
Interaction of Self-Assembling β-Sheet Peptides with Phospholipid Monolayers: The Role of...
8
Interaction of Self-Assembling -Sheet Peptides with Phospholipid Monolayers: The Role of Aggregation State, Polarity, Charge and Applied Field Elisabeth Protopapa, Steven Maude, Amalia Aggeli, and Andrew Nelson* Centre for Self-Organising Molecular Systems (SOMS), School of Chemistry, UniVersity of Leeds, Leeds, LS2 9JT, U.K. ReceiVed October 11, 2008. ReVised Manuscript ReceiVed December 12, 2008 Studies of -sheet peptide/phospholipid interactions are important for an understanding of the folding of -sheet- rich membrane proteins and the action of antimicrobial and toxic peptides. Further, self-assembling peptides have numerous applications in medicine and therefore an insight is required into the relation between peptide molecular structure and biomembrane activity. We previously developed one of the simplest known model peptide systems which, above a critical concentration (c*) in solution, undergoes nucleated one-dimensional self-assembly from a monomeric random coil into a hierarchy of well defined -sheet structures. Here we examine the effects of peptide aggregation, polarity, charge, and applied field on peptide interactions with dioleoyl phosphatidylcholine (DOPC) monolayers using electrochemical techniques. The interactions of six systematically altered 11 residue -sheet tape- forming peptides were investigated. The following findings with respect to 11 residue -sheet peptide-DOPC interaction arose from the study: (i) The solution monomer peptide species is the monolayer active moeity. (ii) Amphiphilic peptides are more monolayer active than polar peptides in the absence of applied electric field. (iii) Positive charge on amphiphilic peptides facilitates monolayer interaction in the absence of applied electric field. (iv) Negative applied electric field facilitates monolayer interaction with positively charged amphiphilic and polar peptides. (v) Neutral amphiphilic peptides permeabilize DOPC layers to ions to the greatest extent. (vi) The -sheet tape forming peptides are shown to be significantly less monolayer disruptive than antimicrobial peptides. These conclusions will greatly contribute to the rational design of new peptide-based biomaterials and biosensors. Introduction The self-assembly of proteins and peptides has attracted a great deal of interest over the past few years. 1-12 We previously developed model peptide systems which, above a critical concentration (c*) in solution, undergo nucleated one-dimensional self-assembly from a monomeric random coil into well-defined, twisted, micrometer-long, -sheet aggregates as a function of peptide concentration. 13-21 A theoretical model was developed which quantitatively describes peptide self-assembly 22,23 and explains the structure and stability of amyloid fibrils formed by more complex proteins in vivo. These self-assembling peptides have been explored as candidates for a variety of applications, such as templates for nanostructured inorganic materials, 24,25 surface nanocoatings, 26 and lubricants for early stage osteoar- thritis. 27 The ability of these simple peptides to self-assemble in a well-defined manner can be utilized to carry out systematic studies of peptide self-assembly in solution. 28-30 It is equally * Corresponding author. Phone: +44 (0)113 343 4992. Fax: +44 (0)113 343 6452. E-mail: [email protected]. (1) Collier, J. H.; Hu, B. H.; Ruberti, J. W.; Zhang, J.; Shum, P.; Thompson, D. H.; Messersmith, P. B. J. Am. Chem. Soc. 2001, 123(38), 9463–9464. (2) de la Paz, M. L.; Goldie, K.; Zurdo, J.; Lacroix, E.; Dobson, C. M.; Hoenger, A.; Serrano, L. Proc. Natl. Acad. Sci. U.S.A. 2002, 99(25), 16052–16057. (3) Hartgerink, J. D.; Beniash, E.; Stupp, S. I. Science 2001, 294(5547), 1684– 1688. (4) Mart, R. J.; Osborne, R. D.; Stevens, M. M.; Ulijn, R. V. Soft Matter 2006, 2(10), 822–835. (5) Matsumura, S.; Uemura, S.; Mihara, H. Supramol. Chem. 2006, 18(5), 397–403. (6) Ozbas, B.; Kretsinger, J.; Rajagopal, K.; Schneider, J. P.; Pochan, D. J. Macromolecules 2004, 37(19), 7331–7337. (7) Reches, M.; Gazit, E. Nat. Nanotechnol. 2006, 1, 195–200. (8) Rogers, S. S.; Venema, P.; Sagis, L. M. C.; van der Linden, E.; Donald, A. M. Macromolecules 2005, 38(7), 2948–2958. (9) Smith, A. M.; Banwell, E. F.; Edwards, W. R.; Pandya, M. J.; Woolfson, D. N. AdV. Funct. Mater. 2006, 16(8), 1022–1030. (10) Smith, J. F.; Knowles, T. P. J.; Dobson, C. M.; MacPhee, C. E.; Welland, M. E. Proc. Natl. Acad. Sci. U.S.A. 2006, 103(43), 15806–15811. (11) Stendahl, J. C.; Rao, M. S.; Guler, M. O.; Stupp, S. I. AdV. Funct. Mater. 2006, 16(4), 499–508. (12) Zhang, S. G.; Holmes, T.; Lockshin, C.; Rich, A. Proc. Natl. Acad. Sci. U.S.A. 1993, 9(8), 3334–3338. (13) Aggeli, A.; Bell, M.; Boden, N.; Keen, J. N.; Knowles, P. F.; McLeish, T. C. B.; Pitkeathly, M.; Radford, S. E. Nature 1997, 386(6622), 259–262. (14) Aggeli, A.; Bell, M.; Boden, N.; Keen, J. N.; McLeish, T. C. B.; Nyrkova, I.; Radford, S. E.; Semenov, A. J. Mat. Chem. 1997, 7(7), 1135–1145. (15) Aggeli, A.; Boden, N.; Zhang, S. Self-Assembling Peptide Systems in Biology, Medicine and Engineering; Kluwer Academic Publishers: Dordecht, 2001. (16) Aggeli, A.; Fytas, G.; Vlassopoulos, D.; McLeish, T. C. B.; Mawer, P. J.; Boden, N. Biomacromolecules 2001, 2(2), 378–388. (17) Aggeli, A.; Nyrkova, I. A.; Bell, M.; Harding, R.; Carrick, L.; McLeish, T. C. B.; Semenov, A. N.; Boden, N. Proc. Natl. Acad. Sci. U.S.A. 2001, 98(21), 11857–11862. (18) Carrick, L.; Tassieri, M.; Waigh, T. A.; Aggeli, A.; Boden, N.; Bell, C.; Fisher, J.; Ingham, E.; Evans, R. M. L. Langmuir 2005, 21(9), 3733–3737. (19) Davies, R. P. W.; Aggeli, A.; Beevers, A. J.; Boden, N.; Carrick, L. M.; Fishwick, C. W. G.; McLeish, T. C. B.; Nyrkova, I.; Semenov, A. N. Supramol. Chem. 2006, 18(5), 435–443. (20) Kayser, V.; Turton, D. A.; Aggeli, A.; Beevers, A.; Reid, G. D.; Beddard, G. S. J. Am. Chem. Soc. 2004, 126(1), 336–343. (21) Fishwick, C. W. G.; Beevers, A. J.; Carrick, L. M.; Whitehouse, C. D.; Aggeli, A.; Boden, N. Nano Lett. 2003, 3(11), 1475–1479. (22) Nyrkova, I. A.; Semenov, A. N.; Aggeli, A.; Bell, M.; Boden, N.; McLeish, T. C. B. Eur. Phys. J. B 2000, 17(3), 499–513. (23) Nyrkova, I. A.; Semenov, A. N.; Aggeli, A.; Boden, N. Eur. Phys. J. B 2000, 17(3), 481–497. (24) Meegan, J. E.; Aggeli, A.; Boden, N.; Brydson, R.; Brown, A. P.; Carrick, L.; Brough, A. R.; Hussain, A.; Ansell, R. J. AdV. Funct. Mater. 2004, 14(1), 31–37. (25) Firth, A.; Aggeli, A.; Burke, J. L.; Yang, X. B.; Kirkham, J. Nanomedicine 2006, 1(2), 189–199. (26) Whitehouse, C.; Fang, J. Y.; Aggeli, A.; Bell, M.; Brydson, R.; Fishwick, C. W. G.; Henderson, J. R.; Knobler, C. M.; Owens, R. W.; Thomson, N. H.; Smith, D. A.; Boden, N. Angew. Chem., Int. Ed. 2005, 44(13), 1965–1968. (27) Bell, C. J.; Carrick, L. M.; Katta, J.; Jin, Z. M.; Ingham, E.; Aggeli, A.; Boden, N.; Waigh, T. A.; Fisher, J. J. Biomed. Mater. Res. A 2006, 78A(2), 236–246. (28) Aggeli, A.; Bell, M.; Boden, N.; Carrick, L. M.; Strong, A. E. Angew. Chem., Int. Ed. 2003, 42(45), 5603–5606. 3289 Langmuir 2009, 25, 3289-3296 10.1021/la803368r CCC: $40.75 2009 American Chemical Society Published on Web 01/27/2009
Transcript of Interaction of Self-Assembling β-Sheet Peptides with Phospholipid Monolayers: The Role of...

Interaction of Self-Assembling -Sheet Peptides with PhospholipidMonolayers: The Role of Aggregation State, Polarity, Charge and
Applied Field
Elisabeth Protopapa, Steven Maude, Amalia Aggeli, and Andrew Nelson*
Centre for Self-Organising Molecular Systems (SOMS), School of Chemistry, UniVersity of Leeds,Leeds, LS2 9JT, U.K.
ReceiVed October 11, 2008. ReVised Manuscript ReceiVed December 12, 2008
Studies of -sheet peptide/phospholipid interactions are important for an understanding of the folding of -sheet-rich membrane proteins and the action of antimicrobial and toxic peptides. Further, self-assembling peptides havenumerous applications in medicine and therefore an insight is required into the relation between peptide molecularstructure and biomembrane activity. We previously developed one of the simplest known model peptide systemswhich, above a critical concentration (c*) in solution, undergoes nucleated one-dimensional self-assembly from amonomeric random coil into a hierarchy of well defined -sheet structures. Here we examine the effects of peptideaggregation, polarity, charge, and applied field on peptide interactions with dioleoyl phosphatidylcholine (DOPC)monolayers using electrochemical techniques. The interactions of six systematically altered 11 residue -sheet tape-forming peptides were investigated. The following findings with respect to 11 residue -sheet peptide-DOPC interactionarose from the study: (i) The solution monomer peptide species is the monolayer active moeity. (ii) Amphiphilicpeptides are more monolayer active than polar peptides in the absence of applied electric field. (iii) Positive chargeon amphiphilic peptides facilitates monolayer interaction in the absence of applied electric field. (iv) Negative appliedelectric field facilitates monolayer interaction with positively charged amphiphilic and polar peptides. (v) Neutralamphiphilic peptides permeabilize DOPC layers to ions to the greatest extent. (vi) The -sheet tape forming peptidesare shown to be significantly less monolayer disruptive than antimicrobial peptides. These conclusions will greatlycontribute to the rational design of new peptide-based biomaterials and biosensors.
Introduction
The self-assembly of proteins and peptides has attracted agreat deal of interest over the past few years.1-12 We previouslydeveloped model peptide systems which, above a criticalconcentration (c*) in solution, undergo nucleated one-dimensionalself-assembly from a monomeric random coil into well-defined,twisted, micrometer-long, -sheet aggregates as a function ofpeptide concentration.13-21 A theoretical model was developed
which quantitatively describes peptide self-assembly22,23 andexplains the structure and stability of amyloid fibrils formed bymore complex proteins in vivo. These self-assembling peptideshave been explored as candidates for a variety of applications,such as templates for nanostructured inorganic materials,24,25
surface nanocoatings,26 and lubricants for early stage osteoar-thritis.27 The ability of these simple peptides to self-assemble ina well-defined manner can be utilized to carry out systematicstudies of peptide self-assembly in solution.28-30 It is equally
* Corresponding author. Phone: +44 (0)113 343 4992. Fax: +44 (0)113343 6452. E-mail: [email protected].
(1) Collier, J. H.; Hu, B. H.; Ruberti, J. W.; Zhang, J.; Shum, P.; Thompson,D. H.; Messersmith, P. B. J. Am. Chem. Soc. 2001, 123(38), 9463–9464.
(2) de la Paz, M. L.; Goldie, K.; Zurdo, J.; Lacroix, E.; Dobson, C. M.; Hoenger,A.; Serrano, L. Proc. Natl. Acad. Sci. U.S.A. 2002, 99(25), 16052–16057.
(3) Hartgerink, J. D.; Beniash, E.; Stupp, S. I. Science 2001, 294(5547), 1684–1688.
(4) Mart, R. J.; Osborne, R. D.; Stevens, M. M.; Ulijn, R. V. Soft Matter 2006,2(10), 822–835.
(5) Matsumura, S.; Uemura, S.; Mihara, H. Supramol. Chem. 2006, 18(5),397–403.
(6) Ozbas, B.; Kretsinger, J.; Rajagopal, K.; Schneider, J. P.; Pochan, D. J.Macromolecules 2004, 37(19), 7331–7337.
(7) Reches, M.; Gazit, E. Nat. Nanotechnol. 2006, 1, 195–200.(8) Rogers, S. S.; Venema, P.; Sagis, L. M. C.; van der Linden, E.; Donald,
A. M. Macromolecules 2005, 38(7), 2948–2958.(9) Smith, A. M.; Banwell, E. F.; Edwards, W. R.; Pandya, M. J.; Woolfson,
D. N. AdV. Funct. Mater. 2006, 16(8), 1022–1030.(10) Smith, J. F.; Knowles, T. P. J.; Dobson, C. M.; MacPhee, C. E.; Welland,
M. E. Proc. Natl. Acad. Sci. U.S.A. 2006, 103(43), 15806–15811.(11) Stendahl, J. C.; Rao, M. S.; Guler, M. O.; Stupp, S. I. AdV. Funct. Mater.
2006, 16(4), 499–508.(12) Zhang, S. G.; Holmes, T.; Lockshin, C.; Rich, A. Proc. Natl. Acad. Sci.
U.S.A. 1993, 9(8), 3334–3338.(13) Aggeli, A.; Bell, M.; Boden, N.; Keen, J. N.; Knowles, P. F.; McLeish,
T. C. B.; Pitkeathly, M.; Radford, S. E. Nature 1997, 386(6622), 259–262.(14) Aggeli, A.; Bell, M.; Boden, N.; Keen, J. N.; McLeish, T. C. B.; Nyrkova,
I.; Radford, S. E.; Semenov, A. J. Mat. Chem. 1997, 7(7), 1135–1145.(15) Aggeli, A.; Boden, N.; Zhang, S. Self-Assembling Peptide Systems in
Biology, Medicine and Engineering; Kluwer Academic Publishers: Dordecht,2001.
(16) Aggeli, A.; Fytas, G.; Vlassopoulos, D.; McLeish, T. C. B.; Mawer, P. J.;Boden, N. Biomacromolecules 2001, 2(2), 378–388.
(17) Aggeli, A.; Nyrkova, I. A.; Bell, M.; Harding, R.; Carrick, L.; McLeish,T. C. B.; Semenov, A. N.; Boden, N. Proc. Natl. Acad. Sci. U.S.A. 2001, 98(21),11857–11862.
(18) Carrick, L.; Tassieri, M.; Waigh, T. A.; Aggeli, A.; Boden, N.; Bell, C.;Fisher, J.; Ingham, E.; Evans, R. M. L. Langmuir 2005, 21(9), 3733–3737.
(19) Davies, R. P. W.; Aggeli, A.; Beevers, A. J.; Boden, N.; Carrick, L. M.;Fishwick, C. W. G.; McLeish, T. C. B.; Nyrkova, I.; Semenov, A. N. Supramol.Chem. 2006, 18(5), 435–443.
(20) Kayser, V.; Turton, D. A.; Aggeli, A.; Beevers, A.; Reid, G. D.; Beddard,G. S. J. Am. Chem. Soc. 2004, 126(1), 336–343.
(21) Fishwick, C. W. G.; Beevers, A. J.; Carrick, L. M.; Whitehouse, C. D.;Aggeli, A.; Boden, N. Nano Lett. 2003, 3(11), 1475–1479.
(22) Nyrkova, I. A.; Semenov, A. N.; Aggeli, A.; Bell, M.; Boden, N.; McLeish,T. C. B. Eur. Phys. J. B 2000, 17(3), 499–513.
(23) Nyrkova, I. A.; Semenov, A. N.; Aggeli, A.; Boden, N. Eur. Phys. J. B2000, 17(3), 481–497.
(24) Meegan, J. E.; Aggeli, A.; Boden, N.; Brydson, R.; Brown, A. P.; Carrick,L.; Brough, A. R.; Hussain, A.; Ansell, R. J. AdV. Funct. Mater. 2004, 14(1),31–37.
(25) Firth, A.; Aggeli, A.; Burke, J. L.; Yang, X. B.; Kirkham, J. Nanomedicine2006, 1(2), 189–199.
(26) Whitehouse, C.; Fang, J. Y.; Aggeli, A.; Bell, M.; Brydson, R.; Fishwick,C. W. G.; Henderson, J. R.; Knobler, C. M.; Owens, R. W.; Thomson, N. H.;Smith, D. A.; Boden, N. Angew. Chem., Int. Ed. 2005, 44(13), 1965–1968.
(27) Bell, C. J.; Carrick, L. M.; Katta, J.; Jin, Z. M.; Ingham, E.; Aggeli, A.;Boden, N.; Waigh, T. A.; Fisher, J. J. Biomed. Mater. Res. A 2006, 78A(2),236–246.
(28) Aggeli, A.; Bell, M.; Boden, N.; Carrick, L. M.; Strong, A. E. Angew.Chem., Int. Ed. 2003, 42(45), 5603–5606.
3289Langmuir 2009, 25, 3289-3296
10.1021/la803368r CCC: $40.75 2009 American Chemical SocietyPublished on Web 01/27/2009

interesting to investigate the interactions of self-assemblingpeptides with phospholipid layers31,32 as model biologicalmembranes.
The potential usage of such peptides for medical applicationsnecessitates a correlation of a peptide’s molecular structure withits biological membrane activity. Peptides with an increasedtendency to interact with the phospholipid bilayer might exhibitcell membrane toxicity, an undesirable property for peptidesdesigned for tissue engineering applications. Studies of theinteractions of model -sheet-forming peptides with lipid layersare also important for the understanding of folding of -sheet-rich integral membrane proteins, such as porins, as well as theaction of some antimicrobial and toxic peptides, such as pore-forming R-hemolysin. Amyloid peptides are another class oftoxic -sheet peptides associated with at least 16 different diseases.More common examples of these diseases with the causativeprotein/peptide in parentheses include: Alzheimer’s disease (-amyloid), type 2 diabetes mellitus (amalyn), Parkinson’s disease(R-synuclein), transmissible spongiform encephalopathy (prion),Huntington’s disease (polyglutamine repeats), medullary car-cinoma of the thyroid (calcitonin), and amyloidosis (immuno-globulin). Most of the amyloid peptides can form transmembraneion channels with a number of common features such asspontaneous membrane insertion and large, heterogeneous singlechannel conductance.33,34 It is possible that amyloidogenic, toxic,and antimicrobial peptides share similar mechanisms of membranedisruption and toxicity. If this is the case, then peptide-lipidinteractions may be crucial in the pathology of amyloid diseasesin vivo.
Here we examine the effects of four factors, namely, peptideaggregation, polarity, net charge, and applied electric field, onpeptide interactions with phospholipid monolayers.31,32 In theseexperiments we employ six systematically altered self-assembling-sheet tape-forming peptides whose structures together withtheir respective hydrophobicities expressed as the Black andMould index35 are displayed in Figure 1. Peptide P11-2(0) wasdesigned as a -strand conformation with one polar and oneapolar side which self-assembles into -tapes that stack intofibrils in water.17 P11-2(0) has two ionizable groups arginine andglutamic acid at positions 3 and 9, respectively, which gives ita net zero charge at pH 7. Peptide P11-4(-2) differs from P11-2(0) in that the glutamine side chains at positions 5 and 7 havebeen replaced by glutamic acid, which gives it a net -2 chargeat pH 7. In contrast, Peptide P11-8(+2) has basic ornithine sidechains at positions 5 and 7, which give the molecule an overall+2 net charge at pH 7. Peptides P11-1(0), P11-20(-2), and P11-19(+2) are the polar versions of P11-2(0), P11-4(-2), and P11-8(+2), respectively. All six peptides were used in experimentsat solution concentrations below 30 µmol dm-3. In the experimentswith peptides at solution concentrations above 30 µmol dm-3,experiments were limited to the five peptides that exemplifiedthe most significant interactions with the DOPC monolayers.
Peptide interaction with biological membranes can disrupt themembrane structure. The barrel-stave model is one of the three
model membrane rupturing mechanisms reported to date.36 Theother feasible mechanisms known are carpet37,38 and toroidal39,40
models. The barrel-stave model describes peptides that traversemembranes and self-assemble in pores. Systems that follow thismodel are exhibited by the antimicrobial peptides melitin andalamethicin. The carpet model describes the behavior of thosepeptides that do not form pores through the membrane but lieparallel to the surface of the membrane. This may lead todestruction and solubilization of the membrane. These peptidesprefer to self-associate with other peptide monomers in a “carpet”-like manner. In the toroidal model, the phospholipid membranebends back on itself like the inside of a torus. This model canexplain the pore formation of some antimicrobial peptides thatcould not be fitted with the barrel-stave model. These threecategories are extreme situations presented as an aid to description,and generally, the actual mechanism of interaction lies somewherein between. In addition, one cannot exclude other possiblemechanisms of interaction. Several molecular properties of thepeptides are known to promote interaction with the phospholipidcomponent of the membrane, such as hydrophobicity,41 thenumber of residues42 and the presence of specific peptide residuessuch as tryptophan.31,42
In order to study the interaction of peptides with biologicalmembranes, biological membrane models are often used.43 Themembrane models used to study peptide-phospholipid interactionhave ranged from free-standing bilayers43 to monolayers at theair-water44 interface to supported monolayers and bilayers.45
One of the most powerful supported membrane models available
(29) Aggeli, A.; Bell, M.; Carrick, L. M.; Fishwick, C. W. G.; Harding, R.;Mawer, P. J.; Radford, S. E.; Strong, A. E.; Boden, N. J. Am. Chem. Soc. 2003,125(32), 9619–9628.
(30) Carrick, L. M.; Aggeli, A.; Boden, N.; Fisher, J.; Ingham, E.; Waigh,T. A. Tetrahedron 2007, 63(31), 7457–7467.
(31) Protopapa, E.; Aggeli, A.; Boden, N.; Knowles, P. F.; Salay, L. C.; Nelson,A. Med. Eng. Phys. 2006, 28(10), 944–955.
(32) Salay, L. C. Self-assembly and conductance of rationally designed -sheetion channels in lipid bilayers. PhD dissertation, University of Leeds, Leeds, 2002.
(33) Kagan, B. L.; Hirakura, Y.; Azimov, R.; Azimova, R.; Lin, M. C. Peptides2002, 23(7), 1311–1315.
(34) Jang, H.; Zheng, J.; Nussinov, R. Biophys. J. 2007, 93(6), 1938–1949.(35) Black, S. D.; Mould, D. R. Anal. Biochem. 1991, 193(1), 72–82.
(36) Juvvadi, P.; Vunnam, S.; Merrifield, R. B. J. Am. Chem. Soc. 1996, 118(38),8989–8997.
(37) Pouny, Y.; Rapaport, D.; Mor, A.; Nicolas, P.; Shai, Y. Biochemistry1992, 31(49), 12416–12423.
(38) Wu, M. H.; Maier, E.; Benz, R.; Hancock, R. E. W. Biochemistry 1999,38(22), 7235–7242.
(39) Ludtke, S. J.; He, K.; Heller, W. T.; Harroun, T. A.; Yang, L.; Huang,H. W. Biochemistry 1996, 35(43), 13723–13728.
(40) Mor, A.; Nicolas, P. J. Biol. Chem. 1994, 269(3), 1934–1939.(41) Tang, Y. C.; Deber, C. M. Biopolymers 2004, 76(2), 110–118.(42) de Planque, M. R. R.; Boots, J. W. P.; Rijkers, D. T. S.; Liskamp, R. M. J.;
Greathouse, D. V.; Killian, J. A. Biochemistry 2002, 41(26), 8396–8404.(43) Salay, L. C.; Procopio, J.; Oliveira, E.; Nakaie, C. R.; Schreier, S. FEBS
Lett. 2004, 565(1-3), 171–175.(44) Maltseva, E.; Brezesinski, G. ChemPhysChem 2004, 5(8), 1185–1190.
Figure 1. Hydrophobicity plots based on the Black and Mould scale35
presenting the amino acid sequence of the individual peptides (namedon each figure) using the one letter code.
3290 Langmuir, Vol. 25, No. 5, 2009 Protopapa et al.

for studying the membrane activity of peptides is that of aphospholipid monolayer/bilayer on a mercury (Hg) electrode.46
The great advantage is its inherent reproducibility, its ease ofuse, and its ability to control the potential and measure the currentvery precisely. The system has acted as a good host for thegramicidin monomolecular channel so that channel function canbe monitored using the Tl+/Tl(Hg) system as a redox probe.47
At the same time, interactions with the phospholipid monolayercan be investigated since the pure monolayer system is virtuallydefect free and self-sealing. Any modification to the structurecan be easily monitored. Electrochemical methods of impedanceare a very sensitive way to monitor the structure and propertiesof the layer, and a novel impedance model has been developedusing the phospholipid-Hg system to test it.48 An investigationof the interaction of gramicidin peptide derivatives with themonolayer was done using voltammetric and impedance tech-niques.49 Results correlated well with those of independentexperiments carried out with membrane models of monolayersat the air-water interface. A recent investigation showed thatthe interaction of the antimicrobial peptide group of CNYderivatives with the DOPC-coated Hg exactly matched thepeptides’ interaction with DOPC bilayer vesicles.50 These findingshave prompted us to use the DOPC-coated Hg system to screenthe interactions of the self-assembling -sheet peptides withphospholipid layers.
Experimental SectionPeptide Synthesis, Purification, and Quality Control. Peptides
were synthesized by NeoMPS SA, Synpep (Dublin, CA), or in-house using standard solid-phase Fmoc chemistry.14 All peptideshad acetylated N- and amide attached C-termini. Purification wascarried out by reversed-phase high performance liquid chromatog-raphy (HPLC), using a Gilson preparative HPLC system and aPhenomenex Jupiter column (15 mm C18 300A) 250 × 10 mm.Negatively charged peptides were purified using 0.1% NH3 in wateras buffer A and 10% buffer A in acetonitrile as buffer B. Positivelycharged peptides were purified using a 20-40% water-acetonitrilegradient in the presence of 0.1% trifluoroacetic acid (TFA). Peptidequality control was carried out using mass spectroscopy, HPLC,amino acid and elemental analysis, and UV spectroscopy. The peptidepurity was between 95 and 98%. All peptides were stored at -4 °Cas freeze-dried powders.
Electrochemical Measurements. Electrochemical methods fo-cused on (a) capacitative elements46,48,49 using impedance techniquesand (b) investigations of the monolayer permeability to Tl+ or Cd2+
ions in which a Faradaic process is involved.46,47 The principle ofthe Tl+ and Cd2+ transport experiments is that in the presence of theDOPC monolayer the electrochemical reduction of Tl+ or Cd2+ issuppressed, because the metal ions are denied access to the mercurysurface. When the DOPC monolayer is permeabilized to either Tl+
or Cd2+ ions, a current due to the reduction of Tl+ or Cd2+ to therespective metal amalgam is observed. The results of both theimpedance and voltammetric measurements are considered in termsof the properties of the phospholipid and modified phospholipidmonolayer.
A homemade potentiostat connected to a function generator andlock-in-amplifier interfaced to a MacLab acquisition board andsoftware(ADInstrumentsLtd.)wasusedtomeasurecapacitance-potential(Cd-E) curves of the coated electrodes. An Autolab system, frequency
response analyzer (FRA), and PGSTAT 30 interface (Ecochemie,Utrecht, Netherlands), controlled with Autolab software, were usedin all the impedance versus frequency measurements of the coatedelectrodes. A homemade potentiostat connected to a MacLabacquisition board and software was used in the sampled currentvoltammetry experiments to measure the reduction of Tl+ and Cd2+.The experiments were performed in a standard three-electrode cell.An Ag/AgCl, 3.5 mol dm-3 KCl reference electrode with a poroussintered glass frit separating the 3.5 mol dm-3 KCl solution fromthe electrolyte served as reference, and a platinum bar served ascounter electrodes located on either side of the working electrode.In the impedance-frequency measurements, a solution resistanceof around 280-300 Ω was recorded for the cell.48,49 Diagnosticplots of the impedance data showed it to be that of an RC seriescircuit as before.48,49
The electrolyte, KCl (0.1 mol dm-3), was prepared from AnalarKCl (Fisher Chemicals Ltd.) calcined at 600 °C and dissolved in18.2 MΩ MilliQ water with added 0.001 mol dm-3 phosphate buffer(pH 7.4). A blanket of argon gas was maintained above the fullydeaerated electrolyte during all experiments. Monolayers of DOPCwere prepared as described earlier46-49 by initially spreading 13 µLof a 2.54 mmol dm-3 solution of DOPC in pentane (HPLC grade,Fisher Scientific Chemicals Ltd.) at the argon-electrolyte interfacein the electrochemical cell.46-49 The working solution of DOPCwas obtained by dilution of the 25.4 mmol dm-3 stock solution(Lipid Products). A fresh mercury drop (area ) 0.0092 cm2) wascoatedwiththespreadphospholipid46-49 layerat theargon-electrolyteinterface prior to each series of experiments to give a monolayer-coated electrode. Peptides were dissolved in 18.2 MΩ MilliQ waterat about 3 mmol dm-3, sonicated until a homogeneous solution wasobtained, immediately distributed in aliquots to different vials, andkept in the freezer. One vial was taken out of the freezer before thestart of an experiment. The remainder of the solution in a vial wasdiscarded at the end of the experiment. Aliquots of the respectiveworking solutions of peptide were injected below the DOPC layerinto the electrolyte, and the solution was gently stirred for five minutes.The phospholipid monolayer was then deposited on the electrode.Such layers in this study are referred to as peptide-modified layers,regardless of the extent of interaction.
For experiments using peptide concentrations of 30 µmol dm-3
and above in the electrolyte, a different experimental protocol wasused and is described in the following. Peptide aliquots were addedto deaerated electrolyte which was then gently stirred for 10 minunder argon followed by the spreading of the phospholipid monolayeron the surface. This was done to remove traces of oxygen from theelectrolyte introduced by the larger peptide aliquot addition and toavoid any disturbance of the peptide by argon bubbling. Impedance-frequency experiments were carried out at 25 °C and all otherelectrochemical data were collected at room temperature.
Electrochemical Impedance. Measurements of capacitance (Cd)versus potential (E) for the coated electrode were carried out bymeasuring the out-of-phase current (I′′ ) at potentials between -0.2and -1.2 V in response to an applied voltage wave of frequency(f) of 75 Hz with ∆V ) 0.0046 V ac amplitude. Cd was calculatedfrom the I′′ value using the equation Cd ) I′′ /(∆Vω), where ω is theangular frequency ()2πf) of the applied waveform assuming RCseries behavior of the cell.48 The Cd-E curve of the DOPC-coatedelectrode was recorded prior to each experiment. At least two Cd-Ecurves following respective depositions on a fresh electrode surfacewere recorded for the individual peptide-phospholipid interactions.Measurements of the impedance (Z) versus frequency of the electrodesystems using frequencies logarithmically distributed from 65 000to 0.1 Hz, with ∆V) 0.005 V ac amplitude at potential-0.4 V werecarried out on the coated electrode systems. The experimentalconditions for the measurement of impedance were used exactly asdescribed previously.48 The impedance data were transformed tothe complex capacitance plane and the complex capacitance axeswere expressed as ReYω-1 and ImYω-1, where Re and Im areabbreviations for real and imaginary, respectively. This was doneusing the EXCEL (Microsoft) spreadsheet. Curve fitting of the data
(45) Kim, J. M.; Patwardhan, A.; Bott, A.; Thompson, D. H. Biochim. Biophys.Acta 2003, 1617(1-2), 10–21.
(46) Bizzotto, D.; Nelson, A. Langmuir 1998, 14(21), 6269–6273.(47) Nelson, A. Biophys. J. 2001, 80(6), 2694–2703.(48) Whitehouse, C.; O’Flanagan, R.; Lindholm-Sethson, B.; Movaghar, B.;
Nelson, A. Langmuir 2004, 20(1), 136–144.(49) Whitehouse, C.; Gidalevitz, D.; Cahuzac, M.; Koeppe, R. E.; Nelson, A.
Langmuir 2004, 20(21), 9291–9298.(50) Ringstad, L.; Protopapa, E.; Lindholm-Sethson, B.; Schmidtchen, A.;
Nelson, A.; Malmsten, M. Langmuir 2008, 24, 208–216.
-Sheet Peptide/Phospholipid Interactions Langmuir, Vol. 25, No. 5, 2009 3291

was carried out using IGOR (Wavemetrics) in the same way asdescribed previously.48,49
Electrochemistry of Tl+/Tl(Hg) and Cd2+/Cd(Hg). The fol-lowing procedure46,47 was used to measure the peptide permeabilizingactivity to Tl+ or Cd2+ of the monolayer. TlNO3 (Sigma Products)and Cd(NO3)2 (BDH Chemicals Ltd.) were employed to prepare thestock solutions (0.1 mol dm-3). Subsequent to deaeration of theelectrolyte, 10-4 mol dm-3 Tl(I) or Cd(II) was added from the stocksolutions. The DOPC layer was then spread on the electrolyte andtransferred to the electrode. A cyclic voltammogram was recordedto check the impermeability of the deposited layer. Following additionof peptide to the electrolyte and stirring, the phospholipid layer wasdeposited on the electrode surface. A series of voltage pulses from-0.2 V to potentials from -0.3 to -0.7 V and back for Tl(I) andfrom -0.3 V to potentials from -0.4 to -0.8 V and back for Cd(II)were initiated and the current transients recorded. The pulses were40 ms long and the currents were sampled at 40 kHz with a 20 kHzlow-pass filter. A delay period of 15 s between each pulse enabledthe establishment of initial concentration conditions. After the pulsingprogram had been performed, an ac out-of-phase voltammogramwas recorded to ensure that the phospholipid layer had not degradedduring the experiment. Values of current were sampled after a timeinterval of 2.5 ms from the beginning of the pulse and plotted againstpotential as a sampled-current voltammogram. The currents weremeasured for the cathodic train of pulses and the anodic train ofpulses. The mean value of current and the range between the twovalues expressed as an error bar were recorded at each potential.
High-Resolution 1H NMR Study of Peptide Self-Assembly.The self-assembly or aggregation state of the peptides in solutionwas examined using 1H NMR spectroscopy. This method has beenpreviously shown to measure the concentration of monomeric peptidepresent in solution.28,30 Solutions of peptides P11-2(0), P11-4(–2),and P11-8(+2) were prepared directly at concentrations between 0and 100 µmol dm-3 in 0.13 mol dm-3 NaCl (Fisher ScientificChemicals Ltd.) in D2O (99.9% D, Fluorochem Ltd.). An NMRinternal reference standard was added to all peptide solutions. ForP11-2(0) and P11-4(–2), solutions contained 0.125 mmol dm-3 (2,2,3,3-d4)-trimethylsilyl-3-propionic acid (TMSP, Cambridge IsotopeLaboratories, chemical shift 0 ppm). However, because of P11-8(+2)’snet positive charge, tetramethylammonium chloride (TMAC,g99%,Sigma-Aldrich, chemical shift 3.18 ppm) was added to P11-8(+2)solutions, at a concentration of 0.125 mmol dm-3. Samples weresonicated in an ultrasonic water bath for 30 min. Solution pD wasmeasured and adjusted to 7.4 ( 0.18 and the relation pD ) pH +0.451 was used. Any adjustment of pD, if necessary, was made withmicroliter volumes of 0.1 mol dm-3 DCl (35% weight DCl, Sigma-Aldrich, diluted with D2O) and/or 0.1 mol dm-3 NaOD (40% weightNaOD, diluted with D2O). Finally, solution vials were closed, sealedwith PTFE tape, and then gently warmed to maximize peptidesolubility. Solutions were left to equilibrate at room temperature.
NMR data were acquired at room temperature with a Bruker DPX300 spectrometer, operating at 300 MHz (5 mm probe, spectralwidth 5995 Hz), controlled by XwinNMR software. A presaturationprogram (Bruker zgpr program) was used to minimize the waterpeak, and 1024 scans were measured per spectrum. A spectrum foreach sample was recorded twice after preparation, at 3-5 and 11-15days, to ensure that the solutions had reached stable equilibrium.MestRe-C 2.3a (Organic Chemistry Department, Santiago deCompostela University) was used to Fourier transform and measureNMR peak integrals. The aromatic multiplet at chemical shift∼7.3-7.5 ppm was integrated, relative to the reference peak. Origin7 (OriginLab Corp.) was used to process and plot data.
Impedance Model. In the absence of any electroactive component,the simplest equivalent circuit model is the uncompensated solutionresistance (Ru) of the cell and the capacitance (C) of the workingelectrode in series.52 Ru can be determined by extrapolating the ImZ
versus ReZ plot to the ReZ axis.53 In the complex capacitance plane,values of ReYω-1 were plotted against ImYω-1 for all values offrequency.53-55 For a series RC circuit, the ReYω-1 versus ImYω-1
plots gives a single semicircle for the RC element, where the capacitorhas no frequency dispersion. The extrapolation of this semicircle tothe ImYω-1 axis at intermediate frequency gives the zero frequencycapacitance (‘ZFC’ ) C) of the RC circuit.54 When applied to thephospholipid-coated electrode, any additional elements to the RCsemicircle at lower frequencies will correspond to properties of thephospholipid layer. Further, if the semicircle representing the RCelement is not perfect,56 the nonideality of the capacitor is indicated.This can be due to dielectric relaxations coupled to the RC chargingprocess and to additional circuit elements at the interface betweenthe capacitor and the solution resistance.57 All the impedance datawere fitted to eq 1 below as done previously:31,48,49,58,59
Y) 1
R+ 1
(iω)ω01-[ Cs -C
1+ (iωτ)R+C]
(1)
In eq 1, Y is the admittance; R is equivalent to the uncompensatedsolution resistance (Ru); C is equivalent to the zero frequencycapacitance (‘ZFC’) of the monolayer, which represents themonolayer capacitance in the absence of an extra capacitative elementrecorded at zero frequency; (Cs -C) is the additional low-frequencycapacitative element with relaxation time constant, τ; R is thecoefficient that represents the distribution of time constants aroundτ; and is the coefficient that relates to nonidealities at the interfacebetween R and C and characterizes a surface “roughness”.56
Generally, approaches unity for a DOPC monolayer or Hg surfaceand decreases below unity when peptides or similar compoundsadsorb on to or penetrate into the DOPC monolayer and roughenthe surface.48,49,59 ω 0 is a dummy constant that is always set atunity. C, Cs, R, and τ characterize the bulk dielectric, in this casethe monolayer, whereas relates to the interface between themonolayer and the solution. Equation 1 shows that the additionallow-frequency element (Cs - C) is significant at low frequenciesbut becomes less significant at intermediate frequencies, where eq1 is dominated by the capacitance, C.
All the experimental data derived from the monolayer on theelectrode were fitted to eq 1, and the fitting procedure was carriedout as follows. The experimental Ru, which is equivalent to R in eq1 was inputted as a fixed parameter. The five variable parameters:C, Cs, R, , and τ were extracted from the subsequent curve fittingof the real component of eq 1 to the ReYω-1 vs log ω plot. This wasthe reported fit and provided a value and SD for each parameter. Theoverall goodness of fit was examined by comparing the experimentaland model, ReYω-1 and ImYω-1 vs log ω plots. Values of parameterswith their errors for two representative interactions are displayed inTable 1.
Results and Discussion
The effects of the peptide-DOPC interactions on thecapacitance (Cd) versus potential (E) plots are displayed in Figure2. Addition of amphiphilic peptide P11-8(+2) and polar peptideP11-19(+2) to electrolyte alters the Cd vs -E profile of DOPCon the cathodic scan. A significant alteration of the Cd vs -E
(51) Glasoe, P. K.; Long, F. A. J. Phys. Chem. 1960, 64(1), 188–190.(52) Wiegand, G.; Arribas-Layton, N.; Hillebrandt, H.; Sackmann, E.; Wagner,
P. J. Phys. Chem. 2002, 106(16), 4245–4254.
(53) Janek, R. P.; Fawcett, W. R.; Ulman, A. J. Phys. Chem. B 1997, 101(42),8550–8558.
(54) Lindholm-Sethson, B. Langmuir 1996, 12(13), 3305–3314.(55) Strasak, L.; Dvorak, J.; Hason, S.; Vetterl, V. Bioelectrochemistry 2002,
56(1-2), 37–41.(56) Peng Diao, P.; Jiang, D. L.; Cui, X. L.; Gu, D. P.; Tong, R. T.; Zhong,
B. J. Electroanal. Chem. 1999, 464(1), 61–67.(57) Lingler, S.; Rubinstein, I.; Knoll, W.; Offenhausser, A. Langmuir 1997,
13(26), 7085–7091.(58) Merrifield, J.; Tattersall, J.; Bird, M.; Nelson, A. Electroanalysis 2007,
19(2-3), 272–279.(59) Weiss, S.; Millner, P.; Nelson, A. Electrochem. Acta 2005, 50(21), 4248–
4256.
3292 Langmuir, Vol. 25, No. 5, 2009 Protopapa et al.
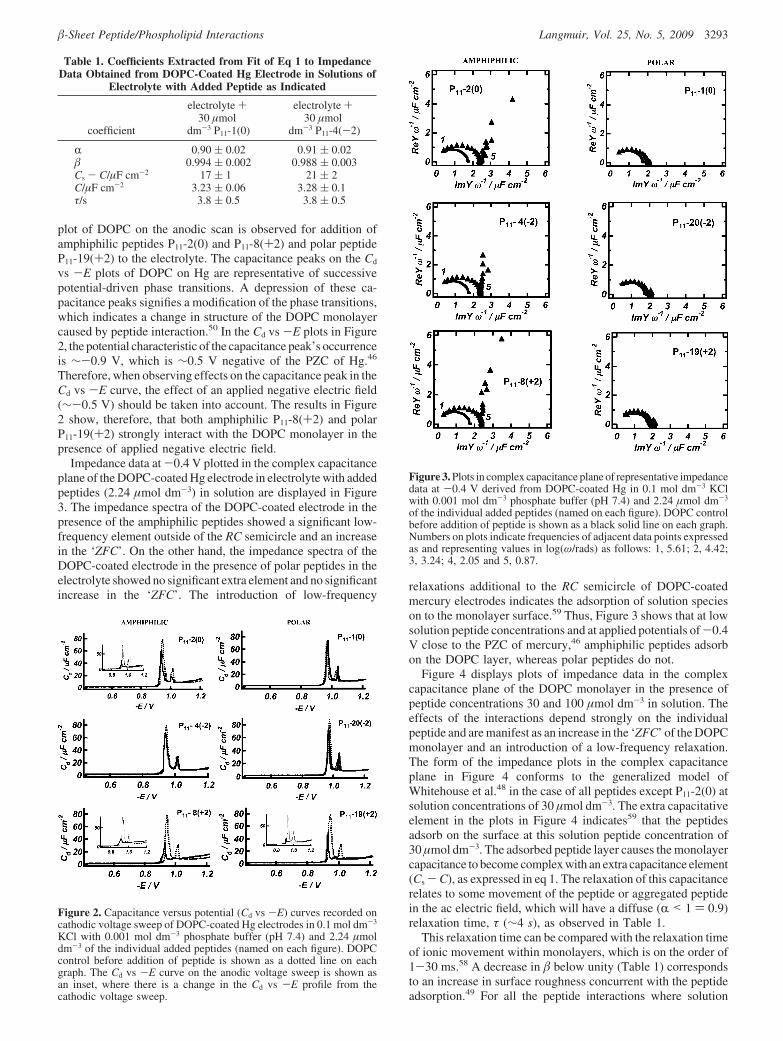
plot of DOPC on the anodic scan is observed for addition ofamphiphilic peptides P11-2(0) and P11-8(+2) and polar peptideP11-19(+2) to the electrolyte. The capacitance peaks on the Cd
vs -E plots of DOPC on Hg are representative of successivepotential-driven phase transitions. A depression of these ca-pacitance peaks signifies a modification of the phase transitions,which indicates a change in structure of the DOPC monolayercaused by peptide interaction.50 In the Cd vs -E plots in Figure2, the potential characteristic of the capacitance peak’s occurrenceis ∼-0.9 V, which is ∼0.5 V negative of the PZC of Hg.46
Therefore, when observing effects on the capacitance peak in theCd vs -E curve, the effect of an applied negative electric field(∼-0.5 V) should be taken into account. The results in Figure2 show, therefore, that both amphiphilic P11-8(+2) and polarP11-19(+2) strongly interact with the DOPC monolayer in thepresence of applied negative electric field.
Impedance data at -0.4 V plotted in the complex capacitanceplane of the DOPC-coated Hg electrode in electrolyte with addedpeptides (2.24 µmol dm-3) in solution are displayed in Figure3. The impedance spectra of the DOPC-coated electrode in thepresence of the amphiphilic peptides showed a significant low-frequency element outside of the RC semicircle and an increasein the ‘ZFC’. On the other hand, the impedance spectra of theDOPC-coated electrode in the presence of polar peptides in theelectrolyte showed no significant extra element and no significantincrease in the ‘ZFC’. The introduction of low-frequency
relaxations additional to the RC semicircle of DOPC-coatedmercury electrodes indicates the adsorption of solution specieson to the monolayer surface.59 Thus, Figure 3 shows that at lowsolution peptide concentrations and at applied potentials of-0.4V close to the PZC of mercury,46 amphiphilic peptides adsorbon the DOPC layer, whereas polar peptides do not.
Figure 4 displays plots of impedance data in the complexcapacitance plane of the DOPC monolayer in the presence ofpeptide concentrations 30 and 100 µmol dm-3 in solution. Theeffects of the interactions depend strongly on the individualpeptide and are manifest as an increase in the ‘ZFC’ of the DOPCmonolayer and an introduction of a low-frequency relaxation.The form of the impedance plots in the complex capacitanceplane in Figure 4 conforms to the generalized model ofWhitehouse et al.48 in the case of all peptides except P11-2(0) atsolution concentrations of 30 µmol dm-3. The extra capacitativeelement in the plots in Figure 4 indicates59 that the peptidesadsorb on the surface at this solution peptide concentration of30 µmol dm-3. The adsorbed peptide layer causes the monolayercapacitance to become complex with an extra capacitance element(Cs -C), as expressed in eq 1. The relaxation of this capacitancerelates to some movement of the peptide or aggregated peptidein the ac electric field, which will have a diffuse (R < 1 ) 0.9)relaxation time, τ (∼4 s), as observed in Table 1.
This relaxation time can be compared with the relaxation timeof ionic movement within monolayers, which is on the order of1-30 ms.58 A decrease in below unity (Table 1) correspondsto an increase in surface roughness concurrent with the peptideadsorption.49 For all the peptide interactions where solution
Table 1. Coefficients Extracted from Fit of Eq 1 to ImpedanceData Obtained from DOPC-Coated Hg Electrode in Solutions of
Electrolyte with Added Peptide as Indicated
coefficient
electrolyte +30 µmol
dm-3 P11-1(0)
electrolyte +30 µmol
dm-3 P11-4(-2)
R 0.90 ( 0.02 0.91 ( 0.02 0.994 ( 0.002 0.988 ( 0.003Cs - C/µF cm-2 17 ( 1 21 ( 2C/µF cm-2 3.23 ( 0.06 3.28 ( 0.1τ/s 3.8 ( 0.5 3.8 ( 0.5
Figure 2. Capacitance versus potential (Cd vs -E) curves recorded oncathodic voltage sweep of DOPC-coated Hg electrodes in 0.1 mol dm-3
KCl with 0.001 mol dm-3 phosphate buffer (pH 7.4) and 2.24 µmoldm-3 of the individual added peptides (named on each figure). DOPCcontrol before addition of peptide is shown as a dotted line on eachgraph. The Cd vs -E curve on the anodic voltage sweep is shown asan inset, where there is a change in the Cd vs -E profile from thecathodic voltage sweep.
Figure 3. Plots in complex capacitance plane of representative impedancedata at -0.4 V derived from DOPC-coated Hg in 0.1 mol dm-3 KClwith 0.001 mol dm-3 phosphate buffer (pH 7.4) and 2.24 µmol dm-3
of the individual added peptides (named on each figure). DOPC controlbefore addition of peptide is shown as a black solid line on each graph.Numbers on plots indicate frequencies of adjacent data points expressedas and representing values in log(ω/rads) as follows: 1, 5.61; 2, 4.42;3, 3.24; 4, 2.05 and 5, 0.87.
-Sheet Peptide/Phospholipid Interactions Langmuir, Vol. 25, No. 5, 2009 3293
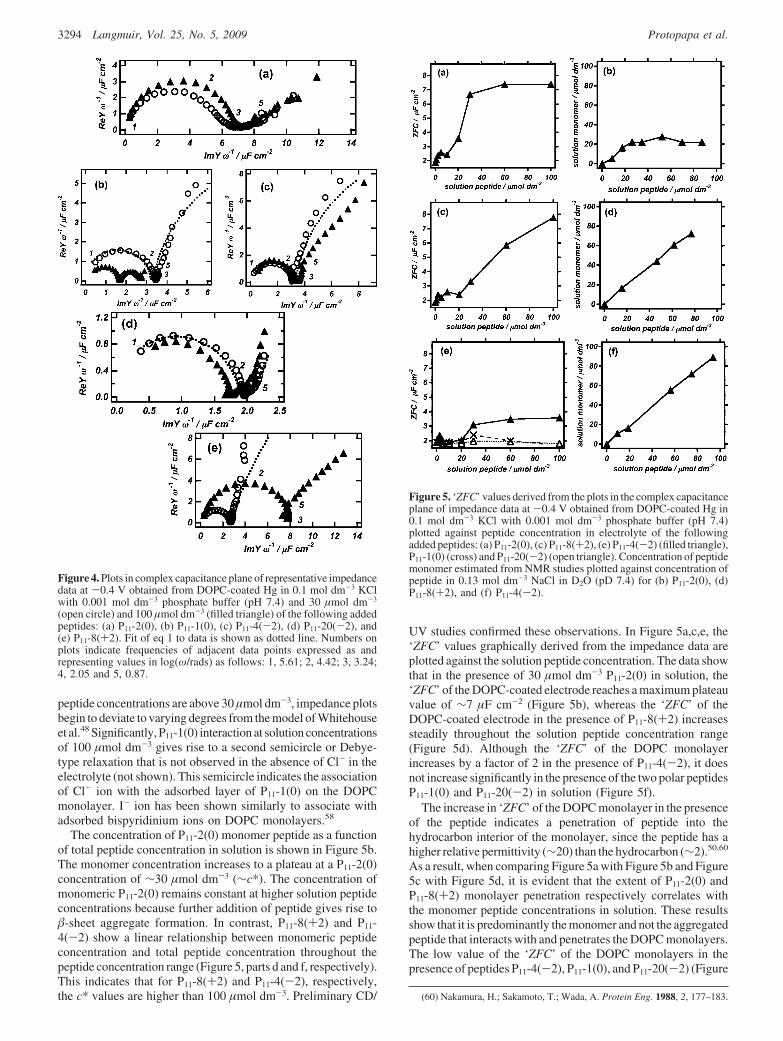
peptide concentrations are above 30 µmol dm-3, impedance plotsbegin to deviate to varying degrees from the model of Whitehouseet al.48 Significantly, P11-1(0) interaction at solution concentrationsof 100 µmol dm-3 gives rise to a second semicircle or Debye-type relaxation that is not observed in the absence of Cl- in theelectrolyte (not shown). This semicircle indicates the associationof Cl- ion with the adsorbed layer of P11-1(0) on the DOPCmonolayer. I- ion has been shown similarly to associate withadsorbed bispyridinium ions on DOPC monolayers.58
The concentration of P11-2(0) monomer peptide as a functionof total peptide concentration in solution is shown in Figure 5b.The monomer concentration increases to a plateau at a P11-2(0)concentration of ∼30 µmol dm-3 (∼c*). The concentration ofmonomeric P11-2(0) remains constant at higher solution peptideconcentrations because further addition of peptide gives rise to-sheet aggregate formation. In contrast, P11-8(+2) and P11-4(-2) show a linear relationship between monomeric peptideconcentration and total peptide concentration throughout thepeptide concentration range (Figure 5, parts d and f, respectively).This indicates that for P11-8(+2) and P11-4(-2), respectively,the c* values are higher than 100 µmol dm-3. Preliminary CD/
UV studies confirmed these observations. In Figure 5a,c,e, the‘ZFC’ values graphically derived from the impedance data areplotted against the solution peptide concentration. The data showthat in the presence of 30 µmol dm-3 P11-2(0) in solution, the‘ZFC’ of the DOPC-coated electrode reaches a maximum plateauvalue of ∼7 µF cm-2 (Figure 5b), whereas the ‘ZFC’ of theDOPC-coated electrode in the presence of P11-8(+2) increasessteadily throughout the solution peptide concentration range(Figure 5d). Although the ‘ZFC’ of the DOPC monolayerincreases by a factor of 2 in the presence of P11-4(-2), it doesnot increase significantly in the presence of the two polar peptidesP11-1(0) and P11-20(-2) in solution (Figure 5f).
The increase in ‘ZFC’ of the DOPC monolayer in the presenceof the peptide indicates a penetration of peptide into thehydrocarbon interior of the monolayer, since the peptide has ahigher relative permittivity (∼20) than the hydrocarbon (∼2).50,60
As a result, when comparing Figure 5a with Figure 5b and Figure5c with Figure 5d, it is evident that the extent of P11-2(0) andP11-8(+2) monolayer penetration respectively correlates withthe monomer peptide concentrations in solution. These resultsshow that it is predominantly the monomer and not the aggregatedpeptide that interacts with and penetrates the DOPC monolayers.The low value of the ‘ZFC’ of the DOPC monolayers in thepresence of peptides P11-4(-2), P11-1(0), and P11-20(-2) (Figure
(60) Nakamura, H.; Sakamoto, T.; Wada, A. Protein Eng. 1988, 2, 177–183.
Figure 4. Plots in complex capacitance plane of representative impedancedata at -0.4 V obtained from DOPC-coated Hg in 0.1 mol dm-3 KClwith 0.001 mol dm-3 phosphate buffer (pH 7.4) and 30 µmol dm-3
(open circle) and 100 µmol dm-3 (filled triangle) of the following addedpeptides: (a) P11-2(0), (b) P11-1(0), (c) P11-4(-2), (d) P11-20(-2), and(e) P11-8(+2). Fit of eq 1 to data is shown as dotted line. Numbers onplots indicate frequencies of adjacent data points expressed as andrepresenting values in log(ω/rads) as follows: 1, 5.61; 2, 4.42; 3, 3.24;4, 2.05 and 5, 0.87.
Figure 5. ‘ZFC’ values derived from the plots in the complex capacitanceplane of impedance data at -0.4 V obtained from DOPC-coated Hg in0.1 mol dm-3 KCl with 0.001 mol dm-3 phosphate buffer (pH 7.4)plotted against peptide concentration in electrolyte of the followingadded peptides: (a) P11-2(0), (c) P11-8(+2), (e) P11-4(-2) (filled triangle),P11-1(0) (cross) and P11-20(-2) (open triangle). Concentration of peptidemonomer estimated from NMR studies plotted against concentration ofpeptide in 0.13 mol dm-3 NaCl in D2O (pD 7.4) for (b) P11-2(0), (d)P11-8(+2), and (f) P11-4(-2).
3294 Langmuir, Vol. 25, No. 5, 2009 Protopapa et al.
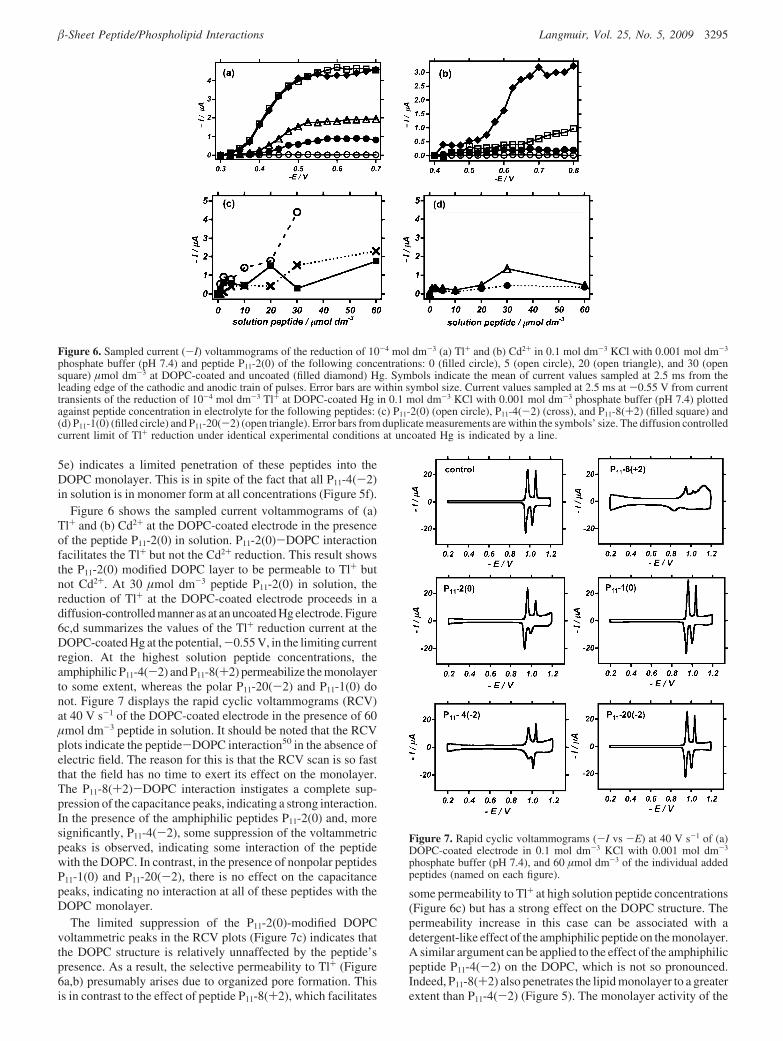
5e) indicates a limited penetration of these peptides into theDOPC monolayer. This is in spite of the fact that all P11-4(-2)in solution is in monomer form at all concentrations (Figure 5f).
Figure 6 shows the sampled current voltammograms of (a)Tl+ and (b) Cd2+ at the DOPC-coated electrode in the presenceof the peptide P11-2(0) in solution. P11-2(0)-DOPC interactionfacilitates the Tl+ but not the Cd2+ reduction. This result showsthe P11-2(0) modified DOPC layer to be permeable to Tl+ butnot Cd2+. At 30 µmol dm-3 peptide P11-2(0) in solution, thereduction of Tl+ at the DOPC-coated electrode proceeds in adiffusion-controlled manner as at an uncoated Hg electrode. Figure6c,d summarizes the values of the Tl+ reduction current at theDOPC-coated Hg at the potential,-0.55 V, in the limiting currentregion. At the highest solution peptide concentrations, theamphiphilic P11-4(-2) and P11-8(+2) permeabilize the monolayerto some extent, whereas the polar P11-20(-2) and P11-1(0) donot. Figure 7 displays the rapid cyclic voltammograms (RCV)at 40 V s-1 of the DOPC-coated electrode in the presence of 60µmol dm-3 peptide in solution. It should be noted that the RCVplots indicate the peptide-DOPC interaction50 in the absence ofelectric field. The reason for this is that the RCV scan is so fastthat the field has no time to exert its effect on the monolayer.The P11-8(+2)-DOPC interaction instigates a complete sup-pression of the capacitance peaks, indicating a strong interaction.In the presence of the amphiphilic peptides P11-2(0) and, moresignificantly, P11-4(-2), some suppression of the voltammetricpeaks is observed, indicating some interaction of the peptidewith the DOPC. In contrast, in the presence of nonpolar peptidesP11-1(0) and P11-20(-2), there is no effect on the capacitancepeaks, indicating no interaction at all of these peptides with theDOPC monolayer.
The limited suppression of the P11-2(0)-modified DOPCvoltammetric peaks in the RCV plots (Figure 7c) indicates thatthe DOPC structure is relatively unnaffected by the peptide’spresence. As a result, the selective permeability to Tl+ (Figure6a,b) presumably arises due to organized pore formation. Thisis in contrast to the effect of peptide P11-8(+2), which facilitates
some permeability to Tl+ at high solution peptide concentrations(Figure 6c) but has a strong effect on the DOPC structure. Thepermeability increase in this case can be associated with adetergent-like effect of the amphiphilic peptide on the monolayer.A similar argument can be applied to the effect of the amphiphilicpeptide P11-4(-2) on the DOPC, which is not so pronounced.Indeed, P11-8(+2) also penetrates the lipid monolayer to a greaterextent than P11-4(-2) (Figure 5). The monolayer activity of the
Figure 6. Sampled current (-I) voltammograms of the reduction of 10-4 mol dm-3 (a) Tl+ and (b) Cd2+ in 0.1 mol dm-3 KCl with 0.001 mol dm-3
phosphate buffer (pH 7.4) and peptide P11-2(0) of the following concentrations: 0 (filled circle), 5 (open circle), 20 (open triangle), and 30 (opensquare) µmol dm-3 at DOPC-coated and uncoated (filled diamond) Hg. Symbols indicate the mean of current values sampled at 2.5 ms from theleading edge of the cathodic and anodic train of pulses. Error bars are within symbol size. Current values sampled at 2.5 ms at -0.55 V from currenttransients of the reduction of 10-4 mol dm-3 Tl+ at DOPC-coated Hg in 0.1 mol dm-3 KCl with 0.001 mol dm-3 phosphate buffer (pH 7.4) plottedagainst peptide concentration in electrolyte for the following peptides: (c) P11-2(0) (open circle), P11-4(-2) (cross), and P11-8(+2) (filled square) and(d) P11-1(0) (filled circle) and P11-20(-2) (open triangle). Error bars from duplicate measurements are within the symbols’ size. The diffusion controlledcurrent limit of Tl+ reduction under identical experimental conditions at uncoated Hg is indicated by a line.
Figure 7. Rapid cyclic voltammograms (-I vs -E) at 40 V s-1 of (a)DOPC-coated electrode in 0.1 mol dm-3 KCl with 0.001 mol dm-3
phosphate buffer (pH 7.4), and 60 µmol dm-3 of the individual addedpeptides (named on each figure).
-Sheet Peptide/Phospholipid Interactions Langmuir, Vol. 25, No. 5, 2009 3295

positively charged amphiphilic P11-8(+2) compared to itsnegatively charged analogue P11-4(-2) is commensurate withthe noted activity of positively charged amphiphilic antimicrobialssuch as magainin 2,61 melittin,62 LL37,63,64 defensin,65 tachyplesinI,66 and synthetic cyclic peptides.67 The increased interaction ofthe amphiphilic P11-8(+2) compared to the amphiphilic P11-4(-2) can be explained as due to the positively charged peptidebeing attracted by the negative dipole potential on the solutionside of the DOPC layer. The dipole potential of a DOPCmonolayer on Hg, which is due to the arrangement of the dipolesof the lipid groups and of water at the monolayer interface, hasbeen measured to be +145 mV toward the hydrocarbon interiorat 25 °C.68 A similar dipole potential also exists across oneleaflet of a PC phospholipid bilayer.69
The results of this paper show that, in the absence of appliedfield, the amphiphilic peptides (P11-2(0), P11-4(-2), and P11-8(+2)) have a greater tendency to interact with the DOPCmonolayer than their polar analogues (P11-1(0), P11-20(-2), andP11-19(+2)). The amphiphilic peptides possess a hydrophobicside consisting of three aromatic residues composed of phenyl-alanine, tryptophan, and phenylalanine, respectively (see Figure1). Aromatic residues, particularly tryptophan and tyrosine, arewell-known to interact specifically with phospholipid headgroups.Tryptophan associates with the carbonyl region of the phos-pholipid head groups and efficiently anchors peptides or proteins
at the lipid-water interface.70-72 This provides a possiblemechanism for the initial binding of amphiphilic peptides to theDOPC monolayer. Also evident in this study is the relatively lowactivity toward phospholipid monolayers of the -sheet self-assembling peptides compared to the antimicrobial peptides.Peptide P11-2(0) fully permeabilized the phospholipid monolayerto Tl+ at a solution concentration of 30 µmol dm-3 gramicidin.A fully permeabilized the phospholipid monolayer to Tl+ atsolution concentrations 3 orders of magnitude lower47 and theCNY variants50 at solution concentrations of about 1 µmol dm-3.
ConclusionsIn the context of the interaction of self-assembling -sheet
11-residue peptides with DOPC monolayers on Hg, the followingpoints can be made:
1 The solution monomer peptide species is the monolayeractive moiety.
2 Amphiphilic peptides are more monolayer active than polarpeptides in the absence of applied electric field.
3 The positive charge on amphiphilic peptides facilitatesmonolayer interaction in the absence of applied electricfield.
4 Negative applied electric field facilitates monolayer interac-tion with positively charged amphiphilic and polar peptides.
5 Neutral amphiphilic peptides permeabilize the DOPC layersto ions to the greatest extent.
6 The -sheet tape forming peptides are shown to besignificantly less monolayer disruptive than antimicrobialpeptides.
Acknowledgment. Funding for this work was provided bythe EU FP 6 SSA “BBMO” and also the School of Chemistry,Leeds. A.A. gratefully acknowledges the financial support of theRoyal Society through a personal Royal Society UniversityResearch Fellowship.
LA803368R
(61) Matsuzaki, K.; Nakamura, A.; Murase, O.; Sugishita, K.; Fujii, N.;Miyajima, K. Biochemistry 1997, 36(8), 2104–2111.
(62) Hristova, K.; Dempsey, C. E.; White, S. H. Biophys. J. 2001, 80(2),801–811.
(63) Henzler-Wildman, K. A.; Martinez, G. V.; Brown, M. F.; Ramamoorthy,A. Biochemistry 2004, 43(26), 8459–8469.
(64) Neville, F.; Gidalevitz, D.; Kale, G.; Nelson, A. Bioelectrochemistry 2007,70(2), 205–213.
(65) Kagan, B. L.; Selsted, M. E.; Ganz, T.; Lehrer, R. I. Proc. Natl. Acad.Sci. U.S.A. 1990, 87(1), 210–214.
(66) Matsuzaki, K.; Yoneyama, S.; Fujii, N.; Miyajima, K.; Yamada, K.; Kirino,Y.; Anzai, K. Biochemistry 1997, 36(32), 9799–9806.
(67) Fernandez-Lopez, S.; Kim, H. S.; Choi, E. C.; Delgado, M.; Granja, J. R.;Khasanov, A.; Kraehenbuehl, K.; Long, G.; Weinberger, D. A.; Wilcoxen, K. M.;Ghadiri, M. R. Nature 2001, 412(6845), 452–455.
(68) Becucci, L.; Moncelli, M. R.; Herrero, R.; Guidelli, R. Langmuir 2000,16(20), 7694–7700.
(69) Clarke, R. J. AdV. Colloid Interface Sci. 2001, 89, 263–281.
(70) de Planque, M. R. R.; Killian, J. A. Mol. Membr. Biol. 2003, 20(4),271–284.
(71) de Planque, M. R. R.; Kruijtzer, J. A. W.; Liskamp, R. M. J.; Marsh, D.;Greathouse, D. V.; Koeppe, R. E.; de Kruijff, B.; Killian, J. A. J. Biol. Chem.1999, 274(30), 20839–20846.
(72) Killian, J. A.; von Heijne, G. Trends Biochem. Sci. 2000, 25(9), 429–434.
3296 Langmuir, Vol. 25, No. 5, 2009 Protopapa et al.




![Assessing Cellular Response to Functionalized α-Helical ...two-component peptide system for making hydrogels, termed hSAFs (hydrogelating self-assembling fi bers). [ 32 ] The peptides](https://static.fdocument.org/doc/165x107/60df4feff816521c5855918c/assessing-cellular-response-to-functionalized-helical-two-component-peptide.jpg)













