
Human renal UOK130 tumor cells: A drug resistant cell line with highly selective over-expression of...
7
Human renal UOK130 tumor cells: A drug resistant cell line with highly selective over-expression of glutathione S-transferase-π isozyme Wei Wang a , Guangxian Liu a , Jiang Zheng a,b, ⁎ a Department of Pharmaceutical Sciences, Northeastern University, Boston, MA 02115, USA b Center for Developmental Therapeutics, Seattle Children's Hospital Research Institute, Division of Gastroenterology, Department of Pediatrics, University of Washington, Seattle, WA 98101, USA Received 3 January 2007; received in revised form 6 April 2007; accepted 12 April 2007 Available online 22 April 2007 Abstract Drug resistance remains one of the primary causes of suboptimal outcomes in cancer chemotherapy. This study reports the development of a drug resistant cell line with over-expression of glutathione S-transferase (GST). The resistant tumor cell model was established by continuous exposure of UOK130, a human renal tumor cell line, to escalating concentrations of cisplatin. By immunoblotting the cisplatin-resistant cells (UOK CR ) were found to express an elevated level of GST-π isozyme. Neither α nor μ isozyme was detected by the corresponding polyclonal antibodies. A significant increase in cellular glutathione (GSH) was also observed in UOK CR cells comparative to the parental cells. In addition, the continuous exposure to cisplatin resulted in decreased cell susceptibility not only to cisplatin (resistant factor: 5.7) but also to melphalan (resistant factor: 2.9) and chlorambucil (resistant factor: 2.3). A transgenic cell line was developed by transfecting of UOK130 cells with GST-π cDNA. The transfection of the GST-π virus into UOK130 cell apparently increased its intracellular GST-π activity. The resistance of the transfectants to cisplatin was consistently increased, compared with that of mock transfectants. A haloenol lactone (HEL) derivative known as a selective inhibitor of GST-π was applied to evaluate the suitableness of the cell model for GST-π-mediated drug resistance studies. The inhibitor was found to potentiate the cytoxicity of cisplatin to both UOK130 and UOK CR cell lines and to reverse their resistance to cisplatin. In conclusion, we developed a multiple drug resistant tumor cell line with selective over-expression of GST-π. The cell model provides a unique tool for mechanistic studies of drug resistance mediated by over-expression of GST-π. © 2007 Elsevier B.V. All rights reserved. Keywords: Cisplatin resistance; Glutathione S-transferase; Haloenol lactone 1. Introduction The study of the mechanisms by which cancers elude treatment has yielded a wealth of information about how to circumvent drug resistance in cancer cells and/or design agents that are not subject to the usual means of resistance (Gottesman, 2002). Studies of cellular mechanisms of multiple drug resistance indicated that exposure to anticancer drugs may lead to the induction and expression of many gene products that protect the cell. Major advances have been made in understanding drug metabolism and deposition and how adaptive phase II metabolism can result in altered cytotoxicity to anticancer drugs (O'Brien et al., 2000; Townsend and Tew, 2003a; Townsend et al., 2003). The glutathione S-transferases (GSTs) are an extended fami- ly of phase II enzymes that catalyze the conjugation of gluta- thione (GSH) to a wide variety of exogenous electrophilic compounds including anticancer drugs, pesticides, and geno- toxic molecules (Zheng et al., 1996). It is now generally ac- cepted that GSTs, especially GST-π class, are over-expressed with high frequency in a wide variety of tumors whereas GST-π levels in normal or surrounding tissues are low. Over-expression of GST-π has been considered to be a marker for drug resistance in cancer treatment (Tew and Ronai, 1999; Townsend and Tew, 2003b). Cisplatin is a widely used chemotherapeutic agent. Cisplatin- based combination chemotherapy displays significant antitumor activity against cancers of the testis, ovary, head, neck, kidney European Journal of Pharmacology 568 (2007) 61 – 67 www.elsevier.com/locate/ejphar ⁎ Corresponding author. Seattle Children's Hospital Research Institute, 1900 9th Avenue, Seattle, WA 98101, USA. Tel.: +1 206 987 7651; fax: +1 206 987 7660. E-mail address: [email protected] (J. Zheng). 0014-2999/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2007.04.021
Transcript of Human renal UOK130 tumor cells: A drug resistant cell line with highly selective over-expression of...

logy 568 (2007) 61–67www.elsevier.com/locate/ejphar
European Journal of Pharmaco
Human renal UOK130 tumor cells: A drug resistant cell line with highlyselective over-expression of glutathione S-transferase-π isozyme
Wei Wang a, Guangxian Liu a, Jiang Zheng a,b,⁎
a Department of Pharmaceutical Sciences, Northeastern University, Boston, MA 02115, USAb Center for Developmental Therapeutics, Seattle Children's Hospital Research Institute, Division of Gastroenterology,
Department of Pediatrics, University of Washington, Seattle, WA 98101, USA
Received 3 January 2007; received in revised form 6 April 2007; accepted 12 April 2007Available online 22 April 2007
Abstract
Drug resistance remains one of the primary causes of suboptimal outcomes in cancer chemotherapy. This study reports the development of adrug resistant cell line with over-expression of glutathione S-transferase (GST). The resistant tumor cell model was established by continuousexposure of UOK130, a human renal tumor cell line, to escalating concentrations of cisplatin. By immunoblotting the cisplatin-resistant cells(UOKCR) were found to express an elevated level of GST-π isozyme. Neither α nor μ isozyme was detected by the corresponding polyclonalantibodies. A significant increase in cellular glutathione (GSH) was also observed in UOKCR cells comparative to the parental cells. In addition,the continuous exposure to cisplatin resulted in decreased cell susceptibility not only to cisplatin (resistant factor: 5.7) but also to melphalan(resistant factor: 2.9) and chlorambucil (resistant factor: 2.3). A transgenic cell line was developed by transfecting of UOK130 cells with GST-πcDNA. The transfection of the GST-π virus into UOK130 cell apparently increased its intracellular GST-π activity. The resistance of thetransfectants to cisplatin was consistently increased, compared with that of mock transfectants. A haloenol lactone (HEL) derivative known as aselective inhibitor of GST-π was applied to evaluate the suitableness of the cell model for GST-π-mediated drug resistance studies. The inhibitorwas found to potentiate the cytoxicity of cisplatin to both UOK130 and UOKCR cell lines and to reverse their resistance to cisplatin. In conclusion,we developed a multiple drug resistant tumor cell line with selective over-expression of GST-π. The cell model provides a unique tool formechanistic studies of drug resistance mediated by over-expression of GST-π.© 2007 Elsevier B.V. All rights reserved.
Keywords: Cisplatin resistance; Glutathione S-transferase; Haloenol lactone
1. Introduction
The study of themechanisms bywhich cancers elude treatmenthas yielded a wealth of information about how to circumvent drugresistance in cancer cells and/or design agents that are not subjectto the usual means of resistance (Gottesman, 2002). Studies ofcellular mechanisms of multiple drug resistance indicated thatexposure to anticancer drugs may lead to the induction andexpression of many gene products that protect the cell. Majoradvances have been made in understanding drug metabolism anddeposition and how adaptive phase II metabolism can result in
⁎ Corresponding author. Seattle Children's Hospital Research Institute, 19009th Avenue, Seattle, WA 98101, USA. Tel.: +1 206 987 7651; fax: +1 206 9877660.
E-mail address: [email protected] (J. Zheng).
0014-2999/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2007.04.021
altered cytotoxicity to anticancer drugs (O'Brien et al., 2000;Townsend and Tew, 2003a; Townsend et al., 2003).
The glutathione S-transferases (GSTs) are an extended fami-ly of phase II enzymes that catalyze the conjugation of gluta-thione (GSH) to a wide variety of exogenous electrophiliccompounds including anticancer drugs, pesticides, and geno-toxic molecules (Zheng et al., 1996). It is now generally ac-cepted that GSTs, especially GST-π class, are over-expressedwith high frequency in a wide variety of tumors whereas GST-πlevels in normal or surrounding tissues are low. Over-expressionof GST-π has been considered to be a marker for drug resistancein cancer treatment (Tew and Ronai, 1999; Townsend and Tew,2003b).
Cisplatin is a widely used chemotherapeutic agent. Cisplatin-based combination chemotherapy displays significant antitumoractivity against cancers of the testis, ovary, head, neck, kidney

62 W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
and lung (Go and Adjei, 1999). However, its effectiveness in thetreatment of cancers is more limited because of acquired orintrinsic drug resistance. Potential biochemical and molecularmechanisms of cisplatin resistance have been proposed andreviewed (Kartalou and Essigmann, 2001; Perez, 1998).Glutathione (GSH), Glutathione-related enzymes, and the gluta-thione conjugates efflux pumps (GS-X pumps) have beenshown to participate in the detoxification of cisplatin and manyother anticancer drugs (Shen et al., 1997; Tew, 1994; Townsendand Tew, 2003b).
In this study, we established a cisplatin-resistant human renalUOK130 cell line (UOKCR) with selective over-expression ofGST-π isozyme. The chemotherapeutic resistance was found tobe reversed by a haloenol lactone derivative known as a GST-πinhibitor. Additionally, we prepared GST-π transgenic cell lineby incorporation of a GST-π cDNA vector into UOK130 cells.The resulting cells revealed resistance to the cytotoxicity in-duced by cisplatin. The cell lines developed in the study willfacilitate the understanding of the role of GST-π expression indrug resistance.
2. Materials and methods
2.1. Chemicals
HEL (3-cinnamyl-5(E)-bromomethylidenetetrahydro-2-fura-none) was synthesized as reported in our earlier paper (Zhenget al., 1996). Cisplatin, melphalan, chlorambucil, GSH andCDNB (1-chloro-2,4-dinitrobenzene) were obtained fromSigma Chemical Co. (St. Louis, MO). The stock solutionswere prepared in either ethanol or DMSO at 10 mM and dilutedwith buffer. The final concentration of ethanol or DMSO in cellculture medium was less than 0.5%.
2.2. Cell culture
UOK130, a human renal carcinoma cell line, was obtainedfromDr.Michael DeGregorio's lab at the University of Californiaat Davis. Cell culture media and supplies were purchased fromMediatech Inc. (Herndon, VA). The serum for cell culture wasfrom Perbio Hyclone Co. (Logan, UT). UOK130, UOKCR
and their resulting transfected cell lines (UOK-T, UOK-VC)were grown asmonolayer culture in improvedminimum essentialmedium supplemented with 10% fetal bovine serum and100 units/ml penicillin/streptomycin (Invitrogen, Carlesbad,CA) at 37 °C, 5% CO2. Cells were held in an incubator chambercontaining humidified 95% air and 5% CO2 at 37 °C. All cellswere used in the log-phase of growth unless otherwise specified.
2.3. Preparation of a cisplatin-resistant UOK130 cell line(UOKCR)
UOK130 cell line was exposed to incrementally increasingcisplatin concentrations to generate resistance to cisplatin (thecisplatin concentration started from 1 μM with a slow increaseat 5, 10, 20, 40, 60, 80, and 100 μM). The duration of exposureto each cisplatin concentration was about 4–6 weeks, depending
on the cell viability. When a cell confluency of 60–80% wasmaintained for about 2 weeks, the next higher cisplatin con-centration was applied to the medium. In an attempt to limit thefounder effects associated with cloning, resistant cell lines arenot strictly clonal; all surviving colonies were pooled aftercisplatin selection to create the respective resistant cell line.When the resistant phenotype was established, UOKCR cell linewas cultured in the medium without cisplatin for 2 weeks andthen tested with cisplatin (100 μM) again to confirm the stabilityof cisplatin resistance. UOKCR cell line was maintained bycontinually cultured with cisplatin (100 μM for 2 weeks) every3 months to ensure the drug resistant phenotype.
2.4. Preparation of GST-π cDNA transfected UOK130 cells
The vector pET17B-GST-π (containing the whole sequenceof human GST-π) was a gift from Dr. David Eaton at theUniversity of Washington. A PCR was performed to amplifyhuman GST-π gene from the pET17B-GST-π plasmid usingprimers (5′-atggaagcttggtacca-ccatgccgccctacaccgtggtc-3′ (up-stream) and 5′-ggcctctagactctcgagtcactgtttcccgttgccatt-gatgg-3′(downstream). The PCR product was first ligated into a pGEM-T Easy Vector (Promega, Madison, WI). The resulting pGEM-GST-π plasmid was digested by HindIII and XbaI to get a GST-π fragment. The GST-π fragment was then inserted intothe expression vector pcDNA™3.1/V5-HisB (Novagen, SanDiego, CA) by HindIII and Xba I. The cloned GST-π wasidentified by DNA sequencing. FuGENE 6 TransfectionReagent (Roche, Indianapolis, IN) was used for transfec-tion. Briefly, prior to transfection, cells were seeded in DMEMin 6-well culture plate for 24 h; cell density was maintained at50–80% confluency. Then FuGENE 6 Reagent 3 μl and well-purified plasmid DNA 1 μg were added accordingly. After 24-h culture at 37 °C, 5% CO2, culture medium was changed andcontinually incubated for 48 h. Then G-418 (600 μg/ml) wasadded for clone selection. High GST-π expressed cloneswere selected according to the expression level of GST-π com-pared to control clones which were transfected with vectorpcDNA™3.1/V5-HisB encoding the neomycin resistance genebut no GST-π and selected with G-418 by means of the mea-surement of CDNB conjugates.
2.5. Cytotoxicity assay
A selection of alkylating agents and HEL, individually or incombination, was evaluated for their cytotoxic effects on cellgrowth in vitro under specified conditions. In vitro cytotxicitywas assessed using modified clonogenic assay (Morgan et al.,1996). Briefly, cells were plated in triplicate 6-well plate at2×105 cells/ml. Twenty-four hours after plating, drugs dilutedwith medium at various concentrations were added into theexperimental wells. The cells were incubated at 37 °C in 5%CO2 and 95% air for 24 h. Cells were collected, centrifuged,counted and then diluted to 5–20 cells/200 μl with IMEM (10%FBS). These diluted cell solutions were added into 96-wellplates (200 μl/well) and incubated for 2 weeks at 37 °C and 5%CO2 in a humidified incubator. Following the incubation period,

Fig. 1. Western blotting of GST isozymes in the UOK130 and UOKCR celllysates. Blots were hybridized using polyclonal goat antibodies (Detroit R&D,Detroit, MI) raised against human GST-π, -α, -μ and mouse polyclonalantibodies I19 for β-actin (Santa Cruz Biotech, Santa Cruz, CA). Bands werevisualized using ECL kit. (UOK: UOK130 cells).
63W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
the plates were examined microscopically, and the wells withclones were counted. The colony-forming efficiency wascalculated as follows (Masters, 2000):
Efficiency ¼ Number of positive wells=Total number of wells inoculatedð Þ�100k
2.6. GST activity assay
Cells were then harvested, washed with ice-cold PBS bufferand sonicated. The cell homogenates were centrifuged at14,000 g for 30 min at 4 °C. The supernatants were collected foranalysis of GST activity according to the method of Habig(Habig et al., 1974). The reaction mixture contained 100 mMpotassium phosphate buffer, pH=6.5, 1.0 mM GSH, and1.0 mM CDNB. The reaction was initiated by the addition ofGST samples prepared as described above. The rate of productformation of the CDNB conjugate with GSH was determinedspectrophotometrically by monitoring the absorbance at 340 nmin a 6-min kinetic model (read per 30 s), using a VERSA MaxUV–visible spectrophotometer (Molecular Device, Sunnyvale,CA). Specific activities were based on protein concentrations asdetermined using BCA protein assay kit from Pierce Inc.(Rockford, IL).
2.7. Assay of cellular GSH content
Intracellular GSH content was measured by ApoGSH™Glutathione Kit (BioVision Research Products, Mountain View,CA). Briefly, cells (1×106) were collected and then centrifugedat 700 g for 5 min. The cell pellets were washed with 1.0 ml ice-cold PBS buffer, and centrifuged again. The cell pellets were re-suspended in 100 μl ice-cold cell lysis buffer, incubated on icefor 10 min, and then centrifuged at 14,000 g for 10 min at 4 °C.The supernatants were transferred to a 96-well plate, followedby addition of 2 μl of 25 mM monochlorobimane (MCB) and2 μl of the 50 U/ml GST reagent into each well. The resultingplate was incubated at 37 °C for 30 min, and the fluorescence atEx/Em=380/460 nm was measured with a Gemini fluorescencephotometer (Molecular Device, Sunnyvale, CA).
2.8. Western blot
The protein bands resolved by SDS-polyacrylamide gel elec-trophoresis (Invitrogen, Carlesbad, CA) using 3.3% stackingand 10% resolving Tris–glycine gels were transferred to
Table 1Resistance profile of UOK/UOKCR cells
Drug (μM) IC50 (UOK130) IC50 (UOKCR) Resistant factor
Cisplatin 15.58±4.67 88.65±6.89 5.7 a
Chlorambucil 44.54±7.22 100.27±5.33 2.3 a
Melphalan 50.37±9.28 146.03±8.43 2.9 a
Cells were exposed to the indicated drugs for a period of 24 h before colony-forming analysis. Data are represented mean±S.D. of three independentexperiments.a Pb0.05 Student's t-test, compared with parental UOK130 cells.
polyvinylidene difluoride microporous membranes (Immobi-lon-P transfer membranes, Millipore, Billerica, MA) byelectroblotter (Invitrogen, Carlesbad, CA). After 1-h transfer(100 mA), blots were blocked by shaking for 1.0 h in 5% non-fatdry milk in PBST buffer. The blotted polyvinylidene difluoridemembranes were incubated for 2 h with 1/500 dilution ofpolyclonal goat antibodies individually (Detroit R&D, Detroit,MI) raised against human GST-π, μ, α isozymes in PBST buffer(0.2 M sodium phosphate containing 0.5% Tween-20, pH 7.4)with 5% non-fat milk. The immunoblots were incubated for 1.0 hwith horseradish peroxidase conjugated rabbit anti-goat IgGantibodies (1/1000 in PBST buffer). The blotted protein bandswere detected by chemiluminescence using ECL westernblotting kits (Amersham International Plc, UK).
2.9. Data analysis
Data were expressed as mean±S.D. Student's t-test wasused to evaluate the significance of the difference between themeans of HEL+drug combination group and drug alone group.Analysis of variance (ANOVA) was applied to evaluate themultiple group data. A value of Pb0.05 was accepted as astatistically significant level. Curve fitting, graphing, and calcu-lating of IC50 were performed using XLfit 4.0 (IDBS, Surrey,UK) and Sigmaplot 9.0 (SYSTAT, Point Richmond, CA).
3. Results
3.1. Resistance of UOKCR cells to chemotherapeutic agents
Cisplatin-resistant UOK130 cells (UOKCR) were developedby continuous exposure of human renal UOK130 tumor cells toescalating concentrations of cisplatin. The resulting tumor cells

64 W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
were found to be resistant to cisplatin with resistant factor of asmuch as 5.7. Table 1 summarizes the resistance profile ofUOKCR cells. Compared with the parental UOK130 cells,UOKCR cells were not only resistant to cisplatin but also cross-resistant to melphalan and chlorambucil (with resistant factorof 2–3).
3.2. Biochemical changes in GST/GSH
Glutathione S-transferases α, μ, and π are the three majorenzymes reported to be responsible for chemotherapeutic resis-tance. The expression of the three GST isozymes was examinedby immunoblotting. We naturally compared GST-π expressionin UOKCR cells and parental UOK130 cells. A dark chemilu-minescent band responsible for GST-π was observed on thepolyvinylidene difluoride membrane blotted with the proteinsample obtained from UOKCR cells (Fig. 1), whereas a lightchemiluminescent band observed on the membrane blotted withthe same amount of protein sample obtained from the parental
Fig. 2. Biochemical analysis of GST-π activity and GSH contents in UOK130and UOKCR cells. (A) UOK130 cells were collected by trypsinization andhomogenized by ultrasonication. The homogenates were centrifuged at 12,000 gfor 30 min and the supernatants were used for GST activity analysis. GSTactivity was measured as nmol CDNB/min/mg protein and expressed aspercentage of blank control. Data are mean±S.D. of three determinations.(B) Cells were collected by trypsinization and dissolved in cell lysis buffer. Thecell lysates were centrifuged at 12,000 g for 30 min and the supernatants wereused for GSH measurement. Fluorescence was measured at 380/460 nm. GSHcontent was expressed as nmol GSH/106 cells. Data are mean±S.D. of threedeterminations.
Fig. 3. Over-expression of GST-π induces resistance to cisplatin in UOK130transgenic cell lines. UOK130 transgenic cells (UOK-T and UOK-VC) wereplated in six-well plates (1×106 cells/well/2 ml of medium). Cisplatin was thenadded to wells at various concentrations (10–400 μM) and incubated for 24 h.After incubation, cell viability was evaluated by colony-forming assay. Data aremean±S.D. of three determinations (□: UOK-T; ▵: UOK-VC).
cells (Fig. 1). This indicates significant induction of GST-π inthe tumor cells after exposure to cisplatin. We also examined theexpression of GST-α and GST-μ in UOKCR and parental cells.The immunoblotting showed that neither GST-α nor GST-μ waspresent in UOK130 cell line, either in UOKCR cells (Fig. 1). Inaddition to immunoblotting, we chemically determined cyto-solic GST activity towards CDNB. As expected, a significantincrease in GST activity was found in UOKCR cells comparativeto the parental cells (Fig. 2A). Meanwhile, we monitoredcellular GSH contents in the two cell lines. As much as 2-foldincrease in GSH level was found in the UOKCR cells relative tothe parental UOK cells (Fig. 2B). These results indicate thatboth expression of GST-π and biosynthesis of GSH in theUOKCR cells were up-regulated to adapt the cisplatin selectionupon cell growth.
3.3. Susceptibility of GST-π transgenic cells to cisplatin
We also developed GST-π over-expressed UOK130 trans-genic cell line (UOK-T) by transfecting UOK130 cells withGST-π cDNA. As expected, immunoblotting showed apprecia-ble increase in GST-π expression in the transgenic cells (Fig. 3).Additionally, the GST assay showed a 2.5-fold increase in GSTactivity in the transgenic cell line relative to that of the wildtype. The cell viability of UOK-T and the wild type (UOK-VC)were examined after exposure to cisplatin. The dose–responseof cisplatin toxicity to the cell lines was shown in Fig. 3. UOK-T (IC50: 71 μM) was found to be less susceptible to cisplatin
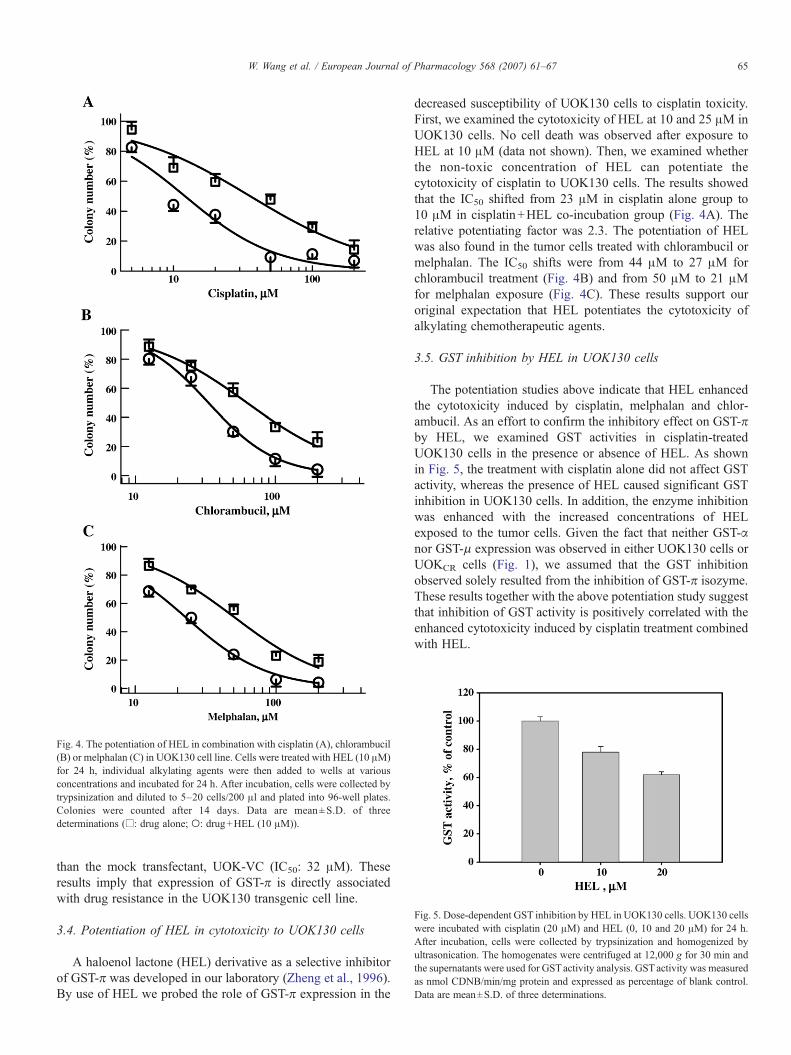
Fig. 4. The potentiation of HEL in combination with cisplatin (A), chlorambucil(B) or melphalan (C) in UOK130 cell line. Cells were treated with HEL (10 μM)for 24 h, individual alkylating agents were then added to wells at variousconcentrations and incubated for 24 h. After incubation, cells were collected bytrypsinization and diluted to 5–20 cells/200 μl and plated into 96-well plates.Colonies were counted after 14 days. Data are mean±S.D. of threedeterminations (□: drug alone; ○: drug+HEL (10 μM)).
Fig. 5. Dose-dependent GST inhibition by HEL in UOK130 cells. UOK130 cellswere incubated with cisplatin (20 μM) and HEL (0, 10 and 20 μM) for 24 h.After incubation, cells were collected by trypsinization and homogenized byultrasonication. The homogenates were centrifuged at 12,000 g for 30 min andthe supernatants were used for GSTactivity analysis. GSTactivity was measuredas nmol CDNB/min/mg protein and expressed as percentage of blank control.Data are mean±S.D. of three determinations.
65W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
than the mock transfectant, UOK-VC (IC50: 32 μM). Theseresults imply that expression of GST-π is directly associatedwith drug resistance in the UOK130 transgenic cell line.
3.4. Potentiation of HEL in cytotoxicity to UOK130 cells
A haloenol lactone (HEL) derivative as a selective inhibitorof GST-π was developed in our laboratory (Zheng et al., 1996).By use of HEL we probed the role of GST-π expression in the
decreased susceptibility of UOK130 cells to cisplatin toxicity.First, we examined the cytotoxicity of HEL at 10 and 25 μM inUOK130 cells. No cell death was observed after exposure toHEL at 10 μM (data not shown). Then, we examined whetherthe non-toxic concentration of HEL can potentiate thecytotoxicity of cisplatin to UOK130 cells. The results showedthat the IC50 shifted from 23 μM in cisplatin alone group to10 μM in cisplatin+HEL co-incubation group (Fig. 4A). Therelative potentiating factor was 2.3. The potentiation of HELwas also found in the tumor cells treated with chlorambucil ormelphalan. The IC50 shifts were from 44 μM to 27 μM forchlorambucil treatment (Fig. 4B) and from 50 μM to 21 μMfor melphalan exposure (Fig. 4C). These results support ouroriginal expectation that HEL potentiates the cytotoxicity ofalkylating chemotherapeutic agents.
3.5. GST inhibition by HEL in UOK130 cells
The potentiation studies above indicate that HEL enhancedthe cytotoxicity induced by cisplatin, melphalan and chlor-ambucil. As an effort to confirm the inhibitory effect on GST-πby HEL, we examined GST activities in cisplatin-treatedUOK130 cells in the presence or absence of HEL. As shownin Fig. 5, the treatment with cisplatin alone did not affect GSTactivity, whereas the presence of HEL caused significant GSTinhibition in UOK130 cells. In addition, the enzyme inhibitionwas enhanced with the increased concentrations of HELexposed to the tumor cells. Given the fact that neither GST-αnor GST-μ expression was observed in either UOK130 cells orUOKCR cells (Fig. 1), we assumed that the GST inhibitionobserved solely resulted from the inhibition of GST-π isozyme.These results together with the above potentiation study suggestthat inhibition of GST activity is positively correlated with theenhanced cytotoxicity induced by cisplatin treatment combinedwith HEL.

Fig. 6. The potentiation of HEL in UOKCR cells. Cells were treated with HEL(10 μM) for 24 h, then cisplatin was added to wells at various concentrations andincubated for 24 h. After incubation, cells were collected by trypsinization anddiluted to 5–20 cells/200 μL and plated into 96-well plates. Colonies werecounted after 14 days. Data are mean±S.D. of three determinations (▵: cisplatinalone; ○: cisplatin+HEL (10 μM)).
66 W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
3.6. Potentiation of HEL in cytotoxicity to UOKCR cells
We also examined the potentiation of HEL in cytotoxicity ofcisplatin to the UOKCR cells we developed. As expected, thepresence of HEL significantly increased the cytotoxicity ofcisplatin to the UOKCR cell line (Fig. 6). The IC50 shifted from109 μM in cisplatin alone group to 40 μM in cisplatin+HEL co-incubation group. This implies that cisplatin resistance isessentially nullified by the addition of the HEL at non-toxicconcentration.
4. Discussion
The major aim of the present study was to develop cell linemodels in support of our mechanistic studies to combat acquiredchemotherapy resistance mediated by over-expression of GST-π. Early studies from this lab reported that UOK130 cellsexpressed GST-π but neither α nor μ isozyme (Zheng et al.,1996; Zheng et al., 1997). Tumor cells, particularly drug resis-tant cancer cells, often express high levels of multiple GSTs,including α, μ, and π isozymes (Tew and Ronai, 1999;Townsend and Tew, 2003b). The selection of GST-π expressionwould make it possible to provide a good cell model to inves-tigate the insight into the biochemical mechanisms of chemo-therapy resistance. We developed a cisplatin-resistant UOK130cell line (UOKCR) by treatment of UOK130 cells with escalatedcisplatin. As expected, an elevated GST-π expression was foundin the UOKCR cells comparative to the parental UOK130 cells.Interestingly, neither α nor μ isozymes was detected by immu-noblotting in either UOK130 or UOKCR cell line (Fig. 1). Thisfinding strongly supports that GST-π may specifically contrib-ute to cisplatin resistance in UOK130 cells. More importantly,the cell line model provides a unique tool to assess GST-π
inhibitors considered as potential agents to fight GST-π-mediated drug resistance.
As the second aim, we tested whether the cell lines aresuitable for our mechanistic studies on chemotherapy resis-tance. Earlier we developed a haloenol lactone derivative (HEL)as a selective inhibitor of GST-π (Zheng et al., 1996). In thepresent study we examined the potentiation of HEL as amodulator on cytotoxicity of cisplatin, chlorambucil and mel-phalan to UOK130 cells, and HEL was found to potentiate thecytotoxicity of all three alkylating agents tested (Fig. 4). Asexpected, HEL showed dose-dependent inhibitory effect oncytosolic GST activity (Fig. 5). Additionally, the enhancementof cytotoxicity observed was positively correlated with GSTinhibition resulting from the exposure to HEL. Furthermore, weapplied HEL in the UOKCR cells for potentiation study. Al-though the UOKCR cells are about 6-fold more resistant tocisplatin than parental UOK130 cells, co-incubation with HELgreatly reversed cisplatin resistance of UOKCR cells (Fig. 6).
Elevated intracellular GSH level was also found in UOKCR
cells (Fig. 2B). Up-regulation of GSH has been reported in drugresistant tumors (Townsend and Tew 2003a,b). It is not sur-prising that the up-regulation of GSH-mediated drug detoxifi-cation is an important factor of cisplatin resistance. HEL hasbeen found to conjugate with GSH as a spontaneous reaction(Zheng et al., 2005). The spontaneous conjugation (depletion)of cellular GSH may be partially responsible for the potentiatingeffect of HEL on the cytotoxicity of cisplatin in UOK130 andUOKCR cells.
Single cDNA transfection experiments offered a more de-fined system for the further mechanistic studies (O'Brien et al.,2000). We manipulated cytosolic GST activity by transfectingGST-π cDNA into UOK130 cells. We successfully preparedstably expressed GST-π transgenic cells, and we found thetransgenic cells are less susceptible to cisplatin than the mocktransfectant. The resistance was found not due to the elevatedGSH contents in these UOK130 transgenic cells (to be pub-lished separately). Therefore we believe that elevated GST-πis one of complicated mechanisms responsible for cisplatinresistance.
In conclusion, we developed a multiple drug resistant humanrenal tumor cell line with selective over-expression of GST-πisozyme. The cell model with isomeric selection of GST ex-pression makes it possible to dissect the complexity of thebiochemical mechanisms of chemotherapy resistance. Our workalso further provided the evidence that GST-π is a therapeutictarget in drug resistance.
Acknowledgement
This work was supported by American Cancer Society GrantRSG-01-059-01-CDD.
Reference
Go, R.S., Adjei, A.A., 1999. Review of the comparative pharmacology andclinical activity of cisplatin and carboplatin. J. Clin. Oncol. 17, 409–422.
Gottesman, M.M., 2002. Mechanisms of cancer drug resistance. Annu. Rev.Med. 53, 615–627.

67W. Wang et al. / European Journal of Pharmacology 568 (2007) 61–67
Habig, W.H., Pabst, M.J., Jakoby, W.B., 1974. Glutathione S-transferases. Thefirst enzymatic step in mercapturic acid formation. J. Biol. Chem. 249,7130–7139.
Kartalou, M., Essigmann, J.M., 2001. Mechanisms of resistance to cisplatin.Mutat. Res. 478, 23–43.
Masters, R.W.J., 2000. Animal Cell Culture: a Practical Approach. OxfordUniversity Press.
Morgan, A.S., Ciaccio, P.J., Tew, K.D., Kauvar, L.M., 1996. Isozyme-specificglutathione S-transferase inhibitors potentiate drug sensitivity in culturedhuman tumor cell lines. Cancer Chemother. Pharmacol. 37, 363–370.
O'Brien, M., Kruh, G.D., Tew, K.D., 2000. The influence of coordinateoverexpression of glutathione phase II detoxification gene products on drugresistance. J. Pharmacol. Exp. Ther. 294, 480–487.
Perez, R.P., 1998. Cellular and molecular determinants of cisplatin resistance.Eur. J. Cancer 34, 1535–1542.
Shen, H., Kauvar, L., Tew, K.D., 1997. Importance of glutathione and associatedenzymes in drug response. Oncol. Res. 9, 295–302.
Tew, K.D., 1994. Glutathione-associated enzymes in anticancer drug resistance.Cancer Res. 54, 4313–4320.
Tew, K.D., Ronai, Z., 1999. GST function in drug and stress response. DrugResist. Updat. 2, 143–147.
Townsend, D., Tew, K., 2003a. Cancer drugs, genetic variation and theglutathione-S-transferase gene family. Am. J. Pharmacogenomics 3,157–172.
Townsend, D.M., Tew, K.D., 2003b. The role of glutathione-S-transferase inanti-cancer drug resistance. Oncogene 22, 7369–7375.
Townsend, D.M., Tew, K.D., Tapiero, H., 2003. The importance of glutathionein human disease. Biomed. Pharmacother. 57, 145–155.
Zheng, J., Mitchell, A.E., Jones, A.D., Hammock, B.D., 1996. Haloenol lactoneis a new isozyme-selective and active site-directed inactivator of glutathioneS-transferase. J. Biol. Chem. 271, 20421–20425.
Zheng, J., Wurz, G.T., Cadman, T.B., Degregorio, M.W., Jones, A.D.,Hammock, B.D., 1997. Haloenol lactone: a new synergist of chemotherapyin vitro. Biochem. Biophys. Res. Commun. 241, 13–17.
Zheng, J., Liu, G., Tozkoparan, B., Wen, D., 2005. Mechanistic studies ofinactivation of glutathione S-transferase Pi isozyme by a haloenol lactonederivative. Med. Chem. 1, 191–198.


















