
Κεφάλαιο Υδατοκαλλιέργειες › bitstream › 11419 › 5088 › 1 ›...
39
Κεφάλαιο 5 Καλλιέργεια ιχθύων: αρχές γενετικής βελτίωσης Αλέξανδρος Τριανταφυλλίδης Υδατοκαλλιέργειες | Καλλιέργεια Ιχθύων
Transcript of Κεφάλαιο Υδατοκαλλιέργειες › bitstream › 11419 › 5088 › 1 ›...

Κεφάλαιο 5
Καλλιέργεια ιχθύων: αρχές γενετικής βελτίωσης
Αλέξανδρος Τριανταφυλλίδης
Υδατ
οκαλ
λιέρ
γειε
ς |
Καλλ
ιέργ
εια
Ιχθύ
ων

2 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
Καλλιέργεια ιχθύων: αρχές γενετικής βελτίωσης Σύνοψη Η γενετική δομή των εκτρεφόμενων ζώων στις υδατοκαλλιέργειες είναι πολύ σημαντική, γιατί είναι αυτή που καθορίζει πρωταρχικά τις παραγωγικές δυνατότητές τους. Στο παρόν Κεφάλαιο θα αναλυθούν κάποιες βασικές αρχές γενετικής, καθώς και οι γενετικές μέθοδοι που χρησιμοποιούνται για την ανάλυση των γονιδίων και των γονιδιωμάτων. Επίσης θα συζητηθούν τα βασικά χαρακτηριστικά της ποσοτικής γενετικής που μελετάει γονίδια που κληρονομούνται με ποσοτικό τρόπο και παρουσιάζουν, συνήθως οικονομικό ενδιαφέρον. Θα αναφερθούμε επίσης σε δύο βασικές τεχνολογίες γενετικής που έχουν ενδιαφέρον στις υδατοκαλλιέργειες, τον καθορισμό του φύλου και τη δημιουργία πολυπλοειδών πληθυσμών. Τέλος, θα αναλυθούν οι δυνατότητες που δημιουργούνται με την ανάπτυξη της βιοτεχνολογίας αλλά και οι βιοηθικοί προβληματισμοί που ανακύπτουν.
Προαπαιτούμενη γνώση Βασικές αρχές γενετικής, κληρονομικότητας και γενετικής μηχανικής καθώς και αρχών υδατοκαλλιέργειας (Κεφάλαιο 1 του παρόντος).
1. Εισαγωγή
Η γενετική σύσταση των ζώων που εκτρέφονται σε υδατοκαλλιέργειες δεν έχει τύχει της ίδιας προσοχής όπως η ποιότητα και η ποσότητα της τροφής που παρέχεται, η καταπολέμηση των ασθενειών τους και η ποιότητα του νερού. Όμως, η γενετική δομή των εκτρεφόμενων ζώων είναι εξίσου σημαντική, επειδή είναι αυτή που καθορίζει πρωταρχικά τις παραγωγικές δυνατότητές τους. Δυστυχώς, οι περισσότεροι υδατοκαλλιεργητές αγνοούσαν την αξία της γενετικής, ίσως γιατί οι γενετιστές δεν κατόρθωναν να εξηγήσουν τις βασικές έννοιες της επιστήμης τους με απλούς και κατανοητούς όρους. Είναι όμως πια γενικά αποδεκτό, ότι όσοι ασχολούνται με υδατοκαλλιέργειες πρέπει να εισάγουν τις βασικές έννοιες της γενετικής βελτίωσης στη διαχείριση της παραγωγικής μονάδας τους. Οι διαχειριστές πρέπει να γνωρίζουν ότι η παραγωγικότητα εξαρτάται, σε μεγάλο βαθμό, από την πρακτική διασταυρώσεων και από το πώς γίνεται η γενετική διαχείριση του πληθυσμού. Σήμερα, στις ιχθυοκαλλιέργειες (του εξωτερικού, αλλά και στην Ελλάδα) εφαρμόζονται διάφορες μέθοδοι γενετικής, με σκοπό να βελτιώσουν ποικίλες βιολογικές παραμέτρους και να επηρεάσουν θετικά το τελικό αποτέλεσμα, είτε αυτό προσμετράται σε ποιοτικά προϊόντα, είτε σε οικονομική απόδοση.
2. Αποκρυπτογράφηση του γονιδιώματος εκτρεφόμενων οργανισμών. Γενετική βάση της κληρονομικότητας και της κυτταρογενετικής
2.1 Ανάλυση γονιδιώματος Το γενετικό υλικό στα ζώα, όπως είναι γνωστό, βρίσκεται στο πυρηνικό και στο μιτοχονδριακό DNA. Το μιτοχονδριακό γονιδίωμα (mtDNA) των ζώων είναι κλειστό δίκλωνο κυκλικό μόριο και αποτελεί ένα πολύ μικρό ποσοστό του συνολικού DNA. Το μέγεθός του ποικίλλει και εξαρτάται από το είδος του οργανισμού. Για παράδειγμα το mtDNA του ανθρώπου αποτελείται από 16.569 bp (ζευγάρια βάσεων), του κυπρίνου από 16.575 bp, και του Αφρικανικού πνευμονόψαρου (Protopterus dollioi) από 16.646 bp. Στα σπονδυλωτά περιέχει 37 γονίδια, από τα οποία τα 22 κωδικοποιούν για μόρια tRNA, δύο για μόρια rRNA και 13 για μόρια mRNA. Τα τελευταία μεταφράζονται σε πρωτεΐνες, οι οποίες συμμετέχουν στη μεταφορά ηλεκτρονίων και στην οξειδωτική φωσφορυλίωση. Οι πρωτεΐνες αυτές είναι το κυτόχρωμα b, επτά υπομονάδες της αφυδρογονάσης του NADH, τρεις υπομονάδες του κυτοχρώματος c και δύο υπομονάδες της μιτοχονδριακής συνθετάσης του ATP. Το μιτοχονδριακό γονιδίωμα περιλαμβάνει, επίσης, και μια μη

3 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
κωδικοποιούσα περιοχή, την περιοχή ελέγχου ή βρόχο εκτόπισης D, όπου βρίσκονται οι προαγωγείς για τη μεταγραφή των δύο πολυνουκλεοτιδικών αλυσίδων, καθώς και η ακολουθία έναρξης της αντιγραφής της μιας αλυσίδας. Ιντρόνια, επαναλαμβανόμενες ακολουθίες DNA και ψευδογονίδια απουσιάζουν συνήθως από το mtDNA. Η διάταξη των γονιδίων στο mtDNA των ζώων παρουσιάζει αξιοσημείωτη σταθερότητα, ιδιαίτερα για τα ψάρια. Το mtDNA κληρονομείται κυρίως μητρικά, από τα θηλυκά άτομα και στα δύο φύλα. Σε λίγες εξαιρέσεις, όπως τα μύδια, έχουν βρεθεί δύο είδη mtDNA. Το ένα από αυτά (F) μεταβιβάζεται από τη μητέρα στους απογόνους και των δύο φύλων, ενώ το άλλο (Μ) κληρονομείται από το αρσενικό άτομο μόνο στους αρσενικούς απογόνους. Πρόσφατα, έχουν διαπιστωθεί αρκετές περιπτώσεις ετεροπλασματικών ατόμων (ατόμων που περιέχουν στα κύτταρα τους μιτοχόνδρια με περισσότερους από έναν τύπους mtDNA, με διαφορετικές δηλαδή μεταλλάξεις). Τέλος, αρκετές ακολουθίες του mtDNA έχουν μεταφερθεί στο πυρηνικό γονιδίωμα. Αυτές οι καταστάσεις δημιουργούν προβλήματα στην ανάλυση του μιτοχονδριακού DNA. Παρ’ όλα αυτά δεν είναι τόσο συχνές για αυτό και το μιτοχονδριακό DNA αποτελούσε για χρόνια το βασικό μοριακό εργαλείο μελέτης της γενετικής ποικιλότητας και διαφοροποίησης των πληθυσμών. Πυρηνικό γονιδίωμα. Η αλματώδης ανάπτυξη των τεχνικών ανάλυσης του DNA επέτρεψε την αποκρυπτογράφηση των πληροφοριών του γενετικού υλικού του πυρήνα, δηλαδή την ανεύρεση της σειράς των βάσεων του DNA, σε αρκετά είδη ψαριών, ανάμεσα στα οποία είναι δύο είδη της οικογένειας Tetraodontidae (Fugu rubripes, Tetraodon nigrobiridis), το ψάρι ζέβρα (Danio rerio) και το ριζόψαρο (Oryzias latipes). Έχει ολοκληρωθεί και το γονιδίωμα σπουδαίων εκτρεφόμενων ειδών, όπως είναι ο σολομός του ατλαντικού, η ιριδίζουσα πέστροφα, η τούρνα (Πίνακας 5.1), ενώ αυτή τη στιγμή είναι σε εξέλιξη προγράμματα για τουλάχιστον άλλα πενήντα είδη ιχθύων, ανάμεσα στα οποία είναι και η τσιπούρα και το λαυράκι, αλλά και για ασπόνδυλα με ενδιαφέρον για τις υδατοκαλλιέργειες όπως το στρείδι του Ειρηνικού (Crassostrea gigas) και διάφορα είδη γαρίδων. Μάλιστα, στο πλαίσιο του προγράμματος Genome 10K (https://genome10k.soe.ucsc.edu/), όπου θα διαβαστούν τα γονιδιώματα 10.000 σπονδυλωτών, αναμένεται να παραχθούν πληροφορίες για 4.000 είδη ψαριών. Η πλήρης αλληλουχία ενός γονιδιώματος αποτελεί τον αναλυτικό χάρτη που μας επιτρέπει να ανακαλύψουμε τα γονίδια και τους πολυμορφισμούς με ενδιαφέρον για την παραγωγή. Από τις πιο σημαντικές πληροφορίες που μπορούμε να βρούμε συγκρίνοντας διαφορετικά άτομα είναι η θέση των απλών νουκλεοτιδικών πολυμορφισμών (single nucleotide polymorphisms, SNPs). Οι πολυμορφισμοί SNPs είναι ο βασικός λόγος που ένα άτομο αποδίδει διαφορετικά από ένα άλλο. Χρειάζεται όμως πρώτα πολλή πειραματική δουλειά για να συσχετιστεί το γενετικό υπόβαθρο με την απόδοση στις υδατοκαλλιέργειες, ώστε να επιλεγούν ως γεννήτορες για τις επόμενες γενιές άτομα με ανώτερο γενετικό υλικό. Μια εξίσου σημαντική πληροφορία που συλλέγεται από την αλληλούχιση του γονιδιώματος είναι η κατανόηση της λειτουργίας και η ταυτοποίηση των γονίδιων που σχετίζονται με την παραγωγή και την απόδοση των ατόμων. Με τον τρόπο αυτό, μπορούμε να έχουμε και καλύτερη διαχείριση των αποθεμάτων αλλά και κατανόηση της αντίστασης σε ασθένειες. Πέρα από την αλληλούχιση του συνολικού DNA (του γονιδιώματος), οι ερευνητές εστιάζουν το ενδιαφέρον τους και στις λειτουργικές πληροφορίες που μπορούν να συλλεχθούν από την ανάλυση του μεταγραφώματος (του συνολικού εκφραζόμενου RNA). Σε αυτό το πλαίσιο, τον Νοέμβριο του 2013 ανακοινώθηκε η έναρξη του προγράμματος 1.000 Fish Transcriptome (Fish-T1K) Project (www.fisht1k.org), που σκοπός του είναι η αλληλούχιση του RNA από 1.000 διαφορετικά είδη ψαριών με υποστήριξη από την Κίνα (CNBG, www.cngb.org) και το Γονιδιωματικό Ινστιτούτο BGI Shenzhen (www.genomics.cn/en/index). Toν Νοέμβριο του 2014 είχαν ήδη συλλεχθεί 7.000 διαφορετικοί ιστοί ιχθύων (εξαιρετικής ποιότητας για αλληλούχιση), που αντιπροσώπευαν το 62% (44/71) των τάξεων και το 28% (112/398) των οικογενειών των Ακτινοπτερυγίων (υπενθυμίζεται ότι σε διαφορετικούς ιστούς εκφράζονται διαφορετικά γονίδια, για αυτό και χρειάζονται τόσες διαφορετικές δειγματοληψίες). Σκοπός είναι να υπάρχουν πληροφορίες από

4 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
τουλάχιστον δύο είδη κάθε τάξης. Οι πληροφορίες για τη λειτουργία των γονιδίων θα έχουν στη συνέχεια πολλαπλές εφαρμογές. Άλλα πεδία στα οποία υπάρχουν εξελίξεις χάρις στη γονιδιωματική είναι ότι είναι δυνατή και η αλληλούχιση γονιδιωμάτων ιών και βακτηρίων παθογόνων για τις ιχθυοκαλλιέργειες, με απώτερο σκοπό να συλλεχθούν πληροφορίες για την καλύτερη αντιμετώπιση τους και τη δημιουργία αντισωμάτων. Επίσης μελετώνται και οι μικροβιακές κοινότητες στο σώμα των ειδών, όπως π.χ. η σύνθεση των μικροβίων στο έντερο του Lates calcarifer και οι αλλαγές της όταν οι ιχθύες είναι σε κατάσταση νηστείας (http://www.biomedcentral.com/1471-2164/15/266). Επίσης μελετάται η σύνθεση των μικροβίων σε διάφορα ανακυκλούμενα συστήματα καλλιέργειας με σκοπό την ανίχνευση παθογόνων οργανισμών (http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0080847). Οι ερευνητικές αυτές δουλειές είναι ακόμη σε πρώτα στάδια (απλής καταγραφής της γονιδιωματικής μικροβιοποικιλότητας) και δεν έχουν ακόμα άμεσες εφαρμογές. Τι πληροφορίες μπορούμε να πάρουμε από τα γονιδιώματα; Το Διεθνές πρόγραμμα αποκωδικοποίησης του γονιδιώματος του είδους Fugu rubripes άρχισε το 1989, με τον καθηγητή Syndney Brenner, και ολοκληρώθηκε το 2002. Το είδος καλλιεργείται στην Ιαπωνία και η σάρκα του χρησιμοποιείται σε υψηλής ποιότητας γεύματα, αν και ορισμένα όργανά του ενδέχεται να περιέχουν θανατηφόρα νευροτοξίνη. Αποτελεί οργανισμό-μοντέλο σε πειράματα διερεύνησης της δομής και της λειτουργίας του γονιδιώματος των οργανισμών. Το γονιδίωμα του F. rubripes, οργανώνεται σε 22 ζευγάρια χρωμοσωμάτων, έχει μέγεθος 365 Mb και είναι 8 φορές μικρότερο από εκείνο του ανθρώπου. Περιέχει λιγότερες επαναλαμβανόμενες αλληλουχίες, γεγονός που κάνει την ανάλυση της λειτουργίας των γονιδίων του σχετικά ευκολότερη από αυτή του ανθρώπου. Η αποκωδικοποίηση του DNA του είδους είναι η πρώτη που ολοκληρώθηκε σε ένα ζωικό είδος. Παρόμοια είναι η οργάνωση του γονιδιώματος του είδους Tetraodon nigrobiridis, που φθάνει τα 385 Mb. Η ανάλυση του γονιδιώματος των δύο αυτών ειδών επιτρέπει να χυθεί άπλετο φως στα μυστικά του γονιδιώματος του ανθρώπου. Κυρίως αποκαλύπτεται η λειτουργία των δομικών γονιδίων και των ρυθμιστικών περιοχών που παρέμειναν συντηρημένες πάνω από 450.000 χρόνια μετά τη διαφοροποίησή τους από κάποιον κοινό πρόγονο. ΄Εχουν αναγνωριστεί περισσότερα από 30.000 γονίδια στα δύο είδη ψαριών, η πλειονότητα των οποίων έχει ταίρι στο γονιδίωμα του ανθρώπου, π.χ. το γονίδιο της δυστροφίνης, μεταλλάξεις του οποίου δημιουργούν την εκφυλιστική νόσο μυϊκή δυστροφία. Οι εξαιρέσεις αφορούν γονίδια του ανοσοποιητικού συστήματος και της ρύθμισης του μεταβολισμού. Η σύγκριση του γονιδιώματος του ανθρώπου με το γονιδίωμα του F. rubripes επέτρεψε ήδη να αποκαλυφθεί η λειτουργία 1.000 γονιδίων του ανθρώπου. Η συγκριτική ανάλυση του γονιδιώματός τους με ηλεκτρονικούς υπολογιστές, έδειξε ότι μεγάλες πολυνουκλεοτιδικές ακολουθίες περιέχουν δεκάδες γονίδια συντηρημένα πάνω στην ίδια χρωμοσωμική περιοχή στον καρυότυπό τους. Το γονιδίωμα του ριζόψαρου έχει μέγεθος 800 εκατομμύρια βάσεις, το ήμισυ του γονιδιώματος του ψαριού ζέβρα. Τα δύο είδη είναι κατάλληλα ως πειραματόζωα για να αποκαλυφθούν μοριακοί μηχανισμοί στις διαδικασίες της ανάπτυξης και της μεταλλαξιγένεσης. Ειδικά το δεύτερο είδος, θεωρείται ιδεώδες πειραματόζωο σε έρευνες για τη γενετική ανάλυσή της ανάπτυξης των σπονδυλωτών. Αυτό οφείλεται στο διαφανές σώμα του εμβρύου που διευκολύνει την οπτική παρατήρηση της ανάπτυξης των ανατομικών δομών του ψαριού.
2.2 Ανάλυση χρωμοσωμάτων Η γενετική σύσταση κάθε είδους ψαριού χαρακτηρίζεται από τη μεγάλη ποσότητα DNA που υπάρχει σε κάθε κύτταρο του. Το DNA, όμως, στον πυρήνα των κυττάρων δεν είναι ένα συνεχές δίκλωνο μόριο, αλλά οργανώνεται σε πολλά ανεξάρτητα ραβδόμορφα χρωμοσώματα. Ο αριθμός των χρωμοσωμάτων ποικίλλει από είδος σε είδος, αλλά είναι σταθερός για κάθε είδος, αν και στα ψάρια έχει παρατηρηθεί κάποια ποικιλότητα ανάμεσα σε άτομα του ίδιου είδους, ακόμη και ανάμεσα και σε κύτταρα του ίδιου ατόμου.

5 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Μερικά είδη ψαριών έχουν 200 χρωμοσώματα. Ο διπλοειδής καρυότυπος της άγριας πέστροφας (Salmo trutta) αποτελείται από 80 χρωμοσώματα, 39 ζευγάρια αυτοσωμάτων και από δύο φυλετικά χρωμοσώματα Χ στα θηλυκά και από ένα Χ και ένα Υ στα αρσενικά άτομα (Εικόνα 5.1).
Πίνακας 5.1 Γονιδιώματα ιχθύων (με ενδιαφέρον για την έρευνα ή τις υδατοκαλλιέργειες) που έχουν δημοσιευθεί μέχρι το 2014. Τα κοινά ελληνικά ονόματα, για όσα από τα είδη υπάρχουν, δίνονται σε παρένθεση δίπλα στο όνομα του είδους).
Είδος Εργασία Δημοσίευσης Χρονολογία Takifugu rubripes http://www.sciencemag.org/content/297/5585/1301.long 2002 Tetraodon nigroviridis http://www.nature.com/nature/journal/v431/n7011/full/nature03025.html 2004 Callorhinchus milii http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0050101 2007 Oryzias latipes http://www.nature.com/nature/journal/v447/n7145/full/nature05846.html 2007 Danio rerio (ψάρι ζέβρα) http://www.nature.com/nature/journal/v496/n7446/full/nature12111.html 2007, 2013 Gadus morhua (μπακαλιάρος Ατλαντικού)
http://www.nature.com/nature/journal/v477/n7363/full/nature10342.html 2011
Lepisosteus oculatus http://www.genetics.org/content/188/4/799.long 2011 Gasterosteus aculeatus (αγκαθερό)
http://www.nature.com/nature/journal/v484/n7392/full/nature10944.html 2012
Petromyzon marinus (λάμπραινα)
http://www.nature.com/ng/journal/v45/n4/full/ng.2568.html 2013
Latimeria chalumnae (κοιλάκανθος)
http://www.nature.com/nature/journal/v496/n7445/full/nature12027.html 2013
Xiphophorus maculatus http://www.nature.com/ng/journal/v45/n5/full/ng.2604.html 2013 Esox lucius (τούρνα) http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0102089 2014 Electrophorus electricus (ηλεκτροφόρο χέλι)
http://www.sciencemag.org/content/344/6191/1522 2014
Oncorhynchus mykiss (ιριδίζουσα πέστροφα)
http://www.nature.com/ncomms/2014/140422/ncomms4657/full/ncomms4657.html 2014
Salmo salar (σολομός) http://www.genomebc.ca/news-events/news-releases/2014/scientific-breakthrough-international-collaboration-has-sequenced-atlantic-salmon-genome/
2014
Η κυτταρογενετική στα ψάρια, ίσως λόγω του μικρού μεγέθους των χρωμοσωμάτων τους, καθιερώθηκε ως επιστήμη αργότερα από αυτήν του ανθρώπου. Σήμερα όμως, στην κυτταρογενετική των ψαριών γίνεται χρήση της χρώσης των χρωμοσωμάτων με ειδικές χρωστικές (π.χ. ζωνώσεις G) ή χρήση μοριακών τεχνικών (FISH), με αποτέλεσμα όχι μόνο να ταυτοποιούνται πλέον όλα τα χρωμοσώματα, αλλά και να αναγνωρίζονται χρωμοσωμικές αναδιατάξεις και άλλες χρωμοσωμικές ανωμαλίες. Επιπλέον, ο συνδυασμός μεθοδολογιών κυτταρογενετικής και μοριακής γενετικής επιτρέπει τη χαρτογράφηση των γονιδίων πάνω στα χρωμοσώματα πολλών ειδών ψαριών.
3. Μοριακή γενετική και εφαρμογές στην ταυτοποίηση και διαχείριση του γενετικού υλικού
3.1 Ανάλυση αλλοενζύμων Οι πρώτες γενετικές έρευνες στα ψάρια άρχισαν τη δεκαετία του 1950 και αφορούσαν παραλλαγές των ομάδων αίματος στον σολομό και τον μπακαλιάρο. Οι μελέτες αυτές έδειξαν την ύπαρξη γενετικά ελεγχόμενης ποικιλότητας, η οποία μπορούσε να χρησιμοποιηθεί στην ανάλυση της γενετικής δομής των πληθυσμών. Μερικά χρόνια αργότερα, μετά την ανακάλυψη της τεχνικής της ηλεκτροφόρησης σε άμυλο και των ιστοχημικών τεχνικών χρώσης, οι έρευνες στους ιχθυοπληθυσμούς συνεχίστηκαν με την ανάλυση των πρωτεϊνικών πολυμορφισμών. Αρχικά, οι μελέτες στηρίζονταν στην ανάλυση πρωτεϊνών, η προσοχή όμως γρήγορα εστιάστηκε στα ένζυμα (ενζυμική ποικιλότητα), στα οποία μετέπειτα βασίστηκαν και οι

6 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
περισσότερες έρευνες. Η εξέταση του γενετικού πολυμορφισμού γινόταν έμμεσα με τη διερεύνηση 10 τουλάχιστον γονιδιακών τόπων που καθορίζουν αλλοένζυμα (τα αλληλόμορφα του ίδιου γονιδίου καθορίζουν διαφορετικές αλλοενζυμικές παραλλαγές με αποτέλεσμα να υπάρχει αλλοενζυμικός πολυμορφισμός) ή άλλες πρωτεϊνικές παραλλαγές (πρωτεϊνικός πολυμορφισμός) σε ηλεκτροφόρηση αμύλου ή πολυακρυλαμιδίου. Ανάμεσα στα πρώτα αποτελέσματα ήταν η περιγραφή παραλλαγών στις αιμοσφαιρίνες του μπακαλιάρου.
Εικόνα 5.1 Τα χρωμοσώματα της άγριας πέστροφας (Salmo trutta) από τον Άγιο Γερμανό (Λίμνη Πρέσπα). ‘Οπου m = μετακεντρικό, a = ακροκεντρικό. Φωτογραφία © Αλέξανδρος Τριανταφυλλίδης.
Η μέθοδος αυτή επέτρεψε την αποκάλυψη αρκετών αλλοενζυμικών και άλλων πρωτεϊνικών πολυμορφισμών και έδωσε τους πρώτους απλούς και εύκολα εφαρμόσιμους γενετικούς δείκτες σε μεγάλης έκτασης έρευνες των φυσικών πληθυσμών των ψαριών. Έτσι, η ηλεκτροφόρηση ενζύμων αποτέλεσε ένα ισχυρό εργαλείο εκτίμησης της γενετικής ποικιλότητας που χαρακτηρίζει κάποιον πληθυσμό, καθώς και των διαφορών στις συχνότητες των αλληλομόρφων ανάμεσα στους διάφορους ιχθυοπληθυσμούς του ίδιου είδους. Μια σύγκριση ανάμεσα στη γενετική διαφοροποίηση των ειδών ψαριών του γλυκού νερού, των ανάδρομων ειδών και των θαλάσσιων ειδών έδειξε ότι η μέση γενετική διαφοροποίηση ανάμεσα σε διάφορες περιοχές είναι της τάξης του 30%, 4% και 1,5%, αντίστοιχα. Η μικρή γενετική διαφοροποίηση στα θαλάσσια ιχθυοαποθέματα ενός είδους μπορεί να αντικατοπτρίζει τη μεγάλη γονιδιακή ροή στο θαλάσσιο περιβάλλον ή τον πολύ μικρό χρόνο που έχει μεσολαβήσει από τον αποχωρισμό τους ώστε να υπάρξει μετρήσιμη γενετική διαφοροποίηση ή ίσως και περιορισμένη διακριτική ικανότητα της μεθοδολογίας. Πράγματι, το μέγεθος της γενετικής ποικιλότητας που αναγνωρίζεται σε έναν πληθυσμό ή είδος με την τεχνική της ηλεκτροφόρησης σε άμυλο ή πολυακρυλαμίδιο είναι ένα ελάχιστο ποσοστό ή με άλλα λόγια είναι υποεκτίμηση της πραγματικής κατάστασης. Γενικά, εκτιμάται ότι η ηλεκτροφόρηση αμύλου μπορεί να αποκαλύψει, κατά προσέγγιση, το 1/3 όλων των πιθανών αντικαταστάσεων των αμινοξέων. Από τη δεκαετία του 1980 και μετά, δίνεται, σχεδόν αποκλειστικά, έμφαση στην άμεση ανάλυση του πολυμορφισμού του DNA. Η άμεση ανάλυση του πολυμορφισμού του DNA αρχικά αφορούσε κυρίως το

7 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
μιτοχονδριακό γονιδίωμα και σε δεύτερο στάδιο το πυρηνικό. Στις επόμενες παραγράφους θα δοθούν συνοπτικά στοιχεία μόνο για τις κυριότερες από τις μεθοδολογίες ανάλυσης του γενετικού υλικού.
3.2 Αλυσιδωτή αντίδραση Πολυμεράσης (PCR) Οι πρώτες έρευνες της ποικιλότητας του mtDNA απαιτούσαν μεγάλες ποσότητες ιστού και χρησιμοποιούσαν χρονοβόρα πρωτόκολλα, ενώ ο αριθμός των ατόμων που εξετάζονταν ήταν περιορισμένος και συχνά ανεπαρκής για αξιόπιστες πληθυσμιακές μελέτες. Η χρησιμοποίηση κατάλληλων εκκινητών του mtDNA και της αλυσιδωτής αντίδρασης της πολυμεράσης (PCR) έκανε την εξέταση της ποικιλότητας του DNA σημαντικά ευκολότερη και γρηγορότερη. Ουσιαστικά, η τεχνική PCR ήταν εκείνη που επέτρεψε την αλματώδη ανάπτυξη της μοριακής γενετικής. Η PCR έχει πολλά πλεονεκτήματα που την καθιστούν σημαντική σε γενετικές μελέτες:
• Είναι πολύ γρήγορη, αφού επιτρέπει την ενίσχυση τμημάτων DNA εκατομμύρια φορές μέσα σε λίγες ώρες.
• Απαιτεί ελάχιστη ποσότητα DNA και, επομένως, ελάχιστη ποσότητα ιστού ή αίματος. Ως εκ τούτου, δεν χρειάζεται να θυσιαστεί ο οργανισμός. Η τεχνική π.χ. είναι τόσο ευαίσθητη, που μπορεί να απομονωθεί και να πολλαπλασιαστεί DNA και από ένα αυγό ψαριού, ή από ένα λέπι.
• Είναι εξαιρετικά ευέλικτη. Αν ο ιστός δεν είναι σε καλή κατάσταση, η τεχνική μπορεί να ενισχύσει άθικτα τμήματα της ακολουθίας του DNA.
Δείτε ακόμη στο διαδίκτυο: https://www.youtube.com/watch?v=DkT6XHWne6E και https://www.youtube.com/watch?v=ZmqqRPISg0g Το μιτοχονδριακό DNA είναι ένα από τα περισσότερο και καλύτερα μελετημένα τμήματα του ζωικού γονιδιώματος. Σε αυτό συνετέλεσαν τα ιδιαίτερα χαρακτηριστικά του, όπως ο υψηλότερος ρυθμός μετάλλαξης του σε σχέση με το πυρηνικό, το μικρό μέγεθός του, κ.λπ. Σήμερα υπάρχουν διαθέσιμοι αρκετοί «παγκόσμιοι εκκινητές», οι οποίοι χρησιμοποιούνται ικανοποιητικά για την ενίσχυση διάφορων περιοχών του mtDNA. Τα ενισχυμένα αυτά τμήματα του DNA αναλύονται στη συνέχεια, είτε με τη βοήθεια ενζύμων περιορισμού (μεθοδολογία PCR-RFLPs), είτε με την εύρεση της νουκλεοτιδικής ακολουθίας τους.
3.3 Μεθοδολογία PCR-RFLPs Η μεθοδολογία του πολυμορφισμού του μήκους των περιοριστικών τμημάτων βασίζεται στη χρήση των ενδονουκλεασών περιορισμού. Τα ένζυμα αυτά αναγνωρίζουν και κόβουν το DNA σε συγκεκριμένες ακολουθίες μήκους συνήθως 4-6 νουκλεοτιδίων. Κάθε ενδονουκλεάση δημιουργεί συγκεκριμένο αριθμό τμημάτων DNA ορισμένου μήκους. Οι αλλαγές στον αριθμό και το μέγεθος των τμημάτων (πολυμορφισμοί) μπορούν να προέλθουν από νουκλεοτιδικές αντικαταστάσεις μέσα στις θέσεις αναγνώρισης, από αναδιατάξεις ακολουθιών και από προσθήκες ή ελλείμματα. Στη μεθοδολογία PCR-RFLPs γίνεται σύγκριση, μεταξύ διαφορετικών ατόμων, των περιοριστικών προτύπων που προκύπτουν από αρκετά ένζυμα περιορισμού, όπως αυτά αναλύονται σε πηκτές ηλεκτροφόρησης αγαρόζης ή πολυακρυλαμιδίου (Εικόνα 5.2). Με τη μεθοδολογία αυτή διερευνάται ο πολυμορφισμός σε αρκετές περιοχές του μιτοχονδριακού γονιδιώματος όπως ο βρόχος εκτόπισης (D-loop), κυτόχρωμα b (cyt-b), οξειδάση του κυτοχρώματος c (COI COII) και 16S ριβοσωμικό DNA (rDNA), ή άλλο μέρος του μιτοχονδριακού γονιδιώματος.
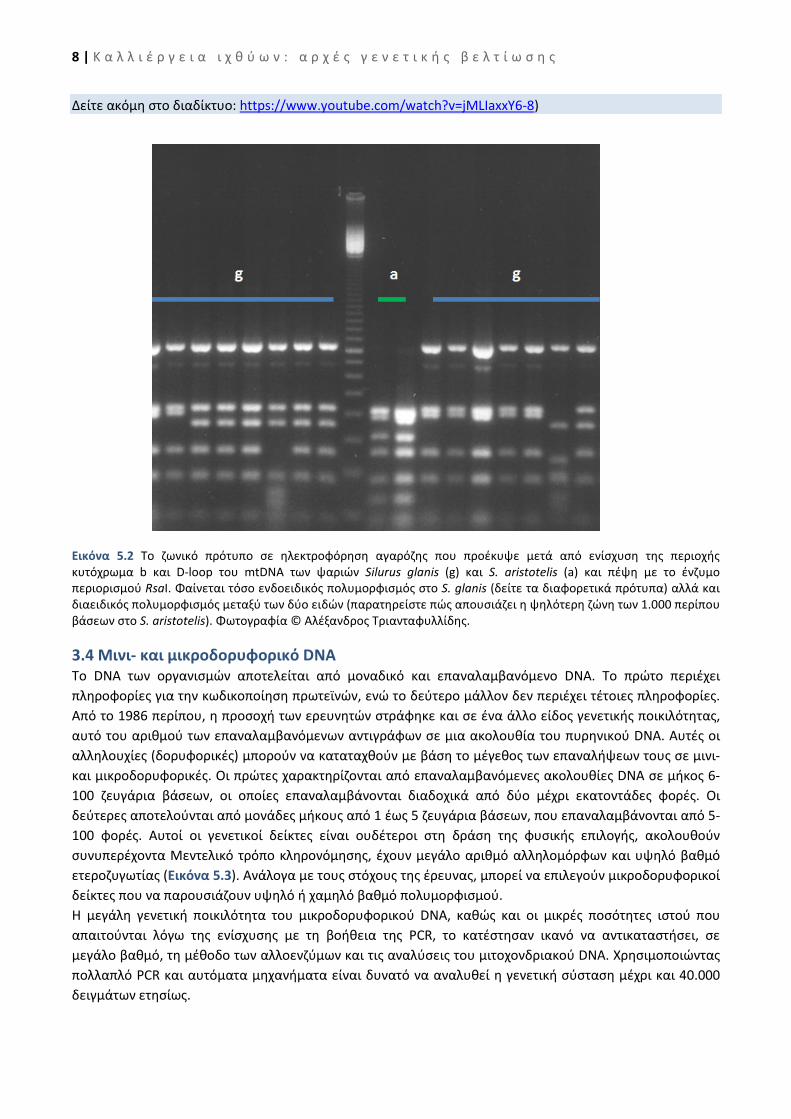
8 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
Δείτε ακόμη στο διαδίκτυο: https://www.youtube.com/watch?v=jMLIaxxY6-8)
Εικόνα 5.2 Το ζωνικό πρότυπο σε ηλεκτρoφόρηση αγαρόζης που προέκυψε μετά από ενίσχυση της περιοχής κυτόχρωμα b και D-loop του mtDNA των ψαριών Silurus glanis (g) και S. aristotelis (a) και πέψη με το ένζυμο περιορισμού RsaI. Φαίνεται τόσο ενδοειδικός πολυμορφισμός στο S. glanis (δείτε τα διαφορετικά πρότυπα) αλλά και διαειδικός πολυμορφισμός μεταξύ των δύο ειδών (παρατηρείστε πώς απουσιάζει η ψηλότερη ζώνη των 1.000 περίπου βάσεων στο S. aristotelis). Φωτογραφία © Αλέξανδρος Τριανταφυλλίδης.
3.4 Μινι- και μικροδορυφορικό DNA Το DNA των οργανισμών αποτελείται από μοναδικό και επαναλαμβανόμενο DNA. Το πρώτο περιέχει πληροφορίες για την κωδικοποίηση πρωτεϊνών, ενώ το δεύτερο μάλλον δεν περιέχει τέτοιες πληροφορίες. Από το 1986 περίπου, η προσοχή των ερευνητών στράφηκε και σε ένα άλλο είδος γενετικής ποικιλότητας, αυτό του αριθμού των επαναλαμβανόμενων αντιγράφων σε μια ακολουθία του πυρηνικού DNA. Αυτές οι αλληλουχίες (δορυφορικές) μπορούν να καταταχθούν με βάση το μέγεθος των επαναλήψεων τους σε μινι- και μικροδορυφορικές. Οι πρώτες χαρακτηρίζονται από επαναλαμβανόμενες ακολουθίες DNA σε μήκος 6-100 ζευγάρια βάσεων, οι οποίες επαναλαμβάνονται διαδοχικά από δύο μέχρι εκατοντάδες φορές. Οι δεύτερες αποτελούνται από μονάδες μήκους από 1 έως 5 ζευγάρια βάσεων, που επαναλαμβάνονται από 5-100 φορές. Αυτοί οι γενετικοί δείκτες είναι ουδέτεροι στη δράση της φυσικής επιλογής, ακολουθούν συνυπερέχοντα Μεντελικό τρόπο κληρονόμησης, έχουν μεγάλο αριθμό αλληλομόρφων και υψηλό βαθμό ετεροζυγωτίας (Εικόνα 5.3). Ανάλογα με τους στόχους της έρευνας, μπορεί να επιλεγούν μικροδορυφορικοί δείκτες που να παρουσιάζουν υψηλό ή χαμηλό βαθμό πολυμορφισμού. Η μεγάλη γενετική ποικιλότητα του μικροδορυφορικού DNA, καθώς και οι μικρές ποσότητες ιστού που απαιτούνται λόγω της ενίσχυσης με τη βοήθεια της PCR, το κατέστησαν ικανό να αντικαταστήσει, σε μεγάλο βαθμό, τη μέθοδο των αλλοενζύμων και τις αναλύσεις του μιτοχονδριακού DNA. Χρησιμοποιώντας πολλαπλό PCR και αυτόματα μηχανήματα είναι δυνατό να αναλυθεί η γενετική σύσταση μέχρι και 40.000 δειγμάτων ετησίως.

9 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Εικόνα 5.3 Το ζωνικό πρότυπο σε πηκτή πολυακρυλαμίδης μικροδορυφορικού τόπου στον γουλιανό. Φωτογραφία © Αλέξανδρος Τριανταφυλλίδης.
3.5 Aπλοί νουκλεοτιδικοί πολυμορφισμοί Τα SNPs μπορούν να οριστούν ως θέσεις απλών ζευγών βάσεων στο γονιδίωμα, στις οποίες συναντώνται διαφορετικές εναλλακτικές ακολουθίες (αλληλόμορφα) στα άτομα του πληθυσμού. Η τεχνική αυτή αποτελεί ένα από τα κυρίαρχα και πιο καινούρια πεδία έρευνας. Τα SNPs θεωρούνται η μέγιστη γενετική πηγή γενοτυπικής ποικιλότητας που διαφοροποιεί τα άτομα μέσα σε ένα είδος. Το έντονο ενδιαφέρον για αυτήν την τεχνική πηγάζει από τη στροφή της έρευνας σε πεδία όπως η μεγάλης κλίμακας ανάλυση του γονιδιώματος αρκετών οργανισμών και η γενετική ανάλυση ασθενειών.
Πίνακας 5.2 Επίπεδα εξελικτικής απόκλισης για τα οποία διάφορες γενετικές μέθοδοι παρέχουν κατάλληλους φυλογενετικούς δείκτες.
Ιεραρχικό Επίπεδο Μιτοχονδριακό DNA
Μικροδορυφορικό DNA
SNPs
Γενετική ταυτότητα - ** ** Πατρότητα * / - ** ** Πληθυσμοί είδους ** ** ** Στενά συγγενικά είδη * * * Ενδιάμεσα ταξινομικά επίπεδα * - - Ανώτερα ταξινομικά επίπεδα * - -
3.6 Πρωτοδιάταξη DNA Σήμερα όλο και περισσότερο χρησιμοποιούνται αυτόματοι αναλυτές (πρώτης, δεύτερης ή τρίτης γενιάς) προσδιορισμού της πρωτοδιάταξης του DNA. Μεθοδολογίες για την αλληλούχιση του DNA με μηχανήματα πρώτης γενιάς (Εικόνα 5.4) έχουν δοθεί σε άλλα μαθήματα. Τα αυτόματα μηχανήματα εύρεσης αλληλουχίας πρώτης γενιάς μπορούν να αναλύουν πια ταυτόχρονα 96 ή ακόμη και 384 δείγματα DNA. Επίσης, έχουν δημιουργηθεί μεγάλα ερευνητικά κέντρα με πολλά αυτόματα μηχανήματα αλληλούχισης, όπου ακόμη και η παραγωγή των DNA τμημάτων προς αλληλούχιση είναι ημι-αυτόματη, αλλά και ο έλεγχος των αποτελεσμάτων είναι αυτοματοποιημένος. Ένα τέτοιο ερευνητικό κέντρο είναι ικανό να παράγει πάνω από 10 Mb δεδομένων την ημέρα. Η πιο επιτυχημένη εταιρεία που εξόπλισε τέτοια ερευνητικά κέντρα είναι

10 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
η Applied Biotechnologies (ABI). Οι βελτιώσεις στην τεχνολογία έχουν επιτρέψει την αύξηση του μέσου μεγέθους ενός διαβάσματος στις 1.000 βάσεις περίπου. Αυτά τα μηχανήματα όμως συνεχίζουν να βασίζονται στην αλληλούχιση μεμονωμένων κλώνων-PCR προϊόντων.
Εικόνα 5.4 Φωτογραφία αυτοραδιογραφίας με τμήμα αλληλουχίας του κυτοχρώματος b μιτοχονδριακού DNA σαυριδιού, ύστερα από αλληλούχηση με μηχάνημα πρώτης γενιάς. Φωτογραφία © Αλέξανδρος Τριανταφυλλίδης.
Οι τεχνολογίες αλληλούχισης του DNA εξελίσσονται συνέχεια. Για την επίτευξη αυτού του σκοπού έχουν εφευρεθεί εντελώς διαφορετικές τεχνολογικές προσεγγίσεις που εξασφαλίζουν ακόμη υψηλότερη ακρίβεια, χαμηλότερο κόστος και γρηγορότερα αποτελέσματα από τα κλασικά ΑΒΙ μηχανήματα τα οποία πια δεν χρησιμοποιούνται στα μεγαλύτερα κέντρα αλληλούχισης. Όντως, χρησιμοποιούνται μηχανήματα επόμενης γενιάς (Next Generation Sequencing, NGS), όπως αυτά των εταιρειών Roche/454 Life Sciences (www.454.com), Solexa/Illumina (www.illumina.com) και SOLiD (της εταιρίας ABI). Η μεγάλη διαφορά τους από τα μηχανήματα πρώτης γενιάς είναι ότι οι αντιδράσεις για την προετοιμασία των προϊόντων προς αλληλούχιση γίνονται παράλληλα (ταυτόχρονα) σε χιλιάδες ή και εκατομμύρια μικροαντιδραστήρες. Τα μηχανήματα 2ης γενιάς αρχικά υπολείπονταν κατά πολύ σε ακρίβεια ανάγνωσης και στο μήκος του τμήματος που διαβαζόταν. Όμως, με συνεχείς βελτιώσεις στη χημεία των αντιδράσεων έχουν ξεπεράσει πια τα αρχικά τους προβλήματα. Έτσι, αρχικά τα νέα μηχάνηματα (Illumina και SOLid) δεν μπορούσαν να διαβάσουν περισσότερα από 35-50 νουκλεοτίδια τη φορά. Το γεγονός αυτό δημιουργούσε προβλήματα στη συναρμολόγηση μεγάλων, πολύπλοκων γονιδιωμάτων. Τώρα είναι δυνατό να φτάσουν άνετα τις 400 (ή και τις 700) βάσεις ανά διάβασμα με 99% ακρίβεια (τα 454 μηχανήματα) και >76 (ή και τις 100) βάσεις με λάθος κάτω από 1% (τα μηχανήματα Illumina). Ταυτόχρονα ήδη εμφανίζονται και τα μηχανήματα αλληλούχισης τρίτης γενιάς (Helicos, Pacific Biosciences & Life Technologies). Η επανάσταση σε αυτά τα μηχανήματα έγκειται στο γεγονός ότι διαβάζουν την αλληλουχία του υπάρχοντος κλώνου και δεν χρειάζονται να ενισχύσουν τον κλώνο για να παίρνουν έντονο σήμα. Και οι τρεις παραπάνω εταιρείες βασίζονται στην ταυτοποίηση του προστιθέμενου νουκλεοτιδίου με βάση εκπομπή φωτός. Εντελώς διαφορετικές προσεγγίσεις, χρησιμοποιούν νανοτεχνολογία και πρωτεΐνες με νανοπόρους. Ανιχνεύουν το κάθε διαφορετικό νουκλεοτίδιο καθώς περνάει μέσα από τους πόρους. Η ανάπτυξη αυτών των μεθόδων θα επιτρέψει την αλληλούχιση αυτής καθαυτής της μητρικής αλυσίδας DNA και θα μειώσει απίστευτα το απαιτούμενο κόστος. Με τα μηχανήματα 3ης γενιάς μπορούμε να κάνουμε πραγματικές ποσοτικές μετρήσεις (π.χ. για τα επίπεδα κάθε RNA ή μεθυλίωσης του DNA). Επίσης, με τον τρόπο αυτόν ξεπερνιούνται τα προβλήματα από την ύπαρξη περιοχών που δεν ενισχύονται (και επομένως δεν αλληλουχίζονται) επειδή έχουν υψηλό ποσοστό GC ή δύσκολες δευτεροταγείς διατάξεις. Επίσης, είναι ακόμα ευκολότερο το διάβασμα από παλαιούς ιστούς στους οποίους έχει καταστραφεί το DNA (από απολιθώματα, μουσεία φυσικής ιστορίας, κ.λπ.) γιατί δεν προστίθεται ακόμη ένα επίπεδο παραγωγής λαθών μέσω της ανάγκης ενίσχυσης του υποστρώματος πριν την αλληλούχιση του.

11 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Δείτε ακόμη στο διαδίκτυο: http://www.youtube.com/watch?v=oYpllbI0qF8, http://www.youtube.com/watch?v=dUjMf2ZezIw, http://www.yourgenome.org/downloads/animations.shtml, https://www.youtube.com/watch?v=womKfikWlxM http://www.youtube.com/watch?v=yVf2295JqUg&feature=BFa&list=PL171604F04548D64E https://nanoporetech.com/technology/the-minion-device-a-miniaturised-sensing-system/the-minion-device-a-miniaturised-sensing-system http://www.dnatube.com/video/3003/SMRT-DNA-Sequencing
3.7 Εφαρμογές Γενετικών Δεικτών Η βασική αρχή όλων των βιοχημικών και μοριακών μεθοδολογιών είναι να χρησιμοποιηθούν κληρονομήσιμοι, καθορισμένοι γενετικοί δείκτες για να αναγνωρίζονται γενότυποι που χαρακτηρίζουν άτομα, πληθυσμούς ή είδη. Οι γενετικοί πολυμορφισμοί που υπάρχουν στους υδρόβιους οργανισμούς μπορούν να χρησιμοποιηθούν ως μοριακοί δείκτες για να προσδιοριστεί ο βαθμός και η κατανομή της γενετικής ποικιλότητας (Πίνακας 5.3). Ο προσδιορισμός αυτής της γενετικής ποικιλότητας στους εκτρεφόμενους οργανισμούς μπορεί να δώσει πληροφορίες σχετικά με τη γενετική σύσταση και τη δομή των πληθυσμών, τη γονιδιακή ροή και τη μετανάστευση πληθυσμών, τη βιολογική ιστορία, το μέγεθος του πληθυσμού, τις φυλογενετικές σχέσεις ανάμεσα σε πληθυσμούς, τη συστηματική των ειδών, τη δημιουργία υβριδίων ανάμεσα σε είδη, την παρακολούθηση της πορείας ενός πληθυσμού ύστερα από εμπλουτισμούς, την αναπαραγωγική συμπεριφορά και τις επιπτώσεις της υπεραλίευσης και της ρύπανσης του περιβάλλοντος στη γενετική ποικιλότητα. Χρήσιμα στοιχεία προκύπτουν επίσης για το μέγεθος της ομομιξίας σε μια μονάδα εκτροφής, την ένταση της φυσικής επιλογής και τη διατήρηση και διαχείριση των αλιευτικών αποθεμάτων. Τέλος, στοιχεία μπορεί να προκύψουν για την ανίχνευση παθογόνων μικροοργανισμών που προσβάλλουν υδατοκαλλιέργειες, μέχρι την ιχνηλάτηση της νοθείας στην εμπορία νωπών ή κατεργασμένων προϊόντων και γενικά στην επίλυση ιατροδικαστικών περιπτώσεων, όπως είναι η γενετική ταυτοποίηση της προέλευσης ενός πληθυσμού ή των γεννητόρων στις υδατοκαλλιέργειες.
3.7.1 Γενετική σύσταση και διαχείριση ιχθύων Η χρήση μοριακών γενετικών δεικτών μπορεί να βοηθήσει στη διερεύνηση πολλών θεμάτων στους φυσικούς ιχθυοπληθυσμούς και τις ιχθυοκαλλιέργειες (Πίνακας 5.3). Η χρήση, για παράδειγμα, τριών μεθόδων γενετικής προσέγγισης βοήθησε στην εξαγωγή ολοκληρωμένων συμπερασμάτων για τη γενετική σύσταση των καλλιεργούμενων και των φυσικών πληθυσμών π.χ. του γουλιανού της Ελλάδας. Η σύγκριση των τριών γενετικών μεθόδων ανάλυσης, οδηγεί στο συμπέρασμα ότι τα υψηλότερα επίπεδα ποικιλομορφίας αποκαλύπτονται με την ανάλυση των μικροδορυφορικών δεικτών και ότι αυτή ενδείκνυται για τη μελέτη σε πληθυσμιακό, αλλά και ατομικό επίπεδο. Επίσης, οι τρεις μεθοδολογίες αποκάλυψαν χαμηλότερα ποσοστά ποικιλομορφίας στους πληθυσμούς γουλιανού από την Ελλάδα, που ίσως οφείλεται στα μικρότερα μεγέθη αυτών των πληθυσμών. Επειδή λοιπόν, η γενετική ποικιλότητα είναι σημαντικότατη για την επιβίωση των πληθυσμών, θεωρείται απαραίτητο να ληφθούν μέτρα που θα προστατέψουν τη γενετική ποικιλομορφία των ειδών γλανιδιού και γουλιανού της Ελλάδας. Ιδιαίτερη αναφορά αξίζουν οι ελληνικοί πληθυσμοί, που εμφανίζουν μοναδικούς γενετικούς χαρακτήρες, γεγονός που πρέπει να ληφθεί υπόψη σε μελλοντικά προγράμματα προστασίας, διαχείρισης και εκμετάλλευσης του γουλιανού. Ένας άλλος, εξίσου σημαντικός, λόγος για τη διατήρηση ή και την αύξηση της γενετικής ποικιλότητας των καλλιεργούμενων ειδών ψαριών, είναι ο ακόλουθος: σε αρκετές περιπτώσεις διαπιστώθηκε θετική συσχέτιση ανάμεσα στη φαινοτυπική ποικιλότητα και τον βαθμό ετεροζυγωτίας γονιδίων που καθορίζουν

12 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
αλλοένζυμα. Αντίθετα, η απώλεια της γενετικής ποικιλότητας έχει αρνητικές επιπτώσεις στην αύξηση, την επιβίωση και τη γονιμότητα τους.
Πίνακας 5.3 Χρήσεις μοριακών γενετικών δεικτών.
Χρήση μοριακών γενετικών δεικτών σε έρευνες φυσικών ιχθυοπληθυσμών Mοναδική γενετική ταυτότητα http://www.tandfonline.com/doi/abs/10.3109/19401736.2010.542242?journalCode=imdn20#.VeDsia1iDS0 Γενετική δομή και κατάσταση των αποθεμάτων (ομομιξία, προσδιορισμός συγγενικών σχέσεων) των ιχθυοπληθυσμών http://www.qub.ac.uk/bb-old/prodohl/GEL/gel.html Αναγνώριση ιχθυοαποθεμάτων που χαρακτηρίζονται από κάποιο βαθμό γενετικής απομόνωσης http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0086809 Εξακρίβωση της φυλογεωγραφικής ιστορίας ειδών και πληθυσμών Π.χ. δείτε τα αποτελέσματα της ομάδας «Conservation genetic management of brown trout (Salmo trutta) in Europe» Αναγνώριση των φυσικών ιχθυοπληθυσμών που χρειάζεται να προστατευτούν http://onlinelibrary.wiley.com/doi/10.1111/1755-0998.12257/abstract Δραστικό μέγεθος ενός ιχθυοπληθυσμού http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1691486/ Χρήση μοριακών γενετικών δεικτών σε έρευνες ιχθυοκαλλιεργειών Αναπαραγωγική αξία κάθε γεννήτορα στις ιχθυοκαλλιέργειες http://www.sciencedirect.com/science/article/pii/S0044848610004576 Επιλογή ή παραγωγή γενετικά βελτιωμένων αποθεμάτων γεννητόρων για ιχθυοκαλλιέργεια http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4264515/ Χαρτογράφηση γονιδίων http://www.biomedcentral.com/1471-2164/15/166 Πραγματοποίηση επιτυχών χειρισμών στο γονιδίωμα διαφόρων ειδών ιχθύων (βλέπε π.χ. τη δουλειά του Dr R. Devlin) http://www.meds-sdmm.dfo-mpo.gc.ca/sdb-bds/profile-profil.do?id=7&lang=eng Ιχνηλάτηση της νοθείας στο εμπόριο νωπών ή κατεργασμένων ιχθύων https://fishpoptrace.jrc.ec.europa.eu/
Χρήση μοριακών γενετικών δεικτών σε έρευνες φυσικών ιχθυοπληθυσμών και ιχθυοκαλλιεργειών Μέτρηση του επιπέδου και των αλλαγών της γενετικής ποικιλότητας στους φυσικούς και τους εκτρεφόμενους πληθυσμούς http://www.sciencedirect.com/science/article/pii/S0044848608006789 Αξιολόγηση των γενετικών επιπτώσεων που επιφέρει η εισαγωγή ή τυχαία απελευθέρωση εκτρεφόμενων ψαριών στους φυσικούς ιχθυοπληθυσμούς. Βλέπε https://aquatrace.eu/ Αξιολόγηση της πορείας των εμπλουτισμών (βλ. παρακάτω)
3.7.2 Γενετική ποικιλότητα και εμπλουτισμοί Πολλοί ιχθυοπληθυσμοί κινδυνεύουν να εξαφανιστούν, ενώ η αφθονία άλλων έχει μειωθεί σημαντικά. Αυτό οφείλεται σε διάφορες ανθρωπογενείς δραστηριότητες, όπως είναι η υποβάθμιση του περιβάλλοντος, η υπεραλίευση και οι εμπλουτισμοί χωρίς σχεδιασμό. Ως εκ τούτου, προτείνονται διάφορα διαχειριστικά σχέδια απελευθέρωσης καλλιεργούμενων οργανισμών σε φυσικούς πληθυσμούς ή μεταφορά ατόμων από έναν τόπο σε άλλον προκειμένου να αποκατασταθούν οι ιχθυοπληθυσμοί. Οι εμπλουτισμοί θεωρούνται απαραίτητοι λόγω μείωσης των ιχθυοαποθεμάτων ή προσδοκιών για αύξηση της αλιείας. Οι

13 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
περισσότεροι θεωρούν ότι οι εμπλουτισμοί έχουν μόνο θετικές επιπτώσεις, μιας και υποβοηθούν τους φυσικούς πληθυσμούς, αλλά αν γίνουν χωρίς τη δέουσα προσοχή μπορεί να οδηγήσουν στη μείωση της ενδοειδικής γενετικής ποικιλότητας, ή ακόμη και στην εξαφάνιση άγριων υδρόβιων οργανισμών, καθώς και των μοναδικών γονιδιακών αποθεμάτων τους. Στην περίπτωση της πέστροφας και του σολομού έχουν γίνει χιλιάδες απελευθερώσεις γόνου, συνήθως σκόπιμες και σπανιότερα τυχαίες. Στη βιβλιογραφία υπάρχει σειρά κανόνων που πρέπει να τηρούνται για τη μείωση των δυσμενών επιπτώσεων από την εισαγωγή εκτρεφόμενων/καλλιεργούμενων οργανισμών στα φυσικά υδάτινα οικοσυστήματα. Στη λίμνη Βόλβη, για παράδειγμα, υπήρχε φυσικός πληθυσμός γριβαδιού. Το 1994 και 1995 έγινε εισαγωγή γόνου γριβαδιού από την κεντρική Ευρώπη. Ανάλυση με μικροδορυφορικούς δείκτες του πληθυσμού, το 2007, έδειξε ότι ο πληθυσμός δεν είναι πια ομοιογενής, αλλά ότι οι εμπλουτισμοί επηρέασαν την αυτόχθονη γενετική σύσταση, επηρεάζοντας και το μέγεθος του. Συμπερασματικά μπορεί να ειπωθεί ότι οι ανεξέλεγκτοι εμπλουτισμοί και η εισαγωγή ξενικών ειδών στα υδάτινα φυσικά οικοσυστήματα θα πρέπει να γίνονται υπό αυστηρό έλεγχο, γιατί μακροπρόθεσμα είναι επιζήμιοι και οδηγούν στα αντίθετα από τα επιδιωκόμενα αποτελέσματα. Καλό είναι να υπάρχει σε κάθε «πείραμα» εμπλουτισμού μια αιτιολογημένη έκθεση σκοπιμότητας, πέρα από την πρόσκαιρη επιχειρηματική διάσταση.
4. Γενετική ποιοτικών χαρακτήρων
4.1 Απλές περιπτώσεις κληρονόμησης ποιοτικών χαρακτήρων Υπάρχουν δύο βασικοί τύποι φαινοτυπικής ποικιλότητας, η ποιοτική και η ποσοτική. Ποιοτική είναι εκείνη που μπορεί να απαριθμηθεί, όχι όμως και να μετρηθεί. Ένα άτομο συνήθως εκφράζει ή όχι έναν ποιοτικό χαρακτήρα, π.χ. αλφικό ή χρωματισμένο γατόψαρο. Η γενετική των ποιοτικών χαρακτηριστικών είναι απλή και αναφέρεται συνήθως ως Μεντελική ή κλασική γενετική. Ένα ποιοτικό χαρακτηριστικό μπορεί να καθορίζεται από ένα ή περισσότερα γονίδια. Τα γονίδια που καθορίζουν ποιοτικά χαρακτηριστικά μπορεί να βρίσκονται σε αυτοσωμικά ή φυλετικά χρωμοσώματα. Ο τρόπος κληρονόμησης ποιοτικών χαρακτηριστικών μπορεί να διαφέρει, π.χ. υπερέχον, υποτελές ή συνυπερέχον αλληλόμορφο.
Πίνακας 5.4 Φαινοτυπικές αναλογίες που προκύπτουν από τη διασταύρωση δύο ετεροζυγωτικών γονέων.
Ένα αυτοσωμικό γονίδιο 3:1 Πλήρης υπεροχή 1:2:1 Ατελής υπεροχή, συνυπεροχή Δύο αυτοσωμικά γονίδια που το κάθε ένα καθορίζει διαφορετικό φαινότυπο 9:3:3:1 Δύο γονίδια με πλήρη υπεροχή 3:6:3:1:2:1 Δύο γονίδια, το ένα με πλήρη υπεροχή και το άλλο με συνυπεροχή 1:2:1:2:4:2:1:2:1 Δύο γονίδια, κάθε γονίδιο δείχνει συνυπεροχή Δύο αυτοσωμικά γονίδια που παράγουν τον φαινότυπο με προσθετική αλληλεπίδραση 1:4:6:4:1 Προσθετική δράση Δύο αυτοσωμικά γονίδια που παράγουν τον φαινότυπο με επιστατική αλληλεπίδραση 12:3:1 Υπερέχουσα επίσταση 9:3:4 Υποτελής επίσταση 9:6:1 Διπλασιασμένα γονίδια με αθροιστική δράση 15:1 Διπλασιασμένα υπερέχοντα γονίδια 9:7 Διπλασιασμένα υποτελή γονίδια 13:3 Υπερέχουσα και υποτελής αλληλεπίδραση
Από τη διασταύρωση ατόμων που διαφέρουν μεταξύ τους σε ένα αυτοσωμικό ποιοτικό χαρακτηριστικό που δείχνει πλήρη υπεροχή ανάμεσα στα δύο αλληλόμορφα, προκύπτει στην F2 γενιά η αναλογία 3:1

14 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
(Εικόνα 5.5). Παρομοίως, από τη διασταύρωση ατόμων που διαφέρουν σε ένα συνυπερέχον ή περισσότερα ποιοτικά χαρακτηριστικά προκύπτουν στην F2 γενιά χαρακτηριστικές φαινοτυπικές αναλογίες, π.χ. 1:2:1, 9:3:3:1, κ.λπ. Το είδος της φαινοτυπικής αναλογίας, όπως είναι γνωστό, εξαρτάται από τον τρόπο κληρονόμησης του χαρακτηριστικού (Εικόνα 5.6) και τον αριθμό των γονιδίων που χρειάζονται για να παραχθεί ο φαινότυπος (Πίνακας 5.4). Η δυνατότητα να τροποποιήσουμε τις συχνότητες των αλληλομόρφων και των φαινοτύπων, για να παραχθεί ένας καθαρός πληθυσμός, είναι πολύ σημαντικό όργανο στη διαχείριση των εκτρεφόμενων πληθυσμών. Με βάση τη Μεντελική γενετική, εύκολα συμπεραίνουμε ότι για ένα αυτοσωμικό γονίδιο με πλήρη υπεροχή, μόνο ομόζυγα (για το υπερέχον ή το υποτελές αλληλόμορφο) άτομα μπορούν να αποτελέσουν καθαρούς πληθυσμούς. Τα ίδια ισχύουν και για τα γονίδια που δείχνουν ατελή υπεροχή. Τα πράγματα όμως είναι πιο περίπλοκα, όταν ένα χαρακτηριστικό εξαρτάται από περισσότερα του ενός γονίδια, ανάμεσα στα οποία ενδέχεται να υπάρχει και αλληλεπίδραση. Παρακάτω δίνουμε ένα παράδειγμα ποιοτικού χαρακτηριστικού, το πρότυπο κατανομής των λεπιών στο κοινό γριβάδι, για να δείξουμε τη συσχέτιση ανάμεσα στη γενετική σύσταση και την οικονομική σπουδαιότητα ενός πληθυσμού.
Εικόνα 5.5 Η κληρονόμηση του αλφικού χρώματος στα γατόψαρα.
4.2 Επιστατική αλληλεπίδραση Η κατανομή των λεπιών ελέγχεται από δύο γονίδια, τα S και N (Πίνακας 5.5). Το N είναι υπερέχον επιστατικό, ενώ ταυτόχρονα με την πλειοτροπική του δράση συμπεριφέρεται και ως υποτελές θανατογόνο γονίδιο. Το N τροποποιεί το πρότυπο των λεπιών. Αντίθετα, το υποτελές αλληλόμορφο n, δεν επηρεάζει το πρότυπό τους. Το γονίδιο S ελέγχει τον αριθμό των λεπιών και δείχνει πλήρη υπεροχή πάνω στο αλληλόμορφο του s. Άτομα της γενοτυπικής σύστασης S/S ή S/s έχουν το κανονικό πρότυπο λεπιών (λέπια διασκορπισμένα σε όλο το σώμα – λεπιδωτή ποικιλία). Ομόζυγα άτομα ss έχουν πολύ μειωμένο αριθμό λεπιών. Μάλιστα τα λίγα λέπια είναι σχετικά μεγάλα και ακανόνιστα διατεταγμένα στο σώμα τους (καθρεπτοειδής ποικιλία). Μία δόση του αλληλόμορφου N μετατρέπει: α) τον λεπιδωτό φαινότυπο του γριβαδιού στον γραμμικό φαινότυπο (μικρά λέπια που περιορίζονται κατά μήκος της ράχης και μεγάλα λέπια κατά μήκος της πλευρικής γραμμής) και β) τον καθρεπτοειδή φαινότυπο στον γυμνό φαινότυπο (σχεδόν πλήρης απουσία λεπιών). Από τα παραπάνω προκύπτει ότι οι πιθανοί 9 γενότυποι, δίνουν 4 φαινοτύπους (Εικόνα 5.7) των διάφορων ποικιλιών του γριβαδιού (μιας και οι τρεις γενότυποι είναι

15 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
θανατογόνοι). Από τη διασταύρωση ατόμων με γραμμικό φαινότυπο που είναι ετερόζυγα και για τα δύο γονίδια: P1 Ss Nn X Ss Nn, προκύπτει η φαινοτυπική αναλογία: 6 γραμμικά : 2 γυμνά : 3 λεπιδωτά : 1 καθρεπτοειδές, αντί της φαινοτυπικής αναλογίας 12 : 3 : 1, επειδή πεθαίνουν 4 άτομα λόγω της θανατογόνου δράσης του αλληλόμορφου Ν.
Εικόνα 5.6 Η κληρονόμηση ενός φυλοσύνδετου γνωρίσματος στο είδος Xiphophorus maculates με σύστημα καθορισμού του φύλου WZ/ZZ.
Από τη μελέτη των γενοτύπων και των φαινοτύπων προκύπτει ότι μόνον άτομα με καθρεπτοειδή φαινότυπο (Πίνακας 5.5), δηλαδή άτομα του γενότυπου ss nn, μπορούν να αποτελέσουν καθαρούς πληθυσμούς. Από την άλλη μεριά, είναι δύσκολο να προκύψει καθαρός λεπιδωτός πληθυσμός, επειδή άτομα με τέτοιο φαινότυπο μπορεί επίσης να είναι της γενοτυπικής σύστασης SS nn ή Ss nn. Μόνο όταν είναι σίγουρο ότι στον γενότυπο του γόνου δεν υπάρχει το αλληλόμορφο s, κάτι που μπορεί να αποδειχθεί με διασταυρώσεις, ο πληθυσμός του ιχθυοτροφείου θα είναι καθαρός.
Πίνακας 5.5 Οι γενότυποι ατόμων γριβαδιών για τα γονίδια που επηρεάζουν την κατανομή των λεπιών και οι αντίστοιχοι φαινότυποι
Γενότυποι Φαινότυποι
SS nn Λεπιδωτός (κανονικός) Ss nn Λεπιδωτός (κανονικός) ss nn Καθρεπτοειδής SS Nn Γραμμικός Ss Nn Γραμμικός ss Nn Γυμνός SS NN Νεκρά άτομα Ss NN Νεκρά άτομα ss NN Νεκρά άτομα

16 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
Εικόνα 5.7 Διαφορετικοί φαινότυποι στο κοινό γριβάδι (κυπρίνο) με βάση το πρότυπο κατανομής των λεπιών. Τροποποιημένη άπό http://www.fao.org/docrep/005/b3310e/B3310E14.htm
Τα άτομα με μειωμένο αριθμό λεπιών αγοράζονται σε ψηλές τιμές στις περισσότερες περιοχές της κεντρικής και βόρειας Ευρώπης. Στην Ελλάδα και την Ασία οι κάτοικοι προτιμούν τα γριβάδια που έχουν λεπιδωτό φαινότυπο. Ο γυμνός φαινότυπος πουλιέται ακριβότερα στην υπόλοιπη Ευρώπη, γιατί το ψάρι καθαρίζεται ευκολότερα . Είναι εύκολο να κατανοηθεί γιατί οι προσπάθειες δημιουργίας καθαρής γυμνής ποικιλίας έχουν αποτύχει (οι γεννήτορες είναι ετερόζυγοι για το υποτελές θανατογόνο γονίδιο Ν). ΄Ισως αξίζει να αναφερθεί ότι, από διαχειριστική σκοπιά, κοστίζει περισσότερο να παραχθούν ο γραμμικός ή γυμνός φαινότυπος, παρά ο καθρεπτοειδής φαινότυπος.
4.3 Πλειοτροπία Ένα γονίδιο είναι δυνατόν να επηρεάζει όχι μόνο ένα, αλλά πολλά και διαφορετικά χαρακτηριστικά, οπότε λέμε ότι το γονίδιο είναι πλειοτροπικό. Για ένα διαχειριστή μονάδας, όταν οι πλειοτροπικές επιδράσεις ενός γενοτύπου επηρεάζουν οικονομικά χαρακτηριστικά, τότε οι επιδράσεις αυτές γίνονται σπουδαιότερες από αυτό καθαυτό τον γενότυπο (και τον φαινότυπο). Στο Ισραήλ π.χ. διαπιστώθηκε ότι γριβάδια με μπλε (bb) και χρυσό (gg) χρώμα έχουν μικρότερη ταχύτητα αύξησης από γριβάδια που έχουν κανονικό χρώμα σώματος. Αντίθετα, τα Πολωνικά μπλε γριβάδια αναπτύσσονται ταχύτερα από τα κανονικά. Οι διαφορετικές πλειοτροπικές επιδράσεις του ίδιου γενοτύπου δείχνουν ή ότι ο μπλε φαινότυπος των γριβαδιών στις δύο χώρες οφείλεται σε διαφορετικά γονίδια ή ότι οφείλεται σε γονίδια που είναι στενά συνδεμένα με το γονίδιο για το μπλε χρώμα. Όπως αναφέρθηκε, πλειοτροπική δράση δείχνουν και τα γονίδια που ελέγχουν το πρότυπο των λεπιών στο γριβάδι. ΄Ατομα π.χ. με λεπιδωτό και καθρεπτοειδή φαινότυπο (γενότυποι S/- nn και ss nn), ζουν 210 λεπτά σε περιβάλλον με απουσία οξυγόνου, ενώ άτομα με γενότυπους S/- Ν/- και ss Ν/- μόνο 32 λεπτά.
4.4 Επιλογή ποιοτικών χαρακτηριστικών Από τα παραπάνω γίνεται φανερό πόσο απαραίτητη είναι η γνώση του τρόπου κληρονόμησης των χαρακτηριστικών. Μόνο όταν αυτός ο τρόπος είναι γνωστός, μπορεί να καταστρωθεί το σωστό πρόγραμμα διαχείρισης ώστε να μεταβληθεί (τροποποιηθεί) η φαινοτυπική, γενοτυπική και γονιδιακή συχνότητα του πληθυσμού σε ένα εκτρεφόμενο είδος. Είμαστε δηλαδή σε θέση να επιτύχουμε επιθυμητούς φαινοτύπους και να περιορίσουμε ή να εξαλείψουμε τους ανεπιθύμητους μέσω επιλογής. Η επιλογή είναι ιδιαίτερα χρήσιμη στις περιπτώσεις που οι ποιοτικοί χαρακτήρες παίζουν σημαντικό ρόλο στην εμπορική αξία του προϊόντος, π.χ. εκτροφή κυπρινοειδών με παρουσία ή απουσία λεπιών ή συγκεκριμένο σχήμα σώματος. Η επιλογή αποτελεί κλασική μέθοδο γενετικής βελτίωσης και χρησιμοποιείται με επιτυχία στις ιχθυοκαλλιέργειες, τόσο των διακοσμητικών όσο και των εδώδιμων ειδών.

17 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Βασική προϋπόθεση για κάθε πρόγραμμα επιλογής είναι η γνώση στοιχείων γενετικής πληθυσμών. Π.χ. σε έναν πληθυσμό γατόψαρου υπάρχουν άτομα με τους δύο φαινοτύπους: αλφικά (ομόζυγα α/α) και κανονικά χρωματισμένα (γενότυπος +/α ή +/+). Είναι εύκολο να συνάγουμε ότι μέσα σε μια γενιά είναι δυνατόν να δημιουργηθεί καθαρό στέλεχος για τον αλφικό φαινότυπο. Ή, με άλλα λόγια, μια γενιά επιλογής ενάντια στο υπερέχον κανονικό αλληλόμορφο θα δημιουργήσει καθαρό αλφικό πληθυσμό. Αντίθετα, η επιλογή ενάντια σε ένα υποτελές αλληλόμορφο (και φαινότυπο) είναι πολύ δύσκολο, αν όχι αδύνατο, να οδηγήσει στην πλήρη εξαφάνιση του υποτελούς και στην πλήρη ομοζυγωτία του υπερέχοντος αλληλομόρφου σε όλα τα άτομα του πληθυσμού. Για αυτό συνήθως το ερώτημα που τίθεται είναι: Πόσες γενιές επιλογής θα χρειαστούν για να μειωθεί η συχνότητα του υποτελούς ανεπιθύμητου αλληλομόρφου σε κάποιο επιθυμητό επίπεδο; Παράδειγμα: Στο γατόψαρο, το αλληλόμορφο α καθορίζει τον αλφικό φαινότυπο. Άτομα με τέτοιο φαινότυπο ωοτοκούν αργότερα από τα κανονικά, παράγουν λιγότερα αυγά και οι απόγονοί τους έχουν μικρότερη βιωσιμότητα. Πόσες, λοιπόν, γενιές επιλογής απαιτούνται ενάντια στον αλφικό φαινότυπο για να μειωθεί η συχνότητα του αλληλόμορφου α σε τέτοιο επίπεδο, ώστε να είναι καθαρός ο πληθυσμός σε ποσοστό 99,99%; Η απάντηση μπορεί να βρεθεί χρησιμοποιώντας τον τύπο: N = 1/qn – 1/q0, όπου Ν είναι ο αριθμός των γενιών, qn είναι η επιθυμητή συχνότητα του υποτελούς αλληλομόρφου στη γενιά Ν, στο παράδειγμά μας 0,01%, και q0 είναι η αρχική συχνότητα του υποτελούς αλληλόμορφου α. ΄Εστω, ότι στο παράδειγμά μας η συχνότητα των αλφικών ατόμων (των ομόζυγων γενοτύπων) ήταν 9%. Λύση: Η συχνότητα του αλληλόμορφου α στη μηδενική γενιά ήταν: q0 =√0,09 και q0 = 0,03. Η συχνότητα του αλληλόμορφου α στη γενιά Ν θα είναι: qn =√0,0001 και qn = 0,01. Ο αριθμός των γενιών που χρειάζονται για να μειωθεί η συχνότητα του α από 0,3 σε 0,01 θα είναι: N = 1/0,01 – 1/0,3 = 96,67 γενιές. Με άλλα λόγια, ο ιχθυοτρόφος θα χρειαστεί 97 γενιές. Αφού η επιλογή ενάντια σε υποτελή αλληλόμορφα δεν φαίνεται και τόσο αποτελεσματική, δεν θα πρέπει να καταλήξουμε στο συμπέρασμα πως είναι αδύνατο να δημιουργηθούν καθαροί πληθυσμοί για υπερέχοντα γονίδια. Συνήθως, αυτό επιτυγχάνεται ελέγχοντας τον γενότυπο ενός γονέα, με βάση τη μελέτη των απογόνων που προκύπτουν από μια διασταύρωση ελέγχου με ομόζυγα άτομα για το υποτελές αλληλόμορφο. Έτσι αν ο γονέας είναι ετερόζυγος, η διασταύρωση (+/α) Χ (α/α) θα μας δώσει και απογόνους με τον υποτελή φαινότυπο, ενώ αν είναι ομόζυγος η διασταύρωση (+/+) Χ (α/α) θα μας δώσει μόνο κανονικούς απογόνους. Στην περίπτωση που τα αλληλόμορφα είναι συνυπερέχοντα έχουμε: Έστω ότι το χρώμα στην πέστροφα καθορίζεται από ένα αυτοσωμικό γονίδιο, το G, με δύο συνυπερέχοντα αλληλόμορφα, τα G και G΄. Το G σε ομοζυγωτία ελέγχει το χρυσαφί χρώμα, το G΄ σε ομοζυγωτία ελέγχει το ασημί χρώμα, ενώ άτομα με γενετική σύσταση G/G΄ έχουν ασημοχρυσαφί χρώμα. Στην περίπτωση αυτή, όπου ο γενότυπος υποδεικνύει και τον φαινότυπο του ψαριού, εύκολα, με απλή επιλογή του φαινοτύπου των γεννητόρων, επιτυγχάνεται η παραγωγή των απογόνων με επιθυμητό φαινότυπο.
5. Γενετική των ποσοτικών χαρακτήρων
5.1 Βασικές αρχές ποσοτικής γενετικής Ποσοτική φαινοτυπική ποικιλότητα είναι εκείνη που μπορεί να μετρηθεί. Περιλαμβάνει χαρακτηριστικά όπως είναι το βάρος, το μήκος, ο αριθμός των ωαρίων ανά θηλυκό, η εκκολαψιμότητα, κ.λπ. Ένα ποσοτικό χαρακτηριστικό μπορεί να καθορίζεται από 2, 3, 10 ή περισσότερα γονίδια. Συνήθως, ο αριθμός των γονιδίων που ελέγχουν τους φαινοτύπους είναι άγνωστος, καθώς επίσης είναι άγνωστος και ο τρόπος της γονιδιακής έκφρασης. Εξαιτίας μάλιστα της απουσίας αυτών των απαραίτητων γενετικών στοιχείων, είναι πολύ δύσκολο να παραχθεί καθαρός πληθυσμός για έναν συγκεκριμένο ποσοτικό φαινότυπο. Δυστυχώς όμως, τα περισσότερα ποσοτικά γνωρίσματα, όπως η βιωσιμότητα και ο ρυθμός αύξησης, είναι εκείνα που

18 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
ενδιαφέρουν την εμπορική υδατοκαλλιέργεια ή την παραγωγή γόνου που προορίζεται για εμπλουτισμούς. Αντίθετα, τα ποιοτικά χαρακτηριστικά είναι εκείνα που ενδιαφέρουν όσους ασχολούνται με την εκτροφή κυρίως καλλωπιστικών ειδών ψαριών. Τα ποσοτικά χαρακτηριστικά δείχνουν συνεχή ποικιλότητα και κατανομή και γι’ αυτό συνήθως περιγράφονται με τη μέση τιμή τους, τη διακύμανση, την τυπική απόκλιση, το τυπικό σφάλμα, το εύρος, κ.λπ. Η επιλογή για ένα ποσοτικό χαρακτηριστικό, δηλαδή η αύξηση ή ελάττωση της φαινοτυπικής μέσης τιμής του, δεν είναι παρά μια μεταφορά της καμπύλης της διασποράς, από τη σημερινή μέση τιμή του πληθυσμού προς τα δεξιά (μεγαλύτερη) ή αριστερά (μικρότερη) της μέσης τιμής κατανομής. Το σημείο επιλογής, δηλαδή η τιμή η οποία θεωρείται ως όριο για την επιλογή, καθορίζεται από τον παραγωγό ή καλύτερα από εδικούς επιστήμονες, με ταυτόχρονη προσπάθεια να εξαλειφθούν οι περισσότεροι ανεπιθύμητοι φαινότυποι. Η συνολική ποικιλομορφία (Vp) που παρατηρείται σε ένα ποσοτικό χαρακτηριστικό οφείλεται κατά ένα μέρος στη γενετική διακύμανση (Vg), κατά ένα άλλο μέρος στην περιβαλλοντική διακύμανση (Ve) και, τέλος, κατά ένα άλλο μέρος στη διακύμανση που οφείλεται στην αλληλεπίδραση περιβάλλοντος και γενετικής σύστασης (Vg-e). Έτσι έχουμε:
Vp = Vg + Ve + Vg-e Από την εξίσωση αυτή προκύπτει ότι τον βελτιωτή θα πρέπει να ενδιαφέρει κυρίως η γενετική διακύμανση. Και αυτό γιατί με το πρόγραμμα βελτίωσης θα προσπαθήσει να διερευνήσει ή να τροποποιήσει τη γενετική σύσταση του πληθυσμού, προκειμένου να αυξήσει την παραγωγικότητα της μονάδας του και φυσικά τα κέρδη του. Και η γενετική όμως διακύμανση είναι σύνθετη. Αποτελείται από την προσθετική ή αθροιστική διακύμανση (Va), τη γενετική διακύμανση που οφείλεται στην υπεροχή, την υποτέλεια ή την υπερκυριαρχία (Vd), και τη διακύμανση που οφείλεται στην επιστατική αλληλεπίδραση των γονιδίων που βρίσκονται σε διαφορετικές θέσεις (Vi), δηλ.
Vg = Va + Vd + Vi Η τιμή του Vi είναι δύσκολο να εκτιμηθεί. Γι’ αυτό στα προγράμματα βελτίωσης συνήθως γίνεται διερεύνηση των παραμέτρων Va και Vd. Η Va είναι το αποτέλεσμα της λειτουργίας των αλληλομόρφων και κληρονομείται με προβλεπόμενο και επαναλήψιμο τρόπο. Αντίθετα, η Vd είναι η λειτουργία όλου του γενοτύπου, αλληλεπίδραση μεταξύ όλων των αλληλομόρφων, και δεν κληρονομείται, μιας και σε κάθε γενιά δημιουργούνται νέοι συνδυασμοί. Από τα προηγούμενα προκύπτει ότι οι δύο παράμετροι είναι εντελώς διαφορετικές συνιστώσες της γενετικής διακύμανσης. Έτσι, δεν είναι παράδοξο πως για να βελτιωθεί η παραγωγικότητα κάποιου πληθυσμού και να διερευνηθεί κάθε τύπος γενετικής διακύμανσης θα πρέπει να χρησιμοποιηθούν διαφορετικά προγράμματα βελτίωσης. Για την Va χρησιμοποιείται η επιλογή, ενώ για την Vd ο υβριδισμός. Ένας από τους σπουδαιότερους παράγοντες για τη διατύπωση προγραμμάτων βελτίωσης των οργανισμών είναι η γνώση της σχετικής συνεισφοράς των γονιδίων στο χαρακτηριστικό που εξετάζεται. Με άλλα λόγια, ο βελτιωτής είναι απαραίτητο να γνωρίζει ποιο μέρος της Vp οφείλεται στην Va για να μπορέσει να εκτιμήσει αν το χαρακτηριστικό θα είναι ευαίσθητο στην επιλογή (αν είναι δηλαδή επιλέξιμο). Ο λόγος της φαινοτυπικής διακύμανσης (Vp) ενός πληθυσμού που οφείλεται σε γενετικούς παράγοντες, ονομάζεται κληρονομησιμότητα. Συμβολίζεται ως h2, είναι δε: h2 = Va/ Vp. Από τη στιγμή που υπολογιστεί ή βρεθεί από τη βιβλιογραφία (Πίνακας 5.6) η τιμή του συντελεστή κληρονομησιμότητας, μπορεί να εκτιμηθεί και ο βαθμός ανταπόκρισης του χαρακτηριστικού στην επιλογή από τον τύπο: R = Sh2, όπου R είναι η ανταπόκριση (γενετικό κέρδος ή απώλεια) ανά γενιά, και S ο διαφορικός συντελεστής επιλογής (η μειονεκτικότητα ή πλεονεκτικότητα) των επιλεγόμενων γεννητόρων σε σχέση με τον μέσο πληθυσμό. Από τον προηγούμενο τύπο βγαίνει πως η επιτυχία ή αποτυχία κάθε προγράμματος επιλογής καθορίζεται από την τιμή του συντελεστή κληρονομησιμότητας του χαρακτηριστικού.
19 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Αν h2 = 1, τότε R = S. Αν h2 = 0, τότε και R = 0 Ας σημειωθεί ότι μπορεί να τροποποιηθούν με επιλογή μόνο εκείνοι οι φαινότυποι των οποίων ο συντελεστής κληρονομησιμότητας είναι μεγαλύτερος ή ίσος με 0,25. Γενικά, οι πληθυσμοί που για συγκεκριμένο ποσοτικό χαρακτηριστικό παρουσιάζουν μεγάλη απόκλιση από τη μέση τιμή, προσφέρονται καλύτερα για προγράμματα επιλογής, από ότι χαρακτηριστικά των οποίων η απόκλιση κυμαίνεται πολύ κοντά στη μέση τιμή. Το ίδιο ισχύει και για τα χαρακτηριστικά που έχουν υψηλό βαθμό κληρονομησιμότητας.
Πίνακας 5.6 Τιμές h2 σε διάφορα είδη ψαριών και οστρακοειδών.
Είδος Γενότυπος h2
Ιριδίζουσα πέστροφα Βάρος σε 150 ημέρες 0,50 Μήκος σε 2 χρόνια 0,26 Ανθεκτικότητα σε DDT 0,06 Γριβάδι Βάρος σε 4 μήνες 0,48 Μήκος σε 2 χρόνια 0,55 Ρυθμός αύξησης 0,25 Ευρωπαϊκό στρείδι (Ostrea edulis) Ρυθμός αύξησης 0,11-0,24 Στρείδι Ειρηνικού (Crassostrea gigas) Επιβίωση «Καλοκαι-
ρινής Θνησιμότητας» 0,63
Χτένι Ατλαντικού (Argopecten irradians) Ρυθμός αύξησης 0,21
Παράδειγμα. Γριβάδια ορισμένης ηλικίας έχουν μέσο βάρος 300 g. Ένας καλλιεργητής επιλέγει 50 θηλυκά άτομα με μέσο βάρος 340 g και 50 αρσενικά με μέσο βάρος 360 g. Ποιο θα είναι το μέσο βάρος του πληθυσμού την επόμενη γενιά; Λύση. Η τιμή h2 για την ταχύτητα αύξησης είναι ίση με 0,25. Η τιμή του διαφορικού συντελεστή επιλογής S υπολογίζεται ως εξής: S = (X επιλεγόμενων θηλυκών + X επιλεγόμενων αρσενικών)/2 - Χ πληθυσμού, (όπου Χ = μέση τιμή) S = (340 – 360)/2 - 300 = 50 g R = S h2 = 50 x 0,25 = 12,5 g Ύστερα από μια γενιά επιλογής το μέσο βάρος του πληθυσμού θα είναι: 300 + 12,5 = 312,5 g. Από τη στιγμή που θα αποφασιστεί ότι για τη βελτίωση ενός πληθυσμού απαιτείται η εφαρμογή προγράμματος επιλογής, το επόμενο στάδιο είναι να αποφασιστεί το είδος του προγράμματος. Από τα μαθήματα της εξέλιξης είναι γνωστό ότι τα προγράμματα επιλογής που μπορούν να εφαρμοστούν είναι: η κατευθύνουσα επιλογή, η σταθεροποιούσα επιλογή, κ.λπ. Αν στο πρόγραμμα επιλέξουμε τη “μη επιλογή”, αυτό σημαίνει διατήρηση της μεγαλύτερης δυνατής διακύμανσης, δηλαδή της πλουσιότερης γενετικής δεξαμενής, έτσι ώστε οι απόγονοι να μεταφέρουν όλες τις γενετικές πληροφορίες. Τέτοιοι απόγονοι έχουν τη δυνατότητα να αντιμετωπίσουν όλες τις πιθανές αντιξοότητες και τα περιβαλλοντικά προβλήματα που ενδέχεται να προκύψουν. Η “μη επιλογή” είναι ιδιαίτερα χρήσιμη, για πληθυσμούς που πρόκειται να χρησιμοποιηθούν για εμπλουτισμούς υδατοσυλλογών.
5.2 Υβριδισμός Αν από τη βιβλιογραφία βρεθεί ότι η τιμή του Va είναι μικρή ή ότι είναι δύσκολο να βελτιωθεί ένας φαινότυπος με επιλογή, η εναλλακτική λύση προκειμένου να αυξηθεί η παραγωγικότητά του πληθυσμού

20 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
είναι ο υβριδισμός (ετερομιξία). Βέβαια, θα πρέπει να γνωρίζουμε ότι ο υβριδισμός μπορεί να χρησιμοποιηθεί για τη βελτίωση ενός πληθυσμού ακόμη και όταν η τιμή του Va είναι μεγάλη. Το Vd είναι η γενετική διακύμανση που παράγεται από την αλληλεπίδραση των αλληλομόρφων κάθε γονιδίου. Επειδή αυτή η μορφή γενετικής διακύμανσης εξαρτάται από αλληλεπιδράσεις αλληλομόρφων, αποδιοργανώνεται σε κάθε μειωτική διαίρεση και γι’ αυτό δεν κληρονομείται από τους γονείς στους απογόνους. Κατά συνέπεια, με τον υβριδισμό επιδιώκεται να βρεθεί ποιοι γεννήτορες θα δώσουν απογόνους με τέτοιους συνδυασμούς αλληλομόρφων, ώστε να αυξάνεται η παραγωγικότητά τους. Με απλά λόγια, ο υβριδισμός έχει ως στόχο τη μεταβολή του μέρους εκείνου της γενετικής διακύμανσης που δεν κληρονομείται, αλλά προκύπτει ως αποτέλεσμα της αλληλεπίδρασης των αλληλομόρφων, η οποία είναι μοναδική σε κάθε ψάρι. Συνεπώς, η προσπάθεια στοχεύει στο να αποκαλυφθούν οι συνδυασμοί που πρέπει να προκληθούν, ώστε να παραχθούν οι κατάλληλοι απόγονοι. Στον υβριδισμό, οι απόγονοι της νέας γενιάς δεν μοιάζουν συνήθως φαινοτυπικά με τους γεννήτορες. Αν μάλιστα πρόκειται για την πρώτη πειραματική προσπάθεια, δεν μπορεί να προβλεφθεί εύκολα το αποτέλεσμα μιας τέτοιας διασταύρωσης. Μπορεί, για παράδειγμα, να διασταυρωθούν δύο βελτιωμένα άτομα ή στελέχη και να παραχθούν μειονεκτικοί απόγονοι ή, αντίθετα, μπορεί να διασταυρωθούν δύο μειονεκτικές σειρές και να παραχθούν απόγονοι με μεγάλη απόδοση. Από τη στιγμή όμως που θα βρεθεί ο άριστος συνδυασμός αλληλομόρφων, μπορεί να επαναλαμβάνεται η ίδια διασταύρωση για να αυξάνεται η παραγωγικότητα. Ωστόσο, δεν θα πρέπει να θεωρηθεί ότι υπάρχει ένας και μοναδικός τρόπος υβριδισμού. Ο υβριδισμός χρησιμοποιείται:
• Ως το πρώτο στάδιο παραγωγής του πληθυσμού στον οποίο θα εφαρμοστεί το πρόγραμμα επιλογής.
• Ως το τελευταίο στάδιο μετά το πέρας του προγράμματος επιλογής. Στην περίπτωση αυτή, με το πρόγραμμα επιλογής επιλέγονται δύο γραμμές, τα υβρίδια των οποίων, σύμφωνα με τα πειράματα, έχουν βελτιωμένη παραγωγή. Με αυτόν τον τρόπο, δημιουργείται μια νέα γενετική δεξαμενή, ανάμεσα σε δύο διαφορετικές επιθυμητές γενεαλογικές γραμμές.
• Για την παραγωγή νέων βελτιωμένων στελεχών. • Για την παραγωγή μονοφυλετικών απογόνων. • Για την παραγωγή ομοιόμορφων απογόνων, μιας και οι καταναλωτές συνήθως προτιμούν προϊόντα
που γνωρίζουν ποιοτικά. • Για την παραγωγή ατόμων στείρων ή με αδυναμία αναπαραγωγής, με σκοπό τη διενέργεια
εμπλουτισμών.
5.2.1 Σχεδιασμός πειραμάτων υβριδισμού Αφού τα αποτελέσματα του υβριδισμού δεν μπορούν να προβλεφθούν, για τον σωστό σχεδιασμό τέτοιων πειραμάτων πρέπει να έχουμε υπόψη μας και τα ακόλουθα:
• Όσο πιο απομακρυσμένοι φυλογενετικά είναι οι δύο γονικοί πληθυσμοί, τόσο μικρότερη είναι η πιθανότητα να γίνει πετυχημένος υβριδισμός ανάμεσά τους και αντίστροφα.
• Όσο πιο όμοιοι είναι οι καρυότυποι των δύο γεννητόρων, τόσο μεγαλύτερη είναι η πιθανότητα να γίνει πετυχημένος υβριδισμός ανάμεσά τους και αντίστροφα.
• Όσο πιο απομακρυσμένη είναι η βιολογία, η οικολογία και η αναπαραγωγική συμπεριφορά δύο γεννητόρων (δύο ειδών), τόσο μικρότερη είναι η πιθανότητα διασταύρωσης τους στο φυσικό περιβάλλον και αντίστροφα. Η χρησιμοποίηση ορμονών ή άλλων πειραματικών μέσων μπορεί να παρακάμψει προβλήματα παρεμπόδισης του υβριδισμού αυτής της μορφής.
• Η αποφυγή πιθανών ανθρώπινων λαθών, όπως η χρησιμοποίηση ακατάλληλων γεννητόρων ή μη χρησιμοποίηση όλων των δυνατών συνδυασμών διασταύρωσης. Η πετυχημένη συνταγή είναι: αν

21 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
αποτύχεις στην πρώτη προσπάθεια υβριδισμού, δοκίμασε πάλι, αν αποτύχεις στη δεύτερη, ξαναδοκίμασε, κ.ο.κ.
Παραδείγματα. Τα ψάρια υβριδίζονται σχετικά εύκολα, ακόμα και στη φύση. Υπάρχει πληθώρα παραδειγμάτων, ακόμη και στην Ελλάδα, δημιουργίας υβριδίων ακόμα και απομακρυσμένων ταξινομικά ειδών. Το γεγονός αυτό έχει μεγάλη σημασία κυρίως σε περιπτώσεις εισαγωγής ειδών και εμπλουτισμού υδατοσυλλογών όπου υπάρχουν είδη στα οποία ο υβριδισμός θα μπορούσε να έχει δυσμενείς επιπτώσεις. Από τη βιβλιογραφία είναι επίσης γνωστά τα αποτελέσματα αρκετών πειραμάτων υβριδισμού για εμπορική εκμετάλλευση, όπως για παράδειγμα ότι η διασταύρωση σολομού του Ατλαντικού με την κοινή πέστροφα δίνει στείρους απογόνους. Η διασταύρωση κυπρίνου ποικιλίας Ευρωπαϊκής με Ιαπωνική δίνει απογόνους με υψηλή παραγωγικότητα. Στην Ελλάδα έχει αποδειχθεί αναπαραγωγική συμβατότητα, τουλάχιστον κάτω από εργαστηριακές συνθήκες, για τη διασταύρωση γουλιανού με γλανίδι. Τα αλλοενζυμικά ηλεκτροφορητικά πρότυπα των γεννητόρων και των γονικών ατόμων, καθώς και το γεγονός ότι ο μορφότυπος των ατόμων ήταν ενδιάμεσος των γονέων επιβεβαίωσαν την υβριδική τους κατάσταση.
5.2.2 Τύποι προγραμμάτων υβριδισμού Υπάρχουν αρκετά προγράμματα υβριδισμού: α) Το πιο κοινό πρόγραμμα είναι η διασταύρωση ανάμεσα σε δύο καθαρές γραμμές ή στελέχη π.χ. P : A X B F1 : AB (Υβρίδιο) β) Η επαναδιασταύρωση είναι ένας άλλος τρόπος υβριδισμού. Στην περίπτωση αυτή τα υβρίδια της πρώτης γενιάς επαναδιασταυρώνονται με έναν από τους γεννήτορες (ή τις γονικές σειρές). Αυτός ο τρόπος υβριδισμού χρησιμοποιείται προκειμένου να δημιουργηθεί υβρίδιο που να έχει περισσότερο γενετικό υλικό από μια συγκεκριμένη γονική σειρά και να μεταβιβαστούν περισσότερα επιθυμητά αλληλόμορφα από το ένα στέλεχος ή είδος σε κάποιο άλλο, π.χ. P : A X B F1 AB (Υβρίδιο) F1 : AB Χ Α Υβρίδια ΑΒ-Α Με αυτόν τον τρόπο, το γονιδίωμα του υβρίδιου ΑΒ-Α αποτελείται σε ποσοστό περίπου 75% από τον γεννήτορα Α και περίπου 25% από τον γεννήτορα Β. γ) Υβριδισμός κυκλικών ανασυνδυασμών. Στον πολλαπλό υβριδισμό διασταυρώνονται τρία ή περισσότερα καθαρά στελέχη για να δημιουργηθούν απόγονοι που θα έχουν διάφορους συνδυασμούς αλληλομόρφων από όλες τις ομάδες, π.χ.
% Α %Β %Γ Α Χ Β Υβρίδια F1 AB 50 50 0 (Υβρίδια F1 AB) Χ Γ Υβρίδια F2 AB-Γ 25 25 50 (Υβρίδια F2 AB-Γ) Χ Α Υβρίδια F3 AB-Γ-Α 62,5 12,5 25
Ομομιξία είναι η διασταύρωση ατόμων με μεγαλύτερη συγγένεια από ότι ο μέσος όρος του πληθυσμού. Η ομομιξία ως όρος είναι γνωστός σε όλους, δυστυχώς όμως λίγοι κατανοούν τις συνέπειες ενός προγράμματος ομομιξίας. Η ομομιξία, όπως ένα οποιοδήποτε άλλο πρόγραμμα βελτίωσης, μπορεί να χρησιμοποιηθεί με σωστό ή λανθασμένο τρόπο. Η ομομιξία δεν αλλάζει τις γονιδιακές συχνότητες από γενιά σε γενιά, κάτι που κάνει η επιλογή, η γενετική παρέκκλιση ή η μετανάστευση. Η ομομιξία απλά αυξάνει το ποσοστό των ομοζυγωτικών ατόμων, σε βάρος των ετεροζυγωτικών και γι’ αυτό αλλάζει τη γενοτυπική συχνότητα από γενιά σε γενιά. Επειδή μάλιστα αυξάνει τη γενοτυπική διακύμανση του πληθυσμού, αυξάνει και τη φαινοτυπική διακύμανσή του. Γιατί όμως γίνεται τόσος λόγος για τις επιπτώσεις της ομομιξίας;

22 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
Τα συγγενικά άτομα είναι γενετικά περισσότερο όμοια από ότι τα ξένα. Έχουν κοινά διάφορα υποτελή αλληλόμορφα σε ετερόζυγη κατάσταση. Τα περισσότερα όμως από αυτά είναι επιβλαβή και όταν έλθουν σε ομοζυγωτία εκφράζονται και παράγουν αυξημένο αριθμό ανώμαλων φαινοτύπων. Αν μάλιστα το είδος πολλαπλασιάζεται ετερομικτικά, η ομομιξία οδηγεί σε απώλεια της ευρωστίας (ομομικτική κατάπτωση) του πληθυσμού. Οι μελέτες ομομιξίας που έχουν γίνει στην ιριδίζουσα πέστροφα έδειξαν ότι οι απόγονοι των νέων ομομικτικών γενιών χαρακτηρίζονταν από μειωμένη ταχύτητα αύξησης, μειωμένη παραγωγή αυγών και αυξημένο ποσοστό απογόνων με ανωμαλίες. Επιπλέον, όταν ομομικτικές σειρές του σολομού του Ατλαντικού χρησιμοποιήθηκαν για εμπλουτισμούς, έδειξαν μικρή πιθανότητα επιβίωσης στο φυσικό περιβάλλον. Στα πειράματα αυτά, το ποσοστό ομομιξίας κυμαινόταν από 25-60%. Άλλες όμως έρευνες έδειξαν ότι, όταν το ποσοστό ομομιξίας δεν υπερβαίνει το 18% (πειράματα έγιναν στην ιριδίζουσα πέστροφα), όχι μόνον δεν παρατηρείται μείωση της προσαρμογής, αλλά αντίθετα διαπιστώνεται μια μικρή, αλλά σημαντική βελτίωση της παραγωγικότητας. Ύστερα από τα προηγούμενα προκύπτει το εύλογο ερώτημα. Αφού η ομομιξία μειώνει την παραγωγικότητα, πώς είναι δυνατόν να χρησιμοποιηθεί για να βελτιωθεί η απόδοση κάποιου είδους ή πληθυσμού; Συνήθως η ομομιξία χρησιμοποιείται στο πρόγραμμα επιλογής που ονομάζεται γραμμική βελτίωση. Στο πρόγραμμα αυτό, και εφόσον υπάρχει ένα άτομο με βελτιωμένα χαρακτηριστικά, αυτό διασταυρώνεται συνεχώς, με κάποιον απόγονο για να αυξηθεί η συνεισφορά του στο γενετικό απόθεμα του πληθυσμού. Παρακάτω δίνονται δύο χαρακτηριστικά παραδείγματα γραμμικής βελτίωσης.
Ήπια ομομιξία Εντατική ομομιξία Α Χ Β Α Χ Β Γ Χ Δ Α Χ Γ Ε Χ Ζ Α Χ Δ Η Χ Θ Α Χ Ε
Σε άλλα προγράμματα βελτίωσης, η ομομιξία χρησιμοποιείται για την παραγωγή καθαρών ομομικτικών σειρών των οποίων όλα τα άτομα θα έχουν τους ίδιους συνδυασμούς αλληλομόρφων. Έπειτα διασταυρώνονται αυτές οι σειρές για να προκύψουν τα βελτιωμένα υβρίδια της πρώτης γενιάς. Ο υβριδισμός μάλιστα δύο ή περισσότερων σειρών είναι ο κλασικός τρόπος παραγωγής ομοιόμορφων απογόνων για καταναλωτικούς σκοπούς.
5.3 Μέγεθος πληθυσμού – γενετική παρέκκλιση Επειδή οι πληθυσμοί στις υδατοκαλλιέργειες είναι μικροί και κλειστοί, είναι επόμενο ότι σε κάθε γενιά συμβαίνει ακούσια, σε κάποιο βαθμό, τόσο ομομιξία όσο και γενετική παρέκκλιση. Αν μάλιστα συμβαίνουν και τα δύο, τότε εύκολα μπορεί να ελαττωθεί σημαντικά η γενετική ποικιλομορφία, να αυξηθεί η ομομιξία με τελικό αποτέλεσμα τη μείωση της παραγωγικότητας της μονάδας, με ταυτόχρονη αύξηση του κόστους παραγωγής. Θεωρητικά, ο ιδεώδης πληθυσμός θα πρέπει να έχει άπειρο μέγεθος. Δυστυχώς, οι υδατοκαλλιεργητές δουλεύουν με πεπερασμένους πληθυσμούς. Σε τέτοια περίπτωση, ο καλύτερος τρόπος περιγραφής του πληθυσμού δεν είναι ο συνολικός αριθμός των ατόμων του, αλλά ο δραστικός (πραγματικός αριθμός των γεννητόρων που συμμετέχουν στην παραγωγή της επόμενης γενιάς) αριθμός (Νe). Η τιμή της παραμέτρου αυτής επηρεάζεται από αρκετούς παράγοντες όπως ο αριθμός των ατόμων και η αναλογία φύλου των ατόμων που συμμετέχουν στις διασταυρώσεις, το σύστημα διασταύρωσης, κ.ά. Όταν ο πληθυσμός είναι παμμικτικός, ο δραστικός αριθμός υπολογίζεται από τον τύπο:
Νe = 4 Θ Α/ (Θ + Α)

23 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
όπου Θ και Α είναι ο αριθμός των θηλυκών και αρσενικών ατόμων που παράγουν βιώσιμους απογόνους. Από τον τύπο προκύπτει ότι η τιμή του Νe μπορεί να αυξηθεί με δύο τρόπους: με αύξηση του αριθμού των γεννητόρων ή με χρήση γεννητόρων στους οποίους η αναλογία φύλου πλησιάζει τη μονάδα. Επειδή ο Νe σχετίζεται αντιστρόφως ανάλογα με την ομομιξία και τη γενετική παρέκκλιση, ο δραστικός αριθμός θεωρείται από τις σπουδαιότερες παραμέτρους που χαρακτηρίζουν έναν πληθυσμό, καθώς δίνει ενδείξεις για τη γενετική σταθερότητά του. Ο μέσος βαθμός ομομιξίας (F) που δημιουργείται ύστερα από μια γενιά διασταυρώσεων σε έναν κλειστό πληθυσμό δίνεται από τον τύπο:
F = 1/2 Νe Γενετική παρέκκλιση, όπως είναι γνωστό, είναι οι τυχαίες αλλαγές στις συχνότητες των αλληλομόρφων ενός πληθυσμού εξαιτίας μικρών δειγμάτων από τη γενετική δεξαμενή κατά τη μετάβαση από τη μια γενιά στην άλλη. Η γενετική παρέκκλιση εκφράζεται ως η διακύμανση της αλλαγής της γονιδιακής συχνότητας και σχετίζεται αντίστροφα με το Νe.
σ2Δq = pq/2Νe όπου p και q είναι οι γονιδιακές συχνότητες των αλληλομόρφων Α και α ενός γονιδίου. Το άμεσο αποτέλεσμα της γενετικής παρέκκλισης είναι η απώλεια κάποιων και η καθιέρωση άλλων αλληλομόρφων. Οι μικρές τιμές Νe οδηγούν στην αμετάκλητη καταστροφή του γενετικού αποθέματος του πληθυσμού, με την απώλεια αλληλομόρφων και τη δημιουργία ομοζυγωτικών ατόμων. Τελικές επιπτώσεις θα είναι η μείωση της προσαρμοστικότητας, της βιωσιμότητας και της παραγωγικότητας του πληθυσμού, καθώς τελικά ο πληθυσμός χάνει την ευελιξία να προσαρμόζεται σε αλλαγές του περιβάλλοντος, μια και έχουν χαθεί αμετάκλητα μερικές από τις γενετικές του δυνατότητες. Ύστερα από όσα αναφέρθηκαν, είναι εύλογο να αναρωτηθούμε ποιες τιμές Νe θεωρούνται ικανές για να αποφεύγεται η ομομιξία και η γενετική παρέκκλιση. Ο FAO, από το 1981, προτείνει ότι η τιμή του Νe, θα πρέπει να είναι τουλάχιστον 50 για βραχυπρόθεσμες εργασίες και 500 για μακροπρόθεσμες εργασίες. Οι αρμόδιες υπηρεσίες των ΗΠΑ προτείνουν αριθμό ίσο με 1.000. Από την άλλη μεριά, οι ειδικοί επιστήμονες προτείνουν τιμές Νe ανάμεσα στο 263 και το 344 για ιχθυοπληθυσμούς που χρησιμοποιούνται στην παραγωγή τροφής και δολωμάτων και Νe ανάμεσα στο 424 και το 685 για πληθυσμούς που θα χρησιμοποιηθούν για εμπλουτισμούς φυσικών υδάτων.
5.4 Άλλοι παράγοντες που επηρεάζουν ποσοτικά χαρακτηριστικά
5.4.1 Διακύμανση που οφείλεται στην αλληλεπίδραση του γενοτύπου με το περιβάλλον Η αλληλεπίδραση αυτή συμβαίνει επειδή ορισμένα αλληλόμορφα, υπεύθυνα για την παραγωγή του φαινοτύπου, εκφράζονται διαφορετικά σε διαφορετικό περιβάλλον. Είναι ευνόητο ότι η σπουδαιότητα του Vg-e ως μέσου τροποποίησης της παραγωγικότητας ενός πληθυσμού είναι μεγάλη. Από πρακτική σκοπιά, για να μελετηθεί η διακύμανση Vg-e, το καλύτερο που έχει να κάνει ο ιχθυοτρόφος είναι να διερευνήσει τις άριστες συνθήκες κάτω από τις οποίες ένα ορισμένο στέλεχος θα έχει την καλύτερη επίδοση. Στο Ισραήλ, για παράδειγμα, ελέγχθηκε η επίδοση τριών υβριδίων γριβαδιού κάτω από διαφορετικά διαχειριστικά προγράμματα (Πίνακας 5.7). Από τα πειράματα αυτά φαίνεται ότι η ημερήσια αύξηση των υβριδίων ανάμεσα σε κινέζικο και ευρωπαϊκό γριβάδι είναι καλύτερη από την απόδοση οποιουδήποτε γεννήτορα, όταν η ιχθυοπυκνότητα είναι υψηλή. Αντίθετα, σε χαμηλή ιχθυοπυκνότητα, η καλύτερη απόδοση παρατηρήθηκε σε υβρίδια της διασταύρωσης ευρωπαϊκού με ευρωπαϊκό γριβάδι.
5.4.2 Περιβαλλοντική διακύμανση Οι υδατοκαλλιεργητές δεν έχουν πρόβλημα να κατανοήσουν ότι αν χρησιμοποιήσουν άριστες συνθήκες περιβάλλοντος, π.χ. θερμοκρασίας ή ποιότητας νερού, τότε αναμένεται βελτίωση της παραγωγικότητας. Μερικοί από τους παράγοντες που συμβάλλουν στη φαινοτυπική διακύμανση είναι η πυκνότητα της
24 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
καλλιέργειας, το μέγεθος των κόκκων της τροφής, το μέγεθος των αυγών, η ηλικία και το μέγεθος των γεννητόρων. Οι παράγοντες αυτοί αναλύονται σε άλλα Κεφάλαια του μαθήματος.
Πίνακας 5.7 Μέση ημερήσια αύξηση (g/ψάρι/ημέρα) τριών υβρίδιων γριβαδιού κάτω από διαφορετικά προγράμματα διαχείρισης. Σύμφωνα με Wohlfarth et al. (1983).
Είδος Διασταύρωσης Υψηλή πυκνότητα Χαμηλή πυκνότητα Κοπριά Κοπριά &
Σάκχαρο Κοπριά & πρωτεΐνη
Κοπριά & πρωτεΐνη
Ευρωπαϊκό x Ευρωπαϊκό 1,69 3,48 5,60 8,43 Ευρωπαϊκό x Κινέζικο 2,30 3,85 5,62 7,70 Κινέζικο x Κινέζικο 2,15 2,54 3,75 4,98
5.5 Προγράμματα γενετικής επιλογής στις ιχθυοκαλλιέργειες Σε μια πρόσφατη έκθεση του FAO με θέμα την αειφόρο υδατοκαλλιέργεια τονίστηκε η σημασία των προγραμμάτων αναπαραγωγής. Δύο θέματα σχετικά με τη γενετική είναι τα πιο σημαντικά: 1) Η γενετική διαχείριση των υπαρχόντων εξημερωμένων καλλιεργούμενων πληθυσμών και 2) η ανάπτυξη βελτιωμένων γεννητόρων. Τα προγράμματα αναπαραγωγής στις ιχθυοκαλλιέργειες είναι πολύ λιγότερο ανεπτυγμένα από αντίστοιχα για χερσαία ζώα. Σε πολλές περιπτώσεις, η παραγωγή βασίζεται ακόμη στη συλλογή άγριων γεννητόρων για τη βελτίωση της απόδοσης. Η εξημέρωση και η γενετική βελτίωση στις υδατοκαλλιέργειες, όμως, έχει πλεονεκτήματα σε σχέση με άλλα καλλιεργούμενα ζώα: Λόγω της υψηλότερης γενετικής ποικιλότητας στα ψάρια, η απόκριση στην επιλογή μπορεί να δώσει καλύτερα αποτελέσματα. Επίσης, λόγω της υψηλής γονιμότητας είναι δυνατή υψηλότερη ένταση επιλογής. Το πρόβλημα στις ιχθυοκαλλιέργειες είναι ότι δεν γίνονται ζευγαρώματα ανά δύο άτομα, οπότε δεν είναι δυνατό να διατηρούνται γενεαλογικά δένδρα. Η χρήση των γενετικών δεικτών επιτρέπει πια την παρακολούθηση των ατόμων και τη συσχέτιση με τα χαρακτηριστικά τους. Μια προηγμένη μέθοδος επιλογής ατόμων για την αναπαραγωγή βασίζεται στη χρήση γενετικών δεικτών (marker-assisted selection - MAS) ή και γονιδίων (gene-assisted selection - GAS). Χρησιμοποιούνται δηλαδή γενετικοί δείκτες ή γονίδια, συγκεκριμένα αλληλόμορφα των οποίων είναι στενά συνδεδεμένα (tight genetic linkage) με ποσοτικούς γενετικούς τόπους (quantitative trait locus - QTL) που αφορούν ένα συγκεκριμένο ποσοτικό χαρακτηριστικό. Η ύπαρξη αυτών των αλληλομόρφων σχετίζεται με την απόδοση των ατόμων. «Επωφελή» αλληλόμορφα είναι επιθυμητά, ενώ «επιβλαβή» (που μειώνουν την απόδοση του χαρακτηριστικού) πρέπει να εξαλειφθούν από τον πληθυσμό. Θα πρέπει να τονιστεί ότι στη μέθοδο MAS (που είναι και η πιο συχνή) δεν γνωρίζουμε αν ο συγκεκριμένος δείκτης που γενοτυπούμε αφορά αυτόν καθαυτόν τον τόπο QTL που επηρεάζει το χαρακτηριστικό, αλλά κάτι τέτοιο δεν μας ενδιαφέρει κιόλας. Αυτό που έχει σημασία είναι ο δείκτης αυτός να είναι στενά συνδεδεμένος με το γονίδιο ώστε να συγκληρονομείται με το χαρακτηριστικό. Αν, για παράδειγμα, το «επωφελές» αλληλόμορφο Q του QTL είναι πιο συχνά συνδεδεμένο με το αλληλόμορφο δείκτη Μ και το «επιβλαβές» αλληλόμορφο q του QTL πιο συχνά συνδεδεμένο με το αλληλόμορφο δείκτη m, στα προγράμματα επιλογής θα φροντίσουμε να επιλέγουμε γεννήτορες με αλληλόμορφα M και όχι m, ελπίζοντας ότι ταυτόχρονα επιλέγουμε το Q και όχι το q. Βεβαίως σημασία έχει και η κληρονομησιμότητα (h) του κάθε χαρακτηριστικού. Αν και, όπως είναι αναμενόμενο, σε υψηλές τιμές h έχουμε καλύτερα αποτελέσματα, οι τεχνικές MAS εφαρμόζονται κυρίως σε χαρακτηριστικά με χαμηλότερες τιμές h, αφού εκεί δεν είναι δυνατό να έχουμε καλά αποτελέσματα με απλές κλασικές μεθόδους επιλογής. Στην τεχνητή επιλογή που γίνεται κατά τη μέθοδο MAS (ή την GAS) αξιολογούνται (σκοράρονται) οι γεννήτορες με βάση τους γενοτύπους τους στο συγκεκριμένο QTL. Σκοπός είναι να εκτιμηθεί η

25 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
αναπαραγωγική αξία των γεννητόρων συνδυάζοντας πληροφορίες και από τον φαινότυπο και τον γενότυπο. Η MAS χρησιμοποιείται κυρίως για πολύπλοκα χαρακτηριστικά, στα οποία η επιλογή είναι δύσκολη είτε επειδή είναι χρονοβόρα ή δαπανηρή (π.χ. ανθεκτικότητα σε ασθένειες, συγκέντρωση συγκεκριμένων λιπαρών οξέων) ή επειδή απαιτείται θανάτωση των ατόμων (π.χ. ποιότητα κρέατος). Στην περίπτωση που ο συγκεκριμένος δείκτης (ή γονίδιο) συμβάλλει σημαντικά στην εμφάνιση του φαινοτύπου (χαρακτηριστικού υπό επιλογή) οι τεχνικές MAS και GAS μπορεί να δώσουν πολύ καλά αποτελέσματα. Πλήθος QTLs έχουν πια ταυτοποιηθεί σε ιχθυοκαλλιέργειες. Για παράδειγμα, πρόσφατα ταυτοποιήθηκε στον σολομό του Ατλαντικού, σε μια περιοχή 4cΜ, ένα QTL που προσφέρει ανθεκτικότητα στην ασθένεια της νέκρωσης του παγκρέατος (infectious pancreatic necrosis – IPN disease), μια από τις σημαντικότερες μολυσματικές ασθένειες για τις ιχθυοκαλλιέργειες. Παρ’ όλα αυτά, οι ερευνητές παραδέχονται ότι, αν και το συγκεκριμένο QTL εξηγεί το 83% της γενετικής διακύμανσης, χρειάζεται ακόμη καλύτερη χαρτογράφηση του, για ακόμη καλύτερα αποτελέσματα σε MAS προγράμματα. Αντίστοιχα κύρια QTLs που σχετίζονται με ανθεκτικότητα σε ασθένειες έχουν βρεθεί και σε πολλά άλλα καλλιεργούμενα είδη. Οι περισσότερες μελέτες ανεύρεσης QTLs έχουν γίνει στα ψάρια της οικογένειας Salmonidae αλλά αυξάνονται με τον καιρό και σε άλλους ιχθείς ή οστρακοειδή. Με τη δυνατότητα ταυτόχρονης γενοτύπησης χιλιάδων δεικτών (κυρίως SNPs) χάρις στις σύγχρονες τεχνολογίες DNA, έχουμε πια περάσει σε προγράμματα γενετικής βελτίωσης που βασίζονται σε γονιδωματική επιλογή (genomic selection). Οι μέθοδοι αυτές αναπτύσσονται ραγδαία στην εποχή μας και απαιτούν τη συσχέτιση χιλιάδων φαινοτυπικών με γενοτυπικά δεδομένα, ώστε να υπολογιστεί η αναπαραγωγική αξία κάθε γεννήτορα σε γονιδιωματικό επίπεδο. Απαιτούν ακόμη επανειλημμένη γενοτύπηση σε πολλές γενιές για να επιβεβαιωθούν τα αποτελέσματα της αλληλομορφικής ποικιλότητας σε χιλιάδες τόπους, σε πολλούς φαινότυπους. Σε αυτή την περίπτωση, αναμένεται ότι οι χιλιάδες SNPs δείκτες είναι στενά συνδεδεμένοι με τα αντίστοιχα QTLs. Στο πλαίσιο του προγράμματος FISHBOOST (http://www.fishboost.eu/) μελετήθηκαν τα προγράμματα επιλογής σε έξι από τα σημαντικότερα ψάρια για τις ευρωπαϊκές Ιχθυοκαλλιέργειες (σολομό Ατλαντικού, ιριδίζουσα πέστροφα, τσιπούρα, λαβράκι, καλκάνι και γριβάδι) που αντιστοιχούν στο 95% της παραγωγής. Χρησιμοποιήθηκαν ερωτηματολόγια σε 36 εταιρείες παραγωγής γόνου. Τα χαρακτηριστικά που επιλέγονται στα προγράμματα επιλογής είναι σχετικά με την ανάπτυξη, τη μορφολογία, την αναπαραγωγή, την αντίσταση σε ασθένειες, την απόδοση κατά τη θρέψη και την επεξεργασία των ψαριών (π.χ. φιλετοποίηση). Συμπεραίνεται ότι πάνω από το 80% της παραγωγής βασίζεται πια σε προγράμματα επιλογής σε αυτά τα ψάρια, με σημαντικά γενετικά κέρδη στην παραγωγή που, όσον αφορά το βάρος των ψαριών, φτάνει στο σολομό και την πέστροφα το 200%. Προγράμματα επιλογής εφαρμόζονται βέβαια και σε άλλα ψάρια (π.χ. το GIFT- Genetically improved farmed tilapia).
6. Καθορισμός του φύλου. Μέθοδοι ελέγχου της αναλογίας φύλου και εφαρμογές. Εφαρμογές του χειρισμού των χρωμοσωμάτων
6.1 Καθορισμός του φύλου Τα ψάρια παρουσιάζουν ομολογουμένως μια απίστευτη ποικιλία μηχανισμών καθορισμού του φύλου, οι οποίοι είναι δυνατόν να διαφέρουν όχι μόνο μεταξύ ειδών του ίδιου γένους αλλά και μεταξύ πληθυσμών του ίδιου είδους, όπως συμβαίνει σε ψάρια γλυκών νερών της νεοτροπικής ζώνης στην κεντρική και νότια Αμερική. Ο μηχανισμός με τον οποίο καθορίζεται το φύλο στα ψάρια είναι γνωστός για αρκετά είδη (Πίνακας 5.8). Σήμερα είναι γνωστοί τουλάχιστον εννιά διαφορετικοί μηχανισμοί καθορισμού του φύλου. Στους 8 από αυτούς τους μηχανισμούς, το φύλο καθορίζεται από τα φυλετικά χρωμοσώματα. Ο πιο κοινός μηχανισμός καθορισμού του φύλου είναι το σύστημα ΧΥ. Τα θηλυκά ψάρια είναι ομογαμετικά (ΧΧ), ενώ τα αρσενικά ετερογαμετικά (ΧΥ). Αυτός ο μηχανισμός καθορισμού του φύλου είναι παρόμοιος με τον

26 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
μηχανισμό καθορισμού του φύλου στον άνθρωπο. Παρατηρείται στην ιριδίζουσα πέστροφα, την Tilapia nilotica και την T. mossambica. Σε άλλα είδη ψαριών, όπως τα Tilapia aurea και T. hornorum, το αρσενικό άτομο είναι ομογαμετικό (ΖΖ), ενώ το θηλυκό είναι ετερογαμετικό (WΖ). Στους δύο μηχανισμούς καθορισμού του φύλου χρησιμοποιούνται διαφορετικοί συμβολισμοί για να μειωθεί η πιθανότητα σφαλμάτων.
Πίνακας 5.8 Παραδείγματα μηχανισμών καθορισμού του φύλου στα ψάρια.
Είδος Μηχανισμός
Ιριδίζουσα Πέστροφα, Tilapia nilotica, T. mossambica ΧΥ Tilapia aurea, T. hornorum WZ Megupsilon aporus, Gobionellus shuleldti X1X1X2X2/X1X2Y Apareiodon affinis ZZ/ZW1W2 Hoplias sp. XY1Y2/XX Πλατύψαρα, γλώσσες WXY Sternophyx diaphana XO Colisa lalius ZO Limia caudofasciata Αυτοσωμικά γονίδια
Στους επόμενους τρεις μηχανισμούς που φαίνονται στον Πίνακα 5.8, ο καθορισμός του φύλου γίνεται με βάση πολλαπλά φυλετικά χρωμοσώματα. Στο τρίτο σύστημα υπάρχουν πολλαπλά χρωμοσώματα Χ. Στο σύστημα αυτό τα θηλυκά άτομα έχουν τη χρωμοσωμική σύσταση Χ1Χ1Χ2Χ2, ενώ τα αρσενικά έχουν τη σύσταση Χ1Χ2Υ, π.χ. τα είδη Gobionellus shuleldti. Στον τέταρτο μηχανισμό καθορισμού του φύλου υπάρχουν πολλαπλά χρωμοσώματα W. Στον μηχανισμό αυτό, τα αρσενικά άτομα έχουν τη χρωμοσωμική σύσταση ΖΖ, ενώ τα θηλυκά τη σύσταση ZW1W2. Στο πέμπτο σύστημα, υπάρχουν πολλαπλά χρωμοσώματα Υ. Το αρσενικό άτομο έχει τη χρωμοσωμική σύσταση ΧΥ1Υ2, ενώ το θηλυκό έχει τη σύσταση ΧΧ. Ο χρωμοσωμικός αριθμός δεν είναι σταθερός στα είδη ψαριών που έχουν οποιοδήποτε από τους προηγούμενους τρεις μηχανισμούς καθορισμού του φύλου. Στους δύο πρώτους μηχανισμούς τα θηλυκά έχουν ένα επιπλέον χρωμόσωμα, ενώ στον τελευταίο τα αρσενικά είναι εκείνα που έχουν ένα επιπλέον χρωμόσωμα. Ο έκτος μηχανισμός καθορισμού που παρατηρείται στις γλώσσες, είναι του τύπου WXY, Σε αυτόν, όταν υπάρχει το χρωμόσωμα Χ παράγονται αρσενικά, εκτός αν ζευγαρώσει με το χρωμόσωμα W. Το χρωμόσωμα W είναι τροποποιημένο χρωμόσωμα Χ που αναστέλλει την αρρενοκαθοριστική λειτουργία του χρωμοσώματος Υ. Έτσι, τα άτομα ΧΥ και ΥΥ είναι αρσενικά, ενώ τα ΧΧ, WX και WY είναι θηλυκά. Στον έβδομο και τον όγδοο μηχανισμό υπάρχει μόνο ένα φυλετικό χρωμόσωμα. Τα συστήματα αυτά είναι: τα ΧΟ και ΖΟ. Στον έβδομο μηχανισμό, που παρατηρείται στο είδος Sternophyx diaphana, τα θηλυκά είναι της χρωμοσωμικής σύστασης ΧΧ, ενώ τα αρσενικά του τύπου ΧΟ. Στον όγδοο, που παρατηρείται στο είδος Colisa lalius, τα αρσενικά είναι του τύπου ΖΖ, ενώ τα θηλυκά του τύπου ΖΟ. Ο ένατος και τελευταίος μηχανισμός καθορισμού του φύλου, παρατηρείται σε είδη που δεν έχουν φυλετικά χρωμοσώματα. Στα είδη αυτά, το φύλο καθορίζεται από αρρενο- ή θηλεο-καθοριστικά γονίδια που εντοπίζονται στα αυτοσώματα. Αν και το φύλο στα προηγούμενα είδη καθορίζεται από φυλετικά χρωμοσώματα, το φύλο ενός ατόμου μπορεί επίσης να επηρεάζεται ή να καθορίζεται και από αυτοσωμικά φυλοτροποποιητικά γονίδια. Έτσι, πολλές φορές, η προσπάθεια να δημιουργηθούν μόνο αρσενικοί ή μόνο θηλυκοί απόγονοι για να αποφευχθεί η αναπαραγωγή κατά την ανάπτυξη, δεν συνοδεύεται από απόλυτη επιτυχία λόγω της ύπαρξης αυτών των φυλοκαθοριστικών γονιδίων. Η ανάπτυξη των σύγχρονων μεθόδων γονιδιωματικής έχει βοηθήσει και εδώ, ώστε να ταυτοποιηθούν είτε αυτά καθαυτά τα γονίδια που καθορίζουν το φύλο ή τουλάχιστον μοριακοί δείκτες που είναι στενά

27 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
συνδεδεμένοι με αυτά τα γονίδια σε πλήθος ιχθύων με εμπορικό ενδιαφέρον, όπως το καλκάνι (Psetta maxima), ο σολομός Oncorhynchus tshawytscha και άλλα είδη της οικογένειας Salmonidae, το στουριόνι (Acipenser fulvescens), το Hippoglossus stenolepsis και άλλα. Το γεγονός αυτό συμβάλλει τα μέγιστα στη μείωση του χρόνου για ταυτοποίηση του φύλου σε προγράμματα επιλογής.
6.2 Μέθοδοι ελέγχου της αναλογίας φύλου και εφαρμογές
6.2.1 Γιατί γίνεται έλεγχος της αναλογίας φύλου Αν και το φύλο στα ψάρια καθορίζεται κυρίως γενετικά, περιβαλλοντικοί παράγοντες, όπως η θερμοκρασία, αλλά και η φωτοπερίοδος, η αλατότητα και η πυκνότητα της καλλιέργειας, μπορούν να συμβάλλουν επίσης στον καθορισμό του φύλου ή να επηρεάσουν κυρίως την αναλογία αρσενικών θηλυκών ατόμων (π.χ. στο λαβράκι). Η δυνατότητα να καθοριστεί το φύλο τροποποιώντας περιβαλλοντικούς παράγοντες έχει σημαντικά οφέλη για τις ιχθυοκαλλιέργειες. Ο πρακτικός έλεγχος της αναλογίας φύλου στις ιχθυοκαλλιέργειες άρχισε το 1977. Αποτελεί, ίσως, την πιο πετυχημένη γενετική παρέμβαση στις σύγχρονες ιχθυοκαλλιεργητικές μονάδες. Η ανάγκη για τον έλεγχο της αναλογίας φύλου ανέκυψε από το γεγονός ότι τα δύο φύλα στα ψάρια παρουσιάζουν διαφορές σε κάποια χαρακτηριστικά, τα οποία έχουν σημαντικό οικονομικό ενδιαφέρον, όπως είναι ο ρυθμός αύξησης. Στα θαλάσσια πλατύψαρα, για παράδειγμα, το θηλυκό αυξάνει γρηγορότερα με αποτέλεσμα το αρσενικό να είναι σημαντικά μικρότερο από το θηλυκό. Στην πέστροφα, η αύξηση των αρσενικών σταματά όταν εμφανιστούν τα δευτερογενή φυλετικά χαρακτηριστικά και το άτομο γίνεται επιθετικό με άσχημη οσμή της σάρκας. Αντίθετα, τα αρσενικά άτομα της τιλάπια είναι επιθυμητά για εκτροφή, διότι έχουν υψηλότερο ρυθμό αύξησης από τα θηλυκά. Στο λαβράκι, οι συνθήκες εκτροφής οδηγούν σε πληθυσμούς με πολύ υψηλό αριθμό αρσενικών. Τα αρσενικά άτομα του λαβρακιού είναι, όμως, μικρότερα από τα θηλυκά, άρα υπάρχουν οικονομικές απώλειες. Η χρήση ορμονών για να παραχθούν μονοφυλετικοί πληθυσμοί για καλλιέργεια και παραγωγή εξακολουθεί να αποτελεί ενεργό ερευνητικό πεδίο. Ένα ακόμα πιο πρόσφατο πεδίο είναι και η τροποποίηση του φύλου με τεχνικές RNAi. Δηλαδή, γίνεται χρήση δίκλωνων μορίων RNA που υβριδίζονται με γονίδια στο πυρηνικό DNA, με αποτέλεσμα να σταματάει η μεταγραφή τους. Τέτοιες τεχνικές χρησιμοποιούνται κυρίως σε καρκινοειδή.
6.2.2 Υβριδισμός και καθορισμός του φύλου Ο έλεγχος της αναλογίας φύλου με τη δημιουργία των κατάλληλων υβριδίων αναπτύχθηκε κυρίως στην τιλάπια. Η παραγωγή μονοφυλετικών πληθυσμών προέρχεται συνήθως από τη διασταύρωση δύο ειδών τιλάπιας που έχουν διαφορετικό μηχανισμό καθορισμού του φύλου. Από την παρακάτω διασταύρωση παράγονται μόνο αρσενικοί απόγονοι επειδή υπερισχύει το χρωμόσωμα Ζ:
P1 Θηλυκό Tilapia nilotica (ΧΧ) x Tilapia hornorum (ZZ) Αρσενικό Γαμέτες Χ Ζ Απόγονοι ΧΖ (Αρσενικοί)
Εναλλακτικά όμως, στην αντίστροφη διασταύρωση δεν παίρνουμε μονοφυλετικό πληθυσμό:
P1 Θηλυκό Tilapia hornorum (WZ) x Tilapia nilotica (XY) Αρσενικό Γαμέτες W, Z X, Y Απόγονοι WX WY, ZX, ZY
Θηλυκοί Αρσενικοί Η παραπάνω μεθοδολογία δεν δίνει 100% αρσενικά ή θηλυκά άτομα εξαιτίας αυτοσωμικών γονιδίων που επηρεάζουν ή τροποποιούν το φύλο. Έτσι, ανάμεσα στα αρσενικά υβρίδια πάντοτε βρίσκονται μερικά θηλυκά άτομα, τα οποία αρχίζουν έναν ανεπιθύμητο κύκλο αναπαραγωγής και υπερπληθυσμού.

28 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
6.2.3 Μεταστροφή του φύλου με επίδραση στεροειδών ορμονών Άλλος τρόπος για τον έλεγχο της αναλογίας του φύλου είναι η χορήγηση στεροειδών ορμονών κατά τα αρχικά στάδια σεξουαλικής ανάπτυξης και διατροφής των ψαριών. Η μεθοδολογία αυτή, επίσης δεν έχει εφαρμοστεί σε εμπορική βάση, για τους παρακάτω λόγους:
• Τα ψάρια κάθε γενιάς πρέπει να διατρέφονται με κατάλληλα στεροειδή και πάντοτε υπό την επίβλεψη ειδικού επιστήμονα.
• Η διατροφή με ορμόνες έχει ως αποτέλεσμα υψηλότερη θνησιμότητα του γόνου. • Οι καταναλωτές είναι απρόθυμοι να αγοράσουν ψάρια που περιέχουν ορμόνες, λόγω των πιθανών
αρνητικών επιδράσεων αυτών των ουσιών στον άνθρωπο. Τα μειονεκτήματα αυτά οδήγησαν στην καθιέρωση της γενετικής μεθοδολογίας ελέγχου του φύλου, με χρήση στεροειδών ορμονών μόνο στα πρώτα στάδια ανάπτυξης των ψαριών, έτσι ώστε να μην υπάρχουν ορμόνες στα προϊόντα που καταλήγουν στην αγορά.
6.2.4 Επαγωγική μεταστροφή του φύλου Η αναστροφή του φύλου στα ψάρια με χορήγηση ορμονών μπορεί να γίνει με ένεση, με εμφύτευση ή με τη διατροφή. Ο τελευταίος τρόπος είναι ο απλούστερος και ο πιο αποτελεσματικός, αρκεί να πραγματοποιηθεί την κατάλληλη περίοδο ανάπτυξης του γόνου, με την κατάλληλη συγκέντρωση της ορμόνης και χρονική διάρκεια επίδρασης. Στη βιβλιογραφία είναι διαθέσιμα αρκετά ειδικά πρωτόκολλα για την πετυχημένη αναστροφή του φύλου σε πολλά είδη ψαριών. Προκειμένου να επιτευχθεί επαγωγική μεταστροφή του φύλου, γόνος κατάλληλης ηλικίας εκτρέφεται για συγκεκριμένο αριθμό ημερών με μεθυλοτεστοστερόνη. Τα γενοτυπικά θηλυκά (ΧΧ) ψάρια, ύστερα από την ορμονική επίδραση με ανδρογόνα, μεταστρέφονται σε φαινοτυπικά αρσενικά. Το βλαστόπλασμα τέτοιων ψαριών αποτελείται αποκλειστικά από σπέρμα που είναι θηλεοκαθοριστικό, αφού τα σπερματοζωάρια έχουν μόνο ένα χρωμόσωμα Χ. Αυγά που γονιμοποιούνται με τέτοια σπερματοζωάρια δίνουν φυσιολογικούς διπλοειδείς, αλλά μόνο θηλυκούς, απογόνους. Η μεθοδολογία αυτή έχει πλέον καθιερωθεί για την παραγωγή αποκλειστικά θηλυκών ατόμων πέστροφας. Επίσης, σε αρσενικά (ΧΥ) άτομα, με χορήγηση οιστρογόνων γίνεται αντιστροφή του φύλου και παράγουν ωάρια. Και στις δύο περιπτώσεις, η επιλογή των κατάλληλων γενοτύπων και η διασταύρωσή τους οδηγεί στην παραγωγή μονοφυλετικών πληθυσμών.
6.2.4.1 Παραγωγή ολο-θηλυκού γόνου σε ψάρια που το φύλο τους ελέγχεται από τον μηχανισμό φυλοκαθορισμού ΧΧ θηλυκό/ΧΥ αρσενικό Στα αρχικά στάδια ανάπτυξης του γόνου (Εικόνα 5.8) χορηγούνται ανδρογόνα, π.χ. 17α-μεθυλοτεστοστερόνη, με αποτέλεσμα από τα παραγόμενα αρσενικά που θα χρησιμοποιηθούν ως πατρική γενιά, τα μισά περίπου να είναι σεξουαλικά ώριμα ψάρια της χρωμοσωμικής σύστασης ΧΥ (φυσιολογικά αρσενικά) και τα άλλα μισά της σύστασης ΧΧ (αρσενικά από αναστροφή του φύλου). Ακολουθεί διασταύρωση ελέγχου με κανονικά θηλυκά (ΧΧ) και τα μεν φυσιολογικά αρσενικά (ΧΥ) δίνουν απογόνους θηλυκούς (ΧΧ) και αρσενικούς (ΧΥ), και γι’ αυτό απομακρύνονται, τα δε “αρσενικά” ΧΧ δίνουν μόνο θηλυκούς (ΧΧ) απογόνους. Στη δεύτερη, τρίτη κ.λπ. γενιά χορηγούνται ανδρογόνα σε ένα ποσοστό του γόνου, προκειμένου να παράγονται συνέχεια “αρσενικά” της σύστασης ΧΧ. Ειδικά στα σολομονοειδή, η επιλογή των αρσενικών διευκολύνεται από το γεγονός ότι τα άτομα αυτά έχουν ατελές σύστημα απελευθέρωσης του σπέρματος και είναι αναγκαίο να θανατωθούν προκειμένου να συλλεχθεί το σπέρμα.
6.2.4.2 Παραγωγή ολο-αρσενικού γόνου σε ψάρια που το φύλο τους ελέγχεται από τον μηχανισμό φυλοκαθορισμού ΧΧ θηλυκό/ΧΥ αρσενικό Η δημιουργία ολο-αρσενικού γόνου είναι πιο χρονοβόρος διαδικασία σε σχέση με την παραγωγή ολο-θηλυκού γόνου, μια και απαιτείται μία επιπλέον γενιά ώστε να εδραιωθεί το σύστημα. Στην πράξη, είναι

29 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
δυνατόν να δημιουργηθεί ολο-αρσενικός γόνος, τόσο χρωμοσωμικής σύστασης ΧΥ, όσο και σύστασης ΥΥ. Η παραγωγή ολο-αρσενικών ατόμων χρωμοσωμικής σύστασης ΥΥ είναι δυνατόν να επιτευχθεί με ανδρογένεση ή με συνδυασμό αναστροφής του φύλου και γυνογένεσης. Η μεθοδολογία παραγωγής ολο-αρσενικών χρησιμοποιείται από τους ιχθυοπαραγωγούς της άγριας πέστροφας, της τιλάπια και του αμερικάνικου γατόψαρου.
Εικόνα 5.8 Πειραματική διαδικασία παραγωγής ολο-θηλυκών πληθυσμών. Μηχανισμός καθορισμού του φύλου: Θηλυκό (ΧΧ), Αρσενικό (ΧΥ).
6.3 Εφαρμογές του χειρισμού των χρωμοσωμάτων Ο χειρισμός των χρωμοσωμάτων στη γεωργική παραγωγή αποδείχθηκε χρήσιμο εργαλείο για τη δημιουργία πολλών νέων φυτικών ποικιλιών. Είναι αδιαμφισβήτητο γεγονός ότι πολλά καλλιεργούμενα φυτά που τρώμε είναι πολυπλοειδή (π.χ. οχταπλοειδείς φράουλες, εξαπλοειδή ακτινίδια και σκόρδα, τριπλοειδείς μπανάνες και πορτοκάλια, κ.λπ.). Από την άλλη, στα ζώα, ο χειρισμός των χρωμοσωμάτων, αν και στα αμφίβια ξεκίνησε στις αρχές του 20ου αιώνα, δεν είχε τόση μεγάλη επιτυχία. Παρ’ όλα αυτά, προέκυψαν τρεις βασικές τεχνικές που εφαρμόζονται στις ιχθυοκαλλιέργειες: η γυνογένεση, η ανδρογένεση και η δημιουργία πολυπλοειδών.
6.3.1 Γυνογένεση Η γυνογένεση (Εικόνα 5.9) περιλαμβάνει την ενεργοποίηση της ανάπτυξης ωαρίου από σπερματοζωάριο, χωρίς όμως το σπερματοζωάριο να συνεισφέρει γενετικά στο σχηματιζόμενο έμβρυο. Οι απόγονοι της γυνογένεσης προέρχονται αποκλειστικά από το γενετικό υλικό του θηλυκού. Υπάρχουν δύο βασικές μεθοδολογίες για την τεχνητή πρόκληση της γυνογένεσης στα ψάρια: η εφαρμογή υδροστατικής πίεσης ή θερμικών πληγμάτων τα πρώτα λεπτά μετά από την ενεργοποίηση του ωαρίου. Το ωάριο, και στις δύο μεθοδολογίες, αρχικά γονιμοποιείται με σπερματοζωάριο του οποίου καταστράφηκε το γενετικό υλικό με επίδραση υπεριώδους ακτινοβολίας ή ακτίνων γ. Τέτοια σπερματοζωάρια έχουν τη δυνατότητα να διαπερνούν τα ωάρια και να προκαλούν έναρξη της εμβρυογένεσης. Με αυτόν τον τρόπο, το γονιμοποιημένο ωάριο αναπτύσσεται σε μη βιώσιμο απλοειδές έμβρυο. Με κατάλληλη όμως επέμβαση σε δεύτερο στάδιο, και στις δύο μεθοδολογίες, επιτυγχάνονται διπλοειδή έμβρυα. Αυτό μπορεί να επιτευχθεί με τρεις τρόπους: με έλεγχο της μείωσης Ι, με σύντηξη του απλοειδούς ωαρίου με το δεύτερο πολικό σωμάτιο ή με παρεμπόδιση της τελόφασης στην πρώτη αυλάκωση του ωαρίου. Με αυτόν τον τρόπο, λαμβάνεται αρχικά διπλοειδές έμβρυο και τελικά βιώσιμο ώριμο άτομο. Τέτοια άτομα είναι θηλυκά (όταν ο μηχανισμός καθορισμού του φύλου είναι ΧΧ/ΧΥ) και ομόζυγα σχεδόν για όλα τα γονίδια του γονιδιώματός τους. Αν και θεωρητικά η δεύτερη μεθοδολογία αναμένεται να δώσει υψηλότερα ποσοστά ομοζυγωτίας

30 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
στα ψάρια που προκύπτουν με γυνογένεση (με αποτέλεσμα την ομομικτική κατάπτωση και το θάνατο πολλών ατόμων), στην πράξη, για διάφορους λόγους, η πρώτη δίνει 8 φορές υψηλότερα ποσοστά γόνιμων, βιώσιμων ομοζυγωτικών ατόμων. Τα άτομα που επιβιώνουν μπορούν να χρησιμοποιηθούν σε προγράμματα υβριδισμού. Γυνογένεση έχει επιτευχθεί στα κυπρινοειδή και σε άλλα είδη, παρ’ όλα αυτά χρησιμοποιείται κυρίως για ερευνητικούς σκοπούς.
Εικόνα 5.9 Πειραματική διαδικασία δημιουργίας γυνογενετικών ατόμων με εφαρμογή σοκ.
6.3.2 Ανδρογένεση Η ανδρογένεση είναι διαδικασία αντίστοιχη της γυνογένεσης. Περιλαμβάνει την παραγωγή απογόνων στους οποίους το γενετικό υλικό προέρχεται αποκλειστικά από το αρσενικό. Αυτό επιτυγχάνεται συνήθως με καταστροφή του γενετικού υλικού του ωαρίου με ακτινοβόληση, πριν από τη γονιμοποίηση από το σπερματοζωάριο. Σε ένα δεύτερο στάδιο, δημιουργούνται διπλοειδή έμβρυα, εξαιτίας κάποιας παρεμβολής στην πρώτη κυτταρική διαίρεση.
6.3.3 Δημιουργία πολυπλοειδών ατόμων Πολυπλοειδή ψάρια, συνήθως τριπλοειδή, δημιουργούνται με διάφορους τρόπους, για παράδειγμα εφαρμόζοντας υδροστατική πίεση ή θερμικά πλήγματα ή χημικά σε γονιμοποιημένα αυγά. Τα θερμικά πλήγματα έχουν το πλεονέκτημα ότι μπορούν να εφαρμοστούν σχετικά εύκολα σε μεγάλες ομάδες αυγών. Τριπλοειδή ψάρια μπορούν επίσης να δημιουργηθούν από τη διασταύρωση διπλοειδών με τετραπλοειδή άτομα. Τα κύρια χαρακτηριστικά αυτών των τριπλοειδών ατόμων είναι ο υψηλός ρυθμός αύξησής τους (λόγω αυξημένης γονιδιακής δόσης και αυξημένης ετεροζυγωτίας) και η στειρότητά τους (λόγω των προβλημάτων σωστού διαχωρισμού των χρωμοσωμάτων κατά τη μείωση). Οι ιδιότητες αυτές καθιστούν τα τριπλοειδή ψάρια κατάλληλα για εμπλουτισμούς, χωρίς να υπάρχουν φόβοι ότι θα συμβάλλουν στην αλλαγή του γονιδιακού αποθέματος των φυσικών πληθυσμών. Όσον αφορά την καλύτερη και γρηγορότερη αύξηση των τριπλοειδών από τα διπλοειδή, αυτό είναι κυρίως δεδομένο στα μαλάκια. Στα ψάρια αν και είναι συχνό το φαινόμενο αυτό, υπάρχουν και είδη όπου τα τριπλοειδή άτομα δεν αναπτύσσονται καλά. Τριπλοειδή άτομα έχουν παραχθεί, έστω και πειραματικά ,σχεδόν σε όλα τα είδη με μεγάλο καλλιεργητικό ενδιαφέρον (όπως γριβάδι, γατόψαρα, τιλάπια, σολομός αλλά και στρείδια, χτένια γυαλιστερές). Εμπορικά στελέχη τριπλοειδών συναντώνται κυρίως σε πέστροφες, γριβάδια και το στρείδι του Ειρηνικού (Beaumont et al. 2010).

31 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
6.4 Κρυοσυντήρηση γεννητικών κυττάρων και κλωνοποίηση στα ψάρια Τα σπερματοζωάρια εύκολα συλλέγονται από αρσενικά ψάρια και χρησιμοποιούνται για υποβοηθούμενη γονιμοποίηση ωαρίων. Τα σπερματοζωάρια μπορεί να χρησιμοποιηθούν άμεσα ή ύστερα από κρυοσυντήρηση σε απεριόριστο χρόνο. Η κρυοσυντήρηση αποτελεί χρήσιμο εργαλείο για την ενδεχόμενη απώλεια ενός υψηλής απόδοσης στελέχους και για να περιοριστεί ο χώρος αποθήκευσης των διάφορων σειρών. Εξαιτίας της υπεραλίευσης και της πιθανής κατάρρευσης των ιχθυοαποθεμάτων αρκετών ειδών ψαριών, όπως του μπακαλιάρου του Ατλαντικού Gadus morhua μουρουνόγαδου, ίσως είναι καιρός να σκεφτούμε τη δημιουργία ενός Κέντρου Κρυοσυντήρησης του γενετικού υλικού απειλούμενων ειδών, ως ένα διαχειριστικό μέτρο για να μπορέσει να διατηρηθεί η γενετική ποικιλότητα στον Ελληνικό χώρο. Η κλωνοποίηση ψαριών (κυρίως Κυπρινοειδών) ξεκίνησε στην Κίνα το 1961. Αργότερα, συνεχίστηκε με επιτυχία στα πειραματόζωα του εργαστηρίου, όπως το ριζόψαρο και το ψάρι ζέβρα. Σε αυτές τις περιπτώσεις εφαρμόστηκε η μεθοδολογία κλωνοποίησης του προβάτου Ντόλι. Παρ΄όλα αυτά, η κλωνοποίηση δεν έχει πολλές εφαρμογές στα ψάρια, αφού κλώνοι μπορούν να συλλεχθούν και με άλλες πιο απλές προσεγγίσεις (π.χ. γυνογένεση).
7. Χειρισμός των γονιδίων και παραγωγή γενετικά τροποποιημένων οργανισμών
7.1 Παραγωγή διαγενετικών ψαριών Από τη δεκαετία του 1980 έχει σημειωθεί ραγδαία πρόοδος στην εφαρμογή της τεχνολογίας του ανασυνδυασμένου DNA για τη βελτίωση των οργανισμών που χρησιμοποιούνται στις υδατοκαλλιέργειες. Αυτό οφείλεται τόσο στην πετυχημένη καλλιέργεια πολλών ειδών, όσο και στην καλή γνώση της βιολογίας τους. Στα είδη αυτά μπορούν εύκολα να συλλεχθούν μεγάλες ποσότητες αυγών, να γονιμοποιηθούν τα αυγά κάτω από αυστηρά ελεγχόμενες συνθήκες περιβάλλοντος και, το σπουδαιότερο, τα γονιμοποιημένα αυγά δεν χρειάζεται να τοποθετηθούν στο αναπαραγωγικό σύστημα της μητέρας για να αναπτυχθούν (όπως συμβαίνει στα θηλαστικά). Εδώ και αρκετά χρόνια, έχουν κατασκευαστεί βιβλιοθήκες γονιδίων από αρκετά είδη ψαριών. Έτσι, δίνεται πλέον η ευκαιρία να τροποποιηθεί το DNA ψαριών με αλληλουχίες DNA από το ίδιο ή άλλα συγγενικά είδη. Ένας επιπλέον λόγος που κάνει τα ψάρια, τα μαλάκια και άλλα ασπόνδυλα κατάλληλο πειραματικό υλικό για την εφαρμογή της τεχνολογίας του ανασυνδυασμένου DNA είναι ότι φαίνεται να έχουν μικρότερη σπουδαιότητα οι ηθικοί φραγμοί που συνοδεύουν τα πειράματα γενετικής μηχανικής στα θηλαστικά και ειδικότερα στον άνθρωπο.
7.1.1 Τι σημαίνει γενετικά τροποποιημένος οργανισμός; Αφού απομονωθεί και κλωνοποιηθεί συγκεκριμένη αλληλουχία γενετικού υλικού (DNA) από έναν οργανισμό, γίνεται εσκεμμένη μεταφορά και ενσωμάτωση του εξωγενούς DNA στο γενετικό υλικό άλλου ψαριού, η έκφρασή του σε αυτό και, τέλος, η κληρονόμησή του στους απογόνους με αποτέλεσμα να δημιουργούνται οι διαγενετικοί οργανισμοί. Με απλά λόγια, ένας γενετικά τροποποιημένος οργανισμός (ΓΤ, G.M.O. = Genetically modified organism) έχει πέρα από το δικό του DNA, επιπλέον DNA άλλου οργανισμού που ενσωματώθηκε στο δικό του. Το τελικό αποτέλεσμα είναι ότι ένας Γ.Τ. οργανισμός πέρα από τα δικά του χαρακτηριστικά έχει ένα ή περισσότερα νέα χαρακτηριστικά άλλου οργανισμού. Απώτερος σκοπός των πειραμάτων είναι η τροποποίηση του γενετικού υλικού των οργανισμών για να αποκτήσουν βελτιωμένα ή εντελώς νέα χαρακτηριστικά ώστε να χρησιμοποιηθούν στις υδατοκαλλιέργειες, τη βιοτεχνολογία, τη φαρμακευτική, την ιατρική ή τη βιομηχανία.

32 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
7.1.2 Από πού μπορεί να γίνει μεταφορά γενετικού υλικού; Σε ένα είδος ψαριού, μεταφορά γενετικού υλικού μπορεί να γίνει από 5 διαφορετικούς τύπους οργανισμών: α) από σειρές (ή διαφορετικά ιχθυοαποθέματα) του ίδιου είδους ψαριού, β) από άλλο είδος ψαριού ή άλλο ζωικό είδος, γ) από ένα φυτικό είδος, δ) από ένα βακτήριο ή ε) από έναν DNA ή RNA ιό.
7.1.3 Από τι αποτελείται η κατασκευή DNA που θα μεταφερθεί; Το τμήμα του DNA που θα μεταφερθεί αποτελείται συνήθως από τέσσερα γενετικά στοιχεία (Εικόνα 5.10),
• το DNA που θα χρησιμεύσει ως όχημα-φορέας μεταφοράς (π.χ. ιός, πλασμίδιο). • το δομικό γονίδιο (DNA) που καθορίζει το επιθυμητό χαρακτηριστικό, π.χ. το γονίδιο που καθορίζει
την αυξητική ορμόνη. • το κατάλληλο ρυθμιστικό στοιχείο (DNA) που θα ρυθμίζει την έκφραση του νεοεισερχόμενου DΝΑ
σε συγκεκριμένους ιστούς (ιστοειδίκευση) και καθορισμένο στάδιο ανάπτυξης. Διαδραματίζει βασικό ρόλο στην επιτυχία των πειραμάτων. Η απλή ενσωμάτωση ενός δομικού γονιδίου στο γονιδίωμα ενός είδους δεν σημαίνει υποχρεωτικά ότι το γονίδιο θα εκφραστεί ή ότι θα αλλάξουν τα μορφολογικά ή φυσιολογικά χαρακτηριστικά του ατόμου.
• το DNA-δείκτη που θα επιτρέπει την εύκολη αναγνώριση και επιλογή του διαγενετικού οργανισμού. Για το σκοπό αυτό συνήθως χρησιμοποιούνται γονίδια που παρέχουν ανθεκτικότητα σε δύο αντιβιοτικά, το αντιβιοτικό καραμυσίνη και το ένζυμο λακταμάση που διασπά το αντιβιοτικό αμπικιλλίνη.
Η ανακάλυψη των ενδονουκλεασών περιορισμού και των βακτηριακών πλασμιδίων επέτρεψε την κλωνική παραγωγή πολλών γονιδίων, καθώς και τη δημιουργία γονιδιακών βιβλιοθηκών σε πολλά ζωικά είδη. Αν έπειτα, κάτω από κατάλληλες πειραματικές συνθήκες, γίνει εισαγωγή πολλαπλών αντιγράφων ενός γονιδίου σε γονιμοποιημένο ωάριο, υπάρχει αυξημένη πιθανότητα ενσωμάτωσης της νέας αλληλουχίας στο γονιδίωμα του δέκτη-ξενιστή. Η τεχνολογία, με χρονολογική σειρά, εφαρμόστηκε πετυχημένα αρχικά στο ποντίκι, τη Drosophila, τα πρόβατα και τους χοίρους. Από το 1984, εργαστήρια σε αρκετές χώρες (Αγγλία, Γαλλία, Ολλανδία, Νορβηγία, ΗΠΑ και Κίνα) ενδιαφέρθηκαν να εφαρμόσουν την τεχνολογία αυτή σε πολλά είδη ψαριών. Πετυχημένες προσπάθειες παραγωγής διαγενετικών ατόμων αναφέρονται αρχικά για τα ακόλουθα είδη ψαριών: ιριδίζουσα πέστροφα, σολομό Ατλαντικού, τιλάπια, αμερικάνικο γατόψαρο, χρυσόψαρο και άλλα είδη του γλυκού νερού. Η τροποποίηση της γενετικής σύστασης βρίσκεται πια σε αρκετά προχωρημένο στάδιο για 40 τουλάχιστον είδη ψαριών, οπότε έχει αρχίσει η πιλοτική εφαρμογή προγραμμάτων εμπορικής εκμετάλλευσής τους. Το μεγαλύτερο ενδιαφέρον εστιάζεται στην παραγωγή φτηνών τροφών και στη δημιουργία βιοαντιδραστήρων, αν και υπάρχει επίσης ενδιαφέρον για την απελευθέρωση στη φύση στείρου διαγενετικού γόνου για αλιεία από ερασιτέχνες ψαράδες. Λόγω των επιπτώσεων από την τυχαία απελευθέρωση διαγενετικών ψαριών στη φύση, υπάρχει προβληματισμός για τις πιθανές αρνητικές συνέπειες στους φυσικούς πληθυσμούς. Ειδικές νομοθετικές ρυθμίσεις για το θέμα αυτό υπάρχουν από την Ευρωπαϊκή Ένωση, όπως η οδηγία 90/220. Η ICES (International Council for the Exploration of the Sea), πριν από την έγκριση των πειραμάτων ή της εμπορικής εφαρμογής, προτείνει να ελέγχονται διεξοδικά τα πρωτόκολλα πιθανών κινδύνων από τη διαχείριση των διαγενετικών ψαριών.
7.2 Βασικά στάδια παραγωγής διαγενετικών ψαριών Η τεχνολογία παραγωγής διαγενετικών ψαριών περιλαμβάνει αρκετά στάδια: Απόκτηση του γονιδίου. Η απομόνωση της κατάλληλης γονιδιακής ακολουθίας μαζί με τις γειτονικές ρυθμιστικές ακολουθίες, από κάποια βιβλιοθήκη γονιδίων ή κάποιο άλλο εργαστήριο, αποτελεί το πρώτο βήμα για την έναρξη της πειραματικής προσπάθειας. Η ρυθμιστική περιοχή θα πρέπει να συμπεριλαμβάνει τις αλληλουχίες του προαγωγέα από την 5’ πλευρά του δομικού γονιδίου, καθώς επίσης και ειδικές ακολουθίες ενισχυτές έκφρασης σε συγκεκριμένο ιστό. Αν υπάρχει δυνατότητα, είναι προτιμότερο να

33 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
επιλεγεί γονιδιωματική ακολουθία (DNA που περιέχει τόσο εξώνια όσο και ιντρόνια) και όχι συμπληρωματική ακολουθία DNA ή cDNA που περιέχει μόνο τα εξώνια. Ο λόγος είναι ευνόητος. Η γονιδιωματική ακολουθία περιέχει όλα τα εξόνια και ιντρόνια και επιπλέον όλες τις απαραίτητες περιοχές για τη ρύθμιση της έκφρασης του γονιδίου.
Εικόνα 5.10 Διαγραμματική απεικόνιση πλασμιδίου που χρησιμοποιείται για τη δημιουργία και την παραγωγή γονιδίου αυξητικής ορμόνης που έχει προαγωγέα από το γονίδιο της μεταλλοθειονίνης και που χρησιμοποιείται για τη δημιουργία διαγενετικών ατόμων.
Κλωνοποίηση του γονιδίου. Η επιθυμητή γονιδιακή ακολουθία που αποκτήθηκε ή παρασκευάστηκε στο εργαστήριο, κλωνοποιείται σε κατάλληλο πλασμίδιο ή ιικό φορέα. Έπειτα πολλαπλασιάζεται σε κατάλληλο βακτηριακό στέλεχος. Σε επόμενα στάδια απομονώνονται εκατομμύρια αντίγραφα της αλληλουχίας από τα βακτηριακά κύτταρα και εάν είναι απαραίτητο ξεχωρίζεται από τον πλασμιδιακό ή ιικό φορέα, χρησιμοποιώντας κατάλληλα ένζυμα περιορισμού. Εισαγωγή του γονιδίου στο ωάριο. Η μεταφορά και η ενσωμάτωση του DΝΑ στα ψάρια γίνεται με:
• Απομόνωση και κλωνοποίηση του γονιδίου και δημιουργία της κατάλληλης αλληλουχίας DNA για άμεση μηχανική μικροέκχυσή της στον προπυρήνα ενός γονιμοποιημένου ωαρίου ή σπερματοζωαρίου ή την εισδοχή της με τη βοήθεια ηλεκτροδιήθησης. Η ηλεκτροδιήθηση επιτρέπει την επεξεργασία μεγάλου αριθμού αυγών, δίνει υψηλότερα ποσοστά διαγενετικών ψαριών και με μεγαλύτερη πιθανότητα επιβίωσης.
• Εισαγωγή της επιθυμητής ακολουθίας DΝΑ σε καλλιέργεια εμβρυακών κυττάρων, και • Μεταφορά του γονιδίου (DNA) χρησιμοποιώντας ως όχημα ιούς ή πλασμίδια.
Γενικά, τα ποσοστά επιτυχίας παραγωγής διαγενετικών ψαριών είναι χαμηλά, της τάξης του 2-5%, αν και παρατηρήθηκαν ποσοστά της τάξης του 50% σε μερικά πειράματα και είδη ψαριών. Ιδεωδώς, η αλληλουχία θα πρέπει να εισάγεται, για καλύτερα αποτελέσματα, με μικροχειριστή στον πυρήνα του ωαρίου. Στα ψάρια, συνήθως, η αλληλουχία εισάγεται στο κυτταρόπλασμα, κοντά στον πυρηνικό φάκελο. Σε πολύ μεγάλα λεκιθικά αυγά, είναι άσκοπο να εισαχθεί η αλληλουχία μέσα στη λέκιθο. Στην ιριδίζουσα πέστροφα

34 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
έχουμε καλύτερα αποτελέσματα όταν η αλληλουχία εισαχθεί από τη μικροπύλη ή μέσα από το χόριο, ακριβώς μετά τη γονιμοποίηση. Από μικρά αυγά που έχουν μαλακό χόριο, όπως, για παράδειγμα, τα αυγά του γριβαδιού, μπορεί να αφαιρεθεί το χόριο πριν από την εισαγωγή της ακολουθίας του γονιδίου. Αξίζει να αναφερθεί ότι θα πρέπει να υπάρχει παράλληλο πείραμα μάρτυρας και με κανονικά γονιμοποιημένα αυγά ώστε να γίνεται συγκριτική μελέτη του βαθμού επιβίωσης των ατόμων που προκύπτουν από τα διάφορα πρωτόκολλα εργασίας. Βέβαια, το μόνιμο ερώτημα είναι: άραγε δημιουργήθηκαν διαγενετικά άτομα; Μετά την εκκόλαψη, τα περισσότερα εργαστήρια χρησιμοποιούν δύο μεθοδολογίες για να απαντήσουν στο ερώτημα αυτό. Με τη μια μεθοδολογία απομονώνεται DNA από ένα μικρό μέρος του γόνου (μικρών ψαριών), ενώ το άλλο μέρος επιτρέπεται να αναπτυχθεί. Εναλλακτικά, επιτρέπεται να αναπτυχθεί όλος ο γόνος και απομονώνεται DNA από βιοψίες μεγάλων ατόμων. Και στις δύο περιπτώσεις, είναι δύσκολο να αποφασιστεί αν τα άτομα είναι πλήρως διαγενετικά ή χίμαιρες. Οι ερευνητές συμπεραίνουν την ύπαρξη μωσαϊκισμού ανάμεσα στους διαγενετικούς απογόνους, όταν διαπιστώνουν για μερικά άτομα χαμηλό ποσοστό υβριδισμού DNA/DNA. Πάντως, για να αποκλειστεί ή να επιβεβαιωθεί ο χιμαιρισμός, πρέπει να απομονωθεί το DNA από όλους τους ιστούς μεγάλων ατόμων και να ελεγχθεί είτε με υβριδισμό με κατάλληλους DNA εκκινητές είτε με αλληλούχιση. Είναι ενδιαφέρον ότι στα αληθινά διαγενετικά άτομα διαπιστώνεται, πολλές φορές, η ενσωμάτωση όχι μόνο μιας αλλά πολλών ακολουθιών από αυτές που εντέθηκαν στο κυτταρόπλασμα. Η ενσωμάτωση του DNA πιστεύεται πως συμβαίνει με μηχανισμό ανασυνδυασμού ανάμεσα στο γονιδίωμα του δέκτη-οργανισμού και στο ξένο DNA. Είναι ευνόητο ότι από το γονιδίωμα των διαγενετικών ζώων απουσιάζει μέρος του γενετικού υλικού ή υπάρχει προσθήκη ξένου DNA. Παρόλα αυτά, δεν αναφέρθηκαν εμφανείς γενετικές ανωμαλίες σε τέτοια άτομα. Ίσως αυτό συμβαίνει επειδή το γονιδίωμα των ευκαρυωτικών οργανισμών είναι τόσο μεγάλο, ώστε τυχαίες απώλειες μέχρι 10 Κb φαίνεται πως δεν συνεπάγονται γενετικές διαταραχές. Δεν μπορεί όμως να αποκλειστεί και η περίπτωση μερικά ελλείμματα του γονιδιώματος, που δημιουργούνται κατά την ενσωμάτωση του DNA, να είναι θανατηφόρα. Η υπόθεση αυτή φαίνεται πιο πιθανή επειδή σε τέτοια πειράματα παρατηρείται μειωμένη βιωσιμότητα τόσο των ωαρίων, όσο και των εμβρύων που εκκολάπτονται από αυτά. Έκφραση του ενσωματωμένου γονιδίου. Η έκφραση του ενσωματωμένου γονιδίου μπορεί να εντοπιστεί με μελέτη του mRNA των διαγενετικών ιστών ή ατόμων. Τεκμηριωμένες αποδείξεις παίρνονται και με ανοσολογικές τεχνικές στο πρωτεϊνικό επίπεδο. Με αυτές τις τεχνικές, στο ριζόψαρο έχει αποδειχθεί η έκφραση του γονιδίου της κρυσταλλίνης των πουλερικών, με χρήση προαγωγέα από κοτόπουλα. Ωστόσο, το πρότυπο της έκφρασης του γονιδίου δεν ήταν φυσιολογικό. Αντίθετα, άλλοι ερευνητές δεν πέτυχαν έκφραση γονιδίου στην ιριδίζουσα πέστροφα χρησιμοποιώντας τον προαγωγέα του γονιδίου της μεταλλοθειονίνης ποντικού. Όταν όμως στα πειράματα χρησιμοποιήθηκε προαγωγέας του γονιδίου της μεταλλοθειονίνης των ψαριών, παρατηρήθηκε τουλάχιστο 10πλάσια έκφραση σε σχέση με την έκφραση του αντίστοιχου γονιδίου των θηλαστικών. Τέτοια πειράματα δείχνουν πόσο απαραίτητο είναι να απομονωθούν προαγωγείς και από άλλα είδη ψαριών. Επιλογή κατάλληλων γονιδίων. Στους αρχικούς πειραματισμούς δημιουργίας διαγενετικών ειδών επιδιωκόταν από τη μια μεριά να αναπτυχθεί η κατάλληλη τεχνολογία και από την άλλη να μελετηθεί ο τρόπος ρύθμισης της έκφρασης των γονιδίων των ευκαρυωτικών οργανισμών. Ακόμη όμως και σ’ αυτό το στάδιο, είχαν επιλεγεί προς πειραματισμό τα γονίδια της αυξητικής ορμόνης και της μεταλλοθειονίνης, με την ελπίδα ότι η ενσωμάτωσή τους θα έδινε κάποια πλεονεκτήματα στα διαγενετικά ζώα. Τώρα όμως που έχει αναπτυχθεί η τεχνολογία, και ορισμένα κέντρα έφθασαν να παράγουν την ανθρώπινη ινσουλίνη στο γάλα διαγενετικών προβάτων, είναι επόμενο να υπάρχει σοβαρός προβληματισμός για το ποιο γονίδιο θα επιλεγεί προς πειραματισμό.

35 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
7.3 Γονίδια που χρησιμοποιούνται κατά τη γενετική τροποποίηση στις υδατο-καλλιέργειες Γενικά, η τροποποίηση του γενετικού υλικού των ψαριών γίνεται με γονίδια που καθορίζουν χαρακτηριστικά, όπως τα παρακάτω: πρωτεΐνες που παρέχουν προστασία σε μόλυνση από ιούς, πρωτεΐνες που οδηγούν στην αύξηση της παραγωγής και τη βελτίωση της ποιότητας των παραγόμενων προϊόντων, και, τέλος, ανθρώπινες πρωτεΐνες για παραγωγή φαρμάκων ή άλλων ανθρώπινων ουσιών. Παρακάτω αναφέρονται συγκεκριμένα παραδείγματα. Γονίδιο αυξητικής ορμόνης (σωματοτροπίνη). Η αυξητική ορμόνη είναι πολυπεπτίδιο. Η σύνθεσή της γίνεται στην υπόφυση των σπονδυλωτών. Συμμετέχει γενικά στην αύξηση και την ανάπτυξη του σώματος ρυθμίζοντας πολλές μεταβολικές λειτουργίες. Ως πρωτεΐνη, αν χορηγηθεί με την τροφή, επέρχεται διάσπασή της στο έντερο των Σπονδυλωτών. Σε νεαρά στάδια όμως ορισμένων ειδών ψαριών, μπορεί να περάσει από τα κύτταρα του εντερικού σωλήνα και να ασκήσει τη δράση της μέσω της διατροφικής οδού. Όπως και άλλες πρωτεϊνικές ορμόνες, π.χ. η ινσουλίνη, είναι λειτουργική όταν χορηγηθεί με ένεση. Έχει, για παράδειγμα, αποδειχθεί ότι όταν η αυξητική ορμόνη των πουλιών και των θηλαστικών ενεθεί στα σολομοειδή, αυτή εξακολουθεί να παραμένει λειτουργική. Το γονίδιο της αυξητικής ορμόνης έχει ενσωματωθεί στο γριβάδι, την ιριδίζουσα πέστροφα, τον σολομό του Ειρηνικού, την τιλάπια και πάρα πολλά άλλα είδη. Έχει δώσει πολύ καλά αποτελέσματα (επίπεδα αύξησης έως και 30 φορές παραπάνω), εκτός από στελέχη όπου με κλασικά προγράμματα επιλογής οι παραγωγοί είχαν ήδη καταφέρει να βελτιώσουν σημαντικά την αύξηση. Γονίδιο εκλυτικού παράγοντα της αυξητικής ορμόνης. Ο εκλυτικός παράγοντας της αυξητικής ορμόνης είναι μια πρωτεΐνη. Η λειτουργία της είναι να διεγείρει τα κύτταρα του κατάλληλου λοβού της υπόφυσης για να εκκρίνουν αυξημένες ποσότητες σωματοτροπίνης. Διαγενετικά ζώα για αυτό το γονίδιο ίσως απελευθερώνουν μεγαλύτερες ποσότητες της φυσικής ορμόνης. Γονίδια μεταλλοθειονινών. Οι μεταλλοθειονίνες είναι πρωτεΐνες που δεσμεύουν βαρέα μέταλλα στα κύτταρα, ειδικότερα κάδμιο, χαλκό, ψευδάργυρο και υδράργυρο. Επιτελούν διττή λειτουργία στο κύτταρο. Η μια, ίσως η κύρια λειτουργία, είναι να παρέχουν ψευδάργυρο στα ένζυμα που δρουν μόνο παρουσία του μετάλλου αυτού. Η άλλη σπουδαία λειτουργία τους είναι η αποτοξίνωση, που περιλαμβάνει τη δέσμευση των προηγούμενων μετάλλων στο κύτταρο και την έκκριση του παραγόμενου συμπλόκου μέσω του ήπατος. Οι μεταλλοθειονίνες είναι επαγόμενες πρωτεΐνες και η σύνθεσή τους αυξάνει σημαντικά με την παρουσία βαρέων μετάλλων. Έτσι, για παράδειγμα, όταν ψάρια ή άλλα ζώα εκτεθούν σε ημιθανατογόνες δόσεις διάφορων μετάλλων, όπως το κάδμιο ή ο χαλκός, είναι ικανά να αυτοπροστατευτούν με αυξημένη σύνθεση μεταλλοθειονινών. Ειδικότερα, οι διαγενετικοί οργανισμοί που ενδέχεται να έχουν αυξημένο αριθμό αυτών των γονιδιακών ακολουθιών, θα μπορέσουν να ζήσουν καλύτερα σε περιβάλλον που έστω και πρόσκαιρα θα ρυπανθεί από βαρέα μέταλλα. Ήδη έχει επιτευχθεί η εισαγωγή του ανασυνδυασμένου προαγωγέα της μεταλλοθειονίνης-σωματοτροπίνης στην ιριδίζουσα πέστροφα και η κληρονόμηση της ακολουθίας αυτής στους απογόνους. Γονίδια που καθορίζουν αντιψυκτικές πρωτεΐνες (AFPs). Αρκετά είδη ψαριών που ζουν στην Αρκτική ή την Ανταρκτική έχουν στο αίμα τους πρωτεΐνες, οι οποίες μειώνουν τη φυσική θερμοκρασία ψύξης του αίματος και των ιστών, δρώντας ως φυσικά αντιψυκτικά. Από τον πλευρονήκτη Pseudopleuronectes americanus έχει ήδη απομονωθεί το υπεύθυνο γονίδιο και έχει επιτευχθεί η ενσωμάτωσή του στον σολομό του Ατλαντικού. Ο τελευταίος δεν έχει φυσικές αντιψυκτικές πρωτεΐνες, με αποτέλεσμα τα ψάρια να πεθαίνουν στους ιχθυοκλωβούς όταν η θερμοκρασία πέσει κάτω από τους -0,7οC. Δυστυχώς τα διαγενετικά ψάρια δεν έδειξαν σημαντικά υψηλότερη αντοχή στο κρύο. Βέβαια, θα χρειαστεί χρόνος για να περάσουν αυτά τα ενδιαφέροντα πειράματα στην εφαρμογή και να αξιοποιηθούν εμπορικά τα αποτελέσματά τους. Γονίδια ανθεκτικότητας σε ασθένειες. Οι μελέτες απομόνωσης ακολουθιών DNA που να παρέχουν ανθεκτικότητα ή ευαισθησία στους φορείς τους βρίσκονται σε αρχικά στάδια στα ψάρια. Είναι ελπιδοφόρο

36 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
ότι ο συνδυασμός DNA μικροσυστοιχιών και αναλύσεων για QTLs ανθεκτικότητας αρχίζει να αποδίδει καρπούς και σε αυτό το θέμα για κάποια εμπορικά ψάρια. Ήδη υπάρχουν εφαρμογές σε γατόψαρα Ictalurus punctatus με ενσωμάτωση γονιδίου που παράγει κεκροπίνες, πρωτεΐνες που προσφέρουν ανθεκτικότητα σε πληθώρα Gram – αρνητικών βακτηρίων. Γονίδια δείκτες. Οι υπηρεσίες που κάνουν εμπλουτισμούς θα ήθελαν να γνωρίζουν ποια είναι η συνεισφορά του γόνου στο γονιδιακό απόθεμα του οικοσυστήματος, ένα ή δύο χρόνια μετά από τον εμπλουτισμό. Για τον σκοπό αυτό χρησιμοποιούν μορφολογικούς ή βιοχημικούς δείκτες ή διάφορα τεχνητά μέσα σήμανσης. Όλες αυτές οι μεθοδολογίες έχουν σοβαρά μειονεκτήματα. Η εναλλακτική μέθοδος που προτείνεται από μερικούς επιστήμονες είναι να δημιουργηθούν διαγενετικά άτομα, που θα έχουν όμως ενσωματωμένες στο γενετικό τους υλικό ανενεργές ακολουθίες DNA. Τα άτομα αυτά μπορούν να καταγραφούν με αξιοπιστία μέσα στο οικοσύστημα και το μεγάλο πλεονέκτημά τους είναι ότι, παρόλο που αποτελούν προϊόντα γενετικής μηχανικής, δεν φαίνεται να συνιστούν κίνδυνο αλλοίωσης της γενετικής σύστασης των οργανισμών του όλου οικοσυστήματος.
8. Εφαρμογές της γενετικής μηχανικής στις υδατοκαλλιέργειες. Προβληματισμοί
8.1 Εφαρμογές της γενετικής μηχανικής στις υδατοκαλλιέργειες Οι υδατοκαλλιέργειες ως βιολογικοί αντιδραστήρες. Στη βιβλιογραφία αναφέρονται ήδη πετυχημένα πειράματα παραγωγής φαρμακευτικών προϊόντων από οικόσιτα διαγενετικά ζώα, όπως η παραγωγή του αντιαιμορροφιλικού παράγοντα στο γάλα διαγενετικών προβάτων. Στα ψάρια θα μπορούσε να γίνει παραγωγή της ανθρώπινης ινσουλίνης στο συκώτι διαγενετικών ψαριών. Δημιουργία ψαριών με βελτιωμένη αύξηση/απόδοση. Η ενσωμάτωση του γονιδίου της αυξητικής ορμόνης της ιριδίζουσας πέστροφας σε κυπρίνο (γριβάδι) δημιούργησε ψάρια που είχαν ταχύτερο ρυθμό αύξησης, με αποτέλεσμα το βάρος τους να είναι κατά 22% μεγαλύτερο από τους μη γενετικά τροποποιημένους μάρτυρες. Πιο εντυπωσιακά αποτελέσματα έδωσε η ενσωμάτωση του γονιδίου της αυξητικής ορμόνης του κόκκινου σολομού και η έκφρασή του στον βλεννογόνο αδένα και το ήπαρ του σολομού του Ειρηνικού. Ο Devlin και οι συνεργάτες του (1994) δημιούργησαν διαγενετικά ψάρια τα οποία στην ηλικία των 14 μηνών ήταν 11 φορές βαρύτερα από τους μη γενετικώς τροποποιημένους μάρτυρες. Πρόσφατα, παρόμοια εντυπωσιακά αποτελέσματα αναφέρονται στην ιριδίζουσα πέστροφα, το γατόψαρο και το χρυσόψαρο. Τέλος, βιοτεχνολογικές εταιρείες ισχυρίζονται ότι ανέπτυξαν νέους μηχανισμούς και νέα οχήματα μεταφοράς επιθυμητών γονιδίων δημιουργίας υπερ-ψαριών (αυξάνονται γρηγορότερα και γίνονται μεγαλύτερα) κατάλληλων για ιχθυοκαλλιέργειες. Δημιουργία ψαριών με επιθυμητή ποιοτική σύσταση. Δημιουργούνται διαγενετικοί οργανισμοί που παράγουν προϊόντα με διαφορετική σύσταση ή βελτιωμένα τεχνολογικά χαρακτηριστικά. Για παράδειγμα, ο διαγενετικός σολομός βρέθηκε ότι περιέχει στο πλάσμα του χαμηλότερη συγκέντρωση τριγλυκεριδίων και παρουσιάζει γρηγορότερο μεταβολισμό των λιπιδίων της τροφής. Ο ρυθμός μεταβολισμού και η κατανάλωση των λιπιδίων των διαγενετικών σολομών μοιάζει περισσότερο με εκείνη της καλλιεργούμενης ιριδίζουσας πέστροφας, παρά με τους άγριους συγγενείς του είδους. Μείωση του κόστους παραγωγής προϊόντων. Ο διαγενετικός σολομός της εταιρείας A/F Proteins (AquaBounty), που καλλιεργείται σε εγκαταστάσεις του Καναδά και της Καλιφόρνιας, αναπτύσσεται ταχύτερα (φθάνει το εμπορικό βάρος των 4 κιλών στους 14 μήνες, ενώ τα φυσικά αδέλφια του χρειάζονται διπλάσιο χρόνο για να φθάσουν το ίδιο βάρος) και καταναλώνει λιγότερη τροφή από τα άγρια αδέλφια του. Ως εκ τούτου, τα διαγενετικά ψάρια θα είναι πολύ φθηνότερα από τα φυσικά αδέλφια τους. Από όλα τα προηγούμενα συνάγεται ότι στην παραγωγή των διαγενετικών οργανισμών υπάρχει και η οικονομική διάσταση, την οποία ίσως δεν θα μπορέσουν να αγνοήσουν μελλοντικά οι ιχθυοκαλλιεργητές, οι βιομηχανίες και οι κυβερνήσεις.

37 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
Παραγωγή εμβολίων με γενετική μηχανική. Οι ιικές και οι βακτηριακές μολύνσεις των ψαριών έχουν ως αποτέλεσμα τη σημαντική μείωση της βιομάζας των ιχθυοπληθυσμών και των ιχθυοκαλλιεργειών της πέστροφας, του σολομού, του γατόψαρου, κ.λπ. Ο ιός της μολυσματώδους αιμοποιητικής νέκρωσης (ΙΗΝV) προκαλεί σοβαρή ασθένεια στους πληθυσμούς της πέστροφας. Με μεθοδολογίες γενετικής μηχανικής αναπτύχθηκε αποτελεσματικό εμβόλιο ενάντια στην υπομονάδα του ιικού γονιδίου της γλυκοπρωτεΐνης κατάλληλο για την αντιμετώπιση του ιού σε ιχθυοκαλλιέργειες. Διαχείριση και απελευθέρωση διαγενετικών ζώων. Μια από τις κύριες φροντίδες των ερευνητών είναι και η αποφυγή της τυχαίας απελευθέρωσης διαγενετικών οργανισμών στο φυσικό περιβάλλον. Οι υδατοκαλλιεργητές, αν χρησιμοποιήσουν διαγενετικά ζώα, θα πρέπει να είναι ιδιαίτερα προσεκτικοί στην τυχαία απελευθέρωση τέτοιων οργανισμών στα φυσικά υδάτινα οικοσυστήματα. Η απελευθέρωσή τους ενδέχεται να έχει απρόβλεπτες συνέπειες στην ισορροπία του οικοσυστήματος. Γι’ αυτό τα διαγενετικά ζώα θα πρέπει να εκτρέφονται σε συνθήκες που να μην επιτρέπουν τη διαφυγή τους σε φυσικά υδάτινα συστήματα. Μια άλλη λύση είναι, τουλάχιστο στα αρχικά πειράματα, η χρησιμοποίηση τριπλοειδών ειδών ή ειδών που η διαφυγή τους στο περιβάλλον δεν προκαλεί κινδύνους.
8.2 Οι πιθανοί οικολογικοί κίνδυνοι από τη διαφυγή καλλιεργούμενων διαγενετικών ψαριών
8.2.1 Ο διαγενετικός σολομός της Aquabounty Η Καναδική Εταιρεία Aqua Bounty Farms (http://aquabounty.com/) υπέβαλλε αίτηση το 2009 στην Υπηρεσία Τροφίμων και Φαρμάκων (FDA) των ΗΠΑ να επιτρέψει την εμπορική καλλιέργεια του ΓΤ στείρου «υπέρ-σολομού», ο οποίος μεγαλώνει στο μισό του χρόνου που χρειάζεται ο άγριος σολομός, όπως φαίνεται στο διάγραμμα της Εικόνας που παρουσιάζεται στην ιστοσελίδα https://aquabounty.com/innovation/technology/. Από την ημερομηνία που ανακοινώθηκε η υποβολή της αίτησης, το τηλεφωνικό κέντρο της εταιρείας άρχισε να δέχεται καταιγιστικά τηλεφωνήματα από δημοσιογράφους οι οποίοι υποβάλλουν “ενοχλητικές” ερωτήσεις για δημιουργία ψαριών Frankenstein. Με απλά λόγια, η εταιρεία βρέθηκε μέσα σε ένα κυκεώνα αντιδικιών που αφορούν τους ΓΤΟ και τα νεοφανή τρόφιμα, που ως γνωστόν κορυφώθηκε στην Ευρώπη και άρχισε να ανακυκλώνεται και από τα ΜΜΕ της βόρεια Αμερικής. Ανεξάρτητη μελέτη στον διαγενετικό σολομό της εταιρείας υποστήριξε ότι ο διαγενετικός σολομός θα καταστρέψει τους άγριους πληθυσμούς του είδους, και αρκετοί επιστήμονες προβληματίζονται για τη μελλοντική χρησιμοποίησή του λόγω οικολογικών συνεπειών. Ο λόγος είναι ότι τα διαγενετικά θηλυκά ψάρια ζουν λιγότερο και λόγω του μεγαλυτέρου μεγέθους τους, προσελκύουν περισσότερα αρσενικά, οπότε ο αρνητικός χαρακτήρας γρήγορα θα εξαπλωθεί στους φυσικούς πληθυσμούς του είδους και ο ιχθυοπληθυσμός θα καταστραφεί. Η αρμόδια αμερικανική υπηρεσία δεν χορήγησε άδεια καλλιέργειας και διάθεσης του προϊόντος στην αγορά και ζήτησε να διεξαχθούν πρόσθετες έρευνες προτού αποφασίσει τελεσίδικα επί του θέματος. Τα ερωτήματα που απαίτησε να διερευνηθούν ήταν αν: α) οι ΓΤ σολομοί είναι κατάλληλοι για κατανάλωση από τον άνθρωπο, β) επηρεάζεται η υγεία των σολομών με την ενσωμάτωση στο γονιδίωμά τους και την έκφραση του γονιδίου της αυξητικής ορμόνης από άλλο είδος ψαριού, και γ) υπάρχουν περιβαλλοντικές επιπτώσεις από την καλλιέργεια και την τυχαία απελευθέρωση των ΓΤ σολομών. Η Εταιρεία Aqua Bounty Farms, προκειμένου να καθησυχάσει τους δικαιολογημένους φόβους μερίδας του τύπου και του κοινού, έχει πια ενσωματώσει εναλλακτικά πρωτόκολλα καλλιέργειας του ΓΤ σολομού, όπως: α) Τη δημιουργία μόνο στείρων και άγονων ΓΤ σολομών και την καλλιέργειά τους σε τεχνητές λίμνες. Ειδικότερα, τα καλλιεργούμενα ψάρια καθώς και τα διαγενετικά ψάρια να είναι τριπλοειδή αντί διπλοειδή, γιατί τα πρώτα δεν αναπαράγονται. Το μόνο πρόβλημα είναι ότι η μεθοδολογία δημιουργίας στείρων και

38 | Κ α λ λ ι έ ρ γ ε ι α ι χ θ ύ ω ν : α ρ χ έ ς γ ε ν ε τ ι κ ή ς β ε λ τ ί ω σ η ς
μη γόνιμων ψαριών δεν είναι 100 % αξιόπιστη. Ακόμη και αν απελευθερωθεί ένα αυγό, μπορεί μεσοπρόθεσμα να προκύψουν περιβαλλοντικά προβλήματα. β) Την καλλιέργεια ή εκτροφή σε χώρους που να μην επικοινωνούν με το φυσικό περιβάλλον. Έτσι, προτάθηκε η καλλιέργειά του ΓΤ σολομού στις δυτικές και όχι τις ανατολικές ακτές της βόρειας Αμερικής, όπου δεν υπάρχουν φυσικοί πληθυσμοί σολομού του Ατλαντικού. Επιπλέον, οι ιχθυοκαλλιέργειες γίνονται στη στεριά, αντί στη θάλασσα. Με αυτόν τον τρόπο η εταιρεία προσπάθησε να της χορηγηθεί η πολυπόθητη άδεια για τη δημιουργία και ανάπτυξη οικονομικά ανταγωνιστικής και βιώσιμης υδατοκαλλιεργητικής μονάδας. Η τελική έγκριση από τον FDA δόθηκε τον Νοέμβριο του 2015. Δείτε ακόμη στο διαδίκτυο: https://www.youtube.com/watch?v=qhMgtwQf9OM και https://www.youtube.com/watch?v=lbBzdnhKdZs
8.2.2 Προβληματισμοί από την εμπορία γενετικά τροποποιημένου διακοσμητικού ψαριού Στις ΗΠΑ από το 2003 πωλείται ελεύθερα το πρώτο ΓΤ τροπικό ψάρι με την εμπορική ονομασία GloFish (http://www.glofish.com/). Το ψάρι αυτό ήδη κολυμπάει σε χιλιάδες ενυδρεία καταστημάτων και οικιών. Το διαγενετικό αυτό ψάρι ζέβρα φέρει στο γονιδίωμά του ένα γονίδιο από κοράλλι που κωδικοποιεί μια πρωτεΐνη, η οποία φωσφορίζει εκπέμποντας κόκκινο φως όταν εκτεθεί σε υπεριώδες φως. Στη συνέχεια, κυκλοφόρησαν μπλε, πορτοκαλί, πράσινα και μωβ φωσφορίζοντα ψάρια (και σε άλλα δύο είδη). Όλες οι σχετικές υπηρεσίες των ΗΠΑ (Υπηρεσία περιβάλλοντος, Διεύθυνση Γεωργίας, Διεύθυνση Άγριας Πανίδας) δήλωσαν αρχικά αναρμόδιες να αποφασίσουν για τους ελέγχους που πρέπει να γίνουν πριν επιτραπεί η πώληση του ΓΤ ψαριού. Πολλοί υποστηρίζουν ότι αυτό ακριβώς το γεγονός δείχνει ότι δεν γίνεται επισταμένη αξιολόγηση για πιθανές αρνητικές επιπτώσεις των τμημάτων του DΝΑ που μεταφέρθηκαν στο ΓΤ ψάρι. Μάλιστα, προσθέτουν ότι ίσως η μεταφορά των νέων τμημάτων DNA σε άγρια στελέχη ενδέχεται να μειώσει την αναπαραγωγική τους ικανότητα με δυσμενή αποτελέσματα για την επιβίωση των πληθυσμών του είδους στο φυσικό περιβάλλον. Παρ’ όλα αυτά, μέχρι και το 2014, έχουν δημοσιευτεί μελέτες που υποστηρίζουν ότι δεν αποτελεί κίνδυνο για το περιβάλλον. Στην ΕΕ, από την άλλη, απαγορεύεται η εισαγωγή και πώληση των ψαριών αυτών, τα οποία, ωστόσο, διακινούνται παράνομα.
8.3 Συμπεράσματα Στον επιστημονικό, αλλά και στον επιχειρηματικό κόσμο, αποτελεί βεβαιότητα, ότι η παραγωγή διαγενετικών οργανισμών και η καλλιέργειά τους θα συμβάλλει θετικά στην ανάπτυξη των υδατοκαλλιεργειών, κάτι που έχει ήδη συμβεί σε άλλα οικόσιτα ζώα με οικονομική σημασία. Ωστόσο, οι υδατοκαλλιέργειες παρουσιάζουν ειδικότερα προβλήματα, όπως είναι η ευκολία διαφυγής των οργανισμών και το γεγονός ότι μπορούν να πωληθούν ως άγρια στους καταναλωτές. Παρόλα τα προβλήματα, το ενδιαφέρον κυβερνήσεων, επενδυτών και ερευνητών φαίνεται πως θα επιτρέψει τη γρήγορη τροποποίηση διάφορων χαρακτηριστικών, όπως είναι ο ρυθμός αύξησης, η μυρωδιά, η ανθεκτικότητα στις ασθένειες και η χρησιμοποίηση τέτοιων βελτιωμένων ή πιο σωστά γενετικά τροποποιημένων ζώων για την αύξηση της παραγωγής. Είναι πλέον γεγονός ότι το θέμα της ανάπτυξης της γεωργικής παραγωγής με μεθοδολογίες γενετικής μηχανικής δημιουργεί πολυποίκιλες συζητήσεις. Ορισμένοι υποστηρίζουν ότι η μεθοδολογία δεν είναι ασφαλής, ενώ άλλοι επιμένουν ότι είναι ένας τρόπος για να καλυφθούν οι ανάγκες του κόσμου σε τροφή. Τώρα, μετά την αποκωδικοποίηση του DNA και ψαριών προέκυψε ένα ακόμη θέμα, που αφορά αυτά καθαυτά τα ψάρια. Το ερώτημα είναι: Ποιος είναι ο κύριος ιδιοκτήτης της γενετικής σύστασης και των αποθεμάτων ενός είδους ψαριού; Αρχίζουν πλέον να ενδιαφέρονται για τις γενετικές πηγές οι ιχθυοκαλλιεργητές, οι βιοτεχνολογικές και φαρμακευτικές εταιρείες, οι ΟΤΑ και οι κυβερνήσεις. Βέβαια, το πραγματικό ερώτημα είναι: Ποιος θα ωφεληθεί από τη χρήση των γονιδίων των ψαριών; Ο ερευνητής που

39 | Υ δ α τ ο κ α λ λ ι έ ρ γ ε ι ε ς
προβαίνει στη δειγματοληψία, η τοπική κοινωνία που αλιεύει επί αιώνες το ιχθυοαπόθεμα, κ.λπ. Το πρόβλημα είναι μεγάλο και δεν έχει βρεθεί ακόμη η λύση του. Δυστυχώς, έχουν εντοπιστεί περιπτώσεις βιοπειρατείας, δηλ. συλλογής δειγμάτων χωρίς την έγκριση των αρμόδιων φορέων. Είναι καιρός λοιπόν να υπάρξει μια διεθνής συμφωνία για τη διαχείριση των γενετικών πηγών. Το θέμα είναι πλέον παγκόσμιας εμβέλειας, αν σκεφτούμε τις σημαντικές εφαρμογές της γενετικής στις υδατοκαλλιέργειες που φθάνουν μέχρι του σημείου της μεταφοράς γονιδίων από ένα είδος σε κάποιο άλλο. Στο πλαίσιο αυτό, υπάρχουν διεθνείς προσπάθειες όπως π.χ. η Κεντρική Αμερικανική Πρωτοβουλία για το Θαλάσσιο Γονιδίωμα (Central American Marine Genomic Initiative) με στόχο την καταγραφή και προστασία της βιοποικιλότητας. Η γενετική, η γονιδιωματική και η βιοτεχνολογία μας έχουν φέρει μπροστά σε έναν κόσμο για τον οποίο δεν είμαστε προετοιμασμένοι. Γι’ αυτό, απαιτείται σύνεση και εξαντλητικός έλεγχος στη χρήση αυτών των τεχνολογιών.
Επιλεγμένη βιβλιογραφία
Avise J.C., 1994. Molecular markers, Natural History and Evolution. Chapman & Hall, New York. Beaumont A., Boudry P., Hoare K., 2010. Biotechnology and Genetics in Fisheries and Aquaculture. Second Edition.
Wiley-Blackwell. Bernardi G., Wiley E.O., Mansour H., Miller M.R., Orti G., Haussler D., O'Brien S.J., Ryder O.A., Venkatesh B., 2012. The
fishes of Genome 10K. Marine Genomics 7: 3-6. Devlin R., Yesaki T.Y., Biagi C.A., Donaldson E.M., Swanson P., Chan W.-K., 1994. Extraordinary salmon growth. Nature
371: 209-210. Falconer D.S., Mackay T.F.C., 1996. Introduction to Quantitative Genetics. Fourth edition. Longman. Fletcher G.L., Rise M.L., 2011. Aquaculture Biotechnology. Wiley-Blackwell. Haffray P., Vauchez C., Vandeputte M., Linhart O., 1998. Different growth and processing traits in males and females of
European catfish, Silurus glanis. Aquatic Living Resources 11: 341-345. Κaraiskou N., Apostolidis A., Triantafyllidis A., Kouvatsi A., Triantaphyllidis C., 2003. Genetic identification and
phylogeny of three species of the genus Trachurus based on mtDNA analysis. Marine Biotechnology 5: 493- 504. Karaiskou Ν., Lappa Μ., Kalomoiris S., Oikonomidis G., Psaltopoulou C., Abatzopoulos T.J., Triantaphyllidis C.,
Triantafyllidis A., 2011. Genetic monitoring and effects of stocking practices on small Cyprinus carpio populations. Conservation Genetics 12:1299-1311.
Niller E., 2000. FDA researchers consider first transgenic fish. Nature Biotechnology, 2:143. Sagi A., Manor R., Ventura T., 2013. Gene Silencing in Crustaceans: From Basic Research to Biotechnologies. Genes 4:
620-645. Saroglia Μ., Zhanjiang L., 2012. Functional Genomics in Aquaculture. Wiley-Blackwell. Triantafyllidis A., Ozouf-Costaz C., Rab P., Suciu R., Karakousis Y., 1999a. Allozyme variation in European silurid
catfishes, Silurus glanis and Silurus aristotelis. Biochemical Systematics & Ecology, 27: 487-498. Triantafyllidis A., Abatzopoulos T.J., Economidis P.S., 1999b. Genetic differentiation and phylogenetic relationships
among Greek Silurus glanis and Silurus aristotelis (Pisces, Siluridae) populations, assessed by PCR-RFLP analysis of mitochondrial DNA segments. Heredity 82: 503-509.
Triantafyllidis A., Krieg F., Cottin C., Abatzopoulos T.J., Triantaphyllidis C., Guyomard R., 2002. Genetic structure and phylogeography of European catfish (Silurus glanis) populations. Molecular Ecology, 11: 1039-1055.
Wohlfarth G.W., Moav R., Hulata G., 1983. A genotype-environment interaction for growth rate in the common carp, growing in intensively manured ponds. Aquaculture, 33: 187-195.
Zhang P.J., Hayat M., Joyce C., Gonzalez-Villaseñor L.I., Lin C.M., Dunham R.A., Chen T.T., Powers D.A., 1990. Gene transfer, expression and inheritance of pRSV – rainbow trout-GH cDNA in the common carp, Cyprinus carpio. Molecular Reproduction & Development, 25: 3-13.
Zhang P.J., Hewit G.M., 1996. Nuclear integrations: Challenges for mitochondrial DNA-markers. TREE, 11: 247-251. Zhanjiang L., 2011. Next Generation Sequencing and Whole Genome Selection in Aquaculture. Wiley-Blackwell.


















