
Kynurenic acid and γ-D-glutamylaminomethylsulfonic acid suppress the compound action potential of...
5
Heuring Research, 25 (1987) 77-81 Elsevier HRR 00840 77 Kynurenic acid and y-D-glutamylaminomethylsulfonic acid suppress the compound action potential of the auditory nerve * Richard P. Bobbin and Gail Ceasar Kresge Hearing Research Laboratory of the South, Department of Otorhinolmygology and Biocommunication, Louisiana State University Medical Center, New Orleans. LA 70119-J 799, U.S.A. (Received 25 April 1986; accepted 8 July 1986) Kynurenic acid and y-D-glutamylaminomethylsulfonic acid, two excitatory amino acid antagonists, were perfused through the guinea pig cochlea while monitoring various cochlear potentials. Both drugs (0.6-10 mM) reduced the magnitude of the compound action potential (CAP) of the cochlear nerve without much effect on other potentials. The results are consistent with the hypothesis that the hair cell transmitter is an excitatory amino acid, possibly L-glutamate. kynurenic acid, y-D-glutamylaminomethylsulfonic acid, cochlea, hair cell. neurotransmitter Introduction Several workers have examined the hypothesis that the excitatory amino acid, L-glutamate, is a neurotransmitter released by hair cells of the mammalian cochlea (see reviews by Bobbin et al., 1984, 1985) and other hair cell systems (see discus- sion by Jenison et al., 1986). Three criteria of transmitter identification seem to have been met by L-glutamate: (1) it is excitatory to all hair cell afferents tested (as cited in Jenison et al., 1986); (2) it is released into the fluid that bathes the hair cells of the cochlea (Jenison et al., 1985) and the lateral line (Bledsoe et al., 1980); and (3) L- glutamine, a L-glutamate precursor, is taken up by the hair cells of the cochlea (Eybalin and Pujol, 1983; Ryan and Schwartz, 1984). However an additional criterion, antagonism of both exoge- nously administered L-glutamate and hair cell transmission with a specific glutamate blocker, has Correspondence address: Kresge Hearing Research Lab., LSUMC, 1100 Florida Ave., Bldg. 124, New Orleans, LA 70119-2799, U.S.A. * A preliminary report of these findings was presented February, 1986, at the 9th Midwinter Research Meeting of the Association for Research in Otolaryngology, Clearwater Beach, Florida. not been demonstrated in the cochlea, although it seems to have been shown in two other hair cell systems (Bledsoe and Bobbin, 1982; Annoni et al., 1984). At present there are thought to be three or more types of excitatory amino acid receptors that may be activated by an endogenous compound such as L-glutamate (Fagni et al., 1983; Bobbin et al., 1985). One classification names three types according to the exogenous substances thought to activate them: N-methyl-D-aspartate (NMDA), quisqualate, and kainate. NMDA and excitatory amino acid antagonists known to antagonize NMDA such as a-aminoadipate and 2-amino-5 phosphonovalerate are notably without effect in the cochlea (Fex and Martin, 1980; Bobbin et al., 1981, 1984; Jenison et al., 1986). In contrast, NMDA antagonists are effective in blocking both NMDA and afferent transmission in a few other hair cell systems, such as the lateral line of Xeno- pus (Bledsoe and Bobbin, 1982). The non-NMDA antagonists that have been tested in the cochlea have little effect on the compound action potential (CAP) of the auditory nerve or have additional actions on the cochlear microphonic potential (CM) (Bobbin et al., 1984, 1985). Therefore we have continued searching for
Transcript of Kynurenic acid and γ-D-glutamylaminomethylsulfonic acid suppress the compound action potential of...

Heuring Research, 25 (1987) 77-81
Elsevier
HRR 00840
77
Kynurenic acid and y-D-glutamylaminomethylsulfonic acid suppress the compound action potential of the auditory nerve *
Richard P. Bobbin and Gail Ceasar Kresge Hearing Research Laboratory of the South, Department of Otorhinolmygology and Biocommunication,
Louisiana State University Medical Center, New Orleans. LA 70119-J 799, U.S.A.
(Received 25 April 1986; accepted 8 July 1986)
Kynurenic acid and y-D-glutamylaminomethylsulfonic acid, two excitatory amino acid antagonists, were perfused through the
guinea pig cochlea while monitoring various cochlear potentials. Both drugs (0.6-10 mM) reduced the magnitude of the compound
action potential (CAP) of the cochlear nerve without much effect on other potentials. The results are consistent with the hypothesis
that the hair cell transmitter is an excitatory amino acid, possibly L-glutamate.
kynurenic acid, y-D-glutamylaminomethylsulfonic acid, cochlea, hair cell. neurotransmitter
Introduction
Several workers have examined the hypothesis that the excitatory amino acid, L-glutamate, is a neurotransmitter released by hair cells of the mammalian cochlea (see reviews by Bobbin et al., 1984, 1985) and other hair cell systems (see discus-
sion by Jenison et al., 1986). Three criteria of transmitter identification seem to have been met by L-glutamate: (1) it is excitatory to all hair cell
afferents tested (as cited in Jenison et al., 1986); (2) it is released into the fluid that bathes the hair cells of the cochlea (Jenison et al., 1985) and the lateral line (Bledsoe et al., 1980); and (3) L- glutamine, a L-glutamate precursor, is taken up by the hair cells of the cochlea (Eybalin and Pujol, 1983; Ryan and Schwartz, 1984). However an additional criterion, antagonism of both exoge- nously administered L-glutamate and hair cell transmission with a specific glutamate blocker, has
Correspondence address: Kresge Hearing Research Lab.,
LSUMC, 1100 Florida Ave., Bldg. 124, New Orleans, LA 70119-2799, U.S.A.
* A preliminary report of these findings was presented
February, 1986, at the 9th Midwinter Research Meeting of
the Association for Research in Otolaryngology, Clearwater Beach, Florida.
not been demonstrated in the cochlea, although it seems to have been shown in two other hair cell systems (Bledsoe and Bobbin, 1982; Annoni et al., 1984).
At present there are thought to be three or more types of excitatory amino acid receptors that may be activated by an endogenous compound such as L-glutamate (Fagni et al., 1983; Bobbin et al., 1985). One classification names three types according to the exogenous substances thought to activate them: N-methyl-D-aspartate (NMDA), quisqualate, and kainate. NMDA and excitatory amino acid antagonists known to antagonize
NMDA such as a-aminoadipate and 2-amino-5 phosphonovalerate are notably without effect in the cochlea (Fex and Martin, 1980; Bobbin et al., 1981, 1984; Jenison et al., 1986). In contrast, NMDA antagonists are effective in blocking both NMDA and afferent transmission in a few other
hair cell systems, such as the lateral line of Xeno- pus (Bledsoe and Bobbin, 1982). The non-NMDA antagonists that have been tested in the cochlea have little effect on the compound action potential (CAP) of the auditory nerve or have additional actions on the cochlear microphonic potential (CM) (Bobbin et al., 1984, 1985).
Therefore we have continued searching for
IX
compounds which might be effective in the cochlea by testing chemicals which have antagonistic
properties on the non-NMDA types of excitatory amino acid receptors. This study describes our
results with two new antagonists, kynurenic acid (kynurenate) and y-D-glutamylaminomethylsul- fonic acid (GAMS), which have significant block- ing effects at non-NMDA receptors in other neu- ronal systems (Ganong et al., 1983; Croucher et al., 1984; Davies and Watkins, 1985; Herrling, 1985; Jahr and Jessell, 1985; Stone and Connick, 1985).
Methods
Experiments were performed on 13 pigmented guinea pigs of both sexes. The methods used are
similar to those previously described (Bobbin and Konishi, 1974; Bledsoe et al., 1981). The animals
were anesthetized with chlorprothixene (Taractan@, 25 mg/kg, i.p.) and pentobarbital (Nembutal@, 30 mg/kg, i.p.), artificially respired, and maintained at a rectal temperature of 37-39°C. During ex-
periments, additional pentobarbital was adminis- tered (25 mg/kg, i.p.) to assure a deep level of anesthesia. The right cochlea of each animal was exposed ventrolaterally and both middle ear muscles were sectioned.
The CAP (Nl-Pl), CM and summating poten- tial (SP) were recorded from a 200 pm enamel-in- sulated silver wire placed in a hole in the Scala
vestibuli of the basal turn, amplified and stored on computer. The measured potentials were average responses (20 samples) obtained to 10 kHz tone bursts having 0.25 ms exponential rise-fall times, 10 ms durations and 100 ms interstimulus inter- vals. Intensity functions were obtained by varying
tone burst intensities (2-104 dB SPL, in 6 dB
steps). The endocochlear potential (EP) and the crossed olivocochlear potential (COCP), the change in the EP evoked by stimulating the cochlear efferents in the midline of the fourth ventricle, were recorded from animals which had received gallamine triethiodide (Flaxedila, Davis and Geck, 8 mg/kg, i.p.) as previously described (Bobbin and Konishi, 1974).
Artificial perilymph solutions were infused into the basal turn Scala tympani and allowed to flow out of the basal turn Scala vestibuli at approxi-
mately 2 pl/min for 10 min through holes made in the cochlea. Two artificial perilymph composi- tions were used. One, a phosphate/bicarbonate buffered solution, has been used previously (Konishi and Kelsey, 1973: Jenison et al., 1986)
and the other was a modification containing a Hepes buffer (Takada and Schacht, 1982). The modified solution had a composition of (mM): NaCl, 137; KCl, 5; CaCl,, 2; NaH,PO,, 0.1; MgCl,, 1; Hepes, 10; glucose, 11; and the pH was adjusted with 1 N NaOH to 7.4. The GAMS (Tocris) and kynurenate (Sigma) were initially dis- solved to make a 10 mM concentration and the pH adjusted to 7.4 with NaOH. Dilutions were then made to yield the concentrations tested (mM): 10, 5, 2.5, 1.25 and 0.625. In all animals, artificial perilymph without drug was the first perfusion.
This was then followed by successive perfusions with increasing concentrations of the drug from 0.625 to 10 mM, except in the animals where the
l Pre KYNA + GAMS
o Post GAMS IlOmM)
A Post KYNA t10mM)
40 80 g0 lb0
dB SPL
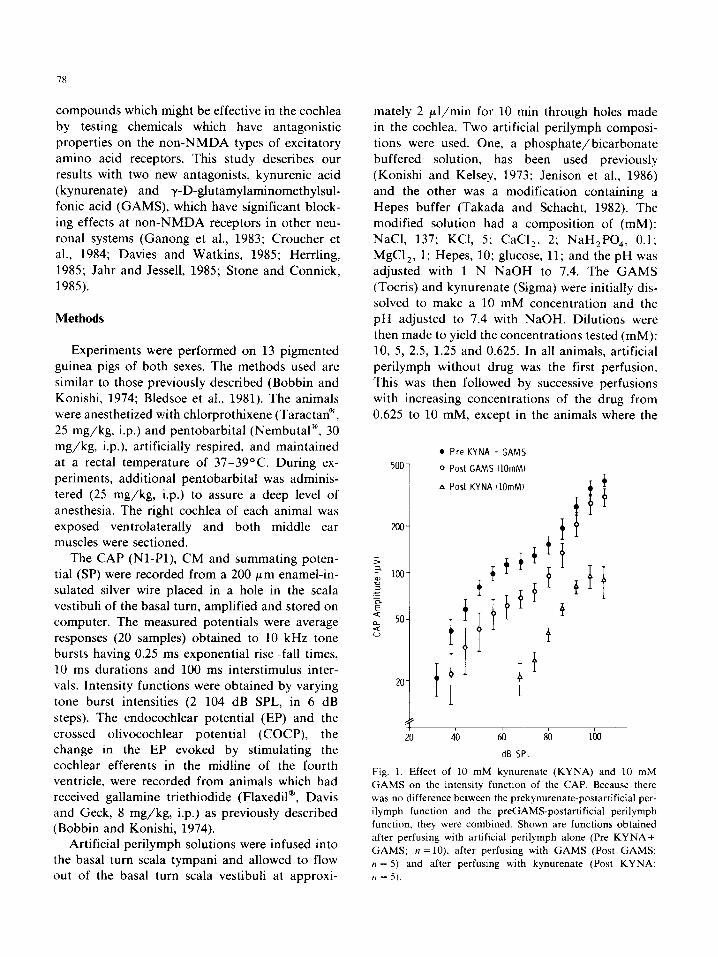
Fig. 1. Effect of 10 mM kynurenate (KYNA) and 10 mM
GAMS on the intensity function of the CAP. Because there
was no difference between the prekynurenate-postartificial per-
ilymph function and the preGAMSpostartificia1 perilymph
function. they were combined. Shown are functions obtained
after perfusing with artificial perilymph alone (Pre KYNA+
GAMS; n =10), after perfusing with GAMS (Post GAMS;
tt = 5) and after perfusing with kynurenate (Post KYNA:
?I = -5).
EP and COCP were monitored when only one concentration was tested. All drug perfusions were followed by another perfusion with artificial per- ilymph without drug. Intensity functions to the
tone bursts were recorded before any perfusions and immediately (within 2 min) after each perfu- sion. It is known empirically that the CAP inten-
sity’ function has a drug sensitive low intensity portion and a drug insensitive high intensity por- tion (Bobbin and Kisiel, 1981). Therefore, to quantify the effects of the drugs, we carried out statistical tests on CAP, CM, and SP data evoked by one intensity from each of these areas, 74 and 104 dB SPL, respectively. Analysis of variance and Newman-Keuls multiple-range tests were used to determine significance (P < 0.05). The data are expressed as mean k S.E.
19
Results
The first perfusion of either artificial perilymph increased the magnitude of all cochlear potentials
(e.g., Hepes, n = 3-5: CAP, 39%; CM, 4%; SP, 56%; EP, 9% and COCP, 17%). The effect on the
CAP, CM, EP, and COCP has been reported previously (Bobbin and Konishi, 1974; Bobbin and Thompson, 1978). In three animals, multiple perfusions of artificial perilymph without drug were carried out (phosphate/ bicarbonate solu- tion, n = 2; Hepes solution, n = 1). Perfusions subsequent to the first produced very little ad- ditional change in the cochlear potentials. There- fore, all drug effects are compared to potentials recorded after the first perfusion with artificial
perilymph.
200 CM74dB SPL
1 75 150 j 1 T
JJV
50 100
25 50
0 R 0.6 1.2 2.5 5 10 R P R 0.6 1.2 2.5 5 10 R 0
P R 0.6 1.2 2.5 5 10 R
CAP74dB SPL
CAP104dB SPL SP104dB SPL CM104dB SPL
P - R 0.6 1.2 2.5 6 10 R ” P R 0.6 1.2 2.6 6 10 R P R 0.6 1.2 2.6 6 10 R
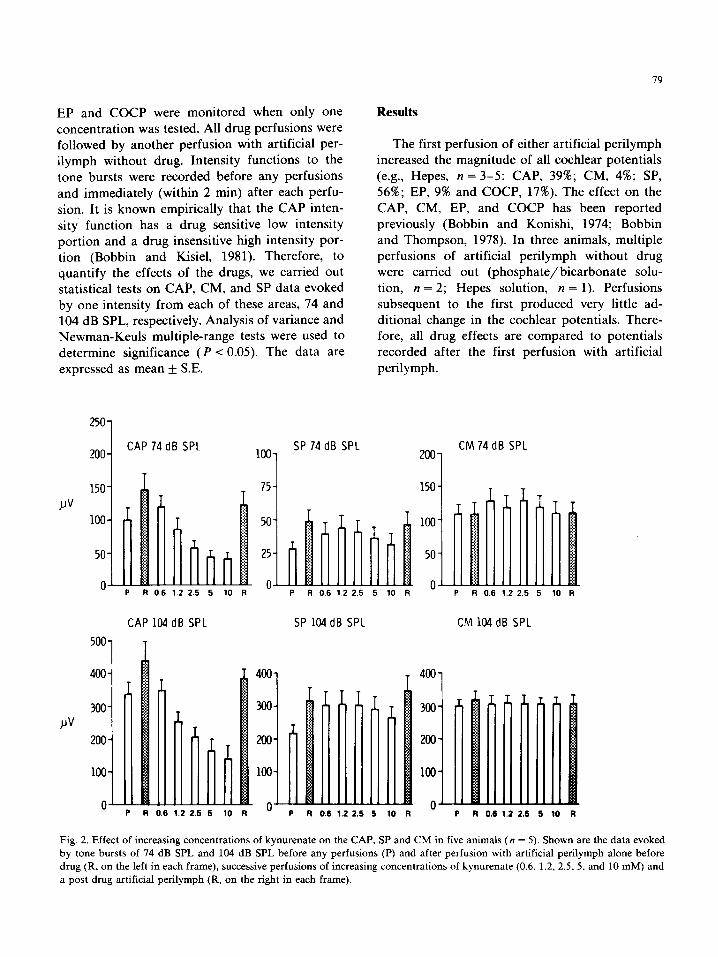
Fig. 2. Effect of increasing concentrations of kynurenate on the CAP. SP and CM in five animals (n = 5). Shown are the data evoked
by tone bursts of 74 dB SPL and 104 dB SPL before any perfusions (P) and after perfusion with artificial perilymph alone before
drug (R, on the left in each frame), successive perfusions of increasing concentrations of kynurenate (0.6, 1.2, 2.5, 5, and 10 mM) and a post drug artificial perilymph (R, on the right in each frame).

X0
Artificial perilymph solutions containing kyn-
urenate (n = 5) or GAMS (n = 5) suppressed the CAP, but did not affect the CM or SP. The suppression induced by both drugs was readily reversed upon perfusing with artificial perilymph without drug. The concentration which caused approximately 50% of the maximum suppressive response (measured at 74 dB SPL) for kynurenate
and GAMS was 1.25 and 2.5 mM, respectively. The greatest suppression occurred near the maxi- mum dosage utilized; therefore only the intensity functions generated before any drug and after 10 mM concentrations of each drug are shown in Fig. 1. The CAP suppression was equal to a lo-20 dB
reduction in sound intensity for GAMS and greater than 40 dB for kynurenate (Fig. 1). Statistically, GAMS did not produce a significant (P r 0.05) change in the CM or SP at 74 or 104 dB SPL and
its effect on CAP was only significant (P < 0.05) at 5 and 10 mM for 74 dB SPL. Since kynurenate gave the largest response on the CAP, its dose response relationship on CAP, CM and SP evoked by tone bursts on the CAP plateau (74 dB SPL) and at the maximal CAP value (104 dB SPL) are shown in Fig. 2. Statistical examination revealed that kynur’enate significantly (P < 0.05) reduced the CAP measured at both 74 and 104 dB SPL, in
concentrations of 1.25 mM and greater, but had no significant effect (P > 0.05) on the CM or SP measured at these intensities.
The effects of kynurenate on the EP and COCP were descriptively evaluated in three animals to examine further its selectivity for the CAP. We
chose to test a concentration of 5 mM since it was at the top of the CAP dose response curve (Fig. 2). After perfusing artificial perilymph, which was the first perfusion in each animal, neither the EP (pre: 82 _rt 7 mV> nor the COCP (pre: 1.5 & 0.3 mV)
were changed (post: EP, 85 Ifr 4 mV; COCP, 1.7 f 0 mV). After a total of five subsequent perfusions with kynurenate (5 mM) alternated with artificial perilymph, the EP (pre: 80 + 7 mV) was decreased slightly and reversibly (post: 70 & 4 mV), but the COCP (pre: 1.7 f 0.2 mv) was not changed (post: 1.6 i 0.1 mV). In each animal, the COCP was readily abolished by a final perfusion of strych- nine sulfate (10 PM), as reported by others (Bob- bin and Konishi, 1974).
Discussion
Our current hypothesis is that the guinea pig cochlea hair cells release L-glutamate as an excita- tory neurotransmitter (Bobbin et al., 1984, 1985). Furthermore, because NMDA and blockers of NMDA have no effect in the cochlea, we have proposed that if L-glutamate is the hair cell neuro- transmitter, then it acts on non-NMDA receptors (Bobbin et al., 1985; Jenison et al., 1986). There- fore we examined two relatively new antagonists which have prominent actions at non-NMDA sites, kynurenate and CAMS.
GAMS has been reported to antagonize kainate
> quisqualate > NMDA > L-glutamate (Croucher et al., 1984; Davies and Watkins, 1985). Kynure-
nate appears to be more of a general ‘broad spectrum antagonist’ that competitively blocks NMDA > kainate > L-glutamate 2 quisqualate (Ganong et al., 1983; Herrling, 1985; Jahr and Jesse& 1985; Stone and Connick, 1985; Ganong and Cotman, 1986). In the present study, GAMS appears selective for the CAP by its lack of effect on CM and SP. Likewise, kynurenate was found to be selective for the CAP, lacking effect on the CM, SP and COCP, and having only a slight
effect on the EP. Therefore the dose-dependent suppression of
the CAP that we observe with GAMS and kynurenate could possibly have resulted from a blockade of the hair cell neurotransmitter at the afferent nerve endings. The effect of kynurenate on EP does suggest that we exercise caution in interpreting the action of the drugs as being solely at this site. Nevertheless. these results are con- sistent with the hypothesis that an excitatory amino acid, such as L-glutamate, is the hair cell neuro- transmitter. Future studies will examine various agonists against each antagonist at the single neu- ron level to determine the site of action more specifically.
Acknowledgements
Thanks to Dr. L. Hughes, Dr. J.K. Cullen, Jr., C. Wiesendanger and A. Florez for designing and building the computer and writing the software which was used to collect the CAP, CM and SP data, and to John Carr, editorial consultant at

81
LSU School of Dentistry. Supported by NSF re- search grant BNS8419241 and the Louisiana Lions Eye Foundation.
References
Annoni, J.M., Co&ran, S.L. and Precht, W. (1984) Pharmacol-
ogy of the vestibular hair cell-afferent fiber synapse in the
frog. J. Neurosci. 4, 2106-2116.
Bledsoe, SC., Jr. and Bobbin, R.P. (1982) Effects of D-a-
aminoadipate on excitation of afferent fibers in the lateral
line of Xenopus laeuis. Neurosci. Lett. 32, 315-320.
Bledsoc, SC., Jr., Bobbin, R.P., Thalmann, R. and Thalmann,
I. (1980) Stimulus-induced release of endogenous amino
acids from the Xenopus laeuis lateral-line organ. Exp. Brain
Res. 40, 97-101.
Bledsoe, S.C., Jr., Bobbin, R.P. and Chihal, D.M. (1981) Kainic
acid: an evaluation of its action on cochlear potentials.
Hear. Res. 4, 109-120.
Bobbin, R.P. and Kisiel, D.L. (1981) Physiology of hearing. In:
Pharmacology of Hearing, pp. 19-50. Editors: R.D. Brown
and E.A. Daigneault. John Wiley and Sons, New York.
Bobbin, R.P. and Konishi, T. (1974) Action of cholinergic and
anticholinergic drugs at the crossed olivocochlear bundle-
hair cell junction. Acta Oto-Laryngol. 77, 56-65.
Bobbin, R.P. and Thompson, M.H. (1978) Effects of putative
transmitters on afferent cochlear transmission. Ann. Otol.
Rhinol. Laryngol. 87, 185-190.
Bobbin, R.P., Bledsoc, S.C., Jr. and Chihal, D.M. (1981) Ef-
fects of various excitatory amino acid antagonists on guinea
pig cochlear potentials. In: Abstr. 4th Midwinter Res.
Meet. Assoc. Res. Otolaryngol. 36, p. 27.
Bobbin, R.P., Bledsoe, S.C., Jr. and Jettison, G.L. (1984)
Neurotransmitters of the cochlea and lateral line organ. In:
Hearing Science: Recent Advances, pp. 159-180. Editor:
C.I. Berlin. College-Hill Press, San Diego.
Bobbin, R.P., Bledsoe, SC. Jr., Winbery, S.L. and Jenison,
G.L. (1985) Actions of putative neurotransmitters and other
relevant compounds on Xenopw laeuis lateral line. In:
Auditory Biochemistry, pp. 102-122. Editor: D.G.
Drescher. Thomas, Springfield, IL.
Croucher, M.J., Meldrum, B.S., Jones, A.W. and Watkins, J.C.
(1984) y-D-Glutamylaminomethylsulphonic acid (GAMS),
a kainate and quisqualate antagonist, prevents sound-in-
duced seizures in DBA/2 mice. Brain Res. 322, 111-114.
Davies, J. and Watkins, J.C. (1985) Depressant actions of
y-D-glutamylaminomethyl sulfonate (GAMS) on amino
acid-induced and synaptic excitation in the cat spinal cord.
Brain Res. 327,113-120.
Eybalin, M. and Pujol, R. (1983) A radioautographic study of
[ 3H]L-glutamate and [3H]L-glutamine uptake in the
guinea-pig cochlea. Neuroscience 9, 863-871.
Fagni, L., Baudry, M. and Lynch, G. (1983) Classification and
properties of acidic amino acid receptors in hippocampus.
J. Neurosci. 3, 1538-1546.
Fex, J. and Martin, M.R. (1980) Lack of effect of DL-a-
aminoadipate, an excitatory amino acid antagonist, on cat
auditory nerve responses to sound. Neuropharmacology 19,
809-811.
Ganong, A.H. and Cotman, C.W. (1986) Kynurenic acid and
quinolinic acid act at N-methyl-D-aspartate receptors in
the rat hippocampus. J. Pharmacol. Expl. Ther. 236,
293-299.
Ganong, A.H., Lanthom, T.H. and Cotman, C.W. (1983)
Kynurenic acid inhibits synaptic and acidic amino acid-
induced responses in the rat hippocampus and spinal cord.
Brain Res. 273,170-174.
Herrling, P.L. (1985) Pharmacology of the corticocaudate exci-
tatory postsynaptic potential in the cat: evidence for its
mediation by quisqualate or kainate-receptors. Neurosci.
14.417-426.
Jahr, C.E. and Jessell, T.M. (1985) Synaptic transmission be-
tween dorsal root ganglion and dorsal horn neurons in
culture: antagonism of monosynaptic excitatory postsyn-
aptic potentials and glutamate excitation by kynurenate. J.
Neurosci. 5, 2281-2289.
Jenison, G.L., Bobbin, R.P. and Thalmann, R. (1985) Potas-
sium-induced release of endogenous amino acids in the
guinea pig cochlea. J. Neurochem. 44, 1845-1853.
Jenison, G.L., Winbery, S. and Bobbin, R.P. (1986) Compara-
tive actions of quisqualate and N-methyl-D-aspartate, exci-
tatory amino acid agonists, on guinea-pig cochlear poten-
tials. Comp. B&hem. Physiol. 84C. 385-389.
Konishi, T. and Kelsey, E. (1973) Effect of potassium de-
ficiency on cochlear potentials and cation contents of the
endolymph. Acta Oto-Laryngol. 76, 410-418.
Ryan, A.F. and Schwartz, I.R. (1984) Preferential glutamine
uptake by cochlear hair cells: implications for the afferent
cochlear transmitter. Brain Res. 290, 376-379.
Stone, T.W. and Conmck, J.H. (1985) Quinolinic acid and
other kynurenines in the central nervous system. Neurosci-
ence 15, 597-617.
Takada, A. and Schacht, J. (1982) Calcium antagonism and
reversibility of gentamicin-induced loss of cochlear micro-
phonics in the guinea pig. Hear. Res. 8, 179-186.


















