
Treatment with α-helical-CRF(9–41) prevents the anorectic effect of 17-β-estradiol
4
Brain Research Bulletin, Vol. 32, pp. 689-692, 1993 0361-9230/93 $6.00 + .OO Printed in the USA. All rights reserved. Copyright0 1993 Pergamon Press Ltd. BRIEF COMMUNICATION Treatment with a-Helical-CRF~9_41j Prevents the Anorectic Effect of 17-P-Estradiol ANNE DAGNAULT, DHAOUI OUERGHI AND DENIS RICHARD’ Department of Physiology, Faculty of Medicine, Lava1 University, Quebec City, GIK7P4, Canada Received 2 1 December 1992; Accepted 14 May 1993 DAGNAULT, A., D. OUERGHI AND D. RICHARD. Treatment with a-helicul-CRF~9_,,, prevents the anorectic e$ect of 17-B- estradiol. BRAIN RES BULL 32(6) 689-692, 1993.-The role of corticotropin-releasing factor (CRF) in the anorexia induced by 1 ‘I-&estradiol (E,) has been assessed in castrated female rats that were trained to eat their daily food ration in three separate meals. Each rat was implanted with a permanent guide cannula that was aimed at the right lateral ventricle of the brain. Seven days after the brain surgery each rat was also subcutaneously implanted with an osmotic minipump containing Buserelin, a potent GnRH agonist that induces reversible castration in rats. Eight rats were used in the study, and each of them underwent four experimental treatments that consisted of a) a subcutaneous (SC) injection of oil combined with an intracerebroventricular (ICY) infusion of saline, b) a SC injection of E2 combined with an ICV infusion of saline c) a SC injection of oil combined with an ICV infusion of a-helical CRF,a_al,. and d) a SC in&ion of E, combined with an ICV iniection of a-helical CRF,.A,,. Subcutaneous ~, ..,_ injections of E2 or oil were carried out the day before the ICV infusions of cy-helical CRFo_,,) or saline. Intr&~broventricular infusions were performed 30 min before the meal for which the interaction effect of E2 and a-helical CRFfe4,, on fbod intake was determined. E2 and a-helical CRFodu interacted on food intake; Ez brought about a 33% reduction in food intake in rats when infused with saline, whereas it was without effect when infused with cY-helical-CRFo_&reated rats. The present results provide evidence that CRF is involved in the anorectic effect of Ez . Food intake Corticotropin-releasing factor Ovariectomy Buserelin Estrogen IT has been known for years that estrogens exert anorectic effects in rodents (30-32) as well as in other species (9,13). Reduced intakes of food have been observed following the preovulatory surge of estradiol (Ez) (9,12,13,28,32), which occurs either at proestrus or at the end of the follicular phase, depending on the species. In addition, the anorectic effects of estrogens have also been emphasized in ovariectomized animals, in which Ez re- placement therapies prevent the orexigenic effect of castration (9,13,16). The mechanisms by which estrogens suppress feeding have yet to be fully described. Interestingly, recent findings lent in- direct support for a role of the anorectic neuropeptide cortico- tropin-releasing factor (CRF) ( 18) in the effect of estrogens on food intake. In fact, during proestrus, when food intake is at its estrus cycle lowest level (12,28,32), ACTH and corticosterone plasma concentrations (5), as well as CRF mRNA in the par- vocellular portion of the paraventricular nucleus (PVN) (3) are elevated. Additionally, it has been recently reported that bilateral adrenalectomy, a procedure known to stimulate CRF release, may significantly attenuate the inducing effects of ovariectomy on energy gain (19). This finding suggests that the estrogen re- moval, which enhance energy deposition primarily by increasing energy intake (20,32), may produce such an action by reducing the brain CRF tone. To assess the role of CRF in the anorectic effects of estrogens, food intake measurements were carried out in castrated female rats concomitantly treated with Ez and cY-helica1-CRF(94,j, a CRF antagonist. METHOD Animal and Diet Female Wistar rats weighing approximately 180 g were housed singly in wire-bottom cages that were suspended above absorbent paper. Upon their arrival in the laboratory, the rats were trained to eat their daily food ration in three meals, the ’ Requests for reprints should be addressed to Dr. Denis Richard, D&rtement de Physiologic, Facult6 de MCdecine, Universit6 Laval, Quebec (QuC), G 1 K 7P4, Canada. 689
Transcript of Treatment with α-helical-CRF(9–41) prevents the anorectic effect of 17-β-estradiol

Brain Research Bulletin, Vol. 32, pp. 689-692, 1993 0361-9230/93 $6.00 + .OO Printed in the USA. All rights reserved. Copyright 0 1993 Pergamon Press Ltd.
BRIEF COMMUNICATION
Treatment with a-Helical-CRF~9_41j Prevents the Anorectic Effect of 17-P-Estradiol
ANNE DAGNAULT, DHAOUI OUERGHI AND DENIS RICHARD’
Department of Physiology, Faculty of Medicine, Lava1 University, Quebec City, GIK7P4, Canada
Received 2 1 December 1992; Accepted 14 May 1993
DAGNAULT, A., D. OUERGHI AND D. RICHARD. Treatment with a-helicul-CRF~9_,,, prevents the anorectic e$ect of 17-B- estradiol. BRAIN RES BULL 32(6) 689-692, 1993.-The role of corticotropin-releasing factor (CRF) in the anorexia induced by 1 ‘I-&estradiol (E,) has been assessed in castrated female rats that were trained to eat their daily food ration in three separate meals. Each rat was implanted with a permanent guide cannula that was aimed at the right lateral ventricle of the brain. Seven days after the brain surgery each rat was also subcutaneously implanted with an osmotic minipump containing Buserelin, a potent GnRH agonist that induces reversible castration in rats. Eight rats were used in the study, and each of them underwent four experimental treatments that consisted of a) a subcutaneous (SC) injection of oil combined with an intracerebroventricular (ICY) infusion of saline, b) a SC injection of E2 combined with an ICV infusion of saline c) a SC injection of oil combined with an ICV infusion of a-helical CRF,a_al,. and d) a SC in&ion of E, combined with an ICV iniection of a-helical CRF,.A,,. Subcutaneous ~, ..,_ injections of E2 or oil were carried out the day before the ICV infusions of cy-helical CRFo_,,) or saline. Intr&~broventricular infusions were performed 30 min before the meal for which the interaction effect of E2 and a-helical CRFfe4,, on fbod intake was determined. E2 and a-helical CRFodu interacted on food intake; Ez brought about a 33% reduction in food intake in rats when infused with saline, whereas it was without effect when infused with cY-helical-CRFo_&reated rats. The present results provide evidence that CRF is involved in the anorectic effect of Ez .
Food intake Corticotropin-releasing factor Ovariectomy Buserelin Estrogen
IT has been known for years that estrogens exert anorectic effects in rodents (30-32) as well as in other species (9,13). Reduced intakes of food have been observed following the preovulatory surge of estradiol (Ez) (9,12,13,28,32), which occurs either at proestrus or at the end of the follicular phase, depending on the species. In addition, the anorectic effects of estrogens have also been emphasized in ovariectomized animals, in which Ez re- placement therapies prevent the orexigenic effect of castration (9,13,16).
The mechanisms by which estrogens suppress feeding have yet to be fully described. Interestingly, recent findings lent in- direct support for a role of the anorectic neuropeptide cortico- tropin-releasing factor (CRF) ( 18) in the effect of estrogens on food intake. In fact, during proestrus, when food intake is at its estrus cycle lowest level (12,28,32), ACTH and corticosterone plasma concentrations (5), as well as CRF mRNA in the par- vocellular portion of the paraventricular nucleus (PVN) (3) are elevated. Additionally, it has been recently reported that bilateral
adrenalectomy, a procedure known to stimulate CRF release, may significantly attenuate the inducing effects of ovariectomy on energy gain (19). This finding suggests that the estrogen re- moval, which enhance energy deposition primarily by increasing energy intake (20,32), may produce such an action by reducing the brain CRF tone.
To assess the role of CRF in the anorectic effects of estrogens, food intake measurements were carried out in castrated female rats concomitantly treated with Ez and cY-helica1-CRF(94,j, a CRF antagonist.
METHOD
Animal and Diet
Female Wistar rats weighing approximately 180 g were housed singly in wire-bottom cages that were suspended above absorbent paper. Upon their arrival in the laboratory, the rats were trained to eat their daily food ration in three meals, the
’ Requests for reprints should be addressed to Dr. Denis Richard, D&rtement de Physiologic, Facult6 de MCdecine, Universit6 Laval, Quebec (QuC), G 1 K 7P4, Canada.
689

690 DAGNAULT. OUERGHI AND RICHARD
TABLE I SEQUENCE OF TREATMENTS TO WHICH RATS WERE SUBJECTED IN THE PRESENT STUDY
Day I Day 3 Day 5 Day 7
(Rats #I and 2) (Rats #3 and 4) (Rats #5 and 6)
(Rats #7 and 8)
Oil-saline Oil-a-helical CRF+,,) Ez-saline
El-a-helical CRF+41)
Oil-a-helical CRFtgA,) E+aline Ez-cu-helical CRF,,_,,) Oil-saline
E,-saline E,-oc-helical CRF+4,j
Oil-saline Oil-cu-helical CRF+4,)
E,-a-helical CRF+,,, Oil-saline
Oil-oc-hehcal CRF,9_41) E+aline
onset of which occurred every 8 h; specifically, rats were allowed to eat at 0700, 1500, and 2300 h. During the meal, rats had free access to a pelleted stock diet (Purina Chow No. 5001) for 1 h. The energy density of the diet was - 17.0 kJ/g wet wt. Rats had continually free access to tap water, and they were maintained on a 12L: 12D cycle (lights off 1400-0200) at an environmental temperature of 23°C.
Experimental Procedures
Rats were anesthetized with a mixture of ketamine hydro- chloride (10 ml) and xylazine (1.5 ml), which was injected in- traperitoneally (0.1 ml per 100 g of bwt.). They were then mounted on a stereotaxy apparatus (David Kopf Instruments,
8- 7-
3 -
& 6- - 2 5. z 4-
8- 7-
s
%
6-
2 5- - z 4-
8 9
31 2- l-
O-
u-helical CRF: P < 0.003
E2 x a-he1 CRF: P < 0.007
q Oil-a-helical CRF
icv infusions
24 hours before icv infusions 24 hours after icv infusions
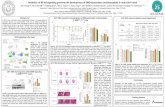
FIG. I. Food intakes in rats subjected to the following four treatments: a) a subcutaneous (SC) injection of oil combined with an intracerebro- ventricular (ICV) infusion of saline, b) a SC injection of E2 combined with an ICV infusion of saline, c) a SC injection of oil combined with an ICV infusion of cu-helical CRF(,,,,, and d) a SC injection of E2 com- bined with an ICV injection of o-helical CRFoA,,. Each rat underwent the four treatments in a specific order (see text). The figure also illustrates the intakes of food both the day preceding and the day following the ICV infusions of saline or a-helical CRF(9-41). In all conditions the food intake measurements were carried out over the midafternoon meal which was provided between 1500 and 1600 h.
Tujunga, CA) and a 23-gauge, IO mm-long guide cannula was vertically inserted into the brain in the direction of the right lateral ventricle. With the incisor bar placed at 3.3 mm below the interaural line, the coordinates from bregma for the guide cannula were as follows: anteropostetior (A-P), - I .O; lateral (L), - 1.2; dorsoventral (D-V), -2.0 mm (14). Animals were allowed to rest 7 days before further experimental treatments. After the recovery, all rats were chemically castrated with Buserelin, a GnRH agonist analog kindly provided by Hoechst Canada (Montreal, Canada). GnRH agonist analogs, which are com- pounds many times more potent than naturally occurring GnRH, paradoxically suppress gonadal function by inducing a reduction in the biologically active gonadotropin secretion through a pituitary desensitization to their own actions (10,25). In addition, Buserehn has proved as efficient as surgical ovariec- tomy to promote energy deposition in rats (2 I). In the present study, 12 pg of Buserelin were delivered daily in the intercellular space from an osmotic minipump (Alzet Model 2002) that was subcutaneously implanted in the middorsal region of the rats. The model of minipump used insured a constant release for 14 days.
Six days after the beginning of the perfusion with Buserelin, each of the eight rats used in the study were assigned to four experimental treatments, which were administered every 2 days in four distinct sequences (Table 1). Subcutaneous injections of E2 or oil were carried out the day before the ICV infusions of a-helical CRF+4,) or saline. lntracerebroventricular infusions were performed 30 min before the midafternoon meal for which the interaction effect of E2 and a-helical CRFc9-41) on food intake was determined. Fifteen micrograms of E2 dissolved in 0.15 ml of corn oil were injected 22 h before the ICV infusions of cy- helical CRFc-41) or saline; preliminary experiments showed that SC injections of Ez in oil did not produce an immediate anorectic effect. Twenty-five micrograms of a-helical CRF were ICV in- jected 30 min before the meal. Alpha-helical CRFc9_41) was dis- solved as follows: 1 mg of the substance was added to a volume of about 200 ~1 of distilled water to which was further added less than 10 ~1 of NaOH 0.1 N that caused a-helical CRFc9-41) to dissolve. Alpha-helical CRF was kindly provided by Dr. J. Rivier from the Clayton Foundation of the Salk Institute. Alpha- helical CRFe+n) infusions were performed in nonanesthetized rats at a rate of 2 &min for 2.5 min. The solution was injected with a 30-gauge, 12-mm-long cannula which was inserted into the guide cannula. After the injection the rats were put back into their respective cages. Half an hour later, they were offered their midafternoon meal. The meal was given in a food dispenser that prevented food spillage and, therefore, allowed careful food intake measurements.
Statistical Analysis
Main and interaction effects of Ez and a-helical CRFc9-41) were assessed by means of a 2 X 2 analysis of variance for re-

CRF AND THE ANORECTIC EFFECTS OF ESTROGENS 691
peated measures. Post hoc comparisons were performed with the Dunn-Sidak procedure (14).
RESULTS AND DISCUSSION
The analysis of the results expressed in Fig. 1, revealed a significant interaction between E2 and a-helical CRF+,,) on food intake; Ez brought about a significant decrease in food intake when combined with saline, whereas it had no significant effect when combined with a-helical CRFc9-41). Twenty-four hours be- fore ICV infusions, measurements of food ingestion during the midafternoon meal provided no significant difference between the rats that were ultimately assigned to the ICV infusions. Sim- ilarly, there was no difference between groups in the food ingested during the meal 24 h after the ICV infusions.
The role of CRF in the control of food intake has been largely studied during the past few years ( 18). Numerous authors have shown that an intracerebroventricular injection of CRF leads to a reduction of food intake in rats ( 1,2,4,22). In addition, evidence has accumulated, which shows that CRF may modulate the an- orectic effects of certain stressful stimuli known to increase CRF secretion (l&23,26). In rats, stressful procedures such as a re- straint stress or a bout of exercise lead to a food intake reduction that is largely prevented by a treatment with a-helical CRFcgA,, (23,26). The present results extend the involvement of CRF in the anorexia caused by EZ, and they support the notion that CRF plays a central role in various forms of anorexia ( 18).
I.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
The mechanisms of action underlying the CRF-mediated an- orectic effects of E2, as well as the brain sites where the interaction may occur, can only be speculatively discussed. Interestingly, it has been recently suggested that Ez treatment may lead to a functional impairment of the glucocorticoid receptor (6). This loss in the ability for glucocorticoid receptors to autoregulate would lead to a stimulation of CRF neurons, which would trans- late into an increase in the plasma concentrations of ACTH and corticosterone. In this respect, it has been recently reported that CRF mRNA is elevated in the parvocellular portion of the PVN on the afternoon of proestrus (3) coincidentally with the proestrus peak of Ez. The proestrus peak of Ez is known to induce, together with a GnRH-dependent surge of gonadotropins, a noticeable reduction in food intake (12,28,32). Interestingly, the increase in CRF mRNA reported by Bohler et al. (3) occurs in the PVN, a brain site acknowledged for its importance in the modula- tion of energy balance ( 11,17,27,29). The PVN contains the highest concentration of CRF-containing nerve cells in the hy- pothalamus (24) and it has been suggested to be involved in the anorectic effects of E2 (7,8). This suggestion has, however, not been corroborated in a long-term energy balance study in which it was shown that PVN lesions do not prevent the effects of Ez in ovariectomized rats (Dagnault and Richard, unpub- lished data). The exact mechanisms by which Ez inhibits food intake through a CRF-modulated process warrant further studies.
REFERENCES
Arase, K.; Shargill, M. S.; Bray, G. A. Effects of intraventricular infusion of corticotropin-releasing factor on VMH-lesioned obese rats. Am. J. Phvsiol. 256:R75 I-R756: 1989. Arase, K.; Shar&l, N. S.; Bray, G. A. Effects of corticotropin releasing factor on genetically obese (fatty) rats. Physiol. Behav. 45565-570; 1989. Bohler, H. C. L.; Zoeller, R. T.; King, J. C.; Rubin, B. S.; Weber, R.; Merriam, G. R. Corticotropin releasing hormone messenger RNA is elevated on the afternoon of proestrus in the parvocellular para- ventricular nuclei of the female rat. Mol. Brain Res. 8:259-262; 1990. Britton, D. R.; Kood, G. F.; Rivier, J.; Vale, W. Intraventricular corticotropin-releasing factor enhances behavioral effects of novelty. Life Sci. 3 1:363-367; 1982. Buckingham, J. C.; Dohler, K. D.; Wilson, C. A. Activity of the pituitary-adrenocortical system and thyroid gland during the oestrous cycle of the rat. J. Endocrinol. 78:359-366; 1978. Burgess, L. H.; Handa, R. J. Chronic estrogen-induced alterations in adrenocorticotropin and corticosterone secretion, and glucocor- ticoid receptor-mediated functions in female rats. Endocrinology 131:1261-1269; 1992. Butera, P. C.; Beikirch, R. J. Central implants of diluted estradiol- independent effects on ingestive and reproductive behaviors of ovariectomized rats. Brain Res. 49 l:266-273; 1989. Butera, P. C.; Willard, D. M.; Raymond, S. A. Effects of PVN lesions on the responsiveness of female rats to estradiol. Brain Res. 576:304-3 IO; 1992. Czaja, J. A.; Goy, R. W. Ovarian hormones and food intake in female guinea pigs and rhesus monkeys. Horm. Behav. 6:329-349; 1975. Fraser, H. M. LHRH analogues: Their clinical physiology and de- livery systems. Bailliere. Clin. Endocrinol. Met. 2:639-658; 1988. Fukushima, M.; Tokunaga, K.; Lupien, J.; Kemnitz, J. W.; Bray, G. A. Dynamic and static phases of obesity following lesions in PVN and VMH. Am. J. Phvsiol. 253:R523-R529: 1990. Haar, M. B. T. Circadian and estrual rhythms in food intake in the rat. Horm. Behav. 3:213-219; 1972.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
Kemnitz, J. W.; Gibber, J. R.; Lindsay, K. A.; Eisele, S. G. Effects of ovarian hormones and eating behaviors, body weight, and glu- coregulation in rhesus monkeys. Harm. Behav. 23:235-250; 1989. Kirk, R. E. Experimental design: Procedures for the behavioral sci- ences. Pacific Grove: Brooks/Cole; 1982. Krahn, D. D.; Gosnell, B. A.; Grace, M.; Levine, A. S. CRF antagonist partially reverses CRF- and stress-induced effect on feeding. Brain Res. Bull. 17:285-289; 1986. Landau, T.; Zucker, I. Estrogenic regulation of body weight in the female rat. Horm. Behav. 7:29-39; 1976. Menendez, J. A.; Atrens, D. M. Insulin and the paraventricular hy- pothalamus-Modulation of energy balance. Brain Res. 555: l93- 201; 1991. Morley, J. E.; Levine, A. S. Corticotropin-releasing factor and inges- tive behaviors. In: De Souza, E. B.; Nemeroff, C. B., eds. Cortico- tropin-releasing factor basic and clinical studies of a neuropeptide. Boca Raton: CRC; 1990:267-274. Ouerghi, D.; Rivest, S.; Richard, D. Adrenalectomy attenuates the effect of chemical castration on energy balance in rats. J. Nutr. 122:369-373; 1992. Richard, D. Effects of ovarian hormones on energy balance and brown adipose tissue thermogenesis. Am. J. Physiol. 250:R245- R249; 1986. Richard, D.; Dekoninck, P.; Lemay, A.; Rivest, S. Reversible increase in energy deposition following castration induced by a gonadotropin releasing hormone agonist. Int. J. Obes. 15:155-161; 1991. Rivest, S.; Richard, D. The effects of corticotropin-releasing factor on energy balance in rats are sex-dependent. Am. J. Physiol. 257:Rl417-R1422; 1989. Rivest, S.; Richard, D. Involvement ofcorticotropin-releasing factor in the anorexia induced by exercise. Brain Res. Bull. 25: 169-172; 1990. Sawchenko, P. E.; Swanson, L. W. Organization of CRF immu- noreactive cells and fibers in the rat brain: Immunohistochemical studies. In: De Souza, E. B.; Nemeroff, C. B., eds. Cotticotropin- releasing factor: Basic and clinical studies of a neuropeptide. Boca Raton: CRC: 1990:30-5 I.

692 DAGNAULT. OUERGHI AND RICHARD
25. Schaison, G. GnRH and its analogs-Structure, mechanism of action and therapeutic uses. J. Steroid Biochem. 33:795-799; 1989.
26. Shibasaki, T.: Yamauchi. N.; Kato, Y.: Masuda, A.; Imaki, T.: Hotta, M.; Demura. H.; Oono, H.; Ling, N.; Shizume, K. Involvement of corticotropin-releasing factor in restraint stress-induced anorexia and reversion of the anorexia by somatostatin in the rat. Life Sci. 43:1103-l 110; 1988.
27. Sims, J. S.; Lorden. J. F. Effect of paraventricular nucleus lesions on body weight, food intake and insulin levels. Behav. Brain Res. 22:265-28 I: 1986.
28. Tarttelin, M. F.: Go&i, R. A. Variations in food and water intake in the normal and acyclic female rat. Physiol. Behav. 7:847-852: 197 I
29. Tokunaga, K.; Fukushima. M.: Kemnitz. J. W.; Bray, G. A. Com- parison of ventromedial and paraventricular lesions in rats that be- come obese. Am. J. Physiol. 25l:R1221-Rl2R7: 1986.
30. Wade. G. N. Sex hormones, regulatory behaviors. and body weight. In: Rosenbiatt. J. S.: Hinde. R. A.: Show, E.: Beer. C. G.. eds. Ad- vances in the study of behavior. New York: Academic: I976:20 I- 279.
31. Wade. G. N.; Ciray. J. M. Gonadal effects on food Intake and adiposity: A metabolic hypothesis. Physiol. Behav. 22:583-593: 1979.
32. Wade, G. N.: Gray. J. M.: Bat-mess, -1. J. Gonadal influences on adiposity. Int. J. Obes. 9:83-92: 1985.










![[1] involuteΣ(Spur and Helical Gear Design) 1.3 Software ...Eng).pdf · [1] involuteΣ(Spur and Helical Gear Design) 1 t Fig..1.1 Calculation Result Screen 1.1 Introduction involute](https://static.fdocument.org/doc/165x107/5a7894b47f8b9a7b698d1836/1-involutespur-and-helical-gear-design-13-software-engpdf1-involutespur.jpg)






