The hydrolytic activity of salivary amylase on starch CLS 331 Experiment No.1.

4057
INTRODUCTION
Limitations to efficient production of meat and milk by cattle are often associated with dietary re-strictions in NE to support productive purposes (Lofgreen and Garrett, 1968; Fox et al., 1992; NRC, 1996). Utilization of dietary energy from α-glucosides (starch) is potentially limited by starch assimilation from the alimentary tract in cattle, and a precise under-
standing of digestion of carbohydrates in the small in-testine of cattle remains equivocal (Huntington, 1997; Huntington et al., 2006; Harmon, 2009). Therefore, most ruminant nutritionists currently adopt feeding strategies designed to increase ruminal starch fermen-tation to improve production efficiencies (Vasconcelos and Galyean, 2007). Increases in fermentable energy supply, however, can lead to metabolic disorders and reductions in DMI (Krehbiel et al., 1995; Owens et al., 1998; Nagaraja and Titgemeyer, 2007), and energy derived by cattle subsequent to fermentation is less than that derived when glucose is absorbed intestinally (Harmon and McLeod, 2001). Therefore, improving small intestinal digestion of starch in cattle may pro-vide great benefit to cattle production systems.
Duodenal supply of glutamate and casein both improve intestinal starch digestion in cattle but by apparently different mechanisms1
D. W. Brake,* E. C. Titgemeyer,*2 and D. E. Anderson†
*Department of Animal Sciences and Industry, and †Department of Clinical Sciences, Kansas State University, Manhattan 66506-1600
ABSTRACT: Greater postruminal flows of protein increase small intestinal starch digestion in cattle. Our objective was to determine if small intestinal starch digestion is increased by duodenal supplementation of AA. We fed 5 duodenally and ileally cannulated steers a low-starch soybean hull–based diet in 5 × 5 Latin square designs and provided continuous duodenal infusion of raw cornstarch in combination with AA or casein and measured small intestinal starch digestion. In Exp. 1 treatments were continuous duodenal infu-sion of 1) no supplement (control), 2) casein (400 g/d), 3) crystalline AA similar in amount and AA composi-tion to the casein (CASAA), 4) crystalline nonessential AA similar to those provided by casein, or 5) crystal-line essential AA similar to those provided by casein. In Exp. 2 treatments were continuous duodenal infusion of 1) no supplement (control), 2) casein (400 g/d), 3) Glu (133 g/d), 4) Phe and Trp plus Met (30.4, 6.5, and 17.5 g/d, respectively; PTM), or 5) a combination of Glu and PTM. Duodenal infusion of casein increased (P ≤
0.05) small intestinal starch digestion. When CASAA was infused, small intestinal starch digestion was simi-lar (P = 0.30) to casein infusion. Infusion of only non-essential AA tended to increase (P = 0.14) small intesti-nal starch digestion relative to the control, but infusion of essential AA alone did not affect (P = 0.84) small intestinal starch digestion. In addition, infusion of casein or CASAA increased ileal flows of ethanol-solu-ble starch (small-chain α-glycosides), but nonessential AA alone were not different than the control. Duodenal infusion of Glu increased (P ≤ 0.05) small intestinal starch digestion, whereas PTM did not. Neither Glu nor PTM increased ileal flow of ethanol-soluble starch, but Glu and PTM provided together tended (P = 0.07) to increase ileal flows of small chain α-glycosides. Our data suggest that Glu alone can increase small intestinal starch digestion in cattle similar to casein, but increases in small intestinal starch digestion in response to Glu are not associated with an increase in ileal flows of small chain α-glycosides.
Key words: amino acids, cattle, digestion, intestine, starch
© 2014 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2014.92:4057–4067 doi:10.2527/jas2014-7909
1Contribution no. 14-324-J from the Kansas Agricultural Experiment Station, Manhattan.
2Corresponding author: [email protected] April 5, 2014.Accepted July 6, 2014.
Published December 12, 2014

Brake et al.4058
Unfortunately, dietary starch per se apparently re-duces secretion of pancreatic α-amylase, which is re-sponsible for the initial hydrolysis of starch during its di-gestion (Kreikemeier et al., 1990; Swanson et al., 2002, 2004). However, several reports have demonstrated that greater flow of protein to the small intestine of cattle can increase small intestinal starch digestion (Streeter and Mathis, 1995; Richards et al., 2002; Brake et al., 2014), portal glucose appearance (Taniguchi et al., 1995), and pancreatic α-amylase secretion (Richards et al., 2003). Many studies that have evaluated effects of small intes-tinal protein flows on small intestinal starch digestion and pancreatic α-amylase secretion used casein, and as a result, little is known about the effects of protein quality or AA flow on small intestinal starch digestion. Indeed, recent reports (Swanson et al., 2004; Liao et al., 2009) suggest that interactions between supplemental proteins and pancreatic α-amylase might be affected by postru-minal flows of α-glucosides and that requirements for nutritionally functional AA (Wu, 2010, 2013) to opti-mize pancreatic secretions may be proportional to starch appearing in the lumen of the small intestine in cattle.
Our studies were designed to evaluate the effects of AA on small intestinal starch digestion in cattle. We hy-pothesized that AA were largely responsible for the effects of postruminal casein supplementation on small intestinal starch digestion in cattle and that responses may be elic-ited by either groups or individual AA found in casein.
MATERIALS AND METHODS
Experiment 1
Five duodenally and ileally cannulated steers (aver-age initial BW = 259 ± 8.9 kg) were placed in a 5 × 5 Latin square with 6-d periods. Animals were tethered to tie-stalls (1.7 by 1.2 m) in a temperature-controlled room (20°C) under 16 h of light (0500 to 2100 h) and 8 h dark. Cattle were provided ad libitum access to water and lim-it fed 4.8 kg/d (DM basis; 0700 and 1900 h) of a soybean hull–based diet (Table 1; about 1.5 × maintenance en-ergy requirement; NRC, 1996). The diet was formulated to provide adequate ruminally available N and to meet or exceed requirements for vitamins and minerals (NRC, 1996). The diet was specifically designed to contain little starch, and it also was designed to provide only modest amounts of RUP so that MP supply would be moderate.
Treatments were continuous duodenal infusion of raw cornstarch alone (1,391 ± 9.9 g DM/d) or cornstarch plus either casein (390 ± 6.6 g DM/d), crystalline AA similar in amount and AA composition to the casein (CASAA; Table 2), crystalline nonessential AA similar to that provided by casein (NEAA; Table 2), or crys-talline essential AA similar to that provided by casein
(EAA; Table 2). Glutamine and Asn were excluded from AA infusions due to instability in solution; amounts of Glu were similar to the content of Glu plus Gln in casein, and Asp was similar to Asp plus Asn in casein.
Cattle were infused through Tygon tubing (i.d. = 2.38 mm; Saint-Gobain North America, Valley Forge, PA) with a peristaltic pump (model CP-78002-10; Cole-Parmer Instrument Company, Vernon Hills, IL) with 12.6 L/d of treatment suspensions. Suspensions (2 containers of 7 L) were prepared daily immediately before infusion for use over 12-h intervals. Cornstarch was maintained in suspension with continuous stirring (Arrow 6000; Arrow Engineering Company, Hillsdale, NJ) during infusion, and the pumping rate was 525 mL/h. Cornstarch suspensions contained CrEDTA (485 mg Cr/12 h; Binnerts et al., 1968), which served as an in-digestible marker, 800 g of raw cornstarch (pure food powder cornstarch; Tate and Lyle Ingredients North Americas, Decatur, IL), and deionized H2O to prepare the final solution weight. The pH of suspensions con-taining CASAA and NEAA were adjusted to near 7 with addition of 39.5 g of 40% (wt/wt) NaOH to each 12-h suspension. Each infusate was prepared daily by weight, and the exact amount infused was determined by record-ing the weight of residual infusate after each 12-h infu-sion period. Suspensions were prepared in slight excess so that infusions were continuous; this approach led to some variation in total infusion amounts. Every 48 h, 100 mL of water was flushed through the lines to prevent accumulation of residual infusate.
Table 1. Composition of soybean hull–based diet fed to steersItem % of DMIngredient
Soybean hulls 72.2Brome hay, late bloom 20.0Corn steep liquor 6.0Limestone 1.0Salt 0.5Mineral and vitamin premix1 0.3
Composition Exp. 1 Exp. 2DM,2 % 88.6 ± 0.38 88.9 ± 0.27OM,2 % DM 92.2 ± 0.17 92.1 ± 0.09CP,2 % DM 14.5 ± 0.06 13.8 ± 0.05Starch,2 % DM 1.8 ± 0.06 2.1 ± 0.05RDP,3 % CP 59.1 59.1NEm,3 Mcal/kg 1.73 1.73
1Provided to diets (per kg diet DM): 50 mg Mn, 50 mg Zn, 10 mg Cu, 0.5 mg I, 0.2 mg Se, 2,200 IU of vitamin A, 275 IU of vitamin D, and 25 IU of vitamin E.
2Based on laboratory analyses; mean ± SEM.3Calculated using the tabular values of NRC (1996).

Starch digestion in cattle 4059
On d 6 of each period, spot samples of ileal digesta and feces were collected 2, 4, 6, 8, 10, and 12 h after the 0700 h feeding. Samples of ileal digesta (200 g) were col-lected by attaching a plastic bag (140 by 229 mm) to the cannula, and feces (200 g) were collected after cattle def-ecated in response to stimulation. Immediately after sam-pling, pH of ileal samples were measured with a mobile pH meter (Portable Meter Model 250A; Orion Research, Beverley, MA). The pH of feces was determined after mixing approximately 5 g of feces with 15 mL of deion-ized H2O. To preserve starch content of digesta and feces, α-amylase was deactivated immediately after measuring pH by increasing sample pH to near 11 by mixing sam-ples with 2.1 mL of 40% (wt/wt) NaOH. Samples were composited and frozen (–20°C) between collections.
Composite digesta and feces were thawed at room temperature (22°C) before analysis. For analyses of DM, starch, glucose, ethanol-soluble starch, and organic acids, pH of an aliquot of previously alkalized ileal di-gesta (75 g) or wet feces (50 g with addition of 25 mL H2O) was neutralized with 1.0 to 1.5 mL of 6 M HCl. A portion of ileal digesta and feces was analyzed for DM by drying at 105°C for 24 h. Starch concentrations were measured using the glucogenic assay described by Herrera-Saldana and Huber (1989) with glucose quan-tified using a glucose oxidase–linked assay (Gochman and Schmitz, 1972). Unpolymerized glucose was deter-mined from assay tubes to which no enzyme was added.
Ethanol-soluble starch was determined using techniques similar to those of Kreikemeier and Harmon (1995) as described by Brake et al. (2014).
The supernatant collected after centrifugation (20,000 × g for 15 min at 4°C) of neutralized ileal digesta and fe-ces was analyzed for Cr concentration by atomic absorp-tion spectrophotometry. A portion of supernatant was also retained, and 1 mL was mixed with 0.25 mL of 25% (wt/vol) metaphosphoric acid and analyzed for VFA and lactic acid by GLC as described by DeFrain et al. (2002).
At 11.5 h after feeding (30 min before final diges-ta sampling), jugular blood (20 mL) was collected by venipuncture into glass vacuum tubes (16 by 100 mm; Monoject Blood Collection Tubes; Sherwood Medical, St. Louis, MO) containing sodium heparin isolated from porcine mucosa (143 United States Pharmacopeia units) to allow measures of circulating glucose, cho-lecystokinin (CCK), α-amino-N, and urea-N concen-trations. Blood was immediately placed on ice, and plasma was harvested by centrifugation (2,200 × g for 15 min at 4°C). Plasma was transferred to plastic tubes (Fisherbrand Premium 2.0 mL MCT Graduated Natural; Fisher Scientific, Waltham, MA), and apro-tinin (500 Kallikrein inhibitor units per mL plasma) was added to a portion of plasma (3 mL). Plasma samples were immediately frozen (–20°C), and plasma contain-ing aprotinin was vortexed for 10 sec before freezing (–80°C). Plasma containing aprotinin was analyzed for CCK with the double antibody RIA described by Benson and Reynolds (2001). Plasma urea (Marsh et al., 1965), plasma glucose (Gochman and Schmitz, 1972), and plasma α-amino-N (Palmer and Peters, 1969) were measured with an AutoAnalyzer (Technicon Analyzer II; Technicon Industrial Systems, Buffalo Grove, IL).
Samples of the diet (200 g/d) were collected 1 and 2 d before sampling of digesta and feces, composited, and stored at room temperature (22°C) until analysis. Feed samples were ground to pass a 1-mm screen (Thomas-Wiley Laboratory Mill Model 4; Thomas Scientific USA, Swedesboro, NJ). Dry matter content was determined by drying samples at 105°C for 24 h in a forced-air oven, and OM was subsequently determined by ashing in a muffle furnace for 8 h at 450°C. Nitrogen content of feed was determined through combustion (Nitrogen Analyzer Model FP-2000; Leco Corporation, St. Joseph, MI), and CP was calculated as 6.25 × N. Starch was analyzed us-ing methods previously described.
Experiment 2
This experiment was conducted to measure response of small intestinal starch digestion by cattle to Glu, Phe plus Trp plus Met (PTM), or a combination of Glu and PTM (Glu+PTM) in amounts 1.5 times those contained
Table 2. Crystalline AA treatments in Exp. 1 and Exp. 2
AA, g/d
Exp. 11 Exp. 22
CASAA NEAA EAA Glu PTMl-Asp 28.9 28.9 – – –l-Thr 17.0 – 17.0 – –l-Ser 22.2 22.2 – – –l-Glu 86.2 86.2 – 133 –l-Pro 41.5 41.5 – – –l-Lys HCl 31.3 – 31.3 – –Gly 7.3 7.3 – – –l-Ala 12.2 12.2 – – –l-Val 25.2 – 25.2 – –l-Met 11.3 – 11.3 – 17.5l-Ile 19.3 – 19.3 – –l-Leu 38.2 – 38.2 – –l-Phe 19.7 – 19.7 – 30.4l-His HCl∙H2O 11.3 – 11.3 – –l-Arg 14.9 – 14.9 – –l-Cys 1.9 1.9 – – –l-Trp 4.2 – 4.2 – 6.5
1CASAA = crystalline AA similar in amount and AA composition to the casein; NEAA = crystalline nonessential AA similar to that provided by casein; EAA = crystalline essential AA similar to that provided by casein. Amounts provided are those infused when 1,391 g/d of cornstarch was infused.
2PTM = Phe plus Trp plus Met. Amounts provided are those infused when 1,429 g/d of cornstarch was infused.
Brake et al.4060
in 400 g casein (Table 3). The amounts of AA used in this experiment were chosen to determine if individu-al AA (i.e., Glu) or a combination of several AA (i.e., PTM) could elicit responses similar to casein. Infusions of raw cornstarch alone (1,429 ± 4.9 g DM/d) and raw cornstarch plus casein (368 ± 1.7 g DM/d) served as negative and positive controls. The pH of suspensions containing Glu were adjusted to near 7 with addition of 40.0 g of 40% (wt/wt) NaOH to each 12-h suspension.
Five duodenally and ileally cannulated steers (av-erage initial BW = 361 kg) were used in a 5 × 5 Latin square with 6-d periods. Animals were limit-fed 5.5 kg/d (DM basis) of the same diet and housed under identical conditions as Exp. 1. Cattle infusions, treatment suspen-sions, sampling, and sample analyses were identical to methods previously described for Exp. 1.
Calculations
Digesta and nutrient flows at the ileum and feces as well as digestibilities were calculated as described by Kreikemeier and Harmon (1995) and detailed by Brake et al. (2014).
Statistical Analyses
Data within each experiment were analyzed using the MIXED procedure of SAS (SAS Inst. Inc., Cary, NC). Fixed effects included treatment and period, and steer was included as a random effect. The LSMEANS option was used to calculate treatment means. For Exp. 1, when the F-statistic for treatment was significant (P ≤ 0.05), all pairwise comparisons of least squares means were evaluated using the PDIFF option. For Exp. 2, effects of Glu and PTM were analyzed by treatment contrasts that considered Glu and PTM as a 2 × 2 factorial; the positive
Table 3. Effect of duodenal infusion of AA or casein on ileal and fecal nutrient flows and small and large intestinal starch disappearance in steers receiving 1.4 kg/d duodenally infused raw cornstarch (Exp. 1)
Item
Treatment1
SEM PControl EAA NEAA CASAA CaseinNo. of observations 5 5 5 5 5Duodenal starch infused, g/d 1,363 1,403 1,396 1,408 1,383 19 –Ileal DM content, % 14.6 15.0 15.4 15.4 16.0 0.59 0.53Ileal pH 6.92a 6.87a 6.84ab 6.80ab 6.72b 0.052 0.04Nutrient flow to ileum, g/d
DM 3,364 3,213 3,195 2,828 3,020 118 0.06Starch 882 895 817 742 788 44 0.08Ethanol-soluble starch 178a 240b 182a 209ab 219b 13 0.01Glucose 47 50 79 55 76 16 0.26Acetate 28.3 28.5 28.5 23.8 25.0 3.7 0.80Propionate 1.4 1.7 1.4 1.1 1.1 0.36 0.70Butyrate 2.3 2.0 2.4 1.7 2.0 0.50 0.74Lactate 41.6 43.6 40.5 33.6 41.0 4.9 0.30
Small intestinal starch disappearance, % of infusion 35.4a 36.1a 41.5ab 47.3b 43.1ab 2.8 0.05Fecal DM content, % 19.7a 19.2a 17.4b 19.8a 19.1a 0.62 0.04Fecal pH 5.59a 5.69a 5.21b 5.59a 5.81a 0.18 0.03Nutrient flow to feces, g/d
DM 2,410a 2,284ab 2,143b 2,155b 2,187b 65 0.03Starch 231a 153ab 226a 90b 69b 45 0.04Ethanol-soluble starch 26 45 34 32 19 11.0 0.54Glucose 46 62 57 56 47 9.1 0.65Acetate 32.9 31.5 36.0 29.4 33.7 2.6 0.16Propionate 11.3 12.9 6.4 11.1 11.8 2.5 0.07Butyrate 10.8a 11.1a 5.1b 11.7a 10.3a 1.4 <0.01Lactate 34.9ab 32.5ab 56.7c 22.5b 39.5a 6.1 <0.01Total organic acids 90.5ab 88.6ab 104.7a 75.3b 95.8a 7.7 0.04
Large intestinal starch disappearance, % of ileal flow 73.3 81.7 70.4 88.4 91.2 5.9 0.06Postruminal starch disappearance, % of infusion 83.0a 89.1ab 83.6a 93.7b 95.0b 2.9 0.03
a–cMeans in same row not bearing a common letter differ (P < 0.05).1EAA = crystalline essential AA similar to that provided by casein (194 g/d AA in profile similar to essential AA of casein); NEAA = crystalline nonessential
AA similar to that provided by casein (203 g/d AA in profile similar to nonessential AA in casein); CASAA = crystalline AA similar in amount and AA composition to the casein (394 g/d AA in profile similar to casein); Casein = 390 g casein DM/d.
Starch digestion in cattle 4061
control (casein) was compared to the negative control by a t test. For pH data of digesta and feces, fixed ef-fects were treatment, period, time, and time × treatment, and steer was included as a random effect. The repeated term was time, with steer × period serving as the subject. The covariance structure was first-order autoregressive. Effects were considered significant at P ≤ 0.05, and ten-dencies were considered at 0.05 < P ≤ 0.15. Pearson’s correlation coefficients (Pearson, 1920) were calculated with the CORR procedure of SAS to determine simple 2-way linear relationships among measures.
RESULTS
Cornstarch used for duodenal infusion in both experi-ments contained 90.3 ± 0.1% DM, and the DM was com-posed entirely of starch (100.7 ± 1.1% starch on a DM basis).
Experiment 1
Nutrient flows and small intestinal starch digest-ibilities are reported in Table 3. Small intestinal starch digestion was different among treatments (P = 0.05). When CASAA was infused, small intestinal starch di-gestion was greater (P < 0.01) than for the control, and both casein (P = 0.07) and NEAA (P = 0.14) tended to increase small intestinal starch digestion compared with the control; however, when cattle received EAA, small intestinal starch digestion was not different (P = 0.84) than the control. Flows of ethanol-soluble starch (small-chain α-glycosides) to the ileum also were affected (P = 0.01) by treatment. Ethanol-soluble starch flows were not different (P = 0.83) than control when NEAA was infused, but infusion of casein (P = 0.03) and EAA (P < 0.01) increased ileal flow of ethanol-soluble starch com-pared with the control. Ethanol-soluble starch flows were intermediate for CASAA and least for the control and NEAA. Ileal flows of unpolymerized glucose did not dif-fer among treatments (P = 0.26) and represented 4.4 ± 0.7% of starch infused daily. Flows of organic acids to
the terminal ileum were not affected (P ≥ 0.30) by treat-ment, and total organic acid flows were 70.7 ± 3.7 g/d.
Total fecal organic acid flows were greater (P = 0.04) for NEAA and casein (105 and 96 g/d, respectively) than for CASAA (75 g/d); total fecal organic acid flows for control and EAA (91 and 89 g/d, respectively) were in-termediate and not different from any other treatment. Interestingly, under the conditions of this experiment, only a weak negative linear association (r = –0.28, P = 0.18) was observed between fecal pH and total flow of organic acids to feces; however, fecal pH was negatively associated (r = –0.57, P < 0.01) with fecal lactate flow. Large intestinal fermentation of starch tended (P = 0.06) to be least for the control and NEAA. Because differ-ences among treatments for large intestinal starch fer-mentation were nearly 2-fold greater than differences in small intestinal starch digestion, fecal starch flows were greatest (P = 0.04) and postruminal starch digestion least (P = 0.03) for control and NEAA; fecal starch flows were least and postruminal starch digestions were greatest for CASAA and casein, and both were intermediate for EAA.
Plasma glucose tended to be increased by casein, CASAA, and EAA (P < 0.11; Table 4). Also, relative to control, plasma CCK concentrations (Table 4) were not affected by treatment (P > 0.16), although the large variation would prevent detection of modest differences. Plasma urea-N (P < 0.01) was least for control, intermedi-ate for EAA, and greatest for NEAA, CASAA, and casein (Table 4). Plasma concentrations (P = 0.03) of α-amino-N were greatest for EAA and casein, least for control and NEAA, and intermediate for CASAA (Table 4).
Experiment 2
Flows of starch to the ileum decreased when Glu (P < 0.01) or casein (P = 0.02) were infused but were not affected by PTM (Table 5). Similarly, both Glu (P < 0.01) and casein (P = 0.02) increased apparent small intestinal digestion of starch, but small intestinal starch digestion was not affected by PTM. We observed a ten-
Table 4. Effect of duodenal infusion of AA or casein on plasma concentrations of circulating metabolites and chole-cystokinin in steers receiving 1.4 kg/d duodenally infused raw cornstarch (Exp. 1)
Plasma
Treatment1
SEM PControl EAA NEAA CASAA CaseinNo. of observations 5 5 5 5 5Cholecystokinin, pmol/L 7.6 4.8 8.2 9.9 4.7 1.65 0.08Glucose, mmol/L 4.70 4.87 4.68 4.91 4.91 0.16 0.07Urea N, mmol/L 1.19a 1.87b 2.54c 2.64c 3.13c 0.27 <0.01α-Amino N, mmol/L 2.67a 2.97b 2.64a 2.92ab 3.08b 0.12 0.03
a–cMeans in same row not bearing a common letter differ (P < 0.05).1EAA = crystalline essential AA similar to that provided by casein (194 g/d AA in profile similar to essential AA of casein); NEAA = crystalline nonessential
AA similar to that provided by casein (203 g/d AA in profile similar to nonessential AA in casein); CASAA = crystalline AA similar in amount and AA composi-tion to the casein (394 g/d AA in profile similar to casein); Casein = 390 g casein DM/d.
Brake et al.4062
dency (P = 0.07) for an interaction between Glu and PTM for ileal flows of ethanol-soluble starch; infusion of Glu (P = 0.86) or PTM (P = 0.97) alone did not affect ileal flow of ethanol-soluble starch compared with the control, but Glu+PTM increased (P = 0.01) ileal flow of ethanol-soluble starch relative to the control. Flow of unpolymerized glucose to the terminal ileum was not affected by treatment (P ≥ 0.48) and represented 1.0 ± 0.2% of the glucose infused as starch.
Flows of acetate, lactate, and total organic acids to the feces were not affected by treatment (P ≥ 0.36). Similarly, fecal pH did not differ among treatments (P ≥ 0.40). Unlike Exp. 1, large intestinal fermentation of starch was not affected (P ≥ 0.37) by treatment. Postruminal starch digestion tended to be greater than the control (P = 0.07) when casein was infused. Average large intestinal fer-mentation of starch was 81.4 ± 2.3% of ileal starch flow.
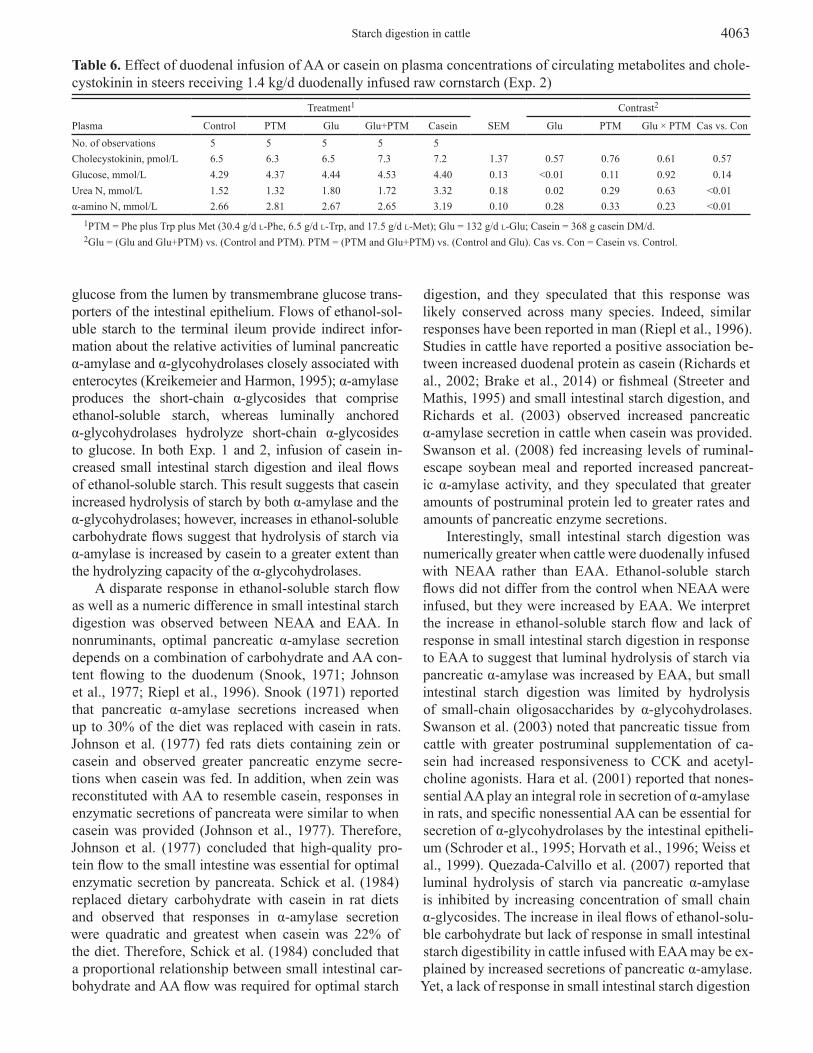
Plasma glucose was increased (P < 0.01) by Glu, and it tended to be slightly increased by both PTM (P = 0.11) and casein (P = 0.14; Table 6). Concentration of CCK did not differ among treatments (P ≥ 0.57). Plasma urea-N was increased by Glu (P = 0.02) and casein (P < 0.01). Casein increased (P < 0.01) concentration of α-amino-N in plasma.
DISCUSSION
Small intestinal starch digestion is thought to occur by similar means in both ruminants and nonruminants, and its digestion can be separated into 3 independent segments (Huntington, 1997): 1) hydrolysis of starch by α-amylase to smaller oligosaccharides, 2) release of glucose from oligosaccharides by α-glycohydrolases produced by the intestinal epithelium, and 3) uptake of
Table 5. Effect of duodenal infusion of AA or casein on ileal and fecal nutrient flows and small and large intestinal starch disappearance in steers receiving 1.4 kg/d duodenally infused raw cornstarch (Exp. 2)
Item
Treatment1
SEM
Contrast2
Control PTM Glu Glu+PTM Casein Glu PTM Glu × PTMCas vs.
ConNo. of observations 5 5 5 5 5Duodenal starch infused, g/d 1,427 1,435 1,417 1,431 1,436 12.6 0.52 0.31 0.78 0.53Ileal DM content, % 14.3 14.2 14.0 14.4 14.5 0.41 0.86 0.81 0.48 0.81Ileal pH 6.69 6.72 6.72 6.73 6.62 0.030 0.47 0.49 0.73 0.05Nutrient flow to ileum, g/d
DM 3,123 3,214 3,348 3,319 3,487 174 0.36 0.86 0.73 0.16Starch 799 754 661 691 684 38 <0.01 0.79 0.22 0.02Ethanol-soluble starch 122 122 124 161 149 19 0.05 0.06 0.07 0.06Glucose 13.7 16.7 15.9 16.2 12.5 2.6 0.71 0.48 0.56 0.72Acetate 24.3 23.6 22.7 23.3 27.0 1.7 0.48 0.99 0.63 0.17Propionate 1.2 0.9 0.9 1.0 0.9 0.16 0.48 0.37 0.16 0.23Butyrate 2.2 2.4 2.2 2.2 2.3 0.46 0.60 0.76 0.76 0.92Lactate 30.8 31.0 32.3 30.5 31.8 2.1 0.80 0.69 0.64 0.72
S mall intestinal starch disappearance, % of infusion
44.1 47.4 53.4 51.8 52.4 2.6 <0.01 0.70 0.28 0.02
Fecal DM content, % 19.0 19.6 19.2 18.6 19.7 0.65 0.44 0.99 0.25 0.34Fecal pH 5.80 5.95 5.97 5.96 5.86 0.17 0.33 0.42 0.40 0.63Nutrient flow to feces, g/d
DM 2,479 2,477 2,493 2,575 2,373 123 0.63 0.73 0.72 0.52Starch 180 151 126 157 121 36 0.29 0.98 0.19 0.08Ethanol-soluble starch 10.6 5.8 6.5 8.8 3.3 3.2 0.84 0.65 0.21 0.08Glucose 35.4 38.4 31.7 25.8 29.5 3.6 0.02 0.63 0.16 0.18Acetate 37.2 35.7 35.2 34.0 34.3 2.7 0.45 0.58 0.94 0.40Propionate 12.7 13.0 15.8 9.8 16.8 2.8 0.95 0.13 0.10 0.13Butyrate 14.7 16.3 15.6 13.5 13.1 1.3 0.45 0.85 0.16 0.40Lactate 42.1 35.5 42.6 40.2 42.5 11.1 0.79 0.64 0.83 0.98Total organic acids 107.6 101.3 110.0 98.5 108.0 12.8 0.99 0.36 0.79 0.98
L arge intestinal starch disappearance, % of ileal flow
77.7 79.9 81.7 77.4 81.2 5.2 0.84 0.77 0.37 0.50
P ostruminal starch disappearance, % of infusion
87.4 89.5 91.2 89.0 91.6 2.5 0.31 0.99 0.18 0.07
1PTM = Phe plus Trp plus Met (30.4 g/d l-Phe, 6.5 g/d l-Trp, and 17.5 g/d l-Met); Glu = 132 g/d l-Glu; Casein = 368 g casein DM/d.2Glu = (Glu and Glu+PTM) vs. (Control and PTM). PTM = (PTM and Glu+PTM) vs. (Control and Glu). Cas vs. Con = Casein vs. Control.
Starch digestion in cattle 4063
glucose from the lumen by transmembrane glucose trans-porters of the intestinal epithelium. Flows of ethanol-sol-uble starch to the terminal ileum provide indirect infor-mation about the relative activities of luminal pancreatic α-amylase and α-glycohydrolases closely associated with enterocytes (Kreikemeier and Harmon, 1995); α-amylase produces the short-chain α-glycosides that comprise ethanol-soluble starch, whereas luminally anchored α-glycohydrolases hydrolyze short-chain α-glycosides to glucose. In both Exp. 1 and 2, infusion of casein in-creased small intestinal starch digestion and ileal flows of ethanol-soluble starch. This result suggests that casein increased hydrolysis of starch by both α-amylase and the α-glycohydrolases; however, increases in ethanol-soluble carbohydrate flows suggest that hydrolysis of starch via α-amylase is increased by casein to a greater extent than the hydrolyzing capacity of the α-glycohydrolases.
A disparate response in ethanol-soluble starch flow as well as a numeric difference in small intestinal starch digestion was observed between NEAA and EAA. In nonruminants, optimal pancreatic α-amylase secretion depends on a combination of carbohydrate and AA con-tent flowing to the duodenum (Snook, 1971; Johnson et al., 1977; Riepl et al., 1996). Snook (1971) reported that pancreatic α-amylase secretions increased when up to 30% of the diet was replaced with casein in rats. Johnson et al. (1977) fed rats diets containing zein or casein and observed greater pancreatic enzyme secre-tions when casein was fed. In addition, when zein was reconstituted with AA to resemble casein, responses in enzymatic secretions of pancreata were similar to when casein was provided (Johnson et al., 1977). Therefore, Johnson et al. (1977) concluded that high-quality pro-tein flow to the small intestine was essential for optimal enzymatic secretion by pancreata. Schick et al. (1984) replaced dietary carbohydrate with casein in rat diets and observed that responses in α-amylase secretion were quadratic and greatest when casein was 22% of the diet. Therefore, Schick et al. (1984) concluded that a proportional relationship between small intestinal car-bohydrate and AA flow was required for optimal starch
digestion, and they speculated that this response was likely conserved across many species. Indeed, similar responses have been reported in man (Riepl et al., 1996). Studies in cattle have reported a positive association be-tween increased duodenal protein as casein (Richards et al., 2002; Brake et al., 2014) or fishmeal (Streeter and Mathis, 1995) and small intestinal starch digestion, and Richards et al. (2003) observed increased pancreatic α-amylase secretion in cattle when casein was provided. Swanson et al. (2008) fed increasing levels of ruminal-escape soybean meal and reported increased pancreat-ic α-amylase activity, and they speculated that greater amounts of postruminal protein led to greater rates and amounts of pancreatic enzyme secretions.
Interestingly, small intestinal starch digestion was numerically greater when cattle were duodenally infused with NEAA rather than EAA. Ethanol-soluble starch flows did not differ from the control when NEAA were infused, but they were increased by EAA. We interpret the increase in ethanol-soluble starch flow and lack of response in small intestinal starch digestion in response to EAA to suggest that luminal hydrolysis of starch via pancreatic α-amylase was increased by EAA, but small intestinal starch digestion was limited by hydrolysis of small-chain oligosaccharides by α-glycohydrolases. Swanson et al. (2003) noted that pancreatic tissue from cattle with greater postruminal supplementation of ca-sein had increased responsiveness to CCK and acetyl-choline agonists. Hara et al. (2001) reported that nones-sential AA play an integral role in secretion of α-amylase in rats, and specific nonessential AA can be essential for secretion of α-glycohydrolases by the intestinal epitheli-um (Schroder et al., 1995; Horvath et al., 1996; Weiss et al., 1999). Quezada-Calvillo et al. (2007) reported that luminal hydrolysis of starch via pancreatic α-amylase is inhibited by increasing concentration of small chain α-glycosides. The increase in ileal flows of ethanol-solu-ble carbohydrate but lack of response in small intestinal starch digestibility in cattle infused with EAA may be ex-plained by increased secretions of pancreatic α-amylase. Yet, a lack of response in small intestinal starch digestion
Table 6. Effect of duodenal infusion of AA or casein on plasma concentrations of circulating metabolites and chole-cystokinin in steers receiving 1.4 kg/d duodenally infused raw cornstarch (Exp. 2)
Plasma
Treatment1
SEM
Contrast2
Control PTM Glu Glu+PTM Casein Glu PTM Glu × PTM Cas vs. ConNo. of observations 5 5 5 5 5Cholecystokinin, pmol/L 6.5 6.3 6.5 7.3 7.2 1.37 0.57 0.76 0.61 0.57Glucose, mmol/L 4.29 4.37 4.44 4.53 4.40 0.13 <0.01 0.11 0.92 0.14Urea N, mmol/L 1.52 1.32 1.80 1.72 3.32 0.18 0.02 0.29 0.63 <0.01α-amino N, mmol/L 2.66 2.81 2.67 2.65 3.19 0.10 0.28 0.33 0.23 <0.01
1PTM = Phe plus Trp plus Met (30.4 g/d l-Phe, 6.5 g/d l-Trp, and 17.5 g/d l-Met); Glu = 132 g/d l-Glu; Casein = 368 g casein DM/d.2Glu = (Glu and Glu+PTM) vs. (Control and PTM). PTM = (PTM and Glu+PTM) vs. (Control and Glu). Cas vs. Con = Casein vs. Control.

Brake et al.4064
may have resulted from an increasing concentration of ethanol-soluble carbohydrate in the small intestinal lu-men if activity of α-glycohydrolases was limiting when EAA were provided. Clearly, pancreatic α-amylase and small intestinal α-glycohydrolases are responsive to lu-minal nutrient flows, and these responses are complex.
Similarly to infusion of 400 g/d casein, infusion of Glu alone increased small intestinal starch digestion; however, Glu alone did not increase flows of ethanol-soluble starch to the ileum. Harmon (1992) reviewed the available literature and concluded that α-glycohydrolase activity associated with the intestinal epithelium was re-sponsive to dietary energy rather than substrate and that cattle were less responsive than their nonruminant coun-terparts. It is well known that Gln, Glu, and Asp are key metabolic fuels for small intestinal mucosa (Windmueller and Spaeth, 1975, 1976; Wu, 1998). Indeed, Harmon (2009) indicated that greater intestinal flows of protein as energy apparently increase small intestinal starch di-gestion in cattle. Our data seem to agree with this con-clusion and indicate that 132 g/d of Glu affected small intestinal starch digestion similar to 400 g/d of casein. Yet, hydrolysis of small-chain α-glycosides apparently increased 32% by NEAA compared with EAA (mea-sured as a decrease in flow of ethanol-soluble starch to the ileum), which suggests that the hydrolytic capacity of bovine small intestine for small chain α-glycosides was appreciably increased in response to nonessential AA.
Yu et al. (2013) reported that duodenal infusion of Phe decreased pancreatic α-amylase secretions in goats. In the current experiment, small intestinal starch digestion was not different from the control when PTM was infused, and flows of ethanol-soluble starch were not different from the control when PTM alone was infused. Flows of ethanol-soluble starch to the terminal ileum were greatest for Glu+PTM. Similarly, casein tended to increase ileal flows of ethanol-soluble starch. We observed a tendency for an interaction between Glu and PTM among flows of ethanol-soluble starch to the ileum. A precise expla-nation of the interaction between Glu and PTM among flows of ethanol-soluble starch to the ileum remains un-clear. It is possible that the increased ileal ethanol-soluble starch flows in response to Glu+PTM were spurious, but responses to Glu+PTM were similar to those observed for casein with regard to ileal flow of ethanol-soluble starch and small intestinal starch digestion.
Relatively less is known about the regulation of postruminal nutritional physiology in ruminants com-pared with that in the reticulorumen of ruminants or in the postgastric compartments of nonruminants (Van Soest, 1994). Indeed, Gressley et al. (2011) recently pointed out the paucity of data related to large intesti-nal or hindgut fermentation in ruminants. Nonetheless, Gressley et al. (2011) summarized studies with steers,
sheep, and lactating cows fitted with cannulas in the duodenum and terminal ileum. They reported that when flow of starch to the ileum was low (≤110 g/d), the large intestinal fermentation of starch was small (≤2% of total tract starch digestion). Similarly, Gressley et al. (2011) noted that when starch flows to the ileum were great-er (i.e., 340 to 1,650 g/d) large intestinal fermentation accounted for a greater proportion of total tract starch disappearance. Large intestinal disappearance of starch was not closely associated with ileal starch flow in Exp. 1 (r = 0.16, P = 0.44) or Exp. 2 (r = –0.04, P = 0.84); how-ever, large intestinal fermentation of starch was strongly related (r = 0.56, P < 0.01) to ethanol-soluble starch flow to the terminal ileum in Exp. 1. Ethanol-soluble starch may be fermented more extensively than ethanol-insol-uble starch in the large intestine of cattle, although fer-mentation of ethanol-insoluble starch was quantitatively the greatest portion of starch that disappeared across the large intestine. Relative to the control, EAA, CASAA, and casein tended to increase large intestinal starch di-gestion. Interestingly, large intestinal disappearance of starch for NEAA was not different from the control, and this contributed to postruminal starch digestibility for NEAA similar to the control, despite a tendency for greater small intestinal starch digestion for NEAA than for the control. Carbohydrates generally are considered to be fermented in the large intestine similarly to in the rumen (Gressley et al., 2011); however, in some cases, large intestinal starch disappearance in ruminants has been reported to be greater than ruminal disappearance (Haïmoud et al., 1995; Callison et al., 2001; Gressley et al., 2011). Theurer (1986) calculated that ruminal di-gestion of starch in whole, dry-rolled, or steamed-whole corn averaged 70%. This value is in close agreement with our observations of large intestinal starch digestion for control and NEAA but is less than large intestinal starch digestibility for EAA, CASAA, and casein.
Gressley et al. (2011) concluded that the acid load from starch in the large intestine of ruminants is a func-tion of total starch flows to and the rate of starch fermen-tation in the hindgut. Total flow of organic acids to feces tended to be related to large intestinal starch fermenta-tion in Exp. 1 (r = 0.34, P = 0.10) and was not related to large intestinal starch fermentation in Exp. 2 (r = 0.23, P = 0.28). Fecal output of total organic acids may not re-flect total production of organic acids in the large intes-tine and may be responsive to location of fermentation in the large intestine.
Total flow of organic acids to feces was greater for NEAA and casein, intermediate for the control and EAA, and least for CASAA in Exp. 1. In addition, in Exp. 2 total flow of organic acids to feces did not differ among treatments, and total flow of organic acids to feces was not related to fecal pH in Exp. 1 (r = –0.07, P = 0.75) or

Starch digestion in cattle 4065
in Exp. 2 (r = 0.03, P = 0.89). Bissell and Hall (2010) in-fused starch into the abomasum of cattle (BW = 804 kg) at more than twice the rate that we infused starch to the duodenum. These authors speculated that differences in fecal pH were related to differences in small intestinal starch digestion and the capability of cattle to adapt to treatment. Small intestinal digestion of starch and fecal pH were not significantly correlated in Exp. 1 (r = 0.26, P = 0.20) or Exp. 2 (r = 0.27, P = 0.19).
Cholecystokinin, a hormone secreted by enteroen-docrine I-cells in response to luminal nutrient flows, is generally thought to modulate pancreatic exocrine se-cretions in mammals (Konturek et al., 2003). Increased dietary protein has been associated with increased plas-ma CCK in both nonruminants (Konturek et al., 1973; Nishi et al., 2003; Veldhorst et al., 2008) and ruminants (Furuse et al., 1992). Swanson et al. (2003) observed direct responses of cattle pancreatic tissues when incu-bated with caerulein, a CCK analog. Circulating concen-trations of CCK were not strongly affected by treatment in Exp. 1 or Exp. 2, and CCK was not closely related to either small intestinal starch digestion (Exp. 1: r = 0.08, P = 0.70; Exp. 2: r = 0.21, P = 0.31) or ileal flow of ethanol-soluble oligosaccharide in Exp. 2 (r = 0.23, P = 0.28). However, jugular concentration of CCK tended to be inversely related (r = –0.38, P = 0.06) to flows of ethanol-soluble oligosaccharides to the terminal il-eum in Exp. 1. Nonetheless, some authors (Morisset et al., 2003; Konturek et al., 2003; Swanson et al., 2003; Zabielski, 2003) have questioned the classic hormone theory associated with CCK, and a preponderance of re-cent findings suggest that the parasympathetic nervous system is often involved in mediating CCK’s stimula-tory effect. Direct reports (Benson and Reynolds, 2001) that hepatic removal of CCK in cattle is related to hepat-ic metabolism further questions CCK’s role as a precise chemical messenger to the pancreas in cattle. Swanson et al. (2003) reported that responses of cattle pancreatic tissue to a CCK analog was dynamic and occurred only in tissues harvested from cattle that had received greater postruminal protein supplies. Clearly, a complete de-scription of CCK’s relationship to small intestinal starch digestion in cattle is yet to be elucidated.
In summary, casein and EAA appeared to increase pancreatic α-amylase secretion, but EAA did not increase small intestinal starch digestion. In contrast, NEAA and Glu increased small intestinal starch digestion but not ileal flows of small-chain α-glycosides. A possible ex-planation for these observations is that EAA may have affected pancreatic α-amylase but not small intestinal α-glycohydrolases. When Harmon (1992) reviewed the available literature on energy supplementation to the small intestine of cattle, he concluded that changes in production of α-glycohydrolases in cattle were limited
and that capacity for small intestinal hydrolysis of small-chain α-glycosides in cattle may be more closely associ-ated with increases in small intestinal length than with changes in α-glycohydrolase secretion. We did not mea-sure small intestinal length, but small intestinal length is unlikely to have changed during our short-term studies. A possible explanation for effects of NEAA and Glu in re-ducing ileal flow of ethanol-soluble starch and in increas-ing small intestinal starch digestion may be that NEAA and Glu increased hydrolysis of small-chain α-glycosides by increasing the α-glycohydrolases. In turn, an increase in the small intestinal hydrolytic capacity of small-chain α-glycosides may indirectly facilitate greater hydrolytic activity of α-amylase. Our manipulations of postrumi-nal Glu (and NEAA) flows may be outside the range of previous manipulations. Dietary manipulations that in-crease postruminal flow of Glu (and NEAA) in relation to postruminal starch flows may increase small intestinal starch digestion in cattle. Increases in small intestinal starch digestion may facilitate improvements in the ef-ficient production of meat and milk by cattle.
LITERATURE CITEDBenson, J. A., and C. K. Reynolds. 2001. Effects of abomasal in-
fusion of long-chain fatty acids on splanchnic metabolism of pancreatic and gut hormones in lactating dairy cows. J. Dairy Sci. 84:1488–1500.
Binnerts, W. T., A. T. Van’t Klooster, and A. M. Frens. 1968. Soluble chromium indicator measured by atomic absorption in diges-tion experiments. Vet. Rec. 81:470.
Bissell, H. A., and M. B. Hall. 2010. Cattle differ in ability to adapt to small intestinal digestion of starch. J. Dairy Sci. 93(E-Suppl. 1):845 (Abstr.).
Brake, D. W., E. C. Titgemeyer, E. A. Bailey, and D. E. Anderson. 2014. Small intestinal digestion of raw cornstarch in cattle con-suming a soybean hull-based diet is improved by duodenal ca-sein infusion. J. Anim. Sci. (in press).
Callison, S. L., J. L. Firkins, M. L. Eastridge, and B. L. Hull. 2001. Site of nutrient digestion by dairy cows fed corn of different particle sizes or steam-rolled. J. Dairy Sci. 84:1458–1467.
DeFrain, J. M., J. E. Shirley, E. C. Titgemeyer, A. F. Park, and R. T. Ethington. 2002. Impact of feeding a raw soybean hull-con-densed corn steep liquor pellet on induced subacute ruminal aci-dosis in lactating cows. J. Dairy Sci. 85:2000–2008.
Fox, D. G., C. J. Sniffen, J. D. O’Connor, J. B. Russell, and P. J. Van Soest. 1992. A net carbohydrate and protein system for evaluat-ing cattle diets: III. Cattle requirements and diet adequacy. J. Anim. Sci. 70:3578–3596.
Furuse, M., M. Kato, S. I. Yang, K. Asakura, and J. Okumura. 1992. Influence of dietary protein concentrations or of duodenal amino acid infusion on cholecystokinin release in goats. Comp. Biochem. Physiol. Comp. Physiol. 101:635–638.
Gochman, N., and J. Schmitz. 1972. Application of a new peroxi-dase indicator reaction to the specific automated determination of glucose with glucose oxidase. Clin. Chem. 18:943–950.
Gressley, T. F., M. B. Hall, and L. E. Armentano. 2011. Ruminant nu-trition symposium: Productivity, digestion and health responses to hindgut acidosis in ruminants. J. Anim. Sci. 89:1120–1130.

Brake et al.4066
Haïmoud, D. A., M. Vernay, C. Bayourthe, and R. Moncoulon. 1995. Avoparcin and monensin effects on the digestion of nutrients in dairy cows fed a mixed diet. Can. J. Anim. Sci. 75:379–385.
Hara, H., N. Akatsuka, and Y. Aoyama. 2001. Non-essential amino acids play an important role in adaptation of the rat exocrine pancreas to high nitrogen feeding. J. Nutr. Biochem. 12:450–457.
Harmon, D. L. 1992. Dietary influences on carbohydrases and small intestinal starch hydrolysis capacity in ruminants. J. Nutr. 122:203–210.
Harmon, D. L. 2009. Understanding starch utilization in the small intestine of cattle. Asian-Australas. J. Anim. Sci. 22:915–922.
Harmon, D. L., and K. R. McLeod. 2001. Glucose uptake and regula-tion by intestinal tissues: Implications and whole-body energet-ics. J. Anim. Sci. 79:E59–E72.
Herrera-Saldana, R., and J. T. Huber. 1989. Influence of varying pro-tein and starch degradabilities on performance of lactating cows. J. Dairy Sci. 72:1477–1483.
Horvath, K., M. Jami, I. D. Hill, J. C. Papadimitriou, L. S. Magder, and S. Chanasongcram. 1996. Isocaloric glutamine-free diet and the morphology and function of rat small intestine. JPEN, J. Parenter. Enteral Nutr. 20:128–134.
Huntington, G. B. 1997. Starch utilization by ruminants: From basics to the bunk. J. Anim. Sci. 75:852–867.
Huntington, G. B., D. L. Harmon, and C. J. Richards. 2006. Sites, rates, and limits of starch digestion and glucose metabolism in growing cattle. J. Anim. Sci. 84:E14–E24.
Johnson, A., R. Hurwitz, and N. Kretchmer. 1977. Adaptation of rat pancreatic amylase and chymotrypsinogen to changes in diet. J. Nutr. 107:87–96.
Konturek, S. J., J. Pepera, K. Zabielski, P. C. Konturek, T. Pawlik, A. Szlachcic, and E. G. Hahn. 2003. Brain-gut axis in pancreatic se-cretion and appetite control. J. Physiol. Pharmacol. 54:293–317.
Konturek, S. J., T. Radecki, P. Thor, and A. Dembinski. 1973. Release of cholecystokinin by amino acids. Proc. Soc. Exp. Biol. Med. 143:305–309.
Krehbiel, C. R., R. A. Britton, D. L. Harmon, T. J. Wester, and R. A. Stock. 1995. The effects of ruminal acidosis on volatile fatty acid absorption and plasma activities of pancreatic enzymes in lambs. J. Anim. Sci. 73:3111–3121.
Kreikemeier, K. K., and D. L. Harmon. 1995. Abomasal glucose, maize starch and maize dextrin infusions in cattle: Small-intestinal disappearance, net portal glucose flux and ileal oligo-saccharide flow. Br. J. Nutr. 73:763–772.
Kreikemeier, K. K., D. L. Harmon, J. P. Peters, K. L. Gross, C. K. Armendariz, and C. R. Krehbiel. 1990. Influence of dietary for-age and feed intake on carbohydrase activities and small intesti-nal morphology of calves. J. Anim. Sci. 68:2916–2929.
Liao, S. F., E. S. Vanzant, D. L. Harmon, K. R. McLeod, J. A. Boling, and J. C. Matthews. 2009. Ruminal and abomasal starch hydro-lysate infusions selectively decrease the expression of cationic amino acid transporter mRNA by small intestinal epithelia of forage-fed beef steers. J. Dairy Sci. 92:1124–1135.
Lofgreen, G. P., and W. N. Garrett. 1968. A system for expressing net energy requirements and feed values for growing and finishing beef cattle. J. Anim. Sci. 27:793–806.
Marsh, W. H., B. Fingerhut, and H. Miller. 1965. Automated and manual direct methods for the determination of blood urea. Clin. Chem. 11:624–627.
Morisset, J., J. Laine, J. Bourassa, M. Lessard, V. Rome, and P. Guilloteau. 2003. Presence and localization of CCK receptor subtypes in calf pancreas. Regul. Pept. 111:103–109.
Nagaraja, T. G., and E. C. Titgemeyer. 2007. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 90:E17–E38.
Nishi, T., H. Hara, and F. Tomita. 2003. The soybean beta-congly-cinin beta 51–63 fragment suppresses appetite by stimulating cholecystokinin release in rats. J. Nutr. 133:2537–2542.
NRC. 1996. Nutrient requirements of beef cattle. 7th rev. ed. National Academy Press Washington, DC.
Owens, F. N., D. S. Secrist, W. J. Hill, and D. R. Gill. 1998. Acidosis in cattle: A review. J. Anim. Sci. 76:275–286.
Palmer, D. W., and T. Peters Jr. 1969. Automated determination of free amino groups in serum and plasma using 2,4,6-trinitroben-zene sulfonate. Clin. Chem. 15:891–901.
Pearson, K. 1920. Notes on the history of correlation. Biometrika 13:25–45.
Quezada-Calvillo, R., C. C. Robayo-Torres, A. R. Opekun, P. Sen, Z. Ao, B. R. Hamaker, A. Quaroni, G. D. Brayer, S. Wattler, M. C. Nehls, E. E. Sterchi, and B. L. Nichols. 2007. Contribution of mucosal maltase-glucoamylase activities to mouse small intes-tinal starch α-glucogenesis. J. Nutr. 137:1725–1733.
Richards, C. J., A. F. Branco, D. W. Bohnert, G. B. Huntington, M. Macari, and D. L. Harmon. 2002. Intestinal starch disappear-ance increased in steers abomasally infused with starch and pro-tein. J. Anim. Sci. 80:3361–3368.
Richards, C. J., K. C. Swanson, S. J. Paton, D. L. Harmon, and G. B. Huntington. 2003. Pancreatic exocrine secretion in steers in-fused postruminally with casein and cornstarch. J. Anim. Sci. 81:1051–1056.
Riepl, R. L., F. Fiedler, M. Ernstberger, J. Teufel, and P. Lehnert. 1996. Effect of intraduodenal taurodeoxycholate and L-phenylalanine on pancreatic secretion and on gastroenteropancreatic peptide release in man. Eur. J. Med. Res. 1:499–505.
Schick, J., R. Verspohl, H. Kern, and G. Scheele. 1984. Two distinct adaptive responses in the synthesis of exocrine pancreatic en-zymes to inverse changes in protein and carbohydrate in the diet. Am. J. Physiol. 247:G611–G616.
Schroder, J., E. Wardelmann, W. Winkler, F. Fandrich, E. Schweizer, and P. Schroeder. 1995. Glutamine dipeptide-supplemented parental nutrition reverses gut atrophy, disaccharidase enzyme activity, and absorption in rats. JPEN, J. Parenter. Enteral Nutr. 19:502–506.
Snook, J. T. 1971. Dietary regulation of pancreatic enzymes in the rat with emphasis on carbohydrate. Am. J. Physiol. 221:1383–1387.
Streeter, M. N., and M. J. Mathis. 1995. Effect of supplemental fish meal protein on site and extent of digestion in beef steers. J. Anim. Sci. 73:1196–1201.
Swanson, K. C., J. A. Benson, J. C. Matthews, and D. L. Harmon. 2004. Pancreatic exocrine secretion and plasma concentra-tion of some gastrointestinal hormones in response to aboma-sal infusion of starch hydrolyzate and/or casein. J. Anim. Sci. 82:1781–1787.
Swanson, K. C., N. Kelly, H. Salim, Y. J. Wang, S. Holligan, M. Z. Fan, and B. W. McBride. 2008. Pancreatic mass, cellularity, and α-amylase and trypsin activity in feedlot steers fed diets differ-ing in crude protein concentration. J. Anim. Sci. 86:909–915.
Swanson, K. C., J. C. Matthews, C. A. Woods, and D. L. Harmon. 2002. Postruminal administration of partially hydrolyzed starch and casein influences pancreatic alpha-amylase expression in calves. J. Nutr. 132:376–381.
Swanson, K. C., J. C. Matthews, C. A. Woods, and D. L. Harmon. 2003. Influence of substrate and/or neurohormonal mimic on in vitro pancreatic enzyme release from calves postruminally infused with partially hydrolyzed starch and/or casein. J. Anim. Sci. 81:1323–1331.
Taniguchi, K., G. B. Huntington, and B. P. Glenn. 1995. Net nutrient flux by visceral tissues of beef steers given abomasal and ru-minal infusions of casein and starch. J. Anim. Sci. 73:236–249.

Starch digestion in cattle 4067
Theurer, C. B. 1986. Grain processing effects on starch utilization by ruminants. J. Anim. Sci. 63:1649–1662.
Van Soest, P. J. 1994. Nutritional ecology of the ruminant. 2nd ed. Cornell Univ. Press, Ithaca, NY.
Vasconcelos, J. T., and M. L. Galyean. 2007. Nutritional recommen-dations of feedlot consulting nutritionists: The 2007 Texas Tech University survey. J. Anim. Sci. 85:2772–2781.
Veldhorst, M., A. Smeets, S. Soenen, A. Hochstenbach-Waelen, R. Hursel, K. Diepvens, M. Lejeune, N. Luscombe-Marsh, and M. Westerterp-Plantenga. 2008. Protein-induced satiety: Effects and mechanisms of different proteins. Physiol. Behav. 94:300–307.
Weiss, M. D., V. DeMarco, D. M. Strauss, D. A. Samuelson, M. E. Lane, and J. Neu. 1999. Glutamine synthetase: A key enzyme for intestinal epithelial differentiation? JPEN, J. Parenter. Enteral Nutr. 23:140–146.
Windmueller, H. G., and A. E. Spaeth. 1975. Intestinal metabolism of glutamine and glutamate from the lumen as compared to gluta-mine from blood. Arch. Biochem. Biophys. 171:662–672.
Windmueller, H. G., and A. E. Spaeth. 1976. Metabolism of absorbed aspartate, asparagine, and arginine by rat small intestine in vivo. Arch. Biochem. Biophys. 175:670–676.
Wu, G. 1998. Intestinal mucosal amino acid catabolism. J. Nutr. 128:1249–1252.
Wu, G. 2010. Functional amino acids in growth, reproduction and health. Adv. Nutr. 1:31–37.
Wu, G. 2013. Functional amino acids in nutrition and health. Amino Acids 45:407–411.
Yu, Z. P., M. Xu, J. H. Yao, K. Liu, F. Li, Y. Liu, F. Wang, F. F. Sun, and N. N. Liu. 2013. Regulation of pancreatic exocrine secre-tion in goats: Differential effects of short- and long-term duo-denal phenylalanine treatment. J. Anim. Physiol. Anim. Nutr. 97:431–438.
Zabielski, R. 2003. Luminal CCK and its neuronal action on exo-crine pancreatic secretion. J. Physiol. Pharmacol. 54:81–94.