


![Hadronic interaction of η mesons with protons · η′ mesons were discovered over fourty years ago [33–35] their hadronic interaction with nucleons has not been established. The](https://static.fdocument.org/doc/165x107/5e8ead87ce94c0335659440f/hadronic-interaction-of-mesons-with-protons-a-mesons-were-discovered-over.jpg)




γλώσσες
Σελίδες
Νομικός


Carsten Münk Clinic for Gastroenterology, Hepatology and Infectiology
Heinrich-Heine-University Düsseldorf
HIV’s interaction with APOBEC3C (a work in progress)
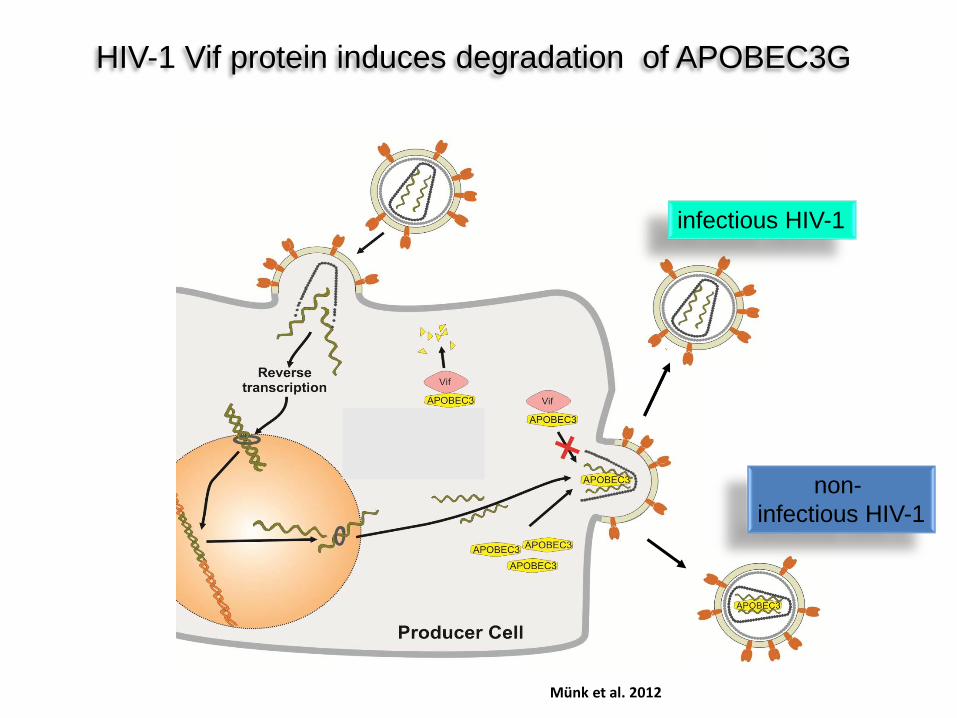
HIV-1 Vif protein induces degradation of APOBEC3G
infectious HIV-1
non- infectious HIV-1
Münk et al. 2012
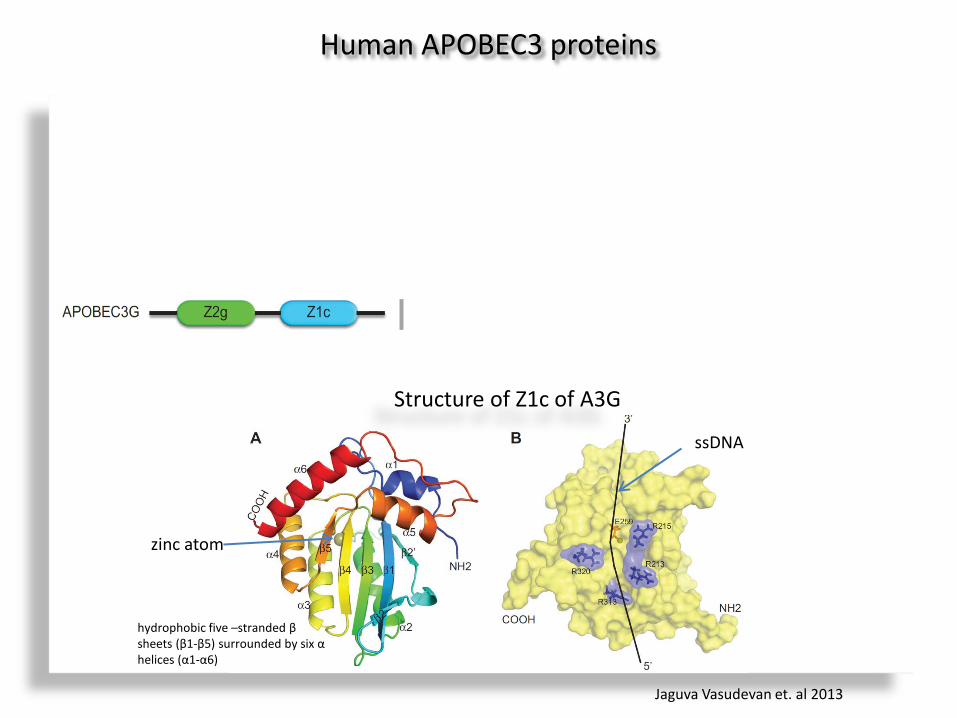
Human APOBEC3 proteins
Vif
Structure of Z1c of A3G
hydrophobic five –stranded β sheets (β1-β5) surrounded by six α helices (α1-α6)
zinc atom
ssDNA
Jaguva Vasudevan et. al 2013
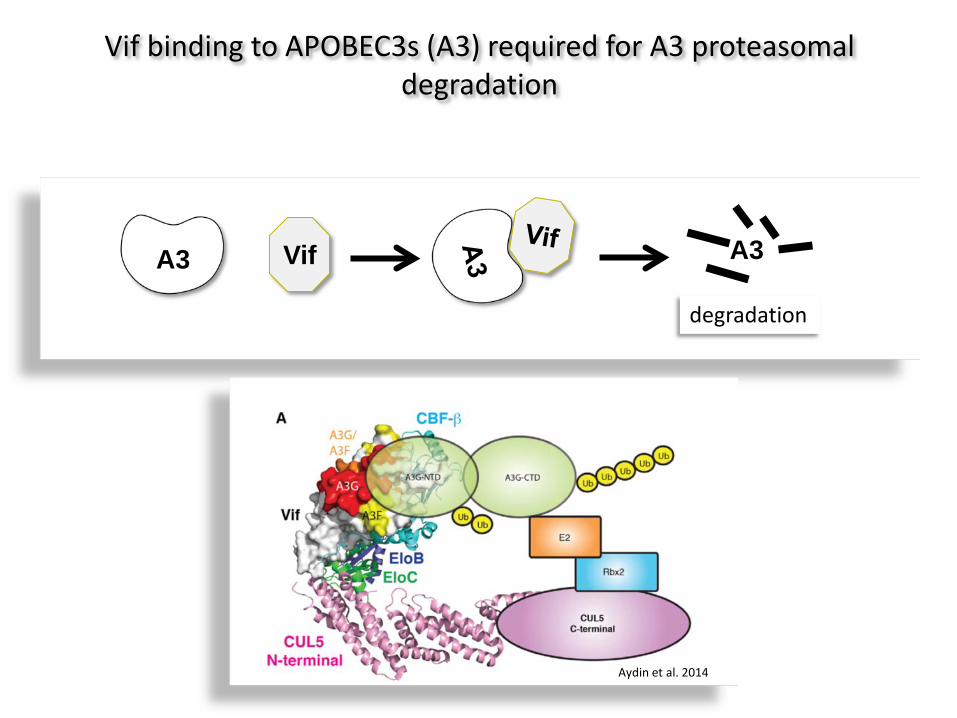
A3 Vif A3
degradation
Vif binding to APOBEC3s (A3) required for A3 proteasomal degradation
Aydin et al. 2014
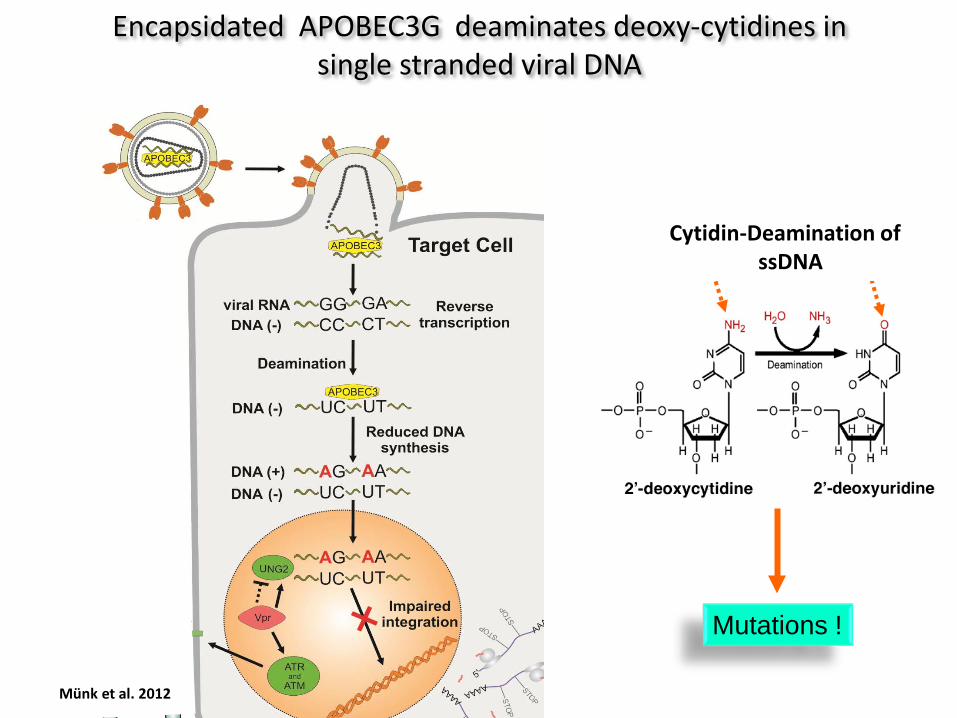
Mutations !
Cytidin-Deamination of ssDNA
Encapsidated APOBEC3G deaminates deoxy-cytidines in single stranded viral DNA
Münk et al. 2012
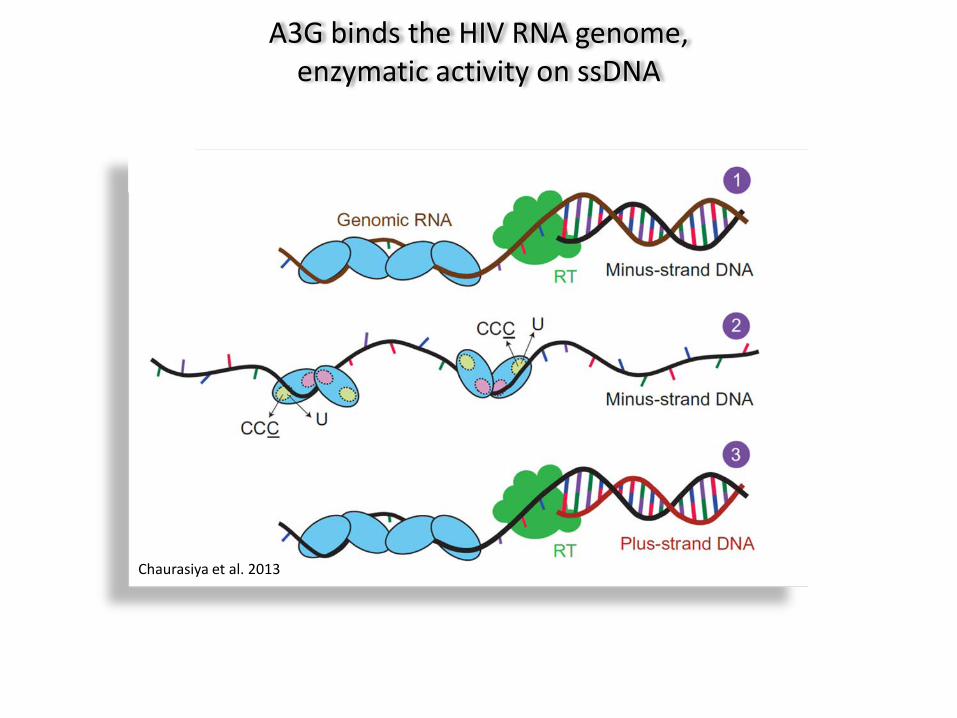
A3G binds the HIV RNA genome, enzymatic activity on ssDNA
Chaurasiya et al. 2013

HIV-1 Δvif
HIV-1 wt
each line represents an individual PCR on viral DNA
Encapsidated APOBEC3G deaminates deoxy-cytidines in single stranded viral DNA
PCR, 10 h post infection
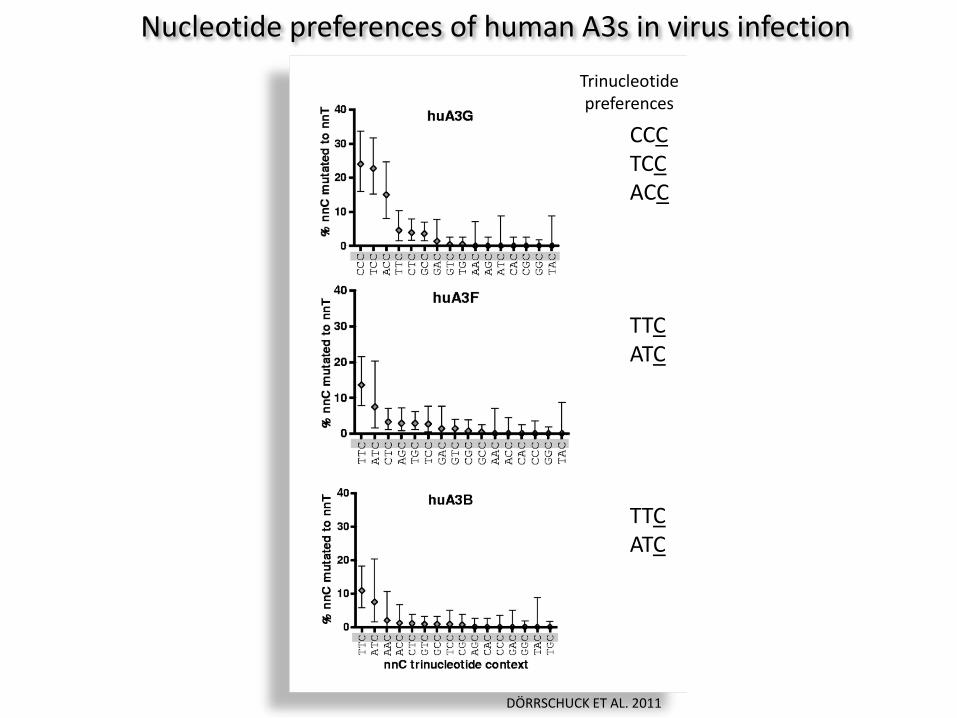
DÖRRSCHUCK ET AL. 2011
Trinucleotide preferences
Nucleotide preferences of human A3s in virus infection
CCC TCC ACC
TTC ATC
TTC ATC
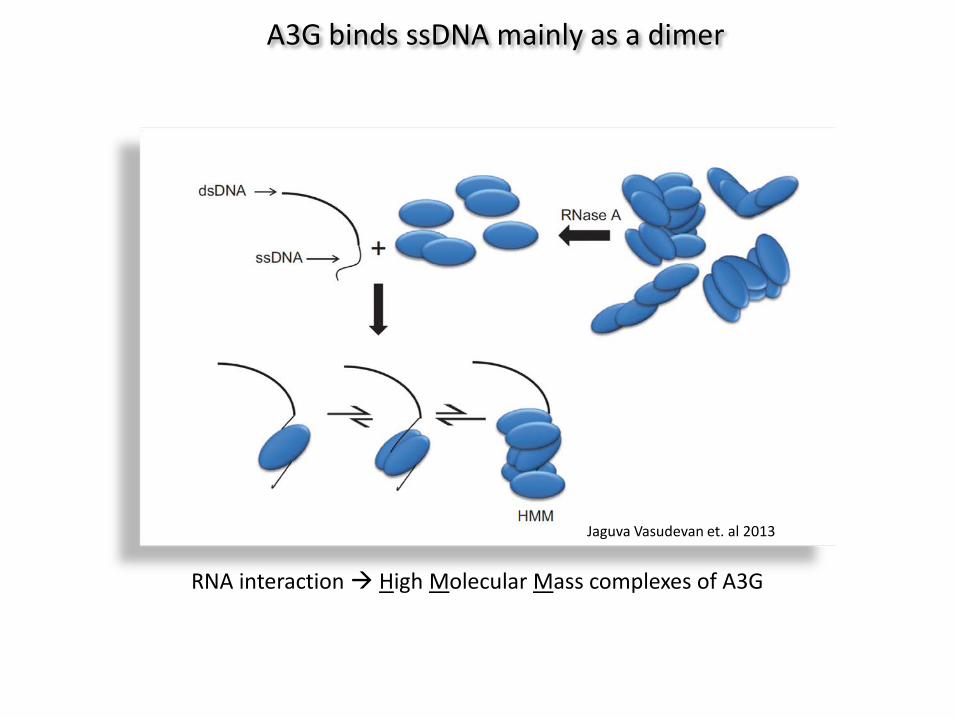
A3G binds ssDNA mainly as a dimer
RNA interaction High Molecular Mass complexes of A3G
Jaguva Vasudevan et. al 2013
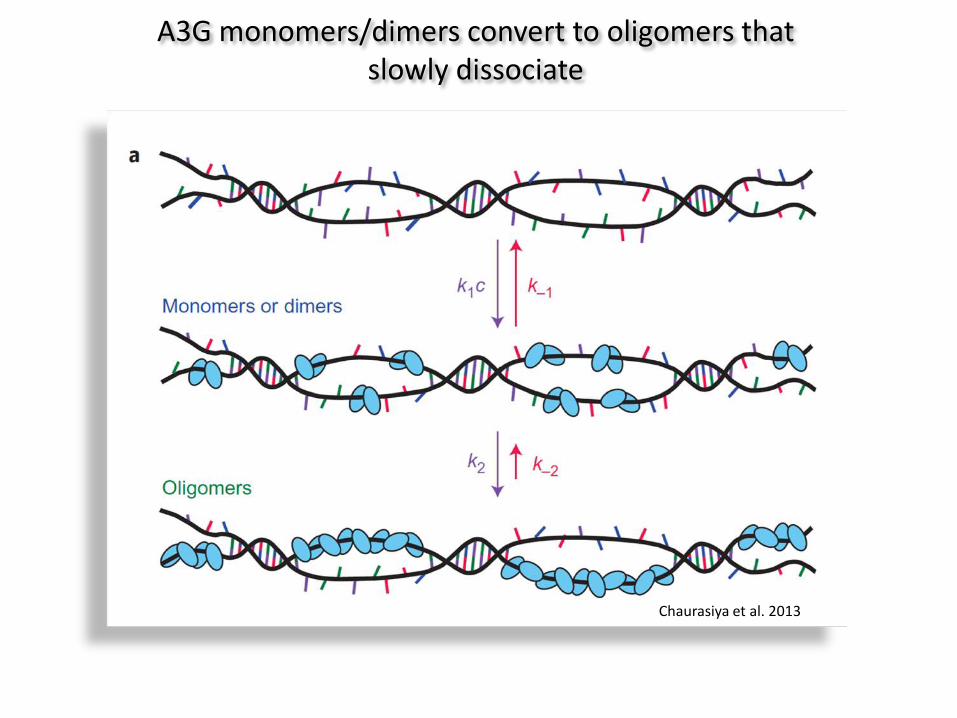
Chaurasiya et al. 2013
A3G monomers/dimers convert to oligomers that slowly dissociate
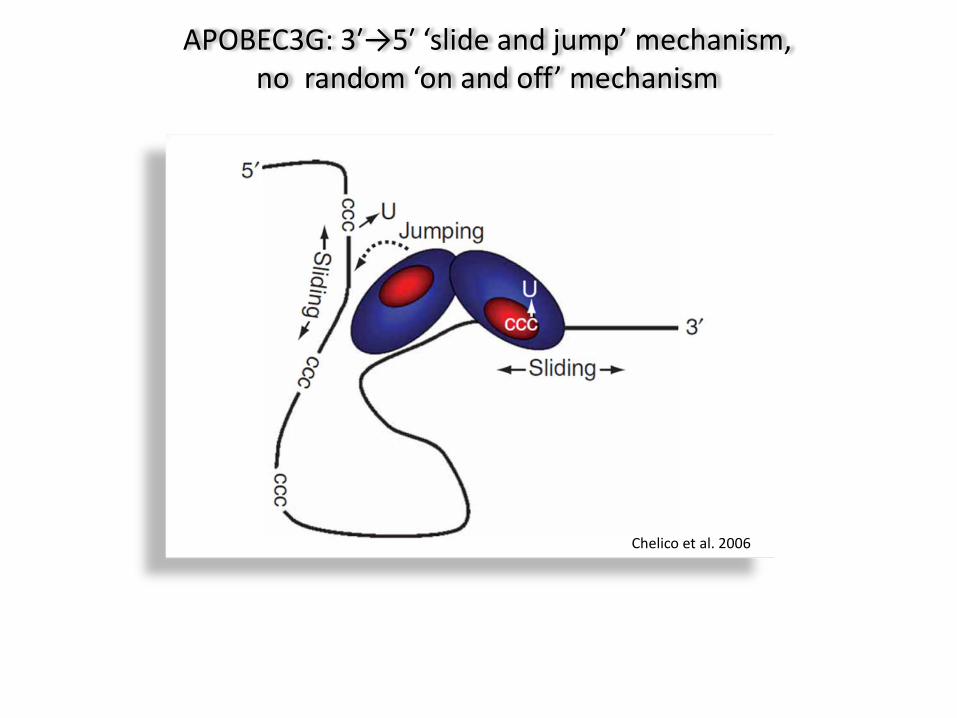
Chelico et al. 2006
APOBEC3G: 3′→5′ ‘slide and jump’ mechanism, no random ‘on and off’ mechanism
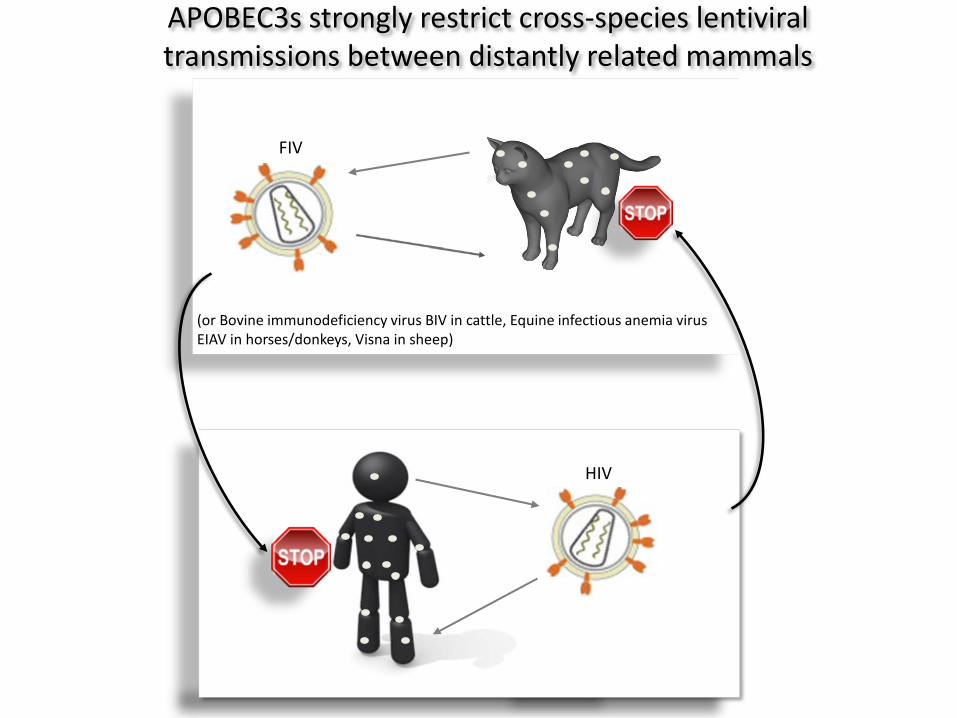
HIV
FIV
(or Bovine immunodeficiency virus BIV in cattle, Equine infectious anemia virus EIAV in horses/donkeys, Visna in sheep)
APOBEC3s strongly restrict cross-species lentiviral transmissions between distantly related mammals
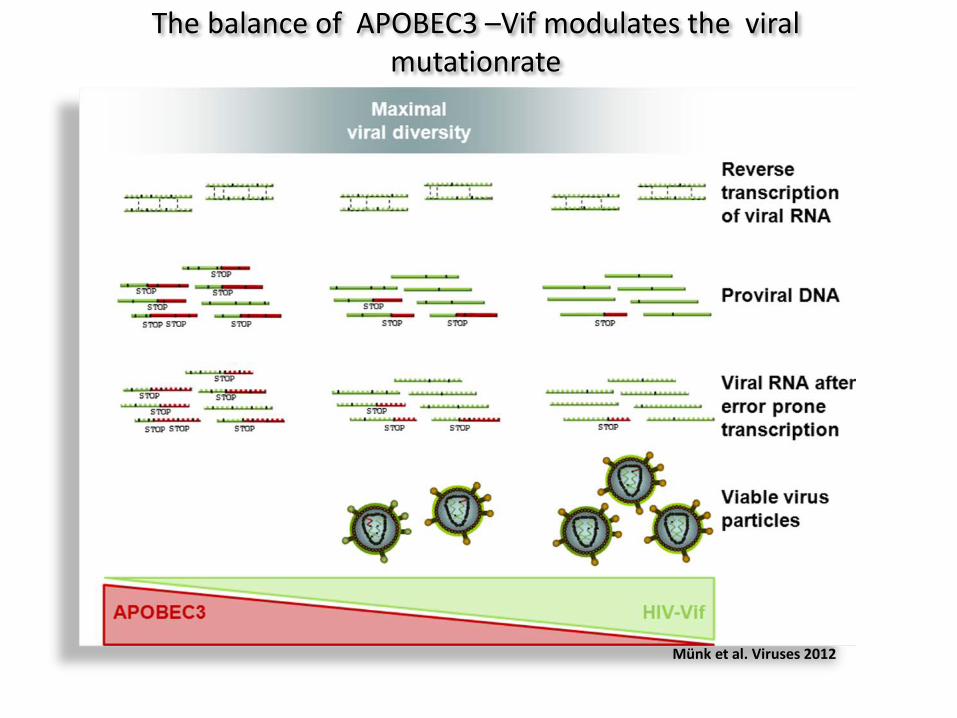
The balance of APOBEC3 –Vif modulates the viral mutationrate
Münk et al. Viruses 2012
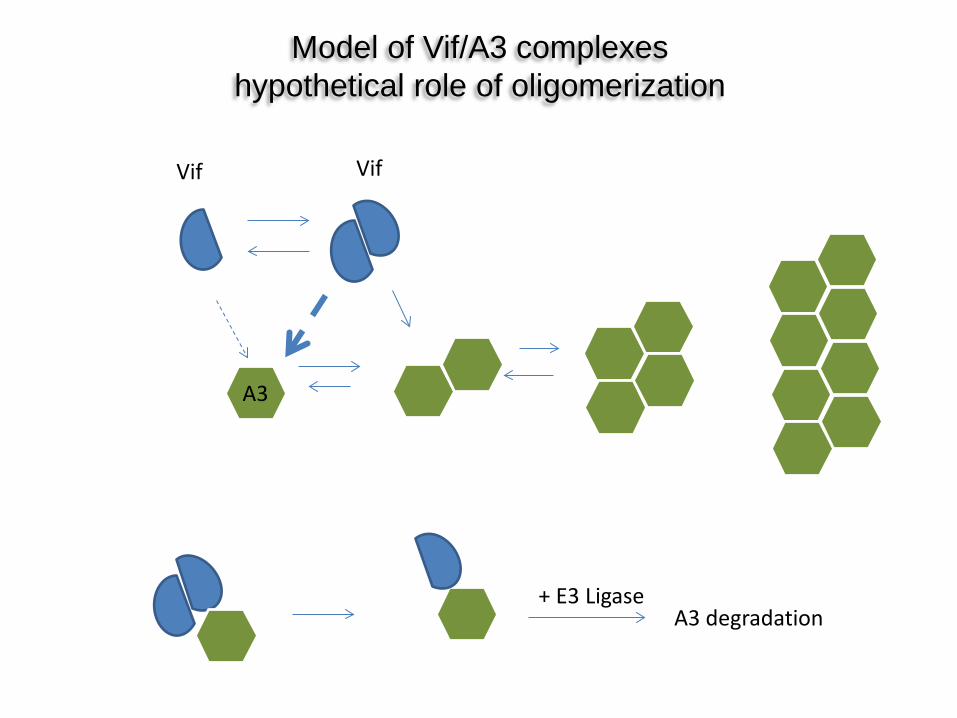
Model of Vif/A3 complexes hypothetical role of oligomerization
Vif Vif
A3
A3 degradation + E3 Ligase
active domain
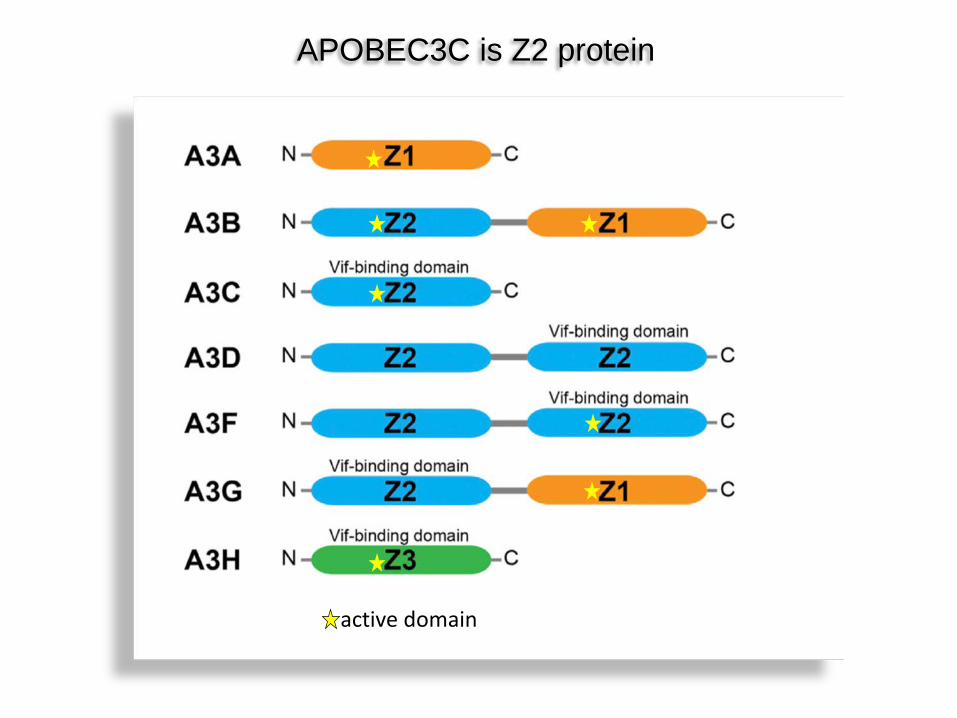
APOBEC3C is Z2 protein

A3C‘s interaction with HIV-1
1. where does Vif bind in A3C?
2. where does A3C bind in Vif
3. antiviral activity of A3C
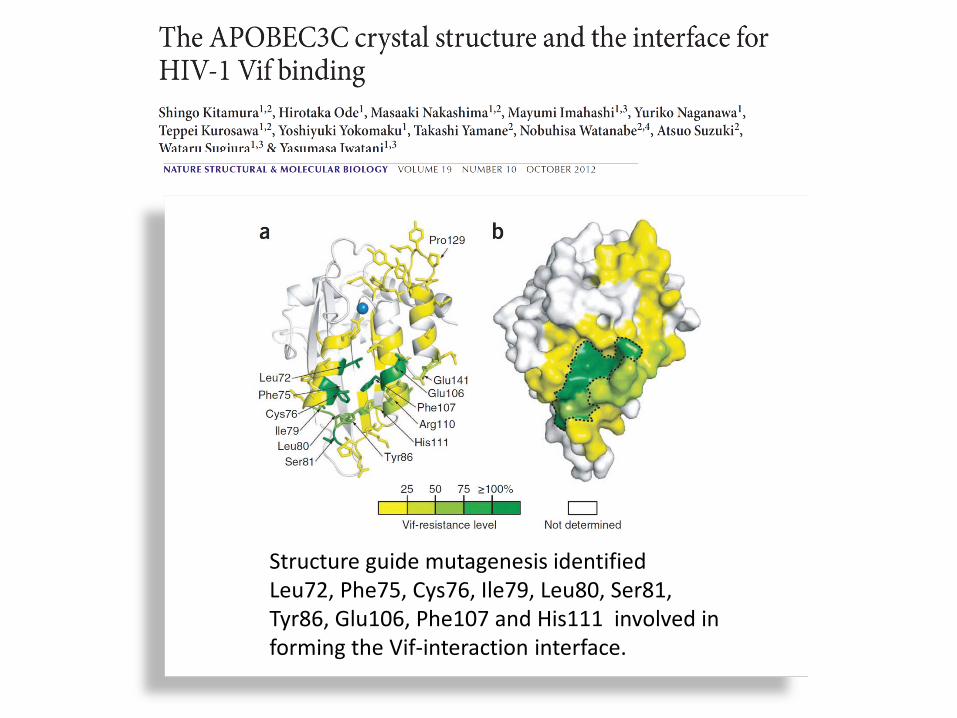
Structure guide mutagenesis identified Leu72, Phe75, Cys76, Ile79, Leu80, Ser81, Tyr86, Glu106, Phe107 and His111 involved in forming the Vif-interaction interface.
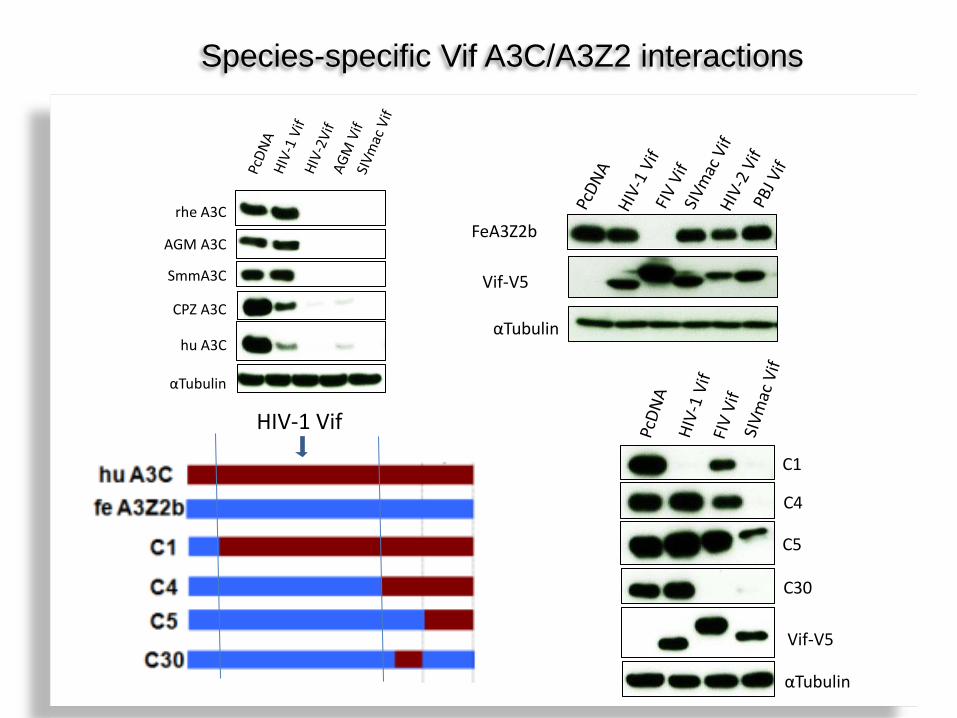
αTubulin
Vif-V5
C1
C4
C5
C30
HIV-1 Vif
αTubulin
hu A3C
CPZ A3C
rhe A3C
AGM A3C
SmmA3C
FeA3Z2b
Vif-V5
αTubulin
Species-specific Vif A3C/A3Z2 interactions
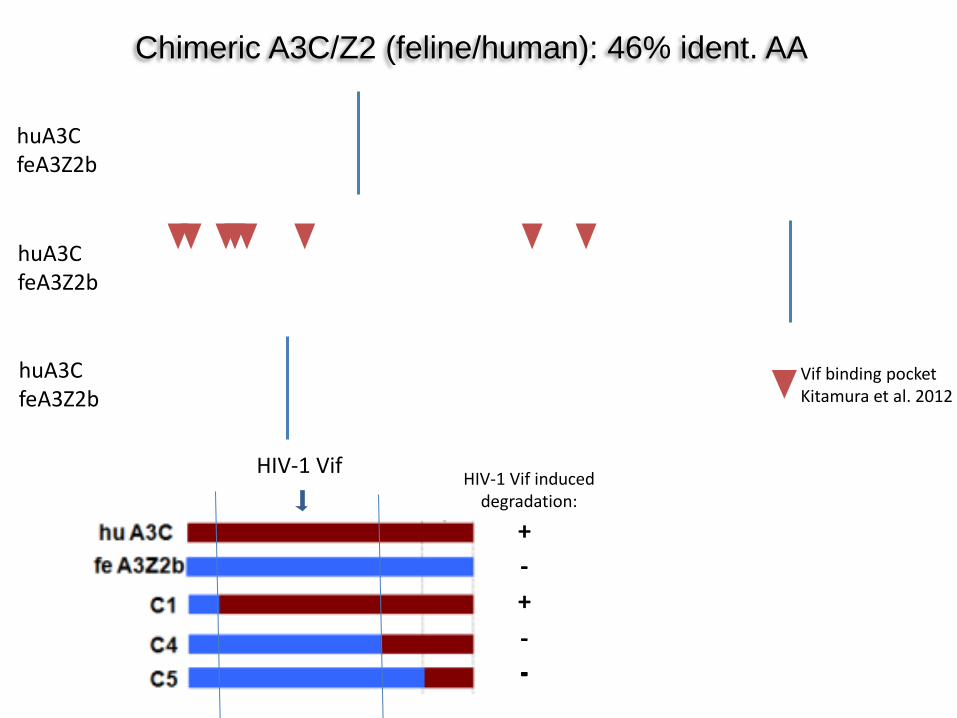
huA3C feA3Z2b
huA3C feA3Z2b
huA3C feA3Z2b
HIV-1 Vif HIV-1 Vif induced
degradation:
+
+ -
- - -
Chimeric A3C/Z2 (feline/human): 46% ident. AA
Vif binding pocket Kitamura et al. 2012
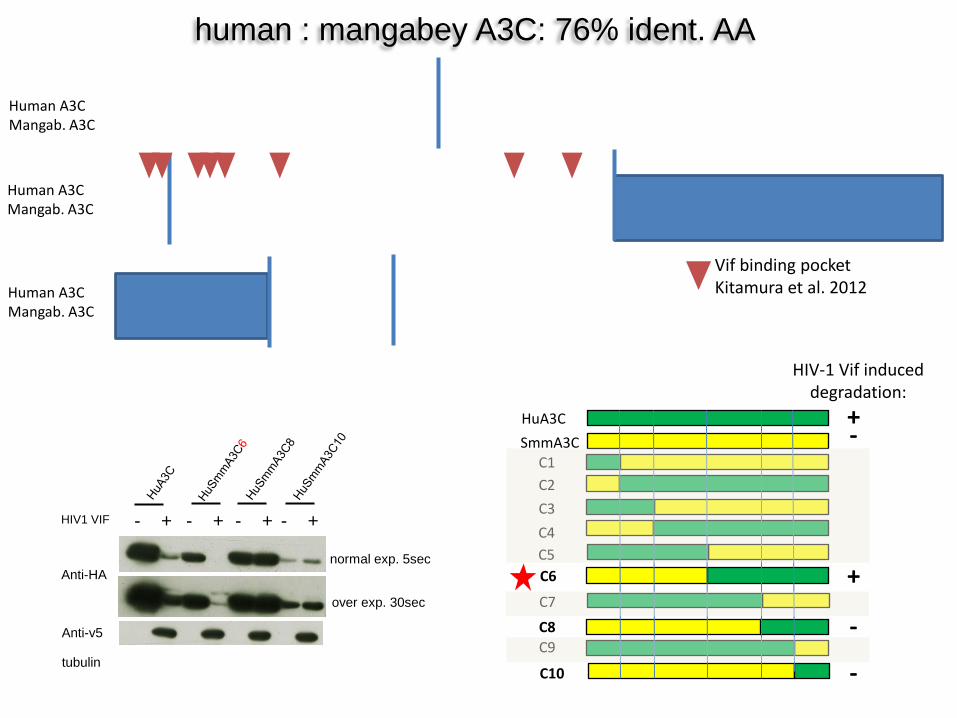
human : mangabey A3C: 76% ident. AA
Human A3C Mangab. A3C
Human A3C Mangab. A3C
Human A3C Mangab. A3C
HuA3C SmmA3C
C5
C1
C6
C2
C4 C3
C7 C8 C9
C10
Anti-HA
tubulin
Anti-v5
HIV1 VIF - + - + - + - +
over exp. 30sec
normal exp. 5sec
HIV-1 Vif induced degradation:
+
+
-
-
-
Vif binding pocket Kitamura et al. 2012
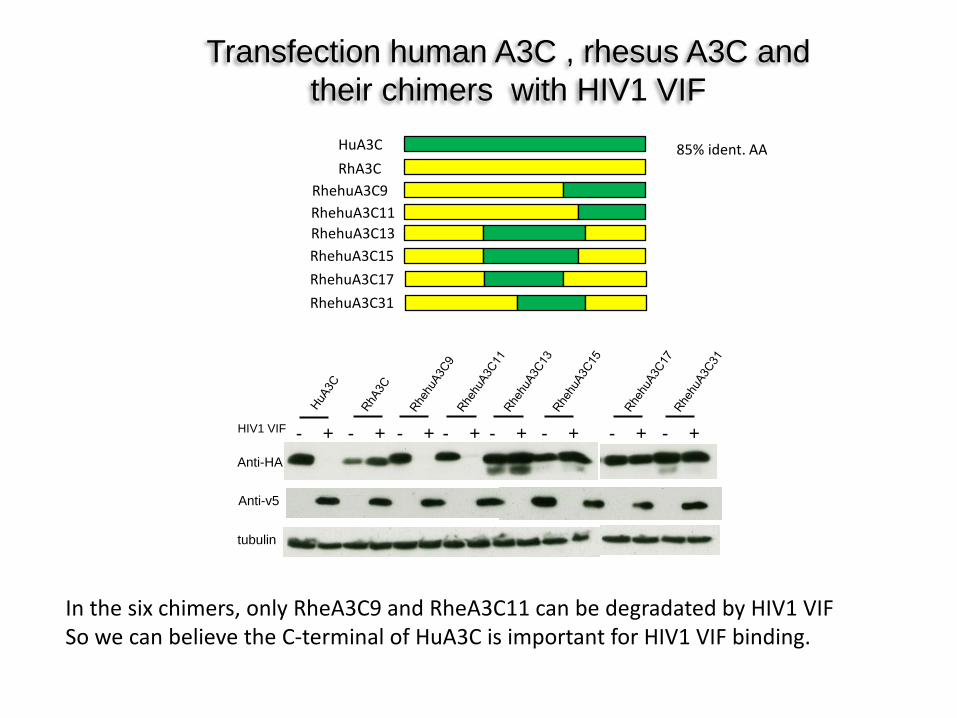
Transfection human A3C , rhesus A3C and their chimers with HIV1 VIF
Anti-HA
tubulin
Anti-v5
HIV1 VIF - + - + - + - + - + - + - + - +
HuA3C RhA3C
RhehuA3C9 RhehuA3C11 RhehuA3C13 RhehuA3C15 RhehuA3C17 RhehuA3C31
In the six chimers, only RheA3C9 and RheA3C11 can be degradated by HIV1 VIF So we can believe the C-terminal of HuA3C is important for HIV1 VIF binding.
85% ident. AA
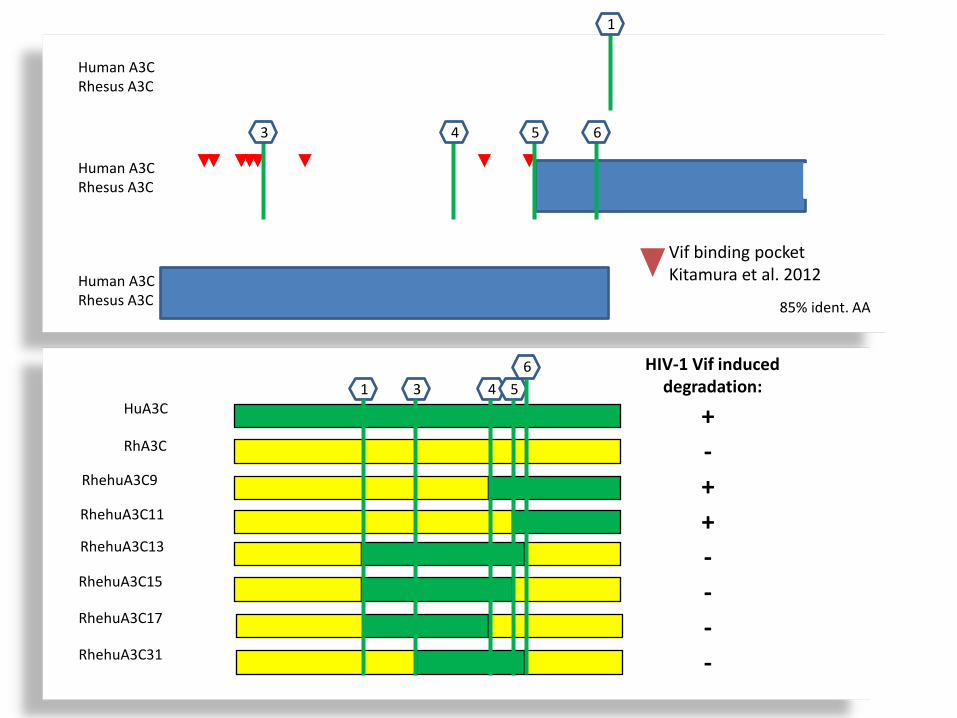
Human A3C Rhesus A3C
Human A3C Rhesus A3C
Human A3C Rhesus A3C
1
3 4 5 6
HuA3C
RhA3C
RhehuA3C9
RhehuA3C11
RhehuA3C13
RhehuA3C15
RhehuA3C17
RhehuA3C31
1 3 4 5 6 HIV-1 Vif induced
degradation:
+
+ +
-
-
- -
-
Vif binding pocket Kitamura et al. 2012
85% ident. AA
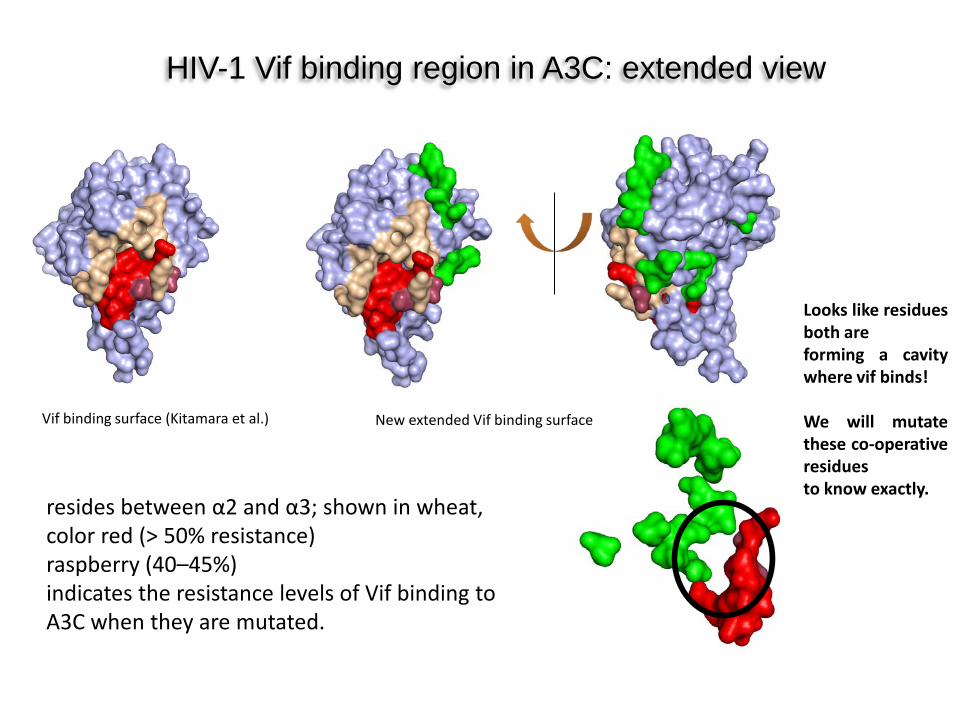
HIV-1 Vif binding region in A3C: extended view
New extended Vif binding surface Vif binding surface (Kitamara et al.)
Looks like residues both are forming a cavity where vif binds! We will mutate these co-operative residues to know exactly.
resides between α2 and α3; shown in wheat, color red (> 50% resistance) raspberry (40–45%) indicates the resistance levels of Vif binding to A3C when they are mutated.
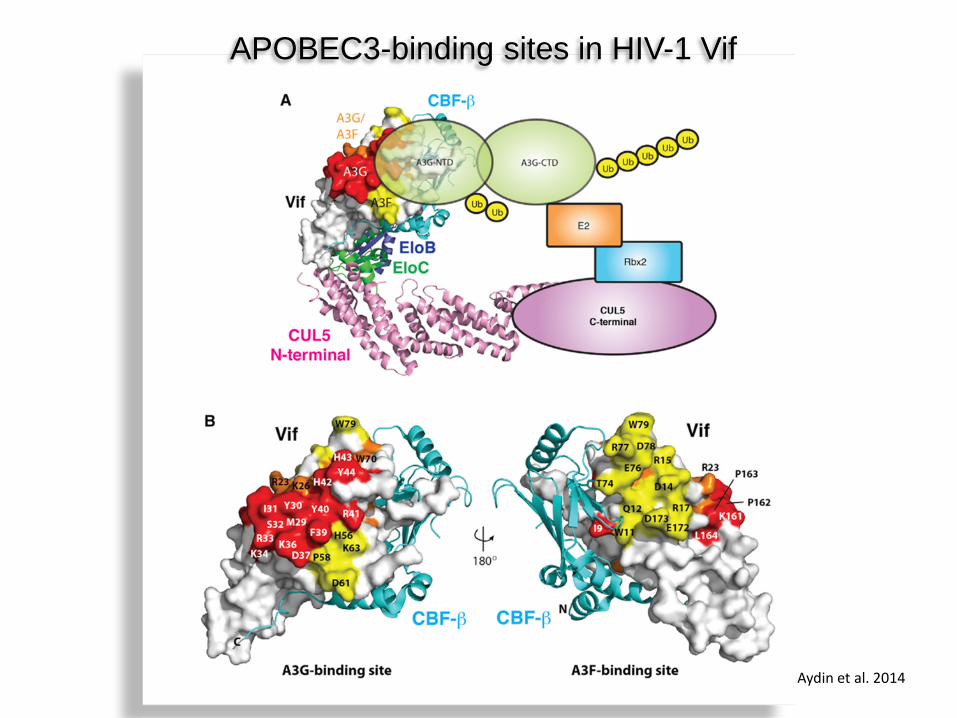
APOBEC3-binding sites in HIV-1 Vif
Aydin et al. 2014
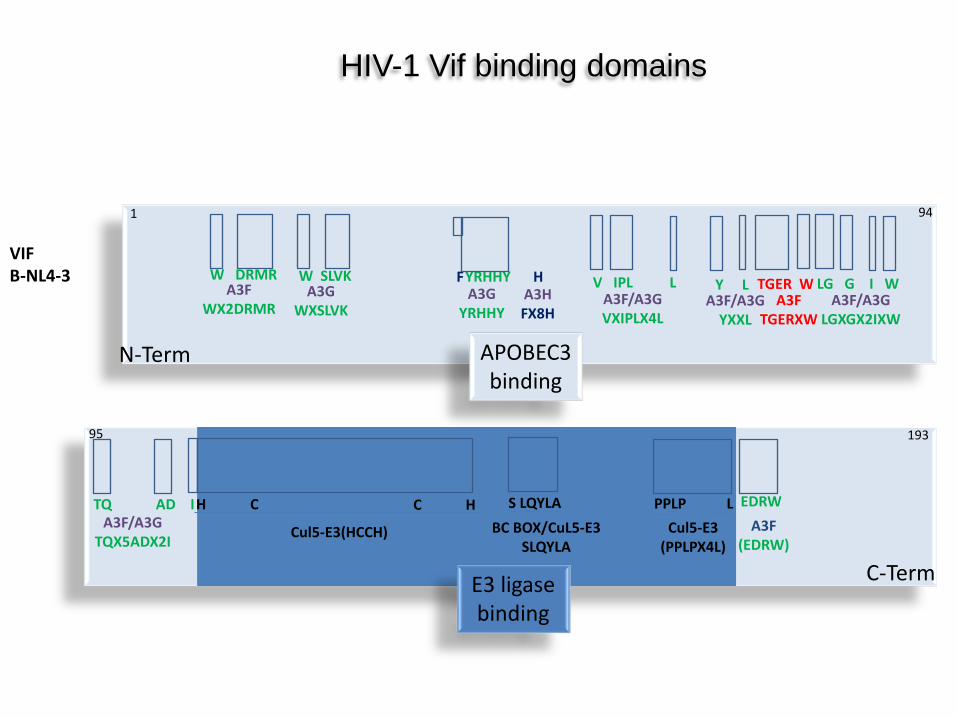
VIF B-NL4-3
A3F WX2DRMR
W DRMR A3G
WXSLVK
W SLVK A3G
YRHHY
F H A3H FX8H
YRHHY V IPL L A3F/A3G VXIPLX4L
Y L A3F/A3G
YXXL
TGER W A3F
TGERXW
LG G I W A3F/A3G
LGXGX2IXW
TQ AD I A3F/A3G
TQX5ADX2I
H C
Cul5-E3(HCCH)
C H S LQYLA BC BOX/CuL5-E3
SLQYLA
PPLP L Cul5-E3
(PPLPX4L)
EDRW A3F
(EDRW)
HIV-1 Vif binding domains
APOBEC3 binding
E3 ligase binding
N-Term
C-Term
1 94
95 193
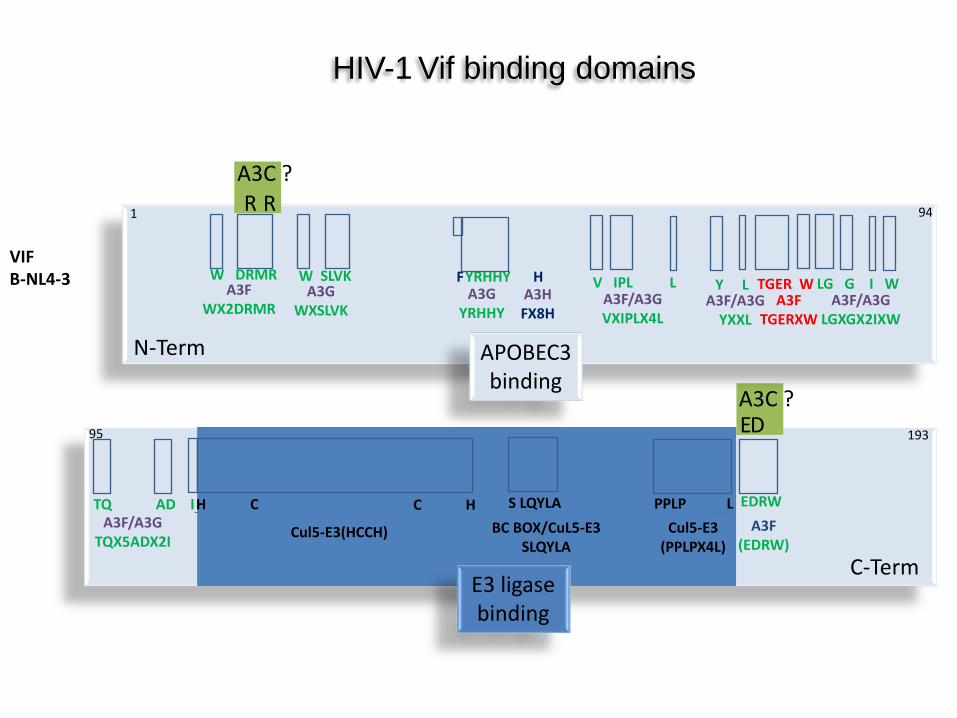
VIF B-NL4-3
A3F WX2DRMR
W DRMR A3G
WXSLVK
W SLVK A3G
YRHHY
F H A3H FX8H
YRHHY V IPL L A3F/A3G VXIPLX4L
Y L A3F/A3G
YXXL
TGER W A3F
TGERXW
LG G I W A3F/A3G
LGXGX2IXW
TQ AD I A3F/A3G
TQX5ADX2I
H C
Cul5-E3(HCCH)
C H S LQYLA BC BOX/CuL5-E3
SLQYLA
PPLP L Cul5-E3
(PPLPX4L)
EDRW A3F
(EDRW)
HIV-1 Vif binding domains
APOBEC3 binding
E3 ligase binding
N-Term
C-Term
1 94
95 193
R R A3C ?
E D A3C ?
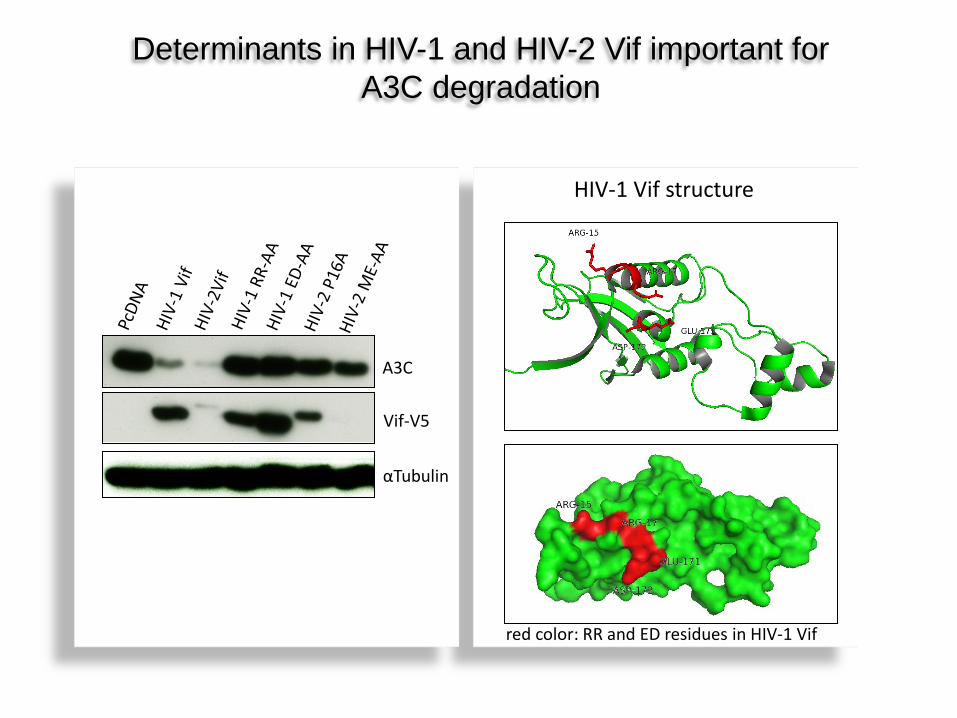
Vif-V5
A3C
αTubulin
Determinants in HIV-1 and HIV-2 Vif important for A3C degradation
red color: RR and ED residues in HIV-1 Vif
HIV-1 Vif structure
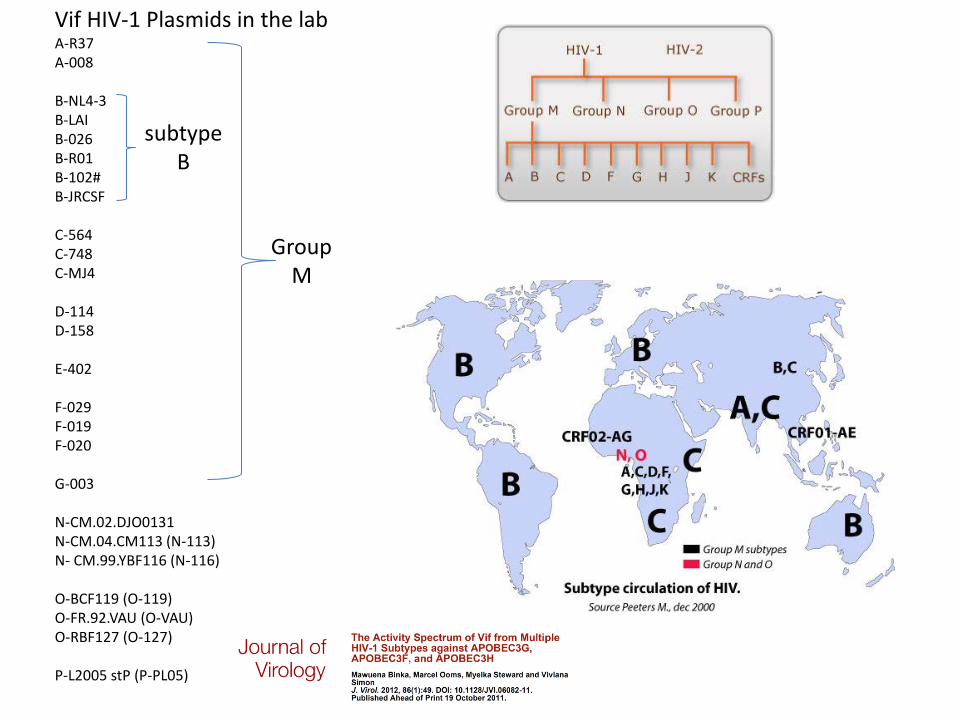
Vif HIV-1 Plasmids in the lab A-R37 A-008 B-NL4-3 B-LAI B-026 B-R01 B-102# B-JRCSF C-564 C-748 C-MJ4 D-114 D-158 E-402 F-029 F-019 F-020 G-003 N-CM.02.DJO0131 N-CM.04.CM113 (N-113) N- CM.99.YBF116 (N-116) O-BCF119 (O-119) O-FR.92.VAU (O-VAU) O-RBF127 (O-127) P-L2005 stP (P-PL05)
subtype B
Group M
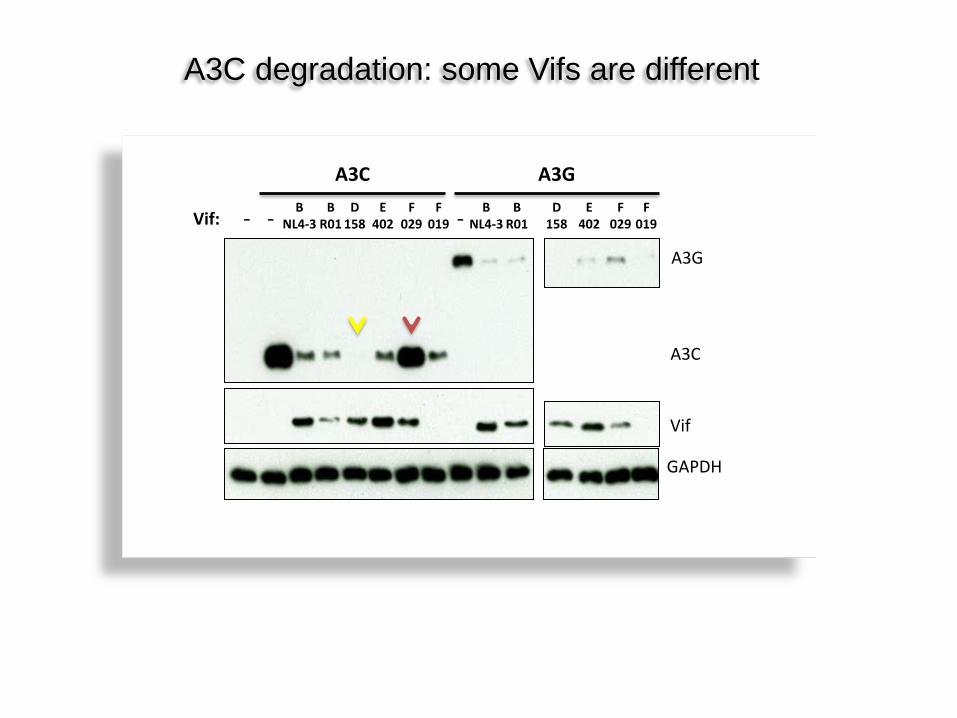
B R01
B NL4-3
D 158
E 402
F 029
F 019
A3C A3G
A3G
Vif
- - -
GAPDH
Vif: B
R01 B
NL4-3 D
158 E
402 F
029 F
019
A3C
A3C degradation: some Vifs are different
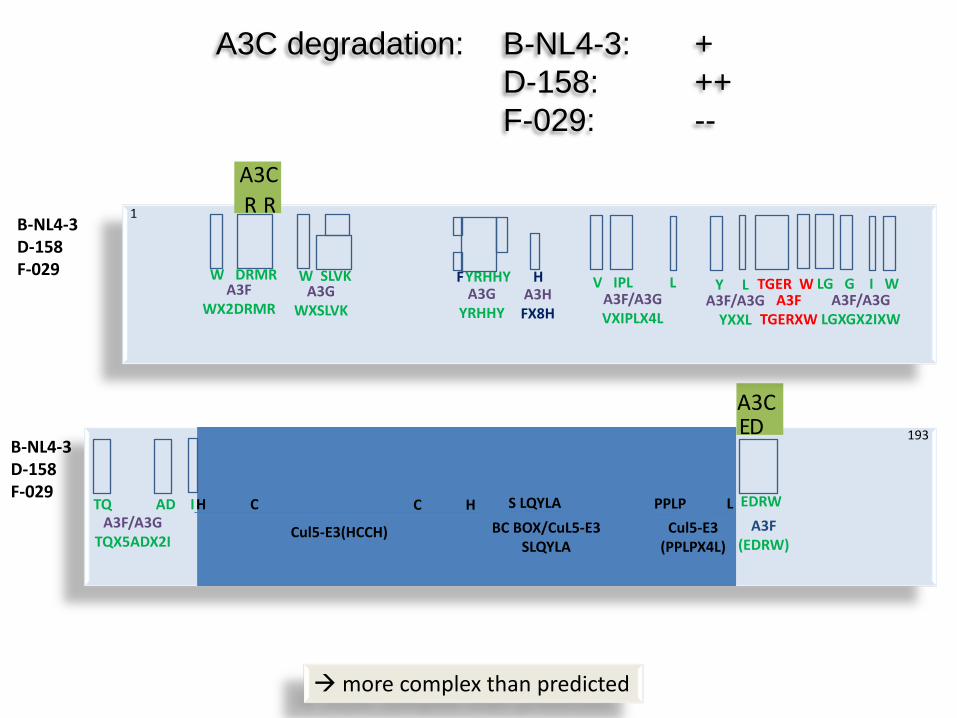
A3C degradation: B-NL4-3: + D-158: ++ F-029: --
B-NL4-3 D-158 F-029
B-NL4-3 D-158 F-029
A3F WX2DRMR
W DRMR A3G
WXSLVK
W SLVK A3G
YRHHY
F H A3H FX8H
YRHHY V IPL L A3F/A3G VXIPLX4L
Y L A3F/A3G
YXXL
TGER W A3F
TGERXW
LG G I W A3F/A3G
LGXGX2IXW
TQ AD I A3F/A3G
TQX5ADX2I
H C
Cul5-E3(HCCH)
C H S LQYLA BC BOX/CuL5-E3
SLQYLA
PPLP L Cul5-E3
(PPLPX4L)
EDRW A3F
(EDRW)
R R A3C
E D A3C
more complex than predicted
193
1
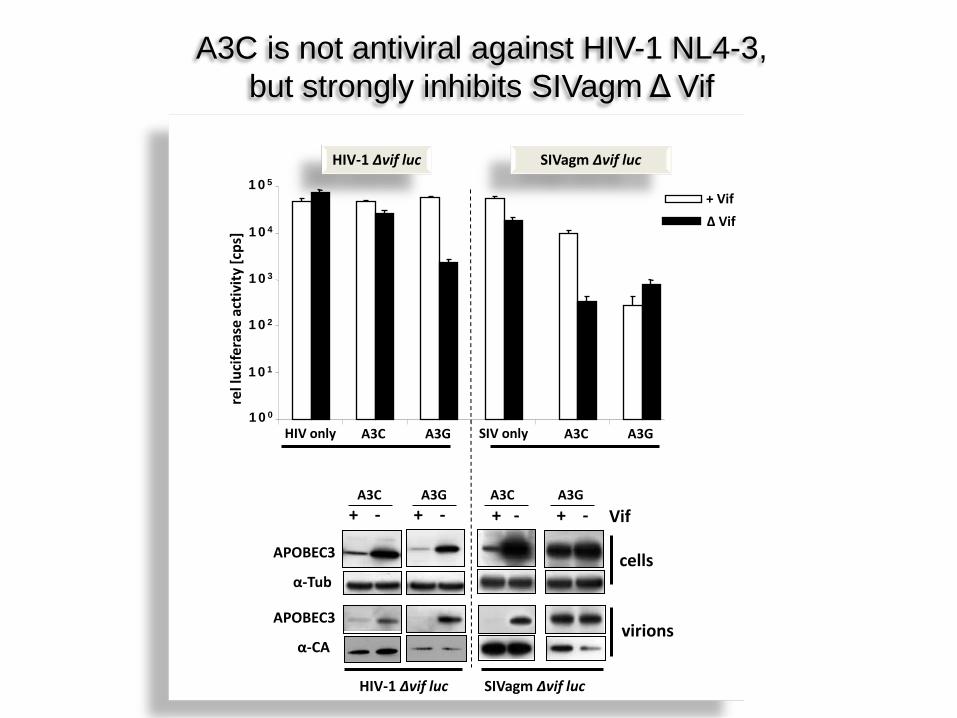
100
101
102
103
104
105
A3C A3G A3C A3G SIV only HIV only
+ Vif ∆ Vif
HIV-1 ∆vif luc SIVagm ∆vif luc
rel l
ucife
rase
act
ivity
[cps
]
+ - + -
virions
cells
+ - + - Vif
APOBEC3
α-Tub
APOBEC3
α-CA
HIV-1 ∆vif luc SIVagm ∆vif luc
A3C A3G A3C A3G
A3C is not antiviral against HIV-1 NL4-3, but strongly inhibits SIVagm Δ Vif
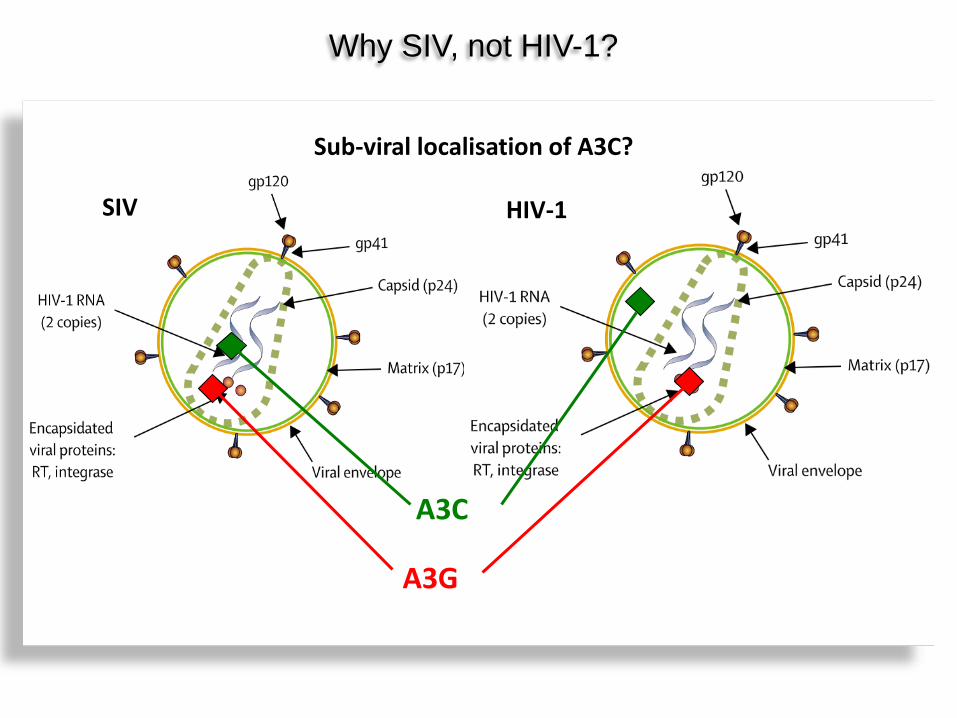
SIV HIV-1
Why SIV, not HIV-1?
A3C
A3G
Sub-viral localisation of A3C?
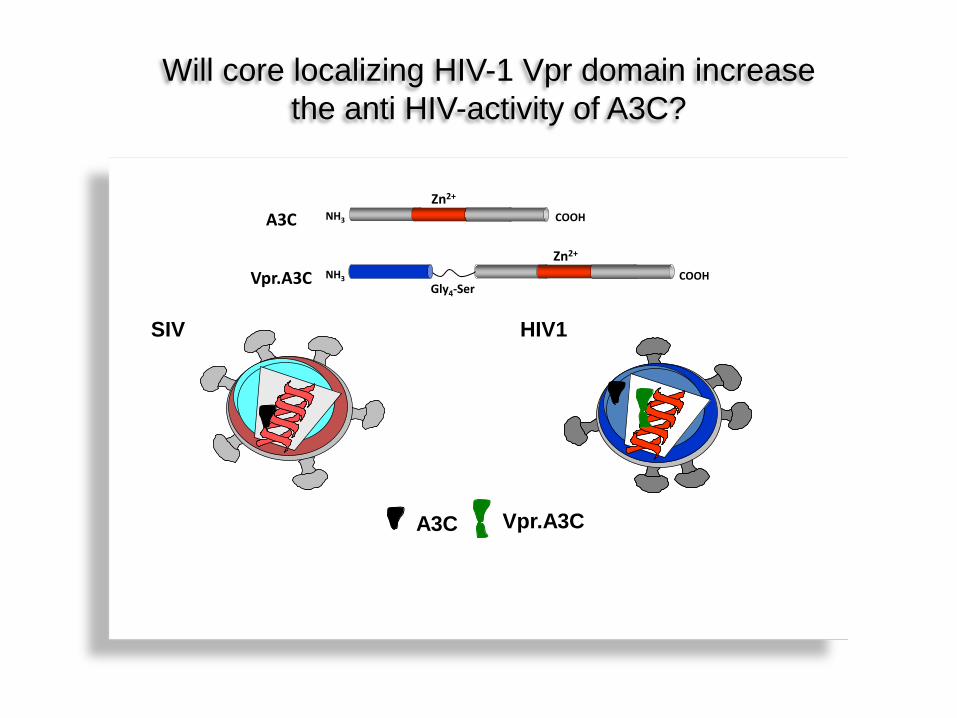
SIV HIV1
A3C Vpr.A3C
Zn2+ COOH NH3
COOH NH3 Gly4-Ser
Zn2+
A3C
Vpr.A3C
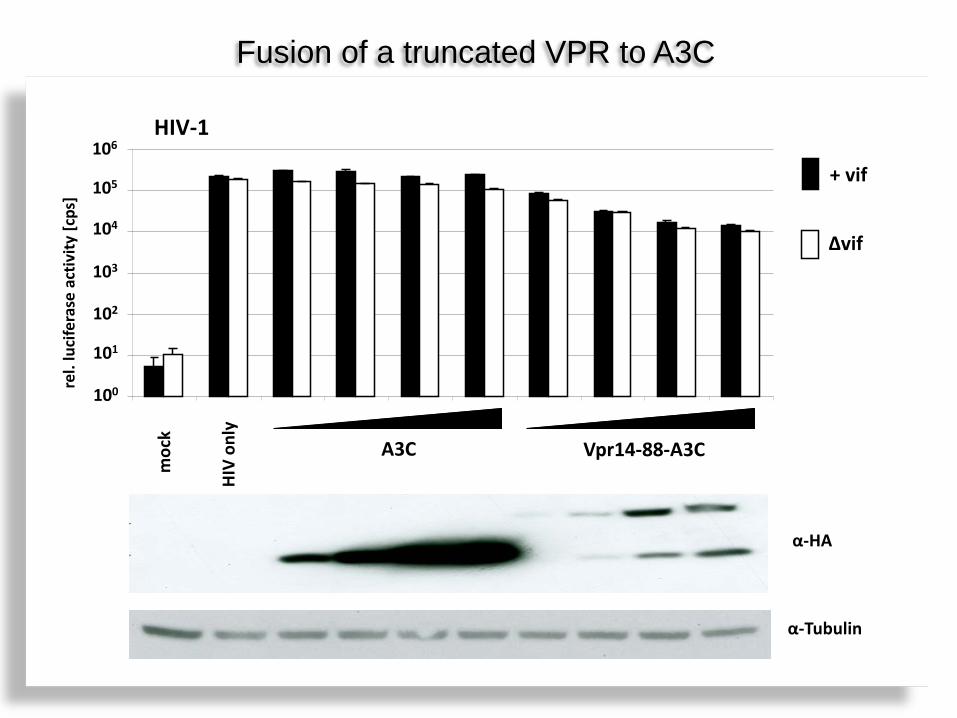
Hypothese I Will core localizing HIV-1 Vpr domain increase the anti HIV-activity of A3C?
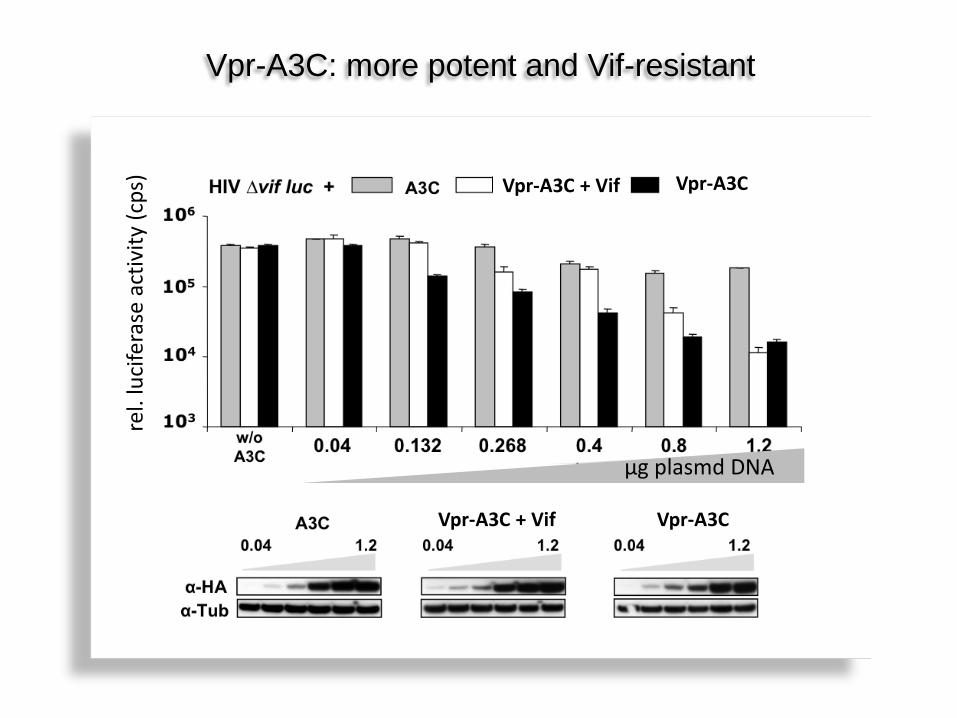
Vpr.3C inhibiert HIV Vif-unabhängig Vpr-A3C: more potent and Vif-resistant
Vpr-A3C + Vif Vpr-A3C
Vpr-A3C Vpr-A3C + Vif
µg plasmd DNA
rel.
luci
fera
se a
ctiv
ity (c
ps)
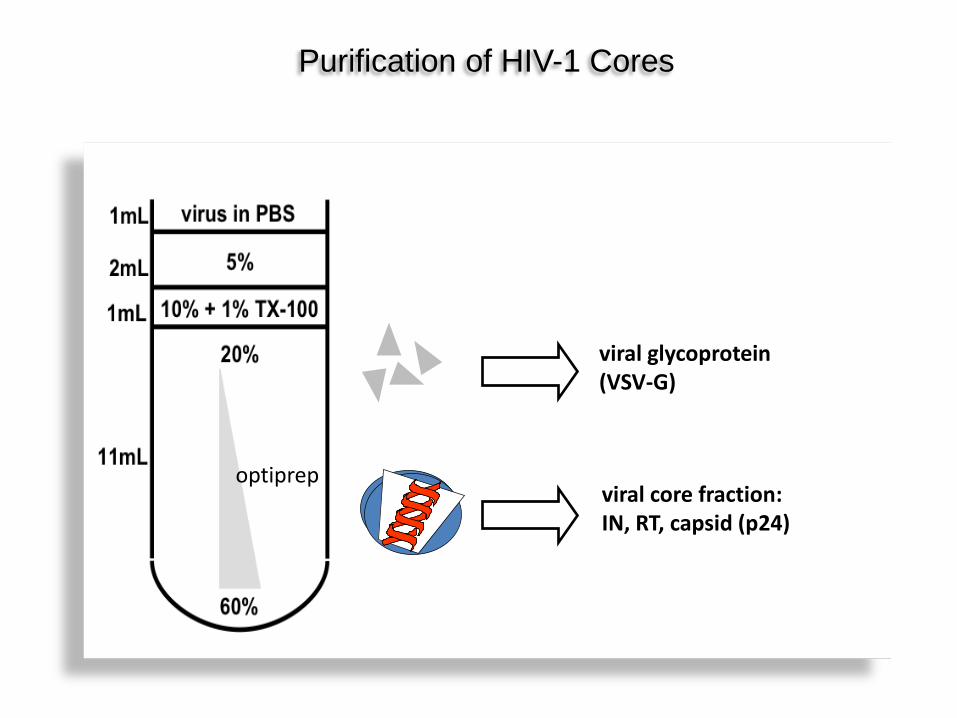
viral glycoprotein (VSV-G)
viral core fraction: IN, RT, capsid (p24)
Purification of HIV-1 Cores
optiprep
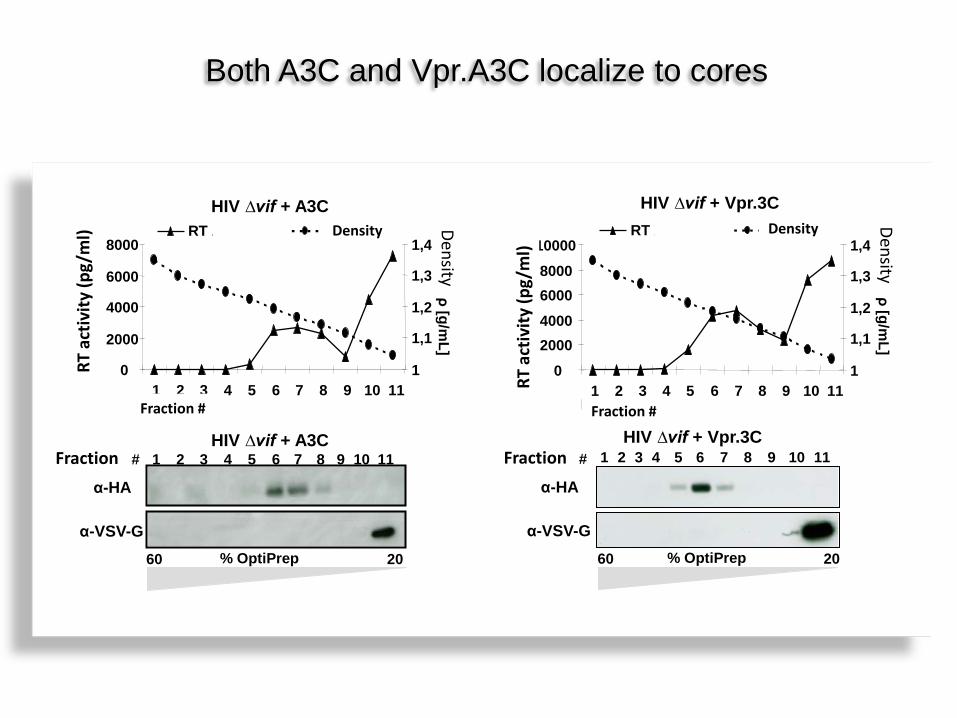
Both A3C and Vpr.A3C localize to cores
α-VSV-G
α-HA
1 2 3 4 5 6 7 8 9 10 11 Fraktion #
% OptiPrep
HIV ∆vif + A3C
60 20
HIV ∆vif + A3C
0
2000
4000
6000
8000
1 2 3 4 5 6 7 8 9 10 11 1
1,1
1,2
1,3
1,4 RT Aktivität Dichte
RT
Aktiv
ität [
pg/m
L] D
ichte ρ [g/mL]
Fraktion #
1 2 3 4 5 6 7 8 9 10 11
α-VSV-G
α-HA
Fraktion # HIV ∆vif + Vpr.3C
% OptiPrep 60 20
HIV ∆vif + Vpr.3C
0 2000 4000 6000 8000
10000
1 2 3 4 5 6 7 8 9 10 11 1
1,1
1,2
1,3
1,4 RT Aktivität Dichte
RT
Aktiv
ität [
pg/m
L] D
ichte ρ [g/mL]
Fraktion #
Density
Density
Fraction Fraction
RT a
ctiv
ity (p
g/m
l)
RT a
ctiv
ity (p
g/m
l)
Density Density
Fraction # Fraction #
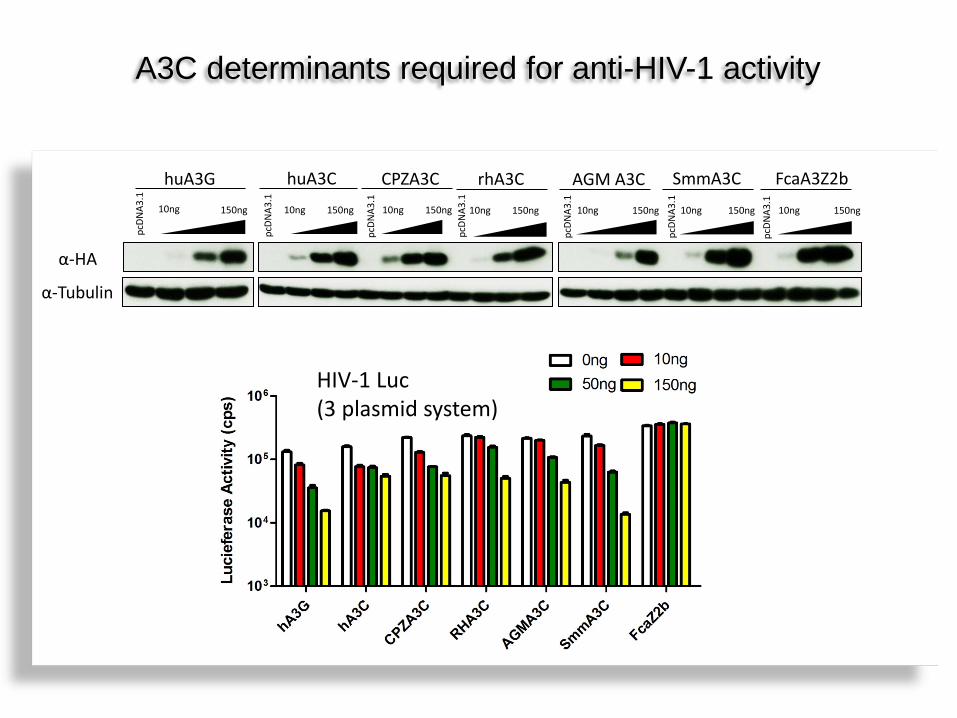
α-Tubulin
α-HA
huA3G huA3C CPZA3C 10ng 150ng
pcDN
A3.1
10ng 150ng
pcDN
A3.1
10ng 150ng
pcDN
A3.1
rhA3C AGM A3C 10ng 150ng
pcDN
A3.1
10ng 150ng
pcDN
A3.1
SmmA3C 10ng 150ng
pcDN
A3.1
FcaA3Z2b 10ng 150ng
pcDN
A3.1
HIV-1 Luc (3 plasmid system)
A3C determinants required for anti-HIV-1 activity
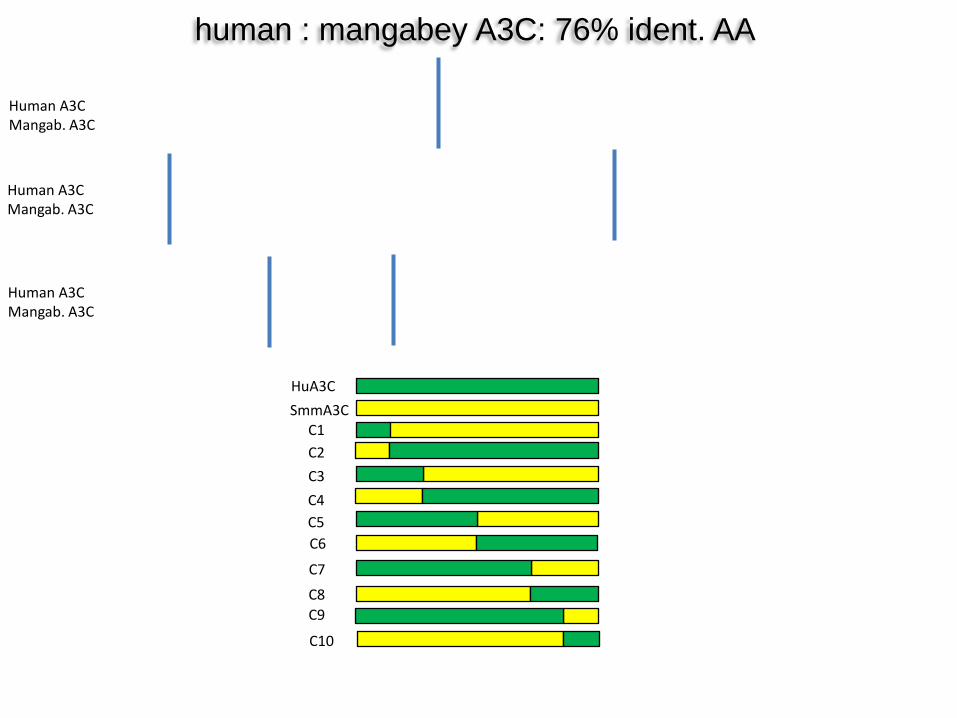
HuA3C SmmA3C
C5
C1
C6
C2
C4 C3
C7 C8 C9
C10
human : mangabey A3C: 76% ident. AA
Human A3C Mangab. A3C
Human A3C Mangab. A3C
Human A3C Mangab. A3C
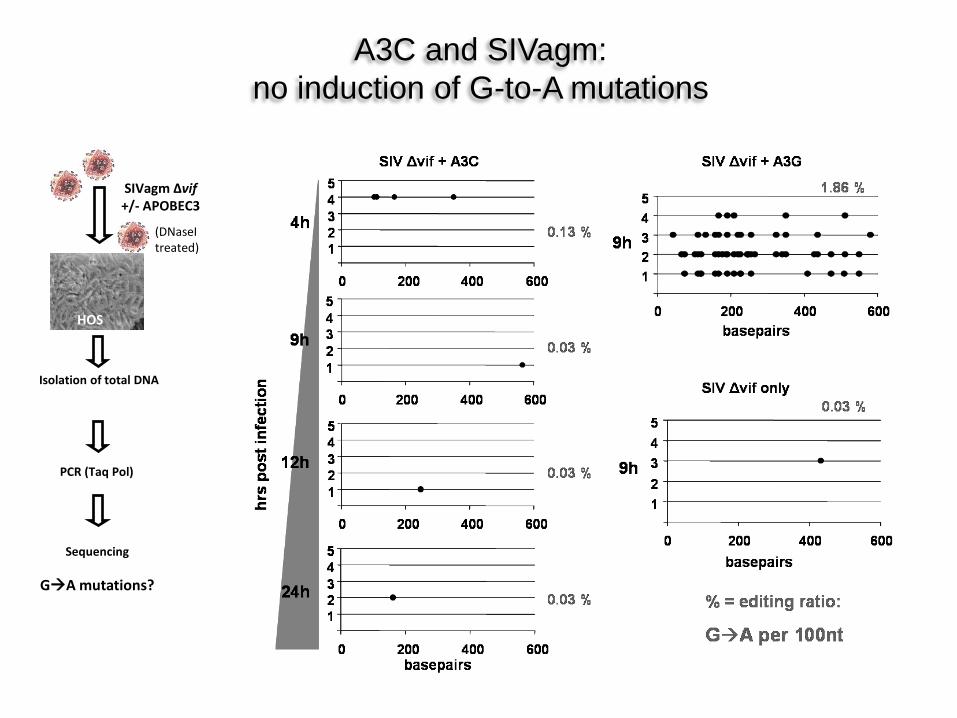
HOS
PCR (Taq Pol)
Sequencing
GA mutations?
SIVagm Δvif +/- APOBEC3
Isolation of total DNA
A3C and SIVagm: no induction of G-to-A mutations
(DNaseI treated)
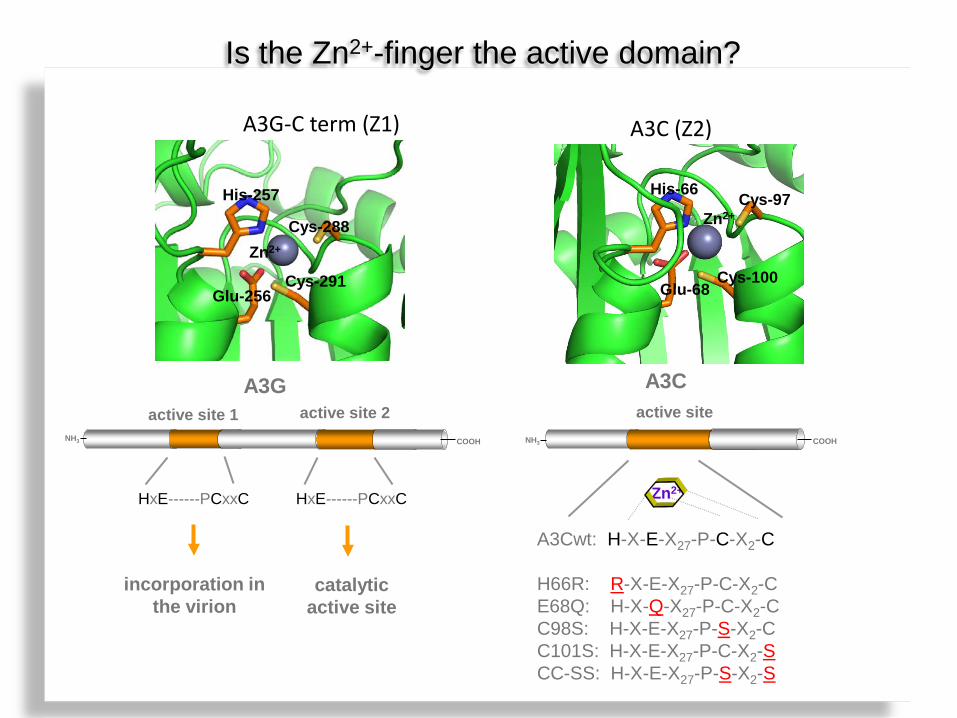
Is the Zn2+-finger the active domain?
His-66 Cys-97
Glu-68 Cys-100
A3C (Z2)
Zn2+
?
A3G-C term (Z1)
His-257
Cys-288
Cys-291 Glu-256
Zn2+
active site 1 active site 2
HxE------PCxxC
HxE------PCxxC
A3G
incorporation in the virion
catalytic active site
NH3
A3Cwt: H-X-E-X27-P-C-X2-C H66R: R-X-E-X27-P-C-X2-C E68Q: H-X-Q-X27-P-C-X2-C C98S: H-X-E-X27-P-S-X2-C C101S: H-X-E-X27-P-C-X2-S CC-SS: H-X-E-X27-P-S-X2-S
active site
Zn2+
A3C
COOH NH3 COOH
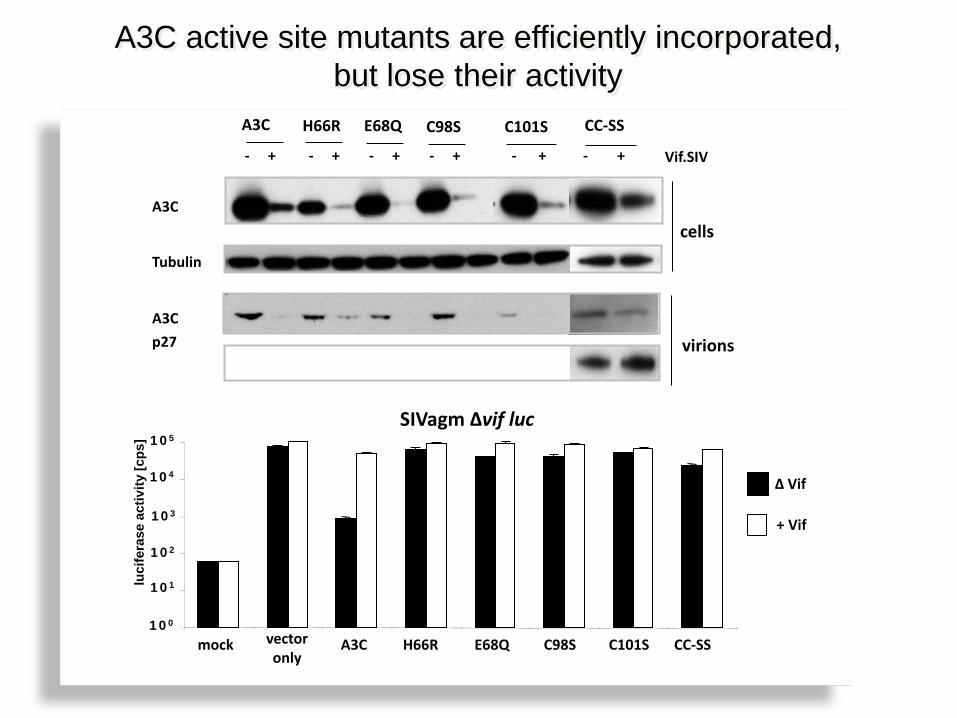
- + - + - + - + - + - +
A3C H66R E68Q C98S C101S
Vif.SIV
virions
cells
CC-SS
A3C
Tubulin
A3C
A3C active site mutants are efficiently incorporated, but lose their activity
p27
mock vector only
A3C H66R E68Q C98S C101S CC-SS 100
101
102
103
104
105
luci
fera
se a
ctiv
ity [c
ps]
+ Vif
Δ Vif
SIVagm ∆vif luc
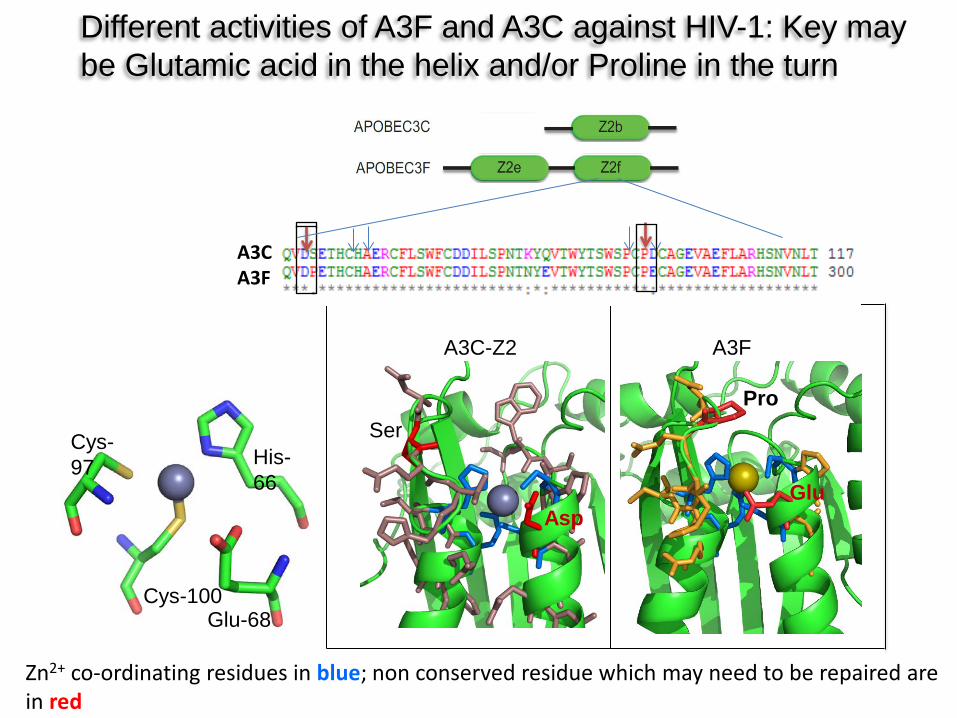
Zn2+ co-ordinating residues in blue; non conserved residue which may need to be repaired are in red
His-66
Cys-100
Cys-97
Glu-68
A3C-Z2 A3F
Ser
Asp Glu
Pro
Different activities of A3F and A3C against HIV-1: Key may be Glutamic acid in the helix and/or Proline in the turn
A3C A3F
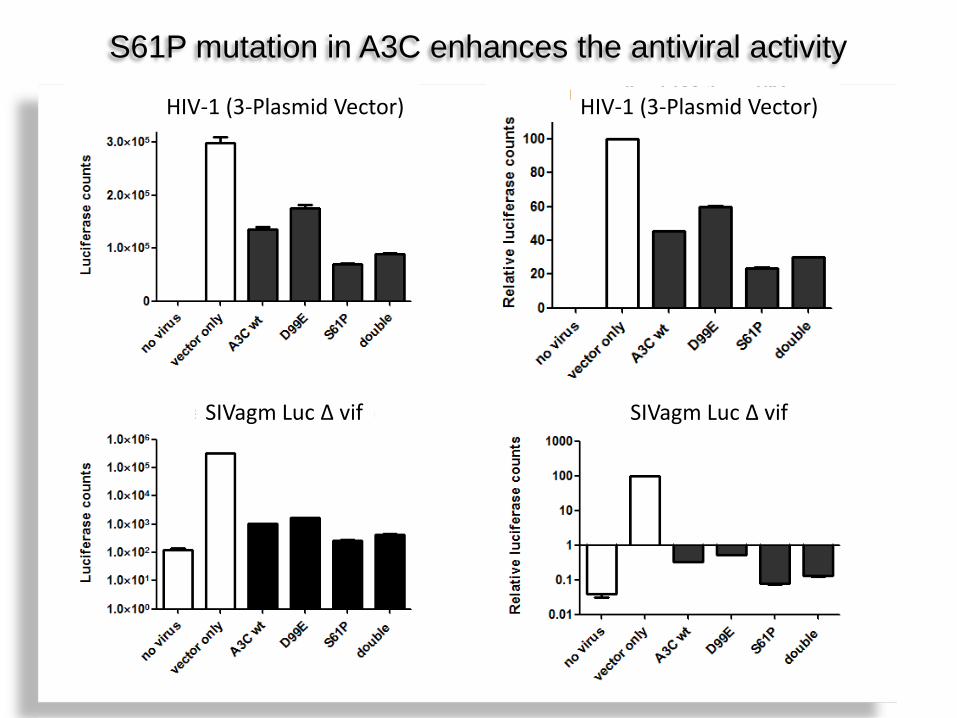
HIV-1 (3-Plasmid Vector) HIV-1 (3-Plasmid Vector)
SIVagm Luc Δ vif SIVagm Luc Δ vif
S61P mutation in A3C enhances the antiviral activity
CCCA and CCCG are specific for A3G, A3C may require others
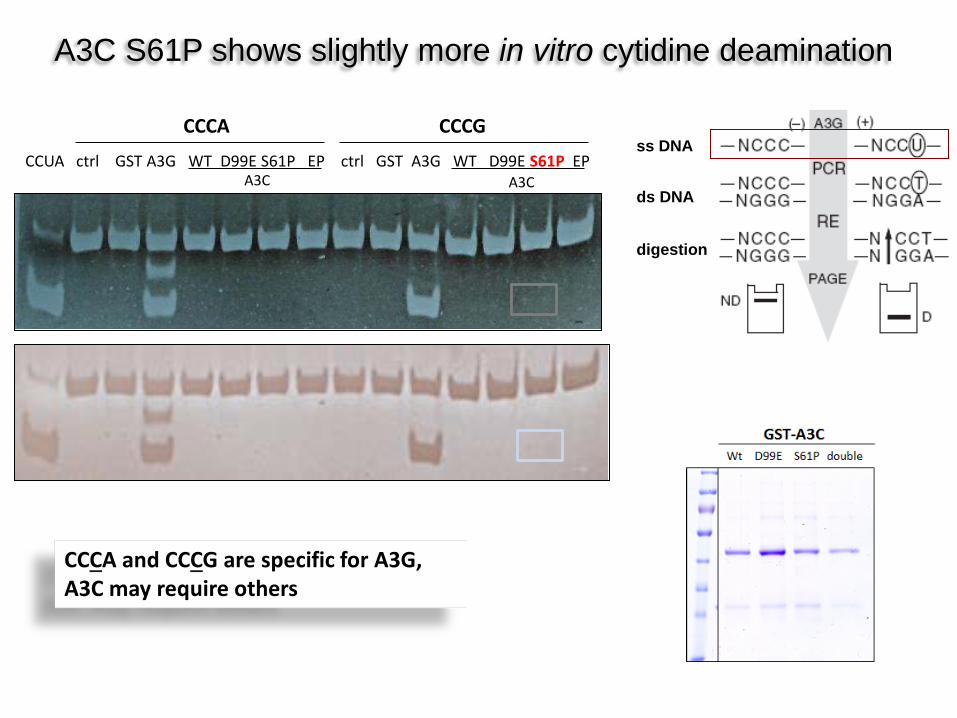
A3C S61P shows slightly more in vitro cytidine deamination
CCUA ctrl GST A3G WT D99E S61P EP ctrl GST A3G WT D99E S61P EP
CCCA CCCG
A3C A3C
ss DNA
ds DNA
digestion
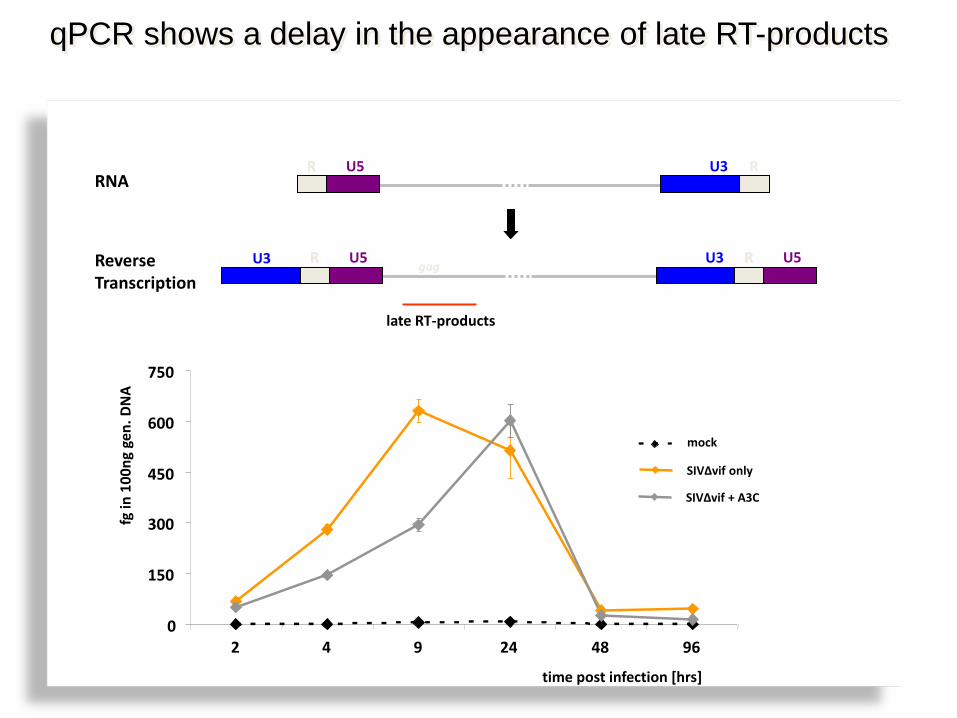
qPCR shows a delay in the appearance of late RT-products
0
150
300
450
600
750
2 4 9 24 48 96
time post infection [hrs]
mock
SIVΔvif only
SIVΔvif + A3C
fg in
100
ng g
en. D
NA
R U5 U3 RNA
Reverse Transcription
R R U5 U3 gag
late RT-products
R R
R R U5 U3
Die Integrase von HIV-1 als weiterer Antagonist von A3C?
moc
k
SIV-
IN
HIV
-IN
Lysat
α-His
moc
k
SIV-
IN
HIV
-IN
Ni-NTA
α-HA
moc
k
SIV-
IN
HIV
-IN
Lysa
t
bound to
Lysa
t
Lysa
t
boun
d
boun
d
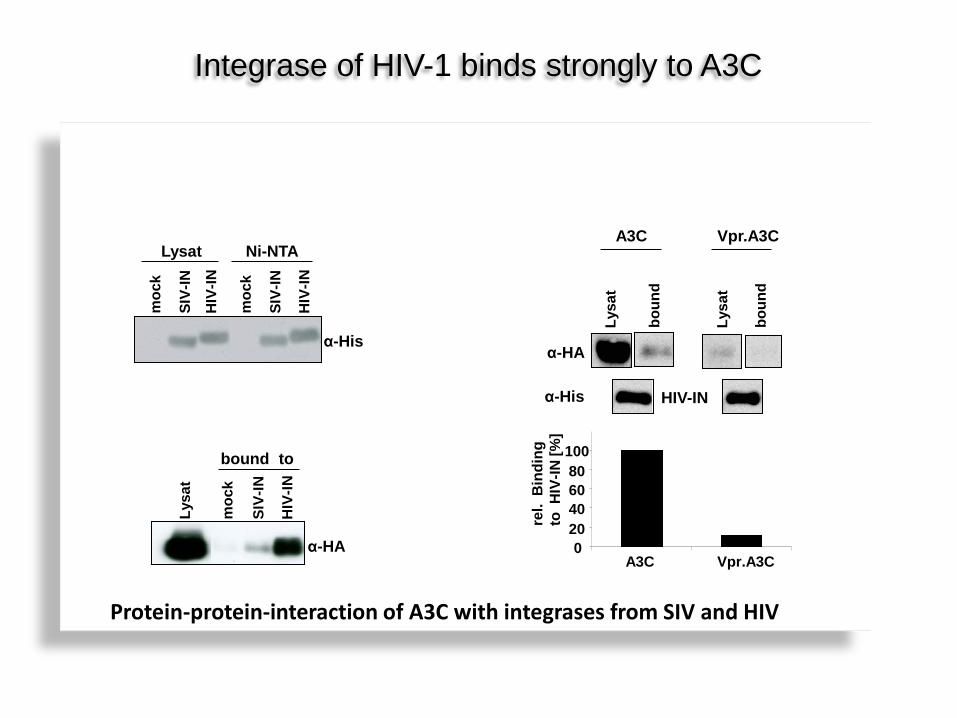
A3C Vpr.A3C
α-HA
α-His HIV-IN
0 20 40 60 80 100
A3C Vpr.A3C re
l. B
indi
ng
to H
IV-IN
[%]
Integrase of HIV-1 binds strongly to A3C
Protein-protein-interaction of A3C with integrases from SIV and HIV
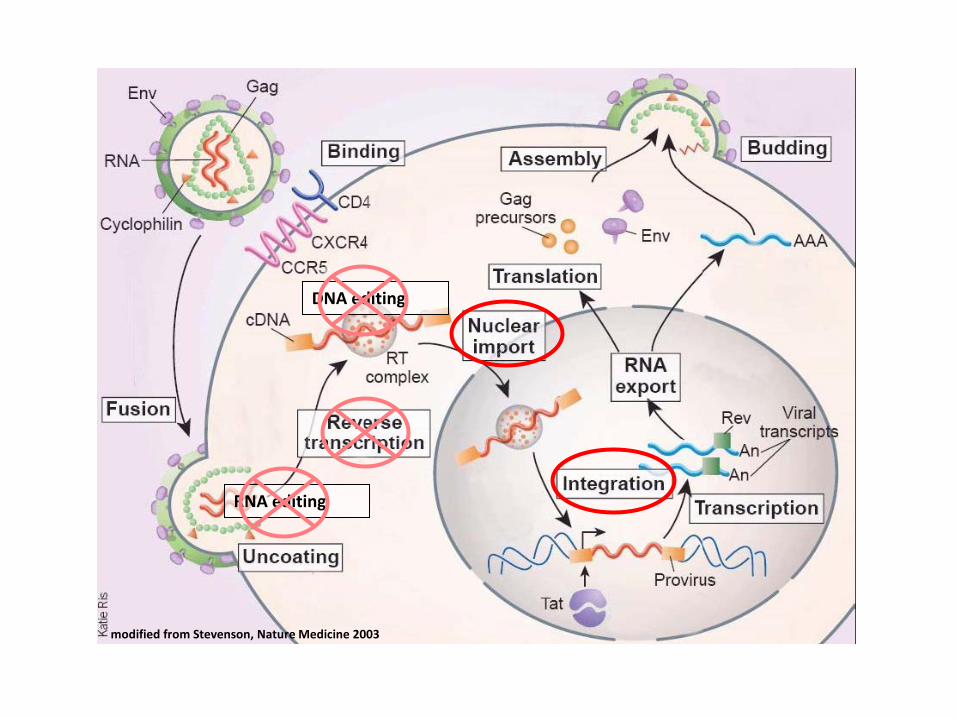
DNA editing
RNA editing
modified from Stevenson, Nature Medicine 2003

Important open questions: - Why is human A3C not antiviral against HIV-1 (at least lab strains)?
- Why is sooty mangabey A3C antiviral against HIV-1?
- What is the antiviral activity of human A3C against SIV?
- Does Vif /A3 binding cause structural changes in either protein (Vif/A3 interface)?
Impacts oligomerization?

Klinik für Gastroenterologie, Hepatologie und Infektiologie Leber- und Infektionszentrum W. Hirsch-Institut für Tropenmedizin, ET Direktor der Klinik Univ.-Prof. Dr. D. Häussinger
Virus
Intrinsic Immune System
Cancer
Host-Virus Arms Races
Experimental results shown from:
Qinyong Gu Henning Hofmann Ananda Ayyappan Jaguva Vasudevan Zeli Zhang Jörg Zielonka
HIV
only
moc
k
A3C Vpr14-88-A3C
+ vif
Δvif
rel.
luci
fera
se a
ctiv
ity [c
ps]
Fusion of a truncated VPR to A3C
100
101
102
103
104
105
106
α-HA
α-Tubulin
HIV-1
Top Related