Changes in 17β,20α-hydroxysteroid dehydrogenase activity supporting an increase in the...
9
Changes in dehydrogenase activity supporting an increase in the estrogen/progesterone ratio of human fetal membranes at parturition Bryan F. Mitchell, MD, and Susan Wong, MSc Edmonton, Alberta, Canada OBJECTIVE: Our purpose was to measure the activity of the reversible enzyme 1713,20a-hydroxysteroid dehydrogenase around parturition with estrogen and progestogen substrates. STUDY DESIGN: Classic kinetic studies and explant cultures were used to determine kinetic parameters and net enzyme activities in both oxidative and reductive directions for both sets of substrates. RESULTS: Affinity constant values for estrone, estradiol, and 20a-dihydroprogesterone were 1 to 8 /-Lmol/L. Affinity constant for progesterone was 9 to 25 /-Lmol/L. Maximal velocities for all substrates in the chorion were 20- to 70-fold higher than in amnion and severalfold higher for estrogen substrates compared with the progestins. Around parturition there was a significant change toward net formation of the stronger estrogen (estradiol) and the weaker progestin (20a-dihydroprogesterone), suggesting an increase in the local estrogen/progesterone ratio. CONCLUSION: The enzyme 1713,20a-hydroxysteroid dehydrogenase may be an important regulator of the local estrogen/progesterone ratio in fetal membranes around the time of parturition. (AM J OBSTET GVNECOL 1993;168:1377-85.) Key words: 17[3,20a-Hydroxysteroid dehydrogenase, fetal membranes, estrogen/progesterone ratio, parturition In most mammalian species estrogen stimulates changes that increase myometrial contractility, whereas progesterone causes myometrial quiescence' and the onset of labor is preceded by an increase in the estro- gen/progesterone ratio in the maternal circulation. These changes occur in species that are dependent on the placenta and in those dependent on the ovary for sex steroid synthesis during pregnancy, suggesting that this may be a common mechanism regulating labor onset in all species. The increase in estrogen/proges- terone ratio leads to several changes that stimulate uterine contractility. These changes include an increase in prostaglandin synthesis, an increased synthesis of myometrial oxytocin receptors, and an increase in my- ometrial gap junction formation. In humans the situation is unclear because measure- From the Department of Obstetrics and Gynecology, Centre for Research in Maternal, Fetal, and Newborn Health, University of Alberta. Supported by grants from the Medical Research Council of Canada (MA773l) and from the Alberta Heritage Foundation for Medical Research. Received for publication July 1, 1992; revised December 1, 1992; accepted December 16, 1992. Reprint requests: B.F. Mitchell, MD, Department of Obstetrics and Gynecology, University of Alberta Hospitals, Edmonton, Alberta, Canada T6G 2R7. Copyright © 1993 by Mosby - Year Book, Inc. 0002-9378/93 $1.00 + .20 6/1/45006 ments of circulating estrogen and progesterone in women during late pregnancy do not demonstrate any significant change in the absolute concentrations or in the ratio of estrogen to progesterone. However, both estrogen and progesterone are synthesized within hu- man amnion, chorion, and decidua. 2 • 3 These "fetal membranes" are also the major sites of synthesis of the stimulatory prostaglandins at parturition" Thus we and others have hypothesized that local changes in estrogen and progesterone synthesis, not reflected in the mater- nal circulation, may significantly influence the process of human parturition. The major estrogen formed by human fetal mem- branes is 'estrone! Estrone can be converted within these tissues to the much more active' estrogen, estra- diol, by the enzyme 17[3-hydroxysteroid dehydroge- nase. Locally synthesized progesterone can be con- verted within these tissues to the completely inactive metabolite 20a-dihydroprogesterone by the enzyme 20a-hydroxysteroid dehydrogenase. Strickler and To- bias 5 have demonstrated that a single site on the same enzyme catalyzes both reactions. They have termed this enzyme 17[3,20a-hydroxysteroid dehydrogenase. An in- crease in the reductive activity of this enzyme would both "activate" estrone to estradiol and "inactivate" progesterone to 20a-dihydroprogesterone, thus causing a marked increase in the "biologically active" estro- gen/progesterone ratio. Further, Gurpide and Marks 6 1377
Transcript of Changes in 17β,20α-hydroxysteroid dehydrogenase activity supporting an increase in the...

Changes in 17~,20a-hydroxysteroid dehydrogenase activity supporting an increase in the estrogen/progesterone ratio of human fetal membranes at parturition
Bryan F. Mitchell, MD, and Susan Wong, MSc
Edmonton, Alberta, Canada
OBJECTIVE: Our purpose was to measure the activity of the reversible enzyme 1713,20a-hydroxysteroid dehydrogenase around parturition with estrogen and progestogen substrates. STUDY DESIGN: Classic kinetic studies and explant cultures were used to determine kinetic parameters and net enzyme activities in both oxidative and reductive directions for both sets of substrates. RESULTS: Affinity constant values for estrone, estradiol, and 20a-dihydroprogesterone were 1 to 8 /-Lmol/L. Affinity constant for progesterone was 9 to 25 /-Lmol/L. Maximal velocities for all substrates in the chorion were 20- to 70-fold higher than in amnion and severalfold higher for estrogen substrates compared with the progestins. Around parturition there was a significant change toward net formation of the stronger estrogen (estradiol) and the weaker progestin (20a-dihydroprogesterone), suggesting an increase in the local estrogen/progesterone ratio. CONCLUSION: The enzyme 1713,20a-hydroxysteroid dehydrogenase may be an important regulator of the local estrogen/progesterone ratio in fetal membranes around the time of parturition. (AM J OBSTET GVNECOL 1993;168:1377-85.)
Key words: 17[3,20a-Hydroxysteroid dehydrogenase, fetal membranes, estrogen/progesterone ratio, parturition
In most mammalian species estrogen stimulates changes that increase myometrial contractility, whereas progesterone causes myometrial quiescence' and the onset of labor is preceded by an increase in the estrogen/progesterone ratio in the maternal circulation. These changes occur in species that are dependent on the placenta and in those dependent on the ovary for sex steroid synthesis during pregnancy, suggesting that this may be a common mechanism regulating labor onset in all species. The increase in estrogen/progesterone ratio leads to several changes that stimulate uterine contractility. These changes include an increase in prostaglandin synthesis, an increased synthesis of myometrial oxytocin receptors, and an increase in myometrial gap junction formation.
In humans the situation is unclear because measure-
From the Department of Obstetrics and Gynecology, Centre for Research in Maternal, Fetal, and Newborn Health, University of Alberta. Supported by grants from the Medical Research Council of Canada (MA773l) and from the Alberta Heritage Foundation for Medical Research. Received for publication July 1, 1992; revised December 1, 1992; accepted December 16, 1992. Reprint requests: B.F. Mitchell, MD, Department of Obstetrics and Gynecology, University of Alberta Hospitals, Edmonton, Alberta, Canada T6G 2R7. Copyright © 1993 by Mosby - Year Book, Inc. 0002-9378/93 $1.00 + .20 6/1/45006
ments of circulating estrogen and progesterone in women during late pregnancy do not demonstrate any significant change in the absolute concentrations or in the ratio of estrogen to progesterone. However, both estrogen and progesterone are synthesized within human amnion, chorion, and decidua. 2
• 3 These "fetal membranes" are also the major sites of synthesis of the stimulatory prostaglandins at parturition" Thus we and others have hypothesized that local changes in estrogen and progesterone synthesis, not reflected in the maternal circulation, may significantly influence the process of human parturition.
The major estrogen formed by human fetal membranes is 'estrone! Estrone can be converted within these tissues to the much more active' estrogen, estradiol, by the enzyme 17[3-hydroxysteroid dehydrogenase. Locally synthesized progesterone can be converted within these tissues to the completely inactive metabolite 20a-dihydroprogesterone by the enzyme 20a-hydroxysteroid dehydrogenase. Strickler and Tobias5 have demonstrated that a single site on the same enzyme catalyzes both reactions. They have termed this enzyme 17[3,20a-hydroxysteroid dehydrogenase. An increase in the reductive activity of this enzyme would both "activate" estrone to estradiol and "inactivate" progesterone to 20a-dihydroprogesterone, thus causing a marked increase in the "biologically active" estrogen/progesterone ratio. Further, Gurpide and Marks6
1377

1378 Mitchell and Wong
have demonstrated that this enzyme is an important regulator of stimulation of estrogen receptor activity, which has a major effect on the biologic potency of sex steroids in target tissues.
In the experiments reported here we investigated these enzyme activities with particular attention to changes that occur around the time of parturition. The experimental approach uses two different in vitro systems with amnion and chorion, two tissues that we previously have shown to have 17~,20a-hydroxysteroid dehydrogenase activity.s, 7 First, we used a biochemically idealized enzyme assay to determine the kinetic parameters of the enzyme with its various substrates. Second, we used a tissue explant culture system where the tissues are studied within their own endogenous matrix and thus may approximate the in vivo situation more closely than other systems." We have reasoned that, because 17~,20a-hydroxysteroid dehydrogenase is a reversible enzyme, rather than analyzing either the oxidative or reductive activity separately, it is the net activity that may have the most biologic relevance. We examined this net activity around the time of parturition with the explant culture system. Our results demonstrate changes supporting the hypothesis of a local paracrine system within human fetal membranes producing an increase in the estrogen/progesterone ratio of these tissues around the time of labor onset.
Material and methods Tissue preparation. These studies were approved by
the Research Ethics Board of the University of Alberta. Fetal membranes were collected from women who were delivered between 38 and 42 weeks' gestation by elective cesarean section before labor or by vaginal delivery after spontaneous onset of labor. There were no significant differences between these two groups with respect to mean gestational age, parity, or sex of the infant. All pregnancies, labors, and newborn infants were normal without evidence of infection or other complications. The placentas and associated fetal membranes were obtained immediately after delivery. The amnion was separated from the underlying choriodecidua and the tissues were rinsed several times with 150 mmol/L sodium chloride before transport to the laboratory. The adherent decidua then was dissected from the chorion.
Enzyme kinetics. To study the enzyme kinetics, the separated amnion and chorion tissues were blotted, minced, and homogenized with a Brinkmann Polytron (Brinkmann Instruments, Westbury, N.Y.; six 10-second bursts) with approximately 1 gm of tissue per 4 ml of 0.25 mol/L sucrose plus 10 mmol/L Tris buffer with 1.5 mmol/L ethylenediaminetetraacetic acid, pH 7.0. The resultant homogenate was centrifuged at 800g for 10 minutes, at 10,000g for 20 minutes, and then at 105,000g for 90 minutes (L8-80 Ultracentrifuge with a 75 Ti rotor, Beckman, Palo Alto, Calif.). The 105,000g
May 1993 Am J Obstet Gynecol
supernatant (cytosol) was kept at 4° C until use. The 105,000g pellet (microsomes) was resuspended in the same buffer at an approximate concentration of 1 gm original tissue per milliliter of buffer. Protein was quantified by the method Lowry et al. 9
Optimal conditions for the assay were established in preliminary experiments. Subsequent experiments were performed with 50 to 200 IJ.g microsomal protein or 200 to 600 IJ.g cytosolic protein in buffer at pH 7.0 for 15 minutes at 37° C. For CIS substrates, microsomal preparations were fourfold to fivefold more active with nicotinamide adenine dinucleotide cofactor than with nicotinamide adenine dinucleotide phosphate. Cytosolic activity was similar with either cofactor, so nicotinamide adenine dinucleotide was used in all experiments. For C2I substrates, nicotinamide adenine dinucleotide phosphate was the preferred cofactor. Cofactors were supplied at saturating concentrations of 1 mmol/L. When reduced cofactor was required, a generating system with glucose-6-phosphate (4 mmol/L) and glucose-6-phosphate dehydrogenase (1 unit) was included in the incubation media. To ensure solubility of the steroid substrate, dimethyl sulfoxide was used up to a final concentration of 5%. Preliminary experiments demonstrated that this concentration did not significantly alter enzyme activity. The reaction was stopped by adding 4 ml diethyl ether and vortexing immediately. Mter centrifugation, the ether phase was separated and the ether evaporated. Reconstituted steroids were separated on thin-layer silica plates (Whatman type LK6DF, Clifton, N.J.) with the solvent system chloroform/methanol, 98: 2. Steroid bands were visualized with 12 vapor. Reaction velocities were calculated by measuring the conversion of tritiumlabeled steroids. All cold steroids were purchased from Sigma, St. Louis, and all radiolabeled steroids were obtained from New England Nuclear, Boston. Corrections for recoveries were performed with carbon 14-labeled products. Tritium and 14C were measured on a Beckman model LS5000TD scintillation counter.
Saturation curves were constructed with seven concentrations in duplicate from 0.1 to 20 IJ.mol/L for estrone, estradiol, and 20a-dihydroprogesterone and 0.1 to 50 IJ.mol/L for progesterone. Kinetic parameters were calculated by means of Lineweaver-Burk analysis.
Tissue explants. The tissue explant experiments were performed as previously reported." Briefly, explants were excised with a sterile cork borer. Duplicate explants were incubated for 3 and 24 hours at 37° C in 1 ml Eagle's minimal essential medium, pH 7.4. Substrates were added to the incubation media in concentrations that reflect endogenous concentrations (2 IJ.mol/L for estrone and estradiol, 0.3 IJ.mol/L for progesterone, and 3.0 IJ.mol/L for 20a-dihydroprogesterone). To avoid end-product inhibition, only one substrate was used per assay tube, and the oxidative and

Volume 168, Number 5 Am J Obstet Gynecol
Mitchell and Wong 1379
Table I. Cross reactivites with other steroids for antisera to estradiol, progesterone, and 20a-dihydroprogesterone
Cross reactivity (%)
Estradiol Progesterone 20a-dihydroprogesterone Steroid antibody
Estrone 3,5 17j3-Estradiol 100 Estriol 3.0 17a-Estradiol 0.1 Estrone-3-sulfate 2.0 17j3-Estradiol-3-sulfate 6.0 Estriol-3-sulfate < 1.0 Cholesterol <0.1 Cortisone 0.2 Corticosterone <0.1 II-Deoxycorticosterone <0.1 Cortisol <0.1 11-Deoxycortisol <0.1 Pregnenolone <0.1 17a-Hydroxypregnenolone <0.1 Pregnenolone sulfate <0.1 20j3-Dihydroprogesterone <0.1 5a-Pregnanedione <0.1 Progesterone <0.1 17a-Hydroxyprogesterone <0.1 20a-Dihydroprogesterone <0.1 Dehydroepiandrosterone 0.1 Dehydroepiandrosterone-3-sulfate 0.1 Testosterone <0.1 17a-Epitestosterone <0.1 5-Androstene-3j3, 16a-diol-17-one <0.3 4-Androstenedione 5-Androstene-3j3. 16a-diol-17 -one-3-acetate
reductive activIties were measured in separate tubes. Steroid products were measured by radioimmunoassay.
Radioimmunoassays. The radioimmunoassays for estrone has been described previously. 7 Antisera to estradiol, progesterone, and 20a-dihydroprogesterone were produced by administering weekly subcutaneous injections of steroid-hemisuccinate-bovine serum albumin conjugates, together with Freund's incomplete adjuvant, to New Zealand White rabbits. The antisera cross reactivities, measured as the ability to displace standard at 50% binding, are shown in Table I. The radioimmunoassays were performed in phosphate-buffered saline solution, pH 7.0, incubated overnight at 4° C. Free and bound steroids were separated with dextran-coated charcoal. In all cases appropriate reagent and substrate blanks were subtracted from the measurements of reaction products. The sensitivities of the assays for estradiol, progesterone, and 20a-dihydroprogesterone were 12, 10, and 12 pg per tube, respectively. For all assays the interassay and intraassay coefficients of variation were < II %.
Preparation of radiolabeled 20a-dihydroprogesterone. Fetal sheep blood was used as a source of 20ahydroxysteroid dehydrogenase. Three milliliters of heparinized fetal sheep blood was centrifuged and the plasma removed. The remaining red blood cells were
0.1 0.1
antibody antibody
<0,1 <0,2 <0,1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2
3.0 <0.2 <0.1 <0.2 <0.1 <0.2
7.0 <0.2 <0.1 <0.2
9.0 <0.2 <0.1 <0.2 22.0 1.0
100 1.0 2.5 0.5 2.4 100
<0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2 <0.1 <0.2
dispersed in 3 ml of saline solution to which tritium- or 14C-progesterone was added. This was incubated at 37° C in a shaking water bath for 90 minutes. Steroids were extracted with 4 x 2 volumes of diethyl ether. Mter centrifugation, the ether fraction was separated and dried under nitrogen. The residue was reconstituted in 50 I.d ethanol and subjected to chromatography on silica gel with the solvent system chloroformdiethyl ether, 100: 30. The band corresponding to 20adihydroprogesterone was scraped and the steroid eluted with ethyl acetate. The ethyl acetate was evaporated under nitrogen and a stock solution of the purified radiolabeled 20a-dihydroprogesterone kept at - 20° C in ethanol. This method allows for approximately 75% of the original radiolabel to be recovered as 20a-dihydroprogesterone.
Statistical analyses. Comparisons between the cesarean section and spontaneous labor groups were performed with the Student unpaired, two-tailed t test. Where Bartlett's test revealed heterogeneity of variance, the Mann-Whitney U test was used.
Results
The characterization data for the subcellular fraction enzyme assays using estrone as substrate are illustrated in Fig. 1. For both the microsomal and cytosol fractions
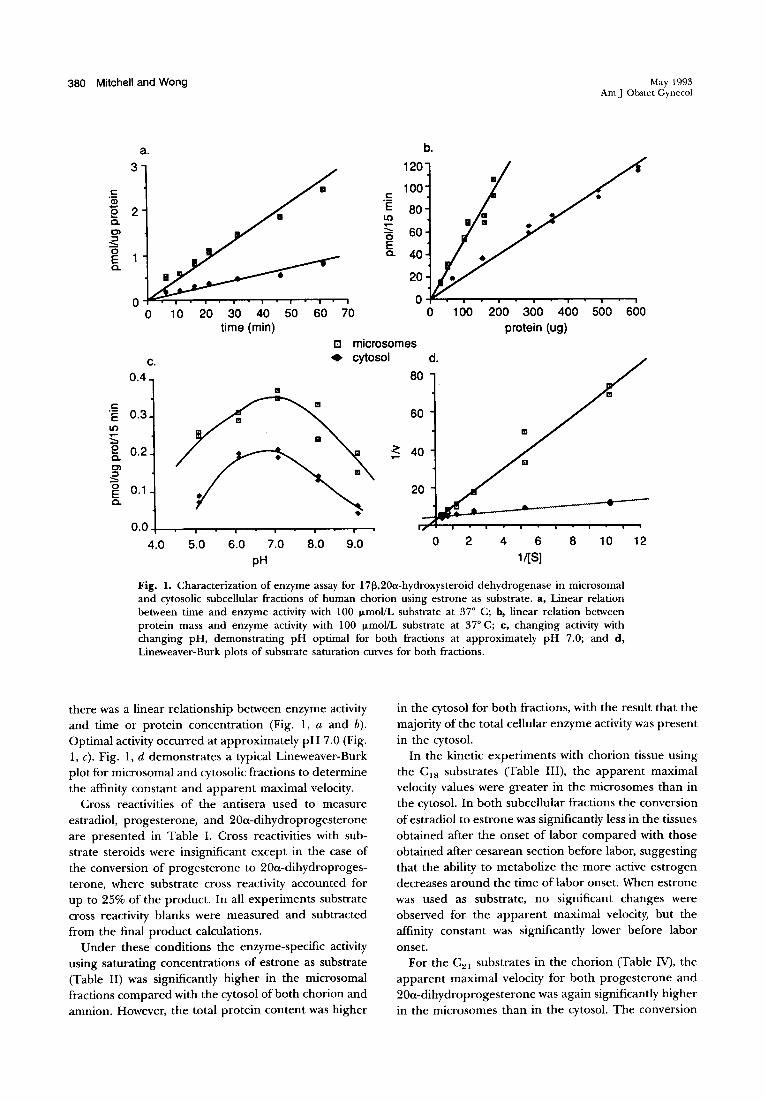
380 Mitchell and Wong
c: 'ij)
a. 3
e 2 a. Ol -2 (5 E a.
o 10 20 30 40 50 60 70 time (min)
c:
120
100
'E 80 LO .... =a 60 E a. 40
20
b.
May 1993 Am J Obstet Gynecol
100 200 300 400 500 600 protein (ug)
c. 0.4
[:J microsomes • cytosol
c: 'E 0.3 LO ;; ~ 0.2 Ol -2 (5 E 0.1 a.
4.0 5.0
I!I
6.0 7.0 pH
8.0
80
60
~ 40
20
9.0 o 2 4 6 1/[8]
8 10 12
Fig. 1. Characterization of enzyme assay for 1713,20a-hydroxysteroid dehydrogenase in microsomal and cytosolic subcellular fractions of human chorion using estrone as substrate. a, Linear relation between time and enzyme activity with 100 f.LmoVL substrate at 37° C; b, linear relation between protein mass and enzyme activity with 100 f.LmoVL substrate at 37° C; c, changing activity with changing pH, demonstrating pH optimal for both fractions at approximately pH 7.0; and d, Lineweaver-Burk plots of substrate saturation curves for both fractions.
there was a linear relationship between enzyme activity and time or protein concentration (Fig. 1, a and b). Optimal activity occurred at approximately pH 7.0 (Fig. 1, c). Fig. 1, d demonstrates a typical Lineweaver-Burk plot for microsomal and cytosolic fractions to determine the affinity constant and apparent maximal velocity.
Cross reactivities of the antisera used to measure estradiol, progesterone, and 20u-dihydroprogesterone are presented in Table 1. Cross reactivities with substrate steroids were insignificant except in the case of the conversion of progesterone to 20u-dihydroprogesterone, where substrate cross reactivity accounted for up to 25% of the product. In all experiments substrate cross reactivity blanks were measured and subtracted from the final product calculations.
Under these conditions the enzyme-specific activity using saturating concentrations of estrone as substrate (Table II) was significantly higher in the microsomal fractions compared with the cytosol of both chorion and amnion. However, the total protein content was higher
in the cytosol for both fractions, with the result that the majority of the total cellular enzyme activity was present in the cytosol.
In the kinetic experiments with chorion tissue using the CIS substrates (Table III), the apparent maximal velocity values were greater in the microsomes than in the cytosol. In both subcellular fractions the conversion of estradiol to estrone was significantly less in the tissues obtained after the onset of labor compared with those obtained after cesarean section before labor, suggesting that the ability to metabolize the more active estrogen decreases around the time of labor onset. When estrone was used as substrate, no significant changes were observed for the apparent maximal velocity, but the affinity constant was significantly lower before labor onset.
For the C21 substrates in the chorion (Table IV), the apparent maximal velocity for both progesterone and 20u-dihydroprogesterone was again significantly higher in the microsomes than in the cytosol. The conversion

Volume 168. Number" Am J Obstet Gynecol
Mitchell and Wong 1381
Table II. Distribution of 1713.20a-hydroxysteroid dehydrogenase activity with estrone used as substrate (20 f.LmoVL) between microsomal and cytosolic subcellular fractions of human chorion and amnion
Specific activity (pmoll JLg Total protein protein/15 min) (mglfraction)
Chorion Microsomes 2.39 ± 0.2S* 21.4 ± 2.1* Cytosol 0.73 ± 0.10 97.6 ± 6.9
Amnion Microsomes 0.97 ± 0.16* 7.4 ± O.S* Cytosol O.SS ± 0.12 S9.1 ± S.4
Data are expressed as mean ± SEM of data from 14 experiments with chorion and II with amnion. *Significant difference (p < O.OS) between microsomal and cytosolic fractions.
Percent of total enzyme activity
41.S SS.2
IS.I S1.9
Table III. Kinetic parameters for interconversion of estrone and estradiol by subcellular fractions of human chorion
Microsomes Cytosol
Substrate
Affinity constant (J.LmoVL) Spontaneous labor Cesarean section Total group
Apparent maximal velocity (pmoVJ.Lg protein/IS min) Spontaneous labor Cesarean section Total group
Estrone
6.0 ± 1.6 (9)* 3.1 ± O.S (S) S.O ± l.l (14)
2.7 ± O.S (9) 3.S ± 1.3 (S) 3.0 ± 0.7 (14)
I Estradiol
1.9 ± 0.2 (7) 2.4 ± 0.4 (S) 2.1 ± 0.2 (12)
2.1 ± O.S (7)* 4.2 ± 1.0 (S) 3.0 ± 0.7 (12)
Estrone
9.S ± 0.6 (10)* 2.7 ± 0.4 (S) 4.1 ± O.S (IS)
1.4 ± 0.3 (10) I.S ± 0.4 (S) 1.4 ± 0.3 (lS)t
Data are expressed as mean ± SEM. Numbers in parentheses indicate the number of experiments. *Significant difference (P < O.OS) between spontaneous labor and cesarean section groups.
I Estradiol
3.3 ± 0.4 (6) 4.1 ± 0.7 (S) 3.7 ± 0.4 (11)t
O.S ± 0.1 (6)* 1.4 ± 0.3 (S) 0.9 ± 0.2 (ll)t
tSignificant difference (P < O.OS) between microsomal and cytosolic fractions using the same substrates.
Table IV. Kinetic parameters for interconversion of progesterone and 20a-dihydroprogesterone by subcellular fractions of human chorion
Substrate Progesterone
Microsomes
I 20a-dihydroprogesterone
Cytosol
Progesterone I 20a-dihydroprogesterone
Affinity constant (J.LmoVL) Spontaneous labor Cesarean section Total group
12.7 ± 3.1 (7) 11.4 ± 3.1 (7) 12.0 ± 2.1 (14)
3.1 ± O.S (S) 4.1 ± 0.9 (7) 3.7 ± 0.6 (12)
19.7 ± 6.4 (7) 12.S ± 2.3 (7) 16.1 ± 3.4 (14)
3.S ± 1.6 (4) S.I ± 0.9 (6) 4.4 ± 0.9 (10)
Apparent maximal velocity (pmoVJ.Lg protein/IS min) Spontaneous labor Cesarean section Total group
0.09 ± 0.02 (7) 0.17 ± O.OS (7) 0.13 ± 0.04 (14)
0.S7 ± 0.22 (S) 0.42 ± 0.12 (7) O.4S ± 0.11 (12)
0.02 ± 0.01 (7) 0.02 ± 0.00 (7) 0.02 ± 0.00 (14)*
0.03 ± 0.01 (4) 0.04 ± 0.01 (6) 0.04 ± O.oJ (10)*
Data are expressed as mean ± SEM. Numbers in parentheses indicate the number of experiments. *Significant difference (p < O.OS) between microsomal and cytosolic fractions using the same substrates.
of 20a-dihydroprogesterone to progesterone was significantly greater than the reverse reaction in both microsomal and cytosol fractions, suggesting that the chorion favors production of the more potent progestogen. The apparent maximal velocity values with the C21
substrates were consistently manyfold lower than with CIS substrates. In both subcellular fractions the affinity
constant for progesterone was significantly higher than for 20a-dihydroprogesterone or for either of the CIS
substrates. No differences in any of the kinetic parameters were detected between the cesarean section and spontaneous labor subgroups with C21 substrates.
It may be noteworthy that, in general, the reduced substrates (estradiol and 20a-dihydroprogesterone)
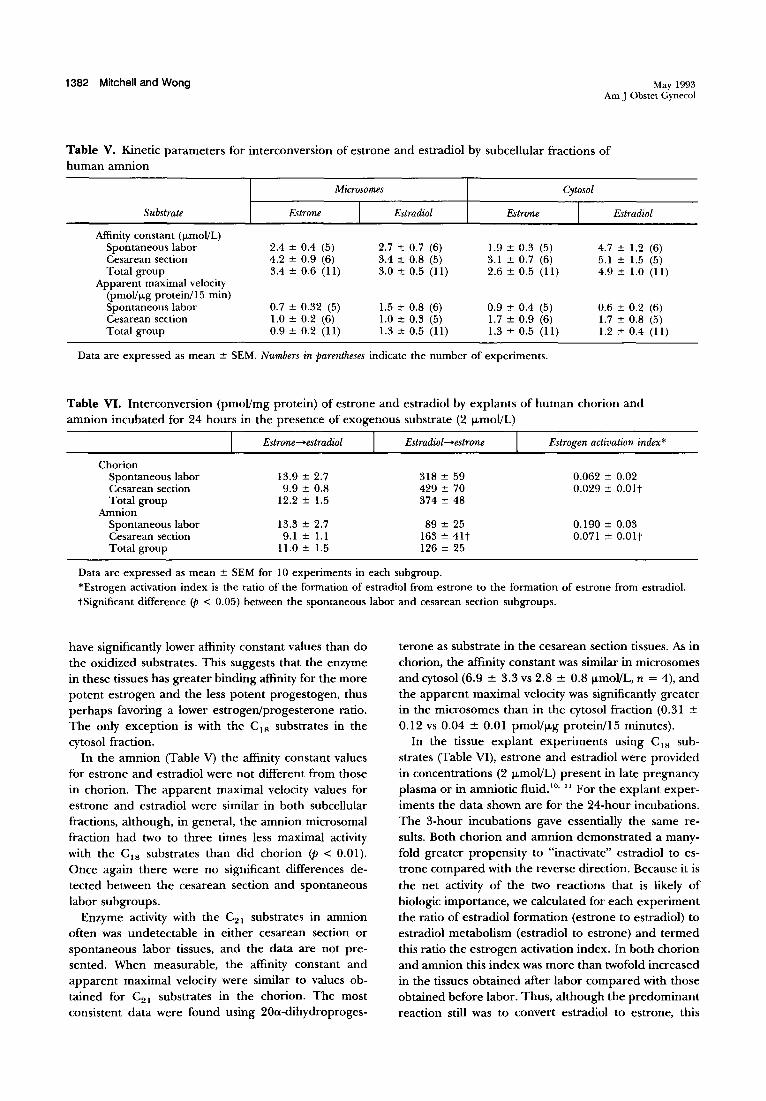
1382 Mitchell and Wong May 1993 Am J Obstet Gynecol
Table V. Kinetic parameters for interconversion of estrone and estradiol by subcellular fractions of human amnion
Microsomes Cytosol
Substrate Estrone I Estradiol Estrone I Estradiol
Affinity constant (ILmoI!L) Spontaneous labor 2.4 ± 0.4 (5) 2.7 ± 0.7 (6) 1.9 ± 0.3 (5) 4.7 ± 1.2 (6) Cesarean section 4.2 ± 0.9 (6) 3.4 ± 0.8 (5) 3.1 ± 0.7 (6) 5.1 ± 1.5 (5) Total group 3.4 ± 0.6 (11) 3.0 ± 0.5 (11) 2.6 ± 0.5 (11) 4.9 ± 1.0 (11)
Apparent maximal velocity (pmoI!lLg protein/I 5 min) Spontaneous labor 0.7 ± 0.32 (5) 1.5 ± 0.8 (6) 0.9 ± 0.4 (5) 0.6 ± 0.2 (6) Cesarean section 1.0 ± 0.2 (6) 1.0 ± 0.3 (5) 1.7 ± 0.9 (6) 1.7 ± 0.8 (5) Total group 0.9 ± 0.2 (II) 1.3 ± 0.5 (11) 1.3 ± 0.5 (11) 1.2 ± 0.4 (11)
Data are expressed as mean ± SEM. Numbers in parentheses indicate the number of experiments.
Table VI. Interconversion (pmoVmg protein) of estrone and estradiol by explants of human chorion and amnion incubated for 24 hours in the presence of exogenous substrate (2 f.\-moVL)
Chorion Spontaneous labor Cesarean section Total group
Amnion Spontaneous labor Cesarean section Total group
Estrone-+estradiol
13.9 ± 2.7 9.9 ± 0.8
12.2 ± 1.5
13.3 ± 2.7 9.1 ± 1.1
11.0± 1.5
Estradiol-+estrone
318 ± 59 429 ± 70 374 ± 48
89 ± 25 163 ± 41t 126 ± 25
Estrogen activation index*
0.062 ± 0.02 0.029 ± O.Olt
0.190 ± 0.03 0.071 ± O.Olt
Data are expressed as mean ± SEM for 10 experiments in each subgroup. *Estrogen activation index is the ratio of the formation of estradiol from estrone to the formation of estrone from estradiol. tSignificant difference (p < 0.05) between the spontaneous labor and cesarean section subgroups.
have significantly lower affinity constant values than do the oxidized substrates. This suggests that the enzyme in these tissues has greater binding affinity for the more potent estrogen and the less potent progestogen, thus perhaps favoring a lower estrogen/progesterone ratio. The only exception is with the CIS substrates in the cytosol fraction.
In the amnion (Table V) the affinity constant values for estrone and estradiol were not different from those in chorion. The apparent maximal velocity values for estrone and estradiol were similar in both subcellular fractions, although, in general, the amnion microsomal fraction had two to three times less maximal activity with the CIS substrates than did chorion <p < 0.01). Once again there were no significant differences detected between the cesarean section and spontaneous labor subgroups.
Enzyme activity with the C21 substrates in amnion often was undetectable in either cesarean section or spontaneous labor tissues, and the data are not presented. When measurable, the affinity constant and apparent maximal velocity were similar to values obtained for C21 substrates in the chorion. The most consistent data were found using 20a-dihydroproges-
terone as substrate in the cesarean section tissues. As in chorion, the affinity constant was similar in microsomes and cytosol (6.9 ± 3.3 vs 2.8 ± 0.8 f.\-moVL, n = 4), and the apparent maximal velocity was significantly greater in the microsomes than in the cytosol fraction (0.31 ±
0.12 vs 0.04 ± 0.01 pmoVf.\-g protein/15 minutes). In the tissue explant experiments using CIS sub
strates (Table VI), estrone and estradiol were provided in concentrations (2 f.\-moVL) present in late pregnancy plasma or in amniotic fiuid.'o, II For the explant experiments the data shown are for the 24-hour incubations. The 3-hour incubations gave essentially the same results. Both chorion and amnion demonstrated a manyfold greater propensity to "inactivate" estradiol to estrone compared with the reverse direction. Because it is the net activity of the two reactions that is likely of biologic importance, we calculated for each experiment the ratio of estradiol formation (estrone to estradiol) to estradiol metabolism (estradiol to estrone) and termed this ratio the estrogen activation index. In both chorion and amnion this index was more than twofold increased in the tissues obtained after labor compared with those obtained before labor. Thus, although the predominant reaction still was to convert estradiol to estrone, this
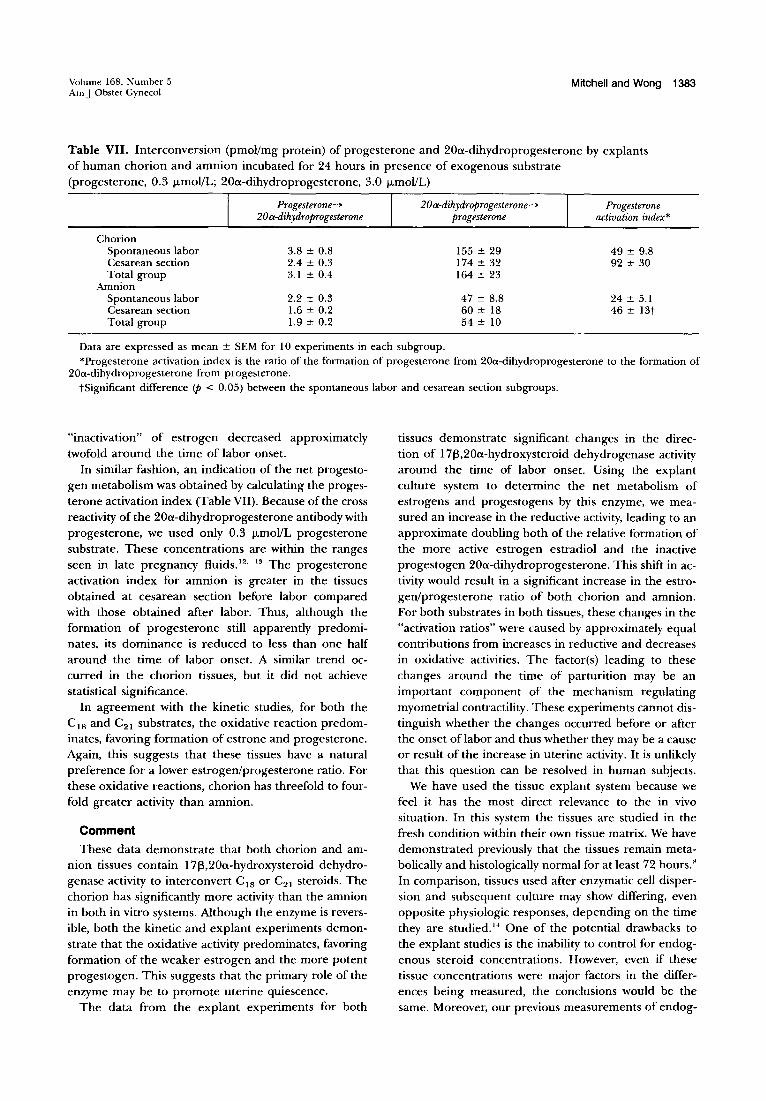
Volume 168. Number 5 Am J Obstet Gynecol
Mitchell and Wong 1383
Table VII. Interconversion (pmoVmg protein) of progesterone and 20a-dihydroprogesterone by explants of human chorion and amnion incubated for 24 hours in presence of exogenous substrate (progesterone, 0.3 /-LmoVL; 20a-dihydroprogesterone, 3.0 /-LmoVL)
Chorion Spontaneous labor Cesarean section Total group
Amnion Spontaneous labor Cesarean section Total group
Progesterone~
20a-dihydroprogesterone
3.8 ± 0.8 2.4 ± 0.3 3.1 ± 0.4
2.2 ± 0.3 1.6 ± 0.2 1.9 ± 0.2
20a-dihydroprogesterone--> progesterone
155 ± 29 174 ± 32 164 ± 23
47 ± 8.8 60 ± 18 54 ± 10
Progesterone activation index*
49 ± 9.8 92 ± 30
24 ± 5.1 46 ± 13t
Data are expressed as mean ± SEM for 10 experiments in each subgroup. *Progesterone activation index is the ratio of the formation of progesterone from 200l-dihydroprogesterone to the formation of
200l-dihydroprogesterone from progesterone. tSignificant difference <p < 0.05) between the spontaneous labor and cesarean section subgroups.
"inactivation" of estrogen decreased approximately twofold around the time of labor onset.
In similar fashion, an indication of the net progestogen metabolism was obtained by calculating the progesterone activation index (Table VII). Because of the cross reactivity of the 20a-dihydroprogesterone antibody with progesterone, we used only 0.3 /-LmoVL progesterone substrate. These concentrations are within the ranges seen in late pregnancy fluids. 12. 13 The progesterone activation index for amnion is greater in the tissues obtained at cesarean section before labor compared with those obtained after labor. Thus, although the formation of progesterone still apparently predominates, its dominance is reduced to less than one half around the time of labor onset. A similar trend occurred in the chorion tissues, but it did not achieve statistical significance.
In agreement with the kinetic studies, for both the CIS and C 21 substrates, the oxidative reaction predominates, favoring formation of estrone and progesterone. Again, this suggests that these tissues have a natural preference for a lower estrogen/progesterone ratio. For these oxidative reactions, chorion has threefold to fourfold greater activity than amnion.
Comment
These data demonstrate that both chorion and amnion tissues contain I 7~,20a-hydroxysteroid dehydrogenase activity to interconvert CIS or C21 steroids. The chorion has significantly more activity than the amnion in both in vitro systems. Although the enzyme is reversible, both the kinetic and explant experiments demonstrate that the oxidative activity predominates, favoring formation of the weaker estrogen and the more potent progestogen. This suggests that the primary role of the enzyme may be to promote uterine quiescence.
The data from the explant experiments for both
tissues demonstrate significant changes in the direction of 17~,20a-hydroxysteroid dehydrogenase activity around the time of labor onset. Using the explant culture system to determine the net metabolism of estrogens and progestogens by this enzyme, we measured an increase in the reductive activity, leading to an approximate doubling both of the relative formation of the more active estrogen estradiol and the inactive progestogen 20a-dihydroprogesterone. This shift in activity would result in a significant increase in the estrogen/progesterone ratio of both chorion and amnion. For both substrates in both tissues, these changes in the "activation ratios" were caused by approximately equal contributions from increases in reductive and decreases in oxidative activities. The factor(s) leading to these changes around the time of parturition may be an important component of the mechanism regulating myometrial contractility. These experiments cannot distinguish whether the changes occurred before or after the onset oflabor and thus whether they may be a cause or result of the increase in uterine activity. It is unlikely that this question can be resolved in human subjects.
We have used the tissue explant system because we feel it has the most direct relevance to the in vivo situation. In this system the tissues are studied in the fresh condition within their own tissue matrix. We have demonstrated previously that the tissues remain metabolically and histologically normal for at least 72 hours." In comparison, tissues used after enzymatic cell dispersion and subsequent culture may show differing, even opposite physiologic responses, depending on the time they are studied. 14 One of the potential drawbacks to the explant studies is the inability to control for endogenous steroid concentrations. However, even if these tissue concentrations were major factors in the differences being measured, the conclusions would be the same. Moreover, our previous measurements of endog-

1384 Mitchell and Wong
enous whole-tissue steroid concentrations do not demonstrate significant changes occurring around the time of parturition. 15 These measurements were made with crude homogenates of whole tissues and could not be corrected for procedural losses. Such measurements may· be of little biologic relevance, because the most important steroid concentrations are likely those at very specific subcellular sites. Concentrations at these critical sites may change significantly without being reflected in whole-tissue concentrations. Although formation Of the weaker estrogen and stronger progestogen still predominates after spontaneous labor onset, our data suggest a shift occurs, favoring a relative increase in the more active estrogen and a decrease in the more potent progestogen. This may be reflected in amniotic fluid, where a significant increase in estradiol and a decrease in progesterone around the time of labor onset occurs.16 This would be compatible with our current findings. Together, these data support the hypothesis of a paracrine system within fetal membranes that increases the local estrogen/progesterone ratio at parturition.
Although the kinetic studies do not reveal major changes in the kinetic parameters of 17~,20a-hydroxysteroid dehydrogenase activity around the time of labor onset, in chorion, the most active of the tissues, the apparent maximal velocity with estradiol as substrate in both micro somes and cytosol was significantly lower after labor than before. This again suggests that a shift in equilibrium toward production of the more active estrogen (estradiol) occurs around the time of labor onset.
Our data demonstrated the enzyme in both the cytosolic and microsomal subcellular fractions of both chorion and amnion. When estrone is used as substrate, the data suggest most of the activity is in the cytosol, although the highest specific activity is in the microsomes. In the placenta, where this enzyme has been studied most extensively, controversy remains about the subcellular distribution of the enzyme. Strickler et al. 17 found 17~,20a-hydroxysteroid dehydrogenase activity only in the cytosol fraction and, in agreement with our data, found the enzyme manyfold more active for C 18
than for C21 substrates.5. 6 Blomquist et aI. IS have also measured the enzyme in placental microsomes and provided compelling kinetic data suggesting that there were two microsomal enzymes - a nonspecific one with similar affinities for C 18' C 19' and C21 steroids and a more specific one interconverting only CI8 substrates and accounting for approximately 30% of the total C 18
activity. Our data do not allow speculation on the presence of
multiple enzymes. However, the apparent maximal velocity for both C I8 and C21 substrates was significantly higher in the microsomes compared with the cytosol.
May 1993 Am J Obstet Gynecol
These findings could be compatible with two separate enzymes or with a conformational change in the microenvironment, leading to an increase in the maximal velocity of a single enzyme. If the latter explanation is correct, then the regulation of translocation of the enzyme between the cytosol and the microsomal membranes may be an important factor in regulating the enzyme activity.
For the placental enzyme the reported affinity constant for estradiol is 10 to 15 /-Lmol!L,6 somewhat higher than our values for the amnion and chorion (1 to 5 /-Lmol!L). When 20a-dihydroprogesterone is used as substrate, the affinity constant of the cytosolic enzyme in the placenta has been reported to be 62 to 208 /-Lmol!U9 compared with our value of 4.4 ± 0.9 mmol!L in chorion. However, the affinity constant of the placental microsomal enzyme has been reported at 2.3 /-Lmol! L, which is similar to our value of 3.7 ± 0.6 /-Lmol!L in chorion. IS
With the cytosol fraction from placenta, Diaz-Zagoya and Arias20 noted a decrease in progesterone conversion to 20a-dihydroprogesterone associated with labor onset, a result compatible with our explant data. Using crude homogenates of placenta, Milewich et aF I measured a fivefold increase in this activity comparing tissues obtained at 12 to 20 weeks' gestation with those obtained after vaginal delivery. However, no estimations of enzyme activity were made immediately before and after labor. In contrast, the same investigators, using crude homogenates of amnion and chorion tissue, noted the enzyme activity decreased 10- 20-fold from mid pregnancy to term, with no change noted between the spontaneous labor and cesarean section groupS.22 These findings suggested the regulation of this enzyme in placenta and fetal membranes may be quite different and thus possibly under local control.
Many studies have addressed the regulation of this enzyme in human placental tissue. Several CI8 , C 19,
and C21 steroids will inhibit the activity of the placental enzyme, usually demonstrating competitive kinetics!' Whereas prostaglandins of the E and F series cause inhibition of the oxidation of 20a-dihydroprogesterone, the reverse reaction is stimulated by prostaglandin F2a.
This increase can be blocked by cycloheximide, suggesting it involves synthesis of new enzyme.24 Additionally, cofactor availability may be a determinant of the net activity.25
Thus it is likely that several locally produced factors may regulate the velocity and direction of 17~,20ahydroxy steroid dehydrogenase. It is not known what factors playa role in regulating the redox environment of this enzyme in human amnion and chorion and hence may influence the estrogen/progesterone ratio in these tissues around the time of parturition.
Abnormalities of the timing of parturition remain a

Volume 168, Number 5 Am J Obstet Gynecol
m£tior problem in clinical obstetrics. It is unlikely that further significant improvements in results will be made in this area until we achieve a better understanding of the physiologic regulation of normal parturition. Animal studies suggest that the estrogen/progesterone ratio within the pregnant uterus is an important factor in regulating myometrial contractility. Our data suggest that this may also apply in the human. If so, it is likely that the changes occur in a paracrine system within intrauterine tissues in which the enzyme 17(3,20a-hydroxysteroid dehydrogenase may play a critical role. Further studies of the function and regulation of this enzyme may yield improved understanding of this important physiologic process.
REFERENCES 1. Challis ]RG, Mitchell BF. Steroid production by the fetal
membranes in relation to the onset of parturition. In: McNellis D, Challis ]RG, MacDonald PC, Nathanielsz PW, Roberts ]M, eds. The onset of labour: cellular and integrative mechanisms. Ithaca, New York: Perinatology Press, 1988:233-53.
2. Romano WM, Lukash LA, Challis JRG, Mitchell BF. Substrate utilization for estrogen synthesis by human fetal membranes and decidua. AM] OBSTET GYNECOL 1986;155: 1170-5.
3. Mitchell BF, Challis JRG, Lukash L. Progesterone synthesis by human amnion, chorion, and decidua at term. AM J OBSTET GYNECOL 1987;157:349-53.
4. Olson DM, Skinner K, Challis JRG. Prostaglandin output in relation to parturition by cells dispersed from human intrauterine tissues.J Clin Endocrinol Metab 1983;57:694-9.
5. Strickler RC, Tobias B. Estradiol 1713-dehydrogenase and 20a-hydroxysteroid dehydrogenase from human placental cytosol: one enzyme with two activities? Steroids 1980;36: 243-53.
6. Gurpide E, Marks C. Influence of endometrial 1713-hydroxysteroid dehydrogenase activity on the binding of estradiol to receptors. J Clin Endocrinol Metab 1981;52: 252-5.
7. Mitchell BF, CrossJ, Hobkirk R, ChallisJRG. Formation of unconjugated estrogens from estrone sulfate by dispersed cells from human fetal membranes and decidua . .1 Clin Endocrinol Metab 1984;58:845-9.
8. Mitchell BF, Powell WA. Progesterone production by human fetal membranes: an in vitro incubation system for studying hormone production and metabolism. AM J OBSTET GYNECOL 1984; 148:303-9.
9. Lowry OH, Rosebrough NJ, Farr LA, Randall RH. Protein measurement with Folin phenol reagent. ] Bioi Chern 1951; 1 93:265-75.
10. Loriaux DL, Ruder HL], Knab DR, Lipsett MB. Estrone sulfate, estrone, estradiol and estriol plasma levels in
Mitchell and Wong 1385
human pregnancy. J Clin Endocrinol Metab 1972;35:887-91.
11. Turnbull AC, Anderson ABM, Flint APF,Jeremy JY, Keirse MJNC, Mitchell MD. Human parturition. In: Knight .1, O'Connor M, eds. The fetus and birth. Ciba Foundation Symposium 47. Amsterdam: Elsevier, 1977 :427 -60.
12. Scommegna A, Burd L, Bieniarz J. Progesterone and pregnenolone sulfate in pregnancy plasma. AM .1 OBSTF:r GYNECOL 1972;113:60-5.
13. Runnebaum B, Stober 1, Zander.J. Progesterone, 20a- and 2013-dihydroprogesterone in mother and child at birth. Acta Endocrinol (Copenh) 1975;80:558-68.
14. Gibb W, Lavoie J C. Effect of glucocorticoids on prostaglandin formation by human amnion. CanJ Physiol Pharmacol 1990;68:671-6.
15. Mitchell BF, Challis JRG. Estrogen and progesterone metabolism in human fetal membranes. In: Mitchell BF, ed. The physiology and biochemistry of human fetal membranes. Ithaca, New York: Perinatology Press, 1988:5-28.
16. Romero R, Scoccia B, Mazor M, Wu YK, Benveniste R. Evidence for a local change in progesterone/estrogen ratio in human parturition at term. AM J OBSTET GYNECOL 1988; 159: 114-9.
17. Strickler RC, Tobias B. 20a-Hydroxysteroid dehydrogenase and 1713-estradiol dehydrogenase localize in cytosol of human term placenta. Am J Physiol 1982;242:EI78-83.
18. Blomquist CH, Lindemann NJ, Hakanson EY. 1713-Hydroxysteroid and 20a-hydroxysteroid dehydrogenase activities of human placental microsomes: kinetic evidence for two enzymes differing in substrate specificity. Arch Biochem Biophys 1985;239:206-15.
19. Rabe T, Kiesel L, Runnebaum B. Partial characterization of the cytoplasmic 20a-hydroxysteroid dehydrogenase (EC 1.1.1.149) of the human placenta at term. J Steroid Biochern 1982;16:737-43.
20. Diaz-Zagoya JC, Arias F. Synthesis and catabolism of progesterone in placentas from normotensive and severely hypertensive patients before and after parturition. AM J OBSTET GYNECOL 1981;141:637-40.
21. Milewich L, Gant NF, Schwarz BE, Chen GT, MacDonald PC. Initiation of human parturition IX. Progesterone metabolism by placentas of early and late human gestation. Obstet Gynecol 1978;51 :278-80.
22. Milewich L, Gant NF, Schwarz BE, Chen GT, MacDonald PC. Initiation of human parturition. VIII. Metabolism of progesterone by fetal membranes of early and late gestation. Obstet Gynecol 1977;50:45-8.
23. Rabe T, Kiesel L, Runnebaum B. Regulation of human placental progesterone synthesis in vitro by naturally occurring steroids. J Steroid Biochem 1985;22:657-64.
24. Kiesel L, Rabe T, Runnebaum B. The role of prostaglandins, cyclic nucleotides and tricarboxylic acids in the regulation of the human placental 20a-hydroxysteroid dehydrogenase in vitro. Steroids 1982;40:99-108.
25. Blomquist CH, Hakanson EY. Pyridine nucleotide levels under conditions of 5a-dihydrotestosterone-stimulated 1713-estradiol formation from estrone and pathway of nicotinamide adenine dinucleotide biosynthesis in placental villi in vitro. J Clin Endocrinol Metab 1991;73:140-5.