
Endovascular repair of traumatic aortic transection six years of experience

November 2012 1947Biol. Pharm. Bull. 35(11) 1947–1955 (2012)
© 2012 The Pharmaceutical Society of Japan
The Antioxidant Effect of Carnosol in Bovine Aortic Endothelial Cells Is Mainly Mediated via Estrogen Receptor α PathwayPi-wen Zhao,a,b David Yue-wei Lee,a Zhong-ze Ma,a Lian-sha Huang,a,b Li-ping Sun,b Ya-dong Li,b Jia-xu Chen,b and Jian-zhao Niu*,b
a Bio-Organic and Natural Products Research Laboratory, Mclean Hospital (Harvard Medical School); Belmont, MA 02478, U.S.A.: and b Department of Cell and Biochemistry, Beijing University of Chinese Medicine; Beijing 100029, China. Received April 10, 2012; accepted July 28, 2012; advance publication released online August 28, 2012
Antioxidant action is critical for maintaining the normal cardiovascular function and vascular endo-thelial cell is an important target of estrogen action through estrogen receptor (ER) pathway. This study is carried out to explore the antioxidant effect of carnosol in bovine aortic endothelial cells (BAECs) via ER pathway. The ER subtype specific estrogenic effect of carnosol was further demonstrated by luciferase reporter gene assay in human embryonic kidney (HEK) 293 cells. Carnosol was extracted from Chinese med-icine Rosmarinus officinalis. ER positive BAECs were employed in cell proliferation assay and cell apoptosis tests. Oxidative stress by intracellular reactive oxygen species (ROS) were measured via 2′7′-dichlorofluo-rescein (DCF) production. ERα and ERβ specific antagonists 1,3-bis(4-hydroxyphenyl)-4-methyl-5-[4-(2-piperidinylethoxy)phenol]-1H-pyrazole (MPP) and 4-[2-phenyl-5,7-bis(trifluoromethyl)pyrazolo[1,5-a]py-rimidine-3-yl]phenol (PHTPP) were employed as tools in the experiment. ER negative HEK 293 cells were employed in luciferase reporter gene assay. The results indicate that carnosol can effectively attenuate H2O2 induced slowing down of cell growth and increasing of cell apoptosis. At the meantime, carnosol pretreating can also effectively reduce the H2O2 induced intracellular ROS elevation in BAECs. ERα and ERβ antago-nist, especially ERα antagonist, can effectively decrease the above antioxidant effects of carnosol. The re-porter gene analysis further demonstrates that the action of carnosol on inducing ERE dependent luciferase expression is realized via ER pathway. The conclusion is that carnosol can exert antioxidant effects towards oxidative stress induced by H2O2 in BAECs. And such effects are realized via ER, especially ERα pathway. The results contribute to explain the mechanism of cardiovascular protective function of carnosol in post-menopausal women.
Key words carnosol; antioxidant effect; estrogen receptor; phytoestrogen; bovine aortic endothelial cell; re-porter gene
Endogenous estrogens play essential roles not only in the hypothalamic-pituitary-gonadal axis, but also in cardiovas-cular and central nervous systems as well as bone and lipid metabolism. It has been reported that the cardiovascular risk rises significantly in recent years. The statistical results indi-cated that cardiovascular disease (CVD) remains one of the main causes of death among women, especially aged women experiencing postmenopausal periods.1–3) Besides the side effect of estrogen replacement therapy (ERT) on leading to higher breast cancer incidence, Women’s Health Initiative (WHI) and the heart and estrogen/progestin replacement study (HERS) found that estrogen did not reduce adverse cardiovas-cular events and, in fact, increased the number of cardiovas-cular events.4) Thus people are paying much more attention to phytoestrogens in recent years.
Phytoestrogens are a group of compounds with chemical structures resembling natural and synthetic estrogens and thus enable them the binding affinity to estrogen receptors (ERs). Through activation or up-regulation and inactivation or down-regulation of certain genes, they can mimic some actions of human estrogens.5) Various kinds of phytoestrogen perform different biological activity due to their structure and deviations in the structure. Phytoestrogen genistein, found in soy products, has been reported to possess antioxidant and estrogenic activity.6) Although estrogen always shows similar high affinity with both ERα and ERβ, phytoestrogens often
have relatively different and lower affinity upon ERα and ERβ. Thus they may display some effects different from those of estrogen. It has been reported that some active components in Chinese medicine such as resveratrol (RVT) possessed anti-oxidant properties.7) As a phytoestrogen, RVT can regulate the transcription of estrogen-responsive target genes via estrogen receptors.8,9) In bovine aortic endothelial cells (BAECs), RVT exerts endothelial improving function through both ERα and ERβ.10)
Carnosol is a dietary diterpene isolated from culinary herbs such as Rosemary, Sage and Oregano.11) The antioxidant prop-erties of rosemary, Rosmarinus officinalis L. (Labiatae), have been known for centuries.12) Carnosol has been proved to be an important antioxidant compound and contributes much to the corresponding function of Rosemary.13) It has also been reported to exert brand functions involving in human health such as anti-angiogenic activity, antiproliferative properties to-wards cancer cells of intestinal, mammary and prostate tumor, anti-inflammatory, anti-infective and anti-human immunode-ficiency virus (HIV) function.9,11,14–18) Carnosol has also been known to be an active anti-metastatic agent against malignant melanocytes.19)
As an essential part in treating postmenopausal syndromes, antioxidant action is critical for maintaining the normal car-diovascular function in women experiencing postmenopausal period. As mentioned above, estrogen receptors are main tar-get points, pathway and of great significance in dealing with menopausal syndromes. But till now, there has been no report
Regular Article
* To whom correspondence should be addressed. e-mail: [email protected]
The authors declare no conflict of interest.

1948 Vol. 35, No. 11
concerning about the correlation between antioxidant effects of carnosol and function of estrogen receptor subtypes. This experiment is designed and conducted to study the mechanism through which carnosol realizes its antioxidant effect via ER pathway. In order to analyse the ER subtype dependent estro-genic effects of carnosol much directly, its action on inducing ERE dependent luciferase expression via ERα or ERβ subtype in HEK 293 cells was also tested in our research. The results will contribute to combine the antioxidant action of carnosol with its clinical application in ER-related postmenopausal syn-drome alleviation.
MATERIALS AND METHODS
Drugs and Chemicals The dried leaves of Rosmarinus officinalis (100 g) were powdered and extracted with acetone for three times at room temperature, and the solution was evaporated under reduced pressure to give a residue. The ace-tone extract (5 g) was chromatographed over silica gel (200 g) using CH2Cl2 (Sigma Chemical Co., St. Louis, MO, U.S.A.) with increasing amounts of MeOH (Sigma Chemical Co.) to give carnosol (12 mg). The chemical structure of carnosol (Fig. 1) was further proved by HPLC and identified by comparison with the published NMR data.
Plasmids Professor Satoshi Inoue at the University of Tokyo, Japan, graciously provided plasmids containing ei-ther ERα or ERβ. Plasmid containing 5× estrogen–response element (ERE) linked to the luciferase reporter gene was recombined and stored by our laboratory. An additional β-ga-lactosidase plasmid (Clontech Laboratories Inc., Mountain View, CA, U.S.A.) was co-transfected in the assay system as an internal standard.
Culture of ER-Positive BAECs and ER-Negative Human Embryonic Kidney (HEK) 293 Cells BAECs and HEK 293 cells (American Type Culture Collection, Manassas, VA, U.S.A.) were maintained and subcultured in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, Grand Island, NY, U.S.A.) with 10% heat-inactivated fetal bovine serum (Hyclone, Logan, UT, U.S.A.), 100 U/mL penicillin, 100 µg/mL streptomycin (Hyclone) under humidified conditions at 5% CO2. Four days prior to each experiment, the media was sub-stituted with phenol red-free DMEM (Gibco) containing 10% charcoal stripped fetal bovine serum (CDT-FBS) (Hyclone). In the following experiments, each control group was treated with 0.1% solvent in DMEM with 10% FBS.
Cell Proliferation Test by 3-(4,5-Dimethylthiazol-2-yl)- 2,5-diphenyltetrazolium Bromide (MTT)-Assay BAECs were seeded into 96-well plates at 3×103 cells/well. Treat-ment media (100 µL/well) with 0.01–1 µM carnosol was added on the following day. After 24 h pre-treating, BAECs were stimulated with oxidative stress by exposed to 1 mM H2O2 for 15, 30, 60 min. In 30 min treating group, 10−8 M ERα or ERβ specific inhibitor 1,3-bis(4-hydroxyphenyl)-4-methyl-5-[4-(2-piperidinylethoxy)phenol]-1H-pyrazole (MPP) or 4-[2-phenyl- 5,7-bis(trifluoromethyl)pyrazolo[1,5-a]pyrimidine-3-yl]phenol (PHTPP) (Thermo Scientific, Park Ellisville, MO, U.S.A.) was added together with carnosol. Cells were harvested and their sensitivity to oxidants was evaluated by proliferation rate calculating via MTT assay. Fifteen micro liter MTT reagent (1 mg/mL) (Sigma Chemical Co.) was added into each well and incubated for an additional 4 h at 37°C. After the medium
was removed, the cells were solubilized by addition of 150 µL dimethyl sulfoxide (DMSO) (Sigma Chemical Co.). The plates were shaken at room temperature for 10 min. MTT is a yellow dye that can be actively converted by mitochondrial oxida-tion-reduction reactions into blue formazan crystals which could be dissolved in DMSO. Thus, the cellular viability could be determined by measuring the absorbance of the converted dye at a wavelength of 570 nm.
Morphological Assessment of Apoptosis by Hoechst 33342 Staining BAECs were cultured on 8-well Chamber slides. After pre-treating with 0.1–1 µM carnosol in the pres-ence or absence of 10−8 M estrogen receptor specific inhibitor MPP or PHTPP for 24 h, the BAECs were stimulated with oxidative stress by 1 mM H2O2 for 30 min. Then the cells were trypsinized, washed with PBS, fixed with 4% paraformalde-hyde for 15 min at room temperature. After stained with 5 µg/mL Hoechst 33342 (Sigma Chemical Co.) at 37°C for 15 min, samples were observed under a DMR fluorescence microscope (Carl Zeiss Imager A1, Germany) with UV illumination. The dye was excited at 340 nm, and fluorescence emission was filtered with a 510 nm barrier filter. To quantify the apoptotic process, cells with fragmented or condensed DNA and normal DNA were counted respectively. Data were expressed as the ratio of apoptotic cells to total cells.
Cell Apoptosis Assay by FACS BAECs were cultured in 6-well plates at 5×104 cells/well. After pretreated with 0.1–1 µM carnosol with or without 10−8 M estrogen receptor specific inhibitor MPP or PHTPP for 24 h, cells were exposed to 1 mM H2O2 for 30 min as oxidative stress stimulation. Sub-sequently, cells were collected and washed with PBS. Cell apoptotic activities were detected by Annexin V-FITC (fluo-rescen isothiocyanate) apoptosis detection kit (BD Biosci-ences, San Diego, CA, U.S.A.). Cells were resuspended in Binding Buffer and stained with 5 µL Annexin V-FITC and 5 µL propidium iodide (PI) for 15 min at RT (25°C) in the dark. After adding additional 400 µL Binding buffer to each tube, cell apoptosis was analyzed by Fluorescence Activated Cell Sorter (FACS) (FACS Calibur, BD Bioscience, CA, U.S.A.) within 1 h.
Measurement of Intracellular Reactive Oxygen Species (ROS) Formation by DCF Production Production of ROS was measured by oxidation sensitive fluorescent probe, 2′7′-di-chlorofluorescein diacetate (DCF-DA) (Sigma Chemical Co.) method, based on the ROS dependent oxidation of DCF-DA to DCF. 1×105 BAECs were plotted into 24 well plates. Cells were grown in DMEM and pretreated with 0.1–1 µM carnosol in the presence or absence of 10−8 M estrogen receptor specific inhibitor MPP or PHTPP for 24 h. Subsequently, medium was removed and cells were washed with PBS twice, followed
Fig. 1. Chemical Structure of Carnosol
November 2012 1949
by incubation with 30 µm DCF-DA (Sigma Chemical Co.) for 30 min at 37°C in the dark. After washed with PBS to remove excess DCF-DA, the oxidative stress stimulation was performed by 1 mM H2O2 for 30 min. After washed two times with PBS, ROS generation was measured by microplate reader as the fluorescence intensity of each well at an excitation wavelength of 495 nm and an emission wavelength of 530 nm.
Plasmid Transfection and Luciferase Reporter Gene Assay in HEK 293 Cells One day before transfection, HEK 293 cells were plotted into 24-well plates at a concentration of 2×104 cells/well. For transfection, maintenance media was aspirated and 100 µL transfection mix was added. Cells were cotransfected with 0.1 µg ERα or 0.1 µg ERβ, 0.8 µg ERE-luc and 0.1 µg β-gal control using lipofectamine 2000 (2 µL/well) (Invitrogen, Carlsbad, CA, U.S.A.). After an incubation of 4 h for transfection according to the manufacturer’s instructions,
experimental medium was added without removal of the trans-fection media. The experimental medium was prepared with phenol red-free DMEM, 10% charcoal-stripped fetal bovine serum (HyClone) and carnosol with the final concentrations from 10−6 to 10−11 M. Each experiment also included an estra-diol standard curve with 10-fold dilutions from 10−8 to 10−13 M. A vehicle control containing 0.1% ethanol, which was the maximal level of ethanol used in the treatment media, was in-cluded in the test. Cells were incubated for 24 h at 37°C. Then media was aspirated and cells were lysed with 100 µL Glo Lysis Buffer (Promega, Madison, WI, U.S.A.). Lysates were transferred to microcentrifuge tubes and stored at −70°C. For the luciferase assay, 50 µL lysate was added to 50 µL steady Glo luciferase assay reagent (Promega). Luminescence was measured with multifunctional microplate reader (Tecan). A β-galactosidase assay kit (Promega) was used to test enzyme
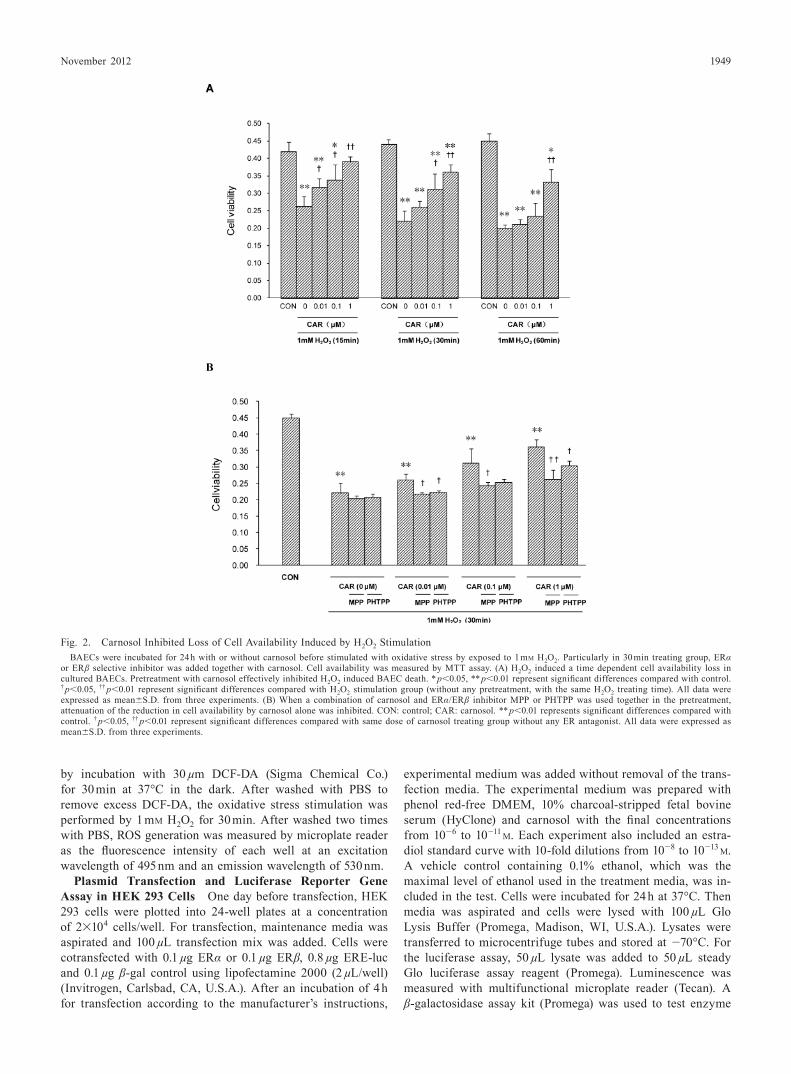
Fig. 2. Carnosol Inhibited Loss of Cell Availability Induced by H2O2 StimulationBAECs were incubated for 24 h with or without carnosol before stimulated with oxidative stress by exposed to 1 mM H2O2. Particularly in 30 min treating group, ERα
or ERβ selective inhibitor was added together with carnosol. Cell availability was measured by MTT assay. (A) H2O2 induced a time dependent cell availability loss in cultured BAECs. Pretreatment with carnosol effectively inhibited H2O2 induced BAEC death. * p<0.05, ** p<0.01 represent significant differences compared with control. † p<0.05, †† p<0.01 represent significant differences compared with H2O2 stimulation group (without any pretreatment, with the same H2O2 treating time). All data were expressed as mean±S.D. from three experiments. (B) When a combination of carnosol and ERα/ERβ inhibitor MPP or PHTPP was used together in the pretreatment, attenuation of the reduction in cell availability by carnosol alone was inhibited. CON: control; CAR: carnosol. ** p<0.01 represents significant differences compared with control. † p<0.05, †† p<0.01 represent significant differences compared with same dose of carnosol treating group without any ER antagonist. All data were expressed as mean±S.D. from three experiments.
1950 Vol. 35, No. 11
activity. The final result of luciferase activity was standard by β-gal value in order to correct for well-to-well variation. The maximal achievable response to 10−8 M estradiol was arbi-trarily set at 100%, luciferase activity in response to carnosol was measured by relative light units (RLUs).
Data Analysis All data were represented as mean val-ues± S.D. Experimental results were analyzed by using facto-rial analysis of variance (ANOVA) with appropriate post hoc test (SPSS version 13.0). Differences were considered statis-tically significant at p values of 0.05 or less. Median effect concentration (EC50) of all samples tested was estimated by nonlinear regression analysis. The computer program Origin was used for the analysis of sigmoidal dose–response curves to obtain estimates of EC50 values.
RESULTS
Carnosol Inhibited Reduction of Cell Viability MTT assay showed that 1 mM H2O2 treating induced a time-de-pendent cell viability reduction in BAECs. After exposure to 1 mM H2O2 for 30 or 60 min, the result indicated a significant reduction of 49.0% and 45.6% viability (relative to control, re-spectively) (Fig. 2). Pretreatment with different concentrations of carnosol attenuated H2O2 induced cell number reduction. As for 30 min in 1 µM carnosol treating group, cell availability remained 69.5% of control. However, if ERα or ERβ antago-nist was added in the pretreatment together with carnosol, the attenuation of the reduction in cell availability was inhibited.
Carnosol Inhibited Cell Apoptosis by Morphological Study and FACS Test Hoechst 33342 staining indicates the condensation in the typical apoptotic cell nucleus. As shown in Fig. 3, control cells had normal round nuclei with dispersed
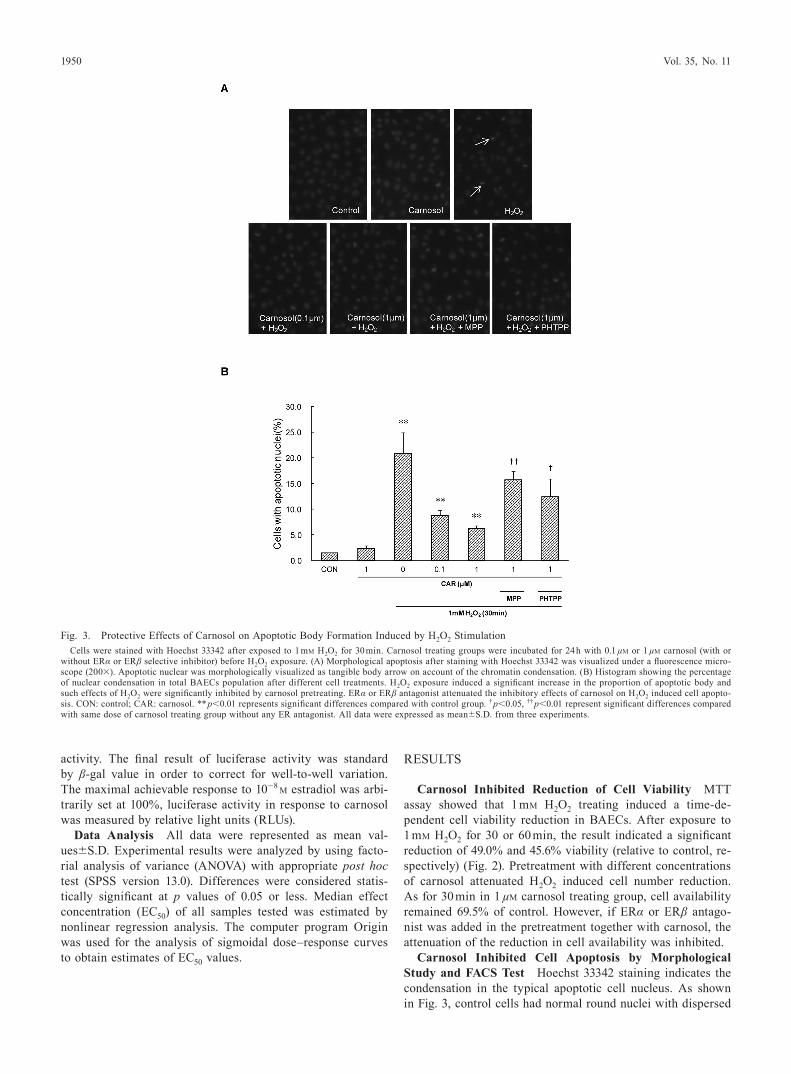
Fig. 3. Protective Effects of Carnosol on Apoptotic Body Formation Induced by H2O2 StimulationCells were stained with Hoechst 33342 after exposed to 1 mM H2O2 for 30 min. Carnosol treating groups were incubated for 24 h with 0.1 µM or 1 µM carnosol (with or
without ERα or ERβ selective inhibitor) before H2O2 exposure. (A) Morphological apoptosis after staining with Hoechst 33342 was visualized under a fluorescence micro-scope (200×). Apoptotic nuclear was morphologically visualized as tangible body arrow on account of the chromatin condensation. (B) Histogram showing the percentage of nuclear condensation in total BAECs population after different cell treatments. H2O2 exposure induced a significant increase in the proportion of apoptotic body and such effects of H2O2 were significantly inhibited by carnosol pretreating. ERα or ERβ antagonist attenuated the inhibitory effects of carnosol on H2O2 induced cell apopto-sis. CON: control; CAR: carnosol. ** p<0.01 represents significant differences compared with control group. † p<0.05, †† p<0.01 represent significant differences compared with same dose of carnosol treating group without any ER antagonist. All data were expressed as mean±S.D. from three experiments.
November 2012 1951
DNA molecule and chromatin. After treated with 1 mM H2O2 for 30 min, BAECs showed a 20.8 percentage of chromatin condensation and nuclear shrinkage. In the carnosol pretreated group, percentage of apoptosis cells was decreased signifi-cantly.
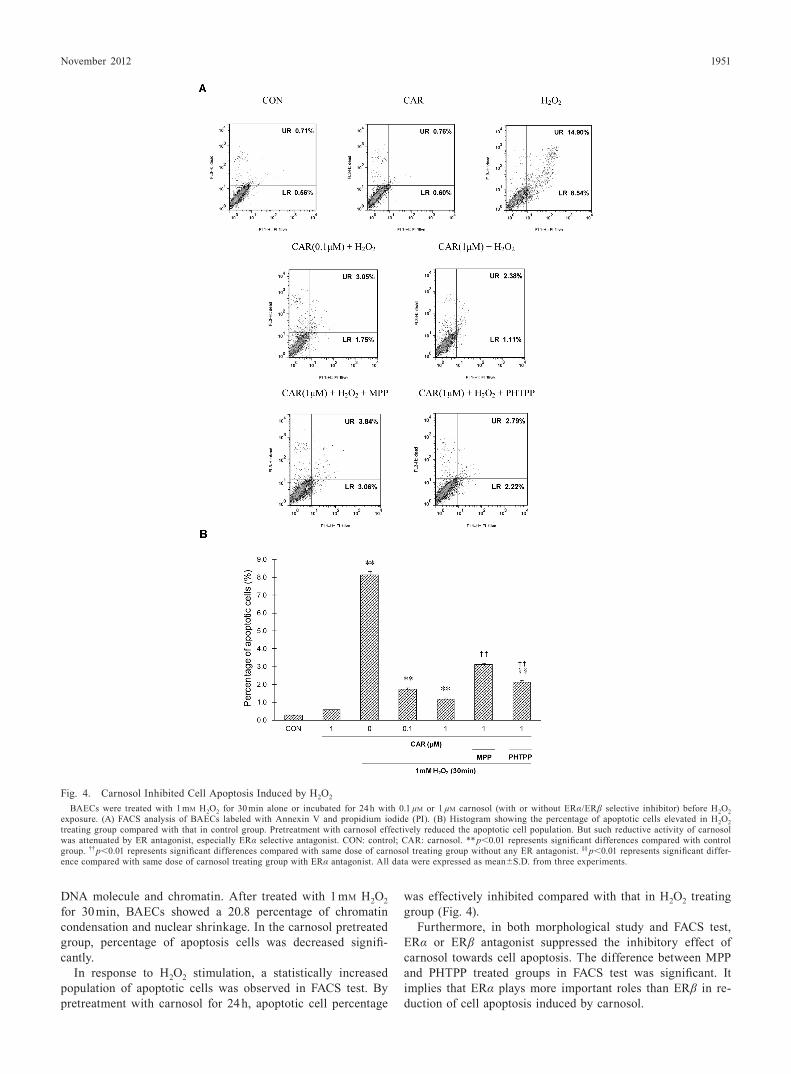
In response to H2O2 stimulation, a statistically increased population of apoptotic cells was observed in FACS test. By pretreatment with carnosol for 24 h, apoptotic cell percentage
was effectively inhibited compared with that in H2O2 treating group (Fig. 4).
Furthermore, in both morphological study and FACS test, ERα or ERβ antagonist suppressed the inhibitory effect of carnosol towards cell apoptosis. The difference between MPP and PHTPP treated groups in FACS test was significant. It implies that ERα plays more important roles than ERβ in re-duction of cell apoptosis induced by carnosol.
Fig. 4. Carnosol Inhibited Cell Apoptosis Induced by H2O2
BAECs were treated with 1 mM H2O2 for 30 min alone or incubated for 24 h with 0.1 µM or 1 µM carnosol (with or without ERα/ERβ selective inhibitor) before H2O2 exposure. (A) FACS analysis of BAECs labeled with Annexin V and propidium iodide (PI). (B) Histogram showing the percentage of apoptotic cells elevated in H2O2 treating group compared with that in control group. Pretreatment with carnosol effectively reduced the apoptotic cell population. But such reductive activity of carnosol was attenuated by ER antagonist, especially ERα selective antagonist. CON: control; CAR: carnosol. ** p<0.01 represents significant differences compared with control group. †† p<0.01 represents significant differences compared with same dose of carnosol treating group without any ER antagonist. §§ p<0.01 represents significant differ-ence compared with same dose of carnosol treating group with ERα antagonist. All data were expressed as mean±S.D. from three experiments.
1952 Vol. 35, No. 11
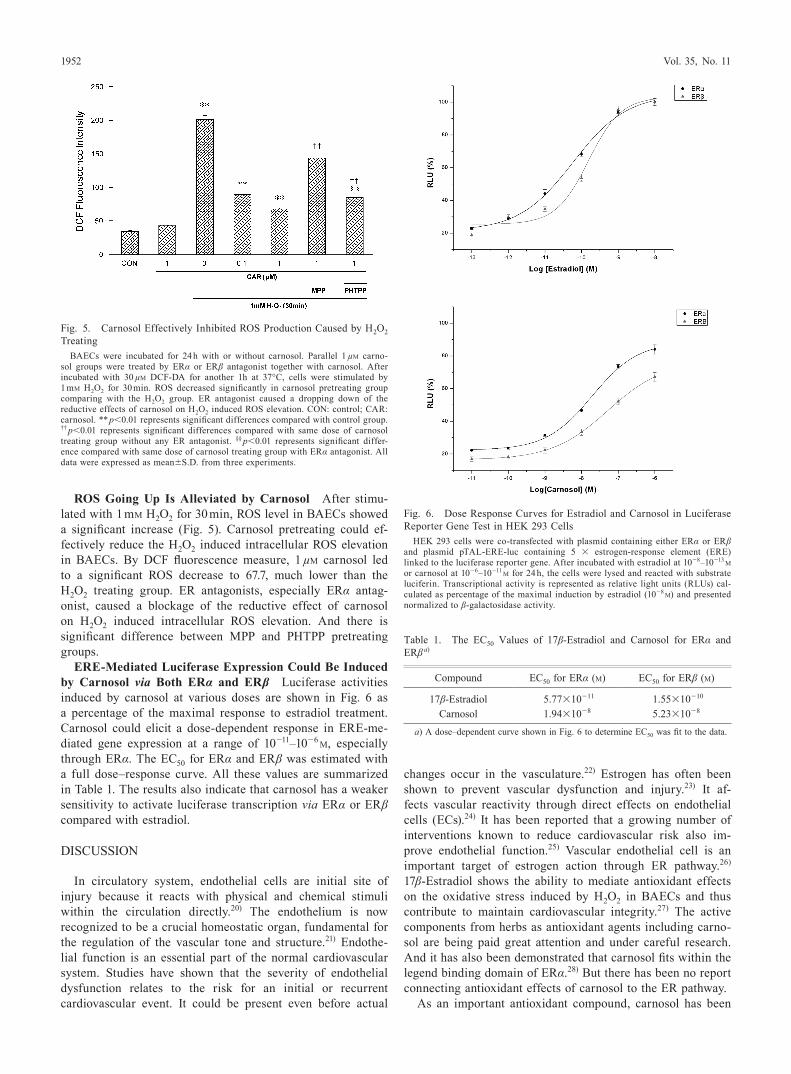
ROS Going Up Is Alleviated by Carnosol After stimu-lated with 1 mM H2O2 for 30 min, ROS level in BAECs showed a significant increase (Fig. 5). Carnosol pretreating could ef-fectively reduce the H2O2 induced intracellular ROS elevation in BAECs. By DCF fluorescence measure, 1 µM carnosol led to a significant ROS decrease to 67.7, much lower than the H2O2 treating group. ER antagonists, especially ERα antag-onist, caused a blockage of the reductive effect of carnosol on H2O2 induced intracellular ROS elevation. And there is significant difference between MPP and PHTPP pretreating groups.
ERE-Mediated Luciferase Expression Could Be Induced by Carnosol via Both ERα and ERβ Luciferase activities induced by carnosol at various doses are shown in Fig. 6 as a percentage of the maximal response to estradiol treatment. Carnosol could elicit a dose-dependent response in ERE-me-diated gene expression at a range of 10−11–10−6 M, especially through ERα. The EC50 for ERα and ERβ was estimated with a full dose–response curve. All these values are summarized in Table 1. The results also indicate that carnosol has a weaker sensitivity to activate luciferase transcription via ERα or ERβ compared with estradiol.
DISCUSSION
In circulatory system, endothelial cells are initial site of injury because it reacts with physical and chemical stimuli within the circulation directly.20) The endothelium is now recognized to be a crucial homeostatic organ, fundamental for the regulation of the vascular tone and structure.21) Endothe-lial function is an essential part of the normal cardiovascular system. Studies have shown that the severity of endothelial dysfunction relates to the risk for an initial or recurrent cardiovascular event. It could be present even before actual
changes occur in the vasculature.22) Estrogen has often been shown to prevent vascular dysfunction and injury.23) It af-fects vascular reactivity through direct effects on endothelial cells (ECs).24) It has been reported that a growing number of interventions known to reduce cardiovascular risk also im-prove endothelial function.25) Vascular endothelial cell is an important target of estrogen action through ER pathway.26) 17β-Estradiol shows the ability to mediate antioxidant effects on the oxidative stress induced by H2O2 in BAECs and thus contribute to maintain cardiovascular integrity.27) The active components from herbs as antioxidant agents including carno-sol are being paid great attention and under careful research. And it has also been demonstrated that carnosol fits within the legend binding domain of ERα.28) But there has been no report connecting antioxidant effects of carnosol to the ER pathway.
As an important antioxidant compound, carnosol has been
Fig. 5. Carnosol Effectively Inhibited ROS Production Caused by H2O2 Treating
BAECs were incubated for 24 h with or without carnosol. Parallel 1 µM carno-sol groups were treated by ERα or ERβ antagonist together with carnosol. After incubated with 30 µM DCF-DA for another 1h at 37°C, cells were stimulated by 1 mM H2O2 for 30 min. ROS decreased significantly in carnosol pretreating group comparing with the H2O2 group. ER antagonist caused a dropping down of the reductive effects of carnosol on H2O2 induced ROS elevation. CON: control; CAR: carnosol. ** p<0.01 represents significant differences compared with control group. †† p<0.01 represents significant differences compared with same dose of carnosol treating group without any ER antagonist. §§ p<0.01 represents significant differ-ence compared with same dose of carnosol treating group with ERα antagonist. All data were expressed as mean±S.D. from three experiments.
Fig. 6. Dose Response Curves for Estradiol and Carnosol in Luciferase Reporter Gene Test in HEK 293 Cells
HEK 293 cells were co-transfected with plasmid containing either ERα or ERβ and plasmid pTAL-ERE-luc containing 5 × estrogen-response element (ERE) linked to the luciferase reporter gene. After incubated with estradiol at 10−8–10−13 M or carnosol at 10−6–10−11 M for 24 h, the cells were lysed and reacted with substrate luciferin. Transcriptional activity is represented as relative light units (RLUs) cal-culated as percentage of the maximal induction by estradiol (10−8 M) and presented normalized to β-galactosidase activity.
Table 1. The EC50 Values of 17β-Estradiol and Carnosol for ERα and ERβ a)
Compound EC50 for ERα (M) EC50 for ERβ (M)
17β-Estradiol 5.77×10−11 1.55×10−10
Carnosol 1.94×10−8 5.23×10−8
a) A dose–dependent curve shown in Fig. 6 to determine EC50 was fit to the data.

November 2012 1953
demonstrated in our experiment to possess a significant action on BAEC proliferation in vasoprotective antioxidant events during cell oxidative stress triggered by H2O2. Carnosol showed a BAEC protective action towards oxidative damage. Specific ER subtype antagonist could effectively attenuate the anti-proliferative action of carnosol. That is to say, carnosol is able to display a phytoestrogenic action in BAECs.
There are two isoforms of estrogen receptors: ERα and ERβ. Both are found in endothelial cells.29) Through activation of ERs and alternation of related gene expression, estrogen exerts multiple vascular effects including antioxidant proper-ties.30) Actions of estrogen are primarily mediated by these two molecular targets. Nuclear ER signaling has been also considered one major mechanism for regulating the growth of endothelial cells.20) Through both ERs, estrogen exerts the function of preventing vascular dysfunction and injury and maintaining the integrity of vascular endothelium in postmenopausal woman.23,27) ERα and ERβ show different effects during that procedure.31) In several cases, ERα has been reported as the main factor in antioxidative process. As mentioned in the beginning, much more attention has been paid to phytoestrogens in recent years, especially some active components in Chinese medicine that show estrogenic effects. As a multifunctional compound, phytoestrogen resveratrol has attracted much attention to exert endothelial improving func-tion through both ERα and ERβ. Our cell proliferation results showed that the antioxidative action of carnosol is also medi-ated through ER pathway.
At the meantime, it has been reported that ERα and ERβ display different functions in mediating cell proliferation. ERβ is reported to play much important rolls in reducing cell proliferation rate in ER positive cells.32) Comparing with es-trogen, phytoestrogen also possesses the ability to activate ER pathway and always shows different binding ability to ERα and ERβ due to their similar but different structure or binding site. Thus the downstream effects of phytoestrogen are always different from estrogen.
According to the morphological observations by Hoechst staining, carnosol showed antioxidant ability towards the going up cell apoptosis rate induced by H2O2. The result of FACS assay also indicated an antioxidant effect of carnosol towards cell apoptosis. And in both tests, a descending of pro-tective effects of carnosol towards oxidant stress was observed with ERα or ERβ antagonist added together with carnosol. The difference between MPP and PHTPP treated groups in FACS analysis was significant. It implies that ER pathway, especially ERα, is involved in the process of anti-apoptosis function of carnosol.
ROS can be produced by several enzyme systems in the vascular wall.33) ROS level and the balance of ROS and anti-oxidants are intracellular signs of oxidant status. It has been reported that resveratrol could decrease the expression of NADPH oxidase which is the predominant source of ROS in endothelial cells.34) By the measurement of intracellular ROS, it was proved in our experiment that content of ROS increased by H2O2 treating was decreased by pretreatment with carno-sol. Decreased ROS level went up again with ER antagonist, especially ERα antagonist. This is a further demonstration of the ER, mainly ERα pathway through which carnosol realizes its antioxidant function.
Both ER subtypes have significant sequence homology in
their DNA-binding domain (96%), but the homology is to a lesser degree in their ligand-binding domain (53%).35) ERα and ERβ are also different in distribution and expression levels in various tissues or cell types.36,37) All these contribute to the different effects induced by binding of different ligands. Both ERα and ERβ possess six functional regions.38) The activation function 1 (AF-1) at the N-terminal and the activation func-tion 2 (AF-2) at the C-terminal are two key regions for bind-ing of various ligands and exerting effects of target genes.39) The lower homology in AF-1 and AF-2 regions may affect the binding mode of estrogen or estrogen-like substances and lead to the consequent change in the conformation of ER-ligand complex. Thus the final effects of different ligands on target gene expression are diverse. Estrogen possesses high affinity with both ERα and ERβ in BAECs, especially ERα. The re-sults of our reporter gene test showed that estradiol could in-duce the expression of luciferase via either ERα or ERβ, with EC50 values of 5.77×10−11 M and 1.55×10−10 M respectively. The affinity of carnosol upon ERα and ERβ is also different. Just like estradiol, carnosol induced greater luciferase expression via ERα than ERβ. It has been reported that some kinds of phytoestrogen possess relatively higher affinity with ERβ.40,41) But our results showed that ERα is a more effective target point for carnosol than ERβ. And the results also demonstrate that the affinity of phytoestrogen with ERα and ERβ differed from that of estradiol owing to their diversity in conformation and the low homology in ligand binding regions of ERs.42) So the ER-ligand complex with specific conformation would bind to ERE and activate certain ERE-induced gene expressions in various degrees.
Above all, our study focused on the antioxidant effects of carnosol via ER pathway in BAECs. The ER subtype, espe-cially ERα dependent estrogenic effect of carnosol has been further demonstrated by reporter gene assay performed in HEK 293 cells. The results contribute to explain the protective function and its pathways of Chinese medicine in treating or alleviating postmenopausal cardiovascular symptoms.
Furthermore, several animal studies have suggested that daily oral administration of carnosol is well tolerated.28) A good prospect of its clinical application can be predicted.
Acknowledgements This work on extraction and explo-ration of the antioxidant effect and its pathway of carnosol has been performed at Bio-Organic and Natural Products Research Laboratory in Mclean Hospital (Harvard Medical School) and is supported by the award No. 30772849 from the National Natural Science Foundation of China (NSFC), 2011CXTD-07 from the program for Innovative Research Team in Beijing University of Chinese Medicine and No. JBYZZ-JS014 from the Natural Science Foundation of Beijing University of Chinese Medicine. The authors thank Dr. Denny Zhao for performing the FACS tests in Massachusetts General Hospital (MGH) and providing valuable suggestions for the FACS results processing.
REFERENCES
1) Grady D, Gebretsadik T, Kerlikowske K, Ernster V, Petitti D. Hormone replacement therapy and endometrial cancer risk: a meta-analysis. Obstet. Gynecol., 85, 304–313 (1995).
2) Weiss LK, Burkman RT, Cushing-Haugen KL, Voigt LF, Simon

1954 Vol. 35, No. 11
MS, Daling JR, Norman SA, Bernstein L, Ursin G, Marchbanks PA, Strom BL, Berlin JA, Weber AL, Doody DR, Wingo PA, McDonald JA, Malone KE, Folger SG, Spirtas R. Hormone replace-ment therapy regimens and breast cancer risk (1). Obstet. Gynecol., 100, 1148–1158 (2002).
3) Yusuf S, Anand S. Hormone replacement therapy: a time for pause. CMAJ, 167, 357–359 (2002).
4) Deschamps AM, Murphy E. Activation of a novel estrogen receptor, GPER, is cardioprotective in male and female rats. Am. J. Physiol. Heart Circ. Physiol., 297, H1806–H1813 (2009).
5) Turner JV, Agatonovic-Kustrin S, Glass BD. Molecular aspects of phytoestrogen selective binding at estrogen receptors. J. Pharm. Sci., 96, 1879–1885 (2007).
6) Widyarini S, Spinks N, Husband AJ, Reeve VE. Isoflavonoid com-pounds from red clover (Trifolium pratense) protect from inflamma-tion and immune suppression induced by UV radiation. Photochem. Photobiol., 74, 465–470 (2001).
7) Kovacic P, Somanathan R. Multifaceted approach to resveratrol bioactivity: Focus on antioxidant action, cell signaling and safety. Oxid. Med. Cell. Longev., 3, 86–100 (2010).
8) Le Corre L, Fustier P, Chalabi N, Bignon YJ, Bernard-Gallon D. Ef-fects of resveratrol on the expression of a panel of genes interacting with the BRCA1 oncosuppressor in human breast cell lines. Clin. Chim. Acta, 344, 115–121 (2004).
9) Aruoma OI, Spencer JP, Rossi R, Aeschbach R, Khan A, Mahmood N, Munoz A, Murcia A, Butler J, Halliwell B. An evaluation of the antioxidant and antiviral action of extracts of rosemary and Provençal herbs. Food Chem. Toxicol., 34, 449–456 (1996).
10) Ylikomi T, Bocquel MT, Berry M, Gronemeyer H, Chambon P. Co-operation of proto-signals for nuclear accumulation of estrogen and progesterone receptors. EMBO J., 11, 3681–3694 (1992).
11) Johnson JJ, Syed DN, Heren CR, Suh Y, Adhami VM, Mukhtar H. Carnosol, a dietary diterpene, displays growth inhibitory effects in human prostate cancer PC3 cells leading to G2-phase cell cycle arrest and targets the 5′-AMP-activated protein kinase (AMPK) pathway. Pharm. Res., 25, 2125–2134 (2008).
12) Richheimer SL, Bernart MW, King GA, Kent MC, Bailey DT. Antioxidant activity of lipid-soluble phenolic diterpenses from rose-mary. J. Am. Oil Chem. Soc., 73, 507–514 (1996).
13) McGovern PE, Mirzoian A, Hall GR. Ancient Egyptian herbal wines. Proc. Natl. Acad. Sci. U.S.A., 106, 7361–7366 (2009).
14) Sagar SM, Yance D, Wong RK. Natural health products that inhibit angiogenesis: a potential source for investigational new agents to treat cancer—Part 1. Curr. Oncol., 13, 14–26 (2006).
15) Visanji JM, Thompson DG, Padfield PJ. Induction of G2/M phase cell cycle arrest by carnosol and carnosic acid is associated with al-teration of cyclin A and cyclin B1 levels. Cancer Lett., 237, 130–136 (2006).
16) Flowers M, Thompson PA. t10c12 conjugated linoleic acid sup-presses HER2 protein and enhances apoptosis in SKBr3 breast cancer cells: possible role of COX2. PLoS ONE, 4, e5342 (2009).
17) Singletary K, MacDonald C, Wallig M. Inhibition by rosemary and carnosol of 7,12-dimethylbenz[a]anthracene (DMBA)-induced rat mammary tumorigenesis and in vivo DMBA-DNA adduct forma-tion. Cancer Lett., 104, 43–48 (1996).
18) Moran AE, Carothers AM, Weyant MJ, Redston M, Bertagnolli MM. Carnosol inhibits beta-catenin tyrosine phosphorylation and prevents adenoma formation in the C57BL/6J/Min/+ (Min/+) mouse. Cancer Res., 65, 1097–1104 (2005).
19) Huang SC, Ho CT, Lin-Shiau SY, Lin JK. Carnosol inhibits the invasion of B16/F10 mouse melanoma cells by suppressing metal-loproteinase-9 through down-regulating nuclear factor-kappa B and c-Jun. Biochem. Pharmacol., 69, 221–232 (2005).
20) Felty Q. Estrogen-induced DNA synthesis in vascular endothelial cells is mediated by ROS signaling. BMC Cardiovasc. Disord., 6, 16 (2006).
21) Versari D, Daghini E, Virdis A, Ghiadoni L, Taddei S. Endothelial dysfunction as a target for prevention of cardiovascular disease. Diabetes Care, 32, S314–S321 (2009).
22) Li H, Förstermann U. Resveratrol: a multifunctional compound improving endothelial function. Editorial to: “Resveratrol supple-mentation gender independently improves endothelial reactivity and suppresses superoxide production in healthy rats,” by S. Soylemez et al. Cardiovasc. Drugs Ther., 23, 425–429 (2009).
23) Kim JK, Levin ER. Estrogen signaling in the cardiovascular sys-tem. Nucl. Recept. Signal., 4, e013 (2006).
24) Miller VM, Mulvagh SL. Sex steroids and endothelial function: translating basic science to clinical practice. Trends Pharmacol. Sci., 28, 263–270 (2007).
25) Widlansky ME, Gokce N, Keaney JF Jr, Vita JA. The clinical implications of endothelial dysfunction. J. Am. Coll. Cardiol., 42, 1149–1160 (2003).
26) Klinge CM, Blankenship KA, Risinger KE, Bhatnagar S, Noisin EL, Sumanasekera WK, Zhao L, Brey DM, Keynton RS. Res-veratrol and estradiol rapidly activate MAPK signaling through estrogen receptor α and β in endothelial cells. J. Biol. Chem., 280, 7460–7468 (2005).
27) Song JY, Kim MJ, Jo HH, Hwang SJ, Chae B, Chung JE, Kwon DJ, Lew YO, Lim YT, Kim JH, Kim JH, Kim MR. Antioxidant effect of estrogen on bovine aortic endothelial cells. J. Steroid Biochem. Mol. Biol., 117, 74–80 (2009).
28) Johnson JJ, Syed DN, Suh Y, Heren CR, Saleem M, Siddiqui IA, Mukhtar H. Disruption of androgen and estrogen receptor activity in prostate cancer by a novel dietary diterpene carnosol: implica-tions for chemoprevention. Cancer Prev. Res. (Phila.), 3, 1112–1123 (2010).
29) O’Lone R, Knorr K, Jaffe IZ, Schaffer ME, Martini PG, Karas RH, Bienkowska J, Mendelsohn ME, Hansen U. Estrogen receptors alpha and beta mediate distinct pathways of vascular gene expres-sion, including genes involved in mitochondrial electron transport and generation of reactive oxygen species. Mol. Endocrinol., 21, 1281–1296 (2007).
30) Smiley DA, Khalil RA. Estrogenic compounds, estrogen receptors and vascular cell signaling in the aging blood vessels. Curr. Med. Chem., 16, 1863–1887 (2009).
31) Zhang QG, Raz L, Wang R, Han D, De Sevilla L, Yang F, Vadl-amudi RK, Brann DW. Estrogen attenuates ischemic oxidative damage via an estrogen receptor α-mediated inhibition of NADPH oxidase activation. J. Neurosci., 29, 13823–13836 (2009).
32) Roger P, Sahla ME, Mäkelä S, Gustafsson JA, Baldet P, Rochefort H. Decreased expression of estrogen receptor β protein in prolif-erative preinvasive mammary tumors. Cancer Res., 61, 2537–2541 (2001).
33) Förstermann U. Oxidative stress in vascular disease: causes, de-fense mechanisms and potential therapies. Nat. Clin. Pract. Cardio-vasc. Med., 5, 338–349 (2008).
34) Spanier G, Xu H, Xia N, Tobias S, Deng S, Wojnowski L, Forster-mann U, Li H. Resveratrol reduces endothelial oxidative stress by modulating the gene expression of superoxide dismutase 1 (SOD1), glutathione peroxidase 1 (GPx1) and NADPH oxidase subunit (Nox4). J. Physiol. Pharmacol., 60 (Suppl. 4), 111–116 (2009).
35) Muramatsu M, Inoue S. Estrogen receptors: how do they control re-productive and nonreproductive functions? Biochem. Biophys. Res. Commun., 270, 1–10 (2000).
36) Taylor AH, Al-Azzawi F. Immunolocalisation of oestrogen receptor beta in human tissues. J. Mol. Endocrinol., 24, 145–155 (2000).
37) Kuiper GG, Carlsson B, Grandien K, Enmark E, Häggblad J, Nils-son S, Gustafsson JA. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology, 138, 863–870 (1997).
38) Shao W, Brown M. Advances in estrogen receptor biology: pros-pects for improvements in targeted breast cancer therapy. Breast

November 2012 1955
Cancer Res., 6, 39–52 (2004).39) Fowler AM, Solodin N, Preisler-Mashek MT, Zhang P, Lee AV,
Alarid ET. Increases in estrogen receptor-alpha concentration in breast cancer cells promote serine 118/104/106-independent AF-1 transactivation and growth in the absence of estrogen. FASEB J., 18, 81–93 (2004).
40) Gutendorf B, Westendorf J. Comparison of an array of in vitro assays for the assessment of the estrogenic potential of natural and synthetic estrogens, phytoestrogens and xenoestrogens. Toxicology,
166, 79–89 (2001).41) Chrzan BG, Bradford PG. Phytoestrogen activate estrogen receptor
beta 1 and estrogenic responses in human breast and bone cancer cell lines. Mol. Nutr. Food Res., 51, 171–177 (2007).
42) Morito K, Hirose T, Kinjo J, Hirakawa T, Okawa M, Nohara T, Ogawa S, Inoue S, Muramatsu M, Masamune Y. Interaction of phy-toestrogens with estrogen receptors α and β. Biol. Pharm. Bull., 24, 351–356 (2001).


















