerobic
microorganisms probably resembles solid-state fermentation (SSF) than a liquid
culture (Holker et al., 2004; Zhu et al., 2009).
1.2.5.2 Solid-state fermentation
Solid-state fermentation is defined as the fermentation in absence or near absence of
free water (Pandey, 1994). SSF is rapidly gaining interest as a cost-effective
technology due to high titres of microbial cellulase production, especially from fungal
cultures (Jha et al., 1995; Cen and Xia. 1999). Filamentous fungi such as Trichoderma
reesei, Aspergillus niger, Penicillium sp. etc. have been employed for cellulase
production using SSF where a basal mineral salts medium is used for moistening the
substrate. Solid state cultures are strongly recommended for producing cellulases than
submerged cultures since the product concentration in SSF remains quite high thereby
reducing the downstream processing steps, in turn reducing the cost of operation
(Vintila et al., 2009). Chahal (1985) had reported a higher yield of cellulases from
Trichoderma reesei in SSF cultures compared to liquid cultures. Tengerdy (1996)
compared cellulase production in SmF and SSF systems and had indicated that there is
about a 10-fold reduction in the production cost when SSF is employed. Nigam and
Singh (1996) have reviewed the use of agricultural wastes as substrates for cellulolytic
enzyme production under SSF and strongly believe that with the appropriate
technology, improved bioreactor design, and operation controls, SSF may become a
competitive method for the production of cellulases. Cellulases produced in solid-state
culture shows remarkable stability towards temperature, pH and metal ions, etc.
Optimization of SSF conditions when attempted may still improve the overall
production economics and also make it an attractive technology for cellulase

Chapter 1 General Introduction …………
- 13 -
production. It offers many advantages over submerged fermentation, including high
volumetric productivity, higher concentration of products, less effluent generation and
low catabolic repression which make it a promising technology in the near future
(Pandey et al., 1999; Singhania et al., 2009).
1.2.5.3 Submerged fermentation
Submerged fermentation is defined as fermentation in the presence of excess water.
Almost all the large-scale enzyme producing facilities are using this proven technology
due to better monitoring and ease of handling. Though bacteria and actinomycetes are
also reported for cellulase production, the titers are very low to make the technology
economically feasible. Most of the cellulases that are produced commercially under
SmF are from filamentous fungi-Trichoderma reesei or Aspergillus niger and can
perform at diverse pH range and temperature (Cherry et al., 2003). A large-scale
production of cellulases requires understanding and proper controlling of the growth
and enzyme production capabilities of the microbe. Cellulase production in cultures is
highly influenced by various parameters including the nature of the cellulosic
substrate, pH of the medium, nutrient availability, inducer supplementation,
fermentation temperature, etc. The media formulation for fermentation is of significant
concern since no general composition can give the optimum growth and cellulase
production which is dependent on the microbe. In Trichoderma reesei, a basal medium
after Mandels and Reese (1957) or Mandel and Weber (1969) has been most frequently
used with or without modifications.
Mostly, pure cellulose preparations like Solka-Floc and Avicel have been used in the
liquid cultures of cellulolytic microbes for production of the enzymes and when natural
cellulosic materials are used as the carbon source the enzyme yields are significantly
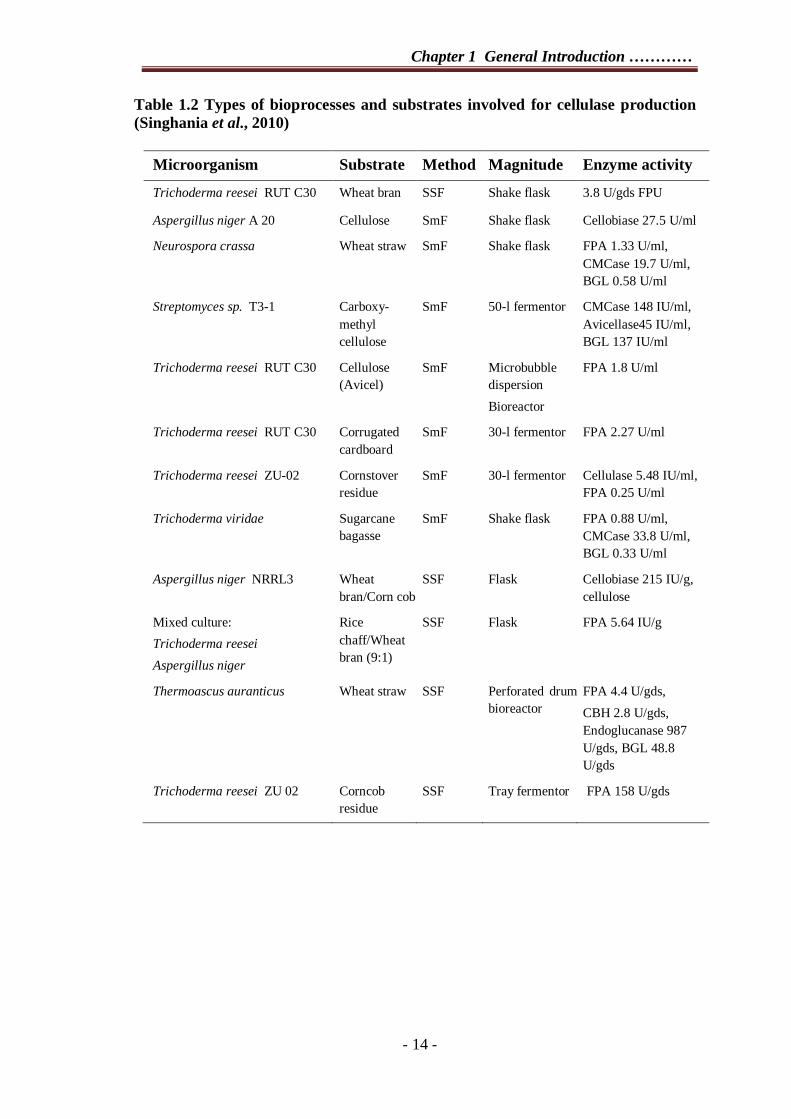
lower (Tangnu et al., 1981). Cellulases produced by SSF and SmF have been listed in
Table 1.2.
Chapter 1 General Introduction …………
- 14 -
Table 1.2 Types of bioprocesses and substrates involved for cellulase production (Singhania et al., 2010)
Microorganism Substrate Method Magnitude Enzyme activity
Trichoderma reesei RUT C30 Wheat bran SSF Shake flask 3.8 U/gds FPU
Aspergillus niger A 20 Cellulose SmF Shake flask Cellobiase 27.5 U/ml
Neurospora crassa Wheat straw SmF Shake flask FPA 1.33 U/ml, CMCase 19.7 U/ml, BGL 0.58 U/ml
Streptomyces sp. T3-1 Carboxy-methyl cellulose
SmF 50-l fermentor CMCase 148 IU/ml, Avicellase45 IU/ml, BGL 137 IU/ml
Trichoderma reesei RUT C30 Cellulose (Avicel)
SmF Microbubble dispersion Bioreactor
FPA 1.8 U/ml
Trichoderma reesei RUT C30 Corrugated cardboard
SmF 30-l fermentor FPA 2.27 U/ml
Trichoderma reesei ZU-02 Cornstover residue
SmF 30-l fermentor Cellulase 5.48 IU/ml, FPA 0.25 U/ml
Trichoderma viridae Sugarcane bagasse
SmF Shake flask FPA 0.88 U/ml, CMCase 33.8 U/ml, BGL 0.33 U/ml
Aspergillus niger NRRL3 Wheat bran/Corn cob
SSF Flask Cellobiase 215 IU/g, cellulose
Mixed culture: Trichoderma reesei Aspergillus niger
Rice chaff/Wheat bran (9:1)
SSF Flask FPA 5.64 IU/g
Thermoascus auranticus Wheat straw SSF Perforated drum bioreactor
FPA 4.4 U/gds, CBH 2.8 U/gds, Endoglucanase 987 U/gds, BGL 48.8 U/gds
Trichoderma reesei ZU 02 Corncob residue
SSF Tray fermentor FPA 158 U/gds

Chapter 1 General Introduction …………
- 15 -
The carbon sources in majority of the commercial cellulase fermentations are
cellulosic biomass ranging from pure cellulose to straw, spent hulls of cereals and
pulses, rice or wheat bran, bagasse, paper industry waste, dairy manure and various
other lignocellulosic residues (Szabo et al., 1996; Reczey et al., 1996; Romero et al.,
1999; Heck et al., 2002; Wen et al., 2005). Rice straw is reported to be an excellent
cheap carbon source for growth of various microorganisms to produce industrially
important microbial enzymes (Gao et al., 2008). Thus, for developing an
economically feasible technology, use of cheaper raw material as a substrate under
SSF for cellulase production could bring down the production costs (Sukumuran et al.,
2005).
1.3 Purification of cellulases and their characterization
Purification of cellulases to homogeneity is necessary for detailed biochemical and
molecular studies and also for the successful determination of primary amino-acid
sequence and three dimensional structure of the protein (Sa-Pereira et al., 2003).
Ideally the final purification process consists of sample preparation, extraction and
clarification followed by three major purification steps i.e. capture (isolate,
concentrate and stabilize), intermediate purification (remove bulk impurities),
polishing (achieve final high level purity). One of the most important steps in
purification of extracellular cellulase is clarification. This step is mainly used to
remove polyphenols, pigments, nucleic acids etc. Clarification can be done by
precipitation, centrifugation or filtration methods, respectively.
Large number of purification protocols for cellulases includes ammonium sulfate
precipitation as the preliminary step in purification process. As an alternative method
to precipitation, ultrafiltration could also be performed where 5-10 kDa molecular cut-
off membranes are generally used. Further, chromatographic methods such as ion
exchange and gel filtration chromatography are used as a final purification step to
obtain high purity cellulase proteins. Microbial cellulases are mainly purified by
chromatographic methods using two to five purification steps. Apart from ion
exchange and gel filtration chromatography, other techniques such as affinity
chromatography and high performance liquid chromatography (HPLC) etc. have also
been used.

Chapter 1 General Introduction …………
- 16 -
Bara et al. (2003) purified exoglucanase by gel filtration and ion exchange
chromatography and obtained 35.7- fold purification. However, a 69-fold purified
protein was obtained using similar techniques by Noronha et al., 2000. For purifying
β-glucosidase Wei et al. (1996) employed ammonium sulfate precipitation, gel
filtration and ion-exchange chromatography step. Using gel filtration chromatography
a 13-fold and 105-fold purified β-glucosidase were obtained from Monascus
purpureus and Paecilomyces thermophila, respectively (Dariot et al., 2008).
Endoglucanases from Aspergillus was purified to homogeneity using ammonium
sulfate precipitation, acetone fractionation and various types of chromatography
methods (gel filtration, sephadex G-100, ion exchange: DEAE-sephadex A-50,
affinity: sepharose-4B). Endoglucanase from Aspergillus terreus M11 was purified
18-fold with a yield of 14% (Gao et al., 2008), 27-fold with a yield of 10.5% from
Aspergillus terreus DSM 826 (Elshafei et al., 2009), 40-fold with a yield of 1.32%
from Aspergillus terreus AN1(Nazir et al., 2009), 2.09-fold with a yield of 18.4%
from Aspergillus niger VTCC F021 (Thai et al., 2010), 12-fold from Aspergillus
awamori F 18 (Nguyen et al., 2010), 5.8-fold with a yield of 3.6% from Penicillium
pinophilum MS 20 (Dipali et al., 2012), respectively.
Many workers have purified and characterized cellulases from different bacteria viz.
Thermomomonospora sp. (George et al., 2001), Pseudomonas fluorescens (Bakare et
al., 2005), Melanocarpus sp. MTCC 3922 (Kaur et al., 2007), Pyrococcus horikoshi
(Kang et al., 2007), Cellulomonas sp. Y15 (Yin et al., 2010), Bacillus sp. (Singh et
al., 2004; Kim et al., 2005; Bischoff et al., 2006; Bajaj et al., 2009; Acharya and
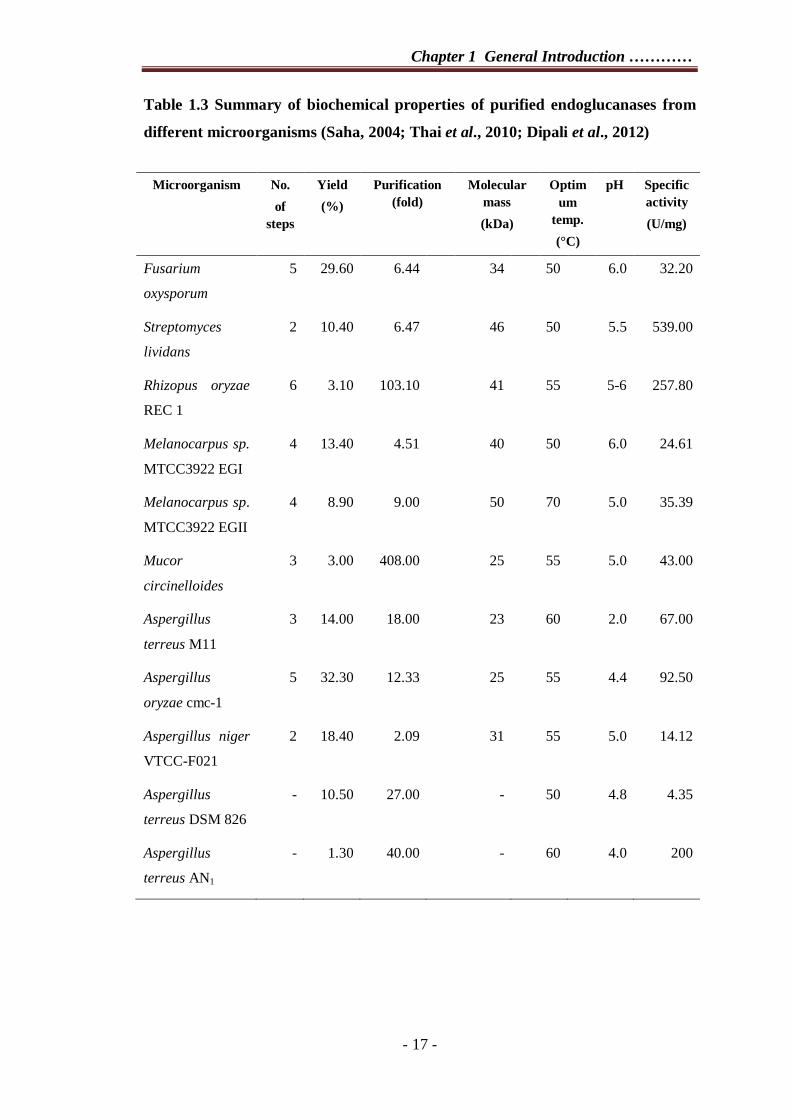
Chaudhary 2011; Ashabil et al., 2011). Table 1.3 shows the summary of biochemical
properties of purified endoglucanases from different microorganisms.
Chapter 1 General Introduction …………
- 17 -
Table 1.3 Summary of biochemical properties of purified endoglucanases from
different microorganisms (Saha, 2004; Thai et al., 2010; Dipali et al., 2012)
Microorganism No. of
steps
Yield (%)
Purification (fold)
Molecular mass (kDa)
Optimum
temp. (°C)
pH Specific activity (U/mg)
Fusarium
oxysporum 5 29.60 6.44 34 50 6.0 32.20
Streptomyces
lividans 2 10.40 6.47 46 50 5.5 539.00
Rhizopus oryzae
REC 1 6 3.10 103.10 41 55 5-6 257.80
Melanocarpus sp.
MTCC3922 EGI 4 13.40 4.51 40 50 6.0 24.61
Melanocarpus sp.
MTCC3922 EGII 4 8.90 9.00 50 70 5.0 35.39
Mucor
circinelloides 3 3.00 408.00 25 55 5.0 43.00
Aspergillus
terreus M11 3 14.00 18.00 23 60 2.0 67.00
Aspergillus
oryzae cmc-1 5 32.30 12.33 25 55 4.4 92.50
Aspergillus niger
VTCC-F021 2 18.40 2.09 31 55 5.0 14.12
Aspergillus
terreus DSM 826 - 10.50 27.00 - 50 4.8 4.35
Aspergillus
terreus AN1 - 1.30 40.00 - 60 4.0 200

Chapter 1 General Introduction …………
- 18 -
1.4 Applications of cellulases
1.4.1 Biofuel industry
Cellulases were initially investigated several decades back for the bioconversion of
biomass to glucose and other fermentable sugars which gave way to research in the
industrial applications of the enzyme in animal feed, food, textiles and detergents and
in the paper industry. With the shortage of fossil fuels and the rising need to feed
alternative source for renewable energy and fuels, there is a renewal interest in the
bioconversion of lignocellulosic biomass using cellulases. Ethanol production of
lignocellulosic biomass comprises the following main steps: hydrolysis of cellulose
and hemicelluloses, sugar fermentation, separation of lignin residue and finally
recovery and purifying the ethanol to meet the fuel specifications. The task of
hydrolyzing lignocellulosic biomass to fermentable sugars is hindered by physico-
chemical, structural compositional factors. Owing to these structural characteristics,
pretreatment is an essential step for obtaining fermentable sugars (Mosier et al.,
2005).
1.4.2 Pretreatment of lignocellulosic biomass
Biomass contains a significant percentage of lignin which has been demonstrated to be
the most important preventive factor in biomass hydrolysis by cellulolytic and
hemicellulolytic enzymes (Dijkerman et al., 1997; Varnai et al., 2010). Dijkerman et
al. (1997) established a correlation between the percentage of lignin and release of
sugars from lignocellulose substrates. It has also been suggested that residual lignin
blocks the progress of cellulase down the cellulose chain (Zhang and Lynd, 2004).
Some researchers indicate that it is not just the presence of lignin, but the type and
distribution that has an impact on enzymatic hydrolysis (Zhang and Lynd, 2004).
Regardless of the cause of reduced bioconversion, the removal or disruption of lignin
is essential for efficient bioconversion of lignocellulose to sugars. During the past few years a large number of pretreatment methods have been
developed which include physical, chemical, physico-chemical and biological
pretreatment etc. (Sun and Cheng, 2002). Pretreatment of lignocellulosic biomass is
costly and crucial for achieving effective hydrolysis of substrates since enzymatic
hydrolysis of native lignocellulose produces less than 20% glucose from the cellulose
fraction (Zhang and Lynd, 2004). But the cost of using un-treated biomass for
Chapter 1 General Introduction …………
- 19 -
enzymatic hydrolysis is much higher when compared to the pretreated substrate
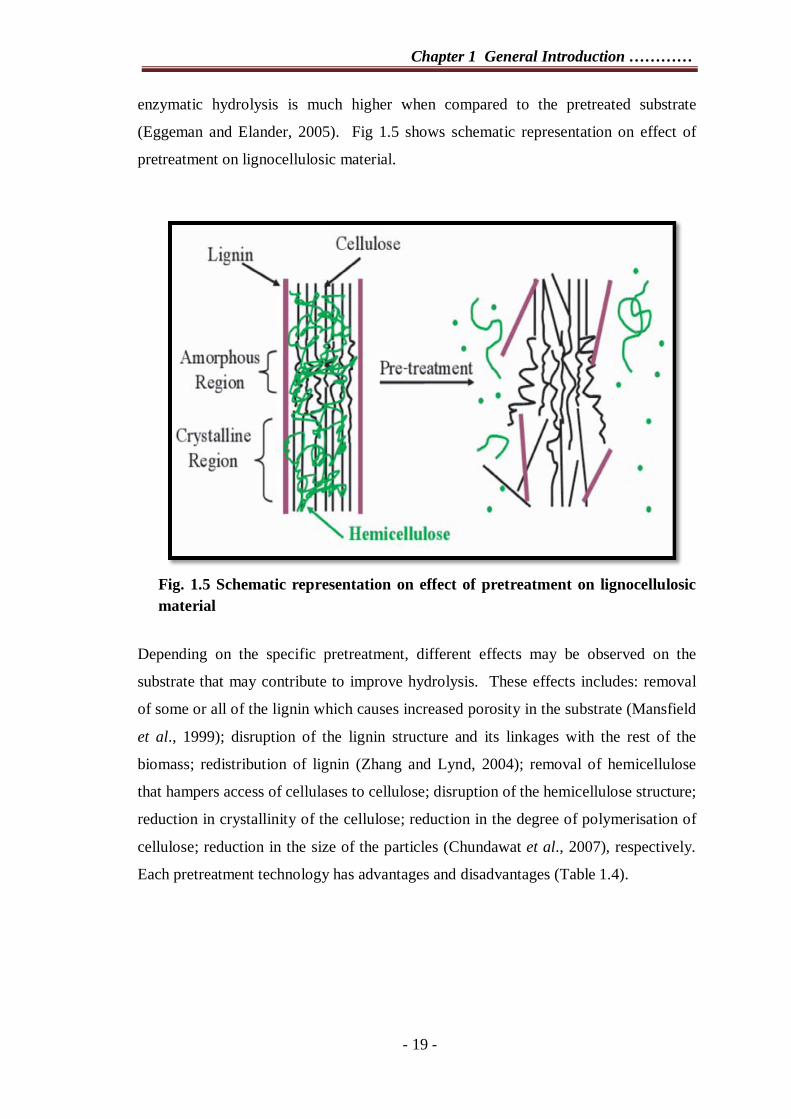
(Eggeman and Elander, 2005). Fig 1.5 shows schematic representation on effect of
pretreatment on lignocellulosic material.
Fig. 1.5 Schematic representation on effect of pretreatment on lignocellulosic material
Depending on the specific pretreatment, different effects may be observed on the
substrate that may contribute to improve hydrolysis. These effects includes: removal
of some or all of the lignin which causes increased porosity in the substrate (Mansfield
et al., 1999); disruption of the lignin structure and its linkages with the rest of the
biomass; redistribution of lignin (Zhang and Lynd, 2004); removal of hemicellulose
that hampers access of cellulases to cellulose; disruption of the hemicellulose structure;
reduction in crystallinity of the cellulose; reduction in the degree of polymerisation of
cellulose; reduction in the size of the particles (Chundawat et al., 2007), respectively.
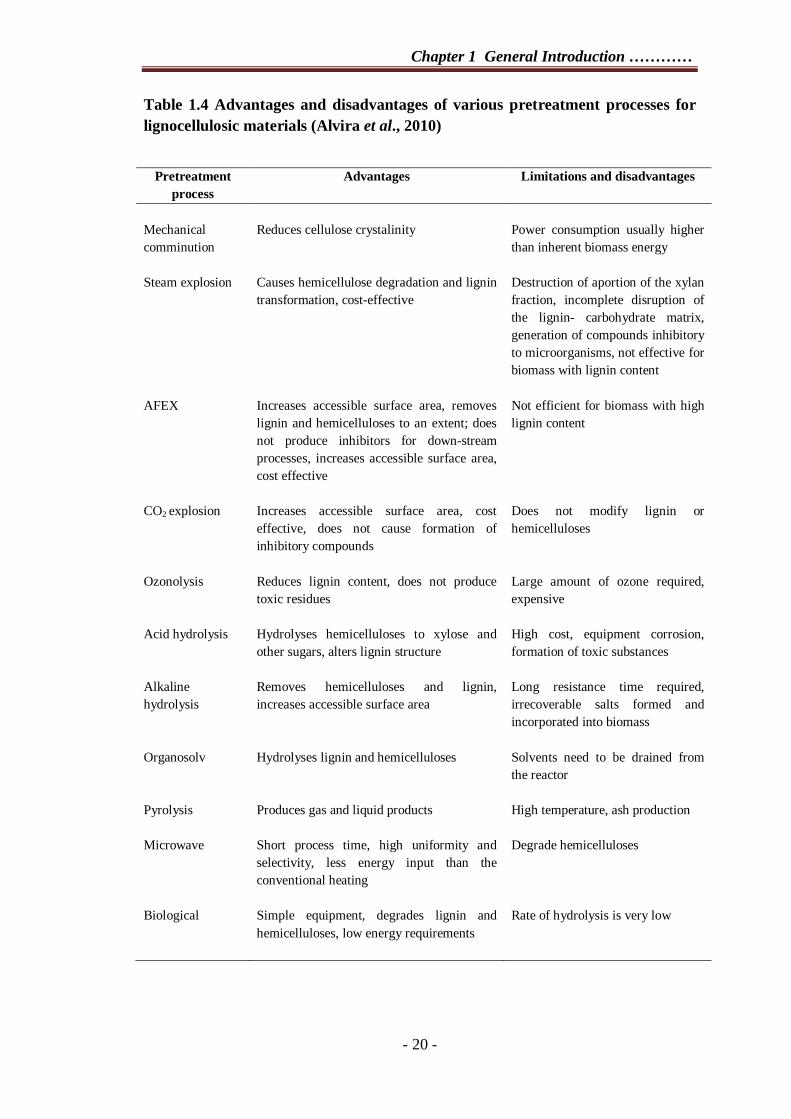
Each pretreatment technology has advantages and disadvantages (Table 1.4).
Chapter 1 General Introduction …………
- 20 -
Table 1.4 Advantages and disadvantages of various pretreatment processes for lignocellulosic materials (Alvira et al., 2010)
Pretreatment process
Advantages Limitations and disadvantages
Mechanical comminution
Reduces cellulose crystalinity
Power consumption usually higher than inherent biomass energy
Steam explosion Causes hemicellulose degradation and lignin transformation, cost-effective
Destruction of aportion of the xylan fraction, incomplete disruption of the lignin- carbohydrate matrix, generation of compounds inhibitory to microorganisms, not effective for biomass with lignin content
AFEX Increases accessible surface area, removes lignin and hemicelluloses to an extent; does not produce inhibitors for down-stream processes, increases accessible surface area, cost effective
Not efficient for biomass with high lignin content
CO2 explosion Increases accessible surface area, cost effective, does not cause formation of inhibitory compounds
Does not modify lignin or hemicelluloses
Ozonolysis Reduces lignin content, does not produce toxic residues
Large amount of ozone required, expensive
Acid hydrolysis Hydrolyses hemicelluloses to xylose and other sugars, alters lignin structure
High cost, equipment corrosion, formation of toxic substances
Alkaline hydrolysis
Removes hemicelluloses and lignin, increases accessible surface area
Long resistance time required, irrecoverable salts formed and incorporated into biomass
Organosolv Hydrolyses lignin and hemicelluloses Solvents need to be drained from the reactor
Pyrolysis Produces gas and liquid products High temperature, ash production
Microwave
Short process time, high uniformity and selectivity, less energy input than the conventional heating
Degrade hemicelluloses
Biological
Simple equipment, degrades lignin and hemicelluloses, low energy requirements
Rate of hydrolysis is very low

Chapter 1 General Introduction …………
- 21 -
Finally, an effective and economical pretreatment should meet the following
requirements: (a) production of reactive cellulose fiber for enzymatic attack (b)
avoiding destruction of hemicelluloses and cellulose (c) avoiding formation of
possible inhibitors for hydrolytic enzymes and fermenting microorganisms (d)
minimizing the energy demand (e) reducing the cost of size reduction of feed stocks
(e) reducing the cost of material for construction of pretreatment reactors and (f)
consumption of little or no chemical and using a cheap chemical.
1.4.3 Enzymatic hydrolysis (Saccharification)
Enzymatic hydrolysis of lignocellulose has long been studied as a method to
depolymerize the biomass into fermentable sugars and conversion to biofuels. It
requires less energy and mild environment conditions (Ferreira et al., 2009). It has
several advantages due to low toxicity and utility cost, less corrosion and no
inhibitory compounds formation as compared to acid hydrolysis (Sun and Cheng,
2002; Taherzadeh et al., 2008; Ferreira et al., 2009). Since, enzymatic hydrolysis is
carried out by cellulase enzymes that are highly substrate specific (Taherzadeh et al.,
2008).
The performance of cellulase mixtures in biomass conversion processes depends on
several properties including enzyme stability, product inhibition, synergism among
the different enzymes, productive binding to the cellulose, physical state as well as the
composition of cellulosic biomass (Heinelman et al., 2009). The choice of the enzyme
preparation for a particular biomass would be more dependent on biomass
characteristics rather than on standard enzyme activities measured (Kabel et al.,
2005), such as filter paper activity (FPU), carboxymethylcellulase (CMCase), β-
glucosidase and xylanase activities. It would not be feasible to predict the efficiency
of cellulases for bioconversion only on the basis of standard cellulase assays, as there
are no clear relationships between cellulase activities on soluble and insoluble
substrates (Zhang et al., 2006). Thus, the soluble substrates should not be used to
predict the efficiency of cellulases for processing relevant solid substrates such as
plant cell walls. Since filter paper is highly crystalline cellulose, preparations having
higher FPUs are desirable for bioconversion and the degradation of which depends on
the combination of activities of EG and CBH (Vincken et al., 1994).

Chapter 1 General Introduction …………
- 22 -
Preparations of cellulase from a single organism may not be very efficient for
hydrolysis of a particular feedstock. Though, the filamentous fungi are the major
source of cellulases and hemicellulases and the mutant strains of Trichoderma
including Trichoderma reesei, Trichoderma viride and Trichoderma longibrachium
are the best known producers, these species have a low level of β-glucosidase activity
(Duff et al., 1996). The key to developing cellulases that are effective towards a
particular biomass feedstock is to artificially construct them either by enzyme
assembly to form cocktails or to engineer the cellulase producers to express desired
combination of cellulase enzymes (Schulein et al., 2000). There are several reports
available where an enzyme cocktail has been employed successfully for biomass
conversion (Zhang et al., 2006; Park et al., 2009; Singh et al., 2009; Sukumuran et
al., 2009). Cellulases for biomass conversion could be a blend or enzyme cocktail
containing endo- and exo-cellulase, xylanase, β-glucosidase, pectinase, etc. which
could vary for different biomass on the basis of their composition. Enzyme cocktails
are also developed by mixing Trichoderma reesei cellulase with other enzymes
including xylanases, pectinases and β-glucosidases and these cocktails have been tried
for hydrolysis of various feedstocks (Xin et al., 1993; Berlin et al., 2007). The
hydrolytic efficiency of a multi enzyme cocktail for lignocellulose saccharification
depends both on biochemical properties of individual enzymes and their ratio
(Gusakov et al., 2007). Some of the following synergies have been reported for
cellulase interaction on complex substrates (degree of synergy shown in brackets
where reported): Endo xylanase and endo-glucanase on sugarcane bagasse (3.59)
(Beukes et al., 2008); Endo xylanase and cellulases on corn cell walls (1.6)
(Murashima et al., 2003); Cellobiohydrolase, endo xylanase, acetyl xylan esterase and
ferulic acid esterase on pretreated corn stover (84% increase) (Selig et al., 2008).
The development of these enzyme cocktails for lignocellulose degradation therefore,
appears to focus on finding the appropriate accessory enzymes to enhance the
degradation of cellulose. There are many factors which influence the enzymatic
hydrolysis of cellulose in lignocellulosic feedstock such as temperature, pH, substrate
load, enzyme load and mixing rate etc. (Olsson et al., 1996), but recently the focus is
on operating at high solid loadings. The process at high solid loading offers many
advantages (≥15% solid, w/w) over conversions performed at low or moderate solid
loads, including increased sugar and ethanol concentrations and decreased capital and

Chapter 1 General Introduction …………
- 23 -
investment costs (Modernbach et al., 2013). The use of high solid operations would
make biofuels more economical and more price-competitive with petroleum.
As solid loading increases, challenges that were negligible in low-solid systems
become more prominent, this has also been noted in high solid pretreatment
(Modernbach et al., 2012). One of the major challenges for enzymatic hydrolysis at
high solid loading is the lack of available water in the reactor. Water is essential for
effective hydrolysis mainly for two reasons: mass transfer and lubricity. Water
increases the effectiveness of the enzymatic and chemical reactions by providing a
medium for solubilizing and aiding in the mass transfer of products. Water also
reduces the viscosity of the slurry by increasing the lubricity of the particles, which
decreases the required shear stress necessary to produce a given shear rate, allowing
lower power input for mixing (Hodge et al., 2009; Kristensen et al., 2009). The
physical and chemical properties of the specific biomass affect the way biomass
absorbs water. As solid loadings approach 20% (w/w), the liquid fraction becomes
fully absorbed into the biomass leaving little free water (Hodge et al., 2009). With
lower amounts of free water, the apparent viscosity of the mixture increases, and
consequently mixing and handling of material become more difficult. Other
challenges specific to high-solid enzymatic hydrolysis include long hydrolysis times.
Enzymatic hydrolysis is typically thought to be the bottleneck of the entire
bioconversion process in terms of both time and money, since the reaction time
needed for most enzymes to convert lignocellulose into sufficient glucose
concentrations for fermentation is on the order of days (usually ≥3 days). Long
hydrolysis times could only be reduced by increasing enzyme loading. Recent studies
have suggested that enzymes can overcrowd accessible cellulose sites. Thus not
reaching the full hydrolytic potential for the given enzyme loading (Xu et al., 2007;
Bommarius et al., 2008). Enzyme is usually added based on per weight of a substrate.
As the solid loading increases the amount of enzyme should also increase
propotionately. The cost of enzyme is still at a level that makes this step in the
conversion process one of the most expensive. Therefore, is important to evaluate the
economics when determining the balance between the loadings applied to the
lignocellulose and the amount of time needed to reach sufficient glucose
concentrations. Researchers are addressing these issues from many angles,
experimenting with different pretreatment methods and various enzyme sources and

Chapter 1 General Introduction …………
- 24 -
cocktails, while modifying operating conditions and slurry properties. Although there
has been some success at the pilot and demonstration scale, many questions must be
resolved before the full potential of high-solids lignocellulose conversion will be
realized. Several studies have been reported on the conversion of cellulosic biomass
to sugars by enzymatic hydrolysis at high solids using a horizontal orientation of the
reactor (Jorgensen et al., 2007; Larsen et al., 2008; Roche et al., 2009). One of the
highest solid loadings in enzymatic hydrolysis reported to date is 40% (w/w)
(Jorgensen et al., 2007 and 2009). A horizontal oriented rotating drum was utilized as
the reactor in these studies in order to effectively mix the solids. The studies found
that cellulose and hemicelulose conversion decreased from 90% to 33% and 70%-
35%, respectively, with the increase in solid loadings from 2% to 40%, but the reactor
was providing adequate mixing as evidenced by the conversion of lignocellulose into
fermentable saccharides (86 g glucose/kg at 40% solids).
1.4.4 Recovery of cellulases
It is necessary to develop efficient methods to produce renewable fuels from
lignocellulosic biomass. Recent commercial cellulase preparations have been shown
to be effective at hydrolyzing cellulose under industrially relevant conditions;
however, the high cost of enzymes remains a significant barrier to the economical
production of ethanol from lignocellulosic biomass (Weiss et al., 2013). It is
therefore necessary to reduce the amount of enzymes required for the enzymatic
hydrolysis step. A variety of methods have been suggested to achieve increased
hydrolysis yields, including surfactant addition (Borjesson et al., 2007), gradual
substrate loading (Rosgaard et al., 2007) or advance reactor configurations coupled
with product removal to avoid inhibition (Andric et al., 2010; Yang et al., 2010).
However, these methods have yet to be shown to be cost effective. One method that
may reduce the amount of enzyme used and increase enzymatic productivity is to
recover/recycle the enzymes (Ramos et al., 1993; Tjerneld et al., 1994). The majority
of cellulase recycling methods that have been reported involved either separating the
enzymes from the solids or liquid phases or recycling of the solid and/or liquid phase
directly. Approaches have been demonstrated where free enzymes were recovered
from the liquid fraction by membrane filtration (Tjerneld et al., 1994). Similarly, the
enzymes associated with the solids have been recovered by washing with excess

Chapter 1 General Introduction …………
- 25 -
volumes of buffer, sometimes with surfactants to desorb the enzymes, which were
then concentrated and added to the fresh substrate (Tu et al., 2009). These methods
have shown different levels of success under controlled laboratory conditions and
with specially prepared feedstocks.
1.4.5 Bioethanol production
Each year a large portion of agricultural residues is disposed of as “waste” can
potentially be used to produce various value added products like biofuels, animal
feeds, chemicals, enzymes etc. Rice straw is one of major crop produced adundantly
all over the world. It is annually produced about 731 million tons which is distributed
in Africa (20.9 million tons), Asia (667.6 million tons), Europe (3.9 million tons),
America (37.2 million tons) and Oceania (1.7 million tons) (Balat et al., 2008). The
options for the disposal of rice straw are limited by the bulk of material, slow
degradation in the soil, harboring of rice stem diseases and high mineral content. Only
a small portion of globally produced rice straw is used as animal feed, the rest is
removed from the field by burning, a common practice all over the world, thus
increasing air pollution and affecting human health (Ogawa et al., 2005; Chen et al.,
2007; Wati et al., 2007). This huge amount of rice straw can potentially produce 205
billion liters bioethanol per year, which is the largest amount from a single biomass
feedstock.
Even though the fermentative process for ethanol production is well known, the
production costs are still the key impediment. Intense research has been carried out
for obtaining efficient fermentative microorganisms, low-cost fermentation substrates
and optimal environmental conditions for fermentation. The ideal strain for ethanol
production of lignocellulosic biomass should possess the following characteristics:
stable conversion of C5 and C6 sugars, resistance against inhibitory compounds,
tolerance concerning temperature, ethanol as well as sugar and industrial stability.
Regardless of the feedstock, the degradation of celluloses or hemicelluloses yields
hexoses and pentoses that need to be fermented to ethanol using yeast strains.
Current approaches are inefficient, since no native microorganisms can convert all
sugars into ethanol at a high yield (Chen, 2011). Therefore, microorganisms that can
efficiently convert both hexoses and pentoses to ethanol would be more useful.

Chapter 1 General Introduction …………
- 26 -
Saccharomyces cerevisiae is the most favoured organism for ethanol production from
hexoses and does not have genes encoded for xylose reductase (XR) and xylitol
dehydrogenase (XDH) and thus cannot utilize xylose. A way to overcome this
obstacle of pentose utilization is through recombinant DNA technology (genetic
engineering). Other approach is to use pentose fermenting microorganisms. Yeasts
such as Pichia stipitis, Candida shehatae and Pachysolen tannophilus are capable of
fermenting both hexose and pentose sugars to ethanol (Chandel et al., 2007; Chen,
2011). Thermophilic bacteria have also been considered for fermentation of pentose
containing lignocellulosic hydrolysates. Thermophilic and saccharolytic Clostridia
are an important group of ethanol producing microorganisms and include species of
Clostridium thermohydrosulfuricum, Thermoanaerobacter (formerly Clostridium)
Thermosaccharolyticum and Clostridium thermocellum. These bacteria may translate
pentoses and aminoacids into ethanol and can synthesize upto 2 mol EtOH/mol
hexose. Having saccharolytic properties, these microorganisms have the ability to
grow on a wide variety of non treated wastes. Clostridium thermocellum can even
directly convert lignocellulosic materials into ethanol (McMillan, 1997). On the other
hand, there are a few cases where the immobilization of these yeasts increases the
ethanol productivity (Chandrakant and Bisaria, 1998), unlike the case of hexose
fermenting yeasts or Zymomonas mobilis.
The processes usually employed in the fermentation of lignocellulosic hydrolysate are
simultaneous saccharification and fermentation (SSF) and separate hydrolysis and
fermentation (SHF). SSF is a single step process whereas SHF is a two step process.
During SSF process the solid fraction of pretreated lignocellulosic material
simultaneously undergoes hydrolysis (saccharification) and once hydrolysis is
completed, the resulting cellulose hydrolysate is fermented and converted into
ethanol. However, one of the main features of the SHF process is that each step can
be performed at its optimal operating conditions. The most important factors to be
taken into account for saccharification step are reaction time, temperature, pH,
enzyme dosage and substrate load. The SHF process has been employed either
conventionally or traditionally but SSF is superior for ethanol production since it can
improve ethanol yields by removing end product inhibition and simultaneously
eliminate the need for separate reactors. In SSF, the cellulases and microorganisms
are added to the same process allowing that the glucose formed during the enzymatic

Chapter 1 General Introduction …………
- 27 -
hydrolysis of cellulose be immediately consumed by the microbial cells converting it
into ethanol. Thus, the inhibition causes by the sugars over the cellulases is
neutralized. Since this process operates at non-optimal conditions for hydrolysis and
requires high enzyme dosage, which positively influence on substrate conversion, but
negatively on process costs. Considering that enzymes account for an important part
of production costs, it is necessary to find methods reducing the cellulase doses to be
utilized (Bjerre et al., 1996; Hamelinck et al., 2005). Apart from SSF or SHF, CBP is
also performed for ethanol production (Cardona et al., 2008). In CBP, mono-or co-
culture of microorganisms is generally used to ferment cellulose directly to ethanol.
Bacteria such as Clostridium thermocellum and some fungi including Neurospora
crassa, Fusarium oxysporum and Paecilomyces sp. have been used in CBP.
However, this process is not efficient because of poor ethanol yield and long
fermentation periods (3-12 days). Sequential fermentation with two different
microorganisms in different time periods of the fermentation process for better
utilization of sugars has also been employed using Saccharomyces cerevisiae in the
first phase for hexose utilization and Candida shehatae in the second phase for
pentose utilization but ethanol yields achieved are not high (Cardona et al., 2008).
Some studies on separate fermentation of individual hydrolysates using C6 specific
yeast strains have been reported (Gupta et al., 2009; Laser et al., 2009). The ethanol
yield obtained from various cellulosic hydrolysate using Saccharomyces cerevisiae
are: Sweet sorghum (0.40 g g-1) (Mamma et al., 1995), Corn cob (0.48 g g-1) (Chen et
al., 2007), Prosopis juliflora (0.49 g g-1) (Gupta et al., 2009) and Lanata camara
(0.33 g g-1) (Kuhad et al., 2010). Moniruzza et al. (1997) reported sequential
fermentation of glucose and xylose from rice straw by using Saccharomyces
cerevisiae ATCC 26603 and Pichia stiptis NRRL Y-7124 and yielded 4 g l-1 and 6 g l-
1 ethanol, respectively. Other microorganisms capable of fermenting a great variety
of sugars including hexoses and pentoses is the fungus Mucor indicus reaching
ethanol yields of 0.46 g g-1 glucose when cultivated under anaerobic conditions (Sues
et al., 2005).

Chapter 1 General Introduction …………
- 28 -
1.4.6 Anaerobic digestion of liquid waste generated during ethanol production
process
While developing an integrated process technology for transport fuel production from
lignocellulosic material, a large amount of solid and liquid wastes are generated,
therefore, it is important to extract energy potential to maximum extent from the
unutilized side streams, which will help us to have a zero waste sustainable process.
Anaerobic digestion (AD) is a process that occurs only under strict anaerobic
conditions where several populations of bacteria react in concert to decompose
complex organic material to methane and carbon dioxide without an external electron
acceptor such as oxygen or nitrate. Anaerobic digestion can be considered as one of
the oldest technologies for waste and wastewater treatment. It is also probably the
major biological process involved in landfill wastes decomposition. There are several
advantages recognized to the process when used for wastewater treatment plants such
as high capacity to treat slowly degradable substrates at high concentrations, very low
sludge production (5-10 times less than in aerobic processes), potentiality for valuable
intermediate metabolites production, low energy requirements (no aeration is
required), reduction of odors in a closed system, pathogens reduction and possibility
for energy recovery through methane combustion or even hydrogen production
(Steyer et al., 2002).
1.4.6.1 Anaerobic treatment systems
All modern high rate biomethanation processes are based on the concept of retaining
highly concentrated viable biomass by some mode of bacterial sludge immobilization.
The processes are attractive alternative to anaerobic lagoon and conventional
digesters. A wide variety of process configurations have been made available which
hold the concept of immobilized cells. The various configurations developed so far
include Upflow anaerobic filter by Young and McCarty (1969), Anaerobic attached
film expanded bed by Switzen Baum and Jewell (1980); Upflow anaerobic sludge
blanket by Lettinga et al., (1980); Anaerobic rotating biological contactor by Tait and
Friedman (1980); Anaerobic fluidized bed by Jeris (1982); and Downflow stationary
fixed film by van den Berg et al., (1985). Switzen Baum and Jewell (1980) developed
the concept of fixed film reactors from the initial idea of anaerobic filters by Young

Chapter 1 General Introduction …………
- 29 -
and McCarty (1969). Anaerobic hybrid reactor which combines advantages of both
anaerobic filter and up flow anaerobic sludge blanket designs, operates with a sludge
blanket in the lower zone and packing media forming a filter in the upper zone, was
first developed in 1981 (Maxham and Wakamiya, 1981). This type of reactor has been
also successfully and widely applied for the treatment of low to medium strength
wastewater (Kumar et al., 2007). The most important aspects in the AHR design are
the selection of appropriate support material. A variety of natural materials such as
smooth quartzite pebbles, shells, granite stones, wooden blocks, brick bats and
synthetic materials like polyvinyl chloride sheets, rasching rings, tyre rubber and
other materials have been used for the attachment and growth of anaerobic biomass
(Gourari and Achkari-Begdouri, 1997; Reyes et al., 1999; Show and Tay, 1999;
Michaud et al., 2002; Melidis et al., 2003; Rovirosa et al., 2004; Yang et al., 2004). It
was reported that the reactors packed with high media porosity obtained better organic
removal efficiency compared to that using non-porous support. Thus, a hybrid
treatment system will reduce investment costs through the use of cheaper local media
and also its flexibility to deal with almost all kinds of wastewater.
1.4.7 Textile industry
Cellulases have become the third largest group of enzymes used in the industry since
their introduction only since a decade. They are used in biostoning of the denim
garments for producing softness and the fade look replacing the use of pumice stones
which were traditionally employed. They act on the cellulose fiber to release the
indigo dye used for coloring the fabric, producing the faded look of denim. Though
use of acidic cellulase from Trichoderma along with the proteases is found to be
equally good, Humicola insolens cellulase is commonly employed in the biostoning
process. Cellulases are utilized for digesting off the small fiber ends protruding from
the fabric resulting in a better finish. Cellulases have also been used in softening,
defibrillation and in processes for providing localized variation in the color density of
fibers.

Chapter 1 General Introduction …………
- 30 -
1.4.8 Laundry and detergents Cellulases, in particular EG III and CBH I, are commonly used in detergents for
cleaning textiles. Several reports disclose that EG III variants, in particular from
Trichoderma reesei and Trichoderma harzianum and Aspergillus niger are
industrially utilized natural sources of cellulases. Cellulase preparations, mainly from
species of Humicola that are active under mild alkaline conditions and at elevated
temperature, are commonly added in washing powders and in detergents.
1.4.9 Food and animal feed In food industry, cellulases are used in extraction and clarification of fruit and
vegetable juices, production of fruit nectars and purees and in the olive oil. Cellulases
are also used in carotenoid extraction in the production of food coloring agents.
Enzyme preparations containing hemicellulases and pectinase addition to cellulases
are used to improve the nutritive quality of forages. Improvements in feed
digestibility and animal performance are reported with the use of cellulases in feed
processing.
1.4.10 Pulp and paper industry In the pulp and paper industry, cellulases and hemicellulases have been employed for
biochemical pulping for modification of coarse mechanical pulp and hand sheet
strength properties, de-inking of recycled fibers and for improving drainage and run
ability of paper mills. Cellulases are employed in the removal of inks, coating and
toners from paper. Bio-characterization of pulp fibers is another application where
microbial cellulases are employed. They are also used in preparation of easily
biodegradable cardboard and in the manufacture of soft paper including paper towels
and sanitary paper and preparations containing cellulases are used to remove adhered
paper.

Chapter 1 General Introduction …………
- 31 -
1.5.0 Cellulase market scenario
Globally, there are two major companies known for cellulase production for biomass
conversion-“Genencor” and “Novozyme”. Both the companies has played a
significant role in bringing down the cost of cellulase several folds by their active
research and are continuing to bring down the cost by adopting novel technologies.
Recently, Genencor has launched Accelerase® 1500, a cellulase complex intended
specifically for lignocellulosic biomass processing industries
(http://www.genecor.com/ wps/wcm/coonect/genecor/products). It is claimed to be
more cost effective and efficient for bioethanol industries than its predecessor-
Accelerase® 1000. Accelerase® 1500 is produced with a genetically modified strain of
Trichoderma reesei. This enzyme preparation is claimed to contain higher levels of β-
glucosidase activity than all other commercial cellulases available today, so as to
ensure almost complete conversion of cellobiose to glucose (http://www.genecor.com/
wps/wcm/coonect/genecor/products).
Genencor has also launched Accelerase® XY accessory xylanase enzyme complex
that enhances both xylan (C5) and glucan (C6) conversion when blended with other
Accelerase® enzyme products. Similarly, Accelerase® XC is an accessory
xylanase/cellulase enzyme complex that contains a broad profile of hemicellulase and
cellulase activities and enhances both xylan (C5) and glucan (C6) conversion when
blended with other Accelerase® enzyme products. Also, Accelerase® BG is an
accessory β-glucosidase enzyme that enhances glucan (C6) conversion when blended
with cellulase products. There are several potential cellulases which may prove to be
effective for biomass hydrolysis when supplemented with β-glucosidase, indicating
the importance of Accelerase® BG.
Novozymes also have a diverse range of cellulase preparations available based on
application as Cellusoft® AP and Cellusoft® CR for bioblasting in textile mills,
Carezyme® and Celluclean for laundry in detergent, Denimax® 601l for stonewash
industry at low temperature as well as many others specific for particular application
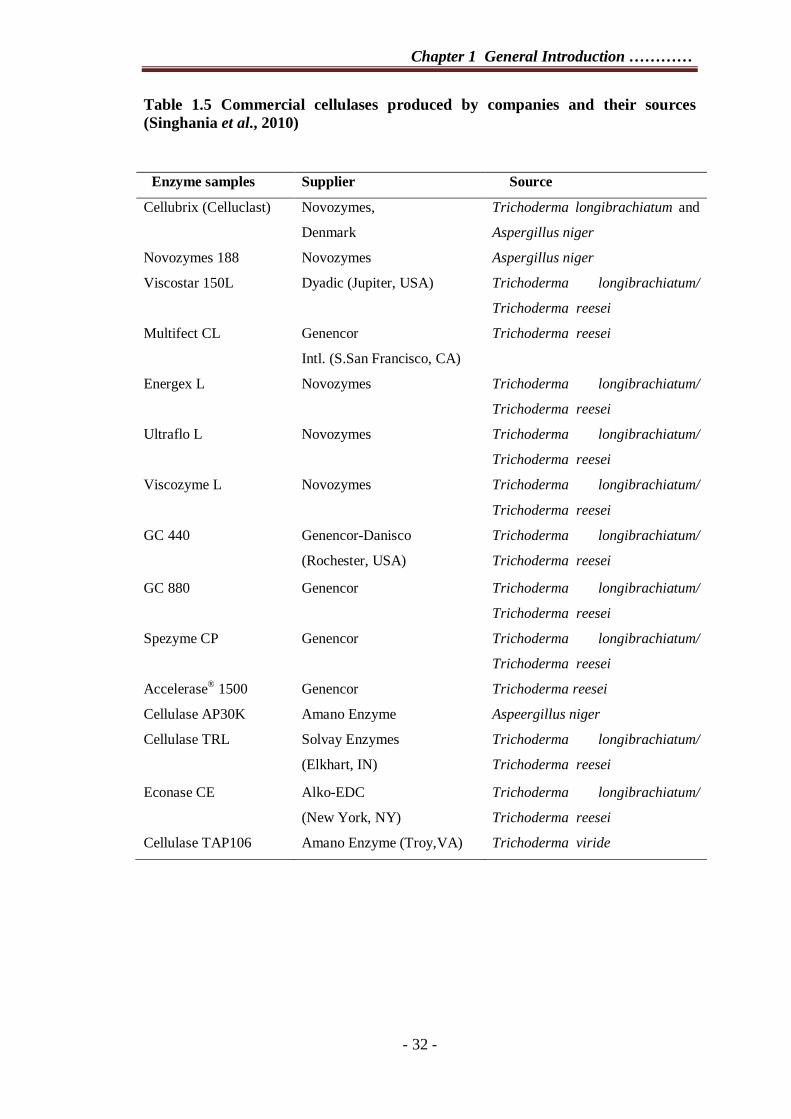
(http://www.bioenergy.novozymes.com). Amano Enzyme Inc. in Japan and MAP’s
India in India are another enzyme industry actively involved in cellulase production
(Table 1.5).
Chapter 1 General Introduction …………
- 32 -
Table 1.5 Commercial cellulases produced by companies and their sources (Singhania et al., 2010)
Enzyme samples Supplier Source Cellubrix (Celluclast) Novozymes,
Denmark Trichoderma longibrachiatum and
Aspergillus niger Novozymes 188 Novozymes Aspergillus niger Viscostar 150L Dyadic (Jupiter, USA) Trichoderma longibrachiatum/
Trichoderma reesei Multifect CL Genencor
Intl. (S.San Francisco, CA) Trichoderma reesei
Energex L Novozymes Trichoderma longibrachiatum/
Trichoderma reesei Ultraflo L Novozymes Trichoderma longibrachiatum/
Trichoderma reesei Viscozyme L Novozymes Trichoderma longibrachiatum/
Trichoderma reesei GC 440
Genencor-Danisco (Rochester, USA)
Trichoderma longibrachiatum/
Trichoderma reesei
GC 880 Genencor Trichoderma longibrachiatum/
Trichoderma reesei Spezyme CP Genencor Trichoderma longibrachiatum/
Trichoderma reesei Accelerase® 1500 Genencor Trichoderma reesei Cellulase AP30K Amano Enzyme Aspeergillus niger Cellulase TRL
Solvay Enzymes (Elkhart, IN)
Trichoderma longibrachiatum/
Trichoderma reesei
Econase CE Alko-EDC (New York, NY)
Trichoderma longibrachiatum/
Trichoderma reesei Cellulase TAP106 Amano Enzyme (Troy,VA) Trichoderma viride

Chapter 1 General Introduction …………
- 33 -
Problem delineated
Energy crisis and environmental pollution from the use of fossil fuels has become a
serious global threat. Thus, we need to move towards a sustainable development path
and explore the use of biofuel as cheaper and cleaner alternate to fossil fuels.
Bioethanol obtained from lignocellulosic materials has received major attention due to
their abundance and immense potential for conversion into sugars and fuels. Typical
bioprocesses for conversion of lignocellulose into biofuels involve production of
enzymes, pretreatment, hydrolysis of biomass and fermentation of sugars released.
Although bioethanol production has been greatly improved by new technologies,
there are still challenges that need further investigation. The major research
challenges are significant reduction in cost of enzyme production, improvement of
enzymatic hydrolysis with effective enzymes, development of novel technologies for
high solid handling during enzymatic hydrolysis and development of robust
fermentating organisms which are more tolerant to inhibitors and ferment all sugars
present in concentrated hydrolysate with high ethanol yields. Sincere efforts are also
required to adapt concepts of integration of the processes, reuse of enzymes and
appropriate technologies for the treatments of the solids and liquid wastes generated.
These efforts will significantly reduce energy and thereby cost of the fuel ethanol.
Thus, the present study was undertaken with the following research objectives:
1. Screening of potential cellulolytic microorganisms from various sources
2. Production of cellulases by solid state fermentation
3. Purification of endoglucanase and its biochemical characterization
4. Pretreatment and enzymatic hydrolysis of rice straw
5. Bioethanol production from enzymatic hydrolysate of rice straw
6. Biomethanation of waste generated during bioconversion of lignocellulosic
material

Chapter 1 General Introduction …………
- 34 -
References Acharya, S., and Chaudhury, A., 2011. Effect of nutritional and environmental
factors on cellulase activity by thermophillic bacteria isolated from hot spring. Journal of Scientific and Industrial Research, 70, 142-148.
Afzal, A., Bano, A., Fatima, M., 2010. Higher soybean yield by inoculation with N-
fixing and P-solubilizing bacteria. Agronomy Sustainable Development, 30, 487-495.
Akcapinar, G.B., Gul, O., Sezerman, U., 2011. Effect of codon optimization on the
expression of Trichoderma reesei endoglucanase 1 in Pichia pastoris. Biotechnology Progress, 27, 1257-1263.
Alper, H., and Stephanopoulos, G., 2009. Engineering for biofuels: exploiting innate
microbial capacity or importing biosynthetic potential? Nature Review in Microbiology, 7, 715-723.
Alvira, P., Tomas-Pejo, E., Ballesteros, M., Negro, M.J., 2010. Pretreatment
technologies for an efficient bioethanol production process based on enzymatic hydrolysis: a review. Bioresource Technology, 101, 4851-4861.
Andric, P., Meyer, A. S., Jensen, P. A., Johansen, K. D., 2010. Reactor design for
minimizing product inhibition during enzymatic lignocellulose hydrolysis I. Significance and mechanism of cellobiose and glucose inhibition on cellulolytic enzymes. Biotechnology in Advances, 28, 308-324.
Aro, N., Saloheimo, A., Ilmen, M., Penttila, M., 2001. ACEII, a novel transcriptional
activator involved in regulation of cellulase and xylanase genes of Trichoderma reesei. Journal of Biological Chemistry, 276, 24309-24314.
Ashabil, A., Lutfiye, K., Burhan, A., 2011. Alkaline thermostable and halophilic
endoglucanase from Bacillus licheniformis C108. African Journal of Biotechnology, 10, 789-796.
Bajaj, B.K., Pangotra, H., Wani, A.M., Sharma, P., Sharma, A., 2009. Partial
purification and characterization of a highly thermostable and pH stable endoglucanase from a newly isolated Bacillus strain M-9. Indian Journal of Chemical Technology, 16, 382-387.
Bakare, M.K., Adewale, I.O., Ajayi, A., Shonukan, O.O., 2005. Purification and
characterization of cellulase from the wild-type and two improved mutants of Pseudomonas fluorescens. African Journal of Biotechnology, 4, 898-904.
Balat, M., Balat, H., Oz, C., 2008. Progress in bioethanol processing. Progress in
Energy and Combustion Science, 4, 551-573.

Chapter 1 General Introduction …………
- 35 -
Bara, M.T.F., Lima, A.L., Ulhoa, C.J., 2003. Purification and characterization of an exo- β-1, 3-glucanase produced by Trichoderma asperellum. FEMS Microbiology Letters, 219, 81-85.
Bayer, E.A., Shimon, L.J.W., Shoham, Y., Lamed, R., 1998. Journal of Structural
Biology. 124, 221-234. Beg, Q.K., Kapoor, M., Mahajan, L., Hoondal, G.S., 2001. Microbial xylanases and
their industrial application: a review. Applied Microbiology and Biotechnology, 56, 326-338.
Berger, E., Zhang, D., Zverlow, V.V., Schwarz, W.H., 2007. Two noncellulosomal
cellulases of Clostridium thermocellum, Cel9I and Cel48Y, hydrolyse crystalline cellulose synergistically, FEMS Microbiology Letters, 268, 194-201.
Berlin, A., Maximenko, V., Gilkes, N., Saddler, J., 2007. Optimization of enzyme
complexes for lignocellulose hydrolysis. Biotechnology and Bioengineering, 97, 287-296.
Beukes, N., Chan, H., Doi, R. H., Pletschke, B.L., 2008. Synergistic associations
between Clostridium cellulovorans enzymes XynA, ManA and EngE against sugarcane bagasse. Enzyme and Microbial Technology, 42, 492-498.
Binod, P., Sindhu, R., Singhania, R.R., Vikram, S., Devi, L., Nagalakshimi, S.,
Kurien, N., Sukumaran, R.K., Pandey, A., 2010. Bioethanol production from rice straw: an overview, Bioresource Technology, 101, 4767-4774.
Bischoff, K.M., Rooney, A.P., Li, X.L., Liu, S., Hughes, S.R., 2006. Purification and
characterization of a family 5 endoglucanase from a moderately thermophilic strain of Bacillus licheniformis. Biotechnology Letters, 28, 1761-1765.
Bjerre, A.B., Olesen, A.B., Fernqvist, T., 1996. Pre-treatment of wheat straw using
combined wet oxidation and alkaline hydrolysis resulting in convertible cellulose and hemicellulose. Biotechnology and Bioengineering, 49, 568-577.
Boisset, C., Fraschini, C., Schulein, M., Henrissat, B., Chanzy, H., 2000. Imaging the
enzymatic digestion of bacterial cellulose ribbons reveals the endo character of the cellobiohydrolase Cel6A from Humicola insolens and its mode of synergy with cellobiohydrolase Cel7A, Applied Environmental Microbiology, 66, 1444-1452.
Bommarius, A.S., Katona, A., Cheben, S.E., Patel, A.S., Ragauskas, A.J., Knudson,
K., 2008. Cellulase kinetics as a function of cellulose pretreatment. Metabolic Engineering, 10, 370-381.
Borjesson, J., Peterson, R., Tjerneld, F., 2007. Enhanced enzymatic conversion of
softwood lignocellulose by polyethylene glycol addition. Enzyme and Microbial Technology, 40, 54-762.

Chapter 1 General Introduction …………
- 36 -
Cadoche, L., and Loopez, G.D., 1989. Assessment of size reduction as a preliminary step in the production of ethanol from lignocellulosic wastes. Biological Wastes, 30, 153-157.
Cardona, C.A., and Sanchez, O.J., 2008. Trends in biotechnological production of
fuel ethanol from different feedstocks. Bioresource Technology, 99, 5270-5295.
Cen, P.L., and Xia, L.M., 1999. Production of cellulase by solid state fermentation.
Advances in Biochemical Engineering and Biotechnology, 65, 69-92. Chahal, D.S., 1985. Solid-state fermentation with Trichoderma reesei for cellulase
production. Applied Environmental Microbiology, 49, 205-210. Chandel, A.K., Chandrasekhar, G., Silva, M.B., Silverio, D., Silva, S., 2012. The
realm of cellulases in biorefinery development. Critical Reviews in Biotechnology, 32, 187-202.
Chandel, A.K., Kapoor, R.K., Singh, A., Kuhad, R.C., 2007. Detoxification of
sugarcane bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501. Bioresource Technology, 98, 1947-1950.
Chandrakant, P., and Bisaria, V.S., 1998. Simultaneous bioconversion of cellulose
and hemicellulose to ethanol. Critical Review of Biotechnology, 18, 295-331. Chen, G., Li, S., Jiao, F., Yuan, Q., 2007. Catalytic dehydration of bioethanol to
ethylene over TiO2/γ-Al2O3 catalysts in micro channel reactors. Catalyst, 125, 111-119.
Chen, Y., 2011. Development and application of co-culture for ethanol production by
co-fermentation of glucose and xylose: a systematic review. Journal of Indian Microbiology and Biotechnology, 38, 581-597.
Cherry, J.R., Fidantsef, A.L., Cherry, Fidantsef, 2003. Directed evolution of industrial
enzymes: an update. Current Opinion in Biotechnology, 14, 438-443. Chundawat, S.P., Venkatesh, B., Dale, B.E., 2007. Effect of particle size based
separation of milled corn stover on AFEX pretreatment and enzymatic digestibility. Biotechnology and Bioengineering, 96, 219-31.
Dariot, D.J., Simonetti, A., Plinho, F., Brandeli, A., 2008. Purification and
characterization of extracellular β-glucosidase from Monascus purpureus. Journal of Microbial Biotechnology, 18, 933-941.
Dijkerman, R., Op den Camp, H.J., Van der Drift, C., Vogels, G.D., 1997. The role of
the cellulolytic high molecular mass (HMM) complex of the anaerobic fungus Piromyces sp. strain E2 in the hydrolysis of microcrystalline cellulose. Archives of Microbiology, 167, 137-142.

Chapter 1 General Introduction …………
- 37 -
Dipali, P., Seeta, L. R., Mala, R., 2012. Purification and biochemical characterization of endoglucanase from Penicillium pinophilum MS 20. Indian Journal of Biochemistry and Biophysics, 49, 189-194.
Divne, C., Stahlberg, J., Teeri, T.T., Jones, T.A., 1998. High-resolution crystal
structures reveal how a cellulose chain is bound in the 50 Å long tunnel of cellobiohydrolase I from Trichoderma reesei. Journal of Molecular Biology, 275, 309-325.
Duff, S.J.B., and Murray, W.D., 1996. Bioconversion of forest products industry
waste cellulosic to fuel ethanol: a review. Bioresource Technology, 55, 1-33. Eggeman, T., and Elander, R.T., 2005. Process and economic analysis of pretreatment
technologies. Bioresource Technology, 96, 2019-2025. Elkins, J.G., Raman, B., Keller, M., 2010. Engineered microbial systems for enhanced
conversion of lignocellulosic biomass. Current Opinion in Biotechnology, 21, 657-662.
Elshafei, A.M., Mohamed, M.H., Bakry, M.H., Osama, M.A., Housam, M.A.,
Abdelmageed, M.O. 2009. Purification and properties of an endoglucanase of Aspergillus terreus DSM 826. Journal of Basic Microbiology, 426-432.
Eriksson, K.E.L., and Bermek, H., 2009. Lignin, lignocellulose, ligninase. Applied
Microbiology, 373-384. Ferreira, S.M.P., Duarte, A.P., Queiroz, J.A., Domingues, F.C., 2009. Influence of
buffer systems on Trichoderma reesei Rut C-30 morphology and cellulase production. Electronic Journal of Biotechnology, 12, 1-9.
Fritscher, C., Messner, R., Kubicek, C.P., 1990. Cellobiose metabolism and
cellobiohydrolase I biosynthesis by Trichoderma reesei. Experimental Mycology, 14, 405-415.
Gao, J., Weng, H., Zhu, D., Yuan, M., Guan, F., Y. Xi, 2008. Production and
characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresource Technology, 99, 7623-7629.
George, P.S., Ahmad, A., Rao, M.B., 2001. Studies on carboxy-methyl cellulase
produced by an alkalothermophilic actinomycete. Bioresource Technology, 77, 171-175.
Gourari, S., and Achkari-Begdouri, A., 1997. Use of baked clay media as biomass
supports for anaerobic filters. Applied Clay Science, 12, 365-375. Gupta, R., Sharma, K.K., Kuhad, R.C., 2009. Separate hydrolysis and fermentation
(SHF) of Prosopis juliflora, a woody substrate for the production of cellulosic ethanol by Saccharomyces cerevisiae and Pichia stipitis-NCIM 3498.

Chapter 1 General Introduction …………
- 38 -
Gusakov, A.V., Salanovich, T.N., Antonov, A.I., Ustinov, B.B., Okunev, O.N., Burlingame, R., 2007. Design of highly efficient cellulase mixtures for enzymatic hydrolysis of cellulose. Biotechnology and Bioenergy, 97, 1028-1038.
Hamelinck, C.N., Hooijdonk, G.V., Faaij. A.P., 2005. Ethanol from lignocellulosic
biomass: techno-economic performance in short-middle-and long-term. Biomass and Bioenergy, 28,384-410.
Heck, J.X., Hertz, P.F., Ayub, M.A.Z., 2002. Cellulase and xylanase production by
isolated Amazon Bacillus strains using soya been industrial residue based solid-state cultivation. Brazilian Journal of Microbiology, 33, 213-218.
Heinelman, P., Snow, C.D., Wu, I., Nguyen, C., Villalobos, A., 2009. A family of
thermostable fungal cellulases created by structure-guided recombination. PNAS, 106, 5451-5452. DOI:10.1073/pnas.0901417106.
Hendriks, A.T.W.M., and Zeeman, G., 2009. Pretreatments to enhance the
digestibility of lignocellulosic biomass. Bioresource Technology, 100, 10-18. Henrissat, B., 1991. A classification of glycosyl hydrolases based on amino acid
sequence similarities. Biochemical Journal, 280, 309-316.1991. Hodge, D.B., Karim, M.N., Schell, D.J., McMillan, J.D., 2009. Model-based fed-
batch for high-solid enzymatic cellulose hydrolysis. Applied Biochemistry and Biotechnology, 152, 88-107.
Holker, U., Hofer, M., Lenz, J., 2004. Biotechnological advantages of laboratory-
scale solid-state fermentation with fungi. Applied Microbiology and Biotechnology, 64, 175-186.
Hoshino, E., Shiroishi, M., Amano, Y., Nomura, M., Kanda, T., 1997. Synergistic
action of exo-type cellulases in the hydrolysis of cellulose with different crystallinities. Journal of Fermentation and Bioengineering, 84, 300-306.
Ilmen, M., Thrane, C., Penttila, M., 1996a. The glucose repressor gene cre 1 of
Trichoderma; isolation and expression of a full-length and a truncated mutant form. Molecular and General Genetics, 251, 451-460.
Ilmen, Onnela, M.L., Klemsadals, Keranans, Pentilla, M., 1996b. Functional analysis
of the cellobiohydrolase I promoter of the filamentous fungus. Trichoderma reesei. Molecular and General Genetics, 253, 303-314.
Ilmen, M., Saloheimo, A., Onnela, M.L., Penttila, M.E., 1997. Regulation of
cellulase gene expression in the filamentous fungus Trichoderma reesei. Applied Environmental Microbiology, 63, 1298-1306.
Jeris, J.S., 1982. Industrial wastewater treatment using anaerobic fluidised bed
reactors. Proceedings of IAWPRC Seminar on Anaerobic Treatment, Copenhagen, Denmark.

Chapter 1 General Introduction …………
- 39 -
Jha, P.K., Nair, S., Gopinathan, M.C, Babu. C.R., 1995. Suitability of rhizobia inoculated wild legumes Argurolobium flaccidum, Astragalus graveolens, Indigofera gangetica and Lespedeza stenocarpa in providing a vegetational cover in unreclaimed limestone quarry. Plant Soil, 177, 139-149.
Jorgensen, H., Kristensen, J.B., Felby, C., 2007. Enzymatic conversion of
lignocellulose into fermentable sugars: Challenges and opportunities, Biofuels, Bioproducts and Biorefining 1, 119-134.
Jorgensen, H., Vibe-Pedersen, J., Larsen, J., Felby, C., 2009. Liquefaction of
lignocellulose at high-solid concentrations. Biotechnology and Bioengineering, 96, 862-870.
Jung, H., Yoon, H.G., Park, W., Choi, C., Wilson, D.B., Shin, D.H., 2008. Effect of
sodium hydroxide treatment of bacterial cellulose on cellulase activity. Cellulose, 15, 465-471.
Jung, J.H., Fouad, W.M., Vermerris, W., Gallo, M., Altpeter, F., 2012. RNAi
suppression of lignin biosynthesis in sugarcane reduces recalcitrance for biofuel production from lignocellulosic biomass. Plant Biotechnology Journal, 10, 1067-1076.
Kabel, M.A., Van der Maarel, M.J.E.C., Klip, G., Voragen, A.G.J., Schols, H.A.,
2005. Standard assays do not predict the efficiency of commercial cellulase preparations towards plant materials. Biotechnology and Bioengineering, 93, 56-63.
Kang, H.J., Uegaki, K., Fukada, H., Ishikawa, K., 2007. Improvement of the
enzymatic activity of the hyperthermophilic cellulase from Pyrococcus horikoshi. Extremophiles, 11, 251- 256.
Kaur, J., Chadha, B.S., Kumar, B.A., Saini, H.S., 2007. Purification and
characterization of two endoglucanases from Melanocarpus sp. MTCC 3922. Bioresource Technology, 98, 74-81.
Kim, J.Y., Hur, S.H., Hong, J.H., 2005. Purification and characterization of an
alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-810. Biotechnology Letters, 27,313- 316.
Koukiekolo, R., Kosugi, A., Inui, M., Yukawa, H., Doi, R.H., 2005. Degradation of
corn fiber by Clostridium cellulovorans cellulases and hemicellulases and contribution of scaffolding protein CbpA. Applied Environmental Microbiology, 71, 3504-3511.
Kristensen, J.B., Felby, C., Jorgensen, H., 2009. Yield-determining factors in high-
solids enzymatic hydrolysis of lignocellulose, Biotechnology and Biofuels, 2. Kubicek, C.P., 1981. Release of carboxy-methyl cellulase and β-glucosidase from the
cell walls of Trichoderma reesei. European Journal of Applied Microbiology and Biotechnology, 13, 226-231.

Chapter 1 General Introduction …………
- 40 -
Kubicek, C.P., and Penttila, M., 1998. Regulation of production of plant polysaccharide degrading enzymes by Trichoderma reesei in Trichoderma and Gliocladium, Volume 2, edition by GE Harman and CP Kubicek (Taylor and Francis, London), 49-72.
Kuhad, R.C., Gupta, R., Khasa, Y.P., Singh, A., 2010. Bioethanol production from
Lantanacamara (red sage): pretreatment, saccharification and fermentation. Bioresource Technology, 101, 8348-8354.
Kumar, G. S, Gupta, S. K., Singh G, 2007. Anaerobic hybrid reactor - a promising
technology for the treatment of distillery spent wash, Journal of Indian School of Mines, 11, 1 25-138.
Kumar, R., and Wyman, C.E., 2009. A Cellulase adsorption and relationship to
features of corn stover solids produced by leading pretreatments. Biotechnology and Bioengineering.103, 252-67.
Larsen, J., Petersen, M.O., Thirup, L., Li, H.W., Iversen, F.K., 2008. The IBUS
process -Lignocellulosic bioethanol close to a commercial reality. Chemical Engineering Technology, 31, 765-772.
Laser, M., Larson, E., Dale, B., Wang, M., Greene, N., Lynd, L.R., 2009.
Comparative analysis of efficiency, environmental impact and process economics for mature biomass refining scenarios Biofuels, Bioproducts and Biorefining, 3, 247-270.
Lettinga, G., van Velsen, A.F.M., Hobma, S.W., de Zeeuw, W.J., Klapwijk, A., 1980.
Use of the upflow sludge blanket (USAB) reactor. Biotechnology and Bioengineering, 22, 699-734.
Linger, J.G., Adney, W.S., Darzins, A., 2010. Heterologous expression and
extracellular secretion of cellulolytic enzymes by Zymomonas mobilis. Applied Environmental Microbiology, 76, 6360-6369.
Liu, D., Zhang, R., Yang, X., Zhang, Z., Song, S., Miao, Y., 2012. Characterization of
a thermo-stable β-glucosidase from Aspergillus fumigatus Z5, and its functional expression in Pichia pastoris X33. Microbial Cell, 11:25.
Maxham, J.V., and Wakamiya, W., 1981. Innovative biological wastewater treatment
technologies applied to the treatment of biomass gasification wastewater. In. Proceedings of the 35th Ind. Waste Conference, Purdue University, 80-94.
Mamma, D., Christakopoulos, P., Koullas, D., Kekos, D., MaMammacris, B.J.,
Koukios, E., 1995. An alternative approach to the bioconversion of sweet sorghum carbohydrates to ethanol. Biomass and Bioenergy, 8, 99-103.
Mandels, M., and Weber, J., 1969. Production of cellulases. Advances in Chemistry
Series, 95, 391-414.

Chapter 1 General Introduction …………
- 41 -
Mandels, M., and Reese, E.T., 1957. Induction of cellulase in Trichoderma viride as influenced by carbon sources and metals. Journal of Bacteriology, 73, 269-278.
Mansfield, S.D., Mooney, C., Saddler, J.N., 1999. Substrate and enzyme
characteristics that limit cellulose hydrolysis. Biotechnology Programmes, 15, 804-816.
Mattanovich, D., Branduardi, P., Dato, L., Gasser, B., Sauer, M., Porro, D., 2012.
Recombinant protein production in yeasts recombinant gene expression. Methods in Molecular Biology, 824, 329-358.
McMillan, J.D., 1997. Biethanol production: Status and prospects. Renewable
Energy, 10, 295,-302. Melidis, P., Georgiou, D., Aivasidis, A., 2003. Scale-up and design optimization of
anaerobic immobilized cell reactors for wastewater treatment. Chemical Engineering and Processing, 42, 897-908.
Michaud, S., Bernet, N., Buffiere, P., Roustan, M., and Moletta, R., 2002. Methane
yield as a monitoring parameter for the start-up of anaerobic fixed film reactors. Water Research, 36, 1385 -1391.
Modenbach, A. A., and Nokes, S.E., 2012. The use of high solids loadings in biomass
pretreatment- a review. Biotechnology and Bioengineering, 109, 1430. Modenbach, A. A., and Nokes, S.E., 2013. Enzymatic hydrolysis of biomass at high
solids loadings- a review. Biomass and Bioenergy, 56, 526-544. Moniruzza, A.M., Dien, B.S., Skory, C.D., Chen, Z.D., Hespell, R.B., Ho, N.W.Y.,
Dale, B.E., Bothast R.J., 1997. Fermentation of corn fibre sugars by an engineered xylose utilizing Saccharomyces yeast strain. World Journal of Microbiology and Biotechnology, 341-346.
Mosier, N., Wyman, C., Dale, B., Elander, R., Lee, Y., Holtzapple, M., Ladisch, M.,
2005. Features of promosing technologies for pretreatment of lignocellulosic biomass. Bioresource Technology, 96, 673-686.
Murashima, K., Kosugi, A., Doi, R., 2003. Synergistic effects of cellulosomal
xylanase and cellulases from Clostridium cellulovorans on plant cell wall degradation. Journal of Bacteriology, 185, 1518-1524.
Nazir, A., Soni, H., Saini, H.S., Manhas, R.K., Chadha, B.S., 2009. Purification and
characterization of an endoglucanase from Aspergillus terreus highly active against barley β-glucan and xyloglucan, World Journal of Microbial Biotechnology, 25, 1189-1197.
Nguyen, V.T., and Quyen, D.T., 2010. Purification and properties of a novel
thermoactive endoglucanase from Aspergillus awamori VTCC-F099. Australian Journal of Basic Applied Sciences, 4, 6211-6216.

Chapter 1 General Introduction …………
- 42 -
Nidetzky, B., Steiner, W., Hayn, M., Claeyssens, M., 1994. Cellulose hydrolysis by the cellulases from Trichoderma reesei: a new model for synergistic interaction. Biochemical Journal, 3, 705-710.
Nigam, P., and Singh, D., 1996. Processing of agriculture wastes in solid state
fermentation for cellulolytic enzyme production. Journal of Scientific and Industrial Research, 55, 457-463.
Noronha, E.F., Kipnis, A., Kipinis, A.P.J., Ulhoa, C.J., 2000. Regulation of 36 kDa 1-
3 glucanase synthesis in Trichoderma harzianum. FEMS Microbiology Letters, 188, 19-22.
Ogawa, S., Lozach, J., Benner, C., Pascual, G., Tangirala, R.K., Westin, S.,
Hoffmann, Subramaniam, A., David, M., Rosenfeld, M.G., 2005. Molecular determinants of crosstalk between nuclear receptors and toll-like receptors. Cell, 122, 707-721.
Olsson, P.A., Baath, E., Jakobsen, I., Soderstrom, B., 1996. Soil bacteria respond to
presence of roots but not to mycelium of Arbuscular mycorrhizal fungi. Soil Biology and Biochemistry, 28, 463-470.
Pandey, A., 1994. Solid state fermentation-an overview. In: Pandey, A. (Ed), Solid
state fermentation. Wiley Eastern Limited, New Delhi, 3-10. Pandey, A., Benjamin, S., Soccol, C.R., Nigam, P., Krieger, N., Soccol, U.T., 1999.
The realm of microbial lipases in biotechnology. Biotechnology and Applied Biochemistry, 29, 119-131.
Park, Y.H., Kim, J., Kim, S.S., Park, Y.K., 2009. Pyrolysis characteristics and kinetics
of oak trees using thermo gravimetric analyzer and micro-tubing reactor. Bioresource Technology, 100, 400-405.
Pottkamper, J., Barthen, P., Ilmberger, N., Schwaneberg, U., Schenk, A., Schulte, M.,
Ignatiev, N., Streit. W. R., 2009. Applying metagenomics for the identification of bacterial cellulases that are stable in ionic liquids. Green Chemistry, 11, 957-965.
Ramos, L.P., Nazhad, M.M., Saddler, J.N., 1993. Effect of enzymatic hydrolysis on
the morphology and fine structure of pretreated cellulosic residues. Enzyme and Microbial Technology, 15, 821-831.
Raven, P.H., Evert, R.F., Eichhorn, S.E., 1999. Biology of plants. New York; W.H.
Freeman and company. Reczey, K., Szengyel, Z.S., Eklund, R., Zacchi, G., 1996. Cellulase production by
Trichoderma reesei. Bioresource Technology, 57, 25-30. Reyes, O., Sanchez, E., Rovirosa, N., Borja, R., Cruz, M., Colmenarejo, M.F.,
Escobedo, R., Ruiz, M., Rodriguez, X., Cprrea, G., 1999. Low strength wastewater treatment by a multistage anaerobic filter packed with waste tyre rubber. Bioresource Technology, 70, 55-60.

Chapter 1 General Introduction …………
- 43 -
Romero, M.D., Aguado, J., González, L., Ladero, M., 1999. Cellulase production by Neurospora crassa on wheat straw. Enzyme and Microbial Technology, 25, 244-250.
Rosgaard, L., Pedersen, S., Meyer, A.S., 2007. Comparison of different pretreatment
strategies or enzymatic hydrolysis of wheat and barley straw. Applied Biochemistry and Biotechnology, 143, 284-296.
Rovirosa, N., Sanchez, E., Cruz, M., Veiga, M.C., Borja, R., 2004. Coliform
concentration reduction and related performance evaluation of a down-flow anaerobic fixed bed reactor treating low-strength saline wastewater. Bioresource Technology, 94, 119-127.
Saha, B.C., 2004. Production, purification and properties of endoglucanase from a
newly isolated strain of Mucor circinelloides. Process Biochemistry, 39, 1871- 1876.
Saha, B.C., 2003. Hemicellulose bioconversion, Journal of Industrial Biotechnology,
30, 279- 291. Sandgren, M., Stahlberg, J., Mitchinson, C., 2005. Structural and biochemical studies
of GH family 12 cellulases: improved thermal stability and ligand complexes. Progress in Biophysics and Molecular Biology, 89, 246-291.
Sa-Pereira, P., Paveia, H., Costa-Ferreira, M., Aires-Barros, M.R., 2003. A new look
at xylanases: An overview of purification strategies. Molecular Biotechnology, 24, 257-281.
Schulein, M., 2000. Protein engineering of cellulases. Biochemica et Biophysica Acta,
1543, 239-252. Schwarz, W.H., 2001. The cellulosome and cellulose degradation by anaerobic
bacteria. Applied Microbiology and Biotechnology, 56, 634-649. Selig, M., Weiss, N., Ji, Y., 2008. NREL laboratory analytical procedure: Enzymatic
saccharification of lignocellulosic biomass, National Renewable Energy Laboratory Retrieved from http://www.nrel.gov/biomass/pdfs/42629.pdf.
Shen, B., Sun, X., Zuo, X., Shilling, T., Apgar, J., Ross, M., 2012. Engineering a
thermo regulated inter-modified xylanase into maize for consolidated lignocellulosic biomass processing. Nature Biotechnology, 30 1131-1136.
Show, K.Y., and Tay, J.H., 1999. Influence of support media on biomass growth and
retention in anaerobic filters, Water Research, 33, 1471-1481. Singh, J., Batra, N., Sobti, R.C., 2004. Purification and characterization of alkaline
cellulose produced by a novel isolate Bacillus sphaericus JS1. Journal of Industrial Microbiology and Biotechnology, 31, 51-56.

Chapter 1 General Introduction …………
- 44 -
Singh, S., Simmons, B.A., Vogel, K.P., 2009. Visualization of biomass solubilization and cellulose regeneration during ionic liquid pretreatment of switch grass, Biotechnology and Bioengineering, 104, 68-75.
Singhania, R.R., Patel, A.K., Soccol, C.R., and Pandey, A., 2009. Recent advances in
solid state fermentation, Biochemical Engineering Journal, 44, 13-18. Singhania, R.R., Sukamaran, R.K., Pandey, A., 2007. Improved cellulose production
by Trichoderma reesei RUT C30 under SSF through process optimization. Applied Biochemistry and Biotechnology, 142, 60-70.
Singhania, R.R., Sukumaran, R.K., Patel, A.K., Larroche, C., Pandey, A., 2010.
Advancement and comparative profiles in the production technologies using solid state and submerged fermentation for microbial cellulases, Enzyme and Microbial Technology, 46, 541-549.
Sternberg, D., and Mandels, G.R., 1979. Induction of cellulolytic enzymes in
Trichoderma reesei by sophorose. Journal of Bacteriology, 139, 761-769. Steyer, J.P., Bouvier J.C., Conte, T., Gras, P., Sousbie, P., 2002. Evaluation of a four
year experience with a fully instrumented anaerobic digestion process, Water Science Technology, 45, 495-502.
Sues, A., Millati, R., Edebo, L., Taherzadeh, M. J., 2005. Ethanol production from
hexoses, pentoses, and dilute-acid hydrolyzate by Mucor indicus. FEMS Yeast Research, 5, 669-676.
Sukumaran, R.K., Singhania, R.R., Mathew, G.M., Pandey, A., 2009. Cellulase
production using biomass feedstock and its application in lignocellulose saccharification of bio-ethanol production, Renewable Energy, 34, 421-424.
Sukumaran, R.K., Singhania, R.R., Pandey, A., 2005. Microbial cellulases production,
applications and challenges. Journal of Scientific and Industrial Research 64, 832 - 844.
Sun, Y., and Cheng, J.Y., 2002. Hydrolysis of lignocellulosic materials for ethanol
production: a review. Bioresource Technology, 83, 1-11. Switzen Baum, M.S., and Jewel, W.J., 1980. Anaerobic attached film expanded bed
reactor treatment. Journal of Water Pollution Control Federation, 52. Szabo, I.J., Johansson, G., Pettersson, G., 1996. Optimized cellulase production by
Phanerochaete chrysosporium: Control of catabolite repression by fed-batch cultivation. Journal of Biotechnology, 48, 221-230.
Tait, S.L., and Friedman, A.A., 1980. Anaerobic rotating biological contractor for
carbonaceous wastewater. Journal of Water Pollution Control Federation, 52, 2257-2269.

Chapter 1 General Introduction …………
- 45 -
Taherzadeh, M.J., and Kamiri, K., 2008. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production. A review. International Journal of Molecular Science, 9, 1621-1651.
Tangnu, S.K., Blanch, H.W., Wilke, C.R., 1981. Enhanced production of cellulase,
hemicellulase, and β-glucosidase by Trichoderma reesei (RUT C-30), Biotechnology and Bioengineering, 23, 1837-1849.
Tengerdy, R.P., 1996. Cellulase production by solid state fermentation. Journal of
Scientific and Industrial Research, 55, 313-316. Thai, H.P., Dinh, T.Q., Ngoc, M.N., 2010. Optimization of endoglucanase production
by Aspergillus niger VTCC-F021. Australian Journal of Basic and Applied Sciences, 4, 4151-4157.
Tjerneld, F., 1994. Enzyme-catalyzed hydrolysis and recycling in cellulose
bioconversion. Methods in Enzymology. G. J. Harry Walter, Academic Press, 228, 549-558.
Tu, M., Zhang, X., Paice, M., MacFarlane, P., Saddler, J.N., 2009. The potential of
enzyme recycling during the hydrolysis of a mixed softwood feedstock. Bioresource Technology, 100, 6407-6415.
Vaheri, M.P., Vaheri, M.E.O., Kauppinen, V.S., 1979. Formation and release of
cellulolytic enzymes during growth of Trichoderma reesei on cellobiose and glycerol, European Journal of Applied Microbiology and Biotechnology, 8, 73-80.
van den Berg, L., Kennedy, K.J., Samson, R., 1985. Anaerobic down flow stationary
fixed film reactors: Performance under steady-state and non-steady state conditions. Water Science and Technology, 17, 89-102.
Van Dyk, J.S., Pletschke, B.I., 2012. A review of lignocellulose bioconversion using
enzymatic hydrolysis and synergistic cooperation between enzymes-factors affecting enzymes, conversion and synergy. Biotechnology Advances, 30, 1458-1480.
Varnai, A., Siika-aho, M., Viikari, L., 2010. Restriction of the enzymatic hydrolysis
of steam pretreated spruce by lignin and hemicellulose. Enzyme and Microbial Technology, 46, 185-193.
Vikari, L., Alapuranen, M., Puranen, T., Vehmannpera, J., Silka-Aho, M., 2007.
Thermostable enzymes in lignocellulosic hydrolysis. Advances in Biochemical Engineering and Biotechnology, 108, 121-145.
Vincken, J.P., Beldman, G., Voragen, A.G.J., 1994. The effect of xylo glucans on the
degradation of cell-wall embedded cellulose by the combined action of cellobiohydrolase and glucanases from Trichoderma viride. Plant Physiology, 104, 99-107.

Chapter 1 General Introduction …………
- 46 -
Vintila, T., Dragomirescu, M., Jurcoane, S., Caprita, R., Maiu, M., 2009. Production of cellulase by submerged and solid-state cultures and yeast selection for conversion of lignocellulose to ethanol. Romanian Biotechnological Letters, 14, 4275-4281.
Walker, L.P., and Wilson, D.B., 1991. Enzymatic hydrolysis of cellulose: an
overview. Bioresource Technology, 36, 3-14. Wati, L., Kumari, S., Kundu, B.S., 2007. Paddy straw as a substrate for ethanol
production. Indian Journal of Microbiology, 47, 26-29. Wei, D.L., Kohtaro, K., hoji, U.S., Hui, L.T., 1996. Purification and characterization
of an extracellular β-glucosidase from the wood-grown fungus Xylaria regalis. Current Microbiology, 33, 297-301.
Weiss, N., Borjesson, J., Pedersen, L.S., and Meyer, A. S., 2013. Enzymatic
lignocellulose hydrolysis: Improved cellulase productivity by insoluble solids recycling. Biotechnology and Biofuels, 6, 5.
Wen, Z., Liao, W., Chen, S., 2005. Production of cellulase/β-glucosidase by the
mixed fungi culture of Trichoderma reesei and Aspergillus phoenicis on dairy manure. Applied Biochemistry and Biotechnology, 121, 93-104.
Wurleitner, E., L. Pera, C., Wacenovsky, A., Cziferszky, S., Zeilinger, C. P., Kubicek,
R. L., 2003. Transcriptional regulation of xyn2 in Hypocrea jecorina. Eukaryotic Cell, 2,150-158.
Xin, Z., Yinbo, Q., Peiji, G., 1993. Acceleration of ethanol production from paper
mill waste fiber by supplementation with b-glucosidase. Enzyme and Microbial Technology. 15, 62-65.
Xu, Q., Adney, W.S., Ding, S.Y., Himmel, M.E., 2007. Cellulase for biomass
conversion. Industrial enzymes, 1st.ed. National Bioenergy Center, National Renewable Energy Laboratory, USA, 35-50.
Yanase, S., Hasunuma, T., Yamada, R., Tanaka, T., Ogino, C., Fukuda, H., Kondo,
A., 2010. Direct ethanol production from cellulosic materials at high temperature using the thermo tolerant yeast Kluyveromyces marxianus displaying cellulolytic enzymes. Applied Microbiology and Biotechnology, 88, 381-388.
Yang, B., and Wyman C.E., 2008. Characterization of the degree of polymerization of
xylooligomers produced by flow through hydrolysis of pure xylan and corn stover with water. Bioresource Technology, 99, 5756-5762.
Yang, J., Zhang, X., Yong, Q., Yu, S., 2010. Three-stage hydrolysis to enhance
enzymatic saccharification of steam-exploded corn stover. Bioresource Technology, 101, 4930-4935.

Chapter 1 General Introduction …………
- 47 -
Yang, S., Jiang, Z., Yan, Q., Zhu, H., 2008. Characterization of a thermostable extracellular a β- glucosdiase with activities of exoglucanse and trans glycosylation from Paecilomyces thermophila. Journal of Agricultural Food Chemistry, 56, 602-608.
Yang, S.F., Tay, J.H., Liu., 2004. Inhibition of free ammonia to the formation of
aerobic granules. Biochemical Engineering Journal, 17, 41-48. Yin, L.J., Huang, P.S., Lin, H.H., 2010. Isolation of cellulase-producing bacteria and
characterization of the cellulase from the isolated bacterium Cellulomonas sp.YJ5. Journal of Agricultural Food Chemistry, 58, 9833-9837.
Young, J.C., and McCarty, P.L., 1969. The anaerobic filter for waste treatment.
Journal of Water Pollution Control Federation, 41, 160-173. Zhang, H.Y., and Lynd, L.R., 2004. Toward an aggregated understanding of
enzymatic hydrolysis of cellulose: Non-complexed cellulase systems. Biotechnology and Bioengineering, 88, 797-824.
Zhang, S., and Lv, X., 2006. Ionic liquids-from basic research to application in
industry. Science Press, ISBN 7-03-016882-8, Beijing. Zhu, Z., Sathitsuksanoh, N., Vinzant, T., Schell, D.J., McMillan, J.D., Zhang, Y.H.P.,
2009. Comparative study of corn stover pretreated by dilute acid and cellulose solvent-based lignocellulose fractionation: enzymatic hydrolysis, supra-molecular structure and substrate accessibility. Biotechnology and Bioengineering, 103, 715-724.


















