
1, 2, 3 1, 7 1 2 3 4 5 6 - Journal of Clinical...
32
1 Identification and subtyping of clinically relevant human and ruminant mycoplasmas using 1 matrix-assisted laser desorption ionization-time of flight mass spectrometry 2 3 S. Pereyre 1, 2, 3 * # , F. Tardy 4, 5 * # , H. Renaudin 1, 2, 3 , E. Cauvin 6 , L. Del Prá Netto Machado 1, 7 , 4 A. Tricot 5, 4 , F. Benoit 6 , M. Treilles 6Δ , C. Bébéar 1, 2, 3 . 5 6 1 Univ. Bordeaux, USC EA371 Infections humaines à mycoplasmes et chlamydiae, 33076 Bordeaux, 7 France. 8 2 INRA, USC Infections humaines à mycoplasmes et chlamydiae, 33076 Bordeaux, France. 9 3 Centre Hospitalier Universitaire de Bordeaux, Laboratoire de Bactériologie, 33076 Bordeaux, 10 France. 11 4 Anses, Laboratoire de Lyon, UMR Mycoplasmoses des Ruminants, 69364 Lyon Cedex 07, France. 12 5 Université de Lyon, VetAgro Sup, UMR Mycoplasmoses des Ruminants, 69280 Marcy L'Etoile, 13 France. 14 6 Laboratoire Départemental de la Manche, Service Santé Animale, 1352 avenue de Paris - 50008 15 Saint-Lô Cedex, France. 16 7 Programa de Pós Graduação em Farmácia, Universidade Federal de Santa Catarina, Florianópolis, 17 Brazil. 18 *These authors contributed equally to this work 19 ΔCurrent address: Laboratoire d’Analyses de Sèvres Atlantique, 210 avenue de la Venise Verte 20 79000 Niort, France 21 22 #Corresponding authors: 23 Copyright © 2013, American Society for Microbiology. All Rights Reserved. J. Clin. Microbiol. doi:10.1128/JCM.01573-13 JCM Accepts, published online ahead of print on 31 July 2013 on September 1, 2018 by guest http://jcm.asm.org/ Downloaded from
Transcript of 1, 2, 3 1, 7 1 2 3 4 5 6 - Journal of Clinical...

1
Identification and subtyping of clinically relevant human and ruminant mycoplasmas using 1
matrix-assisted laser desorption ionization-time of flight mass spectrometry 2
3
S. Pereyre1, 2, 3*#, F. Tardy4, 5*#, H. Renaudin1, 2, 3, E. Cauvin6, L. Del Prá Netto Machado1, 7, 4
A. Tricot5, 4, F. Benoit6, M. Treilles6Δ, C. Bébéar1, 2, 3. 5
6
1Univ. Bordeaux, USC EA371 Infections humaines à mycoplasmes et chlamydiae, 33076 Bordeaux, 7
France. 8
2INRA, USC Infections humaines à mycoplasmes et chlamydiae, 33076 Bordeaux, France. 9
3Centre Hospitalier Universitaire de Bordeaux, Laboratoire de Bactériologie, 33076 Bordeaux, 10
France. 11
4Anses, Laboratoire de Lyon, UMR Mycoplasmoses des Ruminants, 69364 Lyon Cedex 07, France. 12
5Université de Lyon, VetAgro Sup, UMR Mycoplasmoses des Ruminants, 69280 Marcy L'Etoile, 13
France. 14
6Laboratoire Départemental de la Manche, Service Santé Animale, 1352 avenue de Paris - 50008 15
Saint-Lô Cedex, France. 16
7Programa de Pós Graduação em Farmácia, Universidade Federal de Santa Catarina, Florianópolis, 17
Brazil. 18
*These authors contributed equally to this work 19
ΔCurrent address: Laboratoire d’Analyses de Sèvres Atlantique, 210 avenue de la Venise Verte 20
79000 Niort, France 21
22
#Corresponding authors: 23
Copyright © 2013, American Society for Microbiology. All Rights Reserved.J. Clin. Microbiol. doi:10.1128/JCM.01573-13 JCM Accepts, published online ahead of print on 31 July 2013
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

2
Sabine Pereyre: USC EA3671 infections humaines à mycoplasmes et chlamydiae, Université 24
Bordeaux Segalen, 146 rue Léo Saignat, 33076 Bordeaux, France. E-mail: sabine.pereyre@u-25
bordeaux2.fr. Phone number: +33 5 57 57 16 25. Fax number: +33 56 93 29 40. 26
Florence Tardy: Anses, Laboratoire de Lyon, UMR Mycoplasmoses des Ruminants, 31 Avenue 27
Tony Garnier, 69364 Lyon Cedex 07, France. 28
29
Running title: Identification of mycoplasmas by MALDI-TOF MS 30
31
Keywords: matrix-assisted laser desorption ionization-time of flight mass spectrometry, human 32
mycoplasmas, ruminant mycoplasmas, species identification, subtyping 33
34
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

3
35
Abstract 36
Matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) has 37
recently emerged as a new technology for the identification of bacteria. In this study, we aimed to 38
evaluate its applicability to human and ruminant mycoplasma identification, which can be 39
demanding and time-consuming using phenotypic or molecular methods. In addition, MALDI-TOF 40
MS was tested as a subtyping tool for certain species. A total of 29 main spectra (MSP) from 10 41
human and 13 ruminant mycoplasma (sub)species were included in a mycoplasma MSP database to 42
complete the Bruker MALDI Biotyper database. After broth culture and protein extraction, 43
MALDI-TOF MS was applied for identification purposes to 119 human and 143 ruminant clinical 44
isolates previously identified by antigenic or molecular methods and to subcultures of 73 ruminant 45
clinical specimens known to potentially contain several mycoplasma species. MALDI-TOF MS 46
resulted in accurate (sub)species identification, with a score ≥ 1.700 for 96% (251/262) of the 47
isolates. The phylogenetically closest (sub)species were unequivocally distinguished. Although 48
mixtures of strains were reliably detected up to a certain cellular ratio, only the predominant species 49
was identified from the cultures of polymicrobial clinical specimens. For typing purposes, MALDI-50
TOF MS proved to be able to cluster M. bovis and M. agalactiae isolates as a function of their year 51
of isolation and their genome profile, respectively, and M. pneumoniae isolates as a function of their 52
P1-adhesin type. In conclusion, MALDI-TOF MS is a rapid, reliable and cost-effective method for 53
the routine identification of high-density growing mycoplasmas and shows promising prospects for 54
strain typing. 55
56
57
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

4
Introduction 58
In recent years, matrix-assisted laser desorption ionization-time of flight mass spectrometry 59
(MALDI-TOF MS) has emerged as a new technology for species identification. MALDI-TOF MS 60
is a rapid, inexpensive and accurate approach for species identification of Gram-negative and Gram-61
positive bacteria and yeasts (9). It has been recently optimized for routine use in clinical 62
microbiology laboratories (15, 39). This method was further demonstrated to be sufficiently robust 63
to be applicable to fastidious bacteria, such as anaerobic bacteria, Legionella, Bartonella or 64
mycobacteria (4). 65
Mycoplamas are wall-less bacteria with minimal genomes but are nonetheless important pathogens 66
in both human and veterinary medicine (32). The family Mycoplasmataceae of the class Mollicutes 67
regroups two major genera, Mycoplasma and Ureaplasma (19), which will hereafter be indicated by 68
their trivial name, i.e., mycoplasmas. 69
The identification of human and ruminant mycoplasmas is challenging, and phenotypic methods 70
cannot always achieve identification to the species level. Furthermore, in contrast to more classical 71
bacteria, mycoplasmas express few biochemical traits useful for diagnosis. Nonetheless, several 72
biochemical procedures have been standardized, such as the hydrolysis of urea and arginine, 73
fermentation of glucose (41) and phosphatase activity, (29) yet may be time-consuming and not 74
always discriminating. For example, it is not possible to distinguish in culture Ureaplasma parvum 75
from U. urealyticum because they both hydrolyze urea or to distinguish Mycoplasma pneumoniae 76
from M. amphoriforme in culture because they both grow in Hayflick modified broth medium 77
supplemented with glucose (HG medium). Consequently, to identify these species easily, important 78
efforts have been made over the past decades to develop diagnostic tools relying on the antigenic 79
(28, 30) and molecular identification of isolates, mainly based on PCR assays (13, 23, 26, 41, 44). 80
Nevertheless, the identification of certain mycoplasmas, such as subspecies belonging to the 81
M. mycoides cluster, comprising several important ruminant pathogens that are closely related both 82
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

5
antigenically and genetically, has remained problematic (21, 38). Although genes encoding 83
16S rRNA have long been the usual target for universal PCRs followed by sequence/amplicon 84
analysis (18, 24), species-specific PCRs targeting 16S rRNA or other housekeeping genes are often 85
favored because the techniques are more rapid and do not require a specialized laboratory (for 86
reviews see (23, 41)). However, with more than one hundred mycoplasmas currently recognized as 87
pathogens in humans and animals, individual PCR-based systems developed as species-specific 88
tests could be difficult to handle in a laboratory and thus the quest for a single generic test remains 89
pressing. MALDI-TOF MS could be an interesting universal alternative for mycoplasma 90
(sub)species identification, though its use has been hindered thus far by a lack of databases of 91
mycoplasmal MALDI-TOF reference spectra. The technique per se has been assessed only once for 92
the identification of M. pulmonis, a rodent mycoplasma, using a limited database of three MALDI-93
TOF spectra from three rodent pathogen species (16). Indeed, no ruminant or human mycoplasma 94
identification using MALDI-TOF MS has been reported to date, even though these bacteria 95
represent important pathogens for these hosts (32). 96
The aim of this study was to determine whether MALDI-TOF MS could be a useful tool for the 97
identification of ruminant and human mycoplasmas at the species or subspecies level and a 98
subtyping method for certain species. A total of 29 reference peak lists, main spectra (MSPs), from 99
10 human and 13 ruminant mycoplasma (sub)species were first generated to construct a 100
mycoplasma spectral database and complete the database of the Bruker MALDI Biotyper platform. 101
MALDI-TOF MS was then applied for identification purposes to 119 human and 143 ruminant 102
clinical isolates previously identified to the species level using phenotypic, antigenic or molecular 103
techniques and to 73 subcultures of clinical specimens known to potentially contain more than one 104
species of ruminant mycoplasmas. All the analyses were optimized to be compatible with the 105
routine workflow of our laboratories. Subtyping was assessed using clinical strains of 106
M. pneumoniae, M. bovis and M. agalactiae. 107
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

6
108
Material and methods 109
Reference strains, clinical isolates and specimens 110
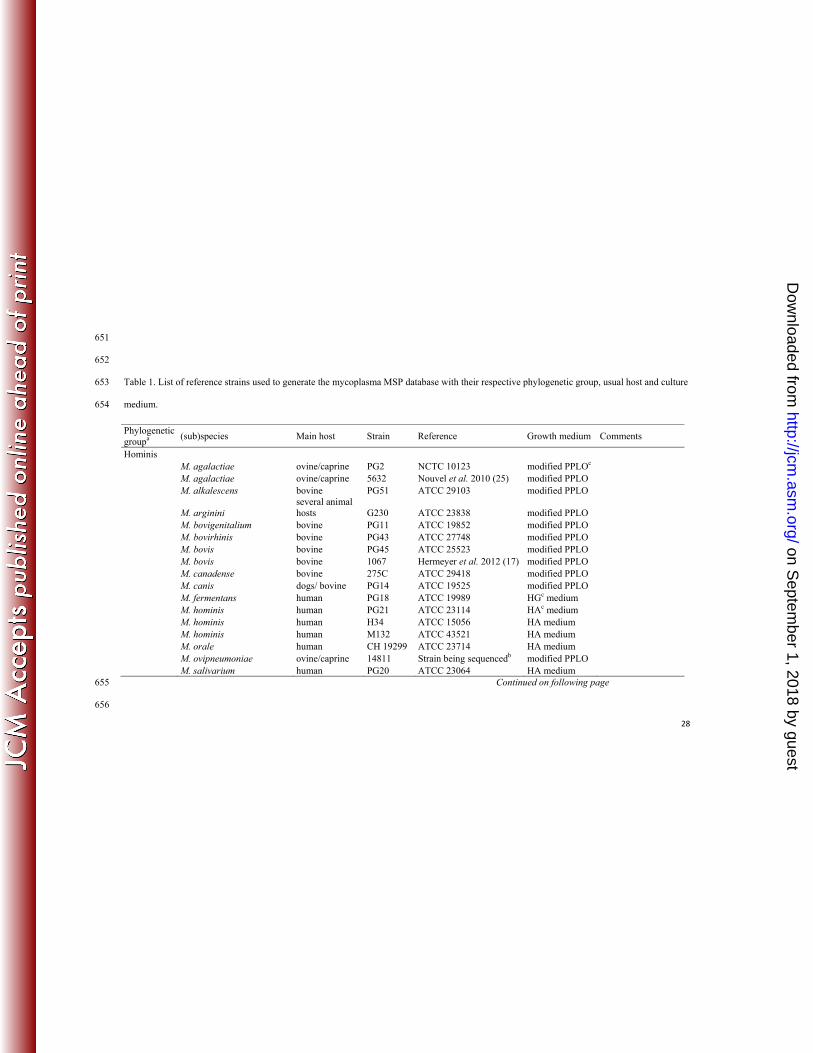
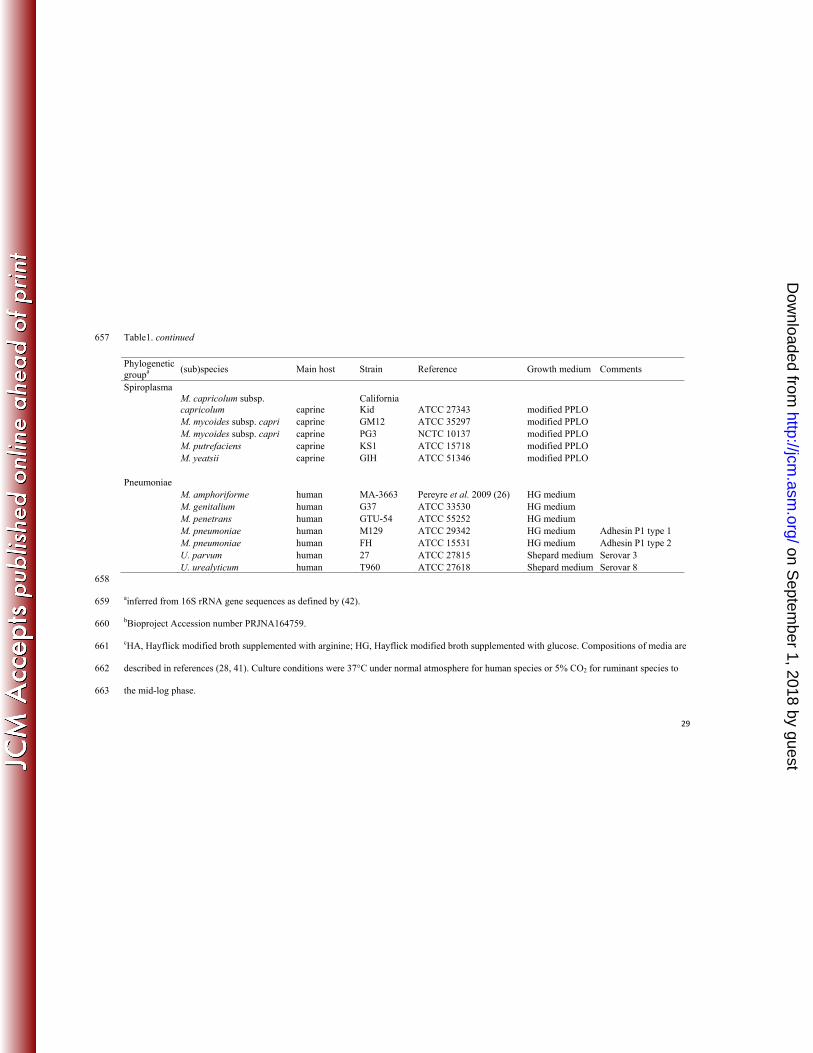
The reference strains used to generate MSPs are listed in Table 1. All strains were grown at 37°C in 111
diverse media, as detailed in the table; several strains were used as reference strains for some 112
species. For comparison purposes, a maximum likelihood phylogenetic tree of the 16S rRNA 113
sequences of these strains or species was computed with MEGA5 software 114
(http://www.megasoftware.net/). All the 16S rRNA sequences were downloaded from the 115
Ribosomal DataBase project (http://rdp.cme.msu.edu/index.jsp) or from the Nucleotide database at 116
NCBI (http://www.ncbi.nlm.nih.gov/nuccore/). 117
Three clinical isolates of M. amphoriforme, one clinical isolate of M. penetrans and 50 clinical 118
isolates of M. pneumoniae collected in Bordeaux University hospital, France, between 2006 and 119
2011, together with four adhesin P1 variants 2a (MP-Ho, MP-Sta, MP-79809 and MP-79692) (10) 120
and one variant 1 (MP-4817) (12) of M. pneumoniae, were used in the study. These isolates had 121
been previously identified using specific real-time PCR targeting the adhesin P1 gene (40) for 122
M. pneumoniae and the 16S rRNA gene for M. amphoriforme (26) and 16S rRNA gene sequencing 123
for M. penetrans. The isolates and variants were cultured at 37°C in HG medium (41). Fifty-five 124
clinical isolates of M. hominis collected in Bordeaux University hospital in 2009 and 2010, 125
previously identified by culture and confirmed by a specific real-time PCR targeting the yidC gene 126
(13), were cultured at 37°C in Hayflick modified broth supplemented with arginine (HA medium) 127
(41). Six clinical isolates of U. parvum and five clinical isolates of U. urealyticum collected in 128
Bordeaux University hospital and previously identified by a real-time PCR assay targeting the 129
urease gene (44) were cultured at 37°C in Shepard broth (41). 130
The clinical isolates from ruminants were grown in PPLO broth (Difco, France), supplemented as 131
previously described (28), at 37°C and 5% CO2. The isolates were first identified using dot 132
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

7
immunobinding on membrane filtration (MF-dot) (28), the current method for routine strain 133
identification in the Anses laboratory of Lyon (38) within the framework of the 134
epidemiosurveillance network VIGIMYC (6). The specimens consisted of i) 48 M. agalactiae 135
isolates collected between 1951 and 2009 in France and Europe from domestic small ruminants and 136
wild ungulates with contagious agalactia, ii) 48 M. bovis isolates collected between 1986 and 2010 137
worldwide from cattle with pneumonia or, to a lesser extent, mastitis (5) or arthritis (1) and iii) 47 138
isolates from the M. mycoides cluster, mainly M. mycoides subsp. capri (Mmc, 19) and 139
M. capricolum subsp. capricolum (Mcc, 19), with a few M. putrefaciens (4) and M. yeatsii (5) 140
isolates, all collected from ruminants with mastitis, arthritis or pneumonia or asymptomatic animals 141
in the case of the M. yeatsii strains. 142
The capacity of MALDI-TOF MS to simultaneously identify two strains in a mixed culture was also 143
assessed by analyzing i) artificially constructed mixtures of strains and ii) a total of 73 subcultures 144
of bovine pneumonia clinical specimens known to often contain more than one mycoplasma species. 145
The artificial mixtures were prepared by mixing variable volumes of culture in HA medium and 146
Shepard medium for the human mycoplasmas or by mixing equal volumes of individual strain 147
cultures after cell counting using a “most probable number” method for the ruminant species (7). 148
The specimen subcultures had been routinely analyzed by MF-dot within the VIGIMYC network 149
and kept frozen at -20°C prior to further analysis by MALDI-TOF MS in a unique run. 150
All the analyses of the ruminant mycoplasmas were performed using a single blind protocol. The 151
blinding was performed in Lyon, France, where the strains were grown, and the specimens were 152
then sent to Saint-Lô, France, for protein extraction and MALDI-TOF MS analysis. Culturing and 153
analysis of the human mycoplasmas were performed in Bordeaux, France. 154
155
Protein extraction for MALDI-TOF MS analysis 156
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

8
Mycoplasma pellets were obtained after centrifugation of variable volumes of culture, depending on 157
the growth capacity of the individual species. For the human mycoplamas, 30 ml of culture of 158
reference strain was used to generate the MSP library, with the exception for Ureaplasma spp. for 159
which 100 ml was required. The culture volumes were the same for the identification of the human 160
clinical isolates using MALDI-TOF MS i.e. 30 ml for the M. pneumoniae clinical isolates and 161
100 ml for the Ureaplasma spp. isolates, except for the M. hominis clinical isolates for which a 1-ml 162
culture was sufficient. For the ruminant mycoplasmas, 1 ml of culture was used for both the MSP 163
construction and identification of clinical isolates. The mycoplasma pellets were washed once for 164
the human mycoplasmas and twice for the ruminant mycoplasmas in 1X PBS prior to being 165
suspended in 300 µl of PBS or water then 900 µl of absolute ethanol was added to precipitate the 166
proteins. After centrifugation at 20,000 g for 2 min, the supernatant was removed, and the pellet 167
was dissolved in an equal volume (20 µl or 50 µl) of 70% formic acid and acetonitrile before 168
centrifugation for 2 min at 20,000 g. One µl of the supernatant was spotted onto an MALDI MSP 169
96 polished target steel plate (Bruker Daltonics, Bremen, Germany). After air-drying at room 170
temperature, each sample was overlaid with 1 µl of matrix solution composed of 10 mg/ml α-cyano-171
4-hydroxycinnamic acid in 50% acetonitrile and 2.5% trifluoroacetic acid and air-dried prior to the 172
MALDI-TOF MS measurement. 173
174
MALDI-TOF mass spectrometry 175
Peptide mass fingerprint product ion spectra were generated using a Microflex LT Biotyper 176
operating system (Bruker Daltonics, Bremen, Germany). The data were analyzed in the automatic 177
mode using Bruker Biotyper 3.0 software and both taxonomy libraries, i.e., that from Bruker and 178
that generated in the present study. A bacterial test standard (BTS; part number 255343, Bruker 179
Daltonics Inc.) was used in each run as a calibrator and for quality control. 180
181
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

9
Mycoplasma MSP database construction and isolate identification by MALDI-TOF MS 182
Reference peak lists were assigned to different reference strains (Table 1) to construct the 183
mycoplasma MSP database, which was used in addition to the Bruker MALDI biotyper MSP 184
database 3.1.2.0 (January 2011) for the species identification of the clinical samples. The MSPs 185
were created according to the manufacturer’s recommendations by using the automated MSP 186
creation functionality of the MALDI Biotyper software package (version 3.0). For each reference 187
strain, 20 individual mass spectra from 20 different spots of protein extracts from human 188
mycoplasma species or three independent mass spectra measurements of eight different spots of 189
protein extracts from ruminant mycoplasma were acquired. The quality of individual raw spectra 190
was carefully checked (absence of intrusive peaks, absence of flat line spectra, low matrix 191
background signal) and a minimum of 18 spectra of high quality were selected for MSP creation. 192
After smoothing, base line correction, and peak-picking, the resulting peak lists were used by the 193
program to calculate and store a MSP containing the information about mean peak masses, mean 194
peak intensities and peak frequency. 195
A total of 20 to 24 spectra from each reference strain (Table 1) were used to create an MSP. The 196
protein extracts from each strain were loaded onto 8 spots, which were analyzed three times for the 197
ruminant mycoplasma species, or onto 20 spots, which were analyzed once for the human 198
mycoplasma species. MSP creation was performed with the default setting of the Biotyper software. 199
For the identification of clinical isolates, the protein extracts were spotted once, twice or 10 times, 200
according to the species to be analyzed and classified by matching MSPs. The degree of spectral 201
concordance is expressed as a logarithmic identification score ranging from 0 to 3 and interpreted 202
according to the manufacturer’s instructions, with a modification of the score acceptable for 203
probable species identification, which was lowered from ≥ 2.000 to ≥ 1.700. 204
The dendrograms were generated using the Biotyper 3.0 software. They were used to infer the 205
relationships of mycoplasma species or strains, which closeness is reflected by an arbitrary distance 206
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

10
level calculated by the software. This distance level is a relative value and thus cannot be compared 207
between dendrograms. 208
209
Results 210
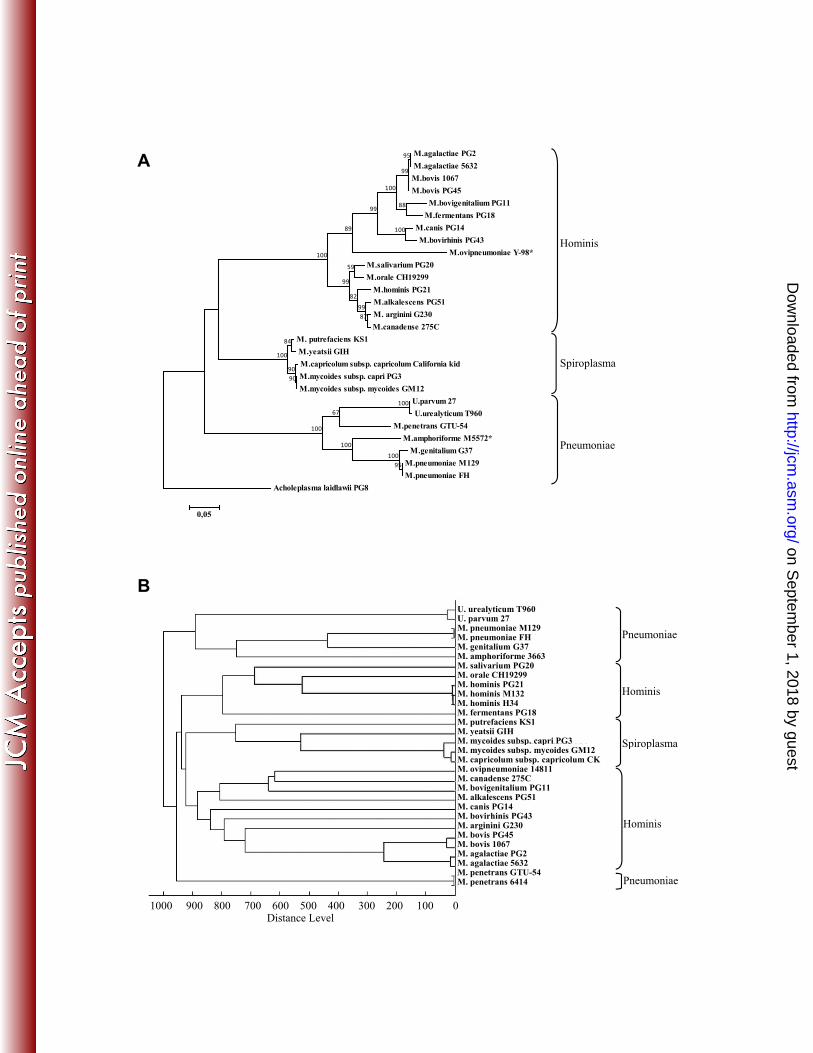
Human and ruminant mycoplasma MSP database construction 211
To create the human and ruminant mycoplasma MSP database, 29 strains, as listed in Table 1, were 212
cultured in an appropriate medium prior to protein extraction, MALDI-TOF MS and MSP 213
generation. These strains represent 10 human and 13 pathogenic or commensal mycoplasma 214
(sub)species belonging to different phylogenetic groups based on 16S rRNA sequences, as 215
illustrated in Figure 1A. To control the quality of this database, the MSPs generated were further 216
used to perform a hierarchical clustering of strains using the “MSP dendrogram” tool of the MALDI 217
biotyper software. The resulting score-oriented dendrogram (Figure 1B) showed an overall topology 218
that was comparable to that of the phylogenetic 16S rRNA tree as defined by Weisburg et al. (42) 219
(Figure 1A). When using an arbitrary distance level of 900 as a cut-off, all the strains were correctly 220
assigned to their respective phylogenetic group in the dendrogram. The Pneumoniae, Spiroplasma 221
and Hominis groups yielded unequivocally separated branches, whereas both the Hominis and 222
Pneumoniae groups were further split into two branches. This was particularly intriguing for 223
M. penetrans GTU-54, which was isolated in a specific branch. We further examined this 224
topological particularity by generating another MSP using a clinical strain of the M. penetrans 225
species, namely strain 6414. In the dendrogram, M. penetrans strain GTU-54 and 6414 remained 226
grouped on a unique branch and separated from each other by a very short distance, as expected for 227
strains belonging to the same species. 228
229
Identification of clinical isolates 230
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

11
The mycoplama colonies are so small and often inlaid in the agar that direct colony deposition often 231
failed in our study to result in species or genus identification, with the exception of (sub)species 232
growing to a high density, such as M. mycoides subsp. capri and some isolates of M. hominis. 233
Moreover, the extraction step was preferred for the mycoplasma species identification because 234
attempts to deposit the mycoplasma pellet directly onto the target plate resulted in weaker scores 235
than when the complete extraction procedure was applied. Thus, a broth culture and protein 236
extraction was performed in all cases in this study. The required culture volume depended on the 237
species to be identified (see below). 238
As the broth media used to grow mycoplasmas contain a large concentration of proteins, non-239
seeded HA, HG, Shepard and PPLO medium batches were analyzed by MALDI-TOF MS as 240
negative controls. These samples were treated like the clinical isolates, as described in Material and 241
Methods, and the resulting spectra were compared to those of the mycoplasma and MALDI 242
Biotyper databases. No spectral concordance was obtained, suggesting that the proteins contained in 243
the different culture media had no significant influence on the identification of the clinical isolates. 244
Moreover, we tested the SP4 medium (41) instead of the usual HG medium to grow M. pneumoniae 245
M129 reference strain before MALDI-TOF MS. The strain was accurately identified as 246
M. pneumoniae M129 with a score >2.000, confirming that changing the culture medium had no 247
influence on the MALDI-TOF MS identification of this species. 248
For the M. hominis species and other species growing in HA medium, preliminary tests revealed 249
that protein extraction from a culture volume of 1 ml was sufficient to achieve species identification. 250
Of the 55 M. hominis clinical isolate extracts spotted twice, 100% were identified to the species 251
level, 95% (52/55) had a score values ≥ 2.000 for both spots, and 5% (3/55) had only one score ≥ 252
2.000 and the other score in the range 1.700-1.999. For M. pneumoniae and other species growing 253
in HG medium and for the Ureaplasma species growing in Shepard medium, a culture volume of 30 254
ml and 100 ml, respectively, was required before protein extraction. Ten spots from the same 255
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

12
protein extract were deposited onto the target plate. Fifty clinical isolates of M. pneumoniae and 256
three clinical isolates of M. amphoriforme were analyzed. In all cases, at least eight spots out of ten 257
yielded a score ≥1.700, corresponding to an accurate species identification. There was no 258
misidentification, and MALDI-TOF MS was able to distinguish M. pneumoniae from M. genitalium, 259
the phylogenetically closest species. Eleven clinical isolates of Ureaplasma spp. were analyzed, 260
consisting of six clinical isolates of U. parvum and five clinical isolates of U. urealyticum. In 100% 261
of the cases, at least eight spots out of ten yield a score ≥1.700, with accurate identification to the 262
species level. 263
For the ruminant mycoplasmas, each clinical sample was only spotted once to develop a procedure 264
compatible with a routine workflow. Of the first set, 96% (46/48) of the M. agalactiae clinical 265
isolates were successfully identified by MALDI-TOF MS, with a score ≥1.700. Thirty-three strains 266
had a score ≥2.000 and 13 a score between 1.700 and 1.999. The only identification failures were 267
due to poor growth of the strains, one of which resulted nonetheless in an accurate identification of 268
M. agalactiae with a 1.5 score. 269
For M. bovis, 98% (47/48) of the analyzed isolates were correctly identified, with 36 yielding a 270
score ≥2.000 and 11 yielding a score between 1.700 and 1.999. The only strain that yielded an 271
unsatisfactory score of 1.6 was a strain (strain 8790) known to have an intermediate position, by 272
both MLST and 16S rRNA sequencing, between M. agalactiae and M. bovis, two highly related 273
species of the Hominis group (22). This strain was nonetheless assigned by MALDI-TOF MS to 274
M. bovis. 275
Forty-seven strains from the M. mycoides cluster were also analyzed, including several species or 276
subspecies. For both M. capricolum subsp. capricolum (Mcc) and M. mycoides subsp. capri (Mmc), 277
95% (18/19) of the strains were correctly assigned to their respective subspecies, with scores ≥1.700. 278
Interestingly, the only strain among the Mmc strains that yielded a score of 1.56 was not Mmc, as 279
was suggested by its MF-dot profile; indeed, this strain was later assigned by house-keeping gene 280
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

13
sequence analysis to the newly described species M. feriruminatoris sp. nov. (14). The 281
identification of the M. putrefaciens strains was congruent with the MF-dot results despite 2/4 282
yielding a score <1.700. In contrast, the identification scores were considered unacceptable for 283
M. yeatsii (<1.300, n=5). 284
285
Analysis of clinical specimens containing more than one mycoplasma (sub)species 286
Because human and ruminant clinical specimens are often polycontaminated by different 287
mycoplasma species, we assessed the capacity of MALDI-TOF MS to address mixtures of strains. 288
When two species, M. bovis and M. bovirhinis, U. parvum and M. hominis or U. urealyticum and 289
M. hominis, were artificially mixed in equivalent cellular concentrations, both species were detected 290
by MALDI-TOF MS, with an approximate equivalent score for each species. However, when the 291
cellular ratio was unbalanced (from 1/5 to 1/100, depending on the species), the predominant 292
species had a score greater than 2.000, whereas the species at the lower concentration was not 293
detected or received a score smaller than 1.300, which was considered to be uninterpretable. 294
A total of 73 subcultures of the bovine pneumonia clinical specimens were also analyzed, 57 of 295
which had previously been identified as a single strain by MF-dot (M. bovis, M. bovirhinis, 296
M. arginini or M. bovigenitalium) and 16 as a mixture of 2 to 3 strains from different species, 297
including M. bovis and one or two other species such as M. alkalescens, M. bovirhinis, 298
M. canadense, M. bovigenitalium and M. arginini. Of the 57 single-strain containing samples, 97% 299
(55/57) were identified by MALDI-TOF MS, with scores ≥1.700 and a species match consistent 300
with that of MF-dot. The 2 single strains for which a discrepancy was observed were identified by 301
MF-dot as M. arginini and by MALDI-TOF MS as M. bovis, with scores ≥1.700. Two subcultures 302
yielded different identification results when examined by MF-dot prior to -20°C storage 303
(identification of M. arginini), and by MALDI-TOF MS after storage (identification of M. bovis, 304
with scores ≥1.700). This discrepancy was not the result of a wrong identification but of a species 305
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

14
ratio modification during the storage of the strain mixtures. For the 16 specimens containing 2 to 3 306
species, as detected by MF-dot, M. bovis was the only component of the mixture that was identified 307
by MALDI-TOF MS. 308
309
Mycoplasma subtyping 310
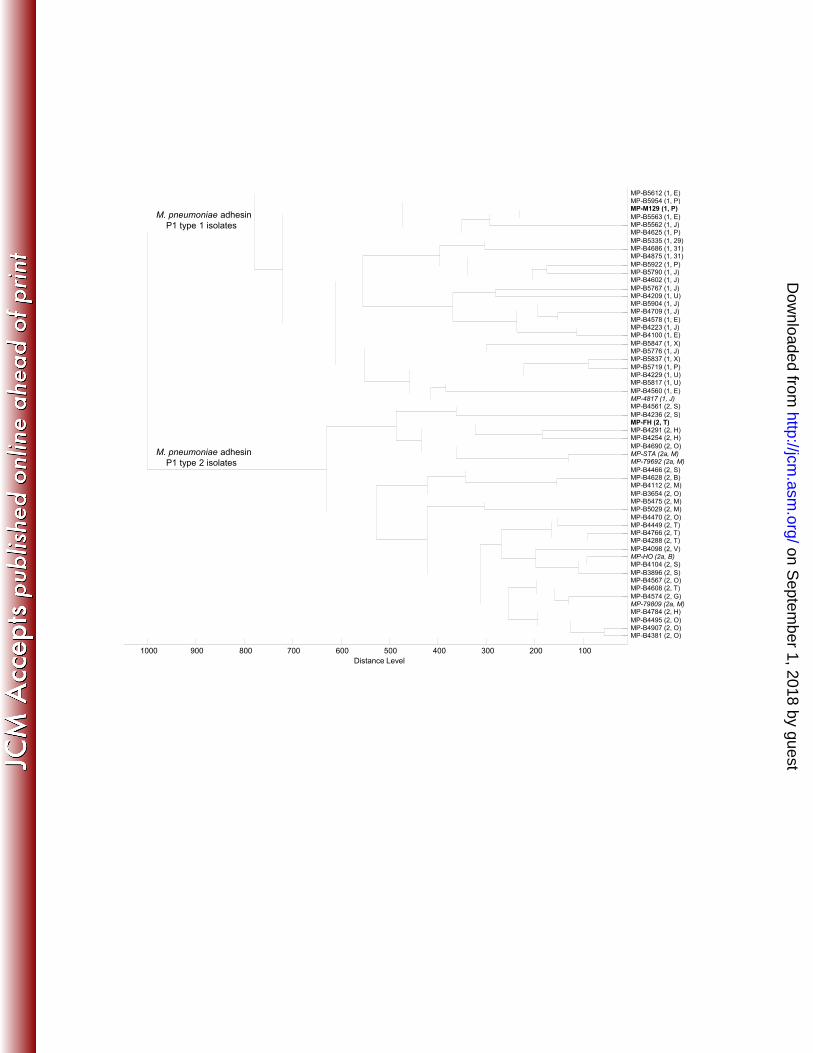
The spectra acquired for species identification purposes were further used to assess the subtyping 311
capacity of MALDI-TOF MS for M. pneumoniae strains, and M. bovis and M. agalactiae strains. 312
Based on the analysis of the gene encoding the P1 protein, M. pneumoniae is known to comprise 313
two subtypes (type 1 and type 2) and a few variants related to each subtype (8). Using MALDI-TOF 314
MS, all the 50 M. pneumoniae spectral profiles were accurately clustered into 2 separate groups 315
corresponding to M. pneumoniae adhesin P1 type 1 (reference strain, M. pneumoniae M129) and 316
M. pneumoniae adhesin P1 type 2 (reference strain, M. pneumoniae FH) (Figure 2). In addition, 317
three M. pneumoniae variant 2a and one variant 1 strains were accurately ranked among the type 2 318
and type 1 strains, respectively (Figure 2). We also searched for an association between the 319
MALDI-TOF MS results and the recently developed Multi-Locus Variable-Number Tandem-320
Repeat (VNTR) analysis (MLVA) typing method based on the analysis of the number of tandem 321
repeats present at five loci of the M. pneumoniae genome (10). Although no association could be 322
drawn because the isolates of the same MLVA type were present in different branches of the 323
dendrogram (Figure 2), all MLVA types J, P, E, U, X, 29 and 31 were clustered in the adhesin P1 324
type 1 strains, and all MLVA types B, T, G, V, M, O, S and H were clustered in the adhesin P1 type 325
2 strains. This finding was expected because a correlation between the MLVA typing results and the 326
type of adhesin P1 gene was previously reported (10), with the former MLVA types being related to 327
adhesin P1 type 1 and the latter MLVA types being related to adhesin P1 type 2. 328
M. bovis and M. agalactiae are two highly related mycoplasma species that are important pathogens 329
of cattle and small ruminants, respectively. An MLST approach was recently proposed as an 330
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

15
unequivocal tool for the strain differentiation, characterization and molecular typing of these two 331
species (22). To infer whether MALDI-TOF MS could be useful to type strains within each species, 332
we generated a dendrogram using 18 strains from each species, all of which, except for the 333
reference strains, were collected in France in different years and from different hosts in the case of 334
M. agalactiae (Figure 3). In the resulting dendrogram, the M. agalactiae strains were separated into 335
two branches that correspond to the two reference strains, namely PG2 and #5632. For M. bovis, the 336
strains were regrouped into two main branches as a function of their year of isolation. In the 337
dendrogram, we also observed that the M. agalactiae group has longer branches than the M. bovis 338
branch, which indicates a more variable species, in agreement with MLST data (22). 339
340
Discussion 341
To assess the feasibility of using MALDI-TOF MS for the identification of clinically relevant 342
mycoplasmas in both human and veterinary medicine we first attempted to enrich an available 343
mycoplasma database that only contained, when this study was initiated, one porcine mycoplasma 344
species, M. hyorhinis (Bruker MALDI Biotyper MSP database 3.1.2.0). For this purpose, 29 MSPs 345
corresponding to 23 mycoplasma (sub)species were constructed, representing most of the human 346
and ruminant pathogenic and commensal species distributed in three phylogenetic groups of the 347
class Mollicute, namely, Spiroplasma, Hominis and Pneumoniae. The inclusion of pathogenic and 348
commensal species in this database had the two following purposes: i) to be able to unequivocally 349
exclude the presence of pathogenic species by the identification of non-pathogenic species and ii) to 350
address a mixture of species in clinical specimens. For example, in ruminant respiratory specimens, 351
M. bovis, a pathogenic mycoplasma, is frequently associated with M. bovirhinis, which is a 352
commensal species. 353
The MSP dendrogram resulted in a clustering of strains that was overall congruent with the 16S 354
rRNA phylogeny, though some groups were further subdivided into several branches (Figure 1). 355
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

16
This observation was an important quality control for further species identification using this 356
mycoplasma database. To limit the potential misidentification of atypical strains from species 357
known to be variable, we included several strains from the same species in the database. For 358
instance, three strains of M. hominis were included, as this species is known to be highly 359
heterogeneous (20). Both M. agalactiae PG2 type strain and #5632 were acquired as well, as they 360
are considered to each be situated at one end of the genetic spectrum encountered in M. agalactiae 361
species (25). This proved successful because, for instance, several strains from wild ungulates 362
(Capra ibex) that are known to be genetically different from domestic ruminant strains (37) were 363
correctly identified as M. agalactiae. For M. bovis, a strain known to be pathogenic (strain 1067) 364
(17) was also used together with the type strain PG45. Lastly, within the recently reclassified Mmc 365
taxon, the former type strain of the MmmLC biotype, namely GM12, was included in addition to the 366
PG3 type strain. Generating several MSPs per species is also a convenient step for subtyping strains 367
once they have been assigned to a species (see below). While this study was being conducted, 368
Bruker released an update of their own database (MALDI Biotyper MSP database 3.3.1.0, April 369
2012), which now includes 10 species of the Mycoplasma genus. However, only six ruminant 370
mycoplasma species were added, none of which from the M. mycoides cluster, and no human 371
mycoplasma species or strains from the Ureaplasma genus are yet included. 372
373
The mycoplasma cultivation and sample preparation were optimized to be compatible with the 374
laboratory workflow for diagnosis. The direct deposition of a colony or a cell pellet on the target 375
plate was shown to occasionally yield species identification, though with scores weaker than those 376
obtained when employing the protein extraction process. As a consequence, as already suggested 377
for other bacteria (9), we chose to apply the protein extraction protocol to increase the rates of 378
identification. This protocol relies on a series of centrifugations and re-suspensions steps that 379
remain, in our opinion, compatible with routine usage. An increased number of washing steps 380
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

17
performed on the mycoplasma pellet prior to extraction improved the spectral quality, and this was 381
mainly due to the removal of the strong protein background generated by the yeast extract and horse 382
serum, two major components of mycoplasma growth media. Nonetheless, increasing the number of 383
washing steps also resulted in a decrease in the quantity of the proteins contained in the final extract. 384
For clinical identification, a 1-ml culture was sufficient to obtain interpretable spectra for most 385
species, except for those growing in HG medium, such as M. pneumoniae, or in Shepard medium, 386
such as Ureaplasma spp., for which 30 ml and 100 ml of culture, respectively, were required. 387
Handling such volumes may be hardly compatible with a routine clinical workflow, however 388
attempts to reduce these volumes failed. These results are consistent with the cell density of 389
individual cultures. For instance, M. hominis, one of the less fastidious human mycoplasmas, 390
typically reaches concentration up to 108 to 109 color changing units (CCU)/ml at the mid-log phase 391
in HA medium, whereas M. pneumoniae and Ureaplasma spp. only yield concentrations of 107-108 392
CCU/ml and 106-107 CCU/ml, respectively. Most of the ruminant species analyzed in this study 393
reached a concentration of 108 to 1010 CFU/ml at the stationary phase, and Stevenson et al. 394
determined that excellent spectra were obtained when a minimum of 106 CFU was spotted onto the 395
target plate (36), which is approximately what we obtained when starting from 1 ml of a 108 396
CFU/ml culture. 397
Bruker Daltonics recommends scores ≥1.700 and ≥2.000 for genus- and species-level identification, 398
respectively. However, as previously demonstrated for other bacteria (1, 2, 5, 39), our results 399
strongly advocate a reduction of the scores acceptable for mycoplasma species identification to 400
≥1.700. With these parameters, the rate of species-level identification was 100%, with no 401
misidentification, even for the low cell density species, i.e., Ureaplasma spp. and M. pneumoniae. 402
Furthermore, several subspecies of the complex M. mycoides cluster known to be highly related 403
were unequivocally identified. Additionally, based on the high rate of identification to the 404
(sub)species level, a reduction in the number of spots deposited on the target plate from the same 405
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

18
isolate was considered when appropriate. For species that grow in HA medium, such as M. hominis, 406
one spot can be utilized for routine diagnosis because it yielded a score ≥1.700 in all cases. In 407
contrast, ten spots were utilized for more fastidious species that grow in HG or Shepard medium. 408
When considering the results obtained using the first three spots for the 50 M. pneumoniae isolates 409
and 11 Ureaplasma spp. isolates, identification to the species level, with a score ≥1.700 for two out 410
of these three spots, was obtained in 98% cases (60/61), suggesting that three spots could be 411
sufficient. For the ruminant mycoplasmas, a single spot was deposited, leading to a correct 412
identification in 95 to 98% cases, with a score >1.700. In cases of a misidentification or a score 413
<1.700, we recommend to either re-launch the analysis with several spots or to increase the quantity 414
of starting material. 415
416
We further demonstrated that MALDI-TOF MS can accommodate in vitro reconstituted mixtures of 417
species up to a certain ratio level. Regarding human urogenital mycoplasma species, it must be 418
noted that Ureaplasma spp. and M. hominis do not grow in the same culture medium, which 419
reduces the need to detect this mixture of species by MALDI-TOF MS because the pre-cultivation 420
step will result in a high predominance of the cultured species. However, such discrimination is of 421
interest for ruminant species that grow in the same medium, as we previously reported that more 422
than 10% of clinical specimens could contain more than one mycoplasma (sub)species (6). 423
However, in the 16 specimens containing two to three ruminant species, MALDI-TOF MS only 424
detected M. bovis, the clinically relevant species, that may have been favored to the detriment of 425
other species during the -20°C storage phase. Another potential explanation for these discrepancies 426
between MF-dot and MALDI-TOF MS identification techniques might be based on the slightly 427
better sensitivity of MF-dot (from 2x104 to 8x106 mycoplasmas per well (28)) versus MALDI-TOF 428
MS (106 CFU per spot (36)), which can result in the co-detection of some non-predominant species 429
when using MF-dot. In this study, different identification results were obtained for two clinical 430
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

19
specimens not considered to be mixture based on MF-dot results, e.g., M. arginini by MF-dot and 431
M. bovis by MALDI-TOF MS. These results might point toward another mixture of strains in which 432
M. bovis was overgrown by M. arginini when the MF-dot was performed but for which the species 433
ratio was modified by a long storage at -20°C prior to the MALDI-TOF MS identification. In 434
conclusion, as already shown by Stevenson et al. (36), not all organisms present in a polymicrobial 435
sample can be reliably detected by MALDI-TOF MS. 436
437
MALDI-TOF MS allowed the identification of Ureaplasma spp. at the species level, which is 438
generally not achieved in routine laboratory diagnosis. Indeed, the former Ureaplasma urealyticum 439
species consisted of a heterogeneous species comprising 2 biovars and 14 serovars. This taxon was 440
split 10 years ago into two distinct species, U. parvum and U. urealyticum, corresponding to the 441
former biovar 1 (including 4 serovars) and biovar 2 (including 10 serovars), respectively (33). 442
Distinction between each species requires either antibody-based phenotyping methods, which are 443
often inconclusive because of multiple cross-reactions, or more accurate molecular techniques (43, 444
44). Although an additional overnight incubation was necessary to obtain a 100-ml culture volume, 445
MALDI-TOF MS allowed an easy identification to the species level of Ureaplasma spp, which is a 446
benefit of the clinical use of MALDI-TOF MS because several studies have reported U. urealyticum 447
to be more pathogenic than U. parvum (11, 31). 448
449
MALDI-TOF MS also proved to accurately distinguish species known to be highly related: 450
M. pneumoniae, M. genitalium and M. amphoriforme, which grow in the same HG medium, and 451
M. bovis and M. agalactiae, which used to be classified as two subspecies of the same species prior 452
to being separated into two different species based on serological, DNA-DNA re-association 453
experiments (3) and 16S rRNA sequence data (27). Furthermore, within individual species, the 454
dendrograms generated from the MALDI-TOF MS spectra achieved epidemiologically relevant 455
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

20
strain clustering. For instance, the M. agalactiae strains were separated into two branches that 456
correspond to the two reference strains, namely PG2 and #5632, which are described as 457
representing each end of the genetic spectrum encountered in M. agalactiae species (25). The 458
strains from wild ungulates, namely Capra ibex or chamois, were interestingly grouped together, as 459
previously reported using partial sequencing of a house-keeping gene (37). For M. bovis, the strains 460
were regrouped into two main branches that correspond to recent and old strains, a sub-clustering 461
that is consistent with the recent data obtained in our group by MLST and MLVA (Becker, personal 462
communication). Although these preliminary typing results are promising, further studies that 463
include a larger number of isolates are needed to confirm these findings. Moreover, MALDI-TOF 464
MS allowed not only the identification of M. pneumoniae isolates but also their adhesin P1 typing 465
in a single step without the need of additional molecular typing techniques (8, 34, 35). However, no 466
direct correlation was found between MALDI-TOF MS and MLVA typing. 467
468
Conclusion 469
In this study, we developed a mycoplasma spectral database of human and ruminant mycoplasmas 470
that proved to be reliable for the identification of clinical isolates. High concordance was found 471
between the MALDI-TOF MS species identification and species identification using biochemical, 472
antigenic or molecular methods. MALDI-TOF MS is a rapid, reliable and cost-effective method, 473
particularly for the routine identification of M. hominis and ruminant mycoplasmas that grow to a 474
high cell density, and could replace conventional identification methods. Moreover, MALDI-TOF 475
MS with no further optimization proved to be useful for the subtyping of several mycoplasma 476
species and may be promising for other typing developments. 477
478
Acknowledgements 479
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

21
We thank Marc Bonneu and Jean-William Dupuy from the genomic platform of the University 480
Bordeaux Segalen for their advice. We are grateful to all VIGIMYC members, François Poumarat 481
and the technical staff in Lyon responsible for managing the collection of the ruminant 482
mycoplasmas strains. We thank Marie Gardette for technical assistance. 483
484
Figure legends 485
Figure 1. Clustering of Mycoplasma and Ureaplasma strains as a function of their 16S rRNA 486
phylogeny (A) or their MSP dendrogram (B). 487
A. Phylogenetic tree showing the position of the strains used in this study. Strains for which a 488
complete 16S rRNA gene sequence was not available were replaced with other strains of the same 489
species and are indicated with a *. Only partial sequences were available for strains H34 and M132 490
of M. hominis, and these strains were consequently not included in the tree. The evolutionary 491
history of the strains was inferred using the Maximum Likelihood method. The tree with the highest 492
likelihood (-9189,0715) is shown, and Acholeplasma laidlawii PG8 was used as an outgroup. A 493
total of 1407 nucleotides from the 16S rRNA gene were used in the analysis. The dataset was 494
resampled 1000 times, and the bootstrap percentage values (>50%) are given at the nodes. 495
Phylogenetic groups as defined by Weiburg et al. (42) are also presented. 496
B. Score-oriented dendrogram of MSPs from human and ruminant mycoplasmas species used to 497
construct the mycoplasma database. The dendrogram was generated with the default settings in 498
MALDI Biotyper (distance measure by correlation and linkage by average). 499
500
Figure 2. Dendrogram of the MALDI-TOF MS profiles generated using MALDI Biotyper 3.0 with 501
50 clinical isolates of M. pneumoniae, M. pneumoniae reference strains FH and M129, four 502
M. pneumoniae variants 2a and 1 M. pneumoniae variant 1: distance measure was set at correlation, 503
and linkage was set at average. The adhesin P1 type and the MLVA type are given in parentheses, 504
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

22
respectively. The reference strains M129 and FH are in bold, and the variants are italicized. MP, 505
M. pneumoniae. 506
507
Figure 3. Dendrogram of MALDI-TOF MS profiles generated using MALDI Biotyper 3.0 with 36 508
clinical isolates of M. agalactiae and M. bovis. The M. agalactiae strains are designated by ‘Maga’, 509
followed by the strain number, the year of isolation and the host species. The M. bovis strains are 510
designated by ‘Mbov’, followed by the strain number and the year of isolation in brackets. 511
512
References 513
1. Alatoom, A. A., C. J. Cazanave, S. A. Cunningham, S. M. Ihde, and R. Patel. 2012. 514
Identification of non-diphtheriae corynebacterium by use of matrix-assisted laser desorption 515
ionization-time of flight mass spectrometry. J. Clin. Microbiol. 50:160-163. 516
2. Alatoom, A. A., S. A. Cunningham, S. M. Ihde, J. Mandrekar, and R. Patel. 2011. 517
Comparison of direct colony method versus extraction method for identification of gram-518
positive cocci by use of Bruker Biotyper matrix-assisted laser desorption ionization-time of 519
flight mass spectrometry. J. Clin. Microbiol. 49:2868-2873. 520
3. Askaa, G., and H. Erno. 1976. Elevation of Mycoplasma agalactiae subsp. bovis to species 521
rank: Mycoplasma bovis (Hale et al.) comb. nov. Int. J. Syst. Bacteriol.:323-325. 522
4. Biswas, S., and J. M. Rolain. 2013. Use of MALDI-TOF mass spectrometry for 523
identification of bacteria that are difficult to culture. J. Microbiol. Methods 92:14-24. 524
5. Bizzini, A., K. Jaton, D. Romo, J. Bille, G. Prod'hom, and G. Greub. 2011. Matrix-525
assisted laser desorption ionization-time of flight mass spectrometry as an alternative to 16S 526
rRNA gene sequencing for identification of difficult-to-identify bacterial strains. J. Clin. 527
Microbiol. 49:693-696. 528
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

23
6. Chazel, M., F. Tardy, D. Le Grand, D. Calavas, and F. Poumarat. 2010. Mycoplasmoses 529
of ruminants in France: recent data from the national surveillance network. BMC Vet. Res. 530
6:32. 531
7. Cochran, W. G. 1950. Estimation of bacterial densities by means of the "most probable 532
number". Biometrics 6:105-116. 533
8. Cousin-Allery, A., A. Charron, B. de Barbeyrac, G. Fremy, J. S. Jensen, H. Renaudin, 534
and C. Bébéar. 2000. Molecular typing of Mycoplasma pneumoniae strains by PCR-based 535
methods and pulsed-field gel electrophoresis. Application to French and Danish isolates. 536
Epidemiol. Infect. 124:103-111. 537
9. Croxatto, A., G. Prod'hom, and G. Greub. 2011. Applications of MALDI-TOF mass 538
spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 36:380-407. 539
10. Dégrange, S., C. Cazanave, A. Charron, H. Renaudin, C. Bébéar, and C. M. Bébéar. 540
2009. Development of multiple-locus variable-number tandem-repeat analysis for molecular 541
typing of Mycoplasma pneumoniae. J. Clin. Microbiol. 47:914-923. 542
11. Deguchi, T., T. Yoshida, T. Miyazawa, M. Yasuda, M. Tamaki, H. Ishiko, and S. I. 543
Maeda. 2004. Association of Ureaplasma urealyticum (Biovar 2) with nongonococcal 544
urethritis. Sexually Transmitted Dis. 31:192-195. 545
12. Dorigo-Zetsma, J. W., B. Wilbrink, J. Dankert, and S. A. Zaat. 2001. Mycoplasma 546
pneumoniae P1 type 1- and type 2-specific sequences within the P1 cytadhesin gene of 547
individual strains. Infect. Immun. 69:5612-5618. 548
13. Férandon, C., O. Peuchant, C. Janis, A. Benard, H. Renaudin, S. Pereyre, and C. 549
Bébéar. 2011. Development of a real-time PCR targeting the yidC gene for the detection of 550
Mycoplasma hominis and comparison with quantitative culture. Clin. Microbiol. Infect. 551
17:155-159. 552
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

24
14. Fischer, A., I. Santana-Cruz, M. Giglio, S. Nadendla, E. Drabek, E. M. Vilei, J. Frey, 553
and J. Jores. 2013. Genome sequence of Mycoplasma feriruminatoris sp. nov., a fast-554
growing mycoplasma species. Genome Announc. 1. 555
15. Ford, B. A., and C. A. Burnham. 2013. Optimization of routine identification of clinically 556
relevant Gram-negative bacteria using MALDI-TOF MS and the Bruker Biotyper. J. Clin. 557
Microbiol. Epub ahead of print. 558
16. Goto, K., M. Yamamoto, M. Asahara, T. Tamura, M. Matsumura, N. Hayashimoto, 559
and K. Makimura. 2012. Rapid identification of Mycoplasma pulmonis isolated from 560
laboratory mice and rats using matrix-assisted laser desorption ionization time-of-flight 561
mass spectrometry. J. Vet. Med. Sci. 74:1083-1086. 562
17. Hermeyer, K., I. Buchenau, A. Thomasmeyer, B. Baum, J. Spergser, R. Rosengarten, 563
and M. Hewicker-Trautwein. 2012. Chronic pneumonia in calves after experimental 564
infection with Mycoplasma bovis strain 1067: characterization of lung pathology, 565
persistence of variable surface protein antigens and local immune response. Acta Vet. Scand. 566
54:9. 567
18. Johansson, K. E., M. Heldtander, and P. B. 1998. Characterization of mycoplasmas by 568
PCR and sequence analysis with universal 16S rDNA primers, p. 145-165. In M. R. J. and R. 569
A. J. Nicholas (ed.), Methods in molecular biology, Mycoplasma protocols, vol. 104. 570
Humana press, Totowa. 571
19. Johansson, K. E., and B. Pettersson. 2002. Taxonomy of Mollicutes, p. 1-29. In S. Razin 572
and R. Herrmann (ed.), Molecular biology and pathogenicity of mycoplasmas. Kluwer 573
Academic/Plenum Publishers, London. 574
20. Ladefoged, S. A., and G. Christiansen. 1992. Physical and genetic mapping of the 575
genomes of five Mycoplasma hominis strains by pulsed-field gel electrophoresis. J. 576
Bacteriol. 174:2199-2207. 577
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

25
21. Le Grand, D., E. Saras, D. Blond, M. Solsona, and F. Poumarat. 2004. Assessment of 578
PCR for routine identification of species of the Mycoplasma mycoides cluster in ruminants. 579
Vet. Res. 35:635-649. 580
22. Manso-Silvan, L., V. Dupuy, I. Lysnyansky, U. Ozdemir, and F. Thiaucourt. 2012. 581
Phylogeny and molecular typing of Mycoplasma agalactiae and Mycoplasma bovis by 582
multilocus sequencing. Vet. Microbiol. 161:104-112. 583
23. Markham, P. F., and A. H. Noormohammadi. 2005. Diagnosis of mycoplasmosis in 584
animals, p. 355-382. In A. Blanchard and G. F. Browning (ed.), Mycoplasmas molecular 585
biology pathogenicity and strategies for control. Horizon bioscience, Norfolk. 586
24. McAuliffe, L., R. J. Ellis, J. R. Lawes, R. D. Ayling, and R. A. Nicholas. 2005. 16S 587
rDNA PCR and denaturing gradient gel electrophoresis; a single generic test for detecting 588
and differentiating Mycoplasma species. J. Med. Microbiol. 54:731-739. 589
25. Nouvel, L. X., P. Sirand-Pugnet, M. S. Marenda, E. Sagne, V. Barbe, S. Mangenot, C. 590
Schenowitz, D. Jacob, A. Barre, S. Claverol, A. Blanchard, and C. Citti. 2010. 591
Comparative genomic and proteomic analyses of two Mycoplasma agalactiae strains: clues 592
to the macro- and micro-events that are shaping mycoplasma diversity. BMC Genomics 593
11:86. 594
26. Pereyre, S., H. Renaudin, A. Touati, A. Charron, O. Peuchant, A. B. Hassen, C. Bébéar, 595
and C. M. Bébéar. 2009. Detection and susceptibility testing of Mycoplasma amphoriforme 596
isolates from patients with respiratory tract infections. Clin. Microbiol. Infect. 16:1007-1009. 597
27. Pettersson, B., M. Uhlen, and K. E. Johansson. 1996. Phylogeny of some mycoplasmas 598
from ruminants based on 16S rRNA sequences and definition of a new cluster within the 599
hominis group. Int. J. Syst. Bacteriol. 46:1093-1098. 600
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

26
28. Poumarat, F., B. Perrin, and D. Longchambon. 1991. Identification of ruminant 601
mycoplasmas by dot immunobinding on membrane filtration (MF dot). Vet. Microbiol. 602
29:329-338. 603
29. Poveda, J. B. 1998. Biochemical characteristics in mycoplasma identification. Methods Mol. 604
Biol. 104:69-78. 605
30. Poveda, J. B., and R. Nicholas. 1998. Serological identification of mycoplasmas by growth 606
and metabolic inhibition tests. Methods Mol. Biol. 104:105-111. 607
31. Povlsen, K., E. Bjornelius, P. Lidbrink, and I. Lind. 2002. Relationship of Ureaplasma 608
urealyticum biovar 2 to nongonococcal urethritis. Eur. J. Clin. Microbiol. Infect. Dis. 21:97-609
101. 610
32. Razin, S., D. Yogev, and Y. Naot. 1998. Molecular biology and pathogenicity of 611
mycoplasmas. Microbiol. Mol. Biol. Rev. 62:1094-1156. 612
33. Robertson, J. A., G. W. Stemke, J. W. Davis, Jr., R. Harasawa, D. Thirkell, F. Kong, M. 613
C. Shepard, and D. K. Ford. 2002. Proposal of Ureaplasma parvum sp. nov. and emended 614
description of Ureaplasma urealyticum (Shepard et al. 1974) Robertson et al. 2001. Int. J. 615
Syst. Evol. Microbiol. 52:587-597. 616
34. Schwartz, S. B., K. A. Thurman, S. L. Mitchell, B. J. Wolff, and J. M. Winchell. 2009. 617
Genotyping of Mycoplasma pneumoniae isolates using real-time PCR and high-resolution 618
melt analysis. Clin. Microbiol. Infect. 15:756-762. 619
35. Spuesens, E. B., T. Hoogenboezem, M. Sluijter, N. G. Hartwig, A. M. van Rossum, and 620
C. Vink. 2010. Macrolide resistance determination and molecular typing of Mycoplasma 621
pneumoniae by pyrosequencing. J. Microbiol. Methods 82:214-222. 622
36. Stevenson, L. G., S. K. Drake, and P. R. Murray. 2010. Rapid identification of bacteria in 623
positive blood culture broths by matrix-assisted laser desorption ionization-time of flight 624
mass spectrometry. J. Clin. Microbiol. 48:444-447. 625
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

27
37. Tardy, F., E. Baranowski, L. X. Nouvel, V. Mick, L. Manso-Silvan, F. Thiaucourt, P. 626
Thebault, M. Breton, P. Sirand-Pugnet, A. Blanchard, A. Garnier, P. Gibert, Y. Game, 627
F. Poumarat, and C. Citti. 2012. Emergence of atypical Mycoplasma agalactiae strains 628
harboring a new prophage and associated with an alpine wild ungulate mortality episode. 629
Appl. Environ. Microbiol. 78:4659-4668. 630
38. Tardy, F., P. Gaurivaud, A. Tricot, L. Maigre, and F. Poumarat. 2009. Epidemiological 631
surveillance of mycoplasmas belonging to the 'Mycoplasma mycoides' cluster: is DGGE 632
fingerprinting of 16S rRNA genes suitable? Lett. Appl. Microbiol. 48:210-217. 633
39. Tekippe, E. M., S. Shuey, D. W. Winkler, M. A. Butler, and C. A. Burnham. 2013. 634
Optimizing identification of clinically relevant Gram-positive organisms using the Bruker 635
Biotyper MALDI-TOF MS system. J. Clin. Microbiol. Epub ahead of print. 636
40. Touati, A., A. Bénard, A. Ben Hassen, C. M. Bébéar, and S. Pereyre. 2009. Evaluation 637
of five commercial real-time PCR assays for the detection of Mycoplasma pneumoniae in 638
respiratory tract specimens. J. Clin. Microbiol. 47:2269-2271. 639
41. Waites, K. B., C. M. Bébéar, J. A. Roberston, D. F. Talkington, and G. E. Kenny (ed.). 640
2001. Cumitech 34, Laboratory diagnosis of mycoplasmal infections. American Society for 641
Microbiology, Washington D. C. 642
42. Weisburg, W. G., J. G. Tully, D. L. Rose, J. P. Petzel, H. Oyaizu, D. Yang, L. Mandelco, 643
J. Sechrest, T. G. Lawrence, J. Van Etten, and et al. 1989. A phylogenetic analysis of the 644
mycoplasmas: basis for their classification. J. Bacteriol. 171:6455-6467. 645
43. Xiao, L., V. Paralanov, J. I. Glass, L. B. Duffy, J. A. Robertson, G. H. Cassell, Y. Chen, 646
and K. B. Waites. 2011. Extensive horizontal gene transfer in ureaplasmas from humans 647
questions the utility of serotyping for diagnostic purposes. J. Clin. Microbiol. 49:2818-2826. 648
44. Yi, J., B. H. Yoon, and E. C. Kim. 2005. Detection and biovar discrimination of 649
Ureaplasma urealyticum by real-time PCR. Mol. Cell Probes 19:255-260. 650
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
28
651
652
Table 1. List of reference strains used to generate the mycoplasma MSP database with their respective phylogenetic group, usual host and culture 653
medium. 654
Continued on following page 655
656
Phylogenetic groupa
(sub)species Main host Strain Reference Growth medium Comments
Hominis M. agalactiae ovine/caprine PG2 NCTC 10123 modified PPLOc M. agalactiae ovine/caprine 5632 Nouvel et al. 2010 (25) modified PPLO M. alkalescens bovine PG51 ATCC 29103 modified PPLO
M. arginini several animal hosts G230 ATCC 23838 modified PPLO
M. bovigenitalium bovine PG11 ATCC 19852 modified PPLO M. bovirhinis bovine PG43 ATCC 27748 modified PPLO M. bovis bovine PG45 ATCC 25523 modified PPLO M. bovis bovine 1067 Hermeyer et al. 2012 (17) modified PPLO M. canadense bovine 275C ATCC 29418 modified PPLO M. canis dogs/ bovine PG14 ATCC 19525 modified PPLO M. fermentans human PG18 ATCC 19989 HGc medium M. hominis human PG21 ATCC 23114 HAc medium M. hominis human H34 ATCC 15056 HA medium M. hominis human M132 ATCC 43521 HA medium M. orale human CH 19299 ATCC 23714 HA medium M. ovipneumoniae ovine/caprine 14811 Strain being sequencedb modified PPLO M. salivarium human PG20 ATCC 23064 HA medium
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
29
Table1. continued 657
658
ainferred from 16S rRNA gene sequences as defined by (42). 659
bBioproject Accession number PRJNA164759. 660
cHA, Hayflick modified broth supplemented with arginine; HG, Hayflick modified broth supplemented with glucose. Compositions of media are 661
described in references (28, 41). Culture conditions were 37°C under normal atmosphere for human species or 5% CO2 for ruminant species to 662
the mid-log phase. 663
Phylogenetic groupa
(sub)species Main host Strain Reference Growth medium Comments
Spiroplasma
M. capricolum subsp. capricolum caprine
California Kid ATCC 27343 modified PPLO
M. mycoides subsp. capri caprine GM12 ATCC 35297 modified PPLO M. mycoides subsp. capri caprine PG3 NCTC 10137 modified PPLO M. putrefaciens caprine KS1 ATCC 15718 modified PPLO M. yeatsii caprine GIH ATCC 51346 modified PPLO Pneumoniae M. amphoriforme human MA-3663 Pereyre et al. 2009 (26) HG medium M. genitalium human G37 ATCC 33530 HG medium M. penetrans human GTU-54 ATCC 55252 HG medium M. pneumoniae human M129 ATCC 29342 HG medium Adhesin P1 type 1 M. pneumoniae human FH ATCC 15531 HG medium Adhesin P1 type 2 U. parvum human 27 ATCC 27815 Shepard medium Serovar 3 U. urealyticum human T960 ATCC 27618 Shepard medium Serovar 8
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
M.agalactiae PG2
M.agalactiae 5632
M.bovis 1067
M.bovis PG45
M b i it li PG11
100
95
99A
M.bovigenitalium PG11
M.fermentans PG18
M.canis PG14
M.bovirhinis PG43
M.ovipneumoniae Y-98*
M.salivarium PG20
M.orale CH19299
M.hominis PG21
M.alkalescens PG51
100
99
82
59
99
89
100
9988
Hominis
M. arginini G230
M.canadense 275C
M. putrefaciens KS1
M.yeatsii GIH
M.capricolum subsp. capricolum California kid
M.mycoides subsp. capri PG3
M.mycoides subsp. mycoides GM12
U.parvum 27
U.urealyticum T960100
67
9090
84
100
8799
Spiroplasma
y
M.penetrans GTU-54
M.amphoriforme M5572*
M.genitalium G37
M.pneumoniae M129
M.pneumoniae FH
Acholeplasma laidlawii PG8
95100
100
100
0,05
Pneumoniae
BU. urealyticum T960U. parvum 27M. pneumoniae M129M i FH PneumoniaeM. pneumoniae FHM. genitalium G37M. amphoriforme 3663M. salivarium PG20M. orale CH19299M. hominis PG21M. hominis M132M. hominis H34M. fermentans PG18M. putrefaciens KS1M. yeatsii GIHM. mycoides subsp. capri PG3
Pneumoniae
Hominis
SpiroplasmaM. mycoides subsp. capri PG3M. mycoides subsp. mycoides GM12M. capricolum subsp. capricolum CKM. ovipneumoniae 14811M. canadense 275CM. bovigenitalium PG11M. alkalescens PG51M. canis PG14M. bovirhinis PG43M. arginini G230M. bovis PG45M. bovis 1067M l ti PG2
Hominis
Spiroplasma
M. agalactiae PG2M. agalactiae 5632M. penetrans GTU-54M. penetrans 6414 Pneumoniae
1000 900 800 700 600 500 400 300 200 100 0 Distance Level
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from
MP B5954 (1 P)MP-B5612 (1, E)
M. pneumoniae adhesin P1 type 1 isolates
MP B4709 (1 J)MP-B5904 (1, J)MP-B4209 (1, U)MP-B5767 (1, J)MP-B4602 (1, J)MP-B5790 (1, J)MP-B5922 (1, P)MP-B4875 (1, 31)MP-B4686 (1, 31)MP-B5335 (1, 29)MP-B4625 (1, P)MP-B5562 (1, J)MP-B5563 (1, E)MP-M129 (1, P)MP-B5954 (1, P)
MP-B4236 (2, S)MP-B4561 (2, S)MP-4817 (1, J)MP-B4560 (1, E)MP-B5817 (1, U)MP-B4229 (1, U)MP-B5719 (1, P)MP-B5837 (1, X)MP-B5776 (1, J)MP-B5847 (1, X)MP-B4100 (1, E)MP-B4223 (1, J)MP-B4578 (1, E)MP-B4709 (1, J)
M. pneumoniae adhesinP1 type 2 isolates
MP-B4449 (2 T)MP-B4470 (2, O)MP-B5029 (2, M)MP-B5475 (2, M)MP-B3654 (2, O)MP-B4112 (2, M)MP-B4628 (2, B)MP-B4466 (2, S)MP-79692 (2a, M)MP-STA (2a, M)MP-B4690 (2, O)MP-B4254 (2, H)MP-B4291 (2, H)MP-FH (2, T)
( , )
MP B4381 (2 O)MP-B4907 (2, O)MP-B4495 (2, O)MP-B4784 (2, H)MP-79809 (2a, M)MP-B4574 (2, G)MP-B4608 (2, T)MP-B4567 (2, O)MP-B3896 (2, S)MP-B4104 (2, S)MP-HO (2a, B)MP-B4098 (2, V)MP-B4288 (2, T)MP-B4766 (2, T)MP B4449 (2, T)
1002003004005006007008009001000Distance Level
MP-B4381 (2, O)
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from

Maga 4206 (1989, sheep)Maga PG2 (Ref., 1952, goat)Maga 14987 (2008, sheep)Maga 14965 (2008, sheep)Maga 14633 (2006 goat)
PG2 branch
Maga 14633 (2006, goat)Maga 5885 (1992, goat)Maga 4023 (1989, sheep)Maga 14557 (2005, sheep)Maga 14968 (2008, goat)Maga 4212 (1989, sheep)Maga 4054 (1989, goat)Maga 14668 (2006, goat)Maga 14628 (2006, wild fauna)Maga 5632 (Ref., before 199, goat)Maga 15310 (2009, wild fauna)Maga 14980 (2008, wild fauna)Maga 15341 (2009, wild fauna)
5632 branch
M. agalactiae
aga 53 ( 009, d au a)Maga 13387 (2003, wild fauna)Mbov PG45 (Ref., 1962, USA)Mbov 1067 (Ref., 1983, FR)Mbov 4698 (1991)Mbov 2199 (1986)Mbov 5700 (1991)Mbov 4785 (1990)Mbov 6129 (1992)Mbov 4904 (1990)Mbov 10269 (1997)Mbov 14956 (2008)Mbov 14746 (2007)
old
recent
M. bovis
Mbov 14611 (2006)Mbov 15508 (2010)Mbov 15711 (2011)Mbov 15176 (2008)Mbov 15210 (2009)Mbov 15355 (2010)Mbov 15345 (2009)
1000 900 600 500 400 300 200 100 0 Distance Level
on Septem
ber 1, 2018 by guesthttp://jcm
.asm.org/
Dow
nloaded from











![ΚΡΙΑΡΑ[13]-ΜΕΣΑΙΩΝΙΚΟ ΛΕΞΙΚΟ [13].pdf](https://static.fdocument.org/doc/165x107/563db810550346aa9a903693/13-13pdf-5661db9bb3055.jpg)






