
JBC Papers in Press. Published on May 9, 2013 as ... · β-Glucosidase with strong...
27
β-Glucosidase with strong transglycosylation activity 1 Characterization of a Novel β-Glucosidase from a Compost Microbial Metagenome with Strong Transglycosylation Activity* Taku Uchiyama, Kentaro Miyazaki, and Katsuro Yaoi Bioproduction Research Institute, National Institute of Advanced Industrial Science and Technology, Central 6, 1-1-1 Higashi, Tsukuba, Japan *Running title: β-Glucosidase with strong transglycosylation activity To whom correspondence should be addressed: Katsuro Yaoi, Bioproduction Research Institute, National Institute of Advanced Industrial Science and Technology, Central 6, 1-1-1 Higashi, Tsukuba 305-8566, Japan, Tel.: (85) 861-6152; Fax: (85) 861-6144; E-mail: [email protected] Keywords: β-glucosidase; product inhibition; transglycosylation http://www.jbc.org/cgi/doi/10.1074/jbc.M113.471342 The latest version is at JBC Papers in Press. Published on May 9, 2013 as Manuscript M113.471342 Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on September 6, 2018 http://www.jbc.org/ Downloaded from
Transcript of JBC Papers in Press. Published on May 9, 2013 as ... · β-Glucosidase with strong...

β-Glucosidase with strong transglycosylation activity
1
Characterization of a Novel β-Glucosidase from a Compost Microbial Metagenome with Strong Transglycosylation Activity* Taku Uchiyama, Kentaro Miyazaki, and Katsuro Yaoi Bioproduction Research Institute, National Institute of Advanced Industrial Science and Technology, Central 6, 1-1-1 Higashi, Tsukuba, Japan *Running title: β-Glucosidase with strong transglycosylation activity To whom correspondence should be addressed: Katsuro Yaoi, Bioproduction Research Institute, National Institute of Advanced Industrial Science and Technology, Central 6, 1-1-1 Higashi, Tsukuba 305-8566, Japan, Tel.: (85) 861-6152; Fax: (85) 861-6144; E-mail: [email protected] Keywords: β-glucosidase; product inhibition; transglycosylation
http://www.jbc.org/cgi/doi/10.1074/jbc.M113.471342The latest version is at JBC Papers in Press. Published on May 9, 2013 as Manuscript M113.471342
Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
2
Background: Search for β-glucosidase which have insensitivity for product inhibition has been demanded. Results: A β-glucosidase, Td2F2 was isolated from a metagenomic library have highly glucose tolerance and its activity is enhanced by various monosaccharides. Conclusion: Td2F2’s highly glucose tolerance is related to highly transglycosylation activity. Significance: This is the first report of the β-glucosidase have highly glucose tolerance and highly transglycosylation activity. SUMMARY The β -glucosidase encoded by the td2f2 gene was isolated from a compost microbial metagenomic library by functional screening. The protein was identified to be a member of the glycoside hydrolase family 1, and was overexpressed in Escherichia coli, purified, and biochemically characterized. The recombinant β -glucosidase, Td2F2, exhibited enzymatic activity with β -glycosidic substrates, with preferences for glucose, fucose, and galactose. Hydrolysis occurred at the nonreducing end and in an exo manner. The order of catalytic efficiency for glucodisaccharides and cellooligosaccharides was sophorose > cellotetraose > cellotriose > laminaribiose > cellobiose > cellopentaose > gentiobiose, respectively. Intriguingly, the p-nitrophenyl β -D-glucopyranoside hydrolysis activity of Td2F2 was activated by various monosaccharides and sugar alcohols. At a D-glucose concentration of 1000 mM, enzyme activity was 6.7-fold higher than that observed in the absence of D-glucose. With 31.3 mM D-glucose, Td2F2 catalyzed transglycosylation to generate sophorose, laminaribiose, cellobiose, and gentiobiose. Transglycosylation products were detected under all activated conditions, suggesting that the activity enhancement induced by monosaccharides and sugar alcohols may be due to the transglycosylation activity of the enzyme. These results show that Td2F2 obtained from a compost microbial metagenome may be a potent candidate for industrial applications.
β-Glucosidases (β-glucoside glucohydrolases, EC 3.2.1.21) are widely distributed in bacteria, fungi, plants, and animals and play vital roles in many biological processes such as biomass conversion by microbes and insects (1, 2),
biogenesis of various functional molecules (e.g., terpenols and flavonoids) from glycoside precursors (3, 4), and cyanide-based biological defense mechanisms (the release of cyanide from cyano glucosides) in plants (5). Recent interest in β-glucosidases arose from their involvement in the biological conversion of cellulose. β-Glucosidases are essential for efficient cellulolysis systems and are important for the degradation of cellooligosaccharides, particularly the breakdown of cellobiose to yield glucose. In addition, β-glucosidases also relieve product inhibition on cellobiohydrolases and endoglucanases, which are two major enzymes responsible for the degradation of cellulose. Both of these catalytic functions of β-glucosidases are critical to various biorefinery processes such as bioethanol production (1). However, most β-glucosidases are very sensitive to the presence of glucose, a primary product of β-glucosidase catalysis (6-8). The search for a β-glucosidase that is insensitive to product inhibition by glucose has newly been reinvigorated. Recently, due to their ability to activate glycosidic bonds, β-glucosidases have been considered as promising biocatalysts for the synthesis of stereo- and regiospecific glycosides or oligosaccharides, which are in turn are potentially useful as functional materials, nutraceuticals, or pharmaceuticals because of their biological recognition, signaling mechanisms, and antibiotic properties (1). In addition to hydrolytic activities, β-glucosidases may catalyze glycosidic bond formation via either a thermodynamically controlled reverse hydrolysis or a kinetically controlled transglycosylation (9). While glycosyltransferases are highly efficient in syntheses of glycosidic linkages, their application to industrial processes is limited by expensive nucleotide sugar precursors, narrow substrate specificity, and low enzyme availability. In contrast, β-glucosidases are attractive for large-scale applications since they are more abundant, commercially available, and exhibit a relatively broad acceptor-substrate specificity with simple substrates (10). However, their use in glycoside synthesis is limited by lower yields to industrial applications. The need for more suitable enzymes has fueled the search for novel β-glucosidases. Here, we report the cloning, heterologous
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
3
expression, and detailed enzymatic characterization of a glycoside hydrolase family 1 (GH1) protein, named Td2F2, from a compost microbial metagenome. According to our results, Td2F2 exhibits the highest potential for use not only in cellulosic ethanol production but also in the synthesis of stereo- and regiospecific glycosides, which is afforded by insensitivity to glucose inhibition and a relatively high transglycosylation activity. EXPERIMENTAL PROCEDURES Materials – Restriction endonucleases, DNA ligase, and DNA polymerase were purchased from Takara Bio (Shiga, Japan). A QIAquick Gel Extraction kit was used to purify DNA restriction fragments and polymerase chain reaction (PCR) products and was obtained from Qiagen (Hilden, Germany). All chromogenic substrates were purchased from Sigma-Aldrich (St. Louis, MO, USA). Cellooligosaccharides, laminarioligosaccharides, xylobiose, and chitobiose were acquired from Seikagaku Kogyo (Tokyo, Japan). Gentiobiose was purchased from Tokyo Chemical Industry (Tokyo, Japan). Isoprimeverose was obtained from Megazyme (Wicklow, Ireland). Curdlan was purchased from Wako Pure Chemical Industries (Osaka, Japan). Phosphoric acid-swollen cellulose was prepared from avicel (Wako Pure Chemical Industries) as described previously (11). Other mono- and disaccharides, including δ-gluconolactone and deoxynojirimycin, were purchased from Sigma-Aldrich. Construction of metagenomic library and screening – High-molecular-weight metagenomic DNA was prepared from compost according to the protocol of Uchiyama et al. (12). The compost we prepared that bark and humus were mixed and fermented for 3 years which fermentation was accelerated by Tricoderma strains inoculation. Metagenomic DNA was partially digested with Sau3AI and separated by gel electrophoresis. DNA fragments of 5 kb were recovered and ligated into a BamHI-digested p18GFP vector (12) and the product was transformed into Escherichia coli DH10B (Invitrogen, Carlsbad, CA, USA). The screening of the clones with β-glucosidase activity was performed on LB agar plates supplemented with 0.1 mM isopropyl-β-D-thiogalactoside, 750 µg mL–1
5-bromo-4-chloro-3-indolyl-β-D-glucopyranoside and 100 µg mL–1 ampicillin. Positive clones were detected by the formation of blue colonies following incubation at 37˚C for 14 h. The colonies were then incubated at 4˚C for 1 week. The resulting clones were reexamined for β-glucosidase activity using the same type of indicator plates. Cloning, expression, and purification of the β-glucosidase Td2F2 – A potential open reading frame of a β-glucosidase gene (termed td2f2) was identified in the inserted DNA fragments. The td2f2 gene was amplified by PCR using the primer pair 5’-TTTTTCATATGGCAGGAGAACGATTCCCG-3’ and 5’-AAAAAAAGCTTAGCCGCGCTGTCCGGCGATG-3’. This introduced NdeI and HindIII sites, respectively (underlined), at each end of the gene. The PCR product was then purified and digested with NdeI and HindIII. The digested fragment was ligated into pET-15b (Novagen, Madison, WI, USA), which was then subjected to the same digestion treatment to generate the plasmid pJTtd2f2 with six histidine residues attached to the N-terminal amino acid of Td2F2. E. coli BL21(DE3) harboring pJTtd2f2 was grown in 1 L of Overnight Express Instant LB Medium (Novagen), supplemented with 100 µg mL–1 ampicillin, at 37˚C. Growth was allowed 24 h. After, the cells were harvested by centrifugation at 4˚C and 5000 × g. The resulting cell pellet was resuspended in 20 mM sodium phosphate buffer (pH 7.4) with BugBuster (Novagen) and Benzonase (Novagen). The crude extract was incubated for 30 min at room temperature. Cell debris was removed by centrifugation at 4˚C and 15,000 × g for 20 min. The supernatant was filtered and run through a Ni-NTA affinity column (Novagen) to purify the recombinant enzyme, Td2F2. Multiple washes with washing buffer [20 mM sodium phosphate buffer (pH 7.4), 0.5 M NaCl, 25 mM imidazole] were performed and the enzyme was eluted with a linear gradient of 20–500 mM imidazole. Enzyme fractions were concentrated using an ultrafiltration membrane (Amicon Ultra; Millipore, Billerica, MA, USA). The concentrated enzyme solutions were then applied to a GE Healthcare (Buckinghamshire, UK) gel filtration column (HiLoad 16/600 Superdex 200 prep grade) and eluted with 50 mM sodium phosphate buffer (pH 6.0) containing 0.5 M NaCl at a flow rate of 1.0 mL min–1.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
4
The enzyme fractions were again concentrated using an ultrafiltration membrane (Amicon Ultra). The purity of the resulting Td2F2 was verified by SDS-PAGE and the enzyme was stored at 4˚C. Protein concentration was estimated from the solution absorbance at 280 nm using a molar extinction coefficient calculated from the amino acid composition (13). Molecular mass estimation of Td2F2 – The molecular mass of native-state Td2F2 was estimated by gel filtration through a GE Healthcare gel filtration column (HiLoad 16/600 Superdex 200 prep grade) in 50 mM sodium phosphate buffer (pH 6.0) containing 0.5 M NaCl at a flow rate of 0.8 mL min–1. The molecular weight standards were aldolase from rabbit muscle (158 kDa), conalbumin from chicken egg white (75 kDa), carbonic anhydrase from bovine erythrocytes (29 kDa), ribonuclease A from bovine pancreas (13.7 kDa), and aprotinin from bovine lung (6.5 kDa). Effects of pH and temperature on enzymatic activity – The optimal pH for Td2F2 activity with p-nitrophenyl β-D-glucopyranoside (pNPGlc) was evaluated at 75°C for 5 min in McIlvaine’s buffer (14) over a pH range of 3.0–8.5. pH stability was determined by measuring residual enzyme activity after incubation at 25°C for 30 min in appropriate buffers. The effect of temperature on enzyme activity for pNPGlc was measured from 20 to 85°C while incubating in buffer at pH 5.5 for 5 min. The thermostability of Td2F2 was determined by incubating the enzyme at various temperatures (20–85°C) in 20 mM Tris–HCl at pH 8.0 for 10 min. Residual activity was then measured in McIlvaine’s buffer at pH 5.5 and 30°C for 5 min. Assays of β-glycosidase activity – The activity of Td2F2 was assayed for 5 min at 75°C in 100 mM sodium acetate buffer (pH 5.5) using substrates containing the nonphysiological chromogenic aglycone p-nitrophenol (pNP) or natural oligosaccharides. The total reaction volume when using the chromogenic substrates was 20 µL and contained 0.05 µg of the enzyme. To stop the reaction, 50 µL of 1.0 M sodium bicarbonate was added to the reaction and the
concentration of pNP was determined by measuring the solution absorbance at 405 nm (Infinite M200 PRO; Tecan, Zurich, Switzerland). For oligosaccharides, the total reaction volume was 10 µL and contained 0.3 µg of the enzyme. The reaction was stopped by heating the sample to 98°C for 10 min. The amount of glucose, liberated as a product of the enzymatic reaction, was measured using a glucose oxidase-peroxidase assay (Amplex Red Glucose/Glucose Oxidase Assay Kit; Invitrogen) in accordance with the manufacturer’s protocol. In all assays, spontaneous hydrolysis of the substrate was accounted for by assay of blank mixtures, which lacked the enzyme. All measurements were performed in triplicate. Kinetic analysis of Td2F2–The kinetic parameters of Td2F2 were determined using pNPGlc, p-nitrophenyl β-D-fucopyranoside (pNPFuc), p-nitrophenyl β-D-galactopyranoside (pNPGal), cellooligosaccharides (di-, tri-, tetra-, and penta-), sophorose, laminarioligosaccharides (di-, tri-, tetra-, and penta-), gentiobiose, and lactose as substrates. The reaction conditions and the methods used to detect enzymatic activity are described above. The reaction velocity was determined at seven different substrate concentrations from 0.5 to 5.0 × Km where possible for each substrate. The kinetic constants Km and kcat were calculated by a nonlinear regression of the Michaelis–Menten equation using GraphPad PRISM Version 5.0 (GraphPad Software, La Jolla, CA, USA). Effects of inhibitors and additives – The inhibitory effects (Ki values) of δ-gluconolactone and deoxynojirimycin on the reaction of the enzyme with pNPGlc were determined at three substrate concentrations (2.5, 5, 10 mM) and five concentrations of inhibitor (5.0–40 mM δ-gluconolactone and 1.6–25 mM deoxynojirimycin). δ-Gluconolactone solutions were always applied within 5 min of preparation. Ki values were determined from Dixon plots. The effects of additives were evaluated by measuring enzyme activity in the presence of various metals, EDTA, ethanol, and DMSO and co-incubating the enzyme solution and 5 mM pNPGlc at 75˚C and pH 5.5 for 5 min following the measurement of pNP levels. Analysis of cellobiose degradation – Cellobiose degradation was analyzed using a high-performance anion exchange chromatograph with pulsed amperometric detection (HPAE-PAD; ICS-3000; Dionex,
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
5
Sunnyvale, CA, USA). The column was prepacked with CarboPac PA-1 (4 × 250 mm; Dionex). Solvents A (100 mM NaOH), B (100 mM NaOH, 1 M sodium acetate), and C (water) were used as eluents (flow rate 1 mL min–1). The eluent program was 10–80% A/(A + D) from 0 to 15 min in gradient mode, 80–98% A/(A + B) from 15 to 20 min in gradient mode, 2–10% B/(A + B) from 20 to 45 min in gradient mode, and 10% B/(A + B) from 45 to 50 min in linear mode. Cellobiose was identified and quantitated by comparing its retention time and peak area with those of an authentic standard. Kinetic analysis of Td2F2 activity with monosaccharides – The kinetic parameters of Td2F2 with monosaccharides were determined using 140 or 1000 mM D-glucose, D-fucose, D-galactose, L-arabinose, D-mannose, and D-xylose with pNPGlc as a substrate. The reaction conditions and the detection of enzymatic activity are described above. The reaction velocity was determined at six to nine pNPGlc concentrations (0.5–5.0 × Km). The calculation of kinetic constants is described above. Analysis of transglycosylation products – Transglycosylation products were analyzed by high-performance liquid chromatography (LC-2000 series; Jasco, Tokyo, Japan) using a corona-charged aerosol detector (Corona CAD; ESA Biosciences, Chelmsord, MA, USA) and a prepacked Shodex Asahipak NH2P-50 column (4.6 × 250 mm; Showa Denko K. K., Kanagawa, Japan). The eluent was water with a linear gradient of CH3CN from 80% to 74% over 30 min. The column was kept at 40˚C and eluted at 1.0 mL min–1. Glucodisaccharides were identified and quantitated by comparing retention times and peak areas with those of authentic standards. The reactants were pretreated for 16 h at 37˚C with glucose oxidase from Aspergillus niger (Wako Pure Chemical Industries) to degrade excess D-glucose prior to analysis by Corona CAD. DNA sequence accession number – The DNA sequence of td2f2 is available from the GenBank database with the accession number of HV538882. RESULTS Screening of clones with β-glucosidase activity from the metagenomic library. Many positive clones exhibiting β-glucosidase activity were screened from the metagenomic library. The plasmid DNA for each positive
clone was re-transformed into E. coli. The transformants exhibited β-glucosidase activity, indicating that the enzyme was encoded in the plasmid inserts. One of these positive clones, designated ptd2f2, was selected for further detailed characterization due to its relatively high β-glucosidase activity (pNPGlc degradation) activated by excess D-glucose. Sequence analysis of Td2F2 – The insert in ptd2f2 was sequenced and revealed one open reading frame (named as td2f2) of 1368 bp with a GC content of 67.2%. Td2F2 consisted of 445 amino acids with a predicted molecular mass of 49,343 Da. Comparisons with the BLAST database (http://www.ncbi.nlm.nih.gov/BLAST/) revealed that Td2F2 was similar to enzymes belonging to the glycoside hydrolase family 1 (GH1) of a class of carbohydrate-active enzymes (http://www.afmb.cnrs-mrs.fr/CAZY/index.html). Td2F2 exhibited the highest identity (61%) with β-glucosidase of Meiothermus ruber DSM 1279 (UniProt accession number D3PLV5), which was revealed from whole genome sequencing and had not been biochemically characterized. The highest identity (52%) with an enzyme with experimentally evidenced β-glucosidase activity was found with GlyB from Alicyclobacillus acidocaldarius (GenBank accession number DQ092439; 15). The alignment of the amino acid sequence of Td2F2 with highly homologous GH1 enzymes is shown in Fig. 1. The region containing the catalytic acid, Asn165-Glu166, is well conserved. The region containing the catalytic nucleophile Glu352 in Td2F2 differs only slightly from the consensus pattern of [LIVMFSTC]-[LIVFYS]-[LIV]-[LIVMST]-E3
52NG-[LIVMFAR]-[CSAGN] as described by the PROSITE motif PS00572, which is well conserved among the GH1 proteins. In Fig. 1, Td2F2 contains a Cys355 residue after the well-conserved catalytic nucleophile region E352NG. This feature is unique to Td2F2; other GH1 proteins have an Ala residue at this position. The amino acid residues related to subsites –1 and +1 are also shown in Fig 1. These residues were identified from structural analyses of Clostridium cellulovorans β-glucosidase, BglA (GenBank accession number AY268940; 16). While subsite –1 is highly conserved, Met413 and Tyr415 residues are shown in Td2F2. No significant set of subsite +1 residues is conserved among the GH1 proteins. This is consistent with the fact
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
6
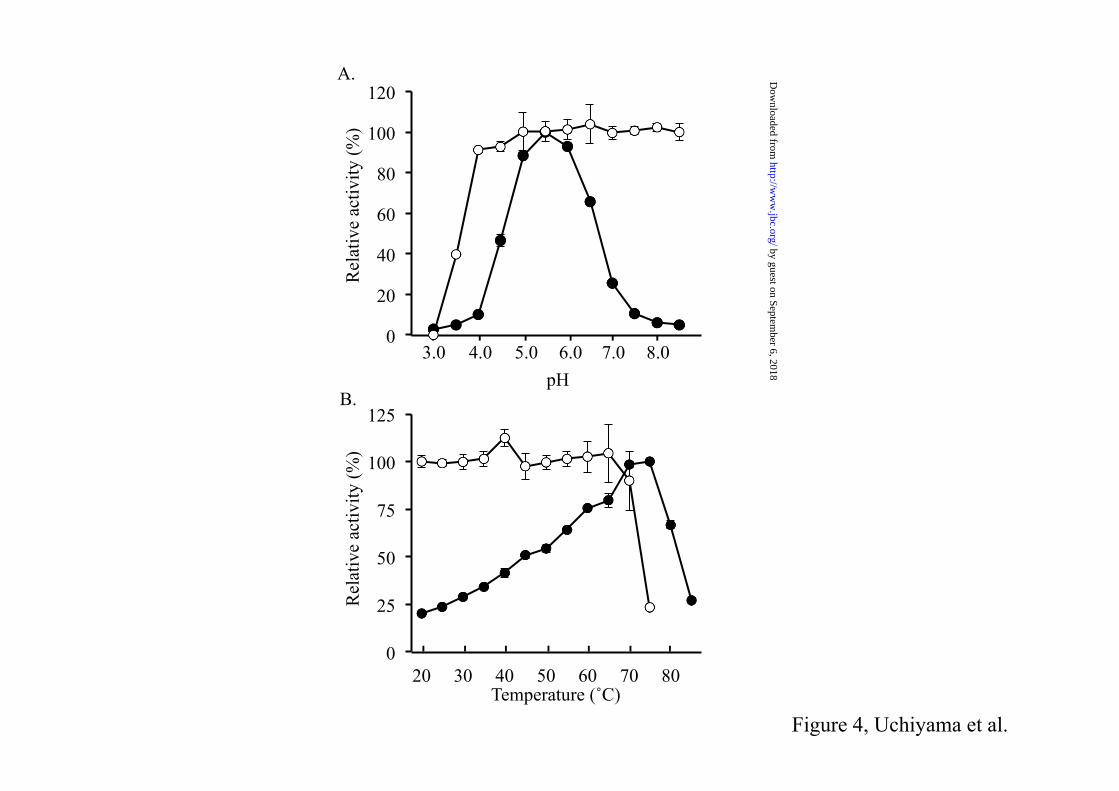
that GH1 proteins have a large range of potential substrates and specificities. Recombinant production of Td2F2 – Recombinant Td2F2 enzyme was efficiently expressed in a soluble protein fraction in E. coli BL21(DE3). N-terminal His-tagged Td2F2 enzyme was purified to homogeneity from the soluble protein fraction by affinity chromatography and gel filtration (Fig. 2). The last purification step yielded about 2.4 mg of pure protein per liter of E. coli culture. Gel filtration with suitable molecular weight standards revealed that the recombinant Td2F2 existed as a monomer of about 52 kDa under native conditions (Fig. 3). A molecular mass of 52 kDa agrees well with the predicted size of Td2F2 (49.3 kDa), plus the His-tag and polylinker regions (2.2 kDa). Properties of Td2F2 – Td2F2 exhibited an optimal activity at pH 5.5 (Fig. 4A) when assayed with 1 mM pNPGlc at 75˚C. Under these conditions, the enzyme had a specific activity of 13.8 ± 0.31 µmol min–1 mg–1. Td2F2 was stable from pH 4.0 to 8.5. Within this pH range, the enzyme maintained more than 85% of its original activity after incubation. Td2F2 showed substantial activity at temperatures from 20˚C to 85˚C and exhibited the highest activity at the optimal temperature of 75˚C (specific activity; 15.2 ± 0.59 µmol min–1 mg–1; Fig. 4B). Thermostability experiments indicated that the enzyme was stable at temperatures lower than 65˚C. The hydrolytic activity of Td2F2 toward various substrates was measured to determine substrate specificity. Among the chromogenic substrates, not only pNPGlc, but also pNPFuc, pNPGal, p-nitrophenyl-β-D-cellobiose, and p-nitrophenyl-β-D-lactopyranoside were good substrates of Td2F2 (Table 1). The enzyme showed very little activity with p-nitrophenyl-β-D-mannopyranoside, p-nitrophenyl-β-D-xylopyranoside, and p-nitrophenyl-α-L-arabinopyranoside, even after prolonged incubation for 1 h. No activity was observed with p-nitrophenyl-N-acetyl-β-D-glucosaminide, p-nitrophenyl-2-acetamide-2-deoxy-β-glucopyranoside, p-nitrophenyl-β-L-arabinopyranoside, p-nitrophenyl-α-D-glucopyranoside, p-nitrophenyl-α-D-galactopyranoside, p-nitrophenyl-α-D-xylopyranoside, p-nitrophenyl-α-L-arabinofuranoside, p-nitrophenyl-α-L-fucopyranoside, and
p-nitrophenyl-α-L-rhamnopyranoside, even after prolonged incubations. The enzyme acted on β-glycosidic linkages, such as those in cellooligosaccharides (cellobiose to cellohexaose), sophorose, laminarioligosaccharides (laminaribiose to laminaripentaose), gentiobiose, and lactose. Td2F2 exhibited a very low activity with laminarihexaose after a prolonged incubation of 1 h. No enzymatic activity was observed with trehalose, kojibiose, nigerose, maltose, and isomaltose, which have α(1,1), α(1,2), α(1,3), α(1,4), and α(1,6) glycosidic linkages, respectively. Insoluble β-glucans, such as phosphoric acid-swollen cellulose [β(1,4)-glucan] and curdlan [β(1,3)-glucan] were likewise not substrates of Td2F2. Analysis of degradation products by HPAE-PAD revealed that sucrose, isoprimeverose, chitobiose, and xylobiose were also not substrates of Td2F2. Only cellotriose and glucose were produced during a time-course HPAE-PAD analysis of cellotetraose hydrolysis. This confirms an exo type of activity for Td2F2 (data not shown). A comparison of enzymatic activity with various substrates confirmed that Td2F2 is specific for β-glycosidic substrates, with preferences for the nonreducing ends of glucose, fucose, and galactose. Kinetic constants of Td2F2 – The steady-state kinetic parameters of Td2F2 were measured for those β-glycosides shown to be good substrates. Table 2 summarizes these data for three aryl β-glycosides, a variety of cellooligosaccharides and laminarioligosaccharides, sophorose, gentiobiose, and lactose. The enzyme showed a greater hydrolytic efficiency for pNPFuc and pNPGlc. Among the glucodisaccharides, sophorose [β(1,2) linked glucose] was the most easily hydrolyzed. Enzymatic activity decreased with the other substrates in the following order: laminaribiose [β(1,3)], cellobiose [β(1,4)], and gentiobiose [β(1,6)]. A different efficacy was observed with cellooligosaccharides with -tetraose being the most easily degraded. Activity then decreased in the following order: -triose, -biose, and -pentaose. The observed hydrolyzing efficacy was again slightly different for laminarioligosaccharides. The -triose was the most easily degraded with activity decreasing in the following order: -biose, -pentaose, and -tetraose.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
7
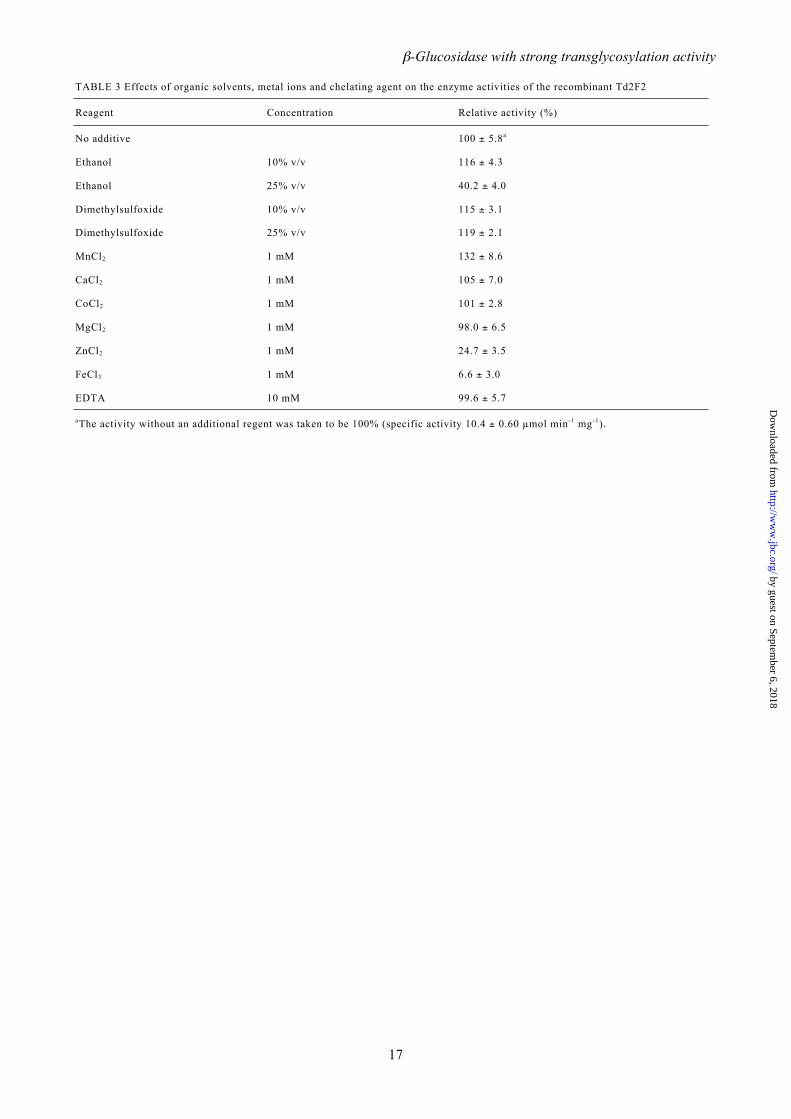
Effects of inhibitors, metals, and other reagents on Td2F2 activity – The kinetics of Td2F2 inhibition were determined using pNPGlc as a substrate and δ-gluconolactone and deoxynojirimycin as inhibitors. Dixon plots indicated competitive inhibition for both inhibitors with Ki values of 4.56 ± 0.64 and 32.5 ± 1.09 mM, respectively. The effects of various metals and regents on Td2F2 activity were also investigated (Table 3). Significant inactivation was observed with ZnCl2 and FeCl3. In contrast, enzyme activity was stimulated by MnCl2. The addition of 10% (v/v) ethanol resulted in a slight enhancement in activity, while 25% (v/v) ethanol dramatically reduced enzyme activity. The addition of 10% or 25% (v/v) DMSO resulted in a slight activity enhancement. EDTA did not affect enzyme activity. D-Glucose activation of Td2F2 – The activity of Td2F2 was enhanced with increasing concentrations of D-glucose (Fig. 5). With 1000 mM D-glucose and 10 mM pNPGlc as substrate, the hydrolysis rate of pNPGlc was 6.7 times that of the control, which did not contain D-glucose (Fig. 5A). This same phenomenon was also observed with 10 mM cellobiose as the substrate. In this case, 1000 mM D-glucose increased the hydrolysis activity by a factor of 2.4 over the control (Fig. 5B). Transglycosylation products, namely gentiobiose, laminaribiose, and sophorose (data not shown), were detected by HPAE-PAD as the products of Td2F2 enzymatic activity with 1.25, 5, or 10 mM cellobiose and 125 mM to 1000 mM D-glucose. In contrast, when Td2F2 was incubated with only cellobiose, the major product was D-glucose, and transglycosylation products were not detected. Kinetic analysis of Td2F2 activity with monosaccharides–The effects of 140 and 1000 mM D-glucose, D-fucose, D-galactose, L-arabinose, D-mannose, and D-xylose on Td2F2 activity were evaluated using pNPGlc as a substrate (Table 4). The addition of monosaccharides increased both Km and kcat. The most significant change in Km was observed with 1000 mM D-fucose, which raised Km by a factor of 14.8. The most significantly change in kcat was observed with the addition of 1000 mM D-glucose, which increased kcat by a factor of 10.8. The ratio of kcat to Km was significantly different for each monosaccharide and
concentration (Table 4). The addition of 1000 mM L-arabinose, D-mannose, and D-xylose increased kcat/Km by factors of 1.8-, 1.4-, and 1.3-fold, respectively, relative to kcat/Km values observed with 140 mM of monosaccharide. However, no significant changes were observed between the kcat/Km values for 140 mM and 1000 mM D-glucose and D-galactose. Notably, the addition of 1000 mM D-fucose decreased kcat/Km by a factor of 0.6 relative to that measured with 140 mM D-fucose, suggesting that D-fucose inhibits the enzymatic degradation of pNPGlc by Td2F2 under 1000 mM condition. Activation of Td2F2 by various monosaccharides and their derivatives – To investigate the effects of various monosaccharides, sugar alcohols, and acid sugars on Td2F2, enzyme activity was assayed in the presence of 140 mM monosaccharide, or a derivative thereof, with 5 mM pNPGlc, pNPFuc, and pNPGal as substrate (Fig. 6). Various monosaccharides and sugar alcohols significantly activated the degradation of pNPFuc and pNPGlc by Td2F2. With pNPFuc as a substrate, Td2F2 activity was strongly activated by the addition of D-glucose, L-glucose, methyl-α-D-glucose, methyl-β-D-glucose, D-fucose, L-fucose, D-arabinose, L-arabinose, D-mannose, D-fructose, D-galactose, L-sorbose, L-rhamnose, D-xylose, sedoheptulose, xylitol, glucitol, and mannitol (340%, 421%, 236%, 224%, 234%, 582%, 423%, 217%, 253%, 229%, 207%, 442%, 310%, 215%, 399%, 301%, 223%, and 340%, respectively). With pNPGlc as a substrate, additions of D-glucose, L-glucose, methyl-α-D-glucose, methyl-β-D-glucose, D-fucose, L-fucose, D-arabinose, L-arabinose, D-mannose, D-galactose, and L-sorbose resulted in strong activations of up to 281%, 420%, 209%, 223%, 216%, 263%, 199%, 209%, 194%, 241%, and 205%, respectively. With pNPGal as the substrate, the addition of L-fucose and sedoheptulose strongly activated Td2F2 by up to 191% and 210%, respectively, whereas L-glucose, D-glucosamine, D-glucuronate, D-gluconate, and D-galacturonate respectively reduced pNPGal degradation to 53%, 41%, 66%, 62%, and 72%. The addition of D-glucosamine also reduced the enzymatic degradation of pNPGlc and pNPFuc to 49% and 71%, respectively. This amino saccharide is a common inhibitor of the enzymatic activity of Td2F2.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
8
Additions of methyl-α-D-glucose and methyl-β-D-glucose showed similar enhancements, which indicates that the differences between α- and β-glycosidic linkages did not affect the enzymatic activity Td2F2. However, D- and L-isomeric differences did affect Td2F2 activity, especially for pNPFuc degradation. Additions of L-glucose, L-fucose, and D-arabinose resulted in enhanced pNPFuc degradation relative to the activities observed with D-glucose, D-fucose, and L-arabinose, respectively. This suggests that the configuration of OH-2 (in the equatorial position) and OH-3 (in the axial position) is important for pNPFuc degradation by Td2F2. The products of reaction mixtures of Td2F2, pNPGlc, and various monosaccharides and their derivatives were examined by HPAE-PAD and TLC. The products included not only glucose, but also the transglycosylation products of D-glucose, L-glucose, methyl-α-D-glucose, methyl-β-D-glucose, D-fucose, L-fucose, D-arabinose, L-arabinose, D-mannose, D-galactose, L-sorbose, L-rhamnose, D-xylose, xylitol, glucitol, mannitol, and arabitol (data not shown). Transglycosylation activity of Td2F2 – Transglycosylation activity was demonstrated using 10 mM pNPGlc as a substrate and 0–1000 mM D-glucose. Corona CAD analyses revealed that the reaction of Td2F2 with pNPGlc plus 31.3 mM D-glucose resulted in the transglycosylation products laminaribiose, cellobiose, sophorose, and gentiobiose (Fig. 7). When Td2F2 was incubated with only pNPGlc, no transglycosylation products were detected. When the D-glucose concentration in was elevated, the dominant transglycosylation product was sophorose, followed by laminaribiose. Figure 8 shows the production of laminaribiose and sophorose by Td2F2 with 0–1000 mM D-glucose analyzed during the degradation 10 mM pNPGlc. Cellobiose and gentiobiose were difficult to detect by Corona CAD at higher D-glucose concentrations. The proportions of pNP, sophorose, and laminaribiose increased with the concentration of D-glucose (Fig. 8). When 1000 mM D-glucose was used as a substrate without the addition of pNPGlc, no transglycosylation or hydrolysis products were observed, indicating that transglycosylation is coordinated with pNPGlc hydrolysis.
DISCUSSION We report here the molecular cloning, expression in E. coli, and functional characterization of the product of the td2f2 gene from the compost microbial metagenome. Amino acid sequencing showed that Td2F2 belongs to GH1, with all of the conserved regions of GH1 represented. Substrate specificity analyses suggested that Td2F2 hydrolyzes aryl-β-glycosides, β-linked disaccharides, and β-linked short chain oligosaccharides, but not aryl-α-glycosides, α-linked disaccharides, and long chain β-glucans. Td2F2 exhibited a broad specificity for β-glucosidases, which are the most commonly observed (1). Kinetic analyses of Td2F2 revealed higher kcat/Km ratios for pNPFuc and pNPGlc than for natural oligosaccharides. Several GH1 enzymes exhibit high specificity toward aryl-β-glucosides, such as chromogenic β-glucosides or arbutin and salicin (15, 17, 18), in which the aglycone group is aromatic. Aromatic β-glucosides are found in plants and the presence of specific enzymatic systems against such substrates suggests a specific interaction of the corresponding microorganism with living plants. Since Td2F2 was cloned from a compost microbial metagenome, its enzyme action likely degrades complex polysaccharides from plant residues. The kcat/Km for hydrolysis of cellotetraose is 3.2- and 1.5-fold greater than that of cellobiose and cellotriose, respectively. This increase is due to the low Km of cellotetraose. This phenomenon is consistent with the kinetic results of GH1 β-glucosidase from Sulfolobus solfataricus (19). The crystal structure of β-glucosidase from S. solfataricus suggests that cellotetraose has a stronger interaction with a tandem array of subsites in the enzyme compared to cellobiose and cellotriose. This hypothesis supports the low Km of cellotetraose (20). The kcat/Km for the hydrolysis of laminaripentaose is sixfold greater than that for laminaritetraose. This increase is due to the high kcat of laminaripentaose, which may be due to steric hindrance at the active site and inhibition of laminaritetraose hydrolysis. Kinetic analyses of Td2F2 showed that the catalytic efficiency for β-linked glucodisaccharides decreased in the order of sophorose > laminaribiose > cellobiose > gentiobiose, which indicates that
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
9
a glycosyl group could be accommodated in several ways at the +1 subsite of Td2F2. Two well-characterized β-glucosidase inhibitors, δ-gluconolactone and deoxynojirimycin, inhibited Td2F2, which suggests that Td2F2 is a β-glucosidase rather than an exo-β-glucanase. The calculated Ki values of 4.56 and 32.5 mM, respectively, suggest that Td2F2 would have a high resistance to these inhibitors (21). Various chemical reagents also affected the activity of Td2F2. Ten percent (v/v) ethanol and DMSO stimulated Td2F2 activity. Alcohol activation of some β-glucosidases due to glycosyltransferase activities has been observed (7, 22). Likewise, activation by organic solvents such as DMSO has been reported (22). The chelating agent EDTA did not inhibit Td2F2, indicating that divalent cations are not required for enzyme activation. However, the activity of Td2F2 was enhanced by the addition of Mn2+. In previous studies, strong activation by Mn2+ was observed with other GH1 β-glucosidases (16), which suggests that Mn2+ ions might assist catalytic function. Td2F2 activity was strongly inhibited by Zn2+ and Fe3+. These metal ions may hinder substrate binding or inhibit the catalytic reaction at the active site. Activity inhibition by glucose is a common characteristic of β-glucosidases (6-8). Most of the β-glucosidases that have been studied are competitively inhibited by their end-product, glucose. However, several β-glucosidases are tolerant to, or may actually be stimulated by, glucose (17, 23-25). Using pNPGlc as a substrate, the enzymatic activity of this type of β-glucosidase was enhanced by factors of 1.2–2.6 in the presence of 50–200 mM glucose. This activity was followed by a gradual inhibition at higher glucose concentrations (17, 23-25). Td2F2 is not only particularly resistant to glucose inhibition, but its enzymatic activity is significantly activated by glucose with no inhibition at higher concentrations. This particular characteristic of Td2F2 is a result of its high transglycosylation activity (Fig. 8). Td2F2 can preferentially utilize glucose rather than water as an acceptor for the glycosyl moiety during the catalytic degradation of pNPGlc and cellobiose, thereby resulting in elevated reaction rates. Td2F2 produced a series of β-linked glucodisaccharides from pNPGlc with D-glucose via transglycosylation. Under these conditions, sophorose was the primary
transglycosylation product. Kinetic studies on the degradation of various substrates agreed with this result; the kcat/Km for the hydrolysis of sophorose was higher than those for the hydrolysis of other β-linked glucodisaccharides. Kinetic analyses of Td2F2 activity with excess monosaccharides in the presence of pNPGlc showed that the addition of monosaccharides increases both Km and kcat. These increases in Km reflect a decreased affinity of pNPGlc for Td2F2 caused by the excess monosaccharides. Increases in kcat may be due to the transglycosylation activity of the enzyme. Differences in the kcat/Km ratios revealed that D-fucose behaved as an inhibitor at a concentration of 1000 mM. D-Glucose and D-galactose were comparatively poor inhibitors. However, L-arabinose, D-mannose, and D-xylose each stimulated enzyme activity. These results suggest that D-glucose, D-fucose, and D-galactose have a greater affinity for docking into the –1 subsite of the enzyme than L-arabinose, D-mannose, and D-xylose. The lower affinity of the latter three sugars for the –1 subsite was demonstrated by the weak activity of Td2F2 with p-nitrophenyl-α-L-arabinopyranoside, p-nitrophenyl-β-D-mannopyranoside, and p-nitrophenyl-β-D-xylopyranoside as substrates. Various monosaccharides and sugar alcohols also stimulated the activity of Td2F2 for the degradation of pNPGlc, pNPFuc, and pNPGal. However, this activity stimulation was not observed with additions of amino sugars or sugar acids. In fact, D-glucosamine inhibited Td2F2 activity. These results indicate that the OH-2 of a monosaccharide may be important for stimulating Td2F2 and that a COOH-5 group may hinder transglycosylation. The activation of Td2F2 by D-glucose was due to the transglycosylation activity of the enzyme. Notably, the combined production rates of sophorose and laminaribiose always exceeded pNP production rates when the reactants included 31.5–1000 mM D-glucose. This phenomenon may be the result of the enzyme catalyzing the reverse reaction of pNPGlc hydrolase, reproducing pNPGlc from pNP and D-glucose. This reproduced pNPGlc might then be consumed by the transglycosylation activity of the enzyme to produce glucodisaccharides.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
10
These experimental results show that Td2F2 may be defined as a β-glucosidase with remarkable transglycosylation activity. Analyses of the X-ray crystal structure of Td2F2 are currently under way. The resulting three-dimensional model of Td2F2 may provide valuable clues as to the presence of glucosyl-binding subsites and may offer an explanation not only for the enzyme’s ability to hydrolyze a broad range of dimeric substrates, but also for its high transglycosylation activity.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
11
REFERENCES
1. Bhatia, Y., Mishra, S., and Bisaria V. S. (2002) Microbial β-glucosidases: cloning, properties, and applications. Crit. Rev. Biotechnol. 22, 375-407
2. Watanabe, H., and Tokuda, G. (2010) Cellulolytic systems in insects. Annu. Rev. Entomol. 55, 609-632
3. Nomura, T., Quesada, A. L., and Kutchan, T. M. (2008) The new β- D-glucosidase in terpenoid-isoquinoline alkaloid biosynthesis in Psychotria ipecacuanha. J. Biol. Chem. 283, 34650-34659
4. Suzuki, H., Takahashi, S., Watanabe, R., Fukushima, Y., Fujita, N., Noguchi, A., Yokoyama,
R., Nishitani, K., Nishino, T., and Nakayama, T. (2006) An isoflavone conjugate-hydrolyzing
β-glucosidase from the roots of soybean (Glycine max) seedlings: purification, gene cloning, phylogenetics, and cellular localization. J. Biol. Chem. 281, 30251-30259
5. Zhou, J., Hartmann, S., Shepherd, B. K., and Poulton, J. E. (2002) Investigation of the
microheterogeneity and aglycone specificity-conferring residues of black cherry prunasin
hydrolases. Plant Physiol. 129, 1252-1264
6. Woodward, J., and Wiseman, A. (1982) Fungal and other β-D-glucosidases–their properties and applications. Enzyme Microb. Technol. 4, 73-79
7. Riou, C., Salmon, J. M., Vallier, M. J., Günata, Z., and Barre, P. (1998) Purification,
characterization, and substrate specificity of a novel highly glucose-tolerant β-glucosidase from Aspergillus oryzae. Appl. Environ. Microbiol. 64, 3607–3614
8. Decker, C. H., Visser, J., and Schreier, P. (2001) β-Glucosidase multiplicity from Aspergillus tubingensis CBS 643.92: purification and characterization of four beta-glucosidases and their
differentiation with respect to substrate specificity, glucose inhibition and acid tolerance.
Appl. Microbiol. Biotechnol. 55, 157–163
9. Hancock, S. M., Corbett, K., Fordham-Skelton, A. P., Gatehouse, J. A., and Davis, B. G.
(2005) Developing promiscuous glycosidases for glycoside synthesis: residues W433 and
E432 in Sulfolobus solfataricus β-glycosidase are important glucoside- and galactoside-specificity determinants. Chembiochem. 6, 866–875
10. Vladimir, K., and Joachim, T. (1997) Glycosylation employing bio-systems: from enzymes to
whole cells. Chem. Soc. Rev. 26, 463–473
11. Wood, T. M. (1998) Preparation of crystalline, amorphous, and dyed cellulase substrates.
Methods Enzymol. 160, 19–25
12. Uchiyama, T., and Watanabe, K. (2008) Substrate-induced gene expression (SIGEX)
screening of metagenome libraries. Nat. Protoc. 3, 1202–1212
13. Pace, C. N., Vajdos, F., Fee, L., Grimsley, G., and Gray, T. (1995) How to measure and
predict the molar absorption coefficient of a protein. Protein Sci. 4, 2411–2423
14. McIlvaine, T. C. (1921) A buffer solution for colorimetric comparison. J. Biol. Chem. 49, 183–186
15. Di Lauro, B., Rossi, M., and Moracci, M. (2006) Characterization of a β-glycosidase from thermoacidophilic bacterium Alicyclobacillus acidocaldarius. Extremophiles 10, 301–310
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
12
16. Jeng, W. Y., Wang, N. C., Lin, M. H., Liaw, Y. C., Chang, W. J., Liu, C. I., Liang, P. H., and
Wang, A. H. (2011) Structural and functional analysis of three β-glucosidases from bacterium Clostridium cellulovorancs, fungus Trichoderma reesei and termite Neotermes koshunensis. J. Struct. Biol. 173, 46–56
17. Fang, Z., Fang, W., Liu, J., Hong, Y., Peng, H., Zhang, X., Sun, B., and Xiao, Y. (2010)
Cloning and characterization of β-glucosidase from marine microbial metagenome with excellent glucose tolerance. J. Microbiol. Biotechnol. 20, 1351–1358
18. Marques, A. R., Coutinho, P. M., Videira, P., Fialho, A. M., and Sá-Correia, I. (2003)
Sphingomonas paucimobilis β-glucosidase Bgl1: a member of a new bacterial subfamily in glycoside hydrolase family 1. Biochem. J. 370, 793–804
19. Nucci, R., Moracci, M., Vaccaro, C., Vespa, N., and Rossi, M. (1993) Exo-glucosidase
activity and substrate specificity of the β-glycosidase isolated from the extreme thermophile Sulfolobus solfataricus. Biotechnol. Appl. Biochem. 17, 239–250
20. Aguilar, C. F., Sanderson, I., Moracci, M., Ciaramella, M., Nucci, R., Rossi, M., and Pearl, L.
H. (1997) Crystal structure of the β-glycosidase from the hyperthermophilic archeon Sulfolobus solfataricus: resilience as a key factor in thermostability. J. Mol. Biol. 271, 789–802
21. Chen, H., Hayn, M., and Esterbauer, H. (1992) Purification and characterization of two
extracellular β-glucosidases from Tricoderma reesei. Biochim. Biophys. Acta. 1121, 54–60 22. Hong, J., Tamaki, H., and Kumagai, H. (2006) Unusual hydrophobic linker region of
β-glucosidase (BGLII) from Thermoascus aurantiacus is required for hyper-activation by organic solvents. Appl. Microbiol. Biotechnol. 73, 80–88
23. Uchima, C. A., Tokuda, G., Watanabe, H., Kitamoto, K., and Arioka, M. (2011)
Heterologous expression and characterization of glucose-stimulated β-glucosidase from the termite Neotermes koshunensis in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 89, 1761–1771
24. Zanoelo, F. F., Polizeli Mde, L., Terenzi, H. F., and Jorge, J. A. (2004) β-Glucosidase activity from the thermophilic fungus Scytalidium thermophilum is stimulated by glucose and
xylose. FEMS Microbiol. Lett. 240, 137–143
25. Pérez-Pons, J. A., Rebordosa, X., and Querol, E. (1995) Properties of a novel
glucose-enhanced β-glucosidase purified from Streptomyces sp. (ATCC 11238). Biochim. Biophys. Acta. 1251, 145–153
26. Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam,
H., Valentin, F., Wallace, I. M., Wilm, A., Lopez, R., Thompson, J. D., Gibson, T. J., and
Higgins, D. G. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948
Acknowledgments We thank Miss T. Kikuch for help in obtaining HPAE-PAD data. This work was supported by in part
by grants from the New Energy and Industrial Technology Development Organization (NEDO).
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
13
FOOTNOTES
The abbreviations used are: GH1, glycoside hydrolase family 1; pNP, p-nitrophenol; pNPGlc,
p-nitrophenyl β-D-glucopyranoside; pNPFuc, p-nitrophenyl β-D-fucopyranoside; pNPGal,
p-nitrophenyl β-D-galactopyranoside; HPAE-PAD, high-performance anion exchange chromatography with pulsed amperometric detection; Corona CAD, corona charged aerosol detector
FIGURE LEGENDS
FIGURE 1. Sequence alignment of Td2F2 against eight similar GH1 β-glucosidases. Full-length
amino acid sequences were aligned by ClustalW2 (26). The identical residues are shown in white
with a black background and conservative changes are shown in black with a gray background. The
two catalytic glutamate residues are indicated by stars. The secondary structure elements of BglA
from Clostridium cellulovorans are also indicated graphically at the bottom: α-helices or 310-helices
are indicated by light-gray cylinders; β-strands are indicated by black arrows. The (β/α)8 barrel structure is indicated by sequential letters. The residues located in the –1 and +1 subsites, which are
described by the structure of BglA from C. cellulovorans (16), are indicated by black and white
triangles. The amino acid sequences used were Mruber (Meiothermus ruber, accession number
D3PLV5), BglA_Cthrm (Clostridium thermocellum, accession number P26208), BglA_Bacil
(Bacillus sp., accession number BAA36160), BglA_Bcirc (Bacillus circulans, accession number
Q03506), BglA_Tetha (Thermoanaerobacter ethanolicus, accession number ADD25173),
GlyB_Aacio (Alicyclobacillus acidocaldarius, accession number AAZ81839), BglB_Tbisp
(Thermobispora bispora, accession number P38645), and BglA_Ccell (C. cellulovorans, accession
number AY268940).
FIGURE 2. SDS-PAGE of purified Td2F2. Lane 1, molecular markers; lane 2, soluble protein
fraction; lane 3, fraction after affinity chromatography; lane 4, fraction after gel-filtration
chromatography.
FIGURE 3. Plot of the molecular weights of protein markers (solid circles) and the elution
volume/void volume ratio. Td2F2 was eluted at 52 kDa (open circle).
FIGURE 4. The enzymatic activity (solid circles) and stability (open circles) of Td2F2 are shown at
various (A) pH and (B) temperatures.
FIGURE 5. Effect of D-glucose on Td2F2 activity is shown for (A) 1 mM pNPGlc (solid triangles),
2 mM pNPGlc (open circles), and 10 mM pNPGlc (solid circles), and (B) 1.25 mM cellobiose (solid
triangles), 5 mM cellobiose (open circles), and 10 mM cellobiose (solid circles). The specific activity
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
14
for cellobiose was measured by the degradation of cellobiose.
FIGURE 6. The effects of various monosaccharides and their derivatives on Td2F2 activity are
shown. Substrates and additives were used at final concentrations of 5 mM and 140 mM, respectively.
The activity without any additives was taken to be 100%. The specific activity for pNPGlc, pNPFuc,
and pNPGal without additives were 9.46 ± 1.11 µmol min–1 mg–1, 19.4 ± 1.66 µmol min–1 mg–1 and
18.2 ± 1.94 µmol min–1 mg–1, respectively. The black, white, and gray bars indicate the relative activities for pNPGlc, pNPFuc, and pNPGal, respectively.
FIGURE 7. Corona CAD analyses of the reaction products of Td2F2. The reaction was performed
with 0.05 µg of Td2F2 incubated for 5 min with 10 mM pNPGlc with or without 31.5 mM D-glucose
in 20 µL of 100 mM sodium acetate buffer (pH 5.5) at 75˚C. Chromatograms are given (A) for
standard glucodisaccharides, (B) in the absence of D-glucose, and (C) in the presence of 31.5 mM
D-glucose. G = glucose, L = laminaribiose, C = cellobiose, S = sophorose, Ge = gentiobiose.
FIGURE 8. Transglycosylation activity of Td2F2. The reaction was performed with 0.05 µg Td2F2
incubated for 5 min with 10 mM pNPGlc with or without 31.5–1000 mM D-glucose in 20 µL of 100
mM sodium acetate buffer (pH 5.5) at 75˚C. The production of pNP (solid triangles), sophorose (open circles), and laminaribiose (solid circles) are shown.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
15
TABLE 1 Substrate specificity of the recombinant Td2F2
Substratea Linkage of glycosyl group Relative activity (%)
Chromogenic substrates (5 mM)
pNPGlc β-Glucose 100 ± 6.7b
pNPFuc β-Fucose 198 ± 3.1
pNPGal β-Galactose 197 ± 7.2
p-Nitrophenyl-β-D-cellobioside β-Cellobiose 24.3 ± 0.3
p-Nitrophenyl-β-D-lactopyranoside β-Lactose 46.8 ± 5.5
Oligosaccharides (5 mM)
Cellobiose β(1,4)Glucose 100 ± 4.3c
Cellotriose β(1,4)Glucose 175 ± 5.4
Cellotetraose β(1,4)Glucose 145 ± 8.4
Cellopentaose β(1,4)Glucose 84.7 ± 5.3
Cellohexaose β(1,4)Glucose 47.7 ± 6.2
Laminaribiose β(1,3)Glucose 112 ± 3.1
Laminaritriose β(1,3)Glucose 146 ± 10
Laminaritetraose β(1,3)Glucose 23.6 ± 2.9
Laminaripentaose β(1,3)Glucose 72.1 ± 7.6
Sophorose β(1,2)Glucose 154 ± 1.6
Gentiobiose β(1,6)Glucose 12.9 ± 1.7
Lactose β(1,4)Galactose 137 ± 8.6
aNo activity or poorly activity was detected with p-nitrophenyl-β-D-mannopyranoside, p-nitrophenyl-β-D-xylopyranoside,
p-nitrophenyl- N-acetyl-β-D-glucosaminide, p-nitrophenyl- 2-acetamide-2-deoxy-β-D-glucopyranoside,
p-nitrophenyl-β-L-arabinopyranoside, p-nitrophenyl-α-L-arabinopyranoside, p-nitrophenyl-α-D-glucopyranoside,
p-nitrophenyl-α-D-galactopyranoside, p-nitrophenyl-α-D-xylopyranoside, p-nitrophenyl-α-L-fucopyranoside,
p-nitrophenyl-α-L-arabinofuranoside, and p-nitrophenyl-α-L-rhamnopyranoside, oligosaccharides, such as laminarihexaose,
trehalose, kojibiose, nigerose, maltose, isomaltose, sucrose, isoprimeverose, chitobiose and xylobiose, insoluble β-glucans (2.0 mg
mL-1), such as phosphoric acid swollen cellulose and curdlan.
bThe specific activity of Td2F2 for pNPGlc was 11.6 ± 0.80 µmol min-1 mg-1, by measuring the release of pNP.
cThe specific activity of Td2F2 for cellobiose was 0.59 ± 0.03 µmol min-1 mg-1, by measuring the release of glucose.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
16
TABLE 2 Steady-state kinetic constants of the recombinant Td2F2
Substrate Km (mM) kcat (s-1) kcat/Km (s-1 mM-1)
Aryl-β-glycosides
pNPGlc 0.39 ± 0.04 12.0 ± 0.33 30.6
pNPFuc 0.33 ± 0.05 22.7 ± 0.96 68.8
pNPGal 4.41 ± 0.66 28.1 ± 1.37 6.38
Oligosaccharides
Cellobiose 4.44 ± 0.39 7.13 ± 0.23 1.61
Cellotriose 3.03 ± 0.37 10.1 ± 0.41 3.35
Cellotetraose 1.53 ± 0.11 7.87 ± 0.15 5.14
Cellopentaose 8.40 ± 1.42 7.91 ± 0.59 0.94
Laminaribiose 2.99 ± 0.20 6.78 ± 0.15 2.27
Laminaritriose 1.60 ± 0.24 6.14 ± 0.26 3.84
Laminaritetraose 6.40 ± 0.90 1.23 ± 0.06 0.19
Laminaripentaose 5.68 ± 0.77 6.48 ± 0.28 1.14
Sophorose 1.28 ± 0.14 7.58 ± 0.25 5.91
Gentiobiose 20.5 ± 3.23 3.30 ± 0.17 0.16
Lactose 11.4 ± 1.41 17.4 ± 0.86 1.53
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from
β-Glucosidase with strong transglycosylation activity
17
TABLE 3 Effects of organic solvents, metal ions and chelating agent on the enzyme activities of the recombinant Td2F2
Reagent Concentration Relative activity (%)
No additive 100 ± 5.8a
Ethanol 10% v/v 116 ± 4.3
Ethanol 25% v/v 40.2 ± 4.0
Dimethylsulfoxide 10% v/v 115 ± 3.1
Dimethylsulfoxide 25% v/v 119 ± 2.1
MnCl2 1 mM 132 ± 8.6
CaCl2 1 mM 105 ± 7.0
CoCl2 1 mM 101 ± 2.8
MgCl2 1 mM 98.0 ± 6.5
ZnCl2 1 mM 24.7 ± 3.5
FeCl3 1 mM 6.6 ± 3.0
EDTA 10 mM 99.6 ± 5.7
aThe activity without an additional regent was taken to be 100% (specific activity 10.4 ± 0.60 µmol min-1 mg-1).
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

β-Glucosidase with strong transglycosylation activity
18
TABLE 4 Effects of sugar addition for pNPGlc degradation efficiencies of the recombinant Td2F2
Additive Concentration Km kcat kcat/Km
(mM) (mM) (s-1) (s-1 mM-1)
No additive 0.39 ± 0.04 12.0 ± 0.33 30.6
D-Glucose 140 0.60 ± 0.05 18.9 ± 0.47 31.4
D-Glucose 1000 4.46 ± 0.23 130 ± 2.23 29.1
D-Fucose 140 1.52 ± 0.10 51.5 ± 1.20 33.7
D-Fucose 1000 5.77 ± 0.41 113 ± 2.80 19.5
D-Galactose 140 1.51 ± 0.22 39.8 ± 2.09 26.5
D-Galactose 1000 2.84 ± 0.41 72.1 ± 3.62 25.4
L-Arabinose 140 0.91 ± 0.10 17.9 ± 0.62 19.8
L-Arabinose 1000 2.55 ± 0.24 92.1 ± 2.95 36.2
D-Mannose 140 0.45 ± 0.04 12.9 ± 0.32 28.4
D-Mannose 1000 2.25 ± 0.14 86.8 ± 1.71 38.6
D-Xylose 140 0.95 ± 0.07 28.4 ± 0.68 29.9
D-Xylose 1000 1.97 ± 0.14 77.5 ± 1.70 39.3
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Td2F2 --------------------------------MAGERFPADFVWGAATAAYQIEGAVREDGRGVSIWDTFSHTPGKIADGTTGDVACDSYHRYGEDIGLLNALGMNAYRFSIAWPRIVPL 88Mruber --------------------------------MKRSDFPANFIWGTATSAYQIEGAVSEDGRGPSIWDTFSHTPGKTKGGDHGDVACDHYHRYPEDIALMKELGVNAYRFSVAWPRILPE 88BglA_Cthrm --------------------------------MSKITFPKDFIWGSATAAYQIEGAYNEDGKGESIWDRFSHTPGNIADGHTGDVACDHYHRYEEDIKIMKEIGIKSYRFSISWPRIFPE 88BglA_Bacil --------------------------------MASIQFPKDFVWGTATASYQIEGAYNEDGRGMSIWDTFSRTPGKVVNGDTGDIACDSYHRYEEDIALLKNLGVKAYRFSIAWPRIYPD 88BglA_Bcirc --------------------------------MSIHMFPSDFKWGVATAAYQIEGAYNEDGRGMSIWDTFSRTPGKVVNGDTGDIACDSYHRYEEDIALLKNLGVKAYRFSIAWPRIYPD 88BglA_Tetha ---------------------------------MEVKFPKDFLWGTATSSYQIEGAVNEDGRTPSIWDTFSKTEGKTYNGHTGDVACDHYHRYKEDVEILKEIGVKAYRFSIAWPRIFPE 87GlyB_Aacio -------------------------------MREMRKFPEGFVWGTATASYQVEGAAREGGRGRSIWDTFSHTPGKVAEGHTGDVACDHYHRYQDDVRLMKELGISSYRFSIAWPRVMPE 89BglB_Tbisp MTESAMTSRAGRGRGADLVAAVVQGHAAASDAAGDLSFPDGFIWGAATAAYQIEGAWREDGRG--LWDVFSHTPGKVASGHTGDIACDHYHRYADDVRLMAGLGDRVYRFSVAWPRIVPD 118BglA_Ccell --------------------------------MEKLRFPKDFIFGTATAAYQIEGAYKEDEKGESIWDRFSHIPGNVAKMHNGDIACDHYHRYKEDVQLLKSLGIKSYRFSIAWPRIFPK 88
Td2F2 ADREVY-PEGLYDILLRLHNDYPFRPLYITENGCALHDEIAEDGGIHDGQRQAFFEAHLAQLQRALAAGVPLKGYFAWSLLDNFEWAMGLSMRYGICYTNFETLERRIKDSGYWLRDFIA 441Mruber MDWEVY-PEGLYDLLIRLGREYRPKAIYITENGAAYPDAVADDGGIHDLERVRYFQRHLALCLEALQHGAPLKGYFAWSLLDNFEWAEGYAKRFGLVYVDFPSQRRRIKASGYWFRDFLR 438BglA_Cthrm MGWIIY-PEGLYDLLMLLDRDYGKPNIVISENGAAFKDEIGSNGKIEDTKRIQYLKDYLTQAHRAIQDGVNLKAYYLWSLLDNFEWAYGYNKRFGIVHVNFDTLERKIKDSGYWYKEVIK 444BglA_Bacil MDWNVY-AEGLYKVLSWVHEEYGDVPIYITENGACYEDELTQEGRVHDAKRADYFKKHFIQCHRLIESGVPLKGYFAWSLLDNFEWAEGYVKRFGIVYTDYKTLKRYPKDSYRFIQSVIE 442BglA_Bcirc IGWEIY-AEGLYDLLRYTADKYGNPTLYITENGACYNDGLSLDGRIHDQRRIDYLAMHLIQASRAIEDGINLKGYMEWSLMDNFEWAEGYGMRFGLVHVDYDTLVRTPKDSFYWYKGVIS 444BglA_Tetha MGWEIS-PESLYDLLKRLDREYTKLPMYITENGAAFKDEVTEDGRVHDDERIEYIKEHLKAAAKFIGERGNLKGYFVWSLMDNFEWAHGYSKRFGIVYVDYETQKRILKDSALWYKEVIQ 442GlyB_Aacio MDWEVY-PDGLYDLLSRLRRDYGDIPIYITENGAAFDDRVQ-DGGVHDADRVAYLAGHFAAAHRFLEEGGNLRGYYVWSLMDNFEWAFGYTKRFGIVYVDYDTLARIPKDSYFWYQRVIR 442BglB_Tbisp TAWPGDRPDGLRTLLLRLSRDYPGVGLIITENGAAFDDRAD-GDRVHDPERIRYLTATLRAVHDAIMAGADLRGYFVWSVLDNFEWAYGYHKR-GIVYVDYTTMRRIPRESALWYRDVVR 465BglA_Ccell MGWEIY-PQGLYDLLTRIHRDYGNIDLYITENGAAFNDMVNRDGKVEDENRLDYLYTHFAAALSAIEAGVPLKGYYIWSFMDNFEWAEGYEKRFGIVHVNYKTQERTIKKSAYWYKELIE 441
Td2F2 GQRG---------- 445Mruber EAVASS-------- 444BglA_Cthrm -NNGF--------- 448BglA_Bacil -NDGFEA------- 448BglA_Bcirc -RGWLDL------- 450BglA_Tetha -KNSMS-------- 447GlyB_Aacio EGGLVPAEPAETAR 456BglB_Tbisp RNGLRNGE------ 473 BglA_Ccell RSNKLE-------- 447
Td2F2 GAGPINQAGLDHYSRMVDALLGAGLQPFVTLYHWDLPQPLEDRLG-WGSRATATVFAEYADIVVRQLGDRVTHWATLNEPWCSAMLGYYLGVHAPGHTDLKRGLEASHNLLLGHGLAVQA 207Mruber GRGRVNPRGLDFYNRLVDALLEQGITPWATLYHWDLPQSLEDQGG-WPSRETAYAFAEYADLVTRHLGDRVKHWITLNEPWCSAYLGYHAGIHAPGQQNFKHSIWASHHLLLAHGLAVPV 207BglA_Cthrm GTGKLNQKGLDFYKRLTNLLLENGIMPAITLYHWDLPQKLQDKGG-WKNRDTTDYFTEYSEVIFKNLGDIVPIWFTHNEPGVVSLLGHFLGIHAPGIKDLRTSLEVSHNLLLSHGKAVKL 207BglA_Bacil GDGELNQKGLDYYAKVIDGLLAAGIEPCVTLYHWDLPQALQDKGG-WDNRDTIRAFVRYAETAFKAFGGKVKQWITFNETWCVSFLSNYIGAHAPGNTDLQLAVNVAHNCMVAHGEAVKA 207BglA_Bcirc GTGEVNRAGLDYYHRLVDELLANGIEPFCTLYHWDLPQALQDQGG-WGSRITIDAFAEYAELMFKELGGKIKQWITFNEPWCMAFLSNYLGVHAPGNKDLQLAIDVSHHLLVAHGRAVTL 207BglA_Tetha -EGKYNSKGMDFYKRLVDELLKKDIMPTATIYHWDLPQWAYDKGGGWLNRDSVKWYVEYATKLFEELGDVIPLWITHNEPWCASILSYGIGEHAPGHKNYREALIAAHHILLSHGEAVKA 206GlyB_Aacio -KGRVWVKGLDFYKRLATALLEHGIRPAATMYHWDLPQWMEDEGG-WNSRETVSRFLEYSEILFRELGDLVPMWITHNEPWCASILGYGIGVHAPGLKDWRRAYRAAHHLLLSHGQAVRL 207BglB_Tbisp GSGPVNPAGLDFYDRLVDELLGHGITPYPTLYHWDLPQTLEDRGG-WAARDTAYRFAEYALAVHRRLGDRVRCWITLNEPWVAAFLATHRG--APGAADVPR-FRAVHHLLLGHGLGLR- 233BglA_Ccell GFGEINQKGIQFYRDLIDELIKNDIEPAITIYHWDLPQKLQDIGG-WANPQVADYYVDYANLLFREFGDRVKTWITHNEPWVASYLGYALGVHAPGIKDMKMALLAAHNILLSHFKAVKA 207
Td2F2 MRAAAPQPLQIGIVLNLTPTYPASDSPEDVAAARRFDGFVNRWFLDPLAGRGYPQDMLDYYGA---AAPQANPEDLTQIAAPLDWLGVNYYERMRAVDAPDAS--LPQAQRLDDPDLPHT 322Mruber IRRNVTG-ARVGITLNLSPGYPASPDPADVAAARRFDGFQNRWYLDPLYGLGYPADMLALYG----EAPSVQGDDLITIAAPTDFLGINYYSRA-VVRNSDLE--PYRFQYVRVGEEHTD 319BglA_Cthrm FREMNIDA-QIGIALNLSYHYPASEKAEDIEAAELSFSLAGRWYLDPVLKGRYPENALKLYKKKGIELSFP-EDDLKLISQPIDFIAFNNYSSEFIKYDPSSESGFSPANSILEKFEKTD 325BglA_Bacil FRALGISG-EIGTTHNLYWFEPYTTKPEDVAAAHRNRAYNNEWFMDPTFKGQYPQFMVDWFKGKGVEVPIQ-PGDMETIAQPIDFIGVNFYSGGFGRYKEGE--GLFDCEEVQVGFDKTF 323BglA_Bcirc FRELGISG-EIGIAPNTSWAVPYRRTKEDMEACLRVNGWSGDWYLDPIYFGEYPKFMLDWYENLGYKPPIV-DGDMELIHQPIDFIGINYYTSSMNRYNPGEAGGMLSSEAISMGAPKTD 325BglA_Tetha FREMNIKGSKIGITLNLTPAYPASEKEEDKLAAQYADGFANRWFLDPIFKGNYPEDMMELYSKIIGEFDFIKEGDLKTISVPIDFLGVNYYTRSIVKYNEDS---MLKAENVPGPGKRTE 323GlyB_Aacio YRELGLPG-EIGITLNLTPVYAATPNPEDLAAADRQDMFQNRWFLDPVLRGEYPEELLHRVDQVVGGFDAVKPGDLDVIATPIDFLGVNYYTRAVVADDPSEP--LLSVRHVPGEGPRTE 324BglB_Tbisp LRSAGAG--QLGLTLSLSPVIEARPGVRG--GGRRVDALANRQFLDPALRGRYPEEVLKIMAG-HARLGHP-GRDLETIHQPVDLLGVNYYSHVRLAAEGEPANRLPGSEGIRFERPTAV 347BglA_Ccell YRELEQDG-QIGITLNLSTCYSNSADEEDIAAAHRSDGWNNRWFLDAALKGTYPEDMIKIFSDTNIMPELP-KELFTEVFETSDFLGINYYTRQVVKNN--SE-AFIGAESVAMDNPKTE 322
β1 β2 α1
β3 β4 α2 α3 α4
β5 β6 α5
β7 β8 α6 α7 α8
Figure 1, Uchiyama et al.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

kDa 1. 2. 3. 4.
25
37
50
75
250
150
100
Figure 2, Uchiyama et al.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 3, Uchiyama et al.
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
1.9 2 2.1 2.2 2.3 2.4 2.5 2.6 2.7
Elution volume/Void volume
Mol
ecul
ar w
eigh
t Aldolase (158 kDa)
Conalbumin (75 kDa)
Carbonic anhydrase (29 kDa) Ribonuclease A (13.7 kDa)
Aprotinin (6.5 kDa)
Td2F2 (52 kDa) z by guest on Septem
ber 6, 2018http://w
ww
.jbc.org/D
ownloaded from
0
25
50
75
100
125
20 30 40 50 60 70 80
Rel
ativ
e ac
tivity
(%)
Rel
ativ
e ac
tivity
(%)
pH
Temperature (˚C)
A.
B.
Figure 4, Uchiyama et al.
0
20
40
60
80
100
120
3.0 4.0 5.0 6.0 7.0 8.0 by guest on Septem
ber 6, 2018http://w
ww
.jbc.org/D
ownloaded from

A. A
ctiv
ity (µ
mol
min
-1 m
g-1)
B.
Act
ivity
(µm
ol m
in-1
mg-1
)
D-Glucose concentration (mM)
0
10
20
30
40
50
60
70
0 200 400 600 800 1000 1200 0
5
10
15
20
25
30
0 200 400 600 800 1000 1200
Figure 5, Uchiyama et al.
D-Glucose concentration (mM)
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Rel
ativ
e ac
tivity
(%)
0
100
200
300
400
500
600
700
Figure 6, Uchiyama et al.
No
addi
tive
D-G
luco
se
L-G
luco
se
D-M
anno
se
D-F
ruct
ose
D-A
rabi
nose
L-A
rabi
nose
D-F
ucos
e
L-Fu
cose
Met
hyl-β
-D-g
luco
se
Met
hyl- α
-D-g
luco
se
D-G
alac
tose
2-D
eoxy
D-R
ibos
e
D-R
ibos
e
D-X
ylos
e
L-R
ham
nose
L-So
rbos
e
N-A
cety
l-D-g
luco
sam
ine
Sedo
hept
ulos
e
D-G
luco
sam
ine
Xyl
itol
Glu
cito
l
Ara
bito
l
Man
nito
l
D-G
lucu
rona
te
D-G
alac
turo
nate
D-G
luco
nate
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5 10 15 20 25
5 10 15 20 25
5 10 15 20 25
500000
400000
300000
200000
100000
0
-100000 0
500000
400000
300000
200000
100000
0 -100000
500000
400000
300000
200000
100000
0 -100000
0
0 Retention time (min)
Retention time (min)
Retention time (min)
Inte
nsity
(µV
) In
tens
ity (µ
V)
Inte
nsity
(µV
)
G
G
G
L S Ge
C
L
S
Ge
C
A.
B.
C.
Figure 7, Uchiyama et al.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

0
200
400
600
800
1000
1200
1400
0 200 400 600 800 1000 1200
Prod
uct c
once
ntra
tion
(µM
)
D-Glucose concentration (mM)
Figure 8, Uchiyama et al.
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Taku Uchiyama, Kentaro Miyazaki and Katsuro Yaoiwith strong transglycosylation activity
-glucosidase from a compost microbial metagenomeβCharacterization of a novel
published online May 9, 2013J. Biol. Chem.
10.1074/jbc.M113.471342Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on September 6, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















