
Enhancement of performance by methyl β-carboline-3-carboxylate, in learning and memory tasks
6
Brain Resrurch Buktin, Vol. 19, pp. 365-370. 0 Pergamon Journals Ltd., 1987.Printed in the U.S.A. 0361-9230187 $3.00 + .OO Enhancement of Performance by Methyl P-Carboline-3Carboxylate, in Learning and Memory Tasks’ PATRICE VENAULT,2 GEORGES CHAPOUTHIER,* JACQUES SIMIAND,? ROBERT H. DODDi: AND JEAN ROSSIER” “Laboratoire de Physiologie Nerveuse, C.N.R.S., 91190 Gif-sur-Yvette, France tSanof Recherche, 31035 Toulouse, France finstitut de Chimie des Substances Naturelles, C.N.R.S., 91190, Gif-sur-Yvette, France VENAULT, P., G. CHAPOUTHIER, J. SIMIAND, R. H. DODD AND J. ROSSIER. Enhancement of performonce by methyl /3-curboline-3-carboxylate in learning and memory tusks. BRAIN RES BULL 19(3) 365-370, 1987.--- Benzodiazepines are known to induce a profound anterograde amnesia in man. In this report, it is shown that methyl P-carboline-3-carboxylate (P-CCM), an inverse agonist of the benzodiazepine receptor, has the opposite effect; it enhances performance in learning and memory tasks. Three different learning models were used: habituation to a new environment and passive avoidance in mice and imprinting in chicks. The opposite effects of both /3-CCM and the benzodiazepine diazepam were blocked by administration of the benzodiazepine receptor antagonist Ro 15-1788, providing evidence that the benzodiazepine receptor is involved in these effects. Methyl /?-carboline-3-carboxylate Diazepam Learning Memory Ro 15-1788 Mouse Chick BENZODIAZEPINES are widely used anxiolytics and anti- convulsants. Their potent sedative properties are routinely used in presurgical anaesthesia. In the sixties, anaesthetists reported that benzodiazepines induced a strong anterograde amnesia in patients [lo]. Specific substances which can block the effects of benzodiazepines have recently been described [ 1,8]. Interestingly, aside from classical antagonists, some of these substances have intrinsic pharmacological properties that are opposite to those of benzodiazepines; they have been called inverse agonists [2,15]. While benzodiazepines usually have an anticonvulsive activity, inverse agonists have been shown to be proconvulsant or convulsant [ 13,161. Inverse agonists are also anxiogenic [S-7, 12, 171, whereas benzodiazepines are anxiolytic. Since benzodiazepines in- duce anterograde amnesia, we have investigated the possibility that inverse agonists might have an opposite ef- fect and therefore might enhance acquisition (learning) and (or) retention (memory). Here we report that, in three differ- ent animal models, an inverse agonist belonging to the p-carboline group, methyl /3-carboline-3-carboxylate @- CCM), enhances animal performance in three different tasks used to investigate learning and memory. The first model, habituation to a new environment in ro- dents, has been used in the past to demonstrate the amnesic properties of benzodiazepines [18]. The second model, a passive avoidance task, has been widely used for the study of memory processes in rodents. Finally, to investigate the effects of P-CCM in a very different learning situation, a third model, imprinting in chicks, was used [9]. Imprinting, a phenomenon first described by Konrad Lorenz, involves newly hatched birds. Effects of /3-CCM were compared to those of diazepam, a typical benzodiazepine. METHOD Animals Subjects were Swiss male mice (25-30 g) (Iffa-Credo, France, for the habituation study, or Charles River, France, for the passive avoidance task) and newly hatched chicks (Hubbard Strain, Bas Bourray, France). Mice were kept 10 per cage in our animal quarters at 25°C with a day-night cycle of 12-12 hr. Hatching of the chicks was monitored in labora- tory incubators. Once dry (1 to 2 hr after hatching) the chicks were placed in individual boxes (20~20x25 cm) thermostatically regulated at 37°C. The boxes were of opaque Plexiglas, ensuring visual isolation. The glass box- covers provided light with a day-night cycle of 12-12 hr. The chicks were given food and water on the second day after ‘Preliminary reports of this work have been presented at the 1984 European Neuroscience Association Meeting in The Hague [20] and at the 1984 American Society for Neuroscience Meeting in Anaheim, CA 131. bequests for reprints should be addressed to: P. Venault, C.N.R.S., L.P.N. II, 91190, Gif-sur-Yvette, France. 365
Transcript of Enhancement of performance by methyl β-carboline-3-carboxylate, in learning and memory tasks

Brain Resrurch Buktin, Vol. 19, pp. 365-370. 0 Pergamon Journals Ltd., 1987. Printed in the U.S.A. 0361-9230187 $3.00 + .OO
Enhancement of Performance by Methyl P-Carboline-3Carboxylate,
in Learning and Memory Tasks’
PATRICE VENAULT,2 GEORGES CHAPOUTHIER,* JACQUES SIMIAND,? ROBERT H. DODDi: AND JEAN ROSSIER”
“Laboratoire de Physiologie Nerveuse, C.N.R.S., 91190 Gif-sur-Yvette, France tSanof Recherche, 31035 Toulouse, France
finstitut de Chimie des Substances Naturelles, C.N.R.S., 91190, Gif-sur-Yvette, France
VENAULT, P., G. CHAPOUTHIER, J. SIMIAND, R. H. DODD AND J. ROSSIER. Enhancement of performonce by methyl /3-curboline-3-carboxylate in learning and memory tusks. BRAIN RES BULL 19(3) 365-370, 1987.--- Benzodiazepines are known to induce a profound anterograde amnesia in man. In this report, it is shown that methyl P-carboline-3-carboxylate (P-CCM), an inverse agonist of the benzodiazepine receptor, has the opposite effect; it enhances performance in learning and memory tasks. Three different learning models were used: habituation to a new environment and passive avoidance in mice and imprinting in chicks. The opposite effects of both /3-CCM and the benzodiazepine diazepam were blocked by administration of the benzodiazepine receptor antagonist Ro 15-1788, providing evidence that the benzodiazepine receptor is involved in these effects.
Methyl /?-carboline-3-carboxylate Diazepam Learning Memory Ro 15-1788 Mouse Chick
BENZODIAZEPINES are widely used anxiolytics and anti- convulsants. Their potent sedative properties are routinely used in presurgical anaesthesia. In the sixties, anaesthetists reported that benzodiazepines induced a strong anterograde amnesia in patients [lo]. Specific substances which can block the effects of benzodiazepines have recently been described [ 1,8]. Interestingly, aside from classical antagonists, some of these substances have intrinsic pharmacological properties that are opposite to those of benzodiazepines; they have been called inverse agonists [2,15]. While benzodiazepines usually have an anticonvulsive activity, inverse agonists have been shown to be proconvulsant or convulsant [ 13,161. Inverse agonists are also anxiogenic [S-7, 12, 171, whereas benzodiazepines are anxiolytic. Since benzodiazepines in- duce anterograde amnesia, we have investigated the possibility that inverse agonists might have an opposite ef- fect and therefore might enhance acquisition (learning) and (or) retention (memory). Here we report that, in three differ- ent animal models, an inverse agonist belonging to the p-carboline group, methyl /3-carboline-3-carboxylate @- CCM), enhances animal performance in three different tasks used to investigate learning and memory.
The first model, habituation to a new environment in ro- dents, has been used in the past to demonstrate the amnesic
properties of benzodiazepines [18]. The second model, a passive avoidance task, has been widely used for the study of memory processes in rodents. Finally, to investigate the effects of P-CCM in a very different learning situation, a third model, imprinting in chicks, was used [9]. Imprinting, a phenomenon first described by Konrad Lorenz, involves newly hatched birds. Effects of /3-CCM were compared to those of diazepam, a typical benzodiazepine.
METHOD
Animals
Subjects were Swiss male mice (25-30 g) (Iffa-Credo, France, for the habituation study, or Charles River, France, for the passive avoidance task) and newly hatched chicks (Hubbard Strain, Bas Bourray, France). Mice were kept 10 per cage in our animal quarters at 25°C with a day-night cycle of 12-12 hr. Hatching of the chicks was monitored in labora- tory incubators. Once dry (1 to 2 hr after hatching) the chicks were placed in individual boxes (20~20x25 cm) thermostatically regulated at 37°C. The boxes were of opaque Plexiglas, ensuring visual isolation. The glass box- covers provided light with a day-night cycle of 12-12 hr. The chicks were given food and water on the second day after
‘Preliminary reports of this work have been presented at the 1984 European Neuroscience Association Meeting in The Hague [20] and at the 1984 American Society for Neuroscience Meeting in Anaheim, CA 131.
bequests for reprints should be addressed to: P. Venault, C.N.R.S., L.P.N. II, 91190, Gif-sur-Yvette, France.
365

366 VENAULT ET AL.
hatching. Except for the imprinting and test sessions, the chicks were kept in these boxes throughout the experiment.
Drugs
Methyl /?-carboline-3-carboxylate (P-CCM), synthesized by one of us, was dissolved in 0.1 N HCl and diluted to volume with saline. Diazepam, provided by Hoffman-La Roche, Paris (1 mg/ml solution), was diluted to volume in saline. Ro 151788, kindly provided by Dr. Haefely (Hoffman-La Roche, Basel, Switzerland), was suspended in saline with a drop of Tween 80. Drugs were administered subcutaneously (SC) or intraperitoneally (IP) in a volume of 0.05 ml/l0 g (body weight).
Habituation to a New Environment Model
In the habituation model, food-deprived mice are placed in an unfamiliar environment (a different cage) with food ad lib. During this training session, they explore the new cage, but eat only a small quantity. When placed in the same en- vironment for a test session a few days later, the same food- deprived animals eat much more than during the training session. This increase in food intake is assumed to indicate that, during the training session, the animals habituate them- selves to the new environment and that during the test ses- sion, they remember it.
In the present experiment, mice food-deprived for 24 hr were injected SC with /3-CCM (0.2, 0.3, 0.4 and 0.6 mg/kg) diazepam (1 and 1.5 mg/kg) or saline. Fifteen min later mice were transferred to the experimental room and placed indi- vidually for 30 set in an unfamiliar metallic cage (32x 24x 15 cm) containing their usual food. During this very short period, food intake was practically nil. Following this train- ing session mice were returned to their polystyrene home cage in the animal quarters, with no food. Food was provided ad lib at the end of the day. Three days later mice were once more deprived of food. After fasting for 24 hr mice were again placed in the metallic cage for a 2 min test session. Food intake was measured. All groups consisted of 10 mice except for the group diazepam 1 mg/kg where 20 mice were used.
Passive Avoidance Model
In the passive avoidance model, mice are trained in a single-trial procedure. The animals are placed in an illumi- nated box connected to a dark box. Entrance into the dark box usually occurs within 30 set and is punished by an elec- tric footshock. On the next day (test session) the same mice are placed again in the illuminated box. Mice staying in the illuminated box for more than 60 set are considered as re- membering the task. Retention can thus be quantified by the percentage of animals avoiding the dark compartment.
In the present experiment, mice were injected with saline (SC or IP), /3-CCM (0.1, 0.2, 0.3 and 0.4 mg/kg SC) or diazepam (1, 2 or 4 mg/kg IP). Fifteen minutes later (saline, /3-CCM) or 30 min later (saline, diazepam), they were placed in an illuminated box (10x 10x 12 cm) connected to a large dark box (23 x 16x 12 cm) having an electrified metallic grid floor. When the mouse stepped into the dark box, a foot- shock was delivered to its paws. In order to obtain clearcut effects we decided to use two different situations: In experi- ments with /3-CCM, where facilitation of retention was in- vestigated, a low shock (0.1 mA for 2 set, unscrambled DC current) was delivered. In experiments with diazepam where
impairment of retention was investigated, a high shock (0.6 mA for 2 set, unscrambled DC current) was delivered. Dur- ing the test session 24 hr later, mice were again placed in the lit box. Mice avoiding the dark compartment for over 60 set were considered as remembering the task. In the /3-CCM treated groups the number of subjects was 20 per group, except for controls 0.2 mg/kg where this number was 40. In the diazepam treated groups and their controls, each group contained 12 subjects.
Imprinting Model
For the imprinting experiments newly hatched chicks (around 24 hr old) are used. Chicks tend to follow a moving decoy presented to them during the imprinting session. When retested in the same situation 24 hr later, imprinted chicks, having memorized the characteristics of the situation and of the decoy, follow the decoy for a longer time than non-imprinted chicks.
In the present experiment, the imprinting apparatus was a circular arena 200 cm in diameter. The stimulus object was a red rubber ball with yellow stripes, 11 cm in diameter, mov- ing at a speed of 385 cm/min and suspended 40 cm from the edge of the arena and 5 cm above floor level. The walls and floor of the enclosed arena were matt black and lit by three 150 W bulbs 2 m off the ground. The stimulus object moved continuously for periods of 16 set with alternating rest periods of 4 set duration, so that well-imprinted chicks were not overtaxed physically. The chicks were observed on a closed-circuit TV screen and their movements, walking towards the ball or standing near it, were automatically re- corded. Recording was based on the detection of the white spot made by the chick on the black background in the video signal produced by the TV camera. The camera was placed vertically 2 m above the center of the arena. The decoy was suspended from a horizontal arm firmly attached to the cam- era. The entire apparatus rotated simultaneously so that the ball always occupied the same position on the TV screen.
An electronic device defined several rectangular zones in the video field surrounding the image of the ball on the screen. There were two main zones. The central zone sur- rounded the ball, and the white spot produced by the chick’s presence was recorded regardless of the movement of the animal or the decoy. A wider zone enveloped the central zone and this was made up of two sub-zones-one in front of the ball and one behind with reference to the direction of rotation of the ball. In these sub-zones, only the movements of the chicks towards the ball were recorded.
The electronic detection technique was based on the presence of the white spot made by the chick during the scanning of each frame. When this spot passed through the subzones at the speed of the decoy, the movement was not recorded because it represents the movement of the decoy alone, the chick remaining motionless. The recording device printed out for each successive minute the total duration of the chick’s following behavior [9].
Chicks were injected IP with saline, P-CCM (2.5 mg/kg) or diazepam (0.25 mg/kg), 5 min before the imprinting ses- sion. After a delay of one minute, the stimulus object began moving and the chick’s behavior was recorded for 5 minutes. Twenty-four hr later the same chicks were retested in the same situation during a 5 min test session, also preceded by a one-minute adaptation period. Number of subjects per group was: /3-CCM treated and their controls (N= 17, N= 18);’ di- azepam treated and their controls (N=39, N=38).
/3-CARBOLINE ENHANCES PERFORMANCE 367
% E - +60 2 nr +40 E
Jj +20 0
c 0 ._
3 -20 $
E ._
q o- CCM !I diazepam
doses (mg/kg)
FIG. 1. Effect of j3-CCM and diazepam on retention of habituation to a new environment in mice. Figure illustrates differences tstan- dard error in food intake of drug-treated groups as compared to saline groups. Hence, ordinate 0 corresponds to the mean of the food intake of saline treated animals during the 2 min of the test (25r4 mg, n=60). kpcO.05; *+p<O.Ol (analysis of variance). (12) m diazepam
.
(12) .* -I
Statistical Analysis
Analysis of variance was used in the habituation to a new environment model and in the imprinting. Chi square with Yates’ correction allowed us to compare the percentages of the passive avoidance model. 0 0 1 2 4 doses W/kg)
RESULTS
Habituation to a New Environment Model
When /3-CCM (0.2 and 0.3 mg/kg SC) was administered to mice before the training session, food intake during the test session was found to be enhanced in comparison with that of controls [for /3-CCM 0.2 mg/kg, F(1,18)=4.16, ~~0.05; for P-CCM 0.3 mgikg, F(1,18)=17,97,p~0.01]. While the aver- age consumption of the controls was 25 mg during the 2 min test session, the P-CCM treated subjects consumed 40 to 60 mg more (Fig. 1). This effect of @CCM disappeared when higher doses (0.4 and 0.6 mg/kg SC), closer to the dose of 1 mg/kg SC, found [17] to be in the anxiogenic dose range, were used. On the other hand, when diazepam f 1.0 and 1.5 mg/kg SC) was administered before the training session, a reduction in food intake was seen during the test session [for diazepam 1.0 mglkg, F(l,38)=4.18,~<0.05; for diazepam 1.5 mgikg, F(1,18)=5.88, p<O.O5].
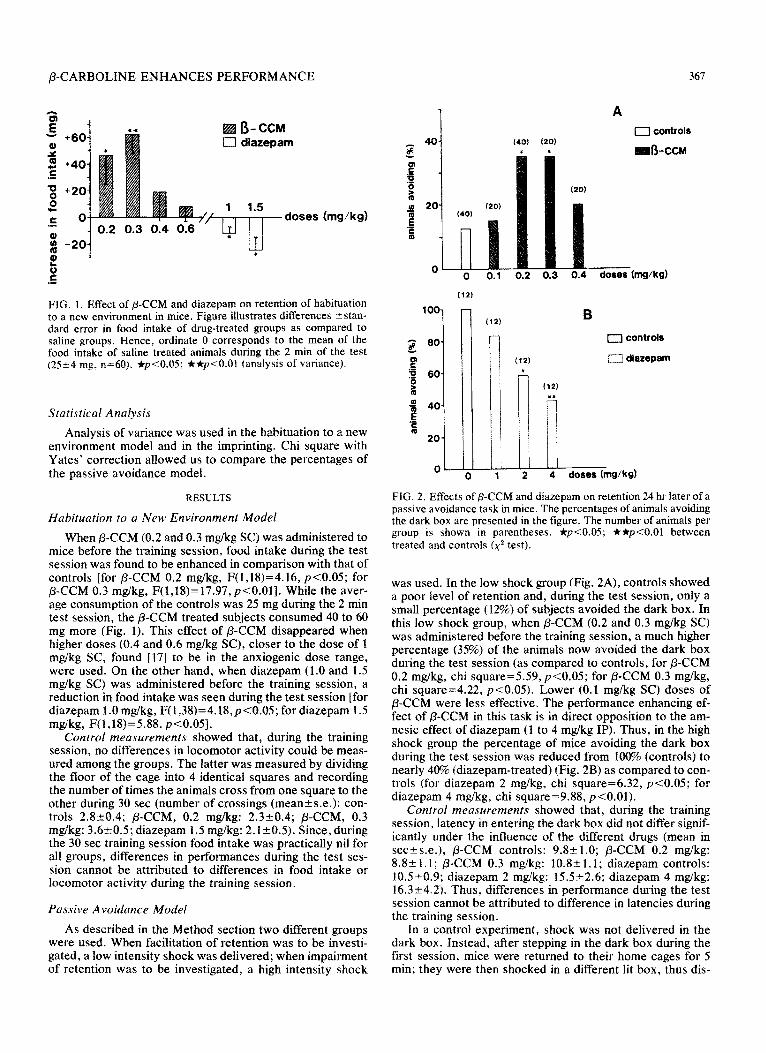
FIG. 2. Effects of P-CCM and diazepam on retention 24 hr later of a passive avoidance task in mice. The percentages of animals avoiding the dark box are presented in the figure. The number of animals per group is shown in parentheses. *p<O.O5; **p<O.Ol between treated and controls (x2 test).
Control measurements showed that, during the training session, no differences in locomotor activity could be meas- ured among the groups. The latter was measured by dividing the floor of the cage into 4 identical squares and recording the number of times the animals cross from one square to the other during 30 set (number of crossings (mean?s.e.): con- trols 2.820.4; @CCM, 0.2 mg/kg: 2.3eO.4; /3-CCM, 0.3 mglkg: 3.6r0.5; diazepam 1.5 mg/kg: 2.1~0.5). Since, during the 30 set training session food intake was practically nil for all groups, differences in performances during the test ses- sion cannot be attributed to differences in food intake or locomotor activity during the training session.
was used. In the low shock group (Fig. ZA), controls showed a poor level of retention and, during the test session, only a small percentage (12%) of subjects avoided the dark box. In this low shock group, when P-CCM (0.2 and 0.3 mgikg SC) was administered before the training session, a much higher percentage (35%) of the animals now avoided the dark box during the test session (as compared to controls, for /3-CCM 0.2 mg/kg, chi square=5.59, ~~0.05; for /3-CCM 0.3 mg/kg, chi square=4.22, ~~0.05). Lower (0.1 mg/kg SC) doses of ,&CCM were less effective. The performance enhancing ef- fect of /3-CCM in this task is in direct opposition to the am- nesic effect of diazepam (1 to 4 mg/kg IP). Thus, in the high shock group the percentage of mice avoiding the dark box during the test session was reduced from 100% (controls) to nearly 40% (diazepam-treated) (Fig. 2B) as compared to con- trols (for diazepam 2 mgikg, chi squarez6.32, ~~0.05; for diazepam 4 mg/kg, chi square=9.88, p<O,Ol).
Passive Avoidance Model
Control measurements showed that, during the training session, latency in entering the dark box did not differ signif- icantly under the influence of the different drugs (mean in sec?s.e.), P-CCM controls: 9.X51.0; /3-CCM 0.2 mg/kg: 8.8-c 1.1; P-CCM 0.3 mg/kg: 10.82 1.1; diazepam controls: lOS+O.P; diazepam 2 mglkg: 15,5t2.6; diazepam 4 mglkg: 16.3~4.2). Thus, differences in performance during the test session cannot be attributed to difference in latencies during the training session.
As described in the Method section two different groups In a control experiment, shock was not delivered in the were used. When facilitation of retention was to be investi- dark box. Instead, after stepping in the dark box during the gated, a low intensity shock was delivered; when impairment first session, mice were returned to their home cages for 5 of retention was to be investigated, a high intensity shock min; they were then shocked in a different lit box, thus dis-
A
0 controls
l3-CCM
0.1 0.2 0.3 0.4 doses bw/kg)
(12)
loo- - (12)
B
0 controls

368 VENAULT ET AL.
l3-CCM
c” +20 Cl diazepam
._ 3- 0 3 =o>, +lO
- 0 .E,O 0.25 0
@Q :a a, 5 -10
FIG. 3. Effect of P-CCM and diazepam on retention of imprinting in chicks. Figure iIIustrates differences ? standard error in time drug- treated chicks spent following the ball as compared to saline treated groups. Hence, ordinate 0 corresponds to the mean of the time spent following the ball by saline-treated chicks (5428 set, N=56). *p<O.O25 between treated and controls (analysis of variance).
sociating dark box from punishment. Twenty-four hr later, during the test session, no difference was observed between saline and treated subjects. Percentages of animals (N=15 per group) avoiding the dark box were saline: 8%; fi-CCM 0.2 mg/kg: 0%; /3-CCM 0.3 mg/kg: 17%. This control experi- ment rules out a non-specific effect of /3-CCM on avoidance of the dark box.
Imprinting Model
When imprinting was accomplished under the effect of /3-CCM (2.5 mg/kg SC) the time the chick spent following the decoy during the test session was increased as compared to non-treated controls (Fig. 3), F(1,33)=6.19, ~~0.025. Thus, in the 5 min period of the test session, controls followed the ball for an average of 53 set whereas P-CCM treated chicks followed it for about 20 set more. On the other hand, when imprinting was performed under the effect of diazepam (0.25 mg/kg SC) the time spent following the ball during the test session was decreased; treated subjects followed the ball for about 20 set less than controls, F(1,75)=4.%, pcO.025. It must be noted here that other doses (P-CCM 1.5 mg/kg; di- azepam 0.5, 1 mg/kg) were tried in the chick with similar though less clearcut results. The present results were ob- tained with a dose of P-CCM (2.5 mgjkg) ten times higher than the dose of diazepam (0.25 mg/kg).
Control measurements showed that during this 5 min im- printing session, the average time the chick spent following the ball was 2423 set with no significant difference between treated subjects and their controls (increase in following be- havior (set) as compared to controls fs.e.: P-CCM 1.88~6.22; diazepam 6.0426.47). Thus, differences in performance dur- ing the test session cannot be attributed to differences in performance during the training session.
DISCUSSION
The results of these three different animal models show that j?-CCM, administered before acquisition, enhances the performance of the task during the retention test. We ver- ified that such an effect cannot be obtained when the drug is administered immediately after acquisition nor before the retention test. For instance, in the study of habituation to a
TABLE I REVERSAL BY Ro 15-1788 OF THE EFFECTS OF P-CCM AND
DIAZEPAM ON PASSIVE AVOIDANCE IN MICE
Drug Injected Animals Avoiding (%)
Low Shock Group Saline 17.5 Ro 15-1788 20 /3-CCM 40* P-CCM + Ro 15-1788 1st
High Shock Group
Saline 89 Ro 15-1788 89 Diazepam l1.t Diazepam + Ro 15-1788 956
*Chi square=3.84, p<O.O5 as compared to saline treated. tChi square=23.61, p<O.OOl as compared to saline treated. $Chi square=9.60, p<O.Ol as compared to subjects treated with
P-CCM alone. Khi square=27.02, p<O.OOl as compared to subjects treated with
diazepam alone.
new environment, when P-CCM (0.3 mg/kg) was adminis- tered just after the training session, no significant difference in food intake of these animals during the test session was seen as compared to the controls. Similarly, if /3-CCM (0.3 mg/kg) was administered 15 min before the test session, the difference in intake of the treated group as compared to that of the controls remained non-significant, We would like, however, to mention other possible alternative interpreta- tions of our results.
Possible Effects on Food Consumption
The effects on the novel environment could find an alter- native explanation in the effects of benzodiazepine receptor ligands on food consumption. The work by Cooper et al. [4] among others provides strong evidence that benzodiazepine inverse agonists consistently induce anorexic effects and that this reduction of food consumption is dose-dependent. In contrast, benzodiazepine receptor agonists produce a large dose-dependent increase of food consumption. Though Cooper’s work involves non-deprived rats fed with a highly palatable food his results could be used as a possible expla- nation of the opposite effects of /3-CCM and diazepam in the habituation to a new environment situation. It can be noticed however that we have chosen a very short training session (30 set) in order to avoid these effects of food consumption during the training session and that our test session is four days later, when the animals are no longer under the infIu- ence of the drugs. Furthermore, in the case of the effects of P-CCM, there is no clear-cut dose-dependent relation. Thus, in our fist model only, the effects on food consumption, though they may play a role that further research could de- tect, cannot account for all the data provided here.
SpeciJicity of the Chick Model
It was interesting to co&ii the effects of fi-CCM and diazepam, obtained in mice, in a different animal species, the chick. It should however be pointed out that many differ- ences in drug response exist between mice and chicks. We

P-CARBOLINE ENHANCES PERFORMANCE 369
TABLE 2
EFFECTS OF @CCM AND DIAZEPAM ON PERFORMANCE DURING A PASSIVE AVOIDANCE TEST 30 SEC AFTER TRAKNING
Drug Injected Animals Avoiding (%)
Low Shock Group Saline 10
P-CCM (0.3 mglkg) 400*
High Shock Group
Saline 87 Diazepam (2 mgikg) 27t
*Chi square=4.80, p<O.OS. iChi square=11.00,p~0.001 between treated and controls.
have already noticed earlier that the dose ratio between P-CCM and diazepam is inverse: in mice we obtained effects opposite to those of /3-CCM 0.3 mg/kg with higher doses of diazepam (1 to 4 mg/kg) whereas in chicks, we obtained the same opposition of effects with a dose of P-CCM (2.5 mgikg) ten times higher than that of diazepam (0.25 mgikg). A lower dose of P-KM (1.5 mgikg) and higher dose of diazepam (OS-1 mgikg) were not effective. This may suggest differ- ences in the drugs’ metabolism and/or mode of action. Little is known of the effects of P-CCM on anxiety in chicks, thus we cannot, at the present time, discuss this point. But we can emphasize another important difference in the effects of @-CCM in chicks. Recent work in our laboratory 1191 has provided evidence that, in the chick, /3-CCM acts both as an inverse agonist (it enhances the convulsive action of pen- tylenetetrazol) and as an agonist. As an agonist, high doses of /3-CCM exert a very powerful sedative action, similar to that of diazepam. A synergism between subliminal doses of diazepam and /3-CCM can also be found for this sedative effect. Though these data make the inte~retation of the ef- fects of /3-CCM imprinting more difficult, it is interesting to note that, in the chick, the opposing effects on imprinting performance are obtained with molecules which are both sedative.
Rel~t~~~ Wit~z the Anxiogenic Action of /MXM
In mice, where anxiogenic and convulsive effects of P-CCM as well as anxiolytic and anticonvulsive effects of diazepam have been extensively studied, we can notice that the performance enhancing effects of P-CCM were not seen in the dose range of the anxiogenic or convulsant effects of this drug, since the optimum dose in the present study (0.2, 0.3 m&g) is much lower than the anxiogenic dose in a con- flict model (1 mglkg) [17] or the convulsive doses (I-10 mg/kg) [ 161, in the same strain of mice. It might however be assumed that the doses used in the present work, insufficient to produce noticeable effects with such classical techniques as conflict models, are still sufficient to act on an emotional component of performance. This explanation is not con- tradictory to an effect on learning and memory processes. It simply assumes that both the improvement of performance in learning and memory tasks and the effects in a conflict situation derive from a common mechanism involving anx- iety. Such an hypothesis should be submitted to further ex- periments in the future.
To investigate whether these effects of &CCM and di- azepam are mediated by the benzodiazepine receptor, we tried to block them using the specific benzodiazepine antagonist, Ro 15-1788 1181. This experiment was under- taken in the passive avoidance situation described earlier and with the same protocol. Thus, low (0.1 mA) and high (0.6 mA) shock groups were used to study the effects of /3-CCM and diazepam, respectively. Groups of mice (N=40 for the low shock groups; N=19 for the high shock groups) were injected before the first session with saline, JKCM (0.3 mgikg SC), diazepam (4 mg/kg IP) or Ro 15-1’788 (15 mg/kg IP) or a combination of /3-CCM + Ro 1.5-1788 or of diazepam + Ro 15-1788. Ro 15-1788 was administered 1.5 min before the first session; the remaining injections were performed as described earlier. Results (Table 1) show that Ro 15-1788 had no effect when administered alone. However, the effects of /XCM or diazepam were completely antagonized by co- administration of Ro 15-1788. This indicates that both the effect of fl-CCM and the opposite effect of diazepam are mediated by the benzodiazepine receptor.
Effect on Acquisition
If we consider that /3-CCM indeed acts on some stage of learning or memory processing, the open question now is to delineate which stages of learning or memory are affected by the drug. If memory can be defined by the retention of in- formation, this information needs to be acquired (learning) and then recalled (retrieval). Since in our experiments, drugs have no effect when administered before the retention test, we can rule out an action on retrieval. This is in agreement with most results obtained with benzodiazepines in human subjects [lo]. A possible explanation of the data would be that /3-CCM increases the level of arousal during the training session. This explanation would concord with the observa- tion that, in rodents, arousal-enhancing drugs improve leam- ing [ 111 and that several /3-carbolines increase arousal [ 141. This interpretation is, however, difficult in the case of chicks where it has been found that, on the contrary, /3-CCM has a sedative effect. In mice, we tried to confirm this arousal- enhancing interpretation with a last experiment using the passive avoidance situation described earlier, the only difference being that the test session was performed 30 set after the end of the training session instead of 24 hr later. High shock groups consisted of 20 subjects and low shock groups of 15 subjects. The data presented in Table 2 show that the results were the same as with a 24 hr interval suggesting an effect on learning. An alternative explanation could be that the drugs act on very short term (30 see) mem- ory processing. This however seems to be ruled out by the fact that no effect was ever obtained when the drugs were administered just after the training session. However, this important question needs further analysis because of the known effect of benzodiazepines on short term memory in man.
Finally, while the fact that we find similar effects in three very different behavioral situations suggests that /3-CCM and diazepam act on learning, the precise mechanisms underly- ing these effects still need to be analysed more closely. Al- ternative explanations could as well be given as partial in- terpretation of the effects observed. It is hoped that further research will shed new light on the results we obtained.

370 VENAULT ET AL.
ACKNOWLEDGEMENTS
This work was supported by C.N.R.S. (A.T.P. Pharmacologic des Neuromediateurs and A.S.P-P.I.R.M.E.D. Neurodrogues), I.N.S.E.R.M. (P.R.C. 121022) and by D.R.E.T. 841039. We would like to thank Dr. P. Potier and Dr. C. Ouannes for valuable help and discussion and Josette Guitard and Jean-Paul Bouillot for technical assistance.
REFERENCES
Braestrup, C., M. Nielsen and C. E. Olsen. Urinary and brain @-carboline-3-carboxylates as potent inhibitors of brain ben- zodiazepine receptors. Froc Nntl Acad Sci USA 77: 2288-2292, 1980. Braestrup, C., R. Schmiechen, G. Neef, M. Nielsen and E. N. Petersen. Interaction of convulsive ligands with benzodiazepine receptors. Science 216: 1241-1243, 1982. Chapouthier, G., P. Venault, L. Prado de Carvalho, J. Simiand and J. Rossier. Possible effects of /I-carbolines on memory. Sot Nearosci Abstr 10: 647, 1984. Cooper, S. J., D. J. Barber, D. B. Gilbert and W. R. Moore% Benzodiazepine receptor ligands and consumption of a highly palatable diet in non-deprived male rats. Psychopharmacology (Berlin) 86: 34%355, 1985.
5. Corda, M. G., W. D. Blaker, W. B. Mendelson, A. Guidotti and E. Costa. &carbolines enhance shock-induced suppression of drinking in rats. Proc Natt Acud Sci USA 80: 2072-2076, 1983.
6. Dorow, R., R. Horowski, G. Paschelke, M. Amin and C. Brae- strup. Severe anxiety induced by FG 7142, a p-carboline ligand for benzodiazepine receptors. Lancer 9: 98-99, 1983.
7. File, S. E., R. G. Lister and D. J. Nutt. The anxiogenic action of benzodiazepine antagonists. Neuropharmacology 21: tO33- 1037, 1982.
8. Hunkeler, W., H. Mohler, L. Pieri, P. Pole, E. P. Bonetti, R. Cumin, R. Schaffner and W. Haefely. Selective antagonists of benzodiazepines. Nature 290: 514-516, 1981.
9. Lecanuet, J. P., T. Alexinsky and G. Chapouthier. The follow- ing response in chicks: conditions for the resistance of consoli- dation to a disruptive agent. Behav Biol 16: 291-304, 1976.
10. Lister, R. G. The amnesic action of benzodiazepines in man. Neurosci Biobehav Rev 9: 87-94, 1985.
II. Martinez, J. L., Jr., R. A. Jensen and J. L. MC Gaugh. Facihta- tion of memory consolidation. in: The Physiological Basis oj Memory, 2nd edition, Chapter 2, edited by J. A. Deutsch. New York: Academic Press, 1983, pp. 49-70.
12.
13.
14.
1.5.
16.
17.
18.
19.
20.
Ninan, P. T., T. M. Insel, R. M. Cohen, J. M. Cook, P. Skolnick and S. M. Paul. Benzodiazepine receptor-mediated experi- mental “anxiety” in primates. Science 218: 1322-1334, 1982. Oakley, N. R. and B. J. Jones. Differential pha~acological effects of P-carboline-3-carboxylic acid esters. Neurophar- macology 21: 587-589, 1982. Ongini, E., C. Barzaghi and M. Marzanatti. Intrinsic and antagonistic effects of P-carboline FG 7142 on behavioral and EEG actions of benzodiazepines and pentobarbital in cats. EwJ Pharmacol95: 125-129, 1983. Pole, P., E. P. Bonetti, R. Schaffner and W. Haefely. A three- state model of the benzodiazepine receptor explains the inter- actions between the benzodiazepine antagonist Ro 15-1788, benzodiazepine tranquilizers, fl-carbolines and phenobarbitone. Naunyn Schmiedebergs Arch Pharmacol321: 260-264, 1982. Prado de Carvalho, L., G. Grecksch, E. A. Cavalheiro, R. H. Dodd, G. Cha~uthier and J. Rossier. Characte~~tion of con- vulsions induced by methyl-~-carboline-3-carboxylate in mice. Eur J Pharmncol 103: 287-293, 1984. Prado de Carvalho, L., G. Grecksch, G. Chapouthier and J. Rossier. Anxiogenic and non-anxiogenic benzodiazepine antagonists. Nature 301: 64-66, 1983. So&&e, P., P. Simon and J. R. Boissier. Amnesic induite chez le rat par les benz~~az~p~es lors dune situation ne faisant pas intervenir de composante nociceptive. C R Acad Sci ID] Paris 283: 203-205, 1976. Venault, P., L. Prado de Carvalho, C. L. Brown, R. H. Dodd, J. Rossier and G. Chapouthier. The benzodiazepine receptor ligand methyl fi-carboline-3-carboxylate is both sedative and oroconvulsant in chicks. Life Sci 39: 1093-1100, 1986. Venault, P., L. Prado de darvalho, J. Simiand, J. Rossier and G. Chapouthier. Effects of &carbolines on memory. Neurosci Lett [Suppl] 18: 119, 1984.






![arXiv:1711.01094v3 [cs.CV] 20 Mar 2018 · tation of cardiac SSFP images in an end-to-end di eren-tiable CNN framework, allowing for thelocalization, align-ment, and segmentation tasks](https://static.fdocument.org/doc/165x107/5fd1f014b0ebb801ad4c6f49/arxiv171101094v3-cscv-20-mar-2018-tation-of-cardiac-ssfp-images-in-an-end-to-end.jpg)






![Technische Universität Chemnitz, Center for ...Preparation of aspheric copper nanoparticles Scheme 1: Synthesis of copper nanoparticles by thermolysis of copper(I) carboxylate 1 [7].](https://static.fdocument.org/doc/165x107/60fcc6b8e53c32273d090db6/technische-universitt-chemnitz-center-for-preparation-of-aspheric-copper.jpg)




