
Preparation and anti-influenza A virus activity of κ-carrageenan oligosaccharide and its sulphated...
9
Preparation and anti-influenza A virus activity of j-carrageenan oligosaccharide and its sulphated derivatives Wei Wang a,b,⇑,1 , Pan Zhang a,b,1 , Guang-Li Yu a,b , Chun-Xia Li a,b , Cui Hao a,b , Xin Qi a , Li-Juan Zhang a,b , Hua-Shi Guan a,b,⇑ a Key Laboratory of Marine Drugs, Ministry of Education, Ocean University of China, Qingdao 266003, PR China b Shandong Provincial Key Laboratory of Glycoscience & Glycoengineering, Ocean University of China, Qingdao 266003, PR China article info Article history: Received 25 October 2011 Received in revised form 16 January 2012 Accepted 31 January 2012 Available online 8 February 2012 Keywords: j-Carrageenan oligosaccharide Sulphated derivative Influenza A virus Structure–activity relationship Pulmonary viral titre Interferon abstract Carrageenans, the sulphated galactans derived from red algae, are attracting increasing interest in devel- oping potential anti-viral drugs. In this study, low molecular weight j-carrageenan oligosaccharides (KCO) and their sulphated derivatives (KCO-S) were prepared, and their anti-influenza A virus (IAV) prop- erties were investigated. The results indicated that KCO and KCO-S could effectively inhibit IAV multipli- cation in MDCK cells in a dose-dependent manner. Furthermore, a structure–activity relationship study showed that the degree of sulphation and molecular weight were the main factors that influenced the anti-IAV activity of j-carrageenan oligosaccharide. The most active j-carrageenan oligosaccharide had a sulphate content of 0.8–1.0 mole/mole of disaccharide and a molecular weight of 1–3 kDa. In addition, KCO and KCO-S could significantly improve survival and decrease pulmonary viral titres in IAV-infected mice. Moreover, the antiviral effect of KCO and KCO-S does not seem to be dependent on the interferon system. In conclusion, carrageenan oligosaccharide and its sulphated derivative have good inhibitory actions on IAV replication in vitro and in vivo. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction Influenza A virus (IAV) is an enveloped virus comprised of eight RNA genomic segments (Lamb & Takeda, 2001). It has been the cause of at least three pandemics in the last century, the most severe leading to more than 40 million fatalities in 1918–1919 (Yewdell & Garcia-Sastre, 2002). Recently, a novel H1N1 IAV strain (H1N1/09 virus) was identified and considered a strong candidate for a novel influenza pandemic (Nguyen et al., 2011). Current anti- IAV drugs are directed against neuraminidase (NA; zanamivir and oseltamivir) and the M2 protein (adamantanes) (Hay, Wolstenholme, Skehel, & Smith, 1985; Mendel et al., 1998). Despite these successes, concerns regarding drug efficacy, resistance, toxic- ity, and cost still remain (Hayden & Pavia, 2006). Hence, the devel- opment of novel antiviral agents that could be used alone or in combination with existing antivirals is of high importance. Carrageenan, the sulphated galactan isolated from marine red algae, has been extensively used in the food, cosmetic and pharmaceutical industries as a thickener and gelling agent (Grassauer et al., 2008). The main three types of carrageenans are k-, j-, and i-carrageenan, and each show different inhibitory ef- fects on different viruses (Pujol et al., 2006; Talarico & Damonte, 2007). Buck et al. (2006) demonstrated that i-carrageenan inhib- ited human papillomavirus (HPV) infection by blocking virus adsorption. Talarico and co-workers reported that k- and i-carra- geenan could inhibit dengue virus replication in mammalian and mosquito cells (Talarico & Damonte, 2007; Talarico, Noseda, Ducatti, Duarte, & Damonte, 2011). Despite having good inhibitory effects on virus infection, the high molecular weight (MW) and poor tissue-penetrating ability of carrageenan polysaccharides have limited their potential antiviral application in humans. Oligosaccharides obtained by chemical or enzymatic hydrolysis of polysaccharides have been attracting increasing interest in developing potential anti-viral drugs (Ji, Wang, Wu, & Luan, 2011). Ekblad et al. (2010) found that heparan sulphate (HS) mi- metic PI-88, a sulphomannan oligosaccharide of low MW, effi- ciently reduced the cell-to-cell spread of herpes simplex virus (HSV). Yamada and co-workers reported that O-acylated carra- geenan oligosaccharides with different MW had increased anti- HIV activity by depolymerisation and sulphation (Yamada, Ogamo, Saito, Uchiyama, & Nakagawa, 2000; Yamada et al., 1997). More- over, the high-solubility of sulphated oligosaccharides at neutral pH enhanced their potential use as anti-viral drugs. 0308-8146/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved. doi:10.1016/j.foodchem.2012.01.108 ⇑ Corresponding authors. Address: School of Medicine and Pharmacy, Ocean University of China, Qingdao 266003, PR China. Tel.: +86 532 8203 1980; fax: +86 532 8203 3054. E-mail addresses: [email protected], [email protected] (W. Wang), [email protected] (H.-S. Guan). 1 These authors contributed equally to this work. Food Chemistry 133 (2012) 880–888 Contents lists available at SciVerse ScienceDirect Food Chemistry journal homepage: www.elsevier.com/locate/foodchem
Transcript of Preparation and anti-influenza A virus activity of κ-carrageenan oligosaccharide and its sulphated...

Food Chemistry 133 (2012) 880–888
Contents lists available at SciVerse ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Preparation and anti-influenza A virus activity of j-carrageenan oligosaccharideand its sulphated derivatives
Wei Wang a,b,⇑,1, Pan Zhang a,b,1, Guang-Li Yu a,b, Chun-Xia Li a,b, Cui Hao a,b, Xin Qi a, Li-Juan Zhang a,b,Hua-Shi Guan a,b,⇑a Key Laboratory of Marine Drugs, Ministry of Education, Ocean University of China, Qingdao 266003, PR Chinab Shandong Provincial Key Laboratory of Glycoscience & Glycoengineering, Ocean University of China, Qingdao 266003, PR China
a r t i c l e i n f o
Article history:Received 25 October 2011Received in revised form 16 January 2012Accepted 31 January 2012Available online 8 February 2012
Keywords:j-Carrageenan oligosaccharideSulphated derivativeInfluenza A virusStructure–activity relationshipPulmonary viral titreInterferon
0308-8146/$ - see front matter � 2012 Elsevier Ltd. Adoi:10.1016/j.foodchem.2012.01.108
⇑ Corresponding authors. Address: School of MedUniversity of China, Qingdao 266003, PR China. Tel.:532 8203 3054.
E-mail addresses: [email protected], wangwWang), [email protected] (H.-S. Guan).
1 These authors contributed equally to this work.
a b s t r a c t
Carrageenans, the sulphated galactans derived from red algae, are attracting increasing interest in devel-oping potential anti-viral drugs. In this study, low molecular weight j-carrageenan oligosaccharides(KCO) and their sulphated derivatives (KCO-S) were prepared, and their anti-influenza A virus (IAV) prop-erties were investigated. The results indicated that KCO and KCO-S could effectively inhibit IAV multipli-cation in MDCK cells in a dose-dependent manner. Furthermore, a structure–activity relationship studyshowed that the degree of sulphation and molecular weight were the main factors that influenced theanti-IAV activity of j-carrageenan oligosaccharide. The most active j-carrageenan oligosaccharide hada sulphate content of 0.8–1.0 mole/mole of disaccharide and a molecular weight of 1–3 kDa. In addition,KCO and KCO-S could significantly improve survival and decrease pulmonary viral titres in IAV-infectedmice. Moreover, the antiviral effect of KCO and KCO-S does not seem to be dependent on the interferonsystem. In conclusion, carrageenan oligosaccharide and its sulphated derivative have good inhibitoryactions on IAV replication in vitro and in vivo.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Influenza A virus (IAV) is an enveloped virus comprised of eightRNA genomic segments (Lamb & Takeda, 2001). It has been thecause of at least three pandemics in the last century, the mostsevere leading to more than 40 million fatalities in 1918–1919(Yewdell & Garcia-Sastre, 2002). Recently, a novel H1N1 IAV strain(H1N1/09 virus) was identified and considered a strong candidatefor a novel influenza pandemic (Nguyen et al., 2011). Current anti-IAV drugs are directed against neuraminidase (NA; zanamivirand oseltamivir) and the M2 protein (adamantanes) (Hay,Wolstenholme, Skehel, & Smith, 1985; Mendel et al., 1998). Despitethese successes, concerns regarding drug efficacy, resistance, toxic-ity, and cost still remain (Hayden & Pavia, 2006). Hence, the devel-opment of novel antiviral agents that could be used alone or incombination with existing antivirals is of high importance.
Carrageenan, the sulphated galactan isolated from marinered algae, has been extensively used in the food, cosmetic and
ll rights reserved.
icine and Pharmacy, Ocean+86 532 8203 1980; fax: +86
pharmaceutical industries as a thickener and gelling agent(Grassauer et al., 2008). The main three types of carrageenans arek-, j-, and i-carrageenan, and each show different inhibitory ef-fects on different viruses (Pujol et al., 2006; Talarico & Damonte,2007). Buck et al. (2006) demonstrated that i-carrageenan inhib-ited human papillomavirus (HPV) infection by blocking virusadsorption. Talarico and co-workers reported that k- and i-carra-geenan could inhibit dengue virus replication in mammalian andmosquito cells (Talarico & Damonte, 2007; Talarico, Noseda, Ducatti,Duarte, & Damonte, 2011). Despite having good inhibitory effectson virus infection, the high molecular weight (MW) and poortissue-penetrating ability of carrageenan polysaccharides havelimited their potential antiviral application in humans.
Oligosaccharides obtained by chemical or enzymatic hydrolysisof polysaccharides have been attracting increasing interest indeveloping potential anti-viral drugs (Ji, Wang, Wu, & Luan,2011). Ekblad et al. (2010) found that heparan sulphate (HS) mi-metic PI-88, a sulphomannan oligosaccharide of low MW, effi-ciently reduced the cell-to-cell spread of herpes simplex virus(HSV). Yamada and co-workers reported that O-acylated carra-geenan oligosaccharides with different MW had increased anti-HIV activity by depolymerisation and sulphation (Yamada, Ogamo,Saito, Uchiyama, & Nakagawa, 2000; Yamada et al., 1997). More-over, the high-solubility of sulphated oligosaccharides at neutralpH enhanced their potential use as anti-viral drugs.

W. Wang et al. / Food Chemistry 133 (2012) 880–888 881
In our previous study, we found that low molecular weight car-rageenan oligosaccharide CO-1 could effectively inhibit IAV repli-cation in vitro, and CO-1 might inhibit a replication step in IAVlife cycle subsequent to viral internalisation but prior to virusrelease (Wang et al., 2011). To further investigate the structure–activity relationship of carrageenan oligosaccharides, different j-carrageenan oligosaccharides and their sulphated derivatives wereprepared, and their anti-IAV activities in vitro and in vivo werecompared in this paper. The results indicated that KCO and its sul-phated derivative KCO-S could effectively inhibit IAV multiplica-tion in MDCK cells in a dose-dependent manner. Moreover, bothKCO and KCO-S could inhibit IAV infection and attenuate pneumo-nia in IAV-infected mice. The antiviral effect of KCO and KCO-Sseemed not to be dependent on the interferon system.
2. Materials and methods
2.1. Compounds and reagents
j-Carrageenan was purchased from Dehui Halobios (Qingdao,China). Ribavirin (50 mg/mL) was purchased from LuKang Cisen(Jining, China). Bio-Gel P-6 (fine) was purchased from Bio-Rad(Hercules, CA). Dextran standards (MW: 180, 1200, 2500, 4600,7100, 10,000, 11,700 Da) were obtained from Sigma–Aldrich (St.Louis, MO). RPM1640 medium, fetal bovine serum (FBS), penicillin,and streptomycin were purchased from Gibco (Grand Island, NY).20-(4-Methylumbelliferyl)-a-D-N-acetylneuraminic acid sodiumsalt hydrate (MUNANA) and 2-(N-morpholino)ethanesulphonicacid (MES) were purchased from Sigma (St. Louis, MO). IFN-c ELISAkit was obtained from Jingtian Biotech (Shanghai, China). Other re-agents were of analytical grade.
2.2. Preparation of j-carrageenan oligosaccharides and theirsulphated derivatives
The low-molecular-weight carrageenan oligosaccharides KCO,KCO-1 and KCO-2 were prepared and depolymerised by mild acidhydrolysis of j-carrageenan followed by fractionation using chro-matographic separation (Yang et al., 2009). In brief, the j-carra-geenan powder dissolved in distilled water was hydrolysed bysulphuric acid at pH 4.0, 60 �C for 1.5, 3.5, and 5 h, respectively.Mixtures were separated using a Bio-Gel P-6 column(4.6 � 120 cm). Samples were eluted at a flow rate of 6 mL/h in0.2 M ammonium bicarbonate, and 1-mL fractions were collected.Fractions were analysed by the carbazole reagent method (Bitter &Muir, 1962), and peaks were pooled and freeze-dried for furtheranalysis.
Sulphated carrageenan oligosaccharides were prepared by themethods described before (Yuan et al., 2005). In brief, 1 g j-carra-geenan oligosaccharides were dissolved in 50 mL DMF by stirringat 30 �C in a water bath for 30 min. Then an equal volume of sul-phur trioxide–pyridine complex reagent (molar ratio of 6:1) wasadded and incubated at 30 �C for 4 h. After that, the reaction solu-tion was precipitated by 80% ethanol–acetone (1:1), and centri-fuged to remove supernatant. The precipitate was washed withethanol, dialysed (100 Da MWCO, Spectrum, Rancho Dominguez,CA) for 24 h, and then freeze-dried. The sulphur content of carra-geenan oligosaccharides and their sulphated derivatives was mea-sured by the barium chloride–gelatin method of Dodgson and Price(1962).
2.3. FT-IR and NMR analysis
The Fourier-transform infrared spectroscopy (FT-IR) and nuclearmagnetic resonance spectroscopy (NMR) analysis of carrageenan
oligosaccharides and their sulphated derivatives were performedaccording to the methods described previously (Yuan et al., 2005).For FT-IR analysis, the samples (1–2 mg) were dried in a P2O5
desiccator for 48 h, then mixed with 100 mg KBr and pressed under7 kg/cm2 to make a transparent film. The film was put into an FT-IRinstrument (Nicolet Nexus 470, Thermo Electron, Waltham, MA)and scanned from 400 to 4000 cm�1.
For NMR analysis, 20 mg KCO or KCO-S was exchanged withdeuterium by lyophilising with 0.5 mL D2O (99.96%) three times.High-resolution 13C NMR of KCO and KCO-S was recorded on a600 MHz NMR spectrometer (JNM-ECA600, JEOL, Tokyo, Japan) at20 �C, using acetone-d6 as the internal standard (dC = 29.5 ppm).
2.4. Cell culture and virus infection
Madin–Darby canine kidney (MDCK) cells were obtained fromCell Bank of Chinese Academy of Sciences (Shanghai, China) andgrown in RPM1640 medium supplemented with 10% FBS,100 units/mL of penicillin and 100 lg/mL of streptomycin. Influ-enza virus (A/Puerto Rico/8/34 [H1N1]; PR/8) was propagated in10-day-old embryonated eggs for 3 days at 36.5 �C.
The virus infection experiments in vitro were performed as de-scribed previously (Wang et al., 2008, 2011). In brief, virus propa-gation solution was diluted in PBS containing 0.2% bovine serumalbumin and was added to cells at the indicated multiplicity ofinfection (MOI). Virus was allowed to adsorb for 60 min at 4 �C.After removing the virus inoculum, cells were maintained in infect-ing media (RPM1640, 4 lg/mL trypsin) at 37 �C in 5% CO2.
2.5. Cytotoxicity assays
The cytotoxicity of compounds was measured by the MTT(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide;Sigma–Aldrich) assay as described previously (Talarico & Damonte,2007; Wang et al., 2011). Confluent MDCK cell cultures in 96-wellplates were exposed to different concentrations of compounds intriplicate, and using incubation conditions equivalent to those usedin the antiviral assays. Next, 10 lL of PBS containing MTT (finalconcentration: 0.5 mg/mL) were added to each well. After 4 h incu-bation at 37 �C, the supernatant was removed and 200 lL of DMSOwere added to each well to solubilise the formazan crystals. Aftervigorous shaking, absorbance values were measured in a micro-plate reader (Bio-Rad) at 570 nm. The CC50 was calculated as thecompound concentration necessary to reduce cell viability by 50%.
2.6. Infectivity antiviral assays
For the determination of antiviral activity, a virus yield reduc-tion assay was performed, as previously described (Leibbrandtet al., 2010). MDCK cell monolayers were firstly incubated withIAV at an MOI of 1.0 for 1 h at 4 �C, then after the virus was re-moved, the infecting media containing different drugs were addedto cells and incubated at 37 �C for 48 h. Then cell supernatantswere collected and the virus yields were determined in MDCK cellsby 50% tissue culture infectious dose (TCID50) assay. Virus titres inTCID50/mL were determined according to Reed and Muench(1938).
Furthermore, the antiviral activity was also evaluated by thecytopathic effects (CPE) inhibition assay as described previously(Hung et al., 2009). MDCK cell cultures in 96-well plates werefirstly infected with IAV (MOI = 1.0), and then treated with differ-ent compounds in triplicate after removal of the virus inoculum.After 48 h incubation, the cells were fixed with 4% formaldehydefor 20 min at room temperature. After removal of the formalde-hyde, the cells were stained with 0.1% crystal violet for 30 min.The plates were washed and dried, and the intensity of crystal

882 W. Wang et al. / Food Chemistry 133 (2012) 880–888
violet staining for each well was measured at 570 nm. The virusinhibition (%) was calculated by the equation:
Virus inhibition ð%Þ ¼ ½ðAsample 570 � Avirus 570Þ=ðAmock 570
� Avirus 570Þ� � 100;
where, Amock 570 was the absorbance without virus infection, Asample
570 was absorbance with virus infection and drug treatment, Avirus
570 was absorbance with virus infection but without drugs.
2.7. Haemagglutination (HA) assay
The haemagglutination (HA) assay was performed as previouslyreported (Sriwilaijaroen et al., 2011). Standardised chicken redblood cell (cRBC) solutions were prepared according to the WHOmanual. Virus propagation solutions were serially diluted 2-foldin round-bottomed 96-well plate and 1% cRBCs were then addedat an equal volume. After 60 min incubation at 4 �C, RBCs in nega-tive wells precipitated and formed red buttons, whereas positivewells had an opaque appearance with no sedimentation. HA titresare given as haemagglutination U/mL (HAU/mL).
2.8. In vivo experiments
Four-week-old female Kunming mice (average weight,16.0 ± 2.0 g) were housed and studied under protocols approvedby the Animal Ethics Committee of Ocean University of China.Briefly, mice were randomly divided into different experimentalgroups (10 mice/group). The drugs administration started 1 dayprior to virus inoculation. Virus-infected control group (virus con-trol) and uninfected control group (normal control) received1 � PBS as a placebo. On the day of virus inoculation, mice werelightly anaesthetised and each was inoculated intranasally withPR8 virus (4 HAU/mouse) diluted in 40 lL of 1 � PBS. Two hoursafter inoculation, mice received intraperitoneal injections of carra-geenan oligosaccharides or ribavirin (40 or 80 mg/kg/day), and thetreatments were repeated once daily for the entire experiment.
Mice were weighed and killed on Day 6 after inoculation, andlungs were removed and weighed. The lung index was calculatedby the following equation using the obtained values:
Lung index ¼ ½Lung weight ðgÞ=Mice weight ðgÞ� � 100:
By this index, the severity of lung injury in pneumonia mice wasevaluated. Subsequently, lung specimens of animals from eachexperimental group were homogenised in serum-free RPMI 1640medium for determination of viral titres.
In the survival experiments, 10 mice per group were intrana-sally infected with PR/8 virus (6 HAU/mouse) at Day 0. IAV-infected mice received intraperitoneal injections of carrageenanoligosaccharides or ribavirin (40 mg/kg/day), and the virus controlgroup and normal control group received PBS as a placebo. Thedrugs administration started 1 day prior to virus infection andwas repeated once daily during the course of the experiment,and survival was assessed in all groups for 14 days after infection.
2.9. Neuraminidase activity assay
Neuraminidase activity assay was performed as described pre-viously (Eichelberger, Hassantoufighi, Wu, & Li, 2008; Nayak &Reichl, 2004) with some modifications. The lung tissue sampleswere firstly snap-frozen upon dissection and subsequently homog-enised in serum-free RPM1640 medium. After centrifugation oflung homogenate, the supernatant (30 lL) of each sample wastransferred to a black 96-well plate, and 70 lL of reaction buffer(33 mM MES buffer (pH 3.5); 4 mM CaCl2; 20 lM MUNANA) were
added. After incubation of the plate at 37 �C for 45 min, 100 lL stopsolution (25% ethanol, 0.1 M glycine, pH 10.7) were added to eachwell and fluorescence intensity of the product 4-MU was measuredusing a SpectraMax M5 plate reader (Molecular Devices, Sunny-vale, CA) with excitation and emission wavelengths of 360 and440 nm, respectively. The mean fluorescence intensity of eachsample is expressed as relative fluorescence units (RFU), which isused to evaluate the amount of IAV particles.
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
The content of interferon-c (IFN-c) in the supernatants ofspleen homogenate was determined by ELISA assay using a com-mercial ELISA kit (Jingtian Biotech, Shanghai, China). In brief, thespleen tissue samples were firstly snap-frozen upon dissectionand subsequently homogenised in 1 � PBS (PH 7.4). After centrifu-gation of spleen homogenate, the supernatant of each sample, po-sitive control and standards diluted in diluent/lysis buffer wereadded to individual wells (50 lL/well), and incubated at 37 �C for30 min with immobilised monoclonal antibody directed againstmouse interferon-c (IFN-c), then were washed three times withwash buffer. Samples were then incubated with horseradish perox-idase (HRP) labelled monoclonal anti-IFN-c antibodies for 30 minat 37 �C. Following three washes, wells were incubated with thesubstrate solution consisting of hydrogen peroxide and tetrameth-ylbenzidine at room temperature for 15 min. The reaction wasstopped by the addition of stop solution (1 N H2SO4) prior to absor-bance reading at 450 nm using a Rainbow microplate reader(Tecan Group Ltd., Männedorf, Switzerland).
2.11. Statistics
All values are presented as mean ± S.D. Statistical significancewas evaluated by unpaired Student’s t-test for comparison ofmeans. Differences were considered statistically significant atp < 0.05.
3. Results and discussion
3.1. Characterisation of j-carrageenan oligosaccharides and theirsulphated derivatives
Our previous study reported that low molecular weightcarrageenan oligosaccharide CO-1 (�2 kDa) effectively inhibitedIAV replication in vitro and its activity was better than that of 3and 5 kDa carrageenan oligosaccharides (Wang et al., 2011). Tofurther explore the structure–activity relationship of carrageenanoligosaccharides, different j-carrageenan oligosaccharides KCO,KCO-1, KCO-2 and their derivatives KCO-S, KCO-S-1, KCO-DS wereprepared. The molecular weights of j-carrageenan oligosaccha-rides KCO, KCO-1 and KCO-2 were determined by high-perfor-mance gel permeation chromatography (HPGPC) as describedbefore (Yamamoto, Nunome, Yamauchi, Kato, & Sone, 1995). Theaverage molecular weight of KCO was about 2.1 kDa, which wasslightly higher than that of CO-1 (MW = 1.7 kDa) in the previousstudy (Wang et al., 2011). The degree of polymerisation (DP) ofKCO was about 3–17, according to the results of HPTLC and PAGEanalysis (Fig. 1B–D). In contrast, the molecular weights of KCO-1and KCO-2 was about 1 and 4 kDa, respectively. The DP of KCO-1was about 3–9 according to the results of HPTLC and PAGE analysis(Fig. 1B and C).
Moreover, j-carrageenan oligosaccharides KCO, KCO-1 andKCO-2 are all odd-numbered oligosaccharides, and all composedby 1,4-linked b-D-3,6-anhydro-galactose (AnG) and 1,3-linked4-O-sulphated-a-D-galactose (G4S) (Fig. 1A) (Yang et al., 2009;
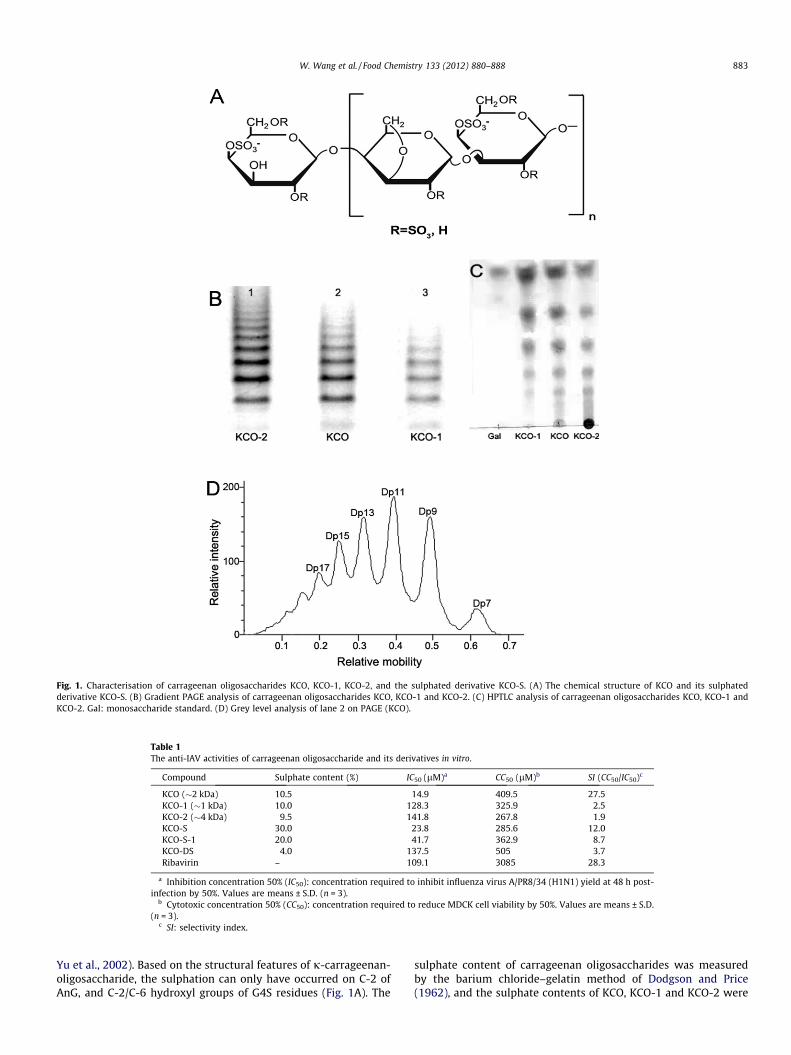
Fig. 1. Characterisation of carrageenan oligosaccharides KCO, KCO-1, KCO-2, and the sulphated derivative KCO-S. (A) The chemical structure of KCO and its sulphatedderivative KCO-S. (B) Gradient PAGE analysis of carrageenan oligosaccharides KCO, KCO-1 and KCO-2. (C) HPTLC analysis of carrageenan oligosaccharides KCO, KCO-1 andKCO-2. Gal: monosaccharide standard. (D) Grey level analysis of lane 2 on PAGE (KCO).
Table 1The anti-IAV activities of carrageenan oligosaccharide and its derivatives in vitro.
Compound Sulphate content (%) IC50 (lM)a CC50 (lM)b SI (CC50/IC50)c
KCO (�2 kDa) 10.5 14.9 409.5 27.5KCO-1 (�1 kDa) 10.0 128.3 325.9 2.5KCO-2 (�4 kDa) 9.5 141.8 267.8 1.9KCO-S 30.0 23.8 285.6 12.0KCO-S-1 20.0 41.7 362.9 8.7KCO-DS 4.0 137.5 505 3.7Ribavirin – 109.1 3085 28.3
a Inhibition concentration 50% (IC50): concentration required to inhibit influenza virus A/PR8/34 (H1N1) yield at 48 h post-infection by 50%. Values are means ± S.D. (n = 3).
b Cytotoxic concentration 50% (CC50): concentration required to reduce MDCK cell viability by 50%. Values are means ± S.D.(n = 3).
c SI: selectivity index.
W. Wang et al. / Food Chemistry 133 (2012) 880–888 883
Yu et al., 2002). Based on the structural features of j-carrageenan-oligosaccharide, the sulphation can only have occurred on C-2 ofAnG, and C-2/C-6 hydroxyl groups of G4S residues (Fig. 1A). The
sulphate content of carrageenan oligosaccharides was measuredby the barium chloride–gelatin method of Dodgson and Price(1962), and the sulphate contents of KCO, KCO-1 and KCO-2 were

884 W. Wang et al. / Food Chemistry 133 (2012) 880–888
all about 10% (Table 1). The degree of substitution (DS) of sulphatein KCO-S is about 1.26 while that of the desulphated oligosaccha-ride KCO-DS is about 0.17.
The FT-IR spectra of KCO and KCO-S looked similar to that ofnatural j-carrageenan (KC) (Fig. 2A). The absorption bands at845, 930 and 1250 cm�1 were the characteristic absorption peaksof 4-sulphate-b-D-galactose, a-D-3,6-anhydrogalactose and sul-phate, respectively. The absorption band at 1200–1000 cm�1 wasthe characteristic absorption peak of pyranose ring ether linkage.All the results indicated that the chain structure of the polysaccha-rides was not destroyed after depolymerisation and modification.Moreover, KCO-S showed a stronger absorption at 1250 cm�1 thanthat of KCO and KC, indicating more sulphate content in KCO-S.
Fig. 2. IR and 13C NMR spectrum of carrageenan oligosaccharide KCO and its derivative Kscanned from 4000 to 400 cm�1 under dry air at RT. (B and C) Carrageenan oligosaccharidlinked-4-sulphated-galactose; A1–A6: six-carbon resonance signals of 1,4-linked-3,6-an
3.2. 13C NMR analysis of KCO and KCO-S
To further confirm the structure of KCO, the 13C NMR analysiswas performed and the results were shown in Fig. 2B. The 13CNMR-spectrum of KCO showed all 12 carbon resonance signals ofj-carrageenan repeating disaccharide as reported before (Yuet al., 2002; Zhang et al., 2010), which suggested that the chemicaldegradation only led to the cleavage of glycosidic bond and did notdestroy the repeating disaccharide structure.
To determine the location of sulphate substitution in KCO-S, acomplete assignment of the 13C NMR spectral signals of KCO andKCO-S were compared according to the previous data reported(Yu et al., 2002; Yuan et al., 2005). The position of sulphation
CO-S. (A) j-carrageenan (KC), carrageenan oligosaccharide KCO and KCO-S spectrae KCO (B) and its derivative KCO-S (C). G1–G6: six-carbon resonance signals of 1,3-
hydrogalactose; A20 , G20: substitution position of sulphate group.

W. Wang et al. / Food Chemistry 133 (2012) 880–888 885
was determined by analysis of the 13C NMR spectra (Fig. 2B and C).Downfield displacements of the signals for C-2 of G4S to d75.4 andC-2 of the AnG to d77.7, as compared with their positions in thecorresponding non-substituted oligosaccharide at d69.7, and 70.7,respectively, confirmed the substitution pattern in KCO-S (Melo,Feitosa, Freitas, & Paula, 2002). The existence of strong signalsand the disappearance of unmodified signals indicated that the hy-droxyl groups at C-2 of the G4S residue were completely sulphated(Fig. 2B and C). The significant signals at d70.7 for the unsubstitut-ed C-2 of the AnG residue and d77.7 for the sulphated C-2 of AnGwere observed, indicating that the C-2 hydroxyl groups of AnG res-idue were partially sulphated in KCO-S.
Fig. 3. The inhibition effect of carrageenan oligosaccharide KCO and its derivatives on(MOI = 1.0). After infection the cells were washed and medium containing KCO was addecrystal violet staining solution. NI: cells without virus infection; PR8: virus infected cellsindicated concentrations of KCO or KCO-S after removed virus. The virus titres in cell supSignificance: ⁄p < 0.05, ⁄⁄p < 0.01 vs. virus control group. (C) IAV (MOI = 1.0) infected MDremoval of the virus inoculum. After 48 h, the antiviral effect was evaluated by CPE inhibPR8 virus (MOI = 1.0), and then treated with indicated concentrations of KCO, KCO-S, KCOby CPE inhibition assay at 48 h p.i. Values are means ± S.D. (n = 3).
3.3. Inhibition of influenza A virus multiplication in vitro by KCO andKCO-S
Carrageenan oligosaccharides and their derivatives were as-sayed for their ability to inhibit IAV multiplication in vitro usinghaemagglutination (HA) assay and virus yield reduction assay,as previously described (Leibbrandt et al., 2010; Sriwilaijaroenet al., 2011). In brief, MDCK cells were infected with A/PR/8/34virus at an MOI of 1.0, and then incubated with or without com-pounds after virus adsorption. After 48 h, the virus titres in cellsupernatants were measured by HA assay and the cell viabilitywas evaluated by crystal violet stain. As shown in Fig. 3A, the
IAV replication in vitro. (A) MDCK cells in 24-well plates were infected with IAVd at the indicated concentrations. After 48 h, living cells were fixed and stained withwithout drug treatment. (B) IAV (MOI = 1.0) infected MDCK cells were treated withernatants were determined by HA assay at 48 h p.i. Values are means ± S.D. (n = 4).CK cells were treated with indicated concentrations of KCO, KCO-1 or KCO-2 after
ition assay. Values are means ± S.D. (n = 3). (D) MDCK cells were firstly infected with-S-1 or KCO-DS after removed virus inoculum. The antiviral activity was determined

Fig. 4. The therapeutic effects of KCO and KCO-S on influenza A virus-infected mice.(A) Viral titres in lungs evaluated by performing neuraminidase activity assay. Themean fluorescence intensity of each sample is shown as relative fluorescence units(RFU). Values are means ± S.D. (n = 3). Significance: ##p < 0.05, ##p < 0.01 vs. normalcontrol group; ⁄p < 0.05, ⁄⁄p < 0.01 vs. virus control group (placebo). (B) Survivalrate. IAV infected mice received intraperitoneal therapy once daily with drugs orplacebo for the entire experiment. Results are expressed as percentage of survival,evaluated daily for 15 days. Significance: ⁄p < 0.05, ⁄⁄p < 0.01 vs. control group(placebo). (C) IFN-c production determined by ELISA assay. Significance: ##p < 0.01vs. normal control group; ⁄p < 0.05 vs. virus control group (placebo).
886 W. Wang et al. / Food Chemistry 133 (2012) 880–888
j-carrageenan-derived oligosaccharide KCO could effectively re-duce the cytopathic effect of the PR/8 virus and promote survivalof IAV-infected MDCK cells when used at the concentration ofmore than 25 lM. Moreover, KCO could also significantly reducethe virus HA titre when used at a concentration of more than50 lM (p < 0.05), and the virus inhibition was dose-dependent atthe concentrations of 12.5–100 lM (Fig. 3B). The IC50 value of oli-gosaccharide KCO was only 14.9 lM, and the selectivity index(CC50/IC50) for KCO was more than 27.0 (SI = 27.5) (Table 1). Fur-thermore, the sulphated derivative KCO-S could also effectivelyinhibit PR8 virus multiplication when used at a concentration ofmore than 50 lM (Fig. 3B), and the IC50 of KCO-S was only about23.8 lM, which was lower than that of ribavirin (IC50 = 109.1 lM)(Table 1). These results indicated that both carrageenan oligosac-charide KCO and its sulphated derivative KCO-S could all effec-tively inhibit the replication of influenza A virus in vitro.
3.4. The structure–activity relationship of j-carrageenanoligosaccharides
The influence of molecular weight on the anti-IAV activities ofj-carrageenan oligosaccharides was firstly investigated by per-forming CPE inhibition assay as previously described (Hung et al.,2009). As shown in Fig. 3C, KCO-treated cells showed betterprotection from virus-induced cell death than cells treated withcarrageenan oligosaccharide KCO-2 with high molecular weight(�4 kDa) at all concentrations tested (6.25–100 lM), which wasin agreement with the results reported previously (Wang et al.,2011). Moreover, the CPE inhibition percent of KCO-1 with verylow molecular weight (�1 kDa) was also lower than that of KCO(�2 kDa) (Fig. 3C). The selectivity index (CC50/IC50) for KCO-1 andKCO-2 was no more than 3.0 (Table 1), which suggested that sugarchains of a certain length might be essential for the anti-IAV activityof j-carrageenan oligosaccharide. Combined with the results of CO-1 reported previously (Wang et al., 2011), we suppose that the mostactive j-carrageenan oligosaccharide should have a molecularweight of about 2 kDa (1–3 kDa).
Furthermore, the antiviral effect of KCO derivatives with differ-ent sulphate contents was also evaluated, to explore the relation-ship between the degree of sulphation and anti-IAV activity. Thesulphate content of these oligosaccharides was measured quantita-tively with chemical gelatin nephelometry (Table 1). As shown inFig. 3D, the protective effect of the sulphated derivatives KCO-Sand KCO-S-1 was all lower than that of KCO at concentrations from6.25 to 100 lM. Moreover, the IC50 values of KCO-S and KCO-S-1were about 23.8 and 41.7 lM, respectively, and the selectivity in-dex (CC50/IC50) for them was no more than 12.0 (Table 1). Further-more, the CPE inhibition percent of the desulphated derivativeKCO-DS was no more than 40% at 100 lM, which was much lessthan that of KCO (>80%) (Fig. 3D). The IC50 of KCO-DS was onlyabout 137.5 lM, and the selectivity index was only 3.7 (Table 1).These results suggested that the anti-IAV activities of j-carra-geenan oligosaccharides were associated with their sulphate con-tents but did not increase simply with increases in the degree ofsulphation, and the optimal sulphate content required for anti-IAV action was about 0.8–1.0 mole/mole of disaccharide.
In conclusion, the degree of sulphation and the molecularweight were major factors that influenced the anti-IAV activity ofj-carrageenan oligosaccharide. The most active j-carrageenan oli-gosaccharide (KCO) was found to have a definite range of sulphatecontents (0.8–1.0 mole/mole of disaccharide) and molecularweights (1–3 kDa). Moreover, we also found marked difference be-tween the anti-IAV neuraminidase activities of i-carrageenan oli-gosaccharide and the sulphated j-carrageenan oligosaccharidewith similar sulphate content (data not shown), which suggestedthat not only the sulphate content but also the positions of the
sulphate groups were important for the anti-IAV actions of carra-geenan oligosaccharides.
3.5. Effects of KCO and its sulphated derivative KCO-S on IAV infectionin vivo
The inhibitory effects of KCO and KCO-S on IAV infection in vivowere also investigated. In brief, IAV-infected mice received intra-peritoneal injections of drugs (40 mg/kg/day) or placebo (PBS) oncedaily for the entire experiment, and then sacrificed at 4 and 6 daysp.i., respectively. Subsequently, the pulmonary viral titres weredetermined by performing neuraminidase activity assay asdescribed previously (Eichelberger et al., 2008; Nayak & Reichl,2004). As shown in Fig. 4A, the mean fluorescence intensity of

Table 2The anti-IAV activities of carrageenan oligosaccharide and its derivatives in vivo.
Groups Dose (mg/kg/day) Body weight (g) Lung indexa (X ± S.D.) Inhibitory rate (%)
Virus control – 16.88 ± 1.15## 1.83 ± 0.39## 0Normal control – 24.05 ± 1.66 0.73 ± 0.09 –Ribavirin 40 20.12 ± 3.63* 1.10 ± 0.37* 39.9Ribavirin 80 20.02 ± 2.28* 0.97 ± 0.17** 47.0KCO 40 22.23 ± 4.43** 0.96 ± 0.34** 47.5KCO 80 20.97 ± 2.91* 1.12 ± 0.45* 38.8KCO-S 40 21.73 ± 1.45** 1.00 ± 0.16** 45.4KCO-S 80 20.28 ± 2.70* 1.09 ± 0.10* 40.4
Values are means ± S.D. (n = 10).## p < 0.01 vs. normal control group; t-test.
* p < 0.05 vs. virus control group; t-test.** p < 0.01 vs. virus control group; t-test.
a Lung index = [lung weight (g)/mice weight (g)] � 100.
W. Wang et al. / Food Chemistry 133 (2012) 880–888 887
samples in KCO and KCO-S treated groups was significantly lowerthan that of the control group at 4 days p.i. (p < 0.01), which sug-gested that the total NA activity in lungs was reduced by KCOand KCO-S treatment. Moreover, there was also shown a significantreduction of NA activity in the lungs of KCO and KCO-S-treatedmice 6 days post infection, as compared to the control group(p < 0.05) (Fig. 4A). It was reported that approximately 50–100neuraminidase (NA) molecules are incorporated into each virionand its activity has previously been used to quantify influenzavirus (Nayak & Reichl, 2004). The data above suggest thatintraperitoneal therapy of infected mice with KCO and KCO-Sresults in a reduction of viral titres in the lung. Moreover, KCO alsosignificantly reduced the lung index in IAV-infected mice with aninhibition rate of 47.5% when used at a dose of 40 mg/kg/day(Table 2), which suggested that KCO treatment attenuated pneu-monia symptoms in mice. Besides that, KCO and KCO-S could sig-nificantly prevent the weight loss of IAV-infected mice comparedto the virus control group (p < 0.05) (Table 2).
Furthermore, influenza A virus can induce lethal infections incertain mouse strains usually within 2 weeks (Barnard, 2009), sothe survival experiments were also performed to evaluate the ef-fect of KCO and KCO-S on the survival of IAV-infected mice. Asshown in Fig. 4B, if treatment with KCO or ribavirin at the doseof 40 mg/kg/day was started 1 day before infection and repeatedevery 24 h for up to 15 days p.i., we noticed significantly increasedsurvival rates in the KCO and ribavirin-treated groups, as comparedto the placebo-treated control group (p < 0.01). In the particularexperiment shown in Fig. 4B, by Day 15 after infection, only 30%of the individuals in the placebo group survived whereas 90% ofthose in the KCO treated group survived. The survival rate was alsosignificantly increased by KCO-S treatment (about 70%), as com-pared to the placebo-treated control group (p < 0.05).
Moreover, ELISA assay of IFN-c was also performed to investi-gate whether the antiviral effect of KCO and KCO-S is associatedwith the interferon system or not. As shown in Fig. 4C, the produc-tion of IFN-c significantly decreased after IAV infection comparedto that of the normal control group (p < 0.01). After treatment withribavirin for 6 days, the production of IFN-c increased a little com-pared to that of the virus control group (placebo). However, KCOtreatment for 6 days could not significantly increase the produc-tion of IFN-c (p > 0.05), which suggested that KCO did not directlyinduce the production of IFN-c to inhibit IAV infection. The produc-tion of IFN-c was slightly decreased by KCO-S treatment comparedto that of the virus control group (Fig. 4C). Furthermore, KCO andKCO-S all had similar antiviral effects on IAV infection of MDCKcells and interferon-deficient Vero cells (data not shown). Takentogether, the antiviral effect of KCO and KCO-S seems not to bedependent on the interferon system.
In conclusion, all these results suggested that KCO and its sulph-ated derivative KCO-S could effectively promote the survival of
IAV-infected mice by directly reducing the pulmonary viral titres,and their antiviral effect seems not to be dependent on the inter-feron system.
4. Conclusions
Low-molecular-weight j-carrageenan oligosaccharide KCO andits sulphated derivative KCO-S could effectively inhibit IAV infec-tion in MDCK cells in a dose-dependent manner. The structure–activity relationship study showed that the specific sugar linkage,a certain sugar length and sulphate content might be essentialfor the anti-IAV activity of j-carrageenan oligosaccharides. Themost active j-carrageenan oligosaccharide had a sulphate contentof 0.8–1.0 mole/mole of disaccharide and a molecular weight of 1–3 kDa. Furthermore, KCO and its derivative KCO-S could signifi-cantly improve survival and decrease pulmonary viral titres inIAV-infected mice. Moreover, the antiviral effect of KCO andKCO-S seems not to be dependent on the interferon system.
In summary, j-Carrageenan oligosaccharide KCO and its sulph-ated derivative KCO-S have good inhibitory effects on IAV replica-tion in vitro and in vivo, which suggests that KCO and KCO-S meritfurther investigation as novel antiviral agents to prevent IAVinfection.
Acknowledgements
This work was supported in part by the Program for ChangjiangScholars and Innovative Research Team in University (IRT0944),National Natural Science Foundation of China (31070724,31000830), The Fundamental Research Funds for the Central Uni-versities (201113013), Shandong Provincial Natural Science Foun-dation (ZR2011HQ012), and Special Fund for Marine ScientificResearch in the Public Interest (201005024).
References
Barnard, D. L. (2009). Animal models for the study of influenza pathogenesis andtherapy. Antiviral Research, 82, A110–A122.
Bitter, M., & Muir, H. (1962). A modified uronic acid reaction. AnalyticalBiochemistry, 4, 330–334.
Buck, C. B., Thompson, C. D., Roberts, J. N., Muller, M., Lowy, D. R., & Schiller, J. T.(2006). Carrageenan is a potent inhibitor of papillomavirus infection. PLoSPathogens, 2, e69.
Dodgson, K. S., & Price, R. G. (1962). A note on the determination of the estersulphate content of sulphated polysaccharides. Biochemical Journal, 84,106–110.
Eichelberger, M. C., Hassantoufighi, A., Wu, M., & Li, M. (2008). Neuraminidaseactivity provides a practical read-out for a high throughput influenza antiviralscreening assay. Virology Journal, 5, 109.
Ekblad, M., Adamiak, B., Bergstrom, T., Johnstone, K. D., Karoli, T., Liu, L., et al. (2010).A highly lipophilic sulfated tetrasaccharide glycoside related to muparfostat (PI-88) exhibits virucidal activity against herpes simplex virus. Antiviral Research,86, 196–203.

888 W. Wang et al. / Food Chemistry 133 (2012) 880–888
Grassauer, A., Weinmuellner, R., Meier, C., Pretsch, A., Prieschl-Grassauer, E., &Unger, H. (2008). Iota-carrageenan is a potent inhibitor of rhinovirus infection.Virology Journal, 5, 107.
Hay, A. J., Wolstenholme, A. J., Skehel, J. J., & Smith, M. H. (1985). The molecularbasis of the specific anti-influenza action of amantadine. EMBO Journal, 4,3021–3024.
Hayden, F. G., & Pavia, A. T. (2006). Antiviral management of seasonal and pandemicinfluenza. Journal of Infectious Diseases, 194(Suppl. 2), S119–S126.
Hung, H. C., Tseng, C. P., Yang, J. M., Ju, Y. W., Tseng, S. N., Chen, Y. F., et al. (2009).Aurintricarboxylic acid inhibits influenza virus neuraminidase. AntiviralResearch, 81, 123–131.
Ji, J., Wang, L. C., Wu, H., & Luan, H. M. (2011). Bio-function summary of marineoligosaccharides. International Journal of Biological Sciences, 3, 74–86.
Lamb, R. A., & Takeda, M. (2001). Death by influenza virus protein. Nature Medicine,7, 1286–1288.
Leibbrandt, A., Meier, C., König-Schuster, M., Weinmüllner, R., Kalthoff, D.,Pflugfelder, B., et al. (2010). Iota-carrageenan is a potent inhibitor of influenzaA virus infection. PLoS One, 5, e14320.
Melo, M. R. S., Feitosa, J. P. A., Freitas, A. L. P., & Paula, R. C. M. (2002). Isolation andcharacterization of soluble sulfated polysaccharide from the red seaweedGracilaria cornea. Carbohydrate Polymers, 49, 491–498.
Mendel, D. B., Tai, C. Y., Escarpe, P. A., Li, W. X., Sidwell, R. W., Huffman, J. H., et al.(1998). Oral administration of a prodrug of the influenza virus neuraminidaseinhibitor GS 4071 protects mice and ferrets against influenza infection.Antimicrobial Agents and Chemotherapy, 42, 640–646.
Nayak, D. P., & Reichl, U. (2004). Neuraminidase activity assays for monitoringMDCK cell culture derived influenza virus. Journal of Virology Methods, 122,9–15.
Nguyen, T. N. A., Dao, T. T., Tung, B. T., Choi, H., Kim, E., Park, J., et al. (2011).Influenza A (H1N1) neuraminidase inhibitors from Vitis amurensis. FoodChemistry, 124, 437–443.
Pujol, C. A., Scolaro, L. A., Ciancia, M., Matulewicz, M. C., Cerezo, A. S., & Damonte, E.B. (2006). Antiviral activity of a carrageenan from Gigartinatina skottsbergiiagainst intraperitoneal murine herpes simplex virus infection. Planta Medica,72, 121–125.
Reed, L., & Muench, H. (1938). A simple method for estimating 50% endpoints.American Journal of Hygiene, 27, 943–947.
Sriwilaijaroen, N., Kadowaki, A., Onishi, Y., Gato, N., Ujike, M., Odagiri, T., et al.(2011). Mumefural and related HMF derivatives from Japanese apricot fruit
juice concentrate show multiple inhibitory effects on pandemic influenza A(H1N1) virus. Food Chemistry, 127, 1–9.
Talarico, L. B., & Damonte, E. B. (2007). Interference in dengue virus adsorption anduncoating by carrageenans. Virology, 363, 473–485.
Talarico, L. B., Noseda, M. D., Ducatti, D. R. B., Duarte, M. E., & Damonte, E. B. (2011).Differential inhibition of dengue virus infection in mammalian and mosquitocells by iota-carrageenan. Journal of General Virology, 92, 1332–1342.
Wang, W., Cui, Z.-Q., Han, H., Zhang, Z.-P., Wei, H.-P., Zhou, Y.-F., et al. (2008).Imaging and characterizing influenza A virus mRNA transport in living cells.Nucleic Acids Research, 36, 4913–4928.
Wang, W., Zhang, P., Hao, C., Zhang, X.-N., Cui, Z.-Q., & Guan, H.-S. (2011). In vitroinhibitory effect of carrageenan oligosaccharide on influenza A H1N1 virus.Antiviral Research, 92, 237–246.
Yamada, T., Ogamo, A., Saito, T., Uchiyama, H., & Nakagawa, Y. (2000). Preparation ofO-acylated low-molecular-weight carrageenans with potent anti-HIV activityand low anticoagulant effect. Carbohydrate Polymer, 41, 115–120.
Yamada, T., Ogamo, A., Saito, T., Watanabe, J., Uchiyama, H., & Nakagawa, Y. (1997).Preparation and anti-HIV activity of low-molecular-weight carrageenans andtheir sulfated derivatives. Carbohydrate Polymer, 32(1), 51–55.
Yamamoto, Y., Nunome, T., Yamauchi, R., Kato, K., & Sone, Y. (1995). Structure of anexocellular polysaccharide of Lactobacillus helveticus TN-4, a spontaneousmutant strain of Lactobacillus helveticus TY1-2. Carbohydrate Research, 275,319–332.
Yang, B., Yu, G. L., Zhao, X., Jiao, G. L., Ren, S. M., & Chai, W. G. (2009). Mechanism ofmild acid hydrolysis of galactan polysaccharides with highly ordereddisaccharide repeats leading to a complete series of exclusively odd-numbered oligosaccharides. FEBS Journal, 276, 2125–2137.
Yewdell, J., & Garcia-Sastre, A. (2002). Influenza virus still surprises. Current Opinionin Microbiology, 5, 414–418.
Yu, G. L., Guan, H. S., Ioanoviciu, A. S., Sikkander, S. A., Thanawiroon, C., Tobacman, J.K., et al. (2002). Structural studies on j-carrageenan derived oligosaccharides.Carbohydrate Research, 337, 433–440.
Yuan, H. M., Zhang, W. W., Li, X. G., Lü, X. X., Li, N., Gao, X. L., et al. (2005).Preparation and in vitro antioxidant activity of j-carrageenan oligosaccharidesand their oversulfated, acetylated, and phosphorylated derivatives.Carbohydrate Research, 340, 685–692.
Zhang, W., Liu, M., Hansen, P. E., Yu, G. L., Yang, B., & Zhao, X. (2010). Sequence andstructure analysis of j-carrageenan-derived oligosaccharides by 2D NMR.Journal of Phycology, 46, 831–838.






![The host immune response in respiratory virus infection ... · (IFN-α/β receptor), activating the JAK/STAT pathway [3]. ... croup, tonsilitis Seasonal influenza Sialic acids Fever,](https://static.fdocument.org/doc/165x107/5c7b1b0809d3f277748b45cf/the-host-immune-response-in-respiratory-virus-infection-ifn-receptor.jpg)











