
Lack of -catenin affects mouse development at gastrulation · bp Bluescript subclone of Otx2cDNA...
9
INTRODUCTION Evidence accumulating during recent years underlines the importance of cadherin cell adhesion molecules in the devel- opment of multicellular organisms (Takeichi, 1991; Kemler, 1992). Although the adhesive function of cadherins has largely been studied in cultured cells, their spatiotemporal expression pattern during early development suggests an essential role in morphogenetic events, such as germ layer formation, gastrula- tion or neural plate induction (Vestweber and Kemler, 1984; Nose and Takeichi, 1986; Hoffman and Balling, 1995). Speci- fically, the requirement of E-cadherin (uvomorulin) for the formation and function of the trophectodermal cell layer at the mouse blastocyst stage has been demonstrated by gene knock- out experiments (Larue et al., 1994). Structure-function analyses of E-cadherin led to the identi- fication of α-, β- and γ-catenin associated with the cytoplas- mic domain of E-cadherin (Ozawa et al., 1989; Nagafuchi and Takeichi, 1989). Catenins are now known to mediate the con- nection of E-cadherin to the actin filament network as part of a higher order submembranous protein complex (for review see Kemler, 1993). Recently, it was shown that β- and γ-catenin become tyrosine-phosphorylated upon epidermal growth factor (EGF) stimulation of epithelial cells. Moreover, a direct asso- ciation of β-catenin with the EGF-receptor (EGF-R) was demonstrated in vitro with recombinant proteins (Hoschuetzky et al., 1994). This and the association of catenins with the tumor suppressor protein APC (Su et al., 1993; Rubinfeld et al., 1993) suggest that catenins are involved in central biolog- ical processes, such as proliferation, adhesion and differen- tiation of cells. Molecular cloning and primary sequence analysis revealed that catenins are homologous to other peripheral cytoplasmic proteins known to be localised in specific cell-cell and/or cell- substratum junctions (Kemler, 1993). β-catenin exhibits high homology to human plakoglobin, a component of desmosomal plaques and adherens junctions (Cowin et al., 1986; Franke et al., 1989), and to the product of the Drosophila segment polarity gene armadillo (Riggleman et al., 1990; Peifer et al., 1992). More recently found members of this protein family include band 6.1 (Hatzfeld et al., 1994), p120 (Reynolds et al., 1994), APC (Kinzler et al., 1991) and SRP1 (Yano et al., 1992). These proteins share a common repeated motif, called the arm repeat, which is thought to be important in mediating specific protein-protein interactions (Peifer et al., 1994a). Studies on the molecular organisation of the E-cadherin- catenin complex in cultured cells (Ozawa and Kemler, 1992) and in vitro using recombinant proteins (Aberle et al., 1994) clearly demonstrated the central role of β-catenin in complex formation. In agreement with this, in the human epithelial cancer cell line, HSC 39, it has been shown that the lack of E- cadherin-catenin complex formation due to the expression of a truncated β-catenin correlates with the loss of cell adhesion (Oyama et al., 1994). Expression of β-catenin in these cells restores both complex formation and cell adhesion (Kawanishi et al., 1995). Plakoglobin is very likely identical to γ-catenin 3529 Development 121, 3529-3537 (1995) Printed in Great Britain © The Company of Biologists Limited 1995 DEV2026 Molecular analysis of the cadherin-catenin complex eluci- dated the central role of β-catenin in this adhesion complex, as it binds to the cytoplasmic domain of E-cadherin and to α-catenin. β-catenin may also function in signalling pathways, given its homology to the gene product of the Drosophila segment polarity gene armadillo, which is known to be involved in the wingless signalling cascade. To study the function of β-catenin during mouse development, gene knock-out experiments were performed in embryonic stem cells and transgenic mice were generated. β-catenin null-mutant embryos formed blastocysts, implanted and developed into egg-cylinder-stage embryos. At day 7 post coitum, the development of the embryonic ectoderm was affected in mutant embryos. Cells detached from the ecto- dermal cell layer and were dispersed into the proamniotic cavity. No mesoderm formation was observed in mutant embryos. The development of extraembryonic structures appeared less dramatically or not at all affected. Our results demonstrate that, although β-catenin is expressed rather ubiquitously, it is specifically required in the ecto- dermal cell layer. Key words: embryonic stem cells, mouse embryogenesis, knock-out mutant, cell adhesion, E-cadherin, β-catenin SUMMARY Lack of β-catenin affects mouse development at gastrulation Hélène Haegel § , Lionel Larue § , Mami Ohsugi, Lev Fedorov † , Kurt Herrenknecht ‡ and Rolf Kemler* Max-Planck-Institut für Immunbiologie, Stübeweg 51, D-79108 Freiburg, Germany *Author for correspondence Present addresses: † Institut für Medizinische Strahlenkunde und Zellforschung (MSZ), Universität Würzburg, Versbacher-Strasse 5, D-97078 Würzburg, Germany ‡ EISAI London Research Laboratories Limited, Bernard Katz Building, UCL, Gower Street, London WC1E 6BT, UK §Both authors contributed equally
Transcript of Lack of -catenin affects mouse development at gastrulation · bp Bluescript subclone of Otx2cDNA...

3529Development 121, 3529-3537 (1995)Printed in Great Britain © The Company of Biologists Limited 1995DEV2026
Lack of β-catenin affects mouse development at gastrulation
Hélène Haegel§, Lionel Larue§, Mami Ohsugi, Lev Fedorov†, Kurt Herrenknecht‡ and Rolf Kemler*
Max-Planck-Institut für Immunbiologie, Stübeweg 51, D-79108 Freiburg, Germany
*Author for correspondencePresent addresses: †Institut für Medizinische Strahlenkunde und Zellforschung (MSZ), Universität Würzburg, Versbacher-Strasse 5, D-97078 Würzburg, Germany‡EISAI London Research Laboratories Limited, Bernard Katz Building, UCL, Gower Street, London WC1E 6BT, UK§Both authors contributed equally
Molecular analysis of the cadherin-catenin complex eluci-dated the central role of β-catenin in this adhesion complex,as it binds to the cytoplasmic domain of E-cadherin and toα-catenin. β-catenin may also function in signallingpathways, given its homology to the gene product of theDrosophila segment polarity gene armadillo, which isknown to be involved in the wingless signalling cascade. Tostudy the function of β-catenin during mouse development,gene knock-out experiments were performed in embryonicstem cells and transgenic mice were generated. β-cateninnull-mutant embryos formed blastocysts, implanted anddeveloped into egg-cylinder-stage embryos. At day 7 post
coitum, the development of the embryonic ectoderm wasaffected in mutant embryos. Cells detached from the ecto-dermal cell layer and were dispersed into the proamnioticcavity. No mesoderm formation was observed in mutantembryos. The development of extraembryonic structuresappeared less dramatically or not at all affected. Ourresults demonstrate that, although β-catenin is expressedrather ubiquitously, it is specifically required in the ecto-dermal cell layer.
Key words: embryonic stem cells, mouse embryogenesis, knock-outmutant, cell adhesion, E-cadherin, β-catenin
SUMMARY
INTRODUCTION
Evidence accumulating during recent years underlines theimportance of cadherin cell adhesion molecules in the devel-opment of multicellular organisms (Takeichi, 1991; Kemler,1992). Although the adhesive function of cadherins has largelybeen studied in cultured cells, their spatiotemporal expressionpattern during early development suggests an essential role inmorphogenetic events, such as germ layer formation, gastrula-tion or neural plate induction (Vestweber and Kemler, 1984;Nose and Takeichi, 1986; Hoffman and Balling, 1995). Speci-fically, the requirement of E-cadherin (uvomorulin) for theformation and function of the trophectodermal cell layer at themouse blastocyst stage has been demonstrated by gene knock-out experiments (Larue et al., 1994).
Structure-function analyses of E-cadherin led to the identi-fication of α-, β- and γ-catenin associated with the cytoplas-mic domain of E-cadherin (Ozawa et al., 1989; Nagafuchi andTakeichi, 1989). Catenins are now known to mediate the con-nection of E-cadherin to the actin filament network as part ofa higher order submembranous protein complex (for review seeKemler, 1993). Recently, it was shown that β- and γ-cateninbecome tyrosine-phosphorylated upon epidermal growth factor(EGF) stimulation of epithelial cells. Moreover, a direct asso-ciation of β-catenin with the EGF-receptor (EGF-R) wasdemonstrated in vitro with recombinant proteins (Hoschuetzkyet al., 1994). This and the association of catenins with thetumor suppressor protein APC (Su et al., 1993; Rubinfeld et
al., 1993) suggest that catenins are involved in central biolog-ical processes, such as proliferation, adhesion and differen-tiation of cells.
Molecular cloning and primary sequence analysis revealedthat catenins are homologous to other peripheral cytoplasmicproteins known to be localised in specific cell-cell and/or cell-substratum junctions (Kemler, 1993). β-catenin exhibits highhomology to human plakoglobin, a component of desmosomalplaques and adherens junctions (Cowin et al., 1986; Franke etal., 1989), and to the product of the Drosophila segmentpolarity gene armadillo (Riggleman et al., 1990; Peifer et al.,1992). More recently found members of this protein familyinclude band 6.1 (Hatzfeld et al., 1994), p120 (Reynolds et al.,1994), APC (Kinzler et al., 1991) and SRP1 (Yano et al.,1992). These proteins share a common repeated motif, calledthe arm repeat, which is thought to be important in mediatingspecific protein-protein interactions (Peifer et al., 1994a).
Studies on the molecular organisation of the E-cadherin-catenin complex in cultured cells (Ozawa and Kemler, 1992)and in vitro using recombinant proteins (Aberle et al., 1994)clearly demonstrated the central role of β-catenin in complexformation. In agreement with this, in the human epithelialcancer cell line, HSC 39, it has been shown that the lack of E-cadherin-catenin complex formation due to the expression ofa truncated β-catenin correlates with the loss of cell adhesion(Oyama et al., 1994). Expression of β-catenin in these cellsrestores both complex formation and cell adhesion (Kawanishiet al., 1995). Plakoglobin is very likely identical to γ-catenin

3530 H. Haegel and others
and, like β-catenin, it binds to the cytoplasmic domain of E-cadherin (Knudsen and Wheelock, 1992; Aberle et al., 1994).Moreover, two distinct E-cadherin-catenin complexes can befound in the same cell, one composed of E-cadherin, α- and β-catenin, the other of E-cadherin, α-catenin and plakoglobin(Butz and Kemler, 1994; Hinck et al., 1994a). The biologicalsignificance of these two distinct cadherin-catenin complexesis not yet well understood, but these experiments suggest thatβ-catenin can be substituted for by plakoglobin.
Analysis of catenin expression during early Xenopus(Schneider et al., 1993; Fagotto and Gumbiner, 1994) andmouse development (Butz and Larue, unpublished data)reveals that catenins have a rather ubiquitous distribution, sug-gesting an association of catenins with the various cell type-specific cadherins during development. More recently apossible signalling function of β-catenin was suggested byexperiments studying the effect of overexpression and under-expression of β-catenin during Xenopus development(Heasman et al., 1994; Funayama et al., 1995). The possibilitythat β-catenin might act as a signalling molecule is supportedby its homology to Drosophila Armadillo, which is known toparticipate in the signalling cascade initiated by Wingless thatleads to establishing segment polarity during Drosophilaembryogenesis (Riggleman et al., 1990; Nordemeer et al.,1994; Peifer et al., 1994b; Peifer, 1995). To gain insights intothe function of β-catenin during mouse embryonic develop-ment, we have inactivated β-catenin by gene targetting exper-iments in embryonic stem (ES) cells and have generated trans-genic mice. We describe here the phenotype of embryosdeficient in β-catenin.
MATERIALS AND METHODS
Construction of the targetting vectorGenomic DNA corresponding to the β-catenin locus was isolatedfrom a λ-EMBL3 phage library made from 129/Sv mouse DNA. A 7kB EcoRI fragment containing exons 2 to 8 of β-catenin wassubcloned in pBluescript KS+ (Stratagene). A restriction map wasestablished, and exon-intron boundaries were determined by sequenceanalysis.
A 1.1 kb SphI fragment was deleted, removing exon 2 and part ofexon 3 of β-catenin. It was replaced by the 4.5 kb XbaI fragment fromplasmid IRES-βgeo (kind gift from Dr A. Smith and Dr P. Soriano),containing the translational initiator IRES (Pelletier and Sonenberg,1988) placed upstream of the promoterless fusion gene β-galactosi-dase-neomycin (β-geo) (Friedrich and Soriano, 1991).
Electroporation, selection and screening of ES cell clonesR1 ES cells (Nagy et al. 1993) were cultured in the presence ofneomycin-resistant embryonic fibroblasts as feeder cells inDulbecco’s Modified Eagle’s Medium (DMEM) supplemented with15% fetal calf serum (FCS), 0.1 mM non-essential amino acids(Gibco), 0.1 mM β-mercaptoethanol, 50 U/ml penicillin, 50 µg/mlstreptomycin and 1000 U/ml leukemia inhibitory factor (LIF)(ESGRO, Gibco). 3×107 cells were electroporated with 30 µg of XbaI-linearized targetting construct using a Bio-Rad Gene Pulser (500 µF,240 V). Recombinant neomycin-resistant clones were selected in thepresence of 400 µg/ml G418 (Gibco) for 10 days, transferred into 24-well plates (Nunc) and expanded in 60 mm culture dishes (Falcon).ES cells were passaged on gelatin-coated dishes in 30% Buffalo RatLiver (BRL) cell-conditioned medium (Smith and Hooper, 1987)without feeder cells, to screen for homologous recombination events
by Southern blot analysis. The probes used were a XbaI-EcoRIfragment containing exons 7 and 8 and a BamHI-XbaI fragment con-taining the neomycin-resistance gene, as indicated in Fig. 1C.
Generation of β-catenin knock-out miceES cells which had undergone homologous recombination at the β-catenin locus were microinjected into blastocysts from (C57BL/6 ×DBA/2) superovulated mice. Male chimeras, as judged from the coatcolor, were bred with C57BL/6 females to test for germline trans-mission. Tail DNA from agouti offspring was used to test for thepresence of the mutation by Southern blot analysis. Germlinechimeras were crossed with 129/Sv females to put the mutation on aninbred background. Offspring were routinely tested for the lacZ geneby X-Gal staining of ear-clips (Felthaus et al., 1994), and those thatwere used for maintenance of the mouse line were also genotyped bya Southern blot of tail DNA.
PCR analysis of β-catenin genotypesExtracts were prepared from either neonate tails, dissected day 7.5p.c. embryos, scraped paraffin sections or in vitro cultured blastocysts.Samples suspended in 10-20 µl of phosphate-buffered saline(PBS)/H2O (1:1) were boiled for 10 minutes, cooled on ice andincubated for 3 hours to overnight in the presence of 1 µg/µl pro-teinase K (Boehringer). After an additional 10 minutes boiling, 5 µlof supernatants were subjected to PCR amplification.
Primers for the PCR reaction were from the neo gene (B: 5′-CTTC-CTCGTGCTTTACGGTA-3′) and the 5′ and 3′ portions of exon 3 ofβ-catenin (A: 5′-ATATTGACGGGCAGTATGCA-3′ and C: 5′-TCAAACTGCGTGGATGGGAT-3′, see scheme in Fig. 3A). ThePCR mix consisted of 250 pM of each oligonucleotide, 400 µMdNTPs (Pharmacia), 1.5 mM MgCl2 and 2.5 U Taq polymerase in Taqbuffer (USB) in a final volume of 50 µl. Temperature cycling con-ditions were at 95°C for 40 seconds, 55°C for 30 seconds and 2minutes at 72°C for 35 cycles. PCR products were resolved on 2%agarose gels.
Histological analysis of mutant embryos and in situhybridisationEmbryos within their deciduae were fixed with 4% paraformaldehydein PBS overnight at 4°C, washed in PBS overnight at 4°C, dehydratedin graded alcohols, embedded in paraffin, sectioned at 7 µm, andstained with hematoxylin and eosin.
For in situ hybridisation, digoxygenin-labelled riboprobes wereprepared from plasmids pMash-2 (gift from Dr F. Guillemot), a 273bp Bluescript subclone of Otx2 cDNA (gift from Dr P. Brûlet) and a1.8 kb Bluescript subclone of T-Brachyury (gift from Dr B.Herrmann), using a DIG-RNA Labelling Kit (Boehringer lot no.1175025). In situ hybridisation was performed as previouslydescribed (Jostarndt and Puntschart 1994) using the DIG Nucleic AcidDetection Kit (Boehringer lot no. 1175041). Slides were lightly coun-terstained with eosin.
In vitro culture of preimplantation embryosβ-cat +/− offspring were crossed inter se after superovulation of 7- to9-week-old females. At day 1.5 or 2.5 p.c. (vaginal plugs were scoredas day 0.5 p.c.), embryos were collected from the oviducts in flushingmedium (Spindle, 1980) and cultured in TE medium (Yamamura andSpindle, 1988) at 37°C in 5% CO2 atmosphere. At day 3.5 p.c.,embryos were transferred to R1 medium (ES cell culture medium)without LIF, in 10% CO2 atmosphere. Photographs were taken every6 hours using an Axiovert 35 phase-contrast microscope (Zeiss).Embryos at day 5.5 or 8.5 p.c. were fixed and immunostained withantibodies directed to β-catenin, E-cadherin or plakoglobin.
For blastocyst outgrowth experiments, embryos were collected atday 2.5 p.c., freed from their zonae pellucidae by treatment with tyroidacid, transferred on gelatin-coated Petriperm dishes (Heraeus) andindividually cultured in ES cell culture medium without LIF in 10%
3531β-catenin and mouse development at gastrulation
IRES betageo
ES 3 4 5
E ES S2 3 4 5 6 7 8
A
B
C
ES
IRES betageo
E S E
neo b-cat
S 3 4 5 6 7 8
1
1
1 kBXX
X
X
6S
E
EE
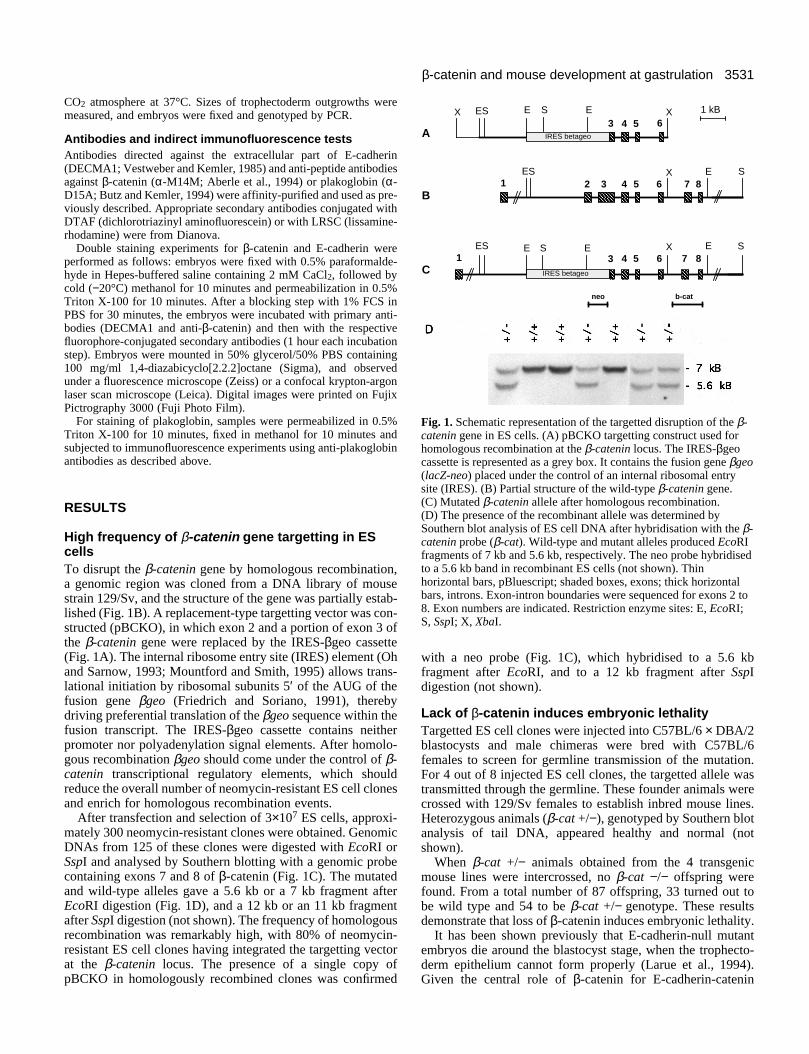
Fig. 1. Schematic representation of the targetted disruption of the β-catenin gene in ES cells. (A) pBCKO targetting construct used forhomologous recombination at the β-catenin locus. The IRES-βgeocassette is represented as a grey box. It contains the fusion gene βgeo(lacZ-neo) placed under the control of an internal ribosomal entrysite (IRES). (B) Partial structure of the wild-type β-catenin gene.(C) Mutated β-catenin allele after homologous recombination.(D) The presence of the recombinant allele was determined bySouthern blot analysis of ES cell DNA after hybridisation with the β-catenin probe (β-cat). Wild-type and mutant alleles produced EcoRIfragments of 7 kb and 5.6 kb, respectively. The neo probe hybridisedto a 5.6 kb band in recombinant ES cells (not shown). Thinhorizontal bars, pBluescript; shaded boxes, exons; thick horizontalbars, introns. Exon-intron boundaries were sequenced for exons 2 to8. Exon numbers are indicated. Restriction enzyme sites: E, EcoRI;S, SspI; X, XbaI.
CO2 atmosphere at 37°C. Sizes of trophectoderm outgrowths weremeasured, and embryos were fixed and genotyped by PCR.
Antibodies and indirect immunofluorescence testsAntibodies directed against the extracellular part of E-cadherin(DECMA1; Vestweber and Kemler, 1985) and anti-peptide antibodiesagainst β-catenin (α-M14M; Aberle et al., 1994) or plakoglobin (α-D15A; Butz and Kemler, 1994) were affinity-purified and used as pre-viously described. Appropriate secondary antibodies conjugated withDTAF (dichlorotriazinyl aminofluorescein) or with LRSC (lissamine-rhodamine) were from Dianova.
Double staining experiments for β-catenin and E-cadherin wereperformed as follows: embryos were fixed with 0.5% paraformalde-hyde in Hepes-buffered saline containing 2 mM CaCl2, followed bycold (−20°C) methanol for 10 minutes and permeabilization in 0.5%Triton X-100 for 10 minutes. After a blocking step with 1% FCS inPBS for 30 minutes, the embryos were incubated with primary anti-bodies (DECMA1 and anti-β-catenin) and then with the respectivefluorophore-conjugated secondary antibodies (1 hour each incubationstep). Embryos were mounted in 50% glycerol/50% PBS containing100 mg/ml 1,4-diazabicyclo[2.2.2]octane (Sigma), and observedunder a fluorescence microscope (Zeiss) or a confocal krypton-argonlaser scan microscope (Leica). Digital images were printed on FujixPictrography 3000 (Fuji Photo Film).
For staining of plakoglobin, samples were permeabilized in 0.5%Triton X-100 for 10 minutes, fixed in methanol for 10 minutes andsubjected to immunofluorescence experiments using anti-plakoglobinantibodies as described above.
RESULTS
High frequency of β-catenin gene targetting in EScellsTo disrupt the β-catenin gene by homologous recombination,a genomic region was cloned from a DNA library of mousestrain 129/Sv, and the structure of the gene was partially estab-lished (Fig. 1B). A replacement-type targetting vector was con-structed (pBCKO), in which exon 2 and a portion of exon 3 ofthe β-catenin gene were replaced by the IRES-βgeo cassette(Fig. 1A). The internal ribosome entry site (IRES) element (Ohand Sarnow, 1993; Mountford and Smith, 1995) allows trans-lational initiation by ribosomal subunits 5′ of the AUG of thefusion gene βgeo (Friedrich and Soriano, 1991), therebydriving preferential translation of the βgeo sequence within thefusion transcript. The IRES-βgeo cassette contains neitherpromoter nor polyadenylation signal elements. After homolo-gous recombination βgeo should come under the control of β-catenin transcriptional regulatory elements, which shouldreduce the overall number of neomycin-resistant ES cell clonesand enrich for homologous recombination events.
After transfection and selection of 3×107 ES cells, approxi-mately 300 neomycin-resistant clones were obtained. GenomicDNAs from 125 of these clones were digested with EcoRI orSspI and analysed by Southern blotting with a genomic probecontaining exons 7 and 8 of β-catenin (Fig. 1C). The mutatedand wild-type alleles gave a 5.6 kb or a 7 kb fragment afterEcoRI digestion (Fig. 1D), and a 12 kb or an 11 kb fragmentafter SspI digestion (not shown). The frequency of homologousrecombination was remarkably high, with 80% of neomycin-resistant ES cell clones having integrated the targetting vectorat the β-catenin locus. The presence of a single copy ofpBCKO in homologously recombined clones was confirmed
with a neo probe (Fig. 1C), which hybridised to a 5.6 kbfragment after EcoRI, and to a 12 kb fragment after SspIdigestion (not shown).
Lack of β-catenin induces embryonic lethalityTargetted ES cell clones were injected into C57BL/6 × DBA/2blastocysts and male chimeras were bred with C57BL/6females to screen for germline transmission of the mutation.For 4 out of 8 injected ES cell clones, the targetted allele wastransmitted through the germline. These founder animals werecrossed with 129/Sv females to establish inbred mouse lines.Heterozygous animals (β-cat +/−), genotyped by Southern blotanalysis of tail DNA, appeared healthy and normal (notshown).
When β-cat +/− animals obtained from the 4 transgenicmouse lines were intercrossed, no β-cat −/− offspring werefound. From a total number of 87 offspring, 33 turned out tobe wild type and 54 to be β-cat +/− genotype. These resultsdemonstrate that loss of β-catenin induces embryonic lethality.
It has been shown previously that E-cadherin-null mutantembryos die around the blastocyst stage, when the trophecto-derm epithelium cannot form properly (Larue et al., 1994).Given the central role of β-catenin for E-cadherin-catenin
3532 H. Haegel and others
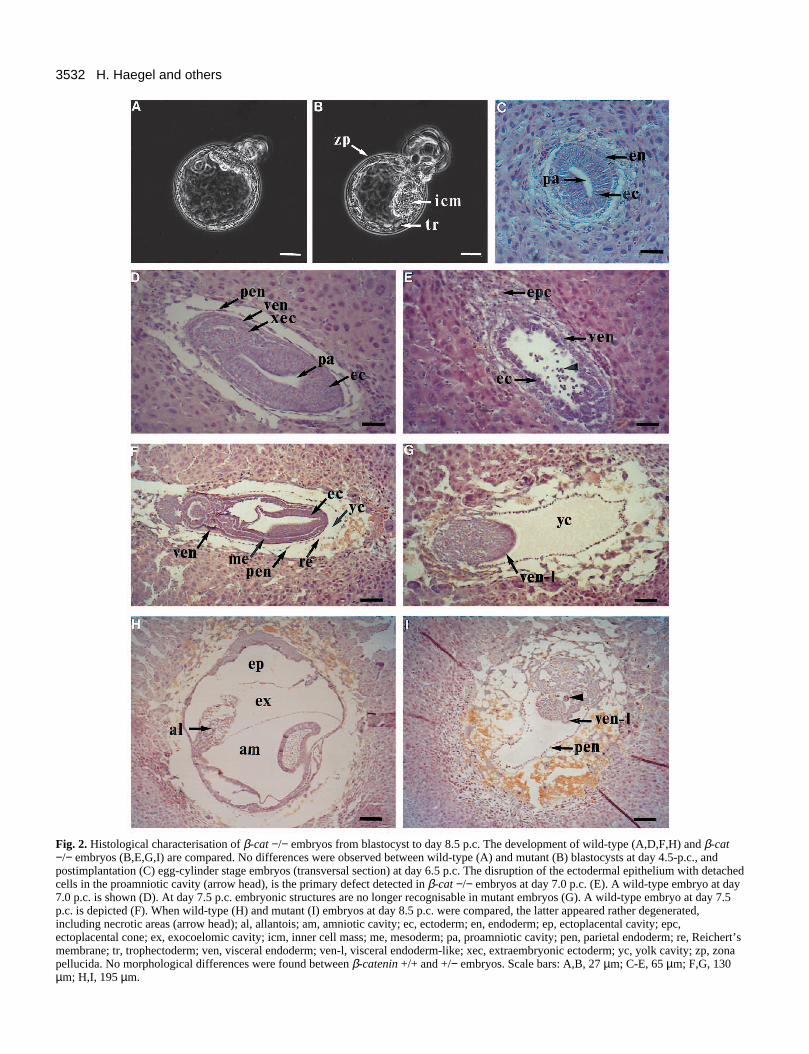
Fig. 2. Histological characterisation of β-cat −/− embryos from blastocyst to day 8.5 p.c. The development of wild-type (A,D,F,H) and β-cat−/− embryos (B,E,G,I) are compared. No differences were observed between wild-type (A) and mutant (B) blastocysts at day 4.5-p.c., andpostimplantation (C) egg-cylinder stage embryos (transversal section) at day 6.5 p.c. The disruption of the ectodermal epithelium with detachedcells in the proamniotic cavity (arrow head), is the primary defect detected in β-cat −/− embryos at day 7.0 p.c. (E). A wild-type embryo at day7.0 p.c. is shown (D). At day 7.5 p.c. embryonic structures are no longer recognisable in mutant embryos (G). A wild-type embryo at day 7.5p.c. is depicted (F). When wild-type (H) and mutant (I) embryos at day 8.5 p.c. were compared, the latter appeared rather degenerated,including necrotic areas (arrow head); al, allantois; am, amniotic cavity; ec, ectoderm; en, endoderm; ep, ectoplacental cavity; epc,ectoplacental cone; ex, exocoelomic cavity; icm, inner cell mass; me, mesoderm; pa, proamniotic cavity; pen, parietal endoderm; re, Reichert’smembrane; tr, trophectoderm; ven, visceral endoderm; ven-l, visceral endoderm-like; xec, extraembryonic ectoderm; yc, yolk cavity; zp, zonapellucida. No morphological differences were found between β-catenin +/+ and +/− embryos. Scale bars: A,B, 27 µm; C-E, 65 µm; F,G, 130µm; H,I, 195 µm.
3533β-catenin and mouse development at gastrulation
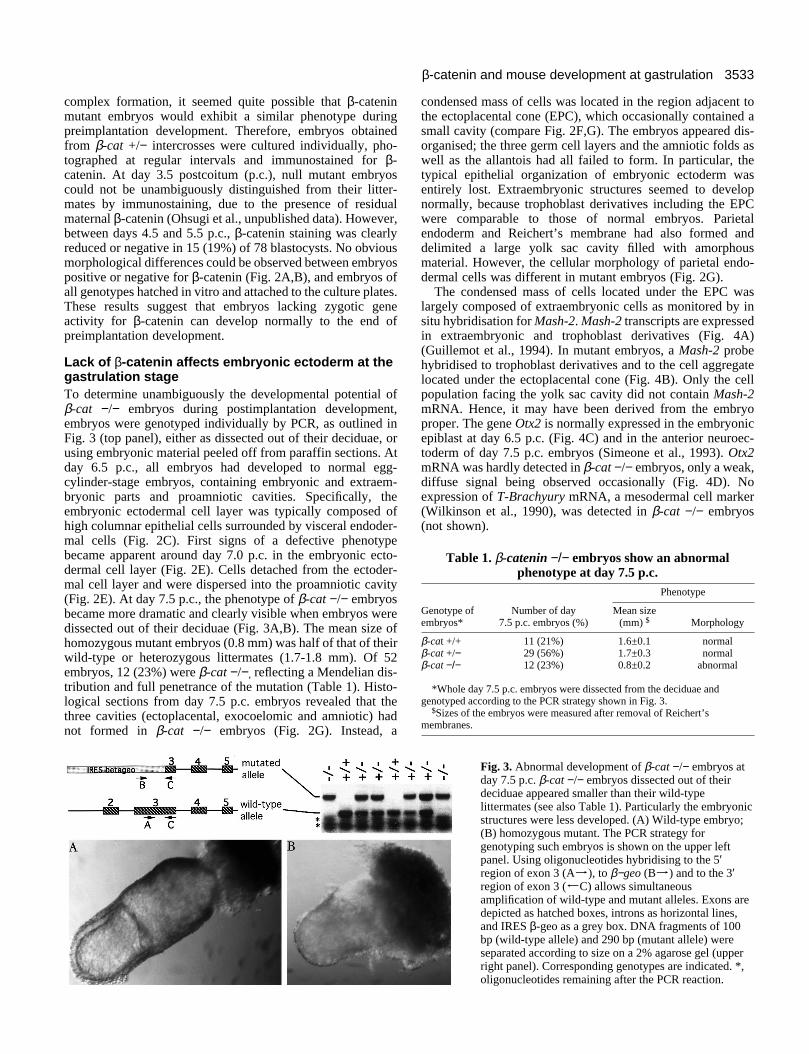
Table 1. β-catenin −/− embryos show an abnormalphenotype at day 7.5 p.c.
Phenotype
Genotype of Number of day Mean sizeembryos* 7.5 p.c. embryos (%) (mm) $ Morphology
β-cat +/+ 11 (21%) 1.6±0.1 normalβ-cat +/− 29 (56%) 1.7±0.3 normalβ-cat −/− 12 (23%) 0.8±0.2 abnormal
*Whole day 7.5 p.c. embryos were dissected from the deciduae andgenotyped according to the PCR strategy shown in Fig. 3.
$Sizes of the embryos were measured after removal of Reichert’smembranes.
complex formation, it seemed quite possible that β-cateninmutant embryos would exhibit a similar phenotype duringpreimplantation development. Therefore, embryos obtainedfrom β-cat +/− intercrosses were cultured individually, pho-tographed at regular intervals and immunostained for β-catenin. At day 3.5 postcoitum (p.c.), null mutant embryoscould not be unambiguously distinguished from their litter-mates by immunostaining, due to the presence of residualmaternal β-catenin (Ohsugi et al., unpublished data). However,between days 4.5 and 5.5 p.c., β-catenin staining was clearlyreduced or negative in 15 (19%) of 78 blastocysts. No obviousmorphological differences could be observed between embryospositive or negative for β-catenin (Fig. 2A,B), and embryos ofall genotypes hatched in vitro and attached to the culture plates.These results suggest that embryos lacking zygotic geneactivity for β-catenin can develop normally to the end ofpreimplantation development.
Lack of β-catenin affects embryonic ectoderm at thegastrulation stageTo determine unambiguously the developmental potential ofβ-cat −/− embryos during postimplantation development,embryos were genotyped individually by PCR, as outlined inFig. 3 (top panel), either as dissected out of their deciduae, orusing embryonic material peeled off from paraffin sections. Atday 6.5 p.c., all embryos had developed to normal egg-cylinder-stage embryos, containing embryonic and extraem-bryonic parts and proamniotic cavities. Specifically, theembryonic ectodermal cell layer was typically composed ofhigh columnar epithelial cells surrounded by visceral endoder-mal cells (Fig. 2C). First signs of a defective phenotypebecame apparent around day 7.0 p.c. in the embryonic ecto-dermal cell layer (Fig. 2E). Cells detached from the ectoder-mal cell layer and were dispersed into the proamniotic cavity(Fig. 2E). At day 7.5 p.c., the phenotype of β-cat −/− embryosbecame more dramatic and clearly visible when embryos weredissected out of their deciduae (Fig. 3A,B). The mean size ofhomozygous mutant embryos (0.8 mm) was half of that of theirwild-type or heterozygous littermates (1.7-1.8 mm). Of 52embryos, 12 (23%) were β-cat −/−, reflecting a Mendelian dis-tribution and full penetrance of the mutation (Table 1). Histo-logical sections from day 7.5 p.c. embryos revealed that thethree cavities (ectoplacental, exocoelomic and amniotic) hadnot formed in β-cat −/− embryos (Fig. 2G). Instead, a
condensed mass of cells was located in the region adjacent tothe ectoplacental cone (EPC), which occasionally contained asmall cavity (compare Fig. 2F,G). The embryos appeared dis-organised; the three germ cell layers and the amniotic folds aswell as the allantois had all failed to form. In particular, thetypical epithelial organization of embryonic ectoderm wasentirely lost. Extraembryonic structures seemed to developnormally, because trophoblast derivatives including the EPCwere comparable to those of normal embryos. Parietalendoderm and Reichert’s membrane had also formed anddelimited a large yolk sac cavity filled with amorphousmaterial. However, the cellular morphology of parietal endo-dermal cells was different in mutant embryos (Fig. 2G).
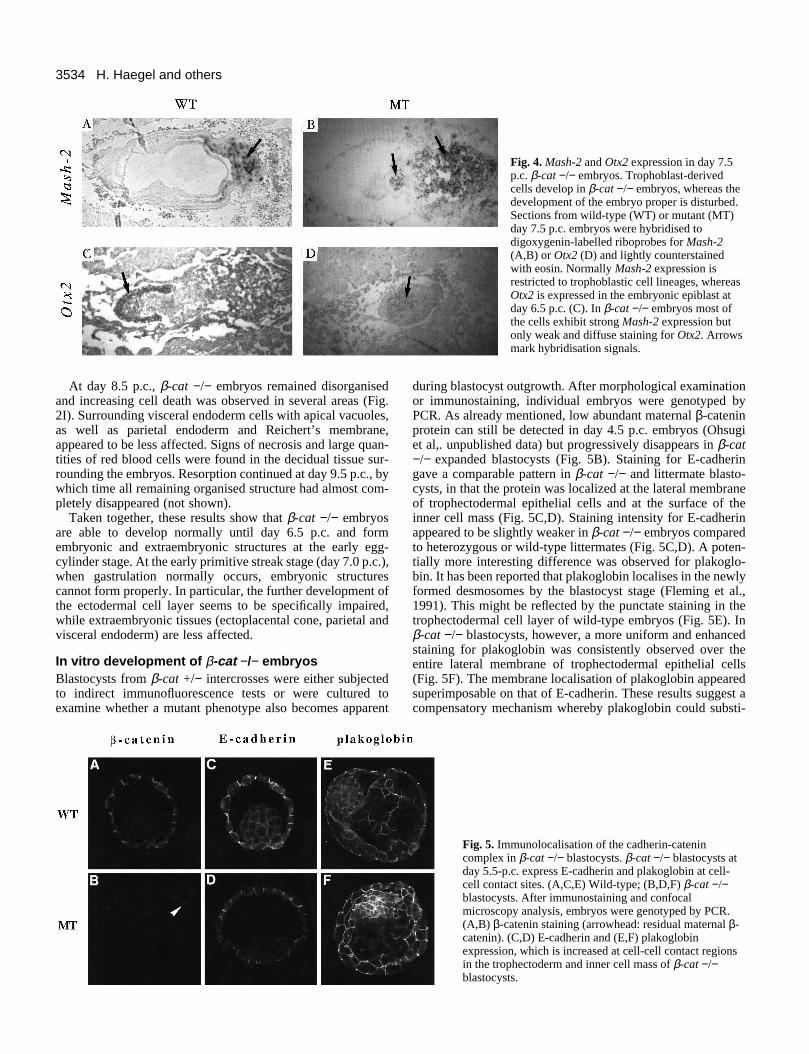
The condensed mass of cells located under the EPC waslargely composed of extraembryonic cells as monitored by insitu hybridisation for Mash-2. Mash-2 transcripts are expressedin extraembryonic and trophoblast derivatives (Fig. 4A)(Guillemot et al., 1994). In mutant embryos, a Mash-2 probehybridised to trophoblast derivatives and to the cell aggregatelocated under the ectoplacental cone (Fig. 4B). Only the cellpopulation facing the yolk sac cavity did not contain Mash-2mRNA. Hence, it may have been derived from the embryoproper. The gene Otx2 is normally expressed in the embryonicepiblast at day 6.5 p.c. (Fig. 4C) and in the anterior neuroec-toderm of day 7.5 p.c. embryos (Simeone et al., 1993). Otx2mRNA was hardly detected in β-cat −/− embryos, only a weak,diffuse signal being observed occasionally (Fig. 4D). Noexpression of T-Brachyury mRNA, a mesodermal cell marker(Wilkinson et al., 1990), was detected in β-cat −/− embryos(not shown).
Fig. 3. Abnormal development of β-cat −/− embryos atday 7.5 p.c. β-cat −/− embryos dissected out of theirdeciduae appeared smaller than their wild-typelittermates (see also Table 1). Particularly the embryonicstructures were less developed. (A) Wild-type embryo;(B) homozygous mutant. The PCR strategy forgenotyping such embryos is shown on the upper leftpanel. Using oligonucleotides hybridising to the 5′region of exon 3 (Aj), to β−geo (Bj) and to the 3′region of exon 3 (JC) allows simultaneousamplification of wild-type and mutant alleles. Exons aredepicted as hatched boxes, introns as horizontal lines,and IRES β-geo as a grey box. DNA fragments of 100bp (wild-type allele) and 290 bp (mutant allele) wereseparated according to size on a 2% agarose gel (upperright panel). Corresponding genotypes are indicated. *,oligonucleotides remaining after the PCR reaction.
3534 H. Haegel and others
Fig. 4. Mash-2 and Otx2 expression in day 7.5p.c. β-cat −/− embryos. Trophoblast-derivedcells develop in β-cat −/− embryos, whereas thedevelopment of the embryo proper is disturbed.Sections from wild-type (WT) or mutant (MT)day 7.5 p.c. embryos were hybridised todigoxygenin-labelled riboprobes for Mash-2(A,B) or Otx2 (D) and lightly counterstainedwith eosin. Normally Mash-2 expression isrestricted to trophoblastic cell lineages, whereasOtx2 is expressed in the embryonic epiblast atday 6.5 p.c. (C). In β-cat −/− embryos most ofthe cells exhibit strong Mash-2 expression butonly weak and diffuse staining for Otx2. Arrowsmark hybridisation signals.
At day 8.5 p.c., β-cat −/− embryos remained disorganisedand increasing cell death was observed in several areas (Fig.2I). Surrounding visceral endoderm cells with apical vacuoles,as well as parietal endoderm and Reichert’s membrane,appeared to be less affected. Signs of necrosis and large quan-tities of red blood cells were found in the decidual tissue sur-rounding the embryos. Resorption continued at day 9.5 p.c., bywhich time all remaining organised structure had almost com-pletely disappeared (not shown).
Taken together, these results show that β-cat −/− embryosare able to develop normally until day 6.5 p.c. and formembryonic and extraembryonic structures at the early egg-cylinder stage. At the early primitive streak stage (day 7.0 p.c.),when gastrulation normally occurs, embryonic structurescannot form properly. In particular, the further development ofthe ectodermal cell layer seems to be specifically impaired,while extraembryonic tissues (ectoplacental cone, parietal andvisceral endoderm) are less affected.
In vitro development of β-cat −/− embryosBlastocysts from β-cat +/− intercrosses were either subjectedto indirect immunofluorescence tests or were cultured toexamine whether a mutant phenotype also becomes apparent
during blastocyst outgrowth. After morphological examinationor immunostaining, individual embryos were genotyped byPCR. As already mentioned, low abundant maternal β-cateninprotein can still be detected in day 4.5 p.c. embryos (Ohsugiet al,. unpublished data) but progressively disappears in β-cat−/− expanded blastocysts (Fig. 5B). Staining for E-cadheringave a comparable pattern in β-cat −/− and littermate blasto-cysts, in that the protein was localized at the lateral membraneof trophectodermal epithelial cells and at the surface of theinner cell mass (Fig. 5C,D). Staining intensity for E-cadherinappeared to be slightly weaker in β-cat −/− embryos comparedto heterozygous or wild-type littermates (Fig. 5C,D). A poten-tially more interesting difference was observed for plakoglo-bin. It has been reported that plakoglobin localises in the newlyformed desmosomes by the blastocyst stage (Fleming et al.,1991). This might be reflected by the punctate staining in thetrophectodermal cell layer of wild-type embryos (Fig. 5E). Inβ-cat −/− blastocysts, however, a more uniform and enhancedstaining for plakoglobin was consistently observed over theentire lateral membrane of trophectodermal epithelial cells(Fig. 5F). The membrane localisation of plakoglobin appearedsuperimposable on that of E-cadherin. These results suggest acompensatory mechanism whereby plakoglobin could substi-
Fig. 5. Immunolocalisation of the cadherin-catenincomplex in β-cat −/− blastocysts. β-cat −/− blastocysts atday 5.5-p.c. express E-cadherin and plakoglobin at cell-cell contact sites. (A,C,E) Wild-type; (B,D,F) β-cat −/−blastocysts. After immunostaining and confocalmicroscopy analysis, embryos were genotyped by PCR.(A,B) β-catenin staining (arrowhead: residual maternal β-catenin). (C,D) E-cadherin and (E,F) plakoglobinexpression, which is increased at cell-cell contact regionsin the trophectoderm and inner cell mass of β-cat −/−blastocysts.

3535β-catenin and mouse development at gastrulation
Fig. 6. Phenotype of β-cat −/− embryos during in vitroculture. Morulae from heterozygous intercrosses werecultured for 6 days and subsequently genotyped byPCR using the primers shown in Fig. 3. ICM cells of β-cat −/− embryos (B,C) had a defect in cell-cell adhesioncompared to β-cat +/− embryos (A). The genotype ofeach of these embryos is shown in D. The two rightlanes of the gel (−/−) correspond to the PCR product ofthe β-cat −/− embryos shown in B and C. The lanelabelled (+/−) corresponds to the PCR products of theβ-cat +/- embryo shown in A. MT, DNA fragmentcorresponding to the mutant allele; WT, fragmentcorresponding to the wild-type allele; nc, negativecontrol; *, oligonucleotides remaining after the PCRreaction; icm, inner cell mass; tr, trophectoderm.
tute for β-catenin in the cadherin-catenin complex of β-cat−/− embryos and enable mutant embryos to form a functionaltrophectoderm.
In another series of experiments, blastocysts were culturedfor 4-6 days to examine the mutant phenotype during in vitroculture. Trophoblast outgrowth appeared equally welldeveloped in mutant and wild-type embryos (Fig. 6). However,in the developing ICMs of β-cat −/− embryos cells lost theircontacts with time and became dissociated; groups of looselyassociated cells detached from the trophoblast, occasionallycontinuing to proliferate in distant areas of the culture dish.This β-cat −/− phenotype during development in vitro corre-lates with that of postimplantation embryos.
DISCUSSION
We describe here the inactivation of the gene encoding for β-catenin by homologous recombination in ES cells and the gen-eration of transgenic mice. We show that lack of β-cateninresults in embryonic lethality; more specifically, our analysispoints to a primary defect in the embryonic ectoderm cell layerof day 6.5-7.5 p.c. embryos.
During early mouse embryonic development, two trueepithelial cell layers are formed: the trophectoderm at the blas-tocyst stage and the embryonic ectoderm at the egg-cylinderstage. Trophectoderm formation is not affected in β-cat −/−embryos, most likely due to the persistence of maternal β-catenin, which might to some extent compensate the lack ofzygotic gene activity. In addition, we observed an enhancedlocalisation of plakoglobin at the lateral membrane of tro-phectodermal epithelial cells, which suggests that plakoglobincan substitute for β-catenin to assure a functional E-cadherin-catenin complex at this developmental stage. It remains unclearwhether the increase of plakoglobin is due to increased tran-scriptional activity or prolonged protein stability.
Our in vivo and in vitro data on β-cat −/− embryos indicatethat the embryo develop into egg cylinders composed ofembryonic and extraembryonic ectoderm and of parietal andvisceral endoderm. Histological examination, as well as theexpression of cell-type-specific genes Mash-2 and Otx-2,
points to a primary defect in the maintenance of embryonicectoderm. Extraembryonic structures, including trophoblast,ectoplacental cone and visceral and parietal endoderm, appearnormal and/or are only affected in consequence of the primarydefect. At early gastrulation stage, the embryonic ectodermalcell layer is subjected to increased cell proliferation and mor-phogenetic movements, such as induction of mesoderm,primitive streak formation, and the establishment of theanterior-posterior axis. This all requires a continuous integra-tion of rapidly dividing cells into the epithelial cell layer. Onthe cellular level, a constantly occurring transition betweenpolarised and dividing, non-polarised epithelial cells has to beassured with a precise control of cell adhesion, cytoskeletalorganisation, and cell polarity.
A possible explanation of the phenotype in β-cat −/−embryos is that the lack of β-catenin affects the adhesive prop-erties of E-cadherin in embryonic ectodermal cells, leading toan improper integration of dividing cells into the ectodermalcell layer. This view is supported by the central role β-cateninplays in complex formation, binding both E-cadherin and α-catenin (Ozawa and Kemler, 1992; Aberle et al., 1994).Although plakoglobin has been reported to bind to E-cadherinand α-catenin (Butz and Kemler, 1994; Hinck et al., 1994a), itapparently can not substitute for the function of β-catenin inembryonic ectoderm. Thus, in spite of the high degree ofhomology between β-catenin and plakoglobin, each proteinmost likely exhibits unique properties, some yet to be discov-ered.
Given the homology of β-catenin and plakoglobin toarmadillo and the known involvement of armadillo in thewingless (Wnt) pathway, these proteins may well be part ofsignalling pathways. First support for such ideas came fromrecent findings that elevated expression levels of β-catenin orplakoglobin in response to wnt-1 expression can increasecadherin-mediated adhesion in cultured cells (Bradley et al.,1993; Hinck et al., 1994b). However, since the embryonicphenotype of wnt-1 deficiency, which leads to defects in braindevelopment (McMahon and Bradley, 1990), and the β-cat−/− phenotype reported here are distinct, we do not currentlyfavor a direct molecular interaction of both proteins. It ispossible that in embryonic ectoderm β-catenin is the target for

3536 H. Haegel and others
another member of the family of wnt signalling molecules.Additional support for a possible signalling function of β-catenin comes from studies of early Xenopus development.Overexpression of β-catenin in the ventral side of earlyXenopus embryos induced the formation of a secondary bodyaxis (Funayama et al., 1995); analysis of β-catenin deletionconstructs suggested that axis duplication might be induced byan intracellular signalling capacity of β-catenin. More infor-mative for the interpretation of our results is the work ofHeasman and colleagues on β-catenin depletion, which led toan inhibition of dorsal mesoderm formation during Xenopusdevelopment. Mesoderm formation was also inhibited in β-cat−/− mouse embryos, as monitored by the absence of T-Brachyury gene expression. Since in Xenopus, depletion of β-catenin had no apparent effect on the calcium-dependent reag-gregation of isolated blastomeres, the effect on dorsalmesoderm induction was explained by a role of β-catenin insignalling pathways (Heasman et al., 1994). It may well be thatβ-catenin exerts a signalling function in the mouse embryonicectoderm in addition to its role in cadherin-catenin complexformation. This could explain why lack of this widelyexpressed protein results in specific defects in the embryonicectodermal cell layer. The availability of β-catenin-negativeES cell lines established from β-cat −/− embryos will behelpful for further analysis of β-catenin function.
We acknowledge the excellent technical assistance of Julia Felthausand Kati Hansen and the help of Lore Lay in preparing the pho-tographs. We thank Drs Philippe Brûlet, François Guillemot,Bernhard Herrmann, Peter Mountford, Philippe Soriano and AustinSmith for gifts of plasmids, and Dr Davor Solter for helpful discus-sions. H. H. is supported by a Human Frontier Science Program long-term fellowship.
REFERENCES
Aberle, H., Butz, S., Stappert, J., Weissig, H., Kemler, R. and Hoschuetzky,H. (1994). Assembly of the cadherin-catenin complex in vitro withrecombinant proteins. J. Cell Sci. 107, 3655-3663.
Bradley, R.S., Cowin, P. and Brown, A.M. (1993). Expression of Wnt-1 inPC12 cells results in modulation of plakoglobin and E-cadherin andincreased cellular adhesion. J. Cell Biol. 123, 1857-1865.
Butz, S. and Kemler, R. (1994). Distinct cadherin-catenin complexes in Ca2+-dependent cell-cell adhesion. FEBS Lett. 355, 195-200.
Cowin, P., Kapprell, H.P., Franke, W.W., Tamkun, J. and Hynes, R.O.(1986). Plakoglobin: a protein common to different kinds of intercellularadhesive junctions. Cell 46, 1063-1073.
Fagotto, F. and Gumbiner, B.M. (1994). β-catenin localization duringXenopus embryogenesis: accumulation at tissue and somite boundaries.Development 120, 3667-3679.
Felthaus, J., Haegel, H. and Larue, L. (1994). Fast screening of mousemutants generated after homologous recombination. Nucleic Acids Res. 22,5504-5505.
Fleming, T.P., Garrod, D. R. and Elsmore, A. J. (1991). Desmosomebiogenesis in the mouse preimplantation embryo. Development. 112, 527-539.
Franke, W.W., Goldschmidt, M.D., Zimbelmann, R., Mueller, H.M.,Schiller, D.L. and Cowin, P. (1989). Molecular cloning and amino acidsequence of human plakoglobin, the common junctional plaque protein.Biochemistry 86, 4027-4031.
Friedrich, G. and Soriano, P. (1991). Promoter traps in embryonic stem cells:a genetic screen to identify and mutate developmental genes in mice. GenesDev. 5, 1513-1523.
Funayama, N., Fagotto, F., McCrea, P. and Gumbiner, B.M. (1995).Embryonic axis induction by the armadillo repeat domain of β-catenin:evidence for intracellular signaling. J. Cell Biol. 128, 959-968.
Guillemot, F., Nagy, A., Auerbach, A., Rossant, J. and Joyner, A.L. (1994).Essential role of Mash-2 in extraembryonic development. Nature 371, 333-336.
Hatzfeld, M., Krisjansson, G.I., Plessmann, U. and Weber, K. (1994). Band6 protein, a major constituent of desmosomes from stratified epithelia, is anovel member of the armadillo multigene family. J. Cell Sci. 107, 2259-2270.
Heasman, J., Crawford, A., Goldstone, K., Garner-Hamrick, P.,Gumbiner, B., McCrea, P., Kintner, C., Yoshida Noro, C. and Wylie, C.(1994). Overexpression of cadherins and underexpression of β-catenininhibit dorsal mesoderm induction in early Xenopus embryos. Cell 79, 791-803.
Hinck, L., Nelson, W.J. and Papkoff, J. (1994a). Wnt-1 modulates cell-celladhesion by stabilizing β-catenin binding to the cell adhesion moleculecadherin. J. Cell Biol. 124, 729-741.
Hinck, L., Näthke, I.S., Papkoff, J. and Nelson, W.J. (1994b). Dynamics ofcadherin-catenin complex formation: novel protein interactions andpathways of complex assembly. J. Cell Biol. 125, 1327-1340.
Hoffmann, I. and Balling, R. (1995). Cloning and expression analysis of anovel mesodermally expressed cadherin. Dev. Biol. (in press).
Hoschuetzky, H., Aberle, H. and Kemler, R. (1994). β-catenin mediates theinteraction of the cadherin-catenin complex with epidermal growth factor. J.Cell Biol. 127, 1375-1380.
Jostarndt, K. and Puntschart, A. (1994). The use of 33P-labelled riboprobesfor in situ hybridizations: localization of myosin alkali light-chain mRNAs inadult human skeletal muscle. Histochemical J. 26, 32-40.
Kawanishi, J., Kato, J., Sasaki, K., Fuji, S., Watanabe, N. and Niitsu, Y.(1995). Loss of E-cadherin-dependent cell-cell adhesion due to mutation ofthe β-catenin gene in a human cancer cell line, HSC-39. Mol. Cell. Biol. 15,1175-1181.
Kemler, R. (1992). Classical cadherins. Semin. Cell Biol. 3,149-155.Kemler, R. (1993). From cadherins to catenins: cytoplasmic protein
interactions and regulation of cell adhesion. T.I.G. 9, 317-321.Kinzler, K.W., Nilbert, M.C., Su, L., Vogelstein, B., Bryan, T.M., Levy,
D.B., Smith, K., Preisinger, A.C., Hedge, P., McKechnie, D., Finniear,R., Markham, A., Groeffen, J., Boguski, M.S., Alstshul, S.F., Horii, A.,Ando, H., Miyoshi, Y., Miki, Y., Nishisho, I. and Nakamura, Y. (1991).Identification of FAP Locus Genes from Chromosome 5q21. Science 253,661-665.
Knudsen, K. and Wheelock, M.J. (1992). Plakoglobin, or an 83-kDhomologue distinct from β-catenin, interacts with E-cadherin and N-cadherin. J. Cell Biol. 118, 671-679.
Larue, L., Ohsugi, M., Hirchenhain, J. and Kemler, R. (1994). E-Cadherin-null mutant embryos fail to form a trophectoderm epithelium. Proc. Natl.Acad. Sci. USA 91, 8263-8267.
McMahon, A.P. and Bradley, A. (1990). The Wnt-1 (int-1) protooncogene isrequired for development of a large region of the mouse brain. Cell 62, 1073-1085.
Mountford, P.S. and Smith, A.G. (1995). Internal ribosome entry sites anddicistronic RNAs in mammalian transgenesis. Trends Genet. 11,179-184.
Nagafuchi, A. and Takeichi, M. (1989). Transmembrane control of cadherin-mediated cell-cell adhesion: a 94 kDa protein functionally associated with aspecific region of the cytoplasmic domain of E-cadherin. Cell Regul. 1, 37-44.
Nagy, A., Rossant, J., Nagy, R., Abramow-Newerly, W. and Roder, J.C.(1993). Derivation of completely cell culture-derived mice from earlypassage embryonic stem cells. Proc. Nat. Acad. Sci. USA 90, 8424-8428.
Noordemeer, J., Klingensmith, J., Perrimon, N. and Nusse, R. (1994).dishevelled and armadillo act in the Wingless signaling pathway inDrosophila. Nature 367, 80-83.
Nose, A. and Takeichi, M. (1986). A novel cadherin cell adhesion molecule:its expression pattern associated with implantation and organogenesis ofmouse embryos. J. Cell Biol, 103, 2649-2658.
Oh, S.K. and Sarnow, P. (1993). Gene regulation: translational initiation byinternal ribosome binding. Curr. Op. Genet. Dev. 3, 295-300.
Oyama, T., Kanai, I., Ochiai, A., Akimoto, S., Oda, T., Yanagihara, K.,Nagafuchi, A., Tsukita, S., Shibamoto, S., Ito, F., Takeichi, M., Matsuda,H. and Hirohashi, S. (1994). A truncated β-catenin disrupts the interactionbetween E-cadherin and α-catenin: A cause of loss of intercellularadhesiveness in human cancer cell lines. Cancer Res. 54, 6282-6287.
Ozawa, M., Baribault, H. and Kemler, R. (1989). The cytoplasmic domain ofthe cell adhesion molecule uvomorulin associates with three independentproteins structurally related in different species. EMBO J. 8, 1711-1717.

3537β-catenin and mouse development at gastrulation
Ozawa, M. and Kemler, R. (1992). Molecular organization of the uvomorulin-catenin complex. J. Cell Biol. 116, 989-996.
Peifer, M., McCrea, P.D., Green, K.J., Wieschaus, E. and Gumbiner, B.M.(1992). The vertebrate adhesive junction proteins β-catenin and plakoglobinand the Drosophila segment polarity gene armadillo form a multigene familywith similar properties. J. Cell Biol. 118, 681-691.
Peifer, M., Berg, S. and Reynolds, A.B. (1994a). A repeating amino acid motifshared by proteins with diverse cellular roles. Cell 76, 789-791.
Peifer, M., Sweeton, D., Casey, M. and Wieschaus, E. (1994b). Winglesssignal and zest-white-3 kinase trigger opposite changes in the intracellulardistribution of Armadillo. Development 120, 369-380.
Peifer, M. (1995). Cell adhesion and signal construction: the Armadilloconnection. Trends Cell Biol. 5, 224-229.
Pelletier, J. and Sonenberg, N. (1988). Internal initiation of translation ofeukaryotic mRNA directed by a sequence derived from Poliovirus RNA.Nature 334, 320-325.
Reynolds, A.B., Daniel, J., McCrea, P., Wheelock, M.J., Wu, J. and Zhang,Z. (1994). Identification of a new catenin: the tyrosine kinase substratep120ras associates with E-cadherin complexes. Mol. Cell. Biol. 12, 8333-8342.
Riggleman, B., Schedl, P. and Wieschaus, E. (1990). Spatial expression of theDrosophila segment polarity gene armadillo is posttranscriptionallyregulated by wingless. Cell 63, 549-560.
Rubinfeld, B., Souza, B., Albert, I, Müller, O., Chamberlain, S.H.,Masiarz, F., Munemitsu, S. and Polaris, P. (1993). Association of the APCgene product with β-catenin. Science 262, 1731-1734.
Schneider, S., Herrenknecht, K., Butz, S., Kemler, R. and Hausen, P.(1993). Catenins in Xenopus embryogenesis and their relation to thecadherin-mediated cell-cell adhesion system. Development 118, 629-640.
Simeone, A., Acampora, D., Mallamaci, A., Stornaiuolo, A., D’Apice, M.R.,
Nigro, V. and Boncinelli, E. (1993). A vertebrate gene related toorthodenticle contains a homeodomain of the bicoid class and demarcatesanterior neuroectoderm in the gastrulating mouse embryo. EMBO J. 12,2735-2747.
Smith, A.G. and Hooper, M.L. (1987). Buffalo Rat Liver cells produce adiffusible activitity which inhibits the differentiation of murine embryonalcarcinoma and embryonic stem cells. Dev. Biol. 121, 1-9.
Spindle, A. (1980). An improved culture medium for mouse blastocysts. InVitro 16, 669-674.
Su, L. K., Vogelstein, B. and Kinzler, K. (1993). Association of the APCtumor suppressor protein with catenins. Science 262, 1734-1737.
Takeichi, M. (1991). Cadherin cell adhesion molecules as a morphogeneticregulator. Science 251, 1451-1455.
Vestweber, D. and Kemler, R. (1984). Rabbit antiserum against a purifiedsurface glycoprotein decompacts mouse preimplantation embryos and reactswith specific adult tissues. Exp. Cell Res. 152, 169-178.
Vestweber, D. and Kemler, R. (1985). Identification of a putative celladhesion domain of uvomorulin. EMBO J. 4, 3393-3398.
Wilkinson, D. G., Bhatt, S. and Herrmann, B. (1990). Expression pattern ofthe mouse T gene and its role in mesoderm formation. Nature 343, 657-659.
Yamamura, H. and Spindle, A. (1988). Stage-specific response ofpreimplantation mouse embryos to W-7, a calmodulin antagonist. J. Exp.Zool. 248, 45-54.
Yano, R., Oakes, M., Yamaghishi, M., Dodd, J.A. and Nomura, M. (1992).Cloning and characterization of SPR1, a suppressor of temperature-sensitiveRNA polymerase I mutations, in Saccharomyces cerevisiae. Mol. Cell. Biol.12, 5640-5651.
(Accepted 31 July 1995)


















