
Enzyme activity patterns of myosin ATPase, α-glycerophosphate dehydrogenase and succinate...
6
Acta histochem. (lena) 96,213-218 (1994) Gustav Fischer Verlag lena' Stuttgart· New York Acta Enzyme activity patterns of myosin ATPase, a-glycerophosphate dehydrogenase and succinate dehydrogenase within different muscle fibre types Veit Wank I, Reinhard Bauer2, Karla Punkt\ and Joseph Ziegan 4 1 Institute of Sports Sciences, Friedrich-Schiller-University lena, Seidelstr. 20, D-07749 lena, 2 Institute of Pathological Physiology, Friedrich-Schiller-University lena, Lobderstr. 3, D-07740 lena, 3 Institute of Anatomy, University of Leipzig, Liebigstr. 13, D-04103 Leipzig and • Institute of Pathology, Bezirks- krankenhaus fur Psychiatrie, Chemnitzer Str. 50, D-04289 Leipzig Accepted 6 February 1994 Summary Muscle fibre compositions of five different rabbit muscles were determined by combining two enzyme-histochemical reactions (NADH tetracolium oxidoreductase and myosin ATPase after alkaline preincubation). The differentiation into the fibre types, fast twitch glycolytic (FTG), fast twitch oxidative (FTO), and slow twitch oxidative (STO) was possi- ble by a reliable staining classification. Aim of the study was the estimation of enzyme activity patterns within the three dif- ferent fibre types. For this purpose, four serial cross-sections with several enzyme histochemical reactions were performed: alkaline combination method for fibre type determination, the reactions of myosin ATPase, a-glycerophosphate dehydrogenase (GPDH), and succinate dehydrogenase (SDH). The measurement procedure for the estimation of enzyme activities was based on the proportionality between the intensity of the enzyme histochemical staining reaction and the degree of enzyme activity. The ac- tivities of GPDH (indicator for glycolytic metabolism) and SDH (oxidative metabolism) were inverse. The calcium-activated myosin ATPase showed only little activity in slow twitch fibres after alkaline preincubation. In contrast to slow twitch fibres, ATPase ac- tivity in fast twitch fibres was relatively high. The results showed that the classification of muscle fibre types due to their myosin ATPase activities and their main metabolisms (oxidative and glycolytic respectively) is justified. Key words: rabbit - muscle fibre - multiple histochemistry - myosin ATPase, a- glycerophosphate dehydrogenase - succinate dehydrogenase Introduction Skeletal muscles are mainly characterized by their specific contraction qualities. The most important qualities, muscle force and contraction velocity, depend on muscle archi- Correspondence to: V. Wank
Transcript of Enzyme activity patterns of myosin ATPase, α-glycerophosphate dehydrogenase and succinate...

Acta histochem. (lena) 96,213-218 (1994) Gustav Fischer Verlag lena' Stuttgart· New York
Acta hislo~hendca
Enzyme activity patterns of myosin ATPase, a-glycerophosphate dehydrogenase and succinate dehydrogenase within different muscle fibre types
Veit Wank I, Reinhard Bauer2, Karla Punkt\ and Joseph Ziegan4
1 Institute of Sports Sciences, Friedrich-Schiller-University lena, Seidelstr. 20, D-07749 lena, 2 Institute of Pathological Physiology, Friedrich-Schiller-University lena, Lobderstr. 3, D-07740 lena, 3 Institute of Anatomy, University of Leipzig, Liebigstr. 13, D-04103 Leipzig and • Institute of Pathology, Bezirkskrankenhaus fur Psychiatrie, Chemnitzer Str. 50, D-04289 Leipzig
Accepted 6 February 1994
Summary
Muscle fibre compositions of five different rabbit muscles were determined by combining two enzyme-histochemical reactions (NADH tetracolium oxidoreductase and myosin ATPase after alkaline preincubation). The differentiation into the fibre types, fast twitch glycolytic (FTG), fast twitch oxidative (FTO), and slow twitch oxidative (STO) was possible by a reliable staining classification.
Aim of the study was the estimation of enzyme activity patterns within the three different fibre types. For this purpose, four serial cross-sections with several enzyme histochemical reactions were performed: alkaline combination method for fibre type determination, the reactions of myosin ATPase, a-glycerophosphate dehydrogenase (GPDH), and succinate dehydrogenase (SDH). The measurement procedure for the estimation of enzyme activities was based on the proportionality between the intensity of the enzyme histochemical staining reaction and the degree of enzyme activity. The activities of GPDH (indicator for glycolytic metabolism) and SDH (oxidative metabolism) were inverse. The calcium-activated myosin ATPase showed only little activity in slow twitch fibres after alkaline preincubation. In contrast to slow twitch fibres, ATPase activity in fast twitch fibres was relatively high. The results showed that the classification of muscle fibre types due to their myosin ATPase activities and their main metabolisms (oxidative and glycolytic respectively) is justified.
Key words: rabbit - muscle fibre - multiple histochemistry - myosin ATPase, aglycerophosphate dehydrogenase - succinate dehydrogenase
Introduction
Skeletal muscles are mainly characterized by their specific contraction qualities. The most important qualities, muscle force and contraction velocity, depend on muscle archi-
Correspondence to: V. Wank

214 V. Wank et al.
tecture (fibre arrangement), contribution of connective tissue, and way of stimulation (Carlson and Wilkie, 1974). Moreover the velocity of force development and the fatigue characteristics are determined by the muscle fibre composition (Barnard et ai., 1970).
There are several ways for the classification of muscle fibre types. Slow twitch and fast twitch fibres can be distinguished because of their different contraction velocity. By histochemical analysis muscle fibre types can be separated on the basis of specific enzyme activity patterns. Finally, it is possible to determine several fibre types by the structure of heavy or light myosin chains and their molecular weights (Billeter et ai., 1981; Gergely and Samaha, 1989).
For practical reasons, muscle fibre determination is often performed histochemically. Therefore, it is necessary to establish the enzyme activity patterns of myosin ATPase and of a key enzyme of the oxidative or glycolytic cell metabolism. Determination of the fast twitch glycolytic (FTG), fast twitch oxidative (FTO), and slow twitch oxidative (STO) fibre types requires the comparison of at least two different enzyme activity patterns.
In this study the histochemically estimated enzyme activity patterns of myosin ATPase, succinate dehydrogenase (SDH), and a-glycerophosphate dehydrogenase (GPDH) within individual muscle fibres were estimated and compared. It could be shown, that a correct muscle fibre determination with two different enzyme reactions within the same cross-section gives similar results in subsequent single stained cross sections.
Material and Methods
Leg muscles from eight rabbits of both sexes at an age of about one year were investigated. Animals were kept in the animal breeding station of the University of Jena under standardized conditions. After anaesthesia with pentobarbital two specimens of lOx 5 x 5 mm muscle tissue were taken from the centre of the widest diameter of each of the following muscles: m.extensor digitorum longus (EDL), m. tibialis anterior (TA). m. gastrocnemius medialis (OM), m. soleus (SOL), m. flexor digitalis pedis superficialis (FDS).
The specimens were mounted in the way that the parallel muscle fibres run vertically to the cross-sectional plane of the tissue block. Every specimen was fixed with tissue adhesive (tragacantha 5 g/IOO g aqua conservata) in vertical position on a piece of cork. Muscle samples of each of the five investigated muscles were mounted together on one cork for simultaneous cutting. To avoid freezing artifacts the tissue pieces were powdered with talcum before freezing in liquid nitrogen. The cork plates with the frozen specimens were wrapped in aluminium foil and stored in liquid nitrogen. Cross-sections (12 11m) were cut using a cryostat microtome at a temperature of - 22°C.
The muscle fibre types fast twitch glycolytic (FTO), fast twitch oxidative (FTO), and slow twitch oxidative (STO) were determined by an alkaline combination staining method according to Ziegan (1979). This preparation combines two enzyme-histochemical reactions (NADH tetrazolium salt reductase and myosin ATPase after alkaline preincubation at pH 10.4) in succession at the same cross-section.
The calcium-activated myosin ATPase (E.C.3.6.1.3. ATPase) produced brown stain (PbS) according to the method of Meijer and Elias (1977). NADH tetrazolium reductase (NADHTR) generated a blue final reaction product (formazan) according to the protocol of Spannhoff (1967). Using these procedures, the three muscle fibre types could be clearly visualized in the same cross-section. The fibres were specifically stained (FTO golden brown, FTO golden brown with fine blue network, and STO blue).
The fibre patterns of each muscle were determined by counting the numbers of FTO, FTO, and STO fibres m measurmg areas of 500 fibres. The fibre type area per cross section was calculated by a computerassisted digital video image processing system. In addition to the alkaline combination method for the fibre type determination described above, enzyme histochemical reactions of myosin-ATPase, succinate dehydrogenase (E.C.1.3.99.1. SDH), and glycerophosphate dehydrogenase (E.C. 1. 1. 1.49. OPDH) were evaluated on successive cross-sections. The following procedures were applied: The activity of myosin ATPase was demonstrated according to Padykula and Herman (1955). The sections were stained blackbrown (CoS). The succinate dehydrogenase (SDH) activity was demonstrated according to Lojda et al. (1976). The fibres were stained blue (formazan). The sections for ATPase and SDH reaction were

Muscle fibre and multiple histochemistry 215
incubated 20 min at 37°C. The incubation medium for the demonstration of a-glycerophosphate dehydrogenase (GPDH) activity consisted of 0.1 M phosphate buffer, pH 7.4, 0.005 M a
glycerophosphate, sodium salt, 0.010J0 menadione and 0.005 M 2,3-p-nitrophenyl-5-phenyltetracoliumcloride (2,3-p-DNTTC). The incubation time was 20 min at 37°C. The fibres are stained blue (formazan).
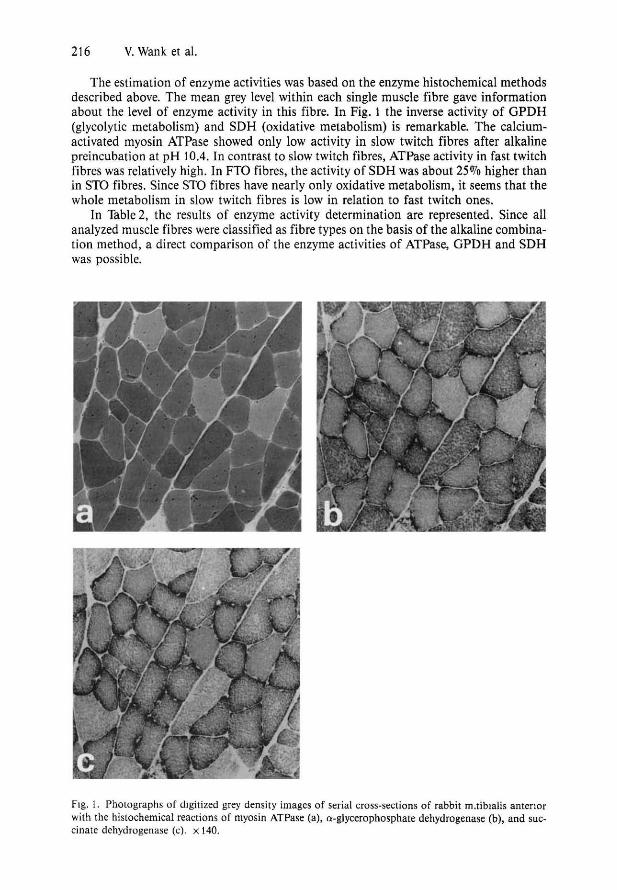
The microscope images of stained cross-sections were taken by a video camera and digitized by a computer-based microscope image analysis system. Photographs of digitized grey density images (serial crosssections with ATPase, GPDH, and SDH enzyme histochemical reactions) are shown in Figure 1.
For analysing the mean enzyme activities within individual muscle fibres, the fibre membranes were marked in the digitized microscope images. The relative enzyme activities could be calculated on the basis of estimation the mean grey densities within every marked fibre. A normalization of the mean grey values of each enzyme was necessary to compare fibres of different muscles. This was accomplished by assigning the absolute minimum and the absolute maximum grey values as 0 and 100 per cent, respectively. The light intensity of the microscope images was scaled between 0 and 255 arbitrary units (S bit) resolution.
For the estimation of the enzyme activities within different fibre types of each of the 40 muscles investigated, four cross-sections were prepared by using different histochemical methods: I) alkaline combination of ATPase and NADHTR reaction on the same cross-section for qualitative muscle fibre type determination, 2) ATPase reaction, 3) GPDH reaction, and 4) SDH reaction (2 - 4 for quantitative estimation of enzyme activities).
In each muscle and in each cross-section the same 25 fibres were analyzed. The selection of the 25 fibres was chosen in such a manner that about one third of each fibre type was represented.
Comparations were made using a t-test. A propability of < 0.05 was assumed to denote significant differences.
Results
The mean fibre type composition of the five investigated muscles of eight rabbits respectively are show in Table 1. The determination of the fibre types was based on the staining reaction of the alkaline combination method according to Ziegan (1979). The difference between the fibre composition of male and female animals was not significant (p < 0.05). The fibre types were homogenously distributed. Merely in sections of m. gastrocnemius medialis (OM), the densities of STO fibres in central regions were found to be higher than in fringe areas.
Table 1. Fibre composition of m.tibialis anterior (TA) , m.extensor digitorum longus (EDL), m.soleus (SOL), m.gastrocnemius medialis (GM), and m.flexor digitalis pedis superficialis (FDS) of eight adult rabbits as determined by a histochemical combination method (Ziegan, 1979). Values are expressed as mean percentages with standard deviations of total fibre surface of the three fibre types (FTG: fast twitch glycolytic, FTO: fast twitch oxidative, STO: slow twitch oxidative). 500 fibers per cross-section were analyzed.
STO FTO FTG
SOL 99.51 0.49 0 (± 1.07) (± 1.07) (0)
GM 25.30 29.45 45.25 (±7.26) (±7.79) (±9.15)
EDL 2.95 20.06 76.99 (± 1.44) (±4.94) (±5.6S)
FDS 9.31 27.14 63.55 (±3.6S) (±9.S7) (± 11.16)
TA S.lO 40.47 51.42 (±2.90) (±S.36) (±9.56)
216 V. Wank et al.
The estimation of enzyme activities was based on the enzyme histochemical methods described above. The mean grey level within each single muscle fibre gave information about the level of enzyme activity in this fibre. In Fig. 1 the inverse activity of GPDH (glycolytic metabolism) and SDH (oxidative metabolism) is remarkable. The calciumactivated myosin ATPase showed only low activity in slow twitch fibres after alkaline preincubation at pH 1004. In contrast to slow twitch fibres, ATPase activity in fast twitch fibres was relatively high. In FTO fibres, the activity of SDH was about 25070 higher than in STO fibres. Since STO fibres have nearly only oxidative metabolism, it seems that the whole metabolism in slow twitch fibres is low in relation to fast twitch ones.
In Table 2, the results of enzyme activity determination are represented. Since all analyzed muscle fibres were classified as fibre types on the basis of the alkaline combination method, a direct comparison of the enzyme activities of ATPase, GPDH and SDH was possible.
Fig. 1. Photographs of digitized grey density images of serial cross-sections of rabbit m.tiblalis antenor with the histochemical reactions of myosin ATPase (a), a-glycerophosphate dehydrogenase (b), and succinate dehydrogenase (e). x 140.
Muscle fibre and multiple histochemistry 217
Table 2. Mean relative enzyme activities with standard deviations of myosin ATPase, a-glycerophosphate dehydrogenase (GPO H), and succinate dehydrogenase (SOH) within different muscle fibre types. The study was performed analysing 25 randomly chosen fibres of each of five rabbit muscles from five animals. Altogether, 625 muscle fibres were investigated: 215 fast twitch glycolytic fibres (FTG) 230 fast twitch oxidative fibres (FTO), and 180 slow twitch oxidative fibres (STO). The determination of muscle fibre types was based on the alkaline combination method of Ziegan (1979). The relative enzyme activities were normalized to the absolute minimum (0) and absolute maximum (100).
ATPase GPOH SOH
FTG fibres 93.6 90.5 14.7 (± 10.7) (±11.1) (±8.1)
FTO fibres 60.7 32.4 88.7 (± 15.2) (±9.1) (± 11.8)
STO fibres 6.3 16.8 64.1 (±4.1) (±8.5) (± 11.1)
Discussion
On the basis of the alkaline enzyme histochemical combination method of Ziegan (1979), the muscle fibre types FTG, FTO, and STO could be determined effectively since only one cross-section is required for analysis.
The calculated standard deviations of the muscle fibre type percentages are in agreement with data of other authors (Bauer et aI., 1987; Brzank et aI., 1986; Henrikson-Larson et aI., 1983; Pieper et aI., 1986). The applied preparation methods to obtain the tissue specimens and the measurement areas in relation to the whole muscle cross-section make more reliable measurement results possible than e. g. analysis of needle biopsies (Bergstrom, 1964; Bauer et aI., 1987; Pongranz, 1976).
The appearance of fast twitch and slow twitch fibres is unequivocally distinguishable. As expected, the differences between FTO and FTG types were not obvious in a few cases. Some muscle fibres had intermediate histochemical qualities between FTO and FTG. It seems that these fibres represent transition forms. In investigations concerning adaptations after strength or endurance training, a shift between FTG and FTO fibre types was observed (Tesch, 1993). There are indications that fast twitch fibres are able to shift their metabolism between oxidative and glycolitic pathways as a result of the training regimen. Similarly, morphological changes are possible (Howald 1982; Pieper et aI., 1986).
The method for the estimation of relative enzyme activities was based on that of Punkt et al. (1984 and 1989), where a histophotometric determination of the extinction rates of the stained myosin-ATPase reaction products in muscle fibres was performed. Punkt et al. (1984) found a significant relationship between the mean extinction rate and the biochemically estimated enzyme activities in muscle tissue.
In contrast to the procedures of Punkt et al. (1984 and 1989), a digital image processing system was used in the present study. Results of estimation of relative enzyme activities on the basis of grey density values measured by a digital video microscope image processing system are comparable with results of histophotometrically estimated relative enzyme activities.
The specific enzyme activities estimated in the present study agree well with the histochemical characterization of the FTG, FTO, and STO fibre types. Apart from the analysis of molecular structures of contractile proteins, the determination of muscle fibre types can also be based on specific enzyme histochemical reactions. The results show that the classification of muscle fibre types using their myosin ATPase activities and their main metabolisms (oxidative or glycolytic) is justified and meaningful.
In addition to the histochemical investigations presented, a series of experiments for

218 V. Wank et al.
biomechanical analysis of the contraction dynamics were carried out by Wank (1993). The comparison of histochemical and biomechanical measurement results confirms the close connections between the muscle metabolism, characterized by the fibre composition, and the velocity of mechanical force development of the muscle.
Acknowledgements
The authors thank Dr. R. M. Abrams (Gainsville, Florida) for his collegial review of the manuscript and his substantial comments.
References
Barnard RJ, Edgerton VR, Furukawa T, and Peter JB (1971) Histochemical, biochemical, and contractile properties of red, white and intermediate fibres. Am J Physiol 220: 410-414
Bauer P, Gabriel MG, Zinner J, and Hassart E (1987) Zur Zuverhissigkeit muskelbioptischer Me13gro13en. Theorie und Praxis Leistungssport 25: 83 - 99
Bergstrom J (1975) Percutaneous needle biopsy of skeletal muscle in physiological and clinical research. Scand J Clin Lab Invest 35: 609 - 616
Billeter R, Heinzmann CW, and Howald H (1981) Analysis of myosin light and heavy chain types in single human skeletal muscle fibers. Eur J Biochem 116: 389 - 395
Brzank KD, Solie G, Pieper KS, and Buhl H (1989) Muskulare Adaptation und Leistungsentwicklung retardlerter und akzelerierter Sportier nach vierwochigem Ausdauertraining. Theorie und Praxis Leistungssport 27: 150 - 161
Carlson FD, and Wilkie RD (1974) Muscle physiology. Prentice-Hall Inc., Englewood Cliffs, New Jersey
Gergely 1., and Samaha F (1989) Biochemistry of muscle and disorders of muscle membranes. In: Siegel GJ et al (Eds) Basic neurochemistry: cellular and medical aspects. Raven Press, New York, pp 609-627
Henrikson-Larson K, Lexell J, and Sjostrom M (1983) Distribution of different fibre types in human skeletal muscles: 1. Method for the preparation and analyses of cross-section of whole tibialis anterior. Histochem J 15: 167 - 178
Howald H (1982) Training induced morphological and functional changes in skeletal muscle. Internat J Sports Med 3: 1 -12
Lojda Z, Gomau R, and Schiebler TH (1976) Enzymhistochemische Methoden. Springer, Berlin, Heidelberg, New York, p 249
Meijer AEFH, and Elias EA (1977) The value of enzyme histochemical techniques in the classification of fibre types of human skeletal muscle. 2. The histochemical demonstrations of myosin adenosine triphosphatase in skeletal muscles from adult patients with no diseases of the neuromuscular system. A comparison between results obtained by calcium salt and lead salt techniques. Histochemistry 52: 45-53
Padykula HA, and Herman E (1955) The specifity of the histochemical method for ATPase. J Histochem Cytochem 3: 170 - 195
Pieper KS, Kriegel M, Bolte B, Ziegan J, and Heilfort U (1986) Zur Ontogenese der Fasertypenverteilung im mensch lichen Skelettmuskel. Mh Vet-Med (Jena) 41: 212-213
Pongratz D (1976) Differentialdiagnose der Erkrankungen der Skelettmuskulatur an Hand von Mmkelbiopsien. Thieme, Stuttgart
Punkt K, Punkt J, Krug H, and Schippel G (1984) Comparison of his to photometric and biochemical myosin ATPase estimations. Histochem J 16: 385 - 387
Punkt K, Erzen J, Krug H, Punkt J, and Seidler E (1989) Histophotometry - the method of choice in quantifying dehydrogenase histochemistry. Acta histochem 87: 63 - 69
Spannhoff L (1967) Einfuhrung in die Praxis der Histochemie. Fischer, Jena Tesch PA (1992) Short· and long-term histochemical and biochemical adaptations in muscle. In: Komi PV
(Ed) Strength and power III sport. Blackwell Scientific Publications, Oxford, pp 239 - 248 Wank V and Gutewort W (1993) Modelling and simulation of muscular contractions with regard to
physiological parameters. Biomechanics, Vol. II, Paris, pp 1450-1451 Ziegan J (1979) Kombinationen enzymhistochemischer Methoden zur Fasertypendifferenzierung und
Beurteilung der Skelettmuskulatur. Acta histochem 65: 34 - 40


![Index [link.springer.com]978-3-319-01008-3/1.pdf · Index β-Hydroxy acyl-CoA dehydrogenase (β-HAD), 117 Álvarez-Sánchez, B., 216, 217 13C labelling, 242, 245, 247 2-Hydroxyisobutyric](https://static.fdocument.org/doc/165x107/5a86029d7f8b9ac96a8cca96/index-link-978-3-319-01008-31pdfindex-hydroxy-acyl-coa-dehydrogenase-had.jpg)















