
p38α MAPK is required for tooth morphogenesis and enamel secretion

www.elsevier.com/locate/jbiosc
Journal of Bioscience and BioengineeringVOL. 117 No. 6, 664e669, 2014
Inhibition of Wnt/b-catenin pathway by Dikkopf-1 affects midfacialmorphogenesis in chick embryo
Masayoshi Kawakami,1,* Hiroaki Okuda,2 Kouko Tatsumi,2 Tadaaki Kirita,1 and Akio Wanaka2
Department of Oral and Maxillofacial Surgery, Nara Medical University, 840 Shijo-cho, Kashihara, Nara 634-8522, Japan1 and Department of Anatomy and Neurosciences,Nara Medical University, 840 Shijo-cho, Kashihara, Nara 634-8522, Japan2
Received 16 July 2013; accepted 25 November 2013Available online 28 December 2013
* CorrespondE-mail ad
naramed-u.ac.naramed-u.ac.
1389-1723/$http://dx.doi
The development of the vertebrate face is regulated by complex interactions among several signaling pathways.Dickkopf-1 (Dkk-1), an inhibitor of the Wnt/b-catenin signaling pathway, can affect midfacial morphogenesis. Thedownstream target genes of the Wnt/b-catenin signaling pathway in morphogenesis of the developing upper jaw and lipremain unknown. To investigate the functional roles of Wnt/b-catenin signaling in facial development, we performed aloss-of-function experiment using local implantation of beads soaked with Dkk-1 during lip fusion at the maxillaryprominence of chick embryos at stage 22(HH22). Antagonism of Wnt/b-catenin signaling by Dkk-1 induced deformitiesof the premaxilla and jugal bone, which are derived from the maxillary mesenchyme. Real-time and semi-quantitativeRT-PCR analysis showed the significant reduction of Lhx8, Msx1 and Msx2 expression levels around the beads in themaxillary mesenchyme at 6 and 24 h after bead implantation. Time course experiments in the HH 22 embryos showedthe effect of Dkk-1 on Lhx8,Msx1 andMsx2 expression was not significant after 48 h of the treatment. At HH 26 when thefusion of facial primordial started, Dkk-1 application did not exhibit any significant reduction of those genes. Ourfindings suggested that Dkk-1 regulates maxillary morphogenesis in chick embryos through Lhx8, Msx1 and Msx2signals. Wnt/b-catenin signaling is responsible for intrinsic upper jaw development before the lip fusion.
� 2013, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Dickkopf-1 (Dkk-1); Lhx8; Wnt/b-catenin signaling; Chick embryo; Craniofacial development]
Morphogenesis of the vertebrate face is a result of theoutgrowth of several facial primordia (1). Cleft lip usually resultsdirectly from failure of fusion of medial nasal prominences with thelateral and maxillary prominences. Production of specific geneproducts or other cellular or regional characteristics can impactselective vulnerability. Several major morphogenetic signalingpathways are involved in dynamic facial morphogenesis. Theb-catenin-dependent or canonical Wnt signaling (Wnt/b-cateninsignaling) pathway is a prominent intercellular signaling pathwayin animals and plays fundamental roles in cellular proliferation,regeneration, differentiation, and function. Coordinated activationand inhibition of Wnt/b-catenin signaling are essential for properantero-posterior formation in the vertebrate embryo, and recentadvances have revealed roles for theWnt/b-catenin signaling in thedevelopment of oral tissues (2,3).
TheWnt/b-catenin signaling pathway functions by regulating theamount of the transcriptional co-activator b-catenin. Wnt ligandsbind to a Frizzled receptor and its co-receptors, low-density lipo-protein receptor-related proteins (LRP5, LRP6). These events lead tothe stabilization of b-catenin, which activates Wnt target geneexpression (4e6). These activities are inhibited by Dickkopf-1 (Dkk-1), a secreted protein that specifically inhibits the Wnt/b-catenin
ing author. Tel.: þ81 744 22 3051x2327; fax: þ81 744 29 8876.dresses: [email protected] (M. Kawakami), okuda@jp (H. Okuda), [email protected] (K. Tatsumi), tkirita@jp (T. Kirita), [email protected] (A. Wanaka).
e see front matter � 2013, The Society for Biotechnology, Japan..org/10.1016/j.jbiosc.2013.11.015
signaling pathwayby interactingwith LRP5/6 co-receptors (7,8). Dkk-1wasoriginally identified asa stronghead inducer inXenopus, due toits potent anti-Wnt effect (9). Dkk-1 knockout mice display severedevelopmental phenotypes including head defects (10), suggestingthat proper head formation in mouse requires an optimal level ofWnt/b-catenin signaling activity (11). Conditional knockout of b-catenin inmouse showed that the ablationofWnt/b-catenin signalingin theoral epitheliumblocks the formationof palatal rugae,which areassociated with antero-posterior extension of the hard palate andprecocious mid-line fusion (2). Studies with Lrp6-deficient micesuggest that lip formation and fusion requires Wnt signaling toregulate the expression of Msx1 and Msx2, members of the Msx ho-meobox gene family that have been implicated in human cleft lip andpalate (12,13). Although these lines of evidence suggest that theWnt/b-catenin signaling pathway and its downstream target genescontribute to morphogenesis of the upper jaw, it is unlikely thatdecreased gene function can explain all of the mechanisms under-lying cleft lip and palate.
Lhx8 belongs to the LIM-homeodomain (LIM-HD) transcriptionfactor family, and its expression overlaps with that of Msx1 andMsx2. Lhx7 (an alias of Lhx8) is expressed in the mesenchyme of thefirst pharyngeal arch and its derivatives, the maxilla and mandible,but not in the epithelium (14e16). Disruption of Lhx8 gene functionin mice causes impairments in palatal shelf contact and fusion thatlead to the formation of a cleft secondary palate (17). Msx1 andMsx2 are expressed in the maxillary and median nasal prominence(18), and act in a variety of cell types to control cell proliferation and
All rights reserved.
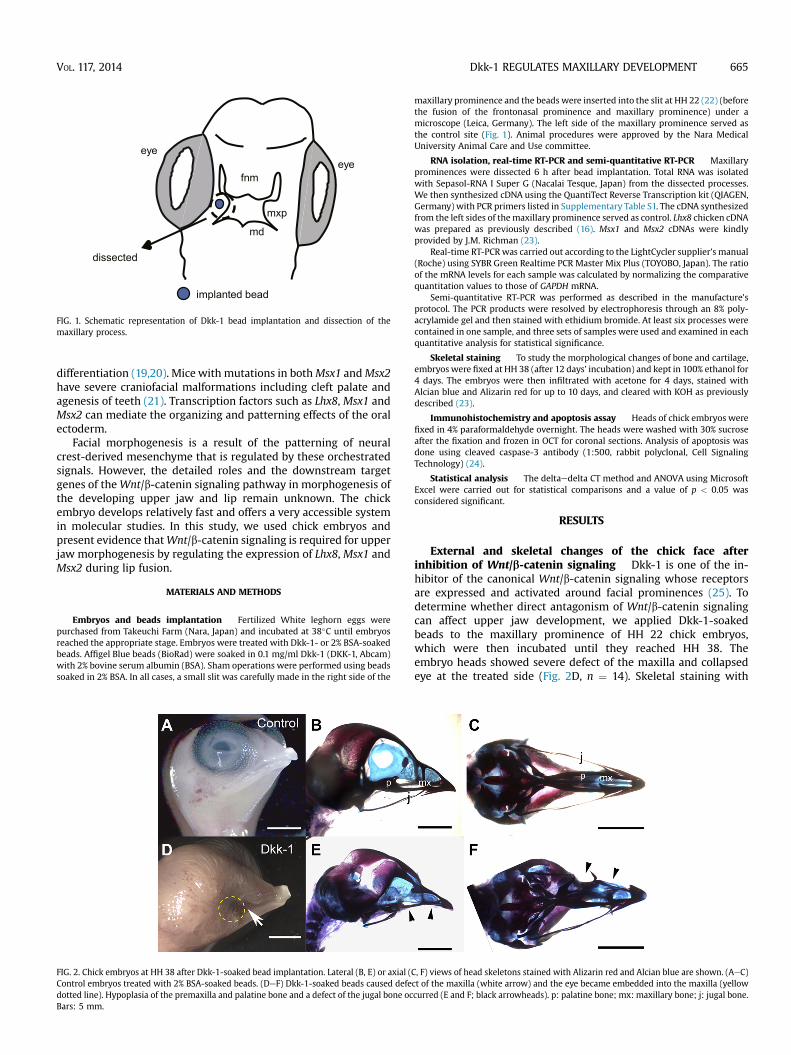
eye
fnm
md
mxp
eye
implanted bead
dissected
FIG. 1. Schematic representation of Dkk-1 bead implantation and dissection of themaxillary process.
VOL. 117, 2014 Dkk-1 REGULATES MAXILLARY DEVELOPMENT 665
differentiation (19,20). Mice with mutations in bothMsx1 andMsx2have severe craniofacial malformations including cleft palate andagenesis of teeth (21). Transcription factors such as Lhx8, Msx1 andMsx2 can mediate the organizing and patterning effects of the oralectoderm.
Facial morphogenesis is a result of the patterning of neuralcrest-derived mesenchyme that is regulated by these orchestratedsignals. However, the detailed roles and the downstream targetgenes of theWnt/b-catenin signaling pathway in morphogenesis ofthe developing upper jaw and lip remain unknown. The chickembryo develops relatively fast and offers a very accessible systemin molecular studies. In this study, we used chick embryos andpresent evidence thatWnt/b-catenin signaling is required for upperjaw morphogenesis by regulating the expression of Lhx8, Msx1 andMsx2 during lip fusion.
MATERIALS AND METHODS
Embryos and beads implantation Fertilized White leghorn eggs werepurchased from Takeuchi Farm (Nara, Japan) and incubated at 38�C until embryosreached the appropriate stage. Embryos were treated with Dkk-1- or 2% BSA-soakedbeads. Affigel Blue beads (BioRad) were soaked in 0.1 mg/ml Dkk-1 (DKK-1, Abcam)with 2% bovine serum albumin (BSA). Sham operations were performed using beadssoaked in 2% BSA. In all cases, a small slit was carefully made in the right side of the
FIG. 2. Chick embryos at HH 38 after Dkk-1-soaked bead implantation. Lateral (B, E) or axial (Control embryos treated with 2% BSA-soaked beads. (DeF) Dkk-1-soaked beads caused defedotted line). Hypoplasia of the premaxilla and palatine bone and a defect of the jugal bone ocBars: 5 mm.
maxillary prominence and the beads were inserted into the slit at HH 22 (22) (beforethe fusion of the frontonasal prominence and maxillary prominence) under amicroscope (Leica, Germany). The left side of the maxillary prominence served asthe control site (Fig. 1). Animal procedures were approved by the Nara MedicalUniversity Animal Care and Use committee.
RNA isolation, real-time RT-PCR and semi-quantitative RT-PCR Maxillaryprominences were dissected 6 h after bead implantation. Total RNA was isolatedwith Sepasol-RNA I Super G (Nacalai Tesque, Japan) from the dissected processes.We then synthesized cDNA using the QuantiTect Reverse Transcription kit (QIAGEN,Germany) with PCR primers listed in Supplementary Table S1. The cDNA synthesizedfrom the left sides of themaxillary prominence served as control. Lhx8 chicken cDNAwas prepared as previously described (16). Msx1 and Msx2 cDNAs were kindlyprovided by J.M. Richman (23).
Real-time RT-PCRwas carried out according to the LightCycler supplier’s manual(Roche) using SYBR Green Realtime PCR Master Mix Plus (TOYOBO, Japan). The ratioof the mRNA levels for each sample was calculated by normalizing the comparativequantitation values to those of GAPDH mRNA.
Semi-quantitative RT-PCR was performed as described in the manufacture’sprotocol. The PCR products were resolved by electrophoresis through an 8% poly-acrylamide gel and then stained with ethidium bromide. At least six processes werecontained in one sample, and three sets of samples were used and examined in eachquantitative analysis for statistical significance.
Skeletal staining To study the morphological changes of bone and cartilage,embryos were fixed at HH 38 (after 12 days’ incubation) and kept in 100% ethanol for4 days. The embryos were then infiltrated with acetone for 4 days, stained withAlcian blue and Alizarin red for up to 10 days, and cleared with KOH as previouslydescribed (23).
Immunohistochemistry and apoptosis assay Heads of chick embryos werefixed in 4% paraformaldehyde overnight. The heads were washed with 30% sucroseafter the fixation and frozen in OCT for coronal sections. Analysis of apoptosis wasdone using cleaved caspase-3 antibody (1:500, rabbit polyclonal, Cell SignalingTechnology) (24).
Statistical analysis The deltaedelta CT method and ANOVA using MicrosoftExcel were carried out for statistical comparisons and a value of p < 0.05 wasconsidered significant.
RESULTS
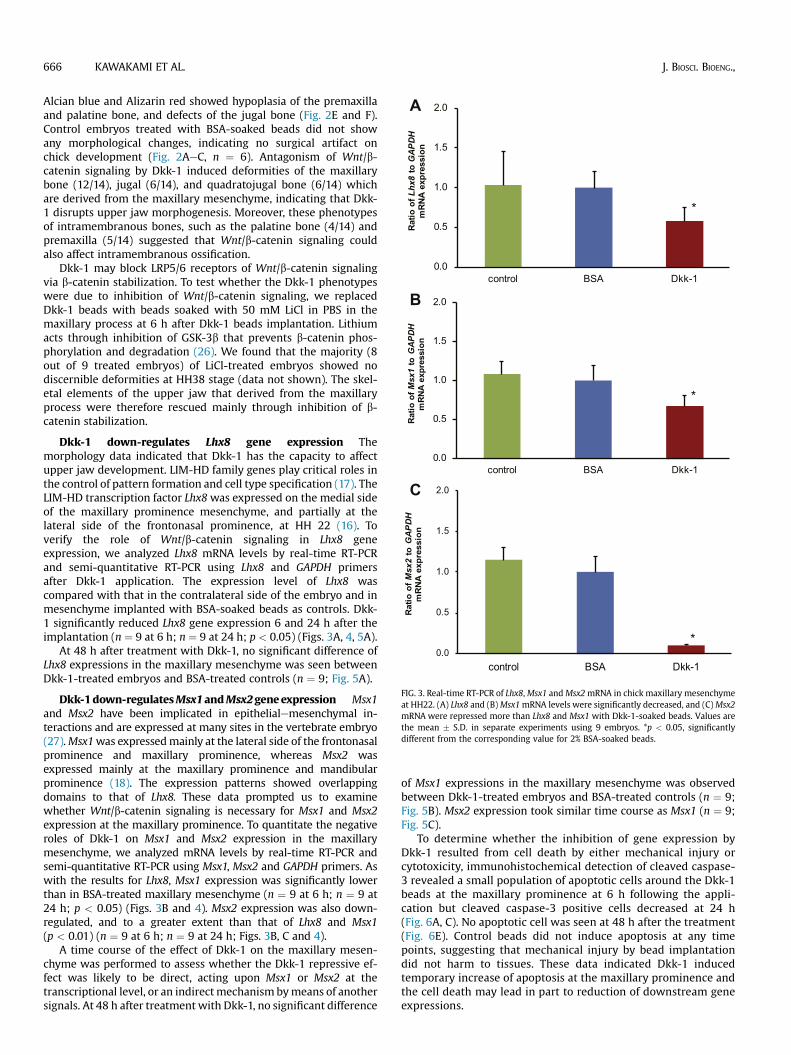
External and skeletal changes of the chick face afterinhibition of Wnt/b-catenin signaling Dkk-1 is one of the in-hibitor of the canonical Wnt/b-catenin signaling whose receptorsare expressed and activated around facial prominences (25). Todetermine whether direct antagonism of Wnt/b-catenin signalingcan affect upper jaw development, we applied Dkk-1-soakedbeads to the maxillary prominence of HH 22 chick embryos,which were then incubated until they reached HH 38. Theembryo heads showed severe defect of the maxilla and collapsedeye at the treated side (Fig. 2D, n ¼ 14). Skeletal staining with
C, F) views of head skeletons stained with Alizarin red and Alcian blue are shown. (AeC)ct of the maxilla (white arrow) and the eye became embedded into the maxilla (yellowcurred (E and F; black arrowheads). p: palatine bone; mx: maxillary bone; j: jugal bone.
0.0
0.5
1.0
1.5
2.0
control BSA Dkk-1
Ratio
o
f L
hx8
to
G
AP
DH
mR
NA
exp
ressio
n
0.0
0.5
1.0
1.5
2.0
control BSA Dkk-1
Ratio
o
f M
sx1
to
G
AP
DH
mR
NA
exp
ressio
n
0.0
0.5
1.0
1.5
2.0
control BSA Dkk-1
Ratio
o
f M
sx2
to
G
AP
DH
mR
NA
exp
ressio
n
*
*
*
A
B
C
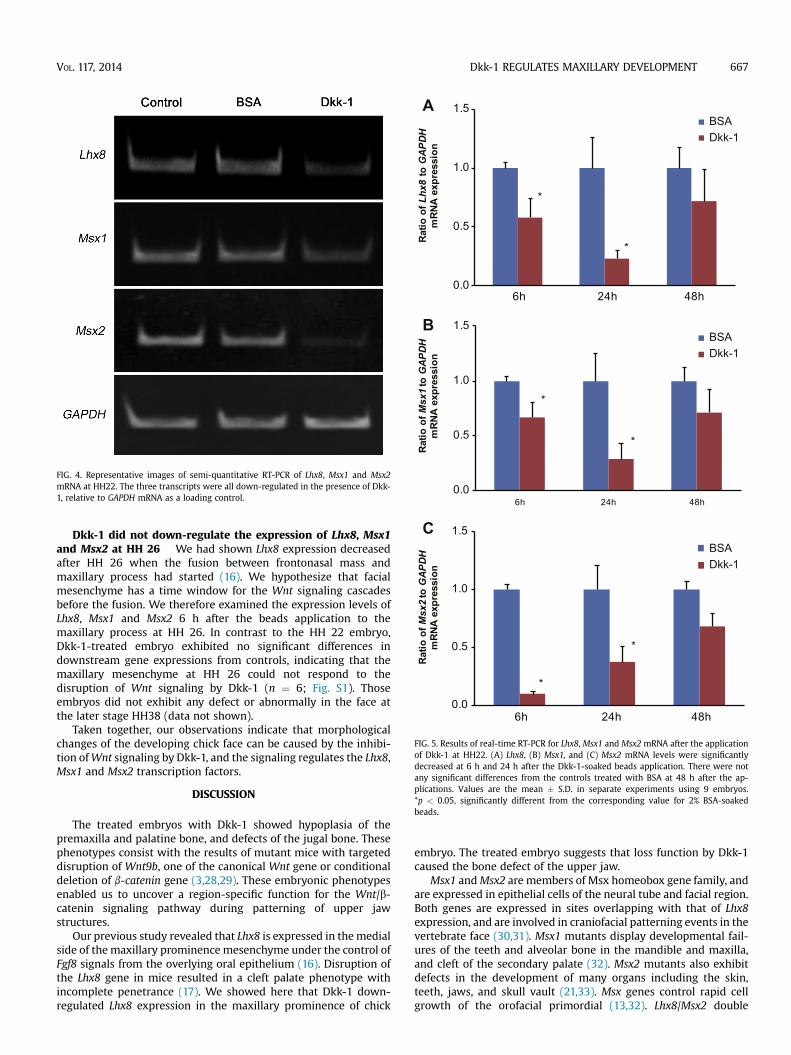
FIG. 3. Real-time RT-PCR of Lhx8,Msx1 andMsx2mRNA in chick maxillary mesenchymeat HH22. (A) Lhx8 and (B)Msx1mRNA levels were significantly decreased, and (C)Msx2mRNA were repressed more than Lhx8 and Msx1 with Dkk-1-soaked beads. Values arethe mean � S.D. in separate experiments using 9 embryos. *p < 0.05, significantlydifferent from the corresponding value for 2% BSA-soaked beads.
666 KAWAKAMI ET AL. J. BIOSCI. BIOENG.,
Alcian blue and Alizarin red showed hypoplasia of the premaxillaand palatine bone, and defects of the jugal bone (Fig. 2E and F).Control embryos treated with BSA-soaked beads did not showany morphological changes, indicating no surgical artifact onchick development (Fig. 2AeC, n ¼ 6). Antagonism of Wnt/b-catenin signaling by Dkk-1 induced deformities of the maxillarybone (12/14), jugal (6/14), and quadratojugal bone (6/14) whichare derived from the maxillary mesenchyme, indicating that Dkk-1 disrupts upper jaw morphogenesis. Moreover, these phenotypesof intramembranous bones, such as the palatine bone (4/14) andpremaxilla (5/14) suggested that Wnt/b-catenin signaling couldalso affect intramembranous ossification.
Dkk-1 may block LRP5/6 receptors of Wnt/b-catenin signalingvia b-catenin stabilization. To test whether the Dkk-1 phenotypeswere due to inhibition of Wnt/b-catenin signaling, we replacedDkk-1 beads with beads soaked with 50 mM LiCl in PBS in themaxillary process at 6 h after Dkk-1 beads implantation. Lithiumacts through inhibition of GSK-3b that prevents b-catenin phos-phorylation and degradation (26). We found that the majority (8out of 9 treated embryos) of LiCl-treated embryos showed nodiscernible deformities at HH38 stage (data not shown). The skel-etal elements of the upper jaw that derived from the maxillaryprocess were therefore rescued mainly through inhibition of b-catenin stabilization.
Dkk-1 down-regulates Lhx8 gene expression Themorphology data indicated that Dkk-1 has the capacity to affectupper jaw development. LIM-HD family genes play critical roles inthe control of pattern formation and cell type specification (17). TheLIM-HD transcription factor Lhx8was expressed on the medial sideof the maxillary prominence mesenchyme, and partially at thelateral side of the frontonasal prominence, at HH 22 (16). Toverify the role of Wnt/b-catenin signaling in Lhx8 geneexpression, we analyzed Lhx8 mRNA levels by real-time RT-PCRand semi-quantitative RT-PCR using Lhx8 and GAPDH primersafter Dkk-1 application. The expression level of Lhx8 wascompared with that in the contralateral side of the embryo and inmesenchyme implanted with BSA-soaked beads as controls. Dkk-1 significantly reduced Lhx8 gene expression 6 and 24 h after theimplantation (n ¼ 9 at 6 h; n ¼ 9 at 24 h; p < 0.05) (Figs. 3A, 4, 5A).
At 48 h after treatment with Dkk-1, no significant difference ofLhx8 expressions in the maxillary mesenchyme was seen betweenDkk-1-treated embryos and BSA-treated controls (n ¼ 9; Fig. 5A).
Dkk-1down-regulatesMsx1andMsx2geneexpression Msx1and Msx2 have been implicated in epithelialemesenchymal in-teractions and are expressed at many sites in the vertebrate embryo(27).Msx1was expressedmainly at the lateral side of the frontonasalprominence and maxillary prominence, whereas Msx2 wasexpressed mainly at the maxillary prominence and mandibularprominence (18). The expression patterns showed overlappingdomains to that of Lhx8. These data prompted us to examinewhether Wnt/b-catenin signaling is necessary for Msx1 and Msx2expression at the maxillary prominence. To quantitate the negativeroles of Dkk-1 on Msx1 and Msx2 expression in the maxillarymesenchyme, we analyzed mRNA levels by real-time RT-PCR andsemi-quantitative RT-PCR using Msx1, Msx2 and GAPDH primers. Aswith the results for Lhx8, Msx1 expression was significantly lowerthan in BSA-treated maxillary mesenchyme (n ¼ 9 at 6 h; n ¼ 9 at24 h; p < 0.05) (Figs. 3B and 4). Msx2 expression was also down-regulated, and to a greater extent than that of Lhx8 and Msx1(p < 0.01) (n ¼ 9 at 6 h; n ¼ 9 at 24 h; Figs. 3B, C and 4).
A time course of the effect of Dkk-1 on the maxillary mesen-chyme was performed to assess whether the Dkk-1 repressive ef-fect was likely to be direct, acting upon Msx1 or Msx2 at thetranscriptional level, or an indirectmechanism bymeans of anothersignals. At 48 h after treatment with Dkk-1, no significant difference
of Msx1 expressions in the maxillary mesenchyme was observedbetween Dkk-1-treated embryos and BSA-treated controls (n ¼ 9;Fig. 5B). Msx2 expression took similar time course as Msx1 (n ¼ 9;Fig. 5C).
To determine whether the inhibition of gene expression byDkk-1 resulted from cell death by either mechanical injury orcytotoxicity, immunohistochemical detection of cleaved caspase-3 revealed a small population of apoptotic cells around the Dkk-1beads at the maxillary prominence at 6 h following the appli-cation but cleaved caspase-3 positive cells decreased at 24 h(Fig. 6A, C). No apoptotic cell was seen at 48 h after the treatment(Fig. 6E). Control beads did not induce apoptosis at any timepoints, suggesting that mechanical injury by bead implantationdid not harm to tissues. These data indicated Dkk-1 inducedtemporary increase of apoptosis at the maxillary prominence andthe cell death may lead in part to reduction of downstream geneexpressions.
0.0
0.5
1.0
1.5
6h 24h 48h
Ratio
o
f L
hx8
to
G
AP
DH
mR
NA
exp
ressio
n
Dkk-1BSA
0.0
0.5
1.0
1.5
6h 24h 48h
Ratio
o
f M
sx
1to
G
AP
DH
mR
NA
exp
ressio
n
Ratio
o
f M
sx
2to
G
AP
DH
mR
NA
exp
ressio
n
A
B
C
Dkk-1BSA
0.0
0.5
1.0
1.5
6h 24h 48h
*
*
*
*
*
*
Dkk-1BSA
FIG. 5. Results of real-time RT-PCR for Lhx8, Msx1 and Msx2 mRNA after the applicationof Dkk-1 at HH22. (A) Lhx8, (B) Msx1, and (C) Msx2 mRNA levels were significantlydecreased at 6 h and 24 h after the Dkk-1-soaked beads application. There were notany significant differences from the controls treated with BSA at 48 h after the ap-plications. Values are the mean � S.D. in separate experiments using 9 embryos.*p < 0.05, significantly different from the corresponding value for 2% BSA-soakedbeads.
FIG. 4. Representative images of semi-quantitative RT-PCR of Lhx8, Msx1 and Msx2mRNA at HH22. The three transcripts were all down-regulated in the presence of Dkk-1, relative to GAPDH mRNA as a loading control.
VOL. 117, 2014 Dkk-1 REGULATES MAXILLARY DEVELOPMENT 667
Dkk-1 did not down-regulate the expression of Lhx8, Msx1and Msx2 at HH 26 We had shown Lhx8 expression decreasedafter HH 26 when the fusion between frontonasal mass andmaxillary process had started (16). We hypothesize that facialmesenchyme has a time window for the Wnt signaling cascadesbefore the fusion. We therefore examined the expression levels ofLhx8, Msx1 and Msx2 6 h after the beads application to themaxillary process at HH 26. In contrast to the HH 22 embryo,Dkk-1-treated embryo exhibited no significant differences indownstream gene expressions from controls, indicating that themaxillary mesenchyme at HH 26 could not respond to thedisruption of Wnt signaling by Dkk-1 (n ¼ 6; Fig. S1). Thoseembryos did not exhibit any defect or abnormally in the face atthe later stage HH38 (data not shown).
Taken together, our observations indicate that morphologicalchanges of the developing chick face can be caused by the inhibi-tion ofWnt signaling by Dkk-1, and the signaling regulates the Lhx8,Msx1 and Msx2 transcription factors.
DISCUSSION
The treated embryos with Dkk-1 showed hypoplasia of thepremaxilla and palatine bone, and defects of the jugal bone. Thesephenotypes consist with the results of mutant mice with targeteddisruption of Wnt9b, one of the canonical Wnt gene or conditionaldeletion of b-catenin gene (3,28,29). These embryonic phenotypesenabled us to uncover a region-specific function for the Wnt/b-catenin signaling pathway during patterning of upper jawstructures.
Our previous study revealed that Lhx8 is expressed in themedialside of the maxillary prominencemesenchyme under the control ofFgf8 signals from the overlying oral epithelium (16). Disruption ofthe Lhx8 gene in mice resulted in a cleft palate phenotype withincomplete penetrance (17). We showed here that Dkk-1 down-regulated Lhx8 expression in the maxillary prominence of chick
embryo. The treated embryo suggests that loss function by Dkk-1caused the bone defect of the upper jaw.
Msx1 andMsx2 are members of Msx homeobox gene family, andare expressed in epithelial cells of the neural tube and facial region.Both genes are expressed in sites overlapping with that of Lhx8expression, and are involved in craniofacial patterning events in thevertebrate face (30,31). Msx1 mutants display developmental fail-ures of the teeth and alveolar bone in the mandible and maxilla,and cleft of the secondary palate (32). Msx2 mutants also exhibitdefects in the development of many organs including the skin,teeth, jaws, and skull vault (21,33). Msx genes control rapid cellgrowth of the orofacial primordial (13,32). Lhx8/Msx2 double
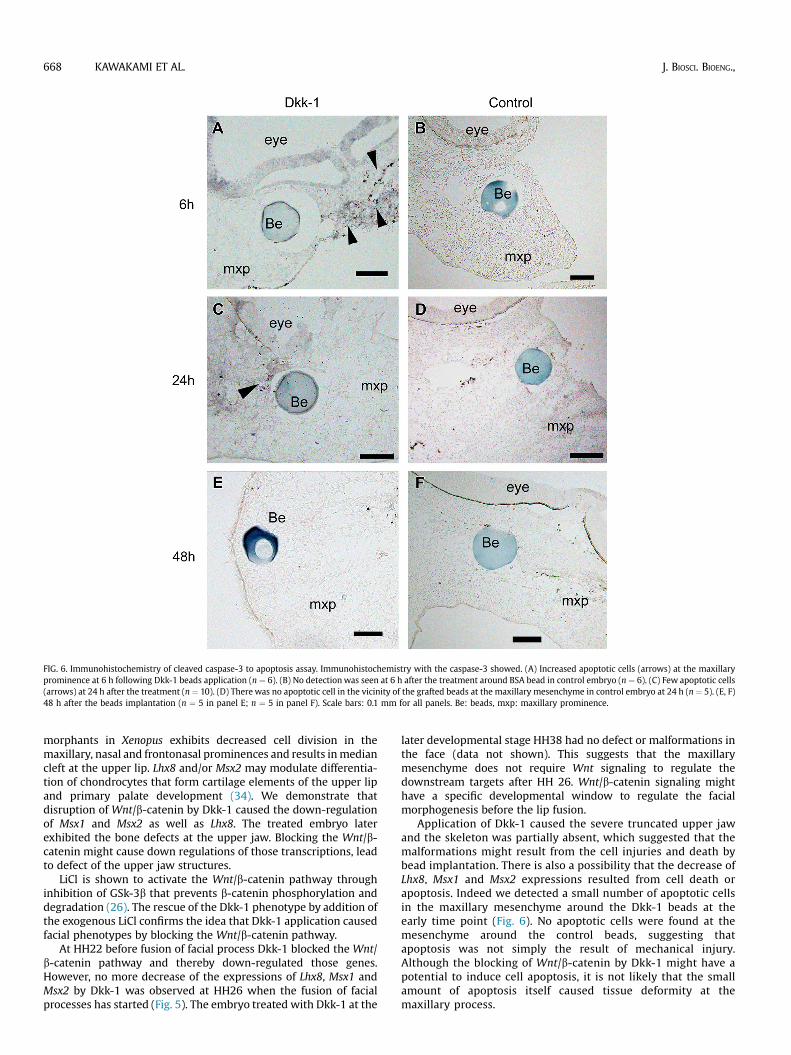
FIG. 6. Immunohistochemistry of cleaved caspase-3 to apoptosis assay. Immunohistochemistry with the caspase-3 showed. (A) Increased apoptotic cells (arrows) at the maxillaryprominence at 6 h following Dkk-1 beads application (n ¼ 6). (B) No detectionwas seen at 6 h after the treatment around BSA bead in control embryo (n ¼ 6). (C) Few apoptotic cells(arrows) at 24 h after the treatment (n ¼ 10). (D) There was no apoptotic cell in the vicinity of the grafted beads at the maxillary mesenchyme in control embryo at 24 h (n ¼ 5). (E, F)48 h after the beads implantation (n ¼ 5 in panel E; n ¼ 5 in panel F). Scale bars: 0.1 mm for all panels. Be: beads, mxp: maxillary prominence.
668 KAWAKAMI ET AL. J. BIOSCI. BIOENG.,
morphants in Xenopus exhibits decreased cell division in themaxillary, nasal and frontonasal prominences and results inmediancleft at the upper lip. Lhx8 and/or Msx2 may modulate differentia-tion of chondrocytes that form cartilage elements of the upper lipand primary palate development (34). We demonstrate thatdisruption of Wnt/b-catenin by Dkk-1 caused the down-regulationof Msx1 and Msx2 as well as Lhx8. The treated embryo laterexhibited the bone defects at the upper jaw. Blocking the Wnt/b-catenin might cause down regulations of those transcriptions, leadto defect of the upper jaw structures.
LiCl is shown to activate the Wnt/b-catenin pathway throughinhibition of GSk-3b that prevents b-catenin phosphorylation anddegradation (26). The rescue of the Dkk-1 phenotype by addition ofthe exogenous LiCl confirms the idea that Dkk-1 application causedfacial phenotypes by blocking the Wnt/b-catenin pathway.
At HH22 before fusion of facial process Dkk-1 blocked the Wnt/b-catenin pathway and thereby down-regulated those genes.However, no more decrease of the expressions of Lhx8, Msx1 andMsx2 by Dkk-1 was observed at HH26 when the fusion of facialprocesses has started (Fig. 5). The embryo treated with Dkk-1 at the
later developmental stage HH38 had no defect or malformations inthe face (data not shown). This suggests that the maxillarymesenchyme does not require Wnt signaling to regulate thedownstream targets after HH 26. Wnt/b-catenin signaling mighthave a specific developmental window to regulate the facialmorphogenesis before the lip fusion.
Application of Dkk-1 caused the severe truncated upper jawand the skeleton was partially absent, which suggested that themalformations might result from the cell injuries and death bybead implantation. There is also a possibility that the decrease ofLhx8, Msx1 and Msx2 expressions resulted from cell death orapoptosis. Indeed we detected a small number of apoptotic cellsin the maxillary mesenchyme around the Dkk-1 beads at theearly time point (Fig. 6). No apoptotic cells were found at themesenchyme around the control beads, suggesting thatapoptosis was not simply the result of mechanical injury.Although the blocking of Wnt/b-catenin by Dkk-1 might have apotential to induce cell apoptosis, it is not likely that the smallamount of apoptosis itself caused tissue deformity at themaxillary process.

VOL. 117, 2014 Dkk-1 REGULATES MAXILLARY DEVELOPMENT 669
Wnt/b-catenin signaling plays an inductive role in the formationof the oral apparatus (2). Disruption ofWnt signaling has a profoundeffect upon the development of facial mesenchyme and their de-rivatives. We used Dkk-1-soaked beads for showing the effect ofWnt/b-catenin signalingon facial development. OurdatawithDkk-1highlight a role for Wnt/b-catenin signaling in the developingmaxillary prominence of chick embryos: a signaling deficiency leadstomidfacial morphological changes of in the chick due to the down-regulationof Lhx8,Msx1 andMsx2geneexpression. These genesmaybe regulatedasdownstreamtargetsofWnt/b-catenin signaling sincethe evidence that Msx1 expression is undetectable in the face in b-catenin mutant mice (3). Our findings suggest that Wnt/b-cateninsignaling has an important function in maxillary morphogenesis byregulating Lhx8, Msx1 and Msx2 transcription factor expression atthe maxillary prominence.
Supplementary information associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.jbiosc.2013.11.015.
ACKNOWLEDGMENTS
We thank Dr. Ryosuke Fujii for real-time RT-PCR and RT-PCRanalysis, and Ian Smith for manuscript editing. This work wassupported by a Grant-in-Aid for Scientific Research from JSPS(21592610, 24593106) to MK. There is no conflict of interest inconnection with this paper.
References
1. Szabo-Rogers, H. L., Smithers, L. E., Yakob, W., and Liu, K. J.: New directionsin craniofacial morphogenesis, Dev. Biol., 341, 84e94 (2010).
2. Lin, C., Fisher, A. V., Yin, Y., Maruyama, T., Veith, M. G., Dhandha, M.,Huang, G. J., Hsu, W., and Ma, L.: The inductive role of Wnt-b-catenin signalingin the formation of oral apparatus, Dev. Biol., 356, 40e50 (2011).
3. Sun, Y., Teng, I., Huo, R., Rosenfeld, M. G., Olson, L. E., Li, X., and Li, X.:Asymmetric requirement of surface epithelial b-catenin during the upper andlower jaw development, Dev. Dyn., 241, 663e674 (2012).
4. Bejsovec, A.: Wnt signaling: an embarrassment of receptors, Curr. Biol., 10,R919eR922 (2000).
5. Gorden,M.D.andNusse,R.:Wntsignaling:multiplepathways,multiple receptors,and multiple transcription factors, J. Biol. Chem., 281, 22429e22433 (2006).
6. MacDonald, B. T., Tamai, K., and He, X.:Wnt/b-catenin signaling: components,mechanisms, and diseases, Dev. Cell, 17, 9e26 (2009).
7. Kawano, Y. and Kypta, R.: Secreted antagonists of the Wnt signaling pathway,J. Cell Sci., 116, 2627e2634 (2003).
8. Liu, F. and Millar, S. E.: Wnt/b-catenin signaling in oral tissue developmentand disease, J. Dent. Res., 89, 318e330 (2010).
9. Grotewold, L. and R}uther, U.: The Wnt antagonist Dickkopf-1 is regulated byBmp signaling and c-Jun and modulates programmed cell death, EMBO J., 21,966e975 (2002).
10. Mukhopadhyay, M., Shtrom, S., Rodriguez-Esteban, C., Chen, L., Tsukui, T.,Gomer,L.,Dorward,D.W.,Glinka,A.,Grinberg,A.,Huang,S.P.,Niehrs,C., IzpisuaBelmonte, J. C., and other 1 author: Dickkopf1 is required for embryonic headinduction and limbmorphogenesis in the mouse, Dev. Cell, 1, 423e434 (2001).
11. Nicolas, F., Jones, V., Khoo, P. L., Bogani, D., Hardy, A., Steiner, K.,Mukhopadhyay, M., Westphal, H., Nolan, P. M., Arkell, R., and Tam, P. P. L.:Stringent requirement of a proper level of canonical Wnt signaling activity forhead formation in mouse embryo, Development, 138, 667e676 (2011).
12. Vieria, A. R., Avila, J. R., Daack-Hirsch, S., Dragan, E., Felix, T. M., Rahimov, F.,Harrington, J., Schultz, R. R., Watanabe, Y., Johnson, M., Fang, J.,O’Brien, S. E., and other 6 authors: Medical sequencing of candidate genes fornonsyndromic cleft lip and palate, PLoS Genet., 1, e64 (2005).
13. Song, L., Li, Y., Wang, K., Wang, Y. Z., Molotkov, A., Gao, L., Zhao, T.,Yamagami, T., Wang, Y., Gan, Q., Pleasure, D. E., and Zhou, C. J.: Lrp-mediated
canonical Wnt signaling is required for lip formation and fusion, Development,136, 3161e3171 (2009).
14. Grigoriou, M., Tucker, A. S., Sharpe, P. T., and Pachnis, V.: Expression andregulation of Lhx6 and Lhx7, a novel subfamily of LIM homeodomain encodinggenes, suggests a role in mammalian head development, Development, 125,2063e2074 (1998).
15. Zhang, Y., Mori, T., Takaki, M., Takeuchi, M., Iseki, K., Hagino, S.,Murakawa, M., Yokoya, S., and Wanaka, A.: Comparison of the expressionpatterns of two LIM-homeodomain genes, Lhx6 and L3/Lh8, in the developingpalate, Orthod. Craniofac. Res., 5, 65e70 (2002).
16. Inoue, M., Kawakami, M., Tatsumi, K., Manabe, T., Makinodan, M.,Matsuyoshi, H., Kirita, T., and Wanaka, A.: Expression and regulation of theLIM homeodomain gene L3/Lhx8 suggests a role in upper lip development ofthe chick embryo, Anat. Embryol. (Berl.), 211, 247e254 (2006).
17. Zhao, Y., Guo, Y. J., Tomac, A. C., Taylor, N. R., Grinberg, A., Lee, E. J.,Huang, S., and Westphal, H.: Isolated cleft palate in mice with a targetedmutation of the LIM homeobox gene Lhx8, Proc. Natl. Acad. Sci. USA, 96,15002e15006 (1999).
18. Brown, J. M., Robertson, K. E., Wedden, S. E., and Tickle, C.: Alterations inMsx1 and Msx2 expression correlate with inhibition of outgrowth of chickfacial primordia induced by retinoic acid, Anat. Embryol. (Berl.), 195, 203e207(1997).
19. Hu, G., Lee, H., Price, S. M., Shen, M. M., and Abate-Shen, C.: Msx homeoboxgenes inhibit differentiation through upregulation of cyclin D1, Development,128, 2373e2384 (2001).
20. Ishii, M., Merrill, A. E., Chan, Y. S., Gitelman, I., Rice, D. P., Sucov, H. M., andMaxon, R. E., Jr.: Msx2 and Twist cooperatively control the development of theneural crest-derived skeletogenic mesenchyme of the murine skull vault,Development, 130, 6131e6142 (2003).
21. Satokata, I., Ma, L., Ohshima, H., Bei, M., Woo, I., Nishizawa, K., Maeda, T.,Takano, Y., Uchiyama, M., Heaney, S., and other 4 authors: Msx2 deficiencyin mice causes pleiotropic defects in bone growth and ectodermal organ for-mation, Nat. Genet., 24, 391e395 (2000).
22. Hamburger, V. and Hamilton, H. L.: A series of normal stages in the devel-opment of the chick embryo, J. Morphol., 88, 49e92 (1951).
23. Ashique, A. M., Fu, K., and Richman, J. M.: Endogenous bone morphogeneticproteins regulate outgrowth and epithelial survival during avian lip fusion,Development, 129, 4647e4660 (2002).
24. Tatsumi, K., Haga, S., Matsuyoshi, H., Inoue, M., Manabe, T., Makinodan, M.,and Wanaka, A.: Characterization of cells with proliferative activity after abrain injury, Neurochem. Int., 46, 381e389 (2005).
25. Jeffs, P., Jaques, K., and Osmond, M.: Cell death in cranial neural crestdevelopment, Anat. Embryol. (Berl.), 185, 583e588 (1992).
26. Klein, P. S. and Melton, D. A.: A molecular mechanism for the effect of lithiumon development, Proc. Natl. Acad. Sci. USA, 93, 8455e8459 (1996).
27. Alappat, S., Zhang, Z. Y., and Chen, Y. P.: Msx homeobox gene family andcraniofacial development, Cell Res., 13, 429e442 (2003).
28. Lan, Y., Ryan, R. C., Zhang, Z., Bullard, S. A., Bush, J. O., Maltby, K. M.,Lidral, A. C., and Jiang, R.: Expression of Wnt9b and activation of canonicalWnt signaling during midfacial morphogenesis in mice, Dev. Dyn., 235,1448e1454 (2006).
29. Carroll, T. J., Park, J. S., Hayashi, S., Majumdar, S., and McMahon, A. P.:Wnt9b plays a central role in the regulation of mesenchymal to epithelialtransitions underlying organogenesis of the mammalian urogenital system,Dev. Cell, 9, 283e292 (2005).
30. Potts, L. F. and Sadler, T. W.: Disruption of msx-1 and msx-2 reveals roles forthese genes in craniofacial, eye, and axial development, Dev. Dyn., 209, 70e84(1997).
31. Levi, G., Mantero, S., Barbieri, O., Cantatore, D., Paleari, L., Beverdam, A.,Genova, F., Robert, B., and Merlo, G. F.: Msx1 and Dlx5 act independently indevelopment of craniofacial skeleton, but converge on the regulation of Bmpsignaling in palate formation, Mech. Dev., 123, 3e16 (2006).
32. Satokata, I. and Mass, R.: Msx1 deficient mice exhibit cleft palate and ab-normalities of craniofacial and tooth development, Nat. Genet., 6, 348e356(1994).
33. Ishii, M., Han, J., Yen, H. Y., Sucov, H. M., Chai, Y., and Maxson, R. E., Jr.:Combined deficiencies of Msx1 and Msx2 cause impaired patterning and sur-vival of the cranial neural crest, Development, 132, 4937e4950 (2005).
34. Kennedy, A. E. and Dickinson, A. J. G.: Median facial clefts in Xenopus laevis:roles of retinoic acid signaling and homeobox genes, Dev. Biol., 365, 229e240(2012).


















