
Loss of epithelial oestrogen receptor α inhibits oestrogen-stimulated prostate proliferation and...
11
ORIGINAL PAPER Journal of Pathology J Pathol 2012; 226: 17–27 Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/path.2949 Loss of epithelial oestrogen receptor α inhibits oestrogen-stimulated prostate proliferation and squamous metaplasia via in vivo tissue selective knockout models Ming Chen, 1† Chiuan-Ren Yeh, 1† Hong-Chiang Chang, 1,2 Spencer Vitkus 1 , Xing-Qiao Wen 3 , Neil A. Bhowmick 4 , Andrew Wolfe 5 and Shuyuan Yeh 1 * 1 Departments of Urology and Pathology, University of Rochester Medical Center, Rochester, NY 14642, USA 2 Department of Urology, National Taiwan University Hospital, Taiwan 3 Department of Urology, Third Affiliated Hospital, Sun Yat-Sen University, Guangzhou, China 4 Samuel Oschin Comprehensive Cancer Institute at the Cedars-Sinai Medical Center, Los Angeles, CA 90048, USA 5 Department of Pediatrics, Johns Hopkins University School of Medicine, Baltimore, Maryland 21287, USA *Correspondence to: Dr Shuyuan Yeh, Department of Urology, University of Rochester Medical Center, 601 Elmwood Avenue, Box 656, Rochester, NY 14642, USA. e-mail: [email protected] † These authors contributed equally to this paper. Abstract Squamous metaplasia (SQM) is a specific phenotype in response to oestrogen in the prostate and oestrogen receptor (ER) α is required to mediate this response. Previous studies utilizing tissue recombination with seminal vesicle (SV) mesenchyme and prostatic ductal tips from wild type and ERαKO mice suggested that both epithelial and stromal ERα are necessary for SQM. However, tissue recombination is conducted in the renal capsule of immune-deficient mice, in which the microenvironment is different from normal prostate microenvironment in the intact mice. Furthermore, whether the requirement of stromal ERα in the SV for developing SQM is the same as in the prostate is unknown. Therefore, there is a clear need to evaluate the respective roles of ERα in prostate epithelial versus stromal compartments in the intact mouse. Here we generated a mouse model that has selectively lost ERα in either stromal (FSP-ERαKO) or epithelial prostate cells (pes-ERαKO) to determine the requirements of ERα for oestrogen-stimulated prostate proliferation and SQM. Our results indicated that FSP-ERαKO prostates develop full and uniform SQM, which suggests that loss of the majority (∼65%) of stromal ERα will not influence oestrogen-mediated SQM. In contrast, loss of epithelial ERα inhibits oestrogen-mediated prostate growth and SQM evidenced by decreasing cytokertin 10 positive squamous cell stratification and differentiation, by reduced ERα protein expression in SQM compared to wild type mice ERα, and by the presence of normal proliferative activities in the oestrogen-treated pes-ERαKO prostates. These in vivo results suggest that epithelial ERα is required for oestrogen-mediated proliferative response and could be an appropriate target for preventing aberrant oestrogen signalling in the prostate. Copyright 2011 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd. Keywords: oestrogen receptor α; squamous metaplasia; prostate; tissue-specific knockout; Cre-loxP Received 31 December 2010; Revised 24 April 2011; Accepted 25 May 2011 No conflicts of interest were declared. Introduction The prostate is primarily considered to be an andro- gen target organ [1], but oestrogen can also influence its development and pathogenesis [2]. Oestrogens exert both indirect and direct effects on the pro- static organ. The indirect oestrogen action is medi- ated at the hypothalamic level to suppress circulating androgens and produce a ‘castration-like’ effect [3–6]. The direct actions of oestrogens are mediated by two distinct receptors, oestrogen receptor (ER) α and β in the prostate [7–10]. Studies using ERα knockout (ERαKO) mouse prostates have consistently shown that ERα is involved in the prostatic branching mor- phogenesis and required for normal prostate develop- ment [11–13]. Studies using conventional ERβKO/neo knock-in mouse prostates showed inconsistent pheno- types in the hyperplastic prostates [14–16]. Recently, an updated Cre-loxP ERβKO mouse model was developed via mating CMV-cre with floxed ERβ mice (CMV-cre-ERβKO) [17]. The CMV-cre-ERβKO males appear to be complete ERβ null mice, yet there is no prostatic phenotype. Taken together, ERβ does not seem to have a critical role in prostate development. Besides the normal physiological roles of oestrogen signalling in prostate development, exposure of peri- natal or neonatal male mice to oestrogens leads to a Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27 Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com
Transcript of Loss of epithelial oestrogen receptor α inhibits oestrogen-stimulated prostate proliferation and...

ORIGINAL PAPERJournal of PathologyJ Pathol 2012; 226: 17–27Published online in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/path.2949
Loss of epithelial oestrogen receptor α inhibitsoestrogen-stimulated prostate proliferation and squamousmetaplasia via in vivo tissue selective knockout modelsMing Chen,1† Chiuan-Ren Yeh,1† Hong-Chiang Chang,1,2 Spencer Vitkus1, Xing-Qiao Wen3, Neil A. Bhowmick4,Andrew Wolfe5 and Shuyuan Yeh1*
1 Departments of Urology and Pathology, University of Rochester Medical Center, Rochester, NY 14642, USA2 Department of Urology, National Taiwan University Hospital, Taiwan3 Department of Urology, Third Affiliated Hospital, Sun Yat-Sen University, Guangzhou, China4 Samuel Oschin Comprehensive Cancer Institute at the Cedars-Sinai Medical Center, Los Angeles, CA 90048, USA5 Department of Pediatrics, Johns Hopkins University School of Medicine, Baltimore, Maryland 21287, USA
*Correspondence to: Dr Shuyuan Yeh, Department of Urology, University of Rochester Medical Center, 601 Elmwood Avenue, Box 656, Rochester,NY 14642, USA. e-mail: [email protected]
†These authors contributed equally to this paper.
AbstractSquamous metaplasia (SQM) is a specific phenotype in response to oestrogen in the prostate and oestrogenreceptor (ER) α is required to mediate this response. Previous studies utilizing tissue recombination with seminalvesicle (SV) mesenchyme and prostatic ductal tips from wild type and ERαKO mice suggested that both epithelialand stromal ERα are necessary for SQM. However, tissue recombination is conducted in the renal capsule ofimmune-deficient mice, in which the microenvironment is different from normal prostate microenvironment inthe intact mice. Furthermore, whether the requirement of stromal ERα in the SV for developing SQM is the sameas in the prostate is unknown. Therefore, there is a clear need to evaluate the respective roles of ERα in prostateepithelial versus stromal compartments in the intact mouse. Here we generated a mouse model that has selectivelylost ERα in either stromal (FSP-ERαKO) or epithelial prostate cells (pes-ERαKO) to determine the requirementsof ERα for oestrogen-stimulated prostate proliferation and SQM. Our results indicated that FSP-ERαKO prostatesdevelop full and uniform SQM, which suggests that loss of the majority (∼65%) of stromal ERα will not influenceoestrogen-mediated SQM. In contrast, loss of epithelial ERα inhibits oestrogen-mediated prostate growth andSQM evidenced by decreasing cytokertin 10 positive squamous cell stratification and differentiation, by reducedERα protein expression in SQM compared to wild type mice ERα, and by the presence of normal proliferativeactivities in the oestrogen-treated pes-ERαKO prostates. These in vivo results suggest that epithelial ERα isrequired for oestrogen-mediated proliferative response and could be an appropriate target for preventing aberrantoestrogen signalling in the prostate.Copyright 2011 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Keywords: oestrogen receptor α; squamous metaplasia; prostate; tissue-specific knockout; Cre-loxP
Received 31 December 2010; Revised 24 April 2011; Accepted 25 May 2011
No conflicts of interest were declared.
Introduction
The prostate is primarily considered to be an andro-gen target organ [1], but oestrogen can also influenceits development and pathogenesis [2]. Oestrogensexert both indirect and direct effects on the pro-static organ. The indirect oestrogen action is medi-ated at the hypothalamic level to suppress circulatingandrogens and produce a ‘castration-like’ effect [3–6].The direct actions of oestrogens are mediated by twodistinct receptors, oestrogen receptor (ER) α and β
in the prostate [7–10]. Studies using ERα knockout(ERαKO) mouse prostates have consistently shown
that ERα is involved in the prostatic branching mor-phogenesis and required for normal prostate develop-ment [11–13]. Studies using conventional ERβKO/neoknock-in mouse prostates showed inconsistent pheno-types in the hyperplastic prostates [14–16]. Recently,an updated Cre-loxP ERβKO mouse model wasdeveloped via mating CMV-cre with floxed ERβ mice(CMV-cre-ERβKO) [17]. The CMV-cre-ERβKO malesappear to be complete ERβ null mice, yet there isno prostatic phenotype. Taken together, ERβ does notseem to have a critical role in prostate development.
Besides the normal physiological roles of oestrogensignalling in prostate development, exposure of peri-natal or neonatal male mice to oestrogens leads to a
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

18 M Chen et al
reduced responsiveness to androgens during adulthoodand an increased incidence of severe dysplasia withageing in the prostate gland, which is similar to high-grade prostatic intraepithelial neoplasia (PIN) in thehuman prostate [18,19]. The effect on prostate develop-ment, following oestrogen exposure in the prenatal orneonatal rodent, is called developmental oestrogeniza-tion of the prostate (also called oestrogen imprintingeffect), a process by which genomic signals generatedin utero can induce a greater propensity for cancer inlater life. In contrast, exposure of adult prostate tooestrogens leads to squamous metaplasia (SQM) inthe prostate gland. SQM is associated with a strat-ified basal epithelial layer, onset of cytokeratin 10(CK10) expression, and enhanced proliferative activi-ties in the oestrogen-treated prostates [20]. Together,developmental oestrogenization of the prostate andSQM are specific responses to oestrogen in the prostategland and appear to be mediated by ERα, but not ERβ
[19,21].The prostate develops in the context of stro-
mal–epithelial cell interaction. The reciprocal inter-action between prostatic epithelial and stromal cellsplays a role in homeostasis and maintaining the growthquiescence in the adult prostatic gland [22]. It hasbeen reported that ERα could be detected in bothprostate epithelial and stromal cells [12,18,19,21].There is some controversy about whether epithelialversus stromal ERα plays a role in the developmentaloestrogenization of the prostate. Based on the immuno-histochemical staining profile of ERα, the results ofone study suggested that stromal ERα, but not epithe-lial ERα, is required for the oestrogen imprinting effecton the prostate [19], while another study showed thatboth epithelial and stromal ERα are required for theoestrogen imprinting effect [12].
In an effort to determine the respective roles ofepithelial versus stromal ERα in oestrogen-mediatedSQM, an earlier study utilized tissue recombinationtechniques with newborn seminal vesicle (SV) mes-enchyme and adult prostatic ductal tips from wild type(Wt) and ERαKO mice, showing that both epithelialand stromal ERα are necessary for oestrogen-mediatedSQM in the prostate [21]. However, tissue recombi-nation is conducted in the renal capsule of immune-deficient mice, in which the microenvironment is dif-ferent from the prostate microenvironment in immune-intact mice. In addition, the experimental method usedprostate duct tips rather than prostate epithelial cells asthe epithelial compartment to prepare the tissue recom-binants; therefore the ductal tips could have been con-taminated by some stromal cells. Furthermore, whetherthe function of stromal ERα in the SV mesenchymeis the same as in prostate mesenchyme in oestrogen-mediated SQM is unknown.
Until recently, there have been no models to elu-cidate the respective roles of epithelial versus stromalERα in vivo within the murine prostate during prostaticpathogenesis, or to demonstrate a direct implication
for understanding prostatic disease and pharmaceuti-cal targets. In the present study, we generated mousemodels that have lost ERα selectively in either stro-mal cells (FSP-ERαKO) or prostate epithelial cells(pes-ERαKO), to determine the temporal and spatialrequirements of ERα in the oestrogen-stimulated pro-liferation and SQM in the prostate.
Materials and methods
Generation of tissue-specific ERαKO miceand hormone treatmentThe whole-body Beta-actin (ACTB)-ERαKO, prostateepithelial specific (pes)-ERαKO, and fibroblast spe-cific protein (FSP)-ERαKO mice were generated bymating floxed ERα female mice with ACTB-Cre,probasin-Cre, and FSP1-Cre transgenic male mice,in which Cre recombinase is under the control ofthe β-actin, probasin, and FSP1 promoter, respec-tively [23–25]. Genomic DNA was used as a tem-plate for PCR genotyping of DNA isolated from tailbiopsies. Genotyping and the primers used were aspreviously described [24]. The adult ACTB-ERαKO,FSP-ERαKO, and pes-ERαKO male mice, or Wt litter-mate controls, were treated with subcutaneous implantsof 20 mg pellets containing 2 mg of diethylstilbe-strol (DES) and 18 mg of cholesterol at 12 weeksof age [21] for each treatment group. Groups offive Wt or ERαKO animals were studied at inter-vals of up to 3 weeks following DES treatment. Themice were sacrificed and prostate tissues were har-vested at 15 weeks of age. All animal procedureswere approved by the Animal Care and Use Com-mittee of the University of Rochester Medical Cen-ter, in accordance with National Institutes of Healthguidelines.
Immunohistochemistry (IHC)and immunofluorescent (IF) stainingIHC and IF staining were carried out as describedpreviously [11,26]. Sections were incubated with thefollowing primary antibodies and dilutions: anti-CK10(DE-K10, 1 : 50), anti-p63 (A4A, 1 : 100), anti-ERα
antibodies (MC-20, 1 : 400), anti-CK5 (AF138, 1 : 200),anti-androgen receptor (AR) (N-20, 1 : 400), anti-p27(C-19, 1 : 200), and anti-Ki67 (NCL-Ki67p, 1 : 1000)in 3% BSA in PBS overnight at 4 ◦C, followed bythe respective secondary antibodies. The stained slideswere mounted and visualized by a bright-field orfluorescent microscope.
Statistical analysisResults were expressed as mean ± SEM. Statisticalanalysis was performed by using an independent Stu-dent t-test for two groups of data and analysis of vari-ance (ANOVA) followed by Scheffe’s test for multiplecomparisons. A p value of less than 0.05 was consid-ered statistically significant.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

ERα and metaplasia in the prostate 19
Results
Loss of epithelial ERα inhibits oestrogen-mediatedSQM in pes-ERαKO prostatesIn order to address the respective roles of epithelialversus stromal ERα in SQM, we generated mice thatconditionally lack ERα in the prostate epithelial cellsor stromal cells by mating floxed ERα female micewith male mice expressing Cre recombinase, whichis under control of probasin or fibroblast-specific pro-tein 1 (FSP1) promoter. The promoter of probasin-Cres is well characterized and has been widely appliedto delete the floxed gene in several mouse models[25,27,28]. The expression of probasin-Cre is post-natal and prostatic epithelium-specific, and graduallyincreases and has nearly 100% activity in adult mice todelete floxed gene expression (Supporting information,Supplementary Figure 1) [25,27–29]. The expressionof FSP1-Cre is first detected in the cells of the fibrob-lastic phenotype after embryonic day 8.5 [31–33].FSP1-Cre is able to knock out ∼65% of floxed genesin the mouse prostate stromal compartment (Sup-porting information, Supplementary Figure 2) and hasbeen widely used to knock out a variety of targetgenes [23,34]. We generated three ERαKO mousemodels: (i) flox ERα gene deletion in every tissueusing ACTB-Cre (ACTB-ERαKO); (ii) selective dele-tion in stromal cells by FSP1-Cre (FSP-ERαKO); and(iii) selective deletion in prostate epithelial cells byprobasin-Cre (pes-ERαKO), to determine the tempo-ral and spatial requirements of ERα in the oestrogen-stimulated proliferation and SQM in the prostate.Our data indicate that FSP-ERαKO prostates haveless branching morphogenesis compared with Wt lit-termates. In contrast, pes-ERαKO mice develop anormal prostate [35]. We chose 10- to 12-week-oldadult mice to perform our experiments since theexpression of Cre develops into full activity in theadult.
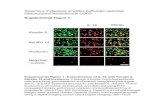
Previous studies have shown that the effects ofDES on the three prostatic lobes reveal a hierar-chy of responses, with the anterior prostate (AP)being the most responsive, the dorso-lateral prostate(DLP) less responsive, and the ventral prostate (VP)the least responsive. Figure 1 shows representativeAP histology sections of cholesterol or DES-treatedWt, ACTB-ERαKO, pes-ERαKO, and FSP-ERαKOmice. The normal prostate ducts were lined by alayer of simple columnar epithelial cells in differentcholesterol-treated mouse genotypes (Figure 1A, a–d).DES treatment resulted in uniform stratified basalepithelial layers in the AP of Wt and FSP-ERαKO mice(Figure 1A, e and g). Those stratified basal epithe-lial cells were positive for the squamous epithe-lial marker CK10 (Figure 1B, e and g), which wasnot present in the cholesterol-treated mouse prostates(Figure 1B, a–d). In contrast, DES-treated ACTB-ERαKO prostates appeared histologically identical tocholesterol-treated ACTB-ERαKO prostates and did
not express CK10 (Figures 1A and 1B, f versus b),which is consistent with previous studies showing thatconventional ERαKO-neo knock-in mice (αERKO)do not respond to DES treatment [19,21]. Impor-tantly, our data show that loss of prostate epithe-lial ERα inhibited SQM in the pes-ERαKO miceas the prostatic ducts from DES-treated pes-ERαKOmice were mainly lined by a single layer of epithe-lial cells (Figure 1A, h versus d), in which CK10was either not detected or only detected focally ina single layer (Figure 1B, h versus d). These resid-ual CK10-positive cells could be due to not enoughprobasin-Cre activity in some prostate basal epithe-lial cells to knock out floxed ERα gene. Our cur-rent studies confirmed that ERα is required for anoestrogen-mediated response in the mouse prostate[19,21]. Furthermore, epithelial ERα, and not stromal,is required to mediate this oestrogenic effect in themurine prostate.
Decreased SQM phenotype and basal cellproliferation in DES-treated pes-ERαKO prostatesTo better characterize SQM defects and cell typechanges in DES-treated pes-ERαKO and Wt mice,we further used IHC and real-time quantitative PCR(Q-PCR, methods in Supporting Information, Sup-plementary materials and methods) assays to detectCK5, CK10, and p63 markers. CK10 is a markerfor SQM cells. CK5 and p63 are markers for basalepithelial cells. As indicated by both IHC and Q-PCR analyses, the percentage of cells expressing CK5,CK10, and p63 increased profoundly in DES-treatedWt mice; however, the changes in DES-treated pes-ERαKO mice were much smaller compared with thosein DES-treated Wt mice (Figures 2A–2D). Mean-while, we also co-stained for CK5/CK10, ERα/p63, andERα/CK10 to examine how these markers co-localizeand spatially change during SQM. The normal mouseprostate contains few basal cells, which are intermit-tently expressed at the basement membrane in bothvehicle (cholesterol)-treated Wt and pes-ERαKO con-trol mice (Figures 3A and 3B, a and f, arrows), andCK10 is not expressed in those basal cells (Figures 3Aand 3B, b). In Wt mice, DES treatment resultedin CK5-positive basal cell expansion, stratification,and multi-layering (Figure 3C, a). These CK5-positivebasal cells also expressed the squamous epithelialmarker CK10 (Figure 3C, b). In contrast, pes-ERαKOmice responses to DES had dramatically decreasedbasal cell expansion compared with DES-treated Wtmice (Figure 3D, a versus Figure 3C, a). DES treat-ment only resulted in a continuous, but not stratified,CK5-positive basal layer in the pes-ERαKO prostates(Figure 3D, a). Consistently, limited expansion ofbasal-type cells in DES pes-ERαKO prostates was alsoobserved when stained with another basal cell marker,p63 (Figure 3D, f versus Figure 3C, f). Furthermore,only a few CK5-positive cells co-expressed CK10 inthe DES-treated pes-ERαKO prostates (Figure 3D, b,
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

20 M Chen et al
Figure 1. PES-ERαKO mice fail to respond to DES treatment. H&E staining (A) and IHC staining of CK10 (B) from Wt, ACTB-ERαKO,FSP-ERαKO, and pes-ERαKO AP tissue treated with vehicle (cholesterol) or DES. Note that DES treatment in Wt and FSP-ERαKO miceresulted in a uniform stratified basal epithelial layer, in which the squamous epithelial marker, CK10, was expressed (A and B, e and g), butnot in the DES-treated ACTB-ERαKO and pes-ERαKO mice (A and B, f and h).
arrowheads), which is consistent with the results inFigure 2C showing that the up-regulation of CK10-positive cells is much less than that in DES-treatedWt prostates. Altogether, our results indicate thatloss of epithelial ERα in the pes-ERαKO prostatesinhibits basal cell expansion and the SQM pheno-type, as evidenced by significantly decreased CK10-positive squamous cells and CK5 (or p63)-positivebasal cells.
Oestrogen exposure stimulates ERα expressionin the luminal and basal epithelial cells in Wtprostates, but not in pes-ERαKO mouse prostates
Previous studies have shown that the induction of SQMis accompanied by the up-regulation of ERα expres-sion in the prostate epithelial cells, which suggeststhat ERα might be involved in oestrogen-mediatedSQM [21]. Therefore, we were interested in examin-ing how ERα changes in the pes-ERαKO prostates inresponse to DES. The prostate epithelial compartmentcontains two major types of cells, luminal cells andbasal cells, and a few intermediate cells. We foundthat ERα is expressed in the luminal cells, but notin the basal cells of Wt prostates, as ERα is not co-expressed with the basal cell marker p63 (Figure 3A, eand f, arrows). As expected, there was almost no
ERα expression in the epithelial compartment of pes-ERαKO prostates (Figure 3B, e and i). In Wt mice,DES treatment resulted in up-regulation of ERα notonly in the luminal cells, but also in the basal cells aswell as squamous cells, as some of the ERα-positivecells also co-expressed p63 or CK10 in the basal layer(Figure 3C, g and k, arrowheads). In contrast, epithe-lial ERα up-regulation was not observed in DES-treatedpes-ERαKO prostates (Figure 3D, e and i). We furthervalidated the expression level and the distributed celltypes of ERα in DES-treated Wt versus pes-ERαKOusing IHC staining. Consistent with the immunoflu-orescence results in Figures 3C and 3D, ERα was up-regulated in Wt mice after DES treatment (Figure 4B, aand b, c and d, arrows), but ERα was rarely expressedand not up-regulated in DES-treated pes-ERαKO mice(Figure 4B, e and f, g and h, arrows). In addition, ourQ-PCR results indicated that although the level of ERα
mRNA was up-regulated in both DES-treated Wt andpes-ERαKO mice, there was significantly less ERα
mRNA in DES-treated pes-ERαKO mice than in DES-treated Wt mice (Figure 4B). Our results indicate thatthe inhibited SQM phenotype is due to loss of prostateepithelial ERα in DES-treated pes-ERαKO mice. Thisconclusion is supported by up-regulation ERα at bothmRNA and protein level in DES-treated pes-ERα KOprostate basal cells.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

ERα and metaplasia in the prostate 21
Figure 2. IHC staining and Q-PCR quantification of the cytokeratin and basal cell markers in the prostates of DES- and vehicle-treatedWt and pes-ERαKO mice. Results were examined and quantified by using both IHC staining (A–C), and real-time Q-PCR assay (D). Thequantified IHC data are presented as % of positive epithelial cells over total epithelial cells. Five independent fields were counted fromeach stained prostate sample. Note the percentage of cells expressing CK5 (A), p63 (B), and CK10 (C) and the relative levels of thosemarkers, by IHC and Q-PCR, are profoundly up-regulated in DES-treated Wt mice; however, the changes in DES-treated pes-ERαKO miceare significantly smaller (A–D). p < 0.05, ∗compared with cholesterol-treated mice; # compared with DES-treated Wt mice.
DES-treated pes-ERαKO prostates have significantlyreduced basal cell growth compared with thatof DES-treated Wt prostatesDES-stimulated prostate proliferation is further charac-terized by IHC staining of Ki67 and p27Kip1. Staining
of the cell proliferation marker Ki67 indicated that theadult prostate is a quiescent organ. Ki67-positive prolif-erative cells were rarely detected in vehicle-treated Wtand pes-ERαKO mice (Figure 4A, a and b, e and f). Inthe Wt mice, DES stimulated the cell proliferation.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

22 M Chen et al
This proliferation was mainly located in the basal cellcompartment (Figure 4A, c and d, arrows). In addition,in DES-treated Wt mice, the stratified squamous/basalcells lacked expression of the cell cycle inhibitorp27Kip1 (Figure 4A, l, arrowheads), which was stillmaintained and expressed in quiescent luminary epithe-lial cells in vehicle-treated prostates (Figure 4A, j,arrows). The results indicate that DES induces the
proliferation of basal cells, with concomitant loss ofp27Kip1. In contrast to DES-treated Wt mice, prostatecell proliferation was rarely observed in the DES-treated pes-ERαKO mice (Figure 4A, g and h), andp27Kip1 still remained positive in the majority ofepithelial cells (Figure 4A, m and n, o and p, arrows).These staining data indicate that the DES-treated pes-ERαKO prostates remain quiescent, suggesting that
Figure 3. Immunofluorescence double staining of cytokeratin marker and ERα in the prostates of Wt and pes-ERαKO mice.Immunofluorescence analyses were carried out using AP tissue sections from vehicle- and DES-treated Wt (A, C) and pes-ERαKOmice (B, D). The tissue sections were double-stained for CK5/CK10, ERα/p63, and ERα/CK10, followed by counterstaining with DAPIto visualize the nuclei. Arrows show the cells that only expressed indicated genes. Arrowheads show the cells that co-express thedouble-staining genes. c, g, and k are merged images. d, h, and i are for DAPI staining. The others are the images for the expression ofspecific genes. Note that in vehicle-treated Wt and pes-ERαKO mice, basal cells are intermittently scattered at the basement membraneas indicated by CK5 and p63 staining (A and B, a and f), and CK10 is not expressed in those basal cells (A and B, b). ERα is expressed in theluminal cells, but not in the basal cells, of Wt prostates, as ERα is not co-expressed with the basal cell marker p63 (A, e–g). There is almostno detectable epithelial ERα expression in pes-ERαKO prostates (B, e and i). In DES-treated Wt prostates, CK5-positive cells are stratifiedand co-express the squamous epithelial marker CK10 (C, a–c). In contrast, in DES-treated pes-ERαKO prostates, DES treatment resultedin a continuous, but not stratified, CK5-positive basal layer, in which only a few cells expressed CK10 (D, a–c). p63 staining results alsoindicated that basal cell expansion occurs focally in the single-layer basal epithelial cells of pes-ERαKO prostates, but it occurs in themulti-layer basal epithelial cells of Wt prostates (D, f versus C, f). ERα is up-regulated in both luminal and basal cells in DES-treated Wtmice (C, e and i, arrows and arrowheads, respectively), but not in DES-treated pes-ERαKO mice (D, e and i).
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

ERα and metaplasia in the prostate 23
Figure 3. (Continued).
epithelial ERα is important for responding to theoestrogen-stimulated proliferative event in the mouseprostate.
Discussion
We generated mouse models that have selectivelylost ERα in either prostate epithelial or stromalcells to study their respective roles in the adultprostate in response to oestrogen. Our unpublished dataacquired during characterization of male reproductiveand normal prostate development in FSP-ERαKO andpes-ERαKO mice indicated that the circulating lev-els of testosterone and fertility are not altered in themale mice of either model. However, FSP-ERαKOprostates have less branching morphogenesis comparedwith Wt littermates. In contrast, pes-ERαKO micemaintain normal homeostasis and develop a normalprostate without changing branching morphogenesis
[35], highlighting the critical role of stromal ERα inprostate development.
In our present study, we found that FSP-ERαKOprostates develop full and uniform SQM as in Wtmice after DES treatment, which indicates that lossof the majority (∼65%) of stromal ERα results indevelopmental defects with reduced prostate branching,yet does not influence oestrogen-stimulated SQM in theprostate. However, full and uniform SQM observedin FSP-ERαKO prostates may also suggest that theremainder of stromal ERα retains sufficient capacityto support oestrogen-mediated SQM in the prostate.Further studies utilizing more efficient stromal Cretransgenic mice are needed to confirm the roles ofstromal ERα in oestrogen-mediated SQM in vivo. Incontrast to FSP-ERαKO mice, loss of epithelial ERα
in the pes-ERαKO prostates causes them to maintaina normal prostate histological structure, yet results ininhibition of oestrogen-stimulated SQM, in which theprostatic ducts were lined by a single layer of columnar
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

24 M Chen et al
Figure 4. The expression of cell growth markers and ERα of Wt and pes-ERαKO prostates was examined by IHC staining. IHC staining ofKi67, p27Kip1, and ERα was carried out using AP tissue sections of cholesterol- (vehicle) or DES-treated Wt and pes-ERαKO mice. Arrowsindicate cells positive for the gene expression. Note that basal cells in DES-treated Wt prostates have increased cell proliferation, asindicated by Ki67 staining (c versus a; d versus b, arrows), but decreased p27Kip1 expression (k versus i; l versus j, arrowheads), in contrastto constantly positive p27Kip1 expression in the luminary epithelial cells (j and l, arrows). However, DES-treated pes-ERαKO prostatesrarely proliferate (g versus e; h versus f) and also constantly maintain high p27Kip1 expression in the prostate epithelial cells before andafter DES treatment (o versus m; p versus n). Up-regulation of ERα was observed in the Wt prostate epithelial cells after DES treatment(B, c versus a; d versus b), but not in the DES-treated pes-ERαKO prostate (B, g versus e; h versus f). The mRNA expression of ERα incholesterol- and DES-treated mouse prostates was examined further by real-time Q-PCR. Note that the level of ERα mRNA is profoundlyup-regulated in DES-treated Wt prostates; the increase in DES-treated pes-ERαKO prostates is much smaller. ∗Compared with control Wtmice; # compared with control Wt and pes-ERKO mice; δ compared with DES-treated Wt mice.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

ERα and metaplasia in the prostate 25
epithelial cells and exhibited rare and focal CK10staining (Figure 1). Together, our Cre-loxP modelsprovide direct in vivo evidence that epithelial ERα isdispensable for prostate development, yet is critical foroestrogen-mediated prostate proliferation and SQM inthe adult prostate.
Further characterization of SQM in Wt mice showedthat DES treatment resulted in CK5-positive basal cellexpansion, stratification, and CK10 squamous cell dif-ferentiation (Figure 3C). However, pes-ERαKO micefailed to develop full and uniform SQM as evidencedby a continuous, but not stratified, CK5-positive basallayer, with very few CK5-positive cells co-expressingCK10 (Figure 3D). These results indicate that loss ofepithelial ERα in the prostate inhibits both basal cellproliferation and squamous cell differentiation. Eventhough both prostatic luminal and basal cells are knownto express significant levels of probasin-Cre [28–30],the few residual CK10-positive cells observed in theDES-treated pes-ERαKO prostates could be due toincomplete knockout of ERα in prostate basal cells byprobasin-Cre.
The other characteristic of SQM is the up-regulationof ERα in both prostatic luminal and basal cells(Figure 3C). One interesting observation is that ERα
expression is not found in the basal cells of normalWt prostates (Figure 3A), but it appears in the basalcells of DES-treated Wt prostates (Figure 3C), whichcould directly contribute to the response of the basalcells to oestrogens and result in basal cell prolifer-ation and metaplasia. In contrast, epithelial ERαKOprostates have a profoundly reduced ERα up-regulationin response to DES treatment, which could contributeto the phenotypic loss of SQM in pes-ERαKO mice(Figures 3D and 4). The underlying mechanisms bywhich ERα promotes basal cell proliferation and squa-mous cell differentiation need to be further investi-gated.
The finding that DES-treated pes-ERαKO mice lackSQM is also supported by SQM biomarker stain-ing. The adult rodent prostate is a growth quiescentorgan. One of the characteristics of SQM is oestrogen-induced basal proliferation. In the Wt males, lumi-nal epithelial cells were positive for the cell cycleinhibitor p27 and were growth quiescent. After DEStreatment, a continuous layer of proliferating basalcells emerged and was positive for Ki67, whereas p27expression was not detected in the multi-layered basalepithelial cells (Figure 4). In contrast, an increase ofKi67-positive basal cells and loss of p27 were notobserved in the basal cells of DES-treated pes-ERαKOmice (Figure 4). The results indicate that the mito-genic effect of oestrogen in the adult mouse prostateis mediated by epithelial ERα. Although increasedproliferation activity is observed in SQM, SQM isalso characterized by down-regulation of AR expres-sion in DES-treated Wt mice (Supporting informa-tion, Supplementary Figure 3, c versus a; d versus b).In the DES-treated pes-ERαKO mice, the expressionof AR was also significantly inhibited compared with
untreated pes-ERαKO mice (Supporting information,Supplementary Figure 3, g versus e; h versus f), whichindicates that AR down-regulation might not contributedirectly to oestrogen-mediated SQM and proliferativeactivities in the mouse prostate.
Previous studies have also shown that the effects ofDES in the rodent prostate lobes reveal a hierarchy ofresponses, with AP being the most responsive, DLPless responsive, and the VP lobe the least responsive[20]. Indeed, we observed that pes-ERαKO prostatesfailed to develop SQM in all lobes compared withWt mice, which was evident by the complete loss ofCK10 in pes-ERαKO VP and DLP after DES treat-ment (data not shown). Together, our current stud-ies indicate that epithelial ERα plays critical roles inoestrogen-mediated SQM and proliferative activities inthe prostate. An earlier study utilizing tissue recombi-nation techniques with mesenchyme of newborn mouseseminal vesicle (SV) and adult prostatic ductal tipsfrom Wt and ERαKO mice stated that both epithelialand stromal ERα are necessary for oestrogen-mediatedSQM in the prostate. In an attempt to validate the rolesof stromal ERα in SQM using an in vivo FSP-ERαKOmodel, we observed that loss of the majority of stro-mal ERα does not influence oestrogen-mediated SQMin the prostate, but our results might be compounded bythe incomplete ablation of stromal ERα in the mouseprostate. As mentioned above, a more efficient stromalCre transgenic mouse model is needed to confirm therole of stromal ERα in SQM. The advantage of ourin vivo assay is that we utilize intact mice, in whicha functional immune system exists, as an experimen-tal tool. The data acquired during prostatic regrowth incastrated adult mice showed that oestrogen exposureresults in cyclin D1-positive cells in the epitheliumand the recruitment of FGF10-positive immune cellsin prostates, suggesting that the immune system is animportant factor contributing to prostatic regrowth andhomeostasis [13].
In our current model of squamous metaplasia(Figure 5), oestrogens, via ERα, exert direct actions onthe prostate by up-regulating ERα signalling, enhanc-ing basal cell proliferation, and promoting the expan-sion of abnormal squamous cells. In contrast, loss ofERα in the prostate epithelial cells results in the fail-ure of the mouse prostate to respond to oestrogen.As a consequence, DES-treated pes-ERαKO prostatescannot develop full and uniform SQM. These in vivoresults suggest that epithelial ERα is important in themediation of the proliferative response to oestrogen.It has been reported that the expression of ERα wasup-regulated during prostatic pathogenesis, includingBPH and prostate cancer [36–38]. The higher expres-sion of ERα suggests the involvement of oestrogensignalling in the prostatic pathogeneses. Studies havealso shown that during prostatic regrowth, there isa window in time when ERα is up-regulated in theprostatic epithelium between days 3 and 5 after testos-terone replacement and selective stimulation of epithe-lial ERα can induce ductal branching, which suggests
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

26 M Chen et al
Figure 5. The oestrogen-stimulated prostate proliferation and SQM compared in Wt and PES-ERαKO mice. Our current studies indicatethat loss of ERα in the prostate epithelial cells results in the failure of an oestrogen response in the mouse prostate, as demonstratedby limited stratifying basal cells, squamous metaplasia, and up-regulated ERα expression. As a consequence, DES-treated pes-ERαKOprostates have a continuous, but not stratified, CK5-positive basal layer, in which only a few cells express CK10, indicating limited andscattered SQM development. The distribution of different types of prostate cells with different expression of ERα, CK10, CK5, Ki67, andp27Kip1 is depicted in the figure. ∗The majority of the luminal epithelial cells of adult prostate are p27Kip1-positive.
that epithelial ERα is essential for ductal branchingduring postnatal prostate growth and homeostasis [13].Since ERα signalling is involved in the prostatic patho-geneses, how to target ERα has a direct implication forunderstanding prostatic disease and the development ofpharmaceuticals for the treatment of BPH and prostatecancer. Our studies suggest that epithelial ERα in theprostate could be an appropriate target for preventingaberrant oestrogen signalling during prostate pathogen-esis.
AcknowledgmentWe thank Dr Chawnshang Chang for helpful sugges-tions; Drs HL Moses and NA Bhowmick for provid-ing FSP 1-cre transgenic mice, and Karen Wolf forassistance with manuscript preparation. This work wassupported by NCI grant CA137474.
Author contribution statement
SY, MC, and HCC conceived the study design andcarried out data interpretation. MC, CRY, and SSconceived and carried out experiments. All the authorswere involved in data discussion, paper preparation,and approved the final version of the manuscript.
References
1. Cunha GR, Donjacour AA, Cooke PS, et al . The endocrinologyand developmental biology of the prostate. Endocr Rev 1987; 8:338–362.
2. Mawhinney MG, Neubauer BL. Actions of estrogen in the male.Invest Urol 1979; 16: 409–420.
3. Jarred RA, Cancilla B, Prins GS, et al . Evidence that estro-gens directly alter androgen-regulated prostate development.Endocrinology 2000; 141: 3471–3477.
4. Prins GS. Neonatal estrogen exposure induces lobe-specificalterations in adult rat prostate androgen receptor expression.Endocrinology 1992; 130: 3703–3714.
5. Rajfer J, Coffey DS. Sex steroid imprinting of the immatureprostate. Long-term effects. Invest Urol 1978; 16: 186–190.
6. Taylor RA, Cowin P, Couse JF, et al . 17beta-estradiol inducesapoptosis in the developing rodent prostate independently ofERalpha or ERbeta. Endocrinology 2006; 147: 191–200.
7. Couse JF, Korach KS. Estrogen receptor null mice: what havewe learned and where will they lead us? Endocr Rev 1999; 20:358–417.
8. Lin Y, Liu G, Zhang Y, et al . Fibroblast growth factor receptor2 tyrosine kinase is required for prostatic morphogenesis andthe acquisition of strict androgen dependency for adult tissuehomeostasis. Development 2007; 134: 723–734.
9. Lazennec G, Bresson D, Lucas A, et al . ER beta inhibits prolif-eration and invasion of breast cancer cells. Endocrinology 2001;142: 4120–4130.
10. Paech K, Webb P, Kuiper GG, et al . Differential ligand activationof estrogen receptors ERalpha and ERbeta at AP1 sites. Science
1997; 277: 1508–1510.11. Chen M, Hsu I, Wolfe A, et al . Defects of prostate development
and reproductive system in the estrogen receptor-alpha null malemice. Endocrinology 2009; 150: 251–259.
12. Omoto Y, Imamov O, Warner M, et al . Estrogen receptor alphaand imprinting of the neonatal mouse ventral prostate by estrogen.Proc Natl Acad Sci U S A 2005; 102: 1484–1489.
13. Omoto Y. Estrogen receptor-alpha signaling in growth of theventral prostate: comparison of neonatal growth and postcastrationregrowth. Endocrinology 2008; 149: 4421–4427.
14. Dupont S, Krust A, Gansmuller A, et al . Effect of single andcompound knockouts of estrogen receptors alpha (ERalpha) andbeta (ERbeta) on mouse reproductive phenotypes. Development
2000; 127: 4277–4291.15. Weihua Z, Makela S, Andersson LC, et al . A role for estrogen
receptor beta in the regulation of growth of the ventral prostate.Proc Natl Acad Sci U S A 2001; 98: 6330–6335.
16. Krege JH, Hodgin JB, Couse JF, et al . Generation and reproduc-tive phenotypes of mice lacking estrogen receptor beta. Proc Natl
Acad Sci U S A 1998; 95: 15677–15682.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com

ERα and metaplasia in the prostate 27
17. Antal MC, Krust A, Chambon P, et al . Sterility and absence ofhistopathological defects in nonreproductive organs of a mouseERbeta-null mutant. Proc Natl Acad Sci U S A 2008; 105:2433–2438.
18. Prins GS, Birch L. Neonatal estrogen exposure up-regulates estro-gen receptor expression in the developing and adult rat prostatelobes. Endocrinology 1997; 138: 1801–1809.
19. Prins GS, Birch L, Couse JF, et al . Estrogen imprinting of thedeveloping prostate gland is mediated through stromal estrogenreceptor alpha: studies with alphaERKO and betaERKO mice.Cancer Res 2001; 61: 6089–6097.
20. Risbridger GP, Wang H, Frydenberg M, et al . The metaplasticeffects of estrogen on mouse prostate epithelium: proliferationof cells with basal cell phenotype. Endocrinology 2001; 142:2443–2450.
21. Risbridger G, Wang H, Young P, et al . Evidence that epithelial andmesenchymal estrogen receptor-alpha mediates effects of estrogenon prostatic epithelium. Dev Biol 2001; 229: 432–442.
22. Cunha GR, Cooke PS, Kurita T. Role of stromal–epithelial inter-actions in hormonal responses. Arch Histol Cytol 2004; 67:417–434.
23. Bhowmick NA, Chytil A, Plieth D, et al . TGF-beta signaling infibroblasts modulates the oncogenic potential of adjacent epithelia.Science 2004; 303: 848–851.
24. Chen M, Wolfe A, Wang X, et al . Generation and characterizationof a complete null estrogen receptor alpha mouse using Cre/LoxPtechnology. Mol Cell Biochem 2009; 321: 145–153.
25. Wu X, Wu J, Huang J, et al . Generation of a prostate epithelialcell-specific Cre transgenic mouse model for tissue-specific geneablation. Mech Dev 2001; 101: 61–69.
26. Chen M, Ni J, Chang HC, et al . CCDC62/ERAP75 functions as acoactivator to enhance estrogen receptor beta-mediated transacti-vation and target gene expression in prostate cancer cells. Carcino-
genesis 2009; 30: 841–850.27. Wu CT, Altuwaijri S, Ricke WA, et al . Increased prostate cell
proliferation and loss of cell differentiation in mice lacking prostateepithelial androgen receptor. Proc Natl Acad Sci U S A 2007; 104:12679–1268428.
SUPPORTING INFORMATION ON THE INTERNET
The following supporting information may be found in the online version of this article.
Supplementary materials and methods.
Figure S1. Comparison of the expression levels of ERα between Wt and ACTB-ERαKO or pes-ERαKO prostates via real-time Q-PCR.
Figure S2. Comparison of the stromal ERα expression between adult Wt and FSP-ERαKO prostates via IHC staining.
Figure S3. Expression of AR in Wt and pes-ERαKO prostates.
28. Wang S, Gao J, Lei Q, et al . Prostate-specific deletion of themurine Pten tumor suppressor gene leads to metastatic prostatecancer. Cancer Cell 2003; 4: 209–221.
29. Niu Y, Altuwaijri S, Lai KP, et al . Androgen receptor is a tumorsuppressor and proliferator in prostate cancer. Proc Natl Acad Sci
U S A 2008; 34: 12182–12187.30. Wang S, Garcia AJ, Wu M, et al . Pten deletion leads to the
expansion of a prostatic stem/progenitor cell subpopulation andtumor initiation. Proc Natl Acad Sci U S A 2006; 103: 1480–1485.
31. Iwano M, Fischer A, Okada H, et al . Conditional abatement oftissue fibrosis using nucleoside analogs to selectively corrupt DNAreplication in transgenic fibroblasts. Mol Ther 2001; 3: 149–159.
32. Iwano M, Plieth D, Danoff TM, et al . Evidence that fibroblastsderive from epithelium during tissue fibrosis. J Clin Invest 2002;110: 341–350.
33. Strutz F, Okada H, Lo CW, et al . Identification and characteriza-tion of a fibroblast marker: FSP1. J Cell Biol 1995; 130: 393–405.
34. Trimboli AJ, Cantemir-Stone CZ, Li F, et al . Pten in stromalfibroblasts suppresses mammary epithelial tumours. Nature 2009;461: 1084–1091.
35. Chen M, Yeh CR, Yeh S. Reduced branching morphogenesis inmice lacking stromal estrogen receptor alpha but not in micelacking epithelial estrogen receptor alpha. Asian J Androl 2011;(in press).
36. Bonkhoff H, Fixemer T, Hunsicker I, et al . Estrogen receptorexpression in prostate cancer and premalignant prostatic lesions.Am J Pathol 1999; 155: 641–647.
37. Royuela M, de Miguel MP, Bethencourt FR, et al . Estrogen recep-tors alpha and beta in the normal, hyperplastic and carcinomatoushuman prostate. J Endocrinol 2001; 168: 447–454.
38. King KJ, Nicholson HD, Assinder SJ. Effect of increasing ratioof estrogen : androgen on proliferation of normal human prostatestromal and epithelial cells, and the malignant cell line LNCaP.Prostate 2006; 66: 105–114.
Copyright 2011 Pathological Society of Great Britain and Ireland. J Pathol 2012; 226: 17–27Published by John Wiley & Sons, Ltd. www.pathsoc.org.uk www.thejournalofpathology.com







![CK HMW [34βE12], 3X · 34βE12 recognizes cytokeratins (CK) 1, 5, 10 and 14 (1). In normal prostate, 34βE12 typically stains the basal cells of the prostate gland. 34βE12 has been](https://static.fdocument.org/doc/165x107/607a25998ae53d3d892e93b9/ck-hmw-34e12-3x-34e12-recognizes-cytokeratins-ck-1-5-10-and-14-1-in.jpg)









