
WA27038-Bone Marrow Niches HSC Fates€¦ · Adipocytes Perilipin+ Caveolin+ Haematopoietic cells...
1
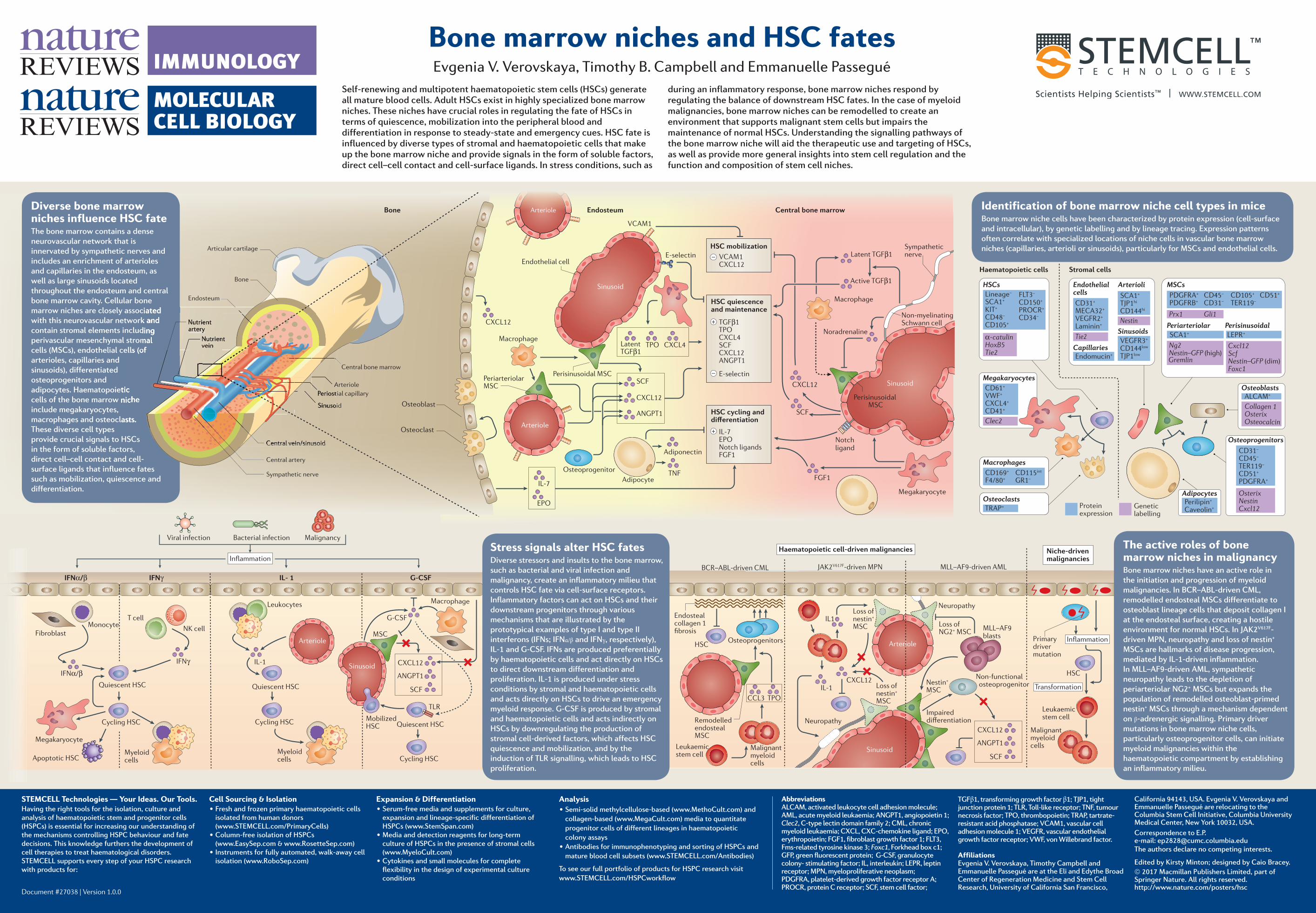
Adipocytes Perilipin + Caveolin + Stromal cells Haematopoietic cells LEPR + SCA1 + Periarteriolar Perisinusoidal CD45 – CD31 – PDGFRA + PDGFRB + CD105 + CD51 + TER119 – Prx1 Gli1 Ng2 Nestin–GFP (high) Cxcl12 Scf Nestin–GFP (dim) Foxc1 MSCs Gremlin Lineage – SCA1 + KIT + CD48 – CD105 + FLT3 – CD150 + PROCR + CD34 – α-catulin HoxB5 Tie2 HSCs Protein expression Genetic labelling Osteoprogenitors Osterix Nestin Cxcl12 CD31 – CD45 – TER119 – CD51 + PDGFRA + Sinusoids VEGFR3 + CD144 low TJP1 low CD31 + MECA32 + VEGFR2 + Laminin + Endothelial cells Capillaries Arterioli SCA1 + TJP1 hi CD144 hi Tie2 Nestin Osteoblasts Collagen 1 Osterix Osteocalcin ALCAM + SCF IL-1 G-CSF CXCL12 ANGPT1 Monocyte Quiescent HSC Cycling HSC Quiescent HSC Cycling HSC Quiescent HSC Cycling HSC Mobilized HSC T cell NK cell Myeloid cells Myeloid cells TLR IFNα/β IFNγ IFNγ IL- 1 G-CSF Viral infection Bacterial infection Malignancy Inflammation Osteoclasts TRAP + CD169 + F4/80 + CD115 int GR1 – Macrophages Macrophage Leukocytes Fibroblast Apoptotic HSC Megakaryocyte IFNα/β CD61 + VWF + CXCL4 + CD41 + Clec2 Megakaryocytes Notch ligand Sinusoid CXCL12 HSC mobilization VCAM1 CXCL12 – TGFβ1 TPO CXCL4 SCF CXCL12 ANGPT1 – + HSC quiescence and maintenance E-selectin + HSC cycling and differentiation IL-7 EPO Notch ligands FGF1 SCF Latent TGFβ1 Active TGFβ1 Noradrenaline FGF1 Macrophage Endothelial cell Non-myelinating Schwann cell Sympathetic nerve Sinusoid Sinusoid Perisinusoidal MSC Bone Central bone marrow Arteriole Sinusoid MSC Osteoclast Osteoblast Sinusoid Periarteriolar MSC Perisinusoidal MSC CXCL12 IL-7 EPO TNF ANGPT1 CXCL12 SCF Latent TGFβ1 TPO CXCL4 Adiponectin Osteoprogenitor Adipocyte Macrophage Arteriole VCAM1 E-selectin Endosteum Arteriole Endothelial cell Megakaryocyte IL-1 IL1 CCL3 TPO CXCL12 Malignant myeloid cells Malignant myeloid cells Loss of NG2 + MSC Non-functional osteoprogenitor Neuropathy Neuropathy Inflammation Transformation HSC Nestin + MSC Loss of nestin + MSC Loss of nestin + MSC Impaired differentiation MLL–AF9 blasts Niche-driven malignancies Haematopoietic cell-driven malignancies MLL–AF9-driven AML JAK2 V617F -driven MPN Primary driver mutation Sinusoid Arteriole Osteoprogenitors Endosteal collagen 1 fibrosis Remodelled endosteal MSC Leukaemic stem cell BCR–ABL-driven CML HSC Leukaemic stem cell SCF CXCL12 ANGPT1 Endomucin + Arteriole Bone marrow niches and HSC fates Evgenia V. Verovskaya, Timothy B. Campbell and Emmanuelle Passegué Self-renewing and multipotent haematopoietic stem cells (HSCs) generate all mature blood cells. Adult HSCs exist in highly specialized bone marrow niches. These niches have crucial roles in regulating the fate of HSCs in terms of quiescence, mobilization into the peripheral blood and differentiation in response to steady-state and emergency cues. HSC fate is influenced by diverse types of stromal and haematopoietic cells that make up the bone marrow niche and provide signals in the form of soluble factors, direct cell–cell contact and cell-surface ligands. In stress conditions, such as during an inflammatory response, bone marrow niches respond by regulating the balance of downstream HSC fates. In the case of myeloid malignancies, bone marrow niches can be remodelled to create an environment that supports malignant stem cells but impairs the maintenance of normal HSCs. Understanding the signalling pathways of the bone marrow niche will aid the therapeutic use and targeting of HSCs, as well as provide more general insights into stem cell regulation and the function and composition of stem cell niches. STEMCELL Technologies — Your Ideas. Our Tools. Having the right tools for the isolation, culture and analysis of haematopoietic stem and progenitor cells (HSPCs) is essential for increasing our understanding of the mechanisms controlling HSPC behaviour and fate decisions. This knowledge furthers the development of cell therapies to treat haematological disorders. STEMCELL supports every step of your HSPC research with products for: Document #27038 | Version 1.0.0 Expansion & Differentiation • Serum-free media and supplements for culture, expansion and lineage-specific differentiation of HSPCs (www.StemSpan.com) • Media and detection reagents for long-term culture of HSPCs in the presence of stromal cells (www.MyeloCult.com) • Cytokines and small molecules for complete flexibility in the design of experimental culture conditions Analysis • Semi-solid methylcellulose-based (www.MethoCult.com) and collagen-based (www.MegaCult.com) media to quantitate progenitor cells of different lineages in haematopoietic colony assays • Antibodies for immunophenotyping and sorting of HSPCs and mature blood cell subsets (www.STEMCELL.com/Antibodies) To see our full portfolio of products for HSPC research visit www.STEMCELL.com/HSPCworkflow Cell Sourcing & Isolation • Fresh and frozen primary haematopoietic cells isolated from human donors (www.STEMCELL.com/PrimaryCells) • Column-free isolation of HSPCs (www.EasySep.com & www.RosetteSep.com) • Instruments for fully automated, walk-away cell isolation (www.RoboSep.com) Abbreviations ALCAM, activated leukocyte cell adhesion molecule; AML, acute myeloid leukaemia; ANGPT1, angiopoietin 1; Clec2, C-type lectin domain family 2; CML, chronic myeloid leukaemia; CXCL, CXC-chemokine ligand; EPO, erythropoietin; FGF1, fibroblast growth factor 1; FLT3, Fms-related tyrosine kinase 3; Foxc1, Forkhead box c1; GFP, green fluorescent protein; G-CSF, granulocyte colony- stimulating factor; IL, interleukin; LEPR, leptin receptor; MPN, myeloproliferative neoplasm; PDGFRA, platelet-derived growth factor receptor A; PROCR, protein C receptor; SCF, stem cell factor; TGFβ1, transforming growth factor β1; TJP1, tight junction protein 1; TLR, Toll-like receptor; TNF, tumour necrosis factor; TPO, thrombopoietin; TRAP, tartrate- resistant acid phosphatase; VCAM1, vascular cell adhesion molecule 1; VEGFR, vascular endothelial growth factor receptor; VWF, von Willebrand factor. Affiliations Evgenia V. Verovskaya, Timothy Campbell and Emmanuelle Passegué are at the Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell Research, University of California San Francisco, California 94143, USA. Evgenia V. Verovskaya and Emmanuelle Passegué are relocating to the Columbia Stem Cell Initiative, Columbia University Medical Center, New York 10032, USA. Correspondence to E.P. e-mail: [email protected] The authors declare no competing interests. Edited by Kirsty Minton; designed by Caio Bracey. © 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved. http://www.nature.com/posters/hsc Stress signals alter HSC fates Diverse stressors and insults to the bone marrow, such as bacterial and viral infection and malignancy, create an inflammatory milieu that controls HSC fate via cell-surface receptors. Inflammatory factors can act on HSCs and their downstream progenitors through various mechanisms that are illustrated by the prototypical examples of type I and type II interferons (IFNs; IFNα/β and IFNγ, respectively), IL-1 and G-CSF. IFNs are produced preferentially by haematopoietic cells and act directly on HSCs to direct downstream differentiation and proliferation. IL-1 is produced under stress conditions by stromal and haematopoietic cells and acts directly on HSCs to drive an emergency myeloid response. G-CSF is produced by stromal and haematopoietic cells and acts indirectly on HSCs by downregulating the production of stromal cell-derived factors, which affects HSC quiescence and mobilization, and by the induction of TLR signalling, which leads to HSC proliferation. The active roles of bone marrow niches in malignancy Bone marrow niches have an active role in the initiation and progression of myeloid malignancies. In BCR–ABL-driven CML, remodelled endosteal MSCs differentiate to osteoblast lineage cells that deposit collagen I at the endosteal surface, creating a hostile environment for normal HSCs. In JAK2 V617F - driven MPN, neuropathy and loss of nestin + MSCs are hallmarks of disease progression, mediated by IL-1-driven inflammation. In MLL–AF9-driven AML, sympathetic neuropathy leads to the depletion of periarteriolar NG2 + MSCs but expands the population of remodelled osteoblast-primed nestin + MSCs through a mechanism dependent on β-adrenergic signalling. Primary driver mutations in bone marrow niche cells, particularly osteoprogenitor cells, can initiate myeloid malignancies within the haematopoietic compartment by establishing an inflammatory milieu. Identification of bone marrow niche cell types in mice Bone marrow niche cells have been characterized by protein expression (cell-surface and intracellular), by genetic labelling and by lineage tracing. Expression patterns often correlate with specialized locations of niche cells in vascular bone marrow niches (capillaries, arterioli or sinusoids), particularly for MSCs and endothelial cells. Diverse bone marrow niches influence HSC fate The bone marrow contains a dense neurovascular network that is innervated by sympathetic nerves and includes an enrichment of arterioles and capillaries in the endosteum, as well as large sinusoids located throughout the endosteum and central bone marrow cavity. Cellular bone marrow niches are closely associated with this neurovascular network and contain stromal elements including perivascular mesenchymal stromal cells (MSCs), endothelial cells (of arterioles, capillaries and sinusoids), differentiated osteoprogenitors and adipocytes. Haematopoietic cells of the bone marrow niche include megakaryocytes, macrophages and osteoclasts. These diverse cell types provide crucial signals to HSCs in the form of soluble factors, direct cell–cell contact and cell- surface ligands that influence fates such as mobilization, quiescence and differentiation. Central bone marrow Bone Endosteum Nutrient artery Nutrient vein Articular cartilage Central artery Central vein/sinusoid Sympathetic nerve Periostial capillary Sinusoid Arteriole
Transcript of WA27038-Bone Marrow Niches HSC Fates€¦ · Adipocytes Perilipin+ Caveolin+ Haematopoietic cells...
AdipocytesPerilipin+
Caveolin+
Stromal cellsHaematopoietic cells
LEPR+
+
SCA1+
Periarteriolar Perisinusoidal
CD45–
CD31–PDGFRA+
PDGFRB+CD105+ CD51+
TER119–
Prx1 Gli1
Ng2Nestin–GFP (high)
Cxcl12ScfNestin–GFP (dim)Foxc1
MSCs
Gremlin
Lineage–
SCA1+
KIT+
CD48–
CD105+
FLT3–
CD150+
PROCR+
CD34–
α-catulinHoxB5Tie2
HSCs
Protein expression
Genetic labelling
Osteoprogenitors
OsterixNestinCxcl12
CD31–
CD45–
TER119–
CD51+
PDGFRA+
SinusoidsVEGFR3+
CD144low
TJP1low
CD31+
MECA32+
VEGFR2+
Laminin+
Endothelialcells
Capillaries
Arterioli
SCA1+
TJP1hi
CD144hi
Tie2
Nestin
Osteoblasts
Collagen 1OsterixOsteocalcin
ALCAM+
SCF
IL-1
G-CSF
CXCL12
ANGPT1
Monocyte
Quiescent HSC
Cycling HSC
Quiescent HSC
Cycling HSC Quiescent HSC
Cycling HSC
Mobilized HSC
T cell
NK cell
Myeloid cells
Myeloid cells
TLR
IFNα/β IFNγ
IFNγ
IL- 1 G-CSF
Viral infection Bacterial infection Malignancy
Inflammation
OsteoclastsTRAP+
CD169+
F4/80+
CD115int
GR1–
Macrophages
Macrophage Leukocytes
Fibroblast
Apoptotic HSC
Megakaryocyte
IFNα/β
CD61+ VWF+ CXCL4+ CD41+ Clec2
Megakaryocytes
Notchligand
SinusoidCXCL12
HSC mobilizationVCAM1CXCL12
–
TGFβ1 TPOCXCL4SCF CXCL12ANGPT1
–
+
HSC quiescence and maintenance
E-selectin
+
HSC cycling and differentiation
IL-7EPONotch ligands FGF1
SCF
Latent TGFβ1
Active TGFβ1
Noradrenaline
FGF1
Macrophage
Endothelial cell
Non-myelinating Schwann cell
Sympathetic nerve
Sinusoid
Sinusoid
Perisinusoidal MSC
Bone Central bone marrow
Arteriole
Sinusoid
MSC
Osteoclast
Osteoblast
Sinusoid
Periarteriolar MSC
Perisinusoidal MSC
CXCL12
IL-7
EPO
TNF
ANGPT1
CXCL12
SCF
LatentTGFβ1
TPO CXCL4
Adiponectin
OsteoprogenitorAdipocyte
Macrophage
Arteriole
VCAM1
E-selectin
Endosteum
Arteriole
Endothelial cell
Megakaryocyte
IL-1
IL1
CCL3 TPO
CXCL12
Malignant myeloid cellsMalignant
myeloid cells
Loss of NG2+ MSC
Non-functional osteoprogenitor
Neuropathy
Neuropathy
Inflammation
Transformation
HSCNestin+ MSC
Loss of nestin+ MSC
Loss of nestin+ MSC
Impaired differentiation
MLL–AF9blasts
Niche-driven malignancies
Haematopoietic cell-driven malignancies
MLL–AF9-driven AMLJAK2V617F-driven MPN
Primary driver mutation
Sinusoid
ArterioleOsteoprogenitors
Endosteal collagen 1 fibrosis
Remodelled endosteal MSC
Leukaemicstem cell
BCR–ABL-driven CML
HSC
Leukaemicstem cell
SCF
CXCL12
ANGPT1
Endomucin+
Arteriole
Bone marrow niches and HSC fatesEvgenia V. Verovskaya, Timothy B. Campbell and Emmanuelle Passegué
Self-renewing and multipotent haematopoietic stem cells (HSCs) generate all mature blood cells. Adult HSCs exist in highly specialized bone marrow niches. These niches have crucial roles in regulating the fate of HSCs in terms of quiescence, mobilization into the peripheral blood and differentiation in response to steady-state and emergency cues. HSC fate is influenced by diverse types of stromal and haematopoietic cells that make up the bone marrow niche and provide signals in the form of soluble factors, direct cell–cell contact and cell-surface ligands. In stress conditions, such as
during an inflammatory response, bone marrow niches respond by regulating the balance of downstream HSC fates. In the case of myeloid malignancies, bone marrow niches can be remodelled to create an environment that supports malignant stem cells but impairs the maintenance of normal HSCs. Understanding the signalling pathways of the bone marrow niche will aid the therapeutic use and targeting of HSCs, as well as provide more general insights into stem cell regulation and the function and composition of stem cell niches.
STEMCELL Technologies — Your Ideas. Our Tools.Having the right tools for the isolation, culture and analysis of haematopoietic stem and progenitor cells (HSPCs) is essential for increasing our understanding of the mechanisms controlling HSPC behaviour and fate decisions. This knowledge furthers the development of cell therapies to treat haematological disorders. STEMCELL supports every step of your HSPC research with products for:
Document #27038 | Version 1.0.0
Expansion & Differentiation• Serum-free media and supplements for culture,
expansion and lineage-specific differentiation ofHSPCs (www.StemSpan.com)
• Media and detection reagents for long-termculture of HSPCs in the presence of stromal cells(www.MyeloCult.com)
• Cytokines and small molecules for completeflexibility in the design of experimental cultureconditions
Analysis• Semi-solid methylcellulose-based (www.MethoCult.com) and
collagen-based (www.MegaCult.com) media to quantitateprogenitor cells of different lineages in haematopoieticcolony assays
• Antibodies for immunophenotyping and sorting of HSPCs andmature blood cell subsets (www.STEMCELL.com/Antibodies)
To see our full portfolio of products for HSPC research visit www.STEMCELL.com/HSPCworkflow
Cell Sourcing & Isolation • Fresh and frozen primary haematopoietic cells
isolated from human donors(www.STEMCELL.com/PrimaryCells)
• Column-free isolation of HSPCs(www.EasySep.com & www.RosetteSep.com)
• Instruments for fully automated, walk-away cellisolation (www.RoboSep.com)
Abbreviations ALCAM, activated leukocyte cell adhesion molecule; AML, acute myeloid leukaemia; ANGPT1, angiopoietin 1; Clec2, C-type lectin domain family 2; CML, chronic myeloid leukaemia; CXCL, CXC-chemokine ligand; EPO, erythropoietin; FGF1, fibroblast growth factor 1; FLT3, Fms-related tyrosine kinase 3; Foxc1, Forkhead box c1; GFP, green fluorescent protein; G-CSF, granulocyte colony- stimulating factor; IL, interleukin; LEPR, leptin receptor; MPN, myeloproliferative neoplasm; PDGFRA, platelet-derived growth factor receptor A; PROCR, protein C receptor; SCF, stem cell factor;
TGFβ1, transforming growth factor β1; TJP1, tight junction protein 1; TLR, Toll-like receptor; TNF, tumour necrosis factor; TPO, thrombopoietin; TRAP, tartrate-resistant acid phosphatase; VCAM1, vascular cell adhesion molecule 1; VEGFR, vascular endothelial growth factor receptor; VWF, von Willebrand factor.
AffiliationsEvgenia V. Verovskaya, Timothy Campbell and Emmanuelle Passegué are at the Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell Research, University of California San Francisco,
California 94143, USA. Evgenia V. Verovskaya and Emmanuelle Passegué are relocating to the Columbia Stem Cell Initiative, Columbia University Medical Center, New York 10032, USA.Correspondence to E.P. e-mail: [email protected] authors declare no competing interests.
Edited by Kirsty Minton; designed by Caio Bracey.© 2017 Macmillan Publishers Limited, part of Springer Nature. All rights reserved.http://www.nature.com/posters/hsc
Stress signals alter HSC fatesDiverse stressors and insults to the bone marrow, such as bacterial and viral infection and malignancy, create an inflammatory milieu that controls HSC fate via cell-surface receptors. Inflammatory factors can act on HSCs and their downstream progenitors through various mechanisms that are illustrated by the prototypical examples of type I and type II interferons (IFNs; IFNα/β and IFNγ, respectively), IL-1 and G-CSF. IFNs are produced preferentially by haematopoietic cells and act directly on HSCs to direct downstream differentiation and proliferation. IL-1 is produced under stress conditions by stromal and haematopoietic cells and acts directly on HSCs to drive an emergency myeloid response. G-CSF is produced by stromal and haematopoietic cells and acts indirectly on HSCs by downregulating the production of stromal cell-derived factors, which affects HSC quiescence and mobilization, and by the induction of TLR signalling, which leads to HSC proliferation.
The active roles of bone marrow niches in malignancyBone marrow niches have an active role in the initiation and progression of myeloid malignancies. In BCR–ABL-driven CML, remodelled endosteal MSCs differentiate to osteoblast lineage cells that deposit collagen I at the endosteal surface, creating a hostile environment for normal HSCs. In JAK2V617F-driven MPN, neuropathy and loss of nestin+ MSCs are hallmarks of disease progression, mediated by IL-1-driven inflammation. In MLL–AF9-driven AML, sympathetic neuropathy leads to the depletion of periarteriolar NG2+ MSCs but expands the population of remodelled osteoblast-primed nestin+ MSCs through a mechanism dependent on β-adrenergic signalling. Primary driver mutations in bone marrow niche cells, particularly osteoprogenitor cells, can initiate myeloid malignancies within the haematopoietic compartment by establishing an inflammatory milieu.
Identification of bone marrow niche cell types in miceBone marrow niche cells have been characterized by protein expression (cell-surface and intracellular), by genetic labelling and by lineage tracing. Expression patterns often correlate with specialized locations of niche cells in vascular bone marrow niches (capillaries, arterioli or sinusoids), particularly for MSCs and endothelial cells.
Diverse bone marrow niches influence HSC fateThe bone marrow contains a dense neurovascular network that is innervated by sympathetic nerves and includes an enrichment of arterioles and capillaries in the endosteum, as well as large sinusoids located throughout the endosteum and central bone marrow cavity. Cellular bone marrow niches are closely associated with this neurovascular network and contain stromal elements including perivascular mesenchymal stromal cells (MSCs), endothelial cells (of arterioles, capillaries and sinusoids), differentiated osteoprogenitors and adipocytes. Haematopoietic cells of the bone marrow niche include megakaryocytes, macrophages and osteoclasts. These diverse cell types provide crucial signals to HSCs in the form of soluble factors, direct cell–cell contact and cell-surface ligands that influence fates such as mobilization, quiescence and differentiation.
Central bone marrow
Bone
Endosteum
Nutrient artery
Nutrientvein
Articular cartilage
Central artery
Central vein/sinusoid
Sympathetic nerve
Periostial capillary
Sinusoid
Arteriole
marrow niches are closely associated with this neurovascular network and contain stromal elements including perivascular mesenchymal stromal cells (MSCs), endothelial cells (of
Central vein/sinusoidCentral vein/sinusoid
Periostial capillary
Sinusoid
Arteriole
Nutrient artery
Periostial capillary
Sinusoid
Arteriole
cells (MSCs), endothelial cells (of
adipocytes. Haematopoietic cells of the bone marrow niche
macrophages and osteoclasts.
Nutrient artery
NutrientveinNutrientNutrientNutrientNutrientNutrientveinveinveinveinvein
cells (MSCs), endothelial cells (of cells (MSCs), endothelial cells (of cells (MSCs), endothelial cells (of
cells of the bone marrow niche
macrophages and osteoclasts.
cells (MSCs), endothelial cells (of


















