
Original Article Resveratrol inhibits TNF-α-induced matrix ... · PDF fileResveratrol...
9
Int J Clin Exp Med 2017;10(2):2764-2772 www.ijcem.com /ISSN:1940-5901/IJCEM0038245 Original Article Resveratrol inhibits TNF-α-induced matrix degradation via the p38/MAPK and Akt pathways in human nucleus pulposus cells Hucheng Liu, Runsheng Guo, Jiangyinzi Shang, Bin Zhang, Min Dai Department of Orthopaedics, The First Affiliated Hospital, Nanchang University, Jiangxi, China Received August 19, 2016; Accepted November 29, 2016; Epub February 15, 2017; Published February 28, 2017 Abstract: Previous studies have proved that resveratrol has beneficial effects on intervertebral disc regeneration. Tumor necrosis factor-α (TNF-α) has been proven to induce matrix degradation, which plays a critical role in inter- vertebral disc degeneration (IVDD). However, the effect of resveratrol on TNF-α-induced matrix degradation remains unknown. The purpose of this study was to find out whether resveratrol can reverse TNF-α-induced matrix degrada- tion in human NP cells and to further study related mechanism involved in the process. The transcriptional activity, mRNA and protein expression of Collagen II and Aggrecan were measured by luciferase reporter assay, RT-qPCR and western blot. Sulfated glycosaminoglycan (sGAG) level was detected to show the production of proteoglycans in NP extracellular matrix. TNF-α reduced Collagen II and Aggrecan production of human NP cells time-dependently. Resveratrol can rescue TNF-α-induced downregulation of Collagen II and Aggrecan production. MAPK, NF-κB and Akt signal pathways were activated after treatment with TNF-α and resveratrol. P38 inhibitor or Akt inhibitor can abol- ished the antagonistic effect of resveratrol on TNF-α-induced catabolic response. Our findings provide novel data supporting the protective effect of resveratrol, which may be used as a therapeutic agent in the treatment of IVDD. Keywords: Resveratrol, tumor necrosis factor-α, matrix degradation, nucleus pulposus cell, signal pathway Introduction Low back pain is a common public health pro- blem worldwide, with a lifetime prevalence of 70-80% [1]. It significantly affects quality of life and burdens the health service to the detri- ment of all industrialized societies. The causes of back pain are multifactorial, intervertebral disc degeneration (IVDD) is one of the predomi- nant factors that leads to this disabling condi- tion [2-4]. Conservative treatments are current- ly aiming to alleviate the symptoms, while sur- geries involve excising the degenerated motion segment and fusion the adjacent vertebral bodies rather than to solve the underlying bio- logical causes involved in disc degeneration. There is a clear need for the development of novel therapeutic procedures that have the ability to interfere with the biological mecha- nism of IVDD. As an alternative to the surgical repair or removal of a diseased disc, biologic treatments to promote intervertebral disc (IVD) repair and restore physiologic function have been considered [5-12]. Resveratrol (trans-3,4’,5-trihydroxystilbene) was first identified in the roots of white hellebore in 1940, mainly found in the skins of grapes and red wines [13-15] and showed powerful biologi- cal functions of anti-inflammatory, antioxidant, cardioprotective, and antitumor properties [16- 24]. Resveratrol has also been reported to pro- vide a protective effect on articular cartilage in rabbit models of osteoarthritis and rheumatoid arthritis [25, 26]. More recently, resveratrol has been found to relieve matrix-degrading enzy- mes upregulation and to facilitate proteoglycan synthesis and accumulation in bovine disc cells [27], as well as to decrease pro-inflammatory cytokines and catabolic enzymes expression in human IVD cells [28]. However, it is not known if resveratrol exerts similar protective effects on TNF-α-induced matrix degradation in human nucleus pulposus (NP) cells. The purpose of this study was to elucidate whether resveratrol can reverse TNF-α-medi- ated matrix degradation in human NP cells and
-
Upload
dinhnguyet -
Category
Documents
-
view
224 -
download
1
Transcript of Original Article Resveratrol inhibits TNF-α-induced matrix ... · PDF fileResveratrol...

Int J Clin Exp Med 2017;10(2):2764-2772www.ijcem.com /ISSN:1940-5901/IJCEM0038245
Original ArticleResveratrol inhibits TNF-α-induced matrix degradation via the p38/MAPK and Akt pathways in human nucleus pulposus cells
Hucheng Liu, Runsheng Guo, Jiangyinzi Shang, Bin Zhang, Min Dai
Department of Orthopaedics, The First Affiliated Hospital, Nanchang University, Jiangxi, China
Received August 19, 2016; Accepted November 29, 2016; Epub February 15, 2017; Published February 28, 2017
Abstract: Previous studies have proved that resveratrol has beneficial effects on intervertebral disc regeneration. Tumor necrosis factor-α (TNF-α) has been proven to induce matrix degradation, which plays a critical role in inter-vertebral disc degeneration (IVDD). However, the effect of resveratrol on TNF-α-induced matrix degradation remains unknown. The purpose of this study was to find out whether resveratrol can reverse TNF-α-induced matrix degrada-tion in human NP cells and to further study related mechanism involved in the process. The transcriptional activity, mRNA and protein expression of Collagen II and Aggrecan were measured by luciferase reporter assay, RT-qPCR and western blot. Sulfated glycosaminoglycan (sGAG) level was detected to show the production of proteoglycans in NP extracellular matrix. TNF-α reduced Collagen II and Aggrecan production of human NP cells time-dependently. Resveratrol can rescue TNF-α-induced downregulation of Collagen II and Aggrecan production. MAPK, NF-κB and Akt signal pathways were activated after treatment with TNF-α and resveratrol. P38 inhibitor or Akt inhibitor can abol-ished the antagonistic effect of resveratrol on TNF-α-induced catabolic response. Our findings provide novel data supporting the protective effect of resveratrol, which may be used as a therapeutic agent in the treatment of IVDD.
Keywords: Resveratrol, tumor necrosis factor-α, matrix degradation, nucleus pulposus cell, signal pathway
Introduction
Low back pain is a common public health pro- blem worldwide, with a lifetime prevalence of 70-80% [1]. It significantly affects quality of life and burdens the health service to the detri-ment of all industrialized societies. The causes of back pain are multifactorial, intervertebral disc degeneration (IVDD) is one of the predomi-nant factors that leads to this disabling condi-tion [2-4]. Conservative treatments are current-ly aiming to alleviate the symptoms, while sur-geries involve excising the degenerated motion segment and fusion the adjacent vertebral bodies rather than to solve the underlying bio-logical causes involved in disc degeneration. There is a clear need for the development of novel therapeutic procedures that have the ability to interfere with the biological mecha-nism of IVDD. As an alternative to the surgical repair or removal of a diseased disc, biologic treatments to promote intervertebral disc (IVD) repair and restore physiologic function have been considered [5-12].
Resveratrol (trans-3,4’,5-trihydroxystilbene) was first identified in the roots of white hellebore in 1940, mainly found in the skins of grapes and red wines [13-15] and showed powerful biologi-cal functions of anti-inflammatory, antioxidant, cardioprotective, and antitumor properties [16-24]. Resveratrol has also been reported to pro-vide a protective effect on articular cartilage in rabbit models of osteoarthritis and rheumatoid arthritis [25, 26]. More recently, resveratrol has been found to relieve matrix-degrading enzy- mes upregulation and to facilitate proteoglycan synthesis and accumulation in bovine disc cells [27], as well as to decrease pro-inflammatory cytokines and catabolic enzymes expression in human IVD cells [28]. However, it is not known if resveratrol exerts similar protective effects on TNF-α-induced matrix degradation in human nucleus pulposus (NP) cells.
The purpose of this study was to elucidate whether resveratrol can reverse TNF-α-medi- ated matrix degradation in human NP cells and

Resveratrol inhibits TNF-α-induced matrix degradation
2765 Int J Clin Exp Med 2017;10(2):2764-2772
to further study related mechanism involved in the process.
Material and methods
Human NP cell isolation and culture
IVD specimens were harvested from patients with scoliosis in spine surgery on the basis of acquiring the informed consent of these patients. This procedure was approved by the Animal Ethics Committee of the First Affili- ated Hospital of Nanchang University. The NP tissues were digested with proteinase (Calbio- chem, CA, USA) and collagenase (Sigma-Aldri- ch, MO, USA). After filtration, the NP samples were cultured in T25 flasks in Dulbecco’s mo- dified Eagle medium (DMEM; Invitrogen, CA, USA) containing 10% fetal bovine serum (FBS; Invitrogen, CA, USA) and antibiotics. NP cells migrated out after one week, and passaged for use when confluent.
Real-time quantitative reverse transcription-PCR (RT-qPCR)
Total RNA was extracted from NP cells using TRIzol reagent (Invitrogen, CA, USA) and con-verted to cDNA using ReverTra Ace qPCR RT Kit (TOYOBO, OSAKA, JAPAN). The cDNA and gene-specific primers were added to SYBR qPCR Mix Kit (TOYOBO, OSAKA, JAPAN) and mRNA expression was quantified by using CFX96 Real-time PCR system (Bio-Rad, CA, USA). The sequences of synthesized primers are as follows: Collagen II forward 5’-TTCATG- CATCCGCTAGTCCCTTCT-3’, Collagen II reverse 5’-CTCCATGTTGCAGAAGACTTTCA-3’; Aggrecan forward 5’-ATTGGCACAAAGGTGGATGCTGTC-3’, Aggrecan reverse 5’-TCACCACCCACTCCGAAG- AAGTTT-3’; β-actin forward 5’-AGCCATGTACG- TAGCCATCC-3’, β-actin reverse 5’-CTCTCAGCT- GTGGTGGTGAA-3’. Endogenous housekeeping gene β-actin was used to normalize gene ex- pression. ΔΔCT method was performed to cal-culate the fold change. Each sample was ana-lyzed in duplicate. All primers used were syn-thesized by Sangon Biotech Co., Ltd (Shanghai, CHINA).
Protein extraction and western blotting
Total cell proteins were extracted using RIPA buffer and measured with a BCA assay (Ther- mo Scientific). Equal amounts of the proteins were resolved on SDS-polyacrylamide gels and transferred to PVDF membranes. Membranes
were blocked with 5% nonfat milk in TBST and incubated overnight at 4°C in 5% nonfat milk in TBST with the anti-Collagen II antibody (1:1000, Abcam); anti-Aggrecan antibody (1:1000), anti-GAPDH (1:1000), anti-p65 antibody (1:1000), anti-phospho-p65 antibody (1:1000), anti-p38 antibody (1:1000), anti-phospho-p38 antibody (1:1000), anti-JNK1/2 antibody (1:1000), anti-phospho-JNK1/2 antibody (1:1000), anti-ERK antibody (1:1000), anti-phospho-ERK antibody (1:1000), anti-Akt antibody (1:1000), anti-phos-pho-Akt antibody (1:1000) all from Cell Signal- ing. The membranes were washed three times next morning and incubated with anti-rabbit secondary antibody (1:2000, Cell Signaling) for 1 h at room temperature. Immunolabeling was measured using the ECL reagent (Invitrogen, CA, USA). Densitometric analysis was perform- ed using Image J analysis software (Bethesda, MD, USA).
Sulfated glycosaminoglycan (sGAG) detection
Proteoglycans in NP extracellular matrix are made up of sulfated glycosaminoglycan (sGAG) and core protein. sGAG expression was detect-ed using a 1,9-dimethylmethylene blue (DM- MB) method with Blyscan Sulfated Glycosami- noglycan Assay Kit (Biocolor, CF, UK) to repre-sent the proteoglycans production of NP cells.
Luciferase reporter assay
Cells were plated to 48-well plates at a density of 2×104 cells cells/well 1 day before transfec-tion. Lipofectamine 2000 (Invitrogen, CA, USA) was used as a transfection reagent. A Collagen II or Aggrecan promoter luciferase construct was utilized to detect the effect of TNF-α and resveratrol on the Collagen II or Aggrecan pro-moter. NP cells were transfected with 250 ng Collagen II-Luc or Aggrecan-Luc and 250 ng pRL-TK plasmids for 6 h. The next day, cells were harvested and Dual-Luciferase Reporter Assay System (Promega, WI, USA) was used for measurements of firefly and Renilla lucifer-ase activities using a TD-20/20 luminometer (Turner Biosystems, CA, USA).
Statistical analysis
All measurements were performed in triplicate. Data are presented as the mean ± standard error. Differences between groups were as- sessed by one-way ANOVA with Bonferroni’s post hoc test for multiple comparisons and
Resveratrol inhibits TNF-α-induced matrix degradation
2766 Int J Clin Exp Med 2017;10(2):2764-2772
Student’s T-test. P-values <0.05 was consid-ered statistically significant.
Results
TNF-α suppresses collagen II and aggrecan expression in NP cells
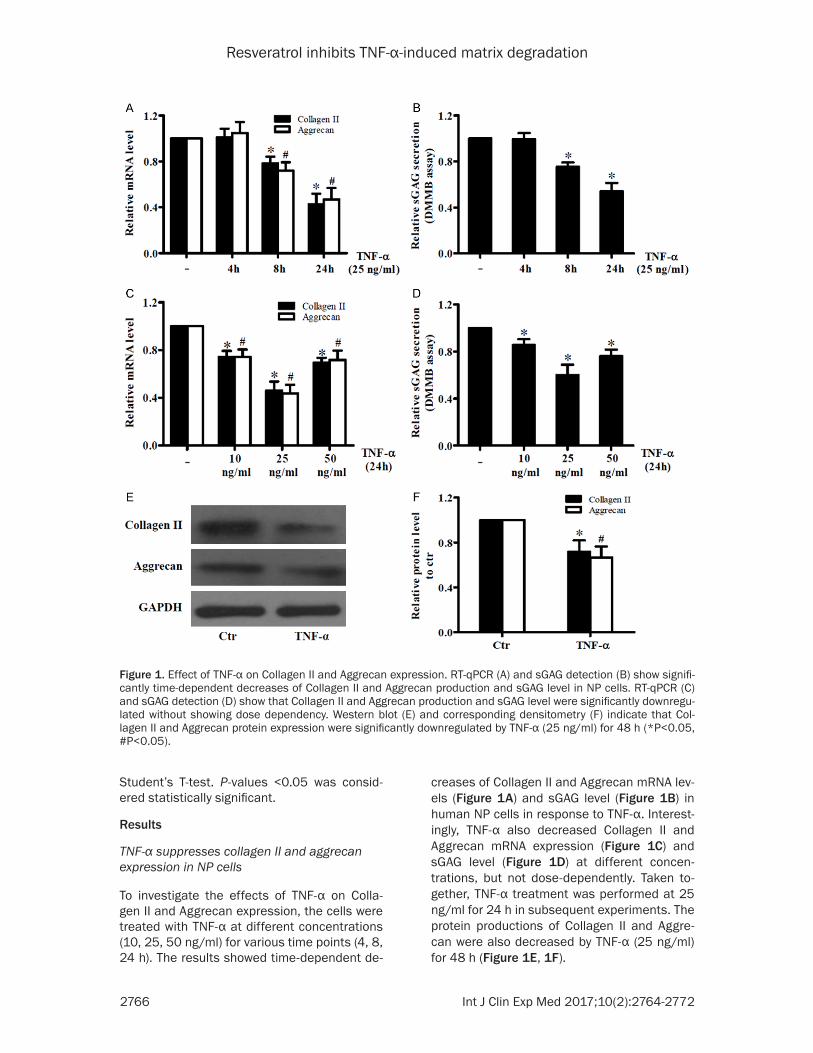
To investigate the effects of TNF-α on Colla- gen II and Aggrecan expression, the cells were treated with TNF-α at different concentrations (10, 25, 50 ng/ml) for various time points (4, 8, 24 h). The results showed time-dependent de-
creases of Collagen II and Aggrecan mRNA lev-els (Figure 1A) and sGAG level (Figure 1B) in human NP cells in response to TNF-α. Interest- ingly, TNF-α also decreased Collagen II and Aggrecan mRNA expression (Figure 1C) and sGAG level (Figure 1D) at different concen- trations, but not dose-dependently. Taken to- gether, TNF-α treatment was performed at 25 ng/ml for 24 h in subsequent experiments. The protein productions of Collagen II and Aggre- can were also decreased by TNF-α (25 ng/ml) for 48 h (Figure 1E, 1F).
Figure 1. Effect of TNF-α on Collagen II and Aggrecan expression. RT-qPCR (A) and sGAG detection (B) show signifi-cantly time-dependent decreases of Collagen II and Aggrecan production and sGAG level in NP cells. RT-qPCR (C) and sGAG detection (D) show that Collagen II and Aggrecan production and sGAG level were significantly downregu-lated without showing dose dependency. Western blot (E) and corresponding densitometry (F) indicate that Col-lagen II and Aggrecan protein expression were significantly downregulated by TNF-α (25 ng/ml) for 48 h (*P<0.05, #P<0.05).
Resveratrol inhibits TNF-α-induced matrix degradation
2767 Int J Clin Exp Med 2017;10(2):2764-2772
Resveratrol restores the catabolic effect of TNF-α on NP cells
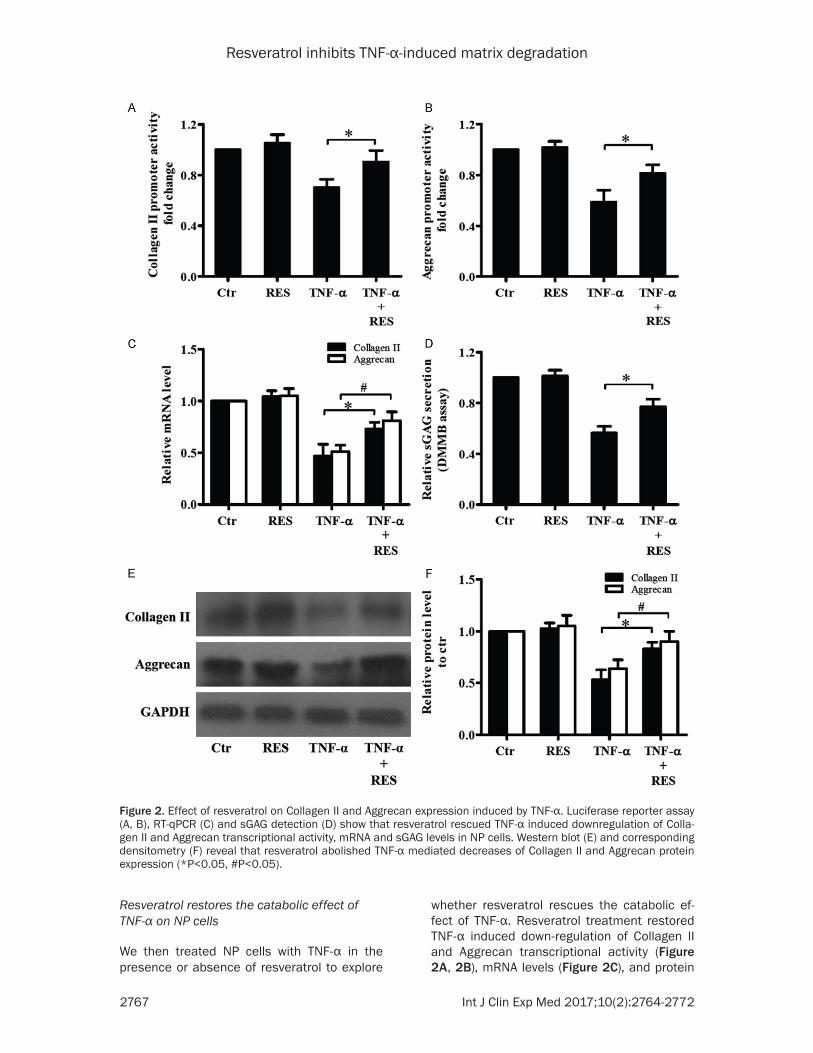
We then treated NP cells with TNF-α in the presence or absence of resveratrol to explore
whether resveratrol rescues the catabolic ef- fect of TNF-α. Resveratrol treatment restored TNF-α induced down-regulation of Collagen II and Aggrecan transcriptional activity (Figure 2A, 2B), mRNA levels (Figure 2C), and protein
Figure 2. Effect of resveratrol on Collagen II and Aggrecan expression induced by TNF-α. Luciferase reporter assay (A, B), RT-qPCR (C) and sGAG detection (D) show that resveratrol rescued TNF-α induced downregulation of Colla-gen II and Aggrecan transcriptional activity, mRNA and sGAG levels in NP cells. Western blot (E) and corresponding densitometry (F) reveal that resveratrol abolished TNF-α mediated decreases of Collagen II and Aggrecan protein expression (*P<0.05, #P<0.05).

Resveratrol inhibits TNF-α-induced matrix degradation
2768 Int J Clin Exp Med 2017;10(2):2764-2772
expression (Figure 2E, 2F). An elevation was also observed in sGAG secretion in NP cells treated with TNF-α and resveratrol (Figure 2D). These results suggest that resveratrol exhi- bits an anti-catabolic effect against TNF-α on NP cells.
Activation of MAPK, NF-κB and Akt signal pathways following treatment with TNF-α and resveratrol
To further determine if MAPK, NF-κB and Akt signal pathways were involved in regulation of Collagen II and Aggrecan induced by TNF-α and resveratrol in NP cells, we evaluated the ac- tivation of p38, JNK1/2, ERK1/2, NF-κB and Akt signal pathways after TNF-α and resvera-trol treatment by western blot.
Immunoblot analysis showed the phosphoryla-tion levels of p38, JNK1/2, ERK1/2, NF-κB and Akt were elevated at 30 min after TNF-α treat-ment, and resveratrol further increased p38 and Akt phosphorylation (Figure 3).
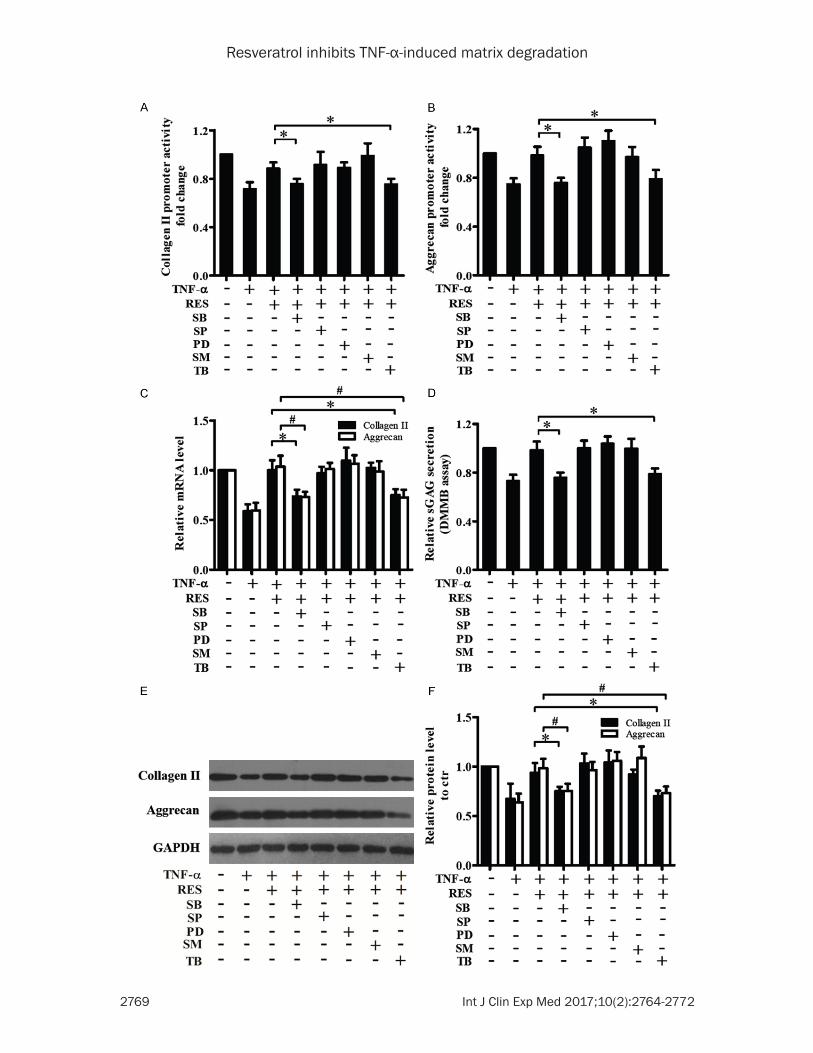
Effect of MAPK, NF-κB and Akt signal path-ways in resveratrol induced anti-catabolic role
To investigate whether MAPK, NF-κB and Akt signal pathways were was involved in resvera-trol-induced anti-catabolic effect towards TNF-α, we treated the NP cells with TNF-α and res-veratrol in the presence or absence of p38 inhibitor (SB203580, 10 µM), JNK1/2 inhibitor (SP600125, 25 µM), ERK1/2 inhibitor (PD98- 059, 20 µM), NF-κB inhibitor (SM7368, 10 µM) and Akt inhibitor (triciribine, 20 µM). Compared to the group of TNF-α plus resveratrol, the Collagen II and Aggrecan transcriptional acti- vity (Figure 4A, 4B), mRNA levels (Figure 4C), protein expression (Figure 4E, 4F) and sGAG secretion (Figure 4D) in the group of TNF-α and resveratrol plus p38 inhibitor or Akt inhibitor were significantly decreased; while inhibition of JNK1/2, ERK1/2 or NF-κB showed no signifi-cant change. These results suggest that the inhibition of p38 or Akt signaling abolished the anti-catabolic effect of resveratrol against TNF-α.
Discussion
The IVD is a major joint of the spine which allows for motion and posture. It is composed of a gelatinous NP, fibrous annulus fibrosus (AF) and sandwiched between two cartilaginous endplates. The degeneration of the IVD is asso-ciated with back pain incidence and severity, and is purported to be one of the major reasons for hospital visits. The causes of degeneration are multifactorial and include both environmen-tal and genetic factors. During degeneration, there are biochemical and cellular changes that occur in the disc including the loss of cells, pro-teoglycans and water, which impacts on the biomechanics and function of the disc.
While the etiology of disc degeneration is not clear, however, one major characteristic in the degeneration is inflammation. Many studies have indicated that degenerated IVDs are in a chronic inflammatory state, and that inflam-matory factors can induce degenerative chang-es in IVD. Human studies have suggested that
Figure 3. Activation of MAPK, NF-κB and Akt signal pathways after TNF-α and resveratrol treatment. Western blot analysis shows that p38, JNK1/2, ERK1/2, NF-κB and Akt pathways were activated af-ter TNF-α after TNF-α treatment, and resveratrol fur-ther increased p38 and Akt phosphorylation.
Resveratrol inhibits TNF-α-induced matrix degradation
2769 Int J Clin Exp Med 2017;10(2):2764-2772

Resveratrol inhibits TNF-α-induced matrix degradation
2770 Int J Clin Exp Med 2017;10(2):2764-2772
increased expression of multiple pro-inflam- matory cytokines is associated with back pain pertaining to IVDD. Examination of surgical- ly removed degenerated disc fragments have shown expression of IL-1, IL-8, TNF-α, IL-10 and PGE2 [29, 30]. Repeated physical injury in rat IVDs can induce persistent inflammation with expression of various pro-inflammatory cyto-kines that are associated with IVDD [31].
Resveratrol has been demonstrated to process antitumor, anti-aging, anti-inflammatory and neuroprotective activities [32]. Csaki C et al [33] and Shakibaei M et al [34] have shown the potential application of resveratrol for the treatment of osteoarthritis. An animal study demonstrated that resveratrol could reduce cartilage degradation and loss of matrix pro- teoglycans on cartilage tissue [25]. Resveratrol has also been reported to decrease transcript and protein production of major pro-inflamma-tory cytokines and matrix catabolic enzymes, and shown analgetic potential to reduce NP- mediated pain in an in vivo animal model of painful radiculopathy [28]. A study perform- ed by Li et al [27] showed that resveratrol fa- cilitated proteoglycan synthesis and reversed IL-1β-mediated proteoglycan loss in bovine IVD cells.
This study illustrates the potent anabolic ef- fects of resveratrol on human IVD homeosta-sis. Treatment with TNF-α reduced Collagen II and Aggrecan at the transcriptional and pro- tein levels, decreased sGAG accumulation in human NP cells. In addition, resveratrol rever- sed the catabolic effects of TNF-α that have been implicated in IVDD.
NF-κB is a key transcription factor that re- gulates various physiological processes includ-ing inflammatory responses and apoptosis, which also plays a critical role in the process of disc degeneration [35, 36]. The MAPK sig- naling pathways have been shown key role in the transduction of extracellular signals to cell responses that may include cell growth, proliferation, differentiation, development, in- flammatory responses, apoptosis and invasion
in mammalian cells [37, 38]. The PI3K/Akt sig-naling pathway is important for cell growth, dif-ferentiation and survival [39]. In the present study, it was found that the phosphorylation levels of p38, JNK1/2, ERK1/2, NF-κB and Akt were increased at 30 min after TNF-α treat-ment, and resveratrol further increased p38 and Akt phosphorylation. In addition, inhibi- tor experiments were performed to determine whether resveratrol can rescue the catabolic effect of TNF-α via MAPK, NF-κB and Akt sig- nal pathways. The results indicated that the inhibition of p38 or Akt signaling can abolish the protective effect of resveratrol against TNF-α.
Conclusions
Our results shed light on the inflammatory response of multiple downstream regulatory molecules after stimulation with resveratrol in human NP cells, which are regulated through p38 and Akt signaling pathways. These data provide us with a better understanding of the anti-inflammatory, antioxidant, and antiprolifer-ative and anabolic effects of resveratrol. In addition, as the mechanisms behind the sup-pressed inflammatory response following expo-sure to resveratrol for long time periods are not known, which still requires further investigation to address. Such study results may provide novel data supporting the protective effect of resveratrol, which may be used as a therapeu-tic agent in the treatment of IVDD.
Disclosure of conflict of interest
None.
Address correspondence to: Min Dai, Department of Orthopaedics, The First Affiliated Hospital, Nan- chang University, No. 17 Yongwai Street, Nan- chang 330006, Jiangxi, China. Tel: 0086-13870- 051522; Fax: 0086-791-88694127; E-mail: [email protected]
References
[1] Balagué F, Mannion AF, Pellisé F, Cedraschi C. Non-specific low back pain. Lancet 2012; 379: 482-491.
Figure 4. Effect of MAPK, NF-κB and Akt signal pathways on Collagen II and Aggrecan expression induced by TNF-α and resveratrol. Luciferase reporter assay (A, B), RT-qPCR (C), sGAG detection (D), Western blot (E) and correspond-ing densitometry (F) indicate that resveratrol inhibited TNF-α mediated downregulation of Collagen II and Aggrecan transcriptional activity, mRNA levels, sGAG level and protein expression. Inhibition of p38 (SB203580, 10 µM) or Akt (triciribine, 20 µM) pathway abolishes the anti-catabolic role of resveratrol against TNF-α (*P<0.05, #P<0.05).

Resveratrol inhibits TNF-α-induced matrix degradation
2771 Int J Clin Exp Med 2017;10(2):2764-2772
[2] Hoy D, Brooks P, Blyth F, Buchbinder R. The epidemiology of low back pain. Best Pract Res Clin Rheumatol, 2010; 24: 769-81.
[3] Freemont AJ. The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain. Rheumatology (Oxford) 2009; 48: 5-10.
[4] Freemont TJ, LeMaitre C, Watkins A, Hoyland JA. Degeneration of intervertebral discs: cur-rent understanding of cellular and molecular events, and implications for novel therapies. Expert Rev Mol Med 2001; 2001: 1-10.
[5] An HS, Thonar EJ, Masuda K. Biological repair of intervertebral disc. Spine 2003; 28: S86-92.
[6] An H, Boden SD, Kang J, Sandhu HS, Abdu W, Weinstein J. Summary statement: emerging techniques for treatment of degenerative lum-bar disc disease. Spine 2003; 28: S24-5.
[7] Zhang Y, An HS, Song S, Toofanfard M, Masuda K, Andersson GB, Thonar EJ. Growth factor os-teogenic protein-1: differing effects on cells from three distinct zones in the bovine inter-vertebral disc. Am J Phys Med Rehabil 2004; 83: 515-21.
[8] Chujo T, An HS, Akeda K, Miyamoto K, Muehle- man C, Attawia M, Andersson G, Masuda K. Effects of growth differentiation factor-5 on the intervertebral disc: in vitro bovine study and in vivo rabbit disc degeneration model study. Spine (Phila Pa 1976) 2006; 31: 2909-17.
[9] Hohaus C, Ganey TM, Minkus Y, Meisel HJ. Cell transplantation in lumbar spine disc degenera-tion disease. Eur Spine J 2008; 17: 492-503.
[10] Kandel R, Roberts S, Urban JP. Tissue engi-neering and the intervertebral disc: the chal-lenges. Eur Spine J 2008; 17: 480-91.
[11] Kwon YJ, Lee JW, Moon EJ, Chung YG, Kim OS, Kim HJ. Anabolic effects of Peniel 2000, a peptide that regulates TGF-β1 signaling on in-tervertebral disc degeneration. Spine (Phila Pa 1976) 2013; 38: E49-58.
[12] Moon SH, Gilbertson LG, Nishida K, Knaub M, Muzzonigro T, Robbins PD, Evans CH, Kang JD. Human intervertebral disc cells are geneti-cally modifiable by adenovirus-mediated gene transfer: implications for theclinical manage-ment of intervertebral disc disorders. Spine (Phila Pa 1976) 2000; 25: 2573-9.
[13] Marques FZ, Markus MA, Morris BJ. Resver- atrol: cellular actions of a potent natural chem-ical that confers a diversity of health benefits. Int J Biochem Cell Biol 2009; 41: 2125-2128.
[14] Shakibaei M, Harikumar KB, Aggarwal BB. Re- sveratrol addiction: to die or not to die. Mol Nutr Food Res 2009; 53: 115-128.
[15] Yu W, Fu YC, Wang W. Cellular and molecular effects of resveratrol in health and disease. J Cell Biochem 2012; 113: 752-759.
[16] Haider UG, Sorescu D, Griendling KK, Vollmar AM, Dirsch VM. Resveratrol suppresses angio-tensin II-induced Akt/protein kinase B and p70 S6 kinase phosphorylation and subsequent hypertrophy in rat aortic smooth muscle cells. Mol Pharmacol 2002; 62: 772-7.
[17] Bhat KPL, Kosmeder JW 2nd, Pezzuto JM. Bio- logical effects of resveratrol. Antioxid Redox Signal 2001; 3: 1041-64.
[18] Bertelli AA, Ferrara F, Diana G, Fulgenzi A, Corsi M, Ponti W, Ferrero ME, Bertelli A. Resveratrol, a natural stilbene in grapes and wine, enhanc-es intraphagocytosis in human promonocytes: a co-factor in antiinflammatory and anticancer chemopreventive activity. Int J Tissue React 1999; 21: 93-104.
[19] Fremont L. Biological effects of resveratrol. Life Sci 2000; 66: 663-73.
[20] Huang KS, Lin M, Cheng GF. Anti-inflamma- tory tetramers of resveratrol from the roots of Vitis amurensis and the conformations of the seven membered ring in some oligostilbenes. Phytochemistry 2001; 58: 357-62.
[21] Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta RG, Moon RC, Pezzuto JM. Cancer chemopreventive activity of resve-ratrol, a natural product derived from grapes. Science 1997; 275: 218-20.
[22] Martinez J, Moreno JJ. Effect of resveratrol, a natural polyphenolic compound, on reactive oxygen species and prostaglandin production. Biochem Pharmacol 2000; 59: 865-70.
[23] Ignatowicz E, Baer-Dubowska W. Resveratrol, a natural chemopreventive agent against de-generative diseases. Pol J Pharmacol 2001; 53: 557-69.
[24] Leiro J, Alvarez E, Arranz JA, Laguna R, Uriarte E, Orallo F. Effects of cis-resveratrol on in- flammatory murine macrophages: antioxidant activity and down-regulation of inflammatory genes. J Leukoc Biol 2004; 75: 1156-65.
[25] Elmali N, Esenkaya I, Harma A, Ertem K, Turkoz Y, Mizrak B. Effect of resveratrol in experimen-tal osteoarthritis in rabbits. Inflamm Res 2005; 54: 158-62.
[26] Elmali N, Baysal O, Harma A, Esenkaya I, Mizrak B. Effects of resveratrol in inflammatory arthritis. Inflammation 2007; 30: 1-6.
[27] Li X, Phillips FM, An HS, Ellman M, Thonar EJ, Wu W, Park D, Im HJ. The action of resveratrol, a phytoestrogen found in grapes, on the inter-vertebral disc. Spine (Phila Pa 1976) 2008; 33: 2586-95.
[28] Wuertz K, Quero L, Sekiguchi M, Klawitter M, Nerlich A, Konno S, Kikuchi S, Boos N. The red wine polyphenol resveratrol shows promising potential for the treatment of nucleus pulpo-

Resveratrol inhibits TNF-α-induced matrix degradation
2772 Int J Clin Exp Med 2017;10(2):2764-2772
sus-mediated pain in vitro and in vivo. Spine (Phila Pa 1976) 2011; 36: E1373-84.
[29] Ahn SH, Cho YW, Ahn MW, Jang SH, Sohn YK, Kim HS. mRNA expression of cytokines and chemokines in herniated lumbar intervertebral discs. Spine (Phila Pa 1976) 2002; 27: 911-7.
[30] O’Donnell JL, O’Donnell AL. Prostaglandin E2 content in herniated lumbar disc disease. Spine (Phila Pa 1976) 1996; 21: 1653-5.
[31] Ulrich JA, Liebenberg EC, Thuillier DU, Lotz JC. ISSLS prize winner: repeated disc injury causes persistent inflammation. Spine (Phila Pa 1976) 2007; 32: 2812-9.
[32] Tang FY, Su YC, Chen NC, Hsieh HS, Chen KS. Resveratrol inhibits migration and invasion of human breast-cancer cells. Mol Nutr Food Res 2008; 52: 683-691.
[33] Csaki C, Keshishzadeh N, Fischer K, Shakibaei M. Regulation of inflammation signalling by resveratrol in human chondrocytes in vitro. Biochem Pharmacol 2008; 75: 677-87.
[34] Shakibaei M, Csaki C, Nebrich S, Mobasheri A. Resveratrol suppresses interleukin-1beta-in-duced inflammatory signaling and apoptosis in human articular chondrocytes: potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem Pharmacol 2008; 76: 1426-39.
[35] Hoyland JA, Le Maitre C, Freemont AJ. Inves- tigation of the role of IL-1 and TNF in matrix degradation in the intervertebral disc. Rheu- matology (Oxford) 2008; 47: 809-14.
[36] Le Maitre CL, Hoyland JA, Freemont AJ. Cata- bolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1beta and TNFalpha expression profile. Arthritis Res Ther 2007; 9: R77.
[37] Widmann C, Gibson S, Jarpe MB and Johnson GL. Mitogen-activated protein kinase: conser-vation of a three-kinase module from yeast to human. Physiol Rev 1999; 79: 143-180.
[38] Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res 2002; 12: 9-18.
[39] Foster FM, Traer CJ, Abraham SM, Fry MJ. The phosphoinositide (PI) 3-kinase family. J Cell Sci 2003; 116: 3037-3040.


















