
Human liver alcohol dehydrogenase: The unique properties of the “atypical” isoenzyme β2β2-Bern...
5
Alcohol. Vol. 2, pp. 47-51, 1985. ©AnkhoInternational Inc. Printed in the U.S.A. 0741-8329/85$3.00 + .00 Human Liver Alcohol Dehydrogenase: The Unique Properties of the "Atypical" Isoenzyme/3z/32-Bern can be Explained by a Single Base Mutation ROLF BOHLER,*t 1 JOHN HEMPEL,t JEAN-PIERRE VON WARTBURG* AND HANS JORNVALLt *Medizinisch-Chemisches Institut der Universitiit Bern, CH-3000 Bern 9, Switzerland and tDepartment of Chemistry I, Karolinska Institutet, S-104 Ol Stockholm, Sweden BUHLER, R., J. HEMPEL, J.-P. VON WARTBURG AND H. JORNVALL. Human liver alcohol dehydrogenase: The unique properties of the "'atypical" isoenzyme fl~flz-Bern can be explained by a single base mutation. ALCOHOL 2(1) 47-51, 1985.--Two allelic variant alcohol dehydrogenase isoenzymes, /3~flz-Bem and/3d3t, coded by the ADH2 locus, were isolated from human livers of Caucasian origin. They represent the "atypical" and "typical" phenotype, respec- tively. ~d3.o-Bernhas a higher specific activity and a lower pH-optimum, has a higher kM for NAD+, is less susceptible to inactivation by iodoacetate, and cannot be activated with chloride ions. In order to define the structural basis for these properties, we determined the amino acid sequence difference between the/3~-Bern and the/31 polypeptide chains. Peptides were prepared by cleavages with trypsin and CNBr, and were purified by exclusion chromatography and reverse phase high performance liquid chromatography. The structural analysis showed that/32-Bern differs at only one position from/3~: Arg-47 in/31 is substituted for His-47 in/32-Bern. This exchange, which is identical to that reported for the/32-Oriental chain, alters the binding of the pyrophosphate group of the coenzyme NAD(H), and also that of iodoacetate, thus explaining the observed differences between flzfl2-Bern and fld3~. Human liver alcohol dehydrogenase Atypical isoenzyme Ailelic variant Amino acid sequence Structure-function relationship Coenzyme binding Inactivation Activation HUMAN liver alcohol dehydrogenase (ADH, E.C.I. 1.1. l) exhibits a large number of pyrazole sensitive (class I) isoen- zymes [23]. Three gene loci, ADHI, ADHz, and AHD3 code for subunits a, 8, and y, respectively, which can combine to the various dimeric isoenzymes. Allelic variants occur at the ADH~ locus which code for/31 and possibly several f12 sub- units [22], and at the ADH:~ locus which code for y~ and y.., [22]. The "atypical" human liver ADH/3.d32-Bern is a variant from the ADH~ locus and can be isolated from livers of Caucasian individuals [25]. Other variants from the ADH~ locus are/3zfl~-Oriental from livers of Oriental origin [12,28] and/32-Indianapolis from livers of African ancestry [2]. The catalytic and functional properties of the "atypical" /32flz-Bern differ significantly from the "typical" /3,/3~. The specific activity is considerably higher and the pH-optimum for the ethanol oxidation is shifted from pH 10.5 to about pH 8.5 [20,21]. In addition, the susceptibility to inhibitors is al- tered [25]. An early report on the primary structure of the fl2-Bern subunit suggested an exchange of Ala-230 in flj for Pro-220 in /32-Bern [21]. Later, it was suggested that/32-Oriental is the end result of changes of two residues resulting in a one posi- tion migration of one of the active site zinc-liganding resi- dues (Cys-46; [28]). However, recently it was shown that fl2-Oriental differs from/3~ by a single Arg/His exchange at position 47 [12]. Structural analysis of/32-Bern [4] showed identity with fl2-Oriental [12] in that it differs from fl, by the same Arg/His mutation at position 47 and not by an Ala/Pro exchange at position 230 as earlier reported [21]. Here, we summarize the isolation, characterization and analysis of the ADH isoenzyme /3d3z-Bern from livers of Caucasian origin, confirming that/32/32-Bern is not only more active at a lower pH, but also that binding of NAD + is weaker. We further show that the "atypical" enzyme is less susceptible to inactivation with iodoacetate, and that it can- not be activated with chloride ions. The structural difference of Arg-47 in/~j for His-47 in/32-Bern affects the general anion binding site that binds the pyrophosphate group of the coen- zyme [7, 15, 29]. NAD + is less tightly bound to the His- containing anion binding site, explaining the higher turnover number, the higher kM for NAD + (since the dissociation of the reduced coenzyme is the rate limiting step), and the ~Requests for reprints should be addressed to Dr. Roif Btihler, Medizinisch-Chemisches Institut der Universitat, Bfihlstrasse 28, CH-3000 Bern 9, Switzerland. 47
Transcript of Human liver alcohol dehydrogenase: The unique properties of the “atypical” isoenzyme β2β2-Bern...

Alcohol. Vol. 2, pp. 47-51, 1985. © Ankho International Inc. Printed in the U.S.A. 0741-8329/85 $3.00 + .00
Human Liver Alcohol Dehydrogenase: The Unique Properties of the "Atypical"
Isoenzyme/3z/32-Bern can be Explained by a Single Base Mutation
R O L F B O H L E R , * t 1 J O H N H E M P E L , t J E A N - P I E R R E V O N W A R T B U R G * A N D H A N S J O R N V A L L t
*Medizinisch-Chemisches Insti tut der Universitiit Bern, CH-3000 Bern 9, Switzerland and tDepar tment o f Chemistry I, Karolinska Institutet, S-104 Ol Stockholm, Sweden
BUHLER, R., J. HEMPEL, J.-P. VON WARTBURG AND H. JORNVALL. Human liver alcohol dehydrogenase: The unique properties of the "'atypical" isoenzyme fl~flz-Bern can be explained by a single base mutation. ALCOHOL 2(1) 47-51, 1985.--Two allelic variant alcohol dehydrogenase isoenzymes, /3~flz-Bem and/3d3t, coded by the ADH2 locus, were isolated from human livers of Caucasian origin. They represent the "atypical" and "typical" phenotype, respec- tively. ~d3.o-Bern has a higher specific activity and a lower pH-optimum, has a higher kM for NAD +, is less susceptible to inactivation by iodoacetate, and cannot be activated with chloride ions. In order to define the structural basis for these properties, we determined the amino acid sequence difference between the/3~-Bern and the/31 polypeptide chains. Peptides were prepared by cleavages with trypsin and CNBr, and were purified by exclusion chromatography and reverse phase high performance liquid chromatography. The structural analysis showed that/32-Bern differs at only one position from/3~: Arg-47 in/31 is substituted for His-47 in/32-Bern. This exchange, which is identical to that reported for the/32-Oriental chain, alters the binding of the pyrophosphate group of the coenzyme NAD(H), and also that of iodoacetate, thus explaining the observed differences between flzfl2-Bern and fld3~.
Human liver alcohol dehydrogenase Atypical isoenzyme Ailelic variant Amino acid sequence Structure-function relationship Coenzyme binding Inactivation Activation
HUMAN liver alcohol dehydrogenase (ADH, E.C.I. 1.1. l) exhibits a large number of pyrazole sensitive (class I) isoen- zymes [23]. Three gene loci, ADHI, ADHz, and AHD3 code for subunits a, 8, and y, respectively, which can combine to the various dimeric isoenzymes. Allelic variants occur at the ADH~ locus which code for/31 and possibly several f12 sub- units [22], and at the ADH:~ locus which code for y~ and y.., [22]. The "atypical" human liver ADH/3.d32-Bern is a variant from the ADH~ locus and can be isolated from livers of Caucasian individuals [25]. Other variants from the ADH~ locus are/3zfl~-Oriental from livers of Oriental origin [12,28] and/32-Indianapolis from livers of African ancestry [2].
The catalytic and functional properties of the "atypical" /32flz-Bern differ significantly from the "typical" /3,/3~. The specific activity is considerably higher and the pH-optimum for the ethanol oxidation is shifted from pH 10.5 to about pH 8.5 [20,21]. In addition, the susceptibility to inhibitors is al- tered [25].
An early report on the primary structure of the fl2-Bern subunit suggested an exchange of Ala-230 in flj for Pro-220 in /32-Bern [21]. Later, it was suggested that/32-Oriental is the
end result of changes of two residues resulting in a one posi- tion migration of one of the active site zinc-liganding resi- dues (Cys-46; [28]). However, recently it was shown that fl2-Oriental differs from/3~ by a single Arg/His exchange at position 47 [12]. Structural analysis of/32-Bern [4] showed identity with fl2-Oriental [12] in that it differs from fl, by the same Arg/His mutation at position 47 and not by an Ala/Pro exchange at position 230 as earlier reported [21].
Here, we summarize the isolation, characterization and analysis of the ADH isoenzyme /3d3z-Bern from livers of Caucasian origin, confirming that/32/32-Bern is not only more active at a lower pH, but also that binding of NAD + is weaker. We further show that the "atypical" enzyme is less susceptible to inactivation with iodoacetate, and that it can- not be activated with chloride ions. The structural difference of Arg-47 in/~j for His-47 in/32-Bern affects the general anion binding site that binds the pyrophosphate group of the coen- zyme [7, 15, 29]. NAD + is less tightly bound to the His- containing anion binding site, explaining the higher turnover number, the higher kM for NAD + (since the dissociation of the reduced coenzyme is the rate limiting step), and the
~Requests for reprints should be addressed to Dr. Roif Btihler, Medizinisch-Chemisches Institut der Universitat, Bfihlstrasse 28, CH-3000 Bern 9, Switzerland.
47
48 BI3HLER ET AL.
TABLE 1
PURIFICATION OF HUMAN LIVER ALCOHOL DEHYDROGENASE ISOENZYMES ~t,8, AND B~-BERN
Total Total Specific protein activity activity Yield
(g) (IU) (IU/mg) (%)
"Typical" liver 1,560.00 - - - - - - Extract 47.80 4,030 0.08 100.0 CapGapp Pool* 1.27 2,744 2.20 68.0 CM-cellulose pool 0.17 69 0.40 1.7
"Atypical" liver 1,370.00 - - - - - - Extract 57.00 14,780 0.26 100.0 CapGapp pool* 0.91 6,300 6.90 43.0 CM-cellulose pool 0.04 416 9.90 2.8
B#32-Bern
*CapGapp pool: pool after double ternary complex affinity chro- matography on 4-[3-(N-6-aminocaproyl)-aminopropyl]pyrazole- Sepharose, containing the mixture of all pyrazole sensitive class I isoenzymes.
lower pH-optimum for ethanol oxidation (His has a lower Pka than Arg). The reduced susceptibility of the "a typ ica l" isoenzyme/3zB2-Bern to inactivation by iodoacetate can also be explained because iodoacetate is directed specifically to its target, Cys-46, by binding to the residue at position 47 [15].
METHOD
Human livers of Caucasian origin were obtained from au- topsies, frozen 10-20 hr after death and stored at -20°C. The isoenzyme /3~/31 was isolated from livers with " ' typical" phenotype and the isoenzyme /32~2-Bern from livers with "a typica l" phenotype. The purification steps included dou- ble ternary complex affinity chromatography followed by CM-cellulose ion exchange chromatography as described [4, 5, 14]. The purity of the isoenzymes was tested by SDS gel electrophoresis and by starch gel electrophoresis [22]. The isoenzymes were identified by their electrophoretic mobilities on starch gel I221. Their subunit composition was verified by monomerization and hybridization with the horse liver isoenzyme EE, followed by starch gel electrophoresis [5,18].
ADH activity was measured photometrically at 340 nm, monitoring the production or utilization of NADH at 25°C. Assay conditions in a total volume of 3 ml were 67 mM glycine-NaOH buffer tpH 10.5 or 8.8), containing 1.6 mM NAD* and 16.7 mM ethanol, when the oxidation reaction was measured or 67 mM sodium phosphate buffer (pH 7.0), 0.1 mM NADH and 5 mM acetaldehyde (freshly distilled), when the reduction reaction was measured. The k~t of NAD ~ was measured by varying the amount of NAD ~ added to the assay mixture while keeping all other parameters constant. The effect of chloride ions was determined at pH 7.0 in 67 mM sodium phosphate buffer in the presence or absence of 50 mM sodium chloride. Activities are expressed as Interna- tional Units (IU) and represent v.mol NADH produced or utilized per rain. Specific activities are expressed as IU/mg protein determined as in [17].
Inactivation by iodoacetate was measured after prein-
TABLE 2 CATALYTIC DIFFERENCES BETWEEN/3~fl~ AND ~#2-BERN
Measured parameter* Unit ,8,,8, /32B.~-Bem
Specific activity IU/mg 0.4 9.9 pH-optimum pH 10.5 8.5 k~t NAD* M 5x10 .... 5x10 Activity in the % 130 90
presence of 50 mM CI 50% inactivation by rain 7 111
0.5 mM iodoacetate (t/2)
*Experimental details are given in the method section.
cubating the isoenzymes in 0.5 ml 20 mM Tris-HCI buffer, pH 7.5, in the presence or absence of 0.5 mM iodoacetate or l mM NAD + at 25°C. The residual enzyme activity of the isoenzymes was determined by removing aliquots and measuring the reductive reaction at pH 7.0.
Tryptic and CNBr peptides of/3t/3~ and fl~2-Bern were prepared after reduction and J*C-carboxymethylation I l l ] , The peptides were fractionated on Sephadex G-50 and further purified by reverse phase high performance liquid chromatography [9]. Manual sequence degradations were performed by the DABITC-Edman method [6] utilizing byproducts to assist identification [24]. Liquid phase se- quencer degradations were performed in a Beckman 890D sequencer with a 0.1 M Quadrol peptide program and with glycine-precycled polybrene as ca rder [13]. Phenylthiohy- dantoin derivatives were analyzed by reverse phase high per- formance liquid chromatography [30], supplemented, for in- dependent residue confirmation, at the positions correspond- ing to the exchanged residues, by amino acid analysis after back hydrolysis in 6 M HCI, containing 0.1% SnCl.,. The presence of '4C-carboxymethylcysteine was monitored by radioactivity measurement.
RESULTS
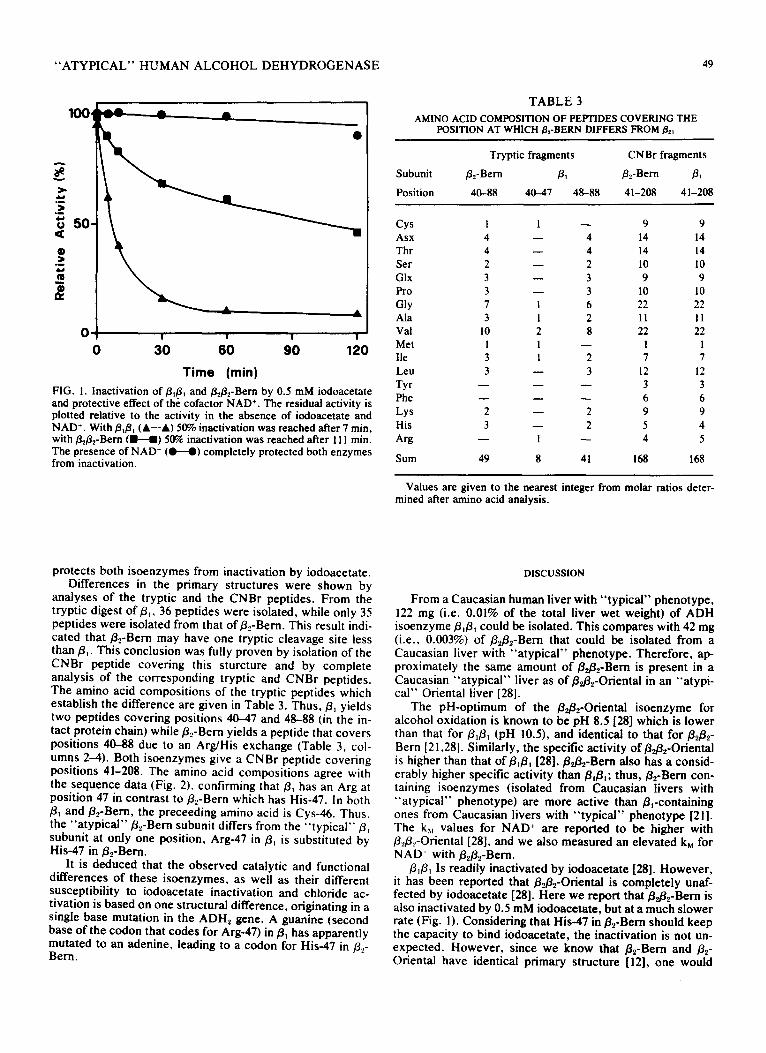
Purification of isoenzyme fld31 from Caucasian human liver with " ' typical" phenotype and of /3~2-Bern from Caucasian liver with "a typ ica l " phenotype resulted in homogeneous enzyme preparations as judged by both SDS- polyacrylamide and starch gel electrophoresis. Based on hybridization experiments, both enzymes were composed of two identical subunits each. Table 1 summarizes the results of the purification. In Table 2, significant characteristics of the two isoenzymes are listed. The specific activity of/32B2- Bern was about 25 times that of/3d3t and was highest at pH 8.5 as compared to pH 10.5 for fl,fl~. The kM value for NAD + also differed between the two isoenzymes. This value was about l0 times higher for /3#~-Bern than for/3j3~. In addi- tion, at pH 7.0, BJ3, was more active in the presence of 50 mM chloride, while/3d3z-Bern remained unaffected or even was slightly inhibited. Figure 1 shows the effect of iodoace- tate on the activity of B#3~ and/3zg2-Bern. The former was readily inactivated in the presence of 0.5 mM iodoacetate (50% inactivation after 7 min) whereas the inactivation of the latter proceeded at a much slower rate (50% inactivation after 111 min). Figure 1 also demonstrates that 1 mM NAD ~
" A T Y P I C A L " H U M A N A L C O H O L DEHYDROGENASE 49
1 0 0 ~ •
"
I I I
0 30 60 90 120
Time (rain) FIG. 1. Inactivation of ~1~1 and/3d32-Bern by 0.5 mM iodoacetate and protective effect of the cofactor NAD +. The residual activity is plotted relative to the activity in the absence of iodoacetate and NAD +. With/31/3t (A--A) 50% inactivation was reached after 7 min, with/3d3~-Bern ( l l - - I ) 50% inactivation was reached after 111 min. The presence of NAD + (O--O) completely protected both enzymes from inactivation.
TABLE 3
AMINO ACID COMPOSITION OF PEPTIDES COVERING THE POSITION AT WHICH/32-BERN DIFFERS FROM/321
Tryptic fragments CNBr fragments
Subunit B.o-Bern fl, fl~-Bern fl,
Position 40-88 40--47 48- -88 41-208 41-208
Cys 1 1 - - 9 9 Asx 4 - - 4 14 14 Thr 4 - - 4 14 14 Ser 2 - - 2 10 10 Glx 3 - - 3 9 9 Pro 3 - - 3 10 10 Gly 7 1 6 22 22 Ala 3 1 2 11 !1 Val 10 2 8 22 22 Met 1 1 - - ! 1 lle 3 1 2 7 7 Leu 3 - - 3 12 12 Tyr - - - - ~ 3 3 Phe - - - - - - 6 6 Lys 2 - - 2 9 9 His 3 - - 2 5 4 Arg - - I ~ 4 5
Sum 49 8 41 168 168
Values are given to the nearest integer from molar ratios deter- mined after amino acid analysis.
protects both isoenzymes from inactivation by iodoacetate. Differences in the primary structures were shown by
analyses of the tryptic and the CNBr peptides. From the tryptic digest o f /3 , 36 peptides were isolated, while only 35 peptides were isolated from that of/3.,-Bern. This result indi- cated that/32-Bern may have one tryptic cleavage site less than/3,. This conclusion was fully proven by isolation of the CNBr peptide covering this sturcture and by complete analysis of the corresponding tryptic and CNBr peptides. The amino acid compositions of the tryptic peptides which establish the difference are given in Table 3. Thus,/3, yields two peptides covering positions 40-47 and 48--88 (in the in- tact protein chain) while/3.,-Bern yields a peptide that covers positions 40-88 due to an Arg/His exchange (Table 3, col- umns 2--4). Both isoenzymes give a CNBr peptide covering positions 41-208. The amino acid compositions agree with the sequence data (Fig. 2). confirming that B, has an Arg at position 47 in contrast to/3~-Bern which has His-47. In both 131 and/3~-Bern, the preceeding amino acid is Cys-46. Thus, the "'atypical"/3..,-Bern subunit differs from the " ' typical" B, subunit at only one position, Arg-47 in/3, is substituted by His-47 in/3.,-Bern.
It is deduced that the observed catalytic and functional differences of these isoenzymes, as well as their different susceptibility to iodoacetate inactivation and chloride ac- tivation is based on one structural difference, originating in a single base mutation in the ADH~ gene. A guanine (second base of the codon that codes for Arg-47) in/31 has apparently mutated to an adenine, leading to a codon for His-47 in/3.,- Bern.
DISCUSSION
From a Caucasian human liver with " typ ica l" phenotype, 122 mg (i.e. 0.01% of the total liver wet weight) of ADH isoenzyme/3,/31 could be isolated. This compares with 42 mg (i.e., 0.003%) of/32/32-Bern that could be isolated from a Caucasian liver with "a typ ica l" phenotype. Therefore, ap- proximately the same amount of/3zB2-Bern is present in a Caucasian "a typ ica l" liver as of/3zB~-Oriental in an "atypi- cal" Oriental liver [28].
The pH-optimum of the flzB2-Oriental isoenzyme for alcohol oxidation is known to be pH 8.5 [28] which is lower than that for/31/3, (pH 10.5), and identical to that for/32f12- Bern 121,28]. Similarly, the specific activity of/3d3.rOriental is higher than that of/3,/3, [28]./3d32-Bern also has a consid- erably higher specific activity than/31/31; thus, B~-Bern con- taining isoenzymes (isolated from Caucasian livers with " 'atypical" phenotype) are more active than /3,-containing ones from Caucasian livers with " typ ica l" phenotype [21]. The k~, values for NAD ÷ are reported to be higher with /3.,_B.,-Oriental [28], and we also measured an elevated kM for NAD ~ with/3d32-Bern.
/3,/3~ Is readily inactivated by iodoacetate [28]. However , it has been reported that/32/3z-Oriental is completely unaf- fected by iodoacetate [28]. Here we report that/3.2flz-Bern is also inactivated by 0.5 mM iodoacetate, but at a much slower rate (Fig. I). Considering that His-47 in/32-Bern should keep the capacity to bind iodoacetate, the inactivation is not un- expected. However, since we know that /32-Bern and /32- Oriental have identical primary structure [12], one would

50 BI ]HLER ET AL .
~o ~5 50 5~
e, - MET - VAt - ALA - '/At GLY - ILE - CYS - ~ - THR - ASP - ASP - HIS - VAL - VAL - SER -
~,-B~rn - MET - VAL - ALA - VAL - GLY - ILE - CYS - HIL~- THR - ASP - ASP - HIS - VAL - VAL - SER -
FIG. 2. Amino acid sequences of/3, and/32-Bern subunit-saround the exchanged amino acid residue (boxed) at position 47.
expect that also/3zS~-Oriental should be slowly inactivated. Other structure differences between the " typ ica l " sub-
unit/3, and the "a typ ica l" allelic variant B2-Bern have been reported earlier, Thus, in/32-Bern, a Pro-230 was reported to substitute Ala-230 in/3, [1]; and in/3z-Oriental, a two-step substitution involving the active site Cys-46 (Cys-46-Arg-47 in r , and His-46-Cys47 in/32-Oriental) was found [28]. It was then believed that fl2-Bern and ~2-Oriental might be different mutations. Later, it was unambiguously shown that/3, has Arg-47 which is replaced by His-47 in/32-Oriental and that Cys-46 is conserved [12]. Here, we show that the sequence reported in [12] also holds true for/32-Bern [10]./32-Bern is identical in primary structure to/32-Oriental but much less frequent [26]. The Ala/Pro exchange in /32-Bern reported earlier [ 1] was not found. Possibly this substitution derives from another subunit (e.g., a or T2) that was contaminating the earlier/32-Bern preparation.
The Arg/I-Iis substitution between B~/BrBern is fully com- patible with the observed differences in catalytic activities. The lower pH-optimum for BnS~-Bern, its hi~aer k~, value for NAD +, and its slower inactivation by iodoacetate are consis- tent with the observed substitution of Arg for His at position 47. Arg-47 in horse liver A D H forms a charge interaction with the pyrophosphate group of the coenzyme [ 15,29]. The lower pk~ value of the imidazole groups of His compared with the guanidino group of Arg is compatible with a lower pH for optimal interaction during coenzyme binding. A His-47 is also compatible with weaker interaction between the imidazole and the pyrophosphate group than between Arg-47 and pyrophosphate. Since the coenzyme dissociation
from the enzyme is the rate limiting step in the ethanol oxi- dation reaction [14], a histidine enzyme form would be ex- pected to have a higher turnover number. A higher turnover number for a His-containing ADH species has also been ob- served in yeast [27]. Thus, the yeast wild type enzyme has a His at the equivalent position, resulting in a turnover number that is much higher than that of ADH isolated from an artifi- cially produced and less active mutant enzyme that turned out to have Arg instead of His at the same position [27]. Also, the inactivator iodoacetate binds to the anion binding site at position 47 in a reversible manner [19,20]. It then is directed to the neighbouring Cys-46 which is specifically and irreversibly alkylated [8, 10, 16]. As is the case with coen- zyme binding, the binding of iodoacetate to the His- containing enzyme is expected to be weaker. The fast inac- tivation of /3,fl, (Arg-47) versus the slow inactivation of /3d32-Bern (His-47) is therefore compatible with this mech- anism.
CONCLUSION
The observed catalytic and functional differences be- tween the "typical"/3, /3j and the "a typ ica l" allelic variant ~ 2 - B e r n , both isolated from human livers of Caucasian origin, are fully explained by the difference in the primary structure of these isoenzymes. Since the codons for Arg and His differ at only the second position of the triplet (guanine for Arg and adenine for His), it is deduced that a one base mutation is likely to be responsible for the differences be- tween/3,/3j (Arg-47) and/32/32-Bern (His-47).
REFERENCES
1. Berger, D., M. Berger and J.-P. yon Wartburg. Structural studies of human liver alcohol-dehydrogenase isoenzymes. Eur J Biochem 50: 215-225, 1974.
2. Bosron, W. F., T. K. Li and B. L. VaUee. New molecular forms of human liver alcohol dehydrogenase: Isolation and charac- terization of ADH-Indianapolis. Proc Natl Acad Sei USA 77: 5784--5788, 1980.
3. Brinden, C.-I., H. Jbrnvall, H. Eklund and B. Furugren. Alco- hol dehydrogenase, In: The Enzymes, voi ! 1, edited by P. D. Boyer. New York: Academic Press, 1975, pp. 103-190.
4. BOlder, R., J. Hempei, J.-P. yon Wartburg and H. Jtrnvail. Atypical human liver alcohol dehydroBenas¢: The ,O2-Beru sub- unit has an amino acid exchange that is identical to the one in the/32-Oriental chain. FEBS Left 173: 360--366, 1984.
5. Biilder, R. and J.-P. yon Wartburg. l)urif'w.ation and substrate specificities of three human liver alcohol dehydrogenase isoen- zymes. FEBS Lett 144: 135-139, 1982.
6. Chang, J. Y., D. Brauer and B. Wittman-Liebold. Micro- sequence analysis of peptides and proteins using 4- NN-dimethylaminoazo-benzene 4'-isothiocyanate/phenylisothio- cyanate double coupling method. FEBS Lett 93: 205-214, 1978.
7. Eklund, H., B. NordstrOm, E. Zeppezauer, G. Stderlund, 1. Ohlsson, T. Boiwe, B.-O. SOderberg, O. Tapia, C.-I. Br~inden and A. Akeson. Three-dimensional structure of horse liver alcohol dehydrogenase at 2.4 A resolution. J Mol Biol 102: 27-59, 1976.
8. Harris, 1. Structure and catalytic activity of alcohol dehydro- genases. Nature 203: 30-34, 1964.
9. Hempel, J., R. BOhler, R. Kaiser, B. Holmquist, C. de Zalenski, J.-P. von Wartburg, B. L. Vallee and H. J6rnvall. Human liver alcohol dehydrogenase: Primary structure of the /3,/3, isoen- zyme. EurJ Biochem 145: 437-445, 1984.
10. J6rnvall, H. Horse liver alcohol dehydrogenase. The primary structure of an N-terminal part of the protein chain of the ethanol active isoenzyme. Eur J Biochem 14: 521-534, 1970.
11. J6rnvall, H. The primary structure of the yeast alcohol dehy- drogenase. Eur J Bioehem 72: 425--442, 1977.
12. J6rnvali, H., J. Hempel, B. L. Vallee, W. F. Bosron and T.-K. Li. Human liver alcohol dehydrogenase: Amino acid substitu- tion in the/3~82-Oriental isozyme explains functional properties, establishes an active site structure, and parallels mutational ex- changes in the yeast enzyme. Proc Natl Aead Sci USA 81: 3024-3028, 1984.

" A T Y P I C A L " H U M A N A L C O H O L D E H Y D R O G E N A S E 51
13. J6rnvall, H. and L. Philipson. Limited proteolysis and a re- active cysteine residue define accessible regions in the native conformation of the adenovirus hexon protein. Eur J Biochem 104: 237-247, 1980.
14. Lange, L. G., A. J. Sytkowski and B. L. VaUee. Human liver alcohol dehydrogenase: Purification, composition and catalytic features. Biochemistry 15: 4687-4693, 1976.
15. Lange, L. G., III, J. F. Riordan, B. L. Vallee and C.-I. Br~in- den. The role of arginyl residues in directing carboxymethyla- tion of horse liver alcohol dehydrogenase. Biochemistry 14: 3497-3502, 1975.
16. Li, T.-K. and B. L. Vallee. Active-center peptides of liver alco- hol dehydrogenase. I. The sequence surrounding the active cys- teinyl residues. Biochemistry 3: 86%873, 1964.
17. Lowry, O. H., N. J. Rosebrough, A. L. Farr and R. J. Randall. Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275, 1951.
18. Lutsdorf, U. M., P. M. Schfirch and J.-P. yon Wartburg. Heterogeneity of horse liver alcohol dehydrogenase. Eur J Biochem 17: 497-508, 1970.
19. Reynolds, C. H. and J. S. McKinley-McKee. Anion-binding to liver alcohol dehydrogenase, studied by rate of alkylation. EurJ Biochem 10: 474-478, 1969.
20. Reynolds, C. H., D. L. Morris and J. S. McKinley-McKee. Complexes of liver alcohol dehydrogenase. Further studies of the rate of inactivation. Eur J Biochem 14: 14-26, 1970.
21 Schenker, T. M., L. J. Teeple and J.-P. yon Wartburg. Heterogeneity and polymorphism of human liver alcohol dehy- drogenase. Eur J Biochem 24: 271-279, 1971.
22. Smith, M., D. A. Hopkinson and H. Harris. Developmental changes and polymorphism in human alcohol dehydrogenase. Ann Hum Genet 34: 251-271, 1971.
23. Strydom, D. J. and B. L. Vallee. Characterization of human alcohol dehydrogenase isoenzymes by high performance liquid chromatographic peptide mapping. Anal Biochem 123: 422--429, 1982.
24. Von Bahr-Lindstr6m, H., J. Hempel and H. J6rnvall. Byproducts as an aid in residue identification during peptide sequence analysis with dimethyl aminobenzene isothiocyanate. J Protein Chem 1: 257-262, 1982.
25. Von Wartburg, J.-P., J. Papenberg and H, Aebi. An atypical human alcohol dehydrogenase. Can J Biochem 43: 88%898, 1965.
26. Von Wartburg, J.-P. and P. M. Schiirch. Atypical human liver alcohol dehydrogenase. Ann N Y Acad Sci 151: 936-956, 1968.
27. Wills, C. and H. J6rnvali. Amino acid substitutions in two func- tional mutants of yeast alcohol dehydrogenase. Nature 279: 734-736, 1979.
28. Yoshida, A., C. C. Impraim and I.-Y. Huang. Enzymatic and structural differences between usual and atypical human liver alcohol dehydrogenase. J Biol Chem 256: 12430-12436, 1981.
29. Zeppezauer, M., H. J6rnvall and I. Ohlsson. Carboxymethyla- tion of horse-liver alcohol dehydrogenase in the crystalline state. The active-site zinc region and general anion-binding site of the enzyme correlated in primary and ternary structures. Eur J Bioehem 58: 95-104, 1975.
30. Zimmermann, C. L., E. Appella and J. J. Pisano. Rapid analysis of amino acid phenylthiohydantoins by high performance liquid chromatography. Anal Biochem 77: 56%573, 1977.










![arXiv:1909.08464v1 [nucl-ex] 18 Sep 2019 · 2019-09-19 · ii) In the quark model [11], the large N branching ratio is explained as a consequence of a dynamical clusterization into](https://static.fdocument.org/doc/165x107/5f8a986ade5074451c6ca2e2/arxiv190908464v1-nucl-ex-18-sep-2019-2019-09-19-ii-in-the-quark-model-11.jpg)







