Cepharanthine and Piperine ameliorate diabetic … Department of Biochemistry, Faculty of Pharmacy,...
13
Cepharanthine and Piperine ameliorate diabetic nephropathy in rats: role of NF-κB and NLRP3 inflammasome Yara A. Samra a, ⁎, Heba S. Said b , Nehal M. Elsherbiny a , Gregory I. Liou c , Mamdouh M. El-Shishtawy a , Laila A. Eissa a, ⁎ a Department of Biochemistry, Faculty of Pharmacy, Mansoura University, Mansoura 35516, Egypt b Department of Microbiology, Faculty of Pharmacy, Mansoura University, Mansoura 35516, Egypt c Department of Ophthalmology, Augusta University, Augusta, GA 30912, USA abstract article info Article history: Received 1 February 2016 Received in revised form 31 May 2016 Accepted 2 June 2016 Available online 3 June 2016 Aims: Hyperglycemia leads to elevation of oxidative stress and proinflammatory cytokines which are the main causes of diabetic nephropathy (DN). NLRP3 inflammasome and thioredoxin-interacting protein (TXNIP) are re- cently assumed to participate in the development of DN. We aimed to investigate the effects of Cepharanthine (CEP), Piperine (Pip) and their combination in streptozotocin (STZ)-induced DN focusing on their role to modu- late NLRP3 and TXNIP induced inflammation. Main methods: Diabetic rats were treated with intraperitoneal (i.p.) injection of CEP (10 mg/kg/day), Pip (30 mg/kg/ day) or their combination for 8 weeks. Nuclear factor kappa B (NF-κB), tumor necrosis factor alpha (TNF-α) and in- terleukin-1β (IL-1β) were assessed by ELISA technique. TXNIP and NLRP3 genes expressions were evaluated by real time-PCR. Key findings: Diabetic rats showed significant increase in renal TXNIP and NLRP3 expression. CEP, Pip or their com- bination significantly decreased TXNIP and NLRP3 expression in diabetic kidneys. Hyperglycemia induced NF-κB ac- tivation leading to increased IL-1β and TNF-α levels. CEP, Pip or their combination showed significant inhibition of NF-κB together with decreased IL-1β and TNF-α levels in diabetic rats. Also, diabetic rats showed significant de- crease in creatinine clearance and increase in blood glucose, serum creatinine, blood urea nitrogen, malondialdehyde, proteinuria, and kidney weight to body Weight ratio. All of these changes were reversed by CEP, Pip or their combination. Significance: The antioxidant and anti-inflammatory effects of CEP and Pip which were accompanied by inhibition of NF-κB and NLRP3 activation might be helpful mechanisms to halt the progression of DN. © 2016 Elsevier Inc. All rights reserved. Keywords: Cepharanthine Piperine Diabetes Renal injury NF-κB TXNIP NLRP3 1. Introduction Diabetes mellitus is a chronic inflammatory disease in which hyper- glycemia enhances glucose oxidation and production of advanced glycation end products (AGEs). Hyperglycemia activates protein kinase C, polyol and hexosamine pathways and upregulates the production of different proinflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin 6 (IL-6) and interleukin-1β (IL-1β) [1]. The pathophysiological changes in diabetes also activate nuclear factor kappa B (NF-κB) which perturbs more inflammatory cytokines produc- tion suggesting that NF-κB has a key role in the progression of diabetes- induced inflammation [2,3]. Diabetic nephropathy (DN) is the main complication of diabetes that is associated with increased cardiovascular mortality. DN is clinically characterized by the hypertension and albuminuria which further pro- gresses to decline in renal function and leads to end-stage renal failure [4]. Current treatment strategy of DN involves glycemic and blood pres- sure control; however, patients continue to suffer from advanced renal damage [5]. Therefore, new therapeutic modalities should be intro- duced to prevent the progression of DN. NLRP3 inflammasome is a typical member of inflammasome large family that is present in mammalian cells directing renal inflammatory processes leading to podocyte injury and glomerulosclerosis [6]. NALP3 inflammasome is composed of NLRP3 (NACHT, LRR and PYD domains- containing protein 3), ASC (apoptosis-associated speck like protein) and caspase-1 [7,8]. Inflammasome is significantly involved in the path- ogenesis of DN through the production of the proinflammatory cyto- kines IL-1β and IL-18 as several NLRs play an important role in the cleavage of pro-IL-1β and pro-IL-18 into mature forms of IL-1β and IL- Life Sciences 157 (2016) 187–199 ⁎ Corresponding author. E-mail addresses: [email protected] (Y.A. Samra), [email protected] (L.A. Eissa). http://dx.doi.org/10.1016/j.lfs.2016.06.002 0024-3205/© 2016 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Life Sciences journal homepage: www.elsevier.com/locate/lifescie
Transcript of Cepharanthine and Piperine ameliorate diabetic … Department of Biochemistry, Faculty of Pharmacy,...

Life Sciences 157 (2016) 187–199
Contents lists available at ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / l i fesc ie
Cepharanthine and Piperine ameliorate diabetic nephropathy in rats: roleof NF-κB and NLRP3 inflammasome
Yara A. Samra a,⁎, Heba S. Said b, Nehal M. Elsherbiny a, Gregory I. Liou c,Mamdouh M. El-Shishtawy a, Laila A. Eissa a,⁎a Department of Biochemistry, Faculty of Pharmacy, Mansoura University, Mansoura 35516, Egyptb Department of Microbiology, Faculty of Pharmacy, Mansoura University, Mansoura 35516, Egyptc Department of Ophthalmology, Augusta University, Augusta, GA 30912, USA
⁎ Corresponding author.E-mail addresses: [email protected] (Y.A. Sam
(L.A. Eissa).
http://dx.doi.org/10.1016/j.lfs.2016.06.0020024-3205/© 2016 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 1 February 2016Received in revised form 31 May 2016Accepted 2 June 2016Available online 3 June 2016
Aims: Hyperglycemia leads to elevation of oxidative stress and proinflammatory cytokines which are the maincauses of diabetic nephropathy (DN). NLRP3 inflammasome and thioredoxin-interacting protein (TXNIP) are re-cently assumed to participate in the development of DN. We aimed to investigate the effects of Cepharanthine(CEP), Piperine (Pip) and their combination in streptozotocin (STZ)-induced DN focusing on their role to modu-late NLRP3 and TXNIP induced inflammation.Mainmethods:Diabetic ratswere treatedwith intraperitoneal (i.p.) injection of CEP (10mg/kg/day), Pip (30mg/kg/day) or their combination for 8weeks. Nuclear factor kappa B (NF-κB), tumor necrosis factor alpha (TNF-α) and in-terleukin-1β (IL-1β) were assessed by ELISA technique. TXNIP andNLRP3 genes expressionswere evaluated by realtime-PCR.Key findings: Diabetic rats showed significant increase in renal TXNIP and NLRP3 expression. CEP, Pip or their com-bination significantly decreased TXNIP andNLRP3 expression in diabetic kidneys. Hyperglycemia inducedNF-κB ac-tivation leading to increased IL-1β and TNF-α levels. CEP, Pip or their combination showed significant inhibition ofNF-κB together with decreased IL-1β and TNF-α levels in diabetic rats. Also, diabetic rats showed significant de-crease in creatinine clearance and increase in blood glucose, serum creatinine, blood urea nitrogen,malondialdehyde, proteinuria, and kidney weight to body Weight ratio. All of these changes were reversed byCEP, Pip or their combination.Significance: The antioxidant and anti-inflammatory effects of CEP and Pipwhichwere accompanied by inhibition ofNF-κB and NLRP3 activation might be helpful mechanisms to halt the progression of DN.
© 2016 Elsevier Inc. All rights reserved.
Keywords:CepharanthinePiperineDiabetesRenal injuryNF-κBTXNIPNLRP3
1. Introduction
Diabetes mellitus is a chronic inflammatory disease in which hyper-glycemia enhances glucose oxidation and production of advancedglycation end products (AGEs). Hyperglycemia activates protein kinaseC, polyol and hexosamine pathways and upregulates the production ofdifferent proinflammatory cytokines such as tumor necrosis factoralpha (TNF-α), interleukin 6 (IL-6) and interleukin-1β (IL-1β) [1]. Thepathophysiological changes in diabetes also activate nuclear factorkappa B (NF-κB) which perturbs more inflammatory cytokines produc-tion suggesting that NF-κB has a key role in the progression of diabetes-induced inflammation [2,3].
ra), [email protected]
Diabetic nephropathy (DN) is themain complication of diabetes thatis associated with increased cardiovascular mortality. DN is clinicallycharacterized by the hypertension and albuminuria which further pro-gresses to decline in renal function and leads to end-stage renal failure[4]. Current treatment strategy of DN involves glycemic and blood pres-sure control; however, patients continue to suffer from advanced renaldamage [5]. Therefore, new therapeutic modalities should be intro-duced to prevent the progression of DN.
NLRP3 inflammasome is a typical member of inflammasome largefamily that is present in mammalian cells directing renal inflammatoryprocesses leading to podocyte injury and glomerulosclerosis [6]. NALP3inflammasome is composed of NLRP3 (NACHT, LRR and PYD domains-containing protein 3), ASC (apoptosis-associated speck like protein)and caspase-1 [7,8]. Inflammasome is significantly involved in the path-ogenesis of DN through the production of the proinflammatory cyto-kines IL-1β and IL-18 as several NLRs play an important role in thecleavage of pro-IL-1β and pro-IL-18 into mature forms of IL-1β and IL-
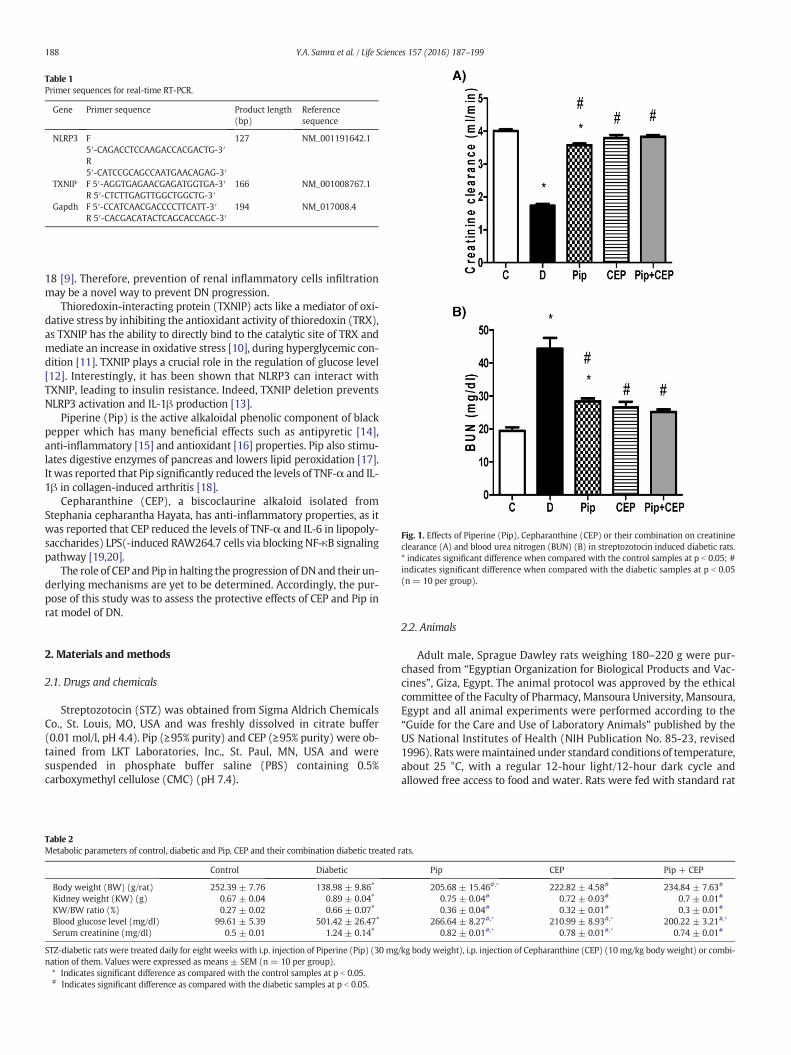
Table 1Primer sequences for real-time RT-PCR.
Gene Primer sequence Product length(bp)
Referencesequence
NLRP3 F5′-CAGACCTCCAAGACCACGACTG-3′
127 NM_001191642.1
R5′-CATCCGCAGCCAATGAACAGAG-3′
TXNIP F 5′-AGGTGAGAACGAGATGGTGA-3′ 166 NM_001008767.1R 5′-CTCTTGAGTTGGCTGGCTG-3′
Gapdh F 5′-CCATCAACGACCCCTTCATT-3′ 194 NM_017008.4R 5′-CACGACATACTCAGCACCAGC-3′
Fig. 1. Effects of Piperine (Pip), Cepharanthine (CEP) or their combination on creatinineclearance (A) and blood urea nitrogen (BUN) (B) in streptozotocin induced diabetic rats.* indicates significant difference when compared with the control samples at p b 0.05; #indicates significant difference when compared with the diabetic samples at p b 0.05(n = 10 per group).
188 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
18 [9]. Therefore, prevention of renal inflammatory cells infiltrationmay be a novel way to prevent DN progression.
Thioredoxin-interacting protein (TXNIP) acts like a mediator of oxi-dative stress by inhibiting the antioxidant activity of thioredoxin (TRX),as TXNIP has the ability to directly bind to the catalytic site of TRX andmediate an increase in oxidative stress [10], during hyperglycemic con-dition [11]. TXNIP plays a crucial role in the regulation of glucose level[12]. Interestingly, it has been shown that NLRP3 can interact withTXNIP, leading to insulin resistance. Indeed, TXNIP deletion preventsNLRP3 activation and IL-1β production [13].
Piperine (Pip) is the active alkaloidal phenolic component of blackpepper which has many beneficial effects such as antipyretic [14],anti-inflammatory [15] and antioxidant [16] properties. Pip also stimu-lates digestive enzymes of pancreas and lowers lipid peroxidation [17].It was reported that Pip significantly reduced the levels of TNF-α and IL-1β in collagen-induced arthritis [18].
Cepharanthine (CEP), a biscoclaurine alkaloid isolated fromStephania cepharantha Hayata, has anti-inflammatory properties, as itwas reported that CEP reduced the levels of TNF-α and IL-6 in lipopoly-saccharides) LPS(-induced RAW264.7 cells via blocking NF-κB signalingpathway [19,20].
The role of CEP and Pip in halting the progression of DNand their un-derlying mechanisms are yet to be determined. Accordingly, the pur-pose of this study was to assess the protective effects of CEP and Pip inrat model of DN.
2. Materials and methods
2.1. Drugs and chemicals
Streptozotocin (STZ) was obtained from Sigma Aldrich ChemicalsCo., St. Louis, MO, USA and was freshly dissolved in citrate buffer(0.01 mol/l, pH 4.4). Pip (≥95% purity) and CEP (≥95% purity) were ob-tained from LKT Laboratories, Inc., St. Paul, MN, USA and weresuspended in phosphate buffer saline (PBS) containing 0.5%carboxymethyl cellulose (CMC) (pH 7.4).
Table 2Metabolic parameters of control, diabetic and Pip, CEP and their combination diabetic treated
Control Diabetic
Body weight (BW) (g/rat) 252.39 ± 7.76 138.98 ± 9.86⁎
Kidney weight (KW) (g) 0.67 ± 0.04 0.89 ± 0.04⁎
KW/BW ratio (%) 0.27 ± 0.02 0.66 ± 0.07⁎
Blood glucose level (mg/dl) 99.61 ± 5.39 501.42 ± 26.47⁎
Serum creatinine (mg/dl) 0.5 ± 0.01 1.24 ± 0.14⁎
STZ-diabetic rats were treated daily for eight weeks with i.p. injection of Piperine (Pip) (30 mgnation of them. Values were expressed as means ± SEM (n= 10 per group).⁎ Indicates significant difference as compared with the control samples at p b 0.05.# Indicates significant difference as compared with the diabetic samples at p b 0.05.
2.2. Animals
Adult male, Sprague Dawley rats weighing 180–220 g were pur-chased from “Egyptian Organization for Biological Products and Vac-cines”, Giza, Egypt. The animal protocol was approved by the ethicalcommittee of the Faculty of Pharmacy, Mansoura University, Mansoura,Egypt and all animal experiments were performed according to the“Guide for the Care and Use of Laboratory Animals” published by theUS National Institutes of Health (NIH Publication No. 85-23, revised1996). Ratsweremaintained under standard conditions of temperature,about 25 °C, with a regular 12-hour light/12-hour dark cycle andallowed free access to food and water. Rats were fed with standard rat
rats.
Pip CEP Pip + CEP
205.68 ± 15.46#,⁎ 222.82 ± 4.58# 234.84 ± 7.63#
0.75 ± 0.04# 0.72 ± 0.03# 0.7 ± 0.01#
0.36 ± 0.04# 0.32 ± 0.01# 0.3 ± 0.01#
266.64 ± 8.27#,⁎ 210.99 ± 8.93#,⁎ 200.22 ± 3.21#,⁎
0.82 ± 0.01#,⁎ 0.78 ± 0.01#,⁎ 0.74 ± 0.01#
/kg bodyweight), i.p. injection of Cepharanthine (CEP) (10mg/kg body weight) or combi-
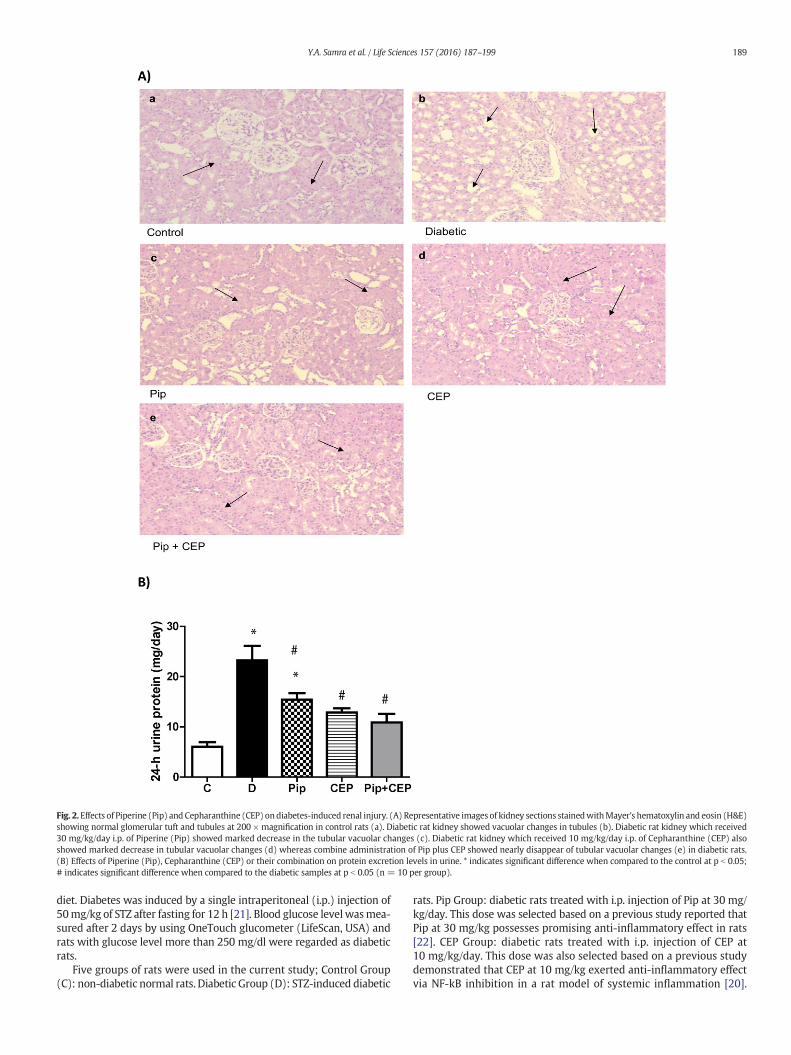
Fig. 2.Effects of Piperine (Pip) and Cepharanthine (CEP) ondiabetes-induced renal injury. (A) Representative images of kidney sections stainedwithMayer's hematoxylin and eosin (H&E)showing normal glomerular tuft and tubules at 200 × magnification in control rats (a). Diabetic rat kidney showed vacuolar changes in tubules (b). Diabetic rat kidney which received30 mg/kg/day i.p. of Piperine (Pip) showed marked decrease in the tubular vacuolar changes (c). Diabetic rat kidney which received 10 mg/kg/day i.p. of Cepharanthine (CEP) alsoshowed marked decrease in tubular vacuolar changes (d) whereas combine administration of Pip plus CEP showed nearly disappear of tubular vacuolar changes (e) in diabetic rats.(B) Effects of Piperine (Pip), Cepharanthine (CEP) or their combination on protein excretion levels in urine. * indicates significant difference when compared to the control at p b 0.05;# indicates significant difference when compared to the diabetic samples at p b 0.05 (n = 10 per group).
189Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
diet. Diabetes was induced by a single intraperitoneal (i.p.) injection of50mg/kg of STZ after fasting for 12 h [21]. Blood glucose level wasmea-sured after 2 days by using OneTouch glucometer (LifeScan, USA) andrats with glucose level more than 250 mg/dl were regarded as diabeticrats.
Five groups of rats were used in the current study; Control Group(C): non-diabetic normal rats. Diabetic Group (D): STZ-induced diabetic
rats. Pip Group: diabetic rats treated with i.p. injection of Pip at 30 mg/kg/day. This dose was selected based on a previous study reported thatPip at 30 mg/kg possesses promising anti-inflammatory effect in rats[22]. CEP Group: diabetic rats treated with i.p. injection of CEP at10 mg/kg/day. This dose was also selected based on a previous studydemonstrated that CEP at 10 mg/kg exerted anti-inflammatory effectvia NF-kB inhibition in a rat model of systemic inflammation [20].
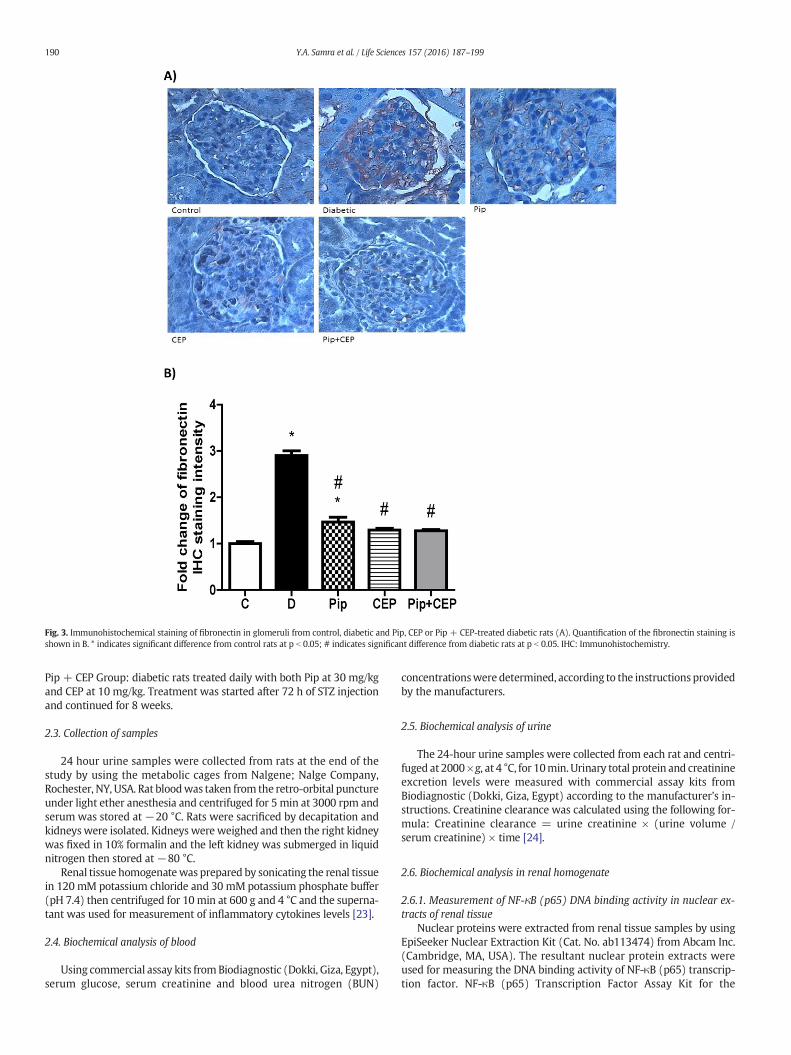
Fig. 3. Immunohistochemical staining of fibronectin in glomeruli from control, diabetic and Pip, CEP or Pip + CEP-treated diabetic rats (A). Quantification of the fibronectin staining isshown in B. * indicates significant difference from control rats at p b 0.05; # indicates significant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
190 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
Pip + CEP Group: diabetic rats treated daily with both Pip at 30 mg/kgand CEP at 10 mg/kg. Treatment was started after 72 h of STZ injectionand continued for 8 weeks.
2.3. Collection of samples
24 hour urine samples were collected from rats at the end of thestudy by using the metabolic cages from Nalgene; Nalge Company,Rochester, NY, USA. Rat bloodwas taken from the retro-orbital punctureunder light ether anesthesia and centrifuged for 5 min at 3000 rpm andserum was stored at −20 °C. Rats were sacrificed by decapitation andkidneyswere isolated. Kidneys were weighed and then the right kidneywas fixed in 10% formalin and the left kidney was submerged in liquidnitrogen then stored at−80 °C.
Renal tissue homogenatewas prepared by sonicating the renal tissuein 120 mM potassium chloride and 30 mM potassium phosphate buffer(pH 7.4) then centrifuged for 10min at 600 g and 4 °C and the superna-tant was used for measurement of inflammatory cytokines levels [23].
2.4. Biochemical analysis of blood
Using commercial assay kits from Biodiagnostic (Dokki, Giza, Egypt),serum glucose, serum creatinine and blood urea nitrogen (BUN)
concentrationswere determined, according to the instructionsprovidedby the manufacturers.
2.5. Biochemical analysis of urine
The 24-hour urine samples were collected from each rat and centri-fuged at 2000×g, at 4 °C, for 10min. Urinary total protein and creatinineexcretion levels were measured with commercial assay kits fromBiodiagnostic (Dokki, Giza, Egypt) according to the manufacturer's in-structions. Creatinine clearance was calculated using the following for-mula: Creatinine clearance = urine creatinine × (urine volume /serum creatinine) × time [24].
2.6. Biochemical analysis in renal homogenate
2.6.1. Measurement of NF-κB (p65) DNA binding activity in nuclear ex-tracts of renal tissue
Nuclear proteins were extracted from renal tissue samples by usingEpiSeeker Nuclear Extraction Kit (Cat. No. ab113474) from Abcam Inc.(Cambridge, MA, USA). The resultant nuclear protein extracts wereused for measuring the DNA binding activity of NF-κB (p65) transcrip-tion factor. NF-κB (p65) Transcription Factor Assay Kit for the
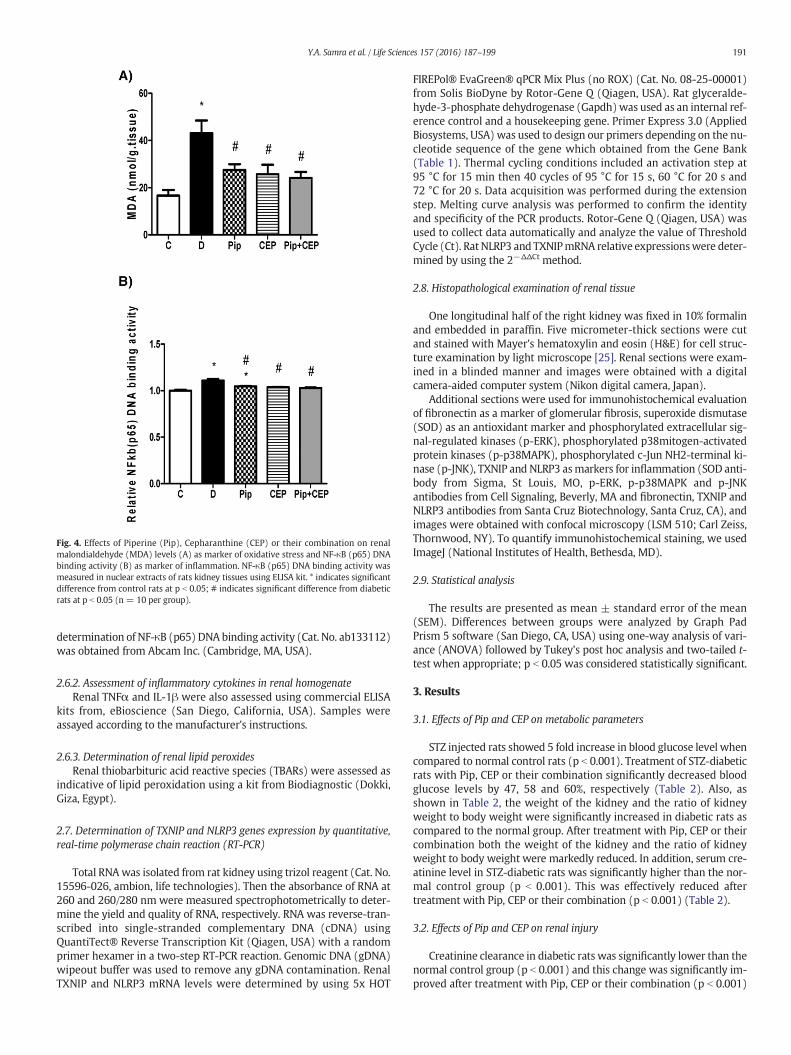
Fig. 4. Effects of Piperine (Pip), Cepharanthine (CEP) or their combination on renalmalondialdehyde (MDA) levels (A) as marker of oxidative stress and NF-κB (p65) DNAbinding activity (B) as marker of inflammation. NF-κB (p65) DNA binding activity wasmeasured in nuclear extracts of rats kidney tissues using ELISA kit. * indicates significantdifference from control rats at p b 0.05; # indicates significant difference from diabeticrats at p b 0.05 (n = 10 per group).
191Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
determination of NF-κB (p65) DNA binding activity (Cat. No. ab133112)was obtained from Abcam Inc. (Cambridge, MA, USA).
2.6.2. Assessment of inflammatory cytokines in renal homogenateRenal TNFα and IL-1β were also assessed using commercial ELISA
kits from, eBioscience (San Diego, California, USA). Samples wereassayed according to the manufacturer's instructions.
2.6.3. Determination of renal lipid peroxidesRenal thiobarbituric acid reactive species (TBARs) were assessed as
indicative of lipid peroxidation using a kit from Biodiagnostic (Dokki,Giza, Egypt).
2.7. Determination of TXNIP and NLRP3 genes expression by quantitative,real-time polymerase chain reaction (RT-PCR)
Total RNAwas isolated from rat kidney using trizol reagent (Cat. No.15596-026, ambion, life technologies). Then the absorbance of RNA at260 and 260/280 nm were measured spectrophotometrically to deter-mine the yield and quality of RNA, respectively. RNA was reverse-tran-scribed into single-stranded complementary DNA (cDNA) usingQuantiTect® Reverse Transcription Kit (Qiagen, USA) with a randomprimer hexamer in a two-step RT-PCR reaction. Genomic DNA (gDNA)wipeout buffer was used to remove any gDNA contamination. RenalTXNIP and NLRP3 mRNA levels were determined by using 5x HOT
FIREPol® EvaGreen® qPCR Mix Plus (no ROX) (Cat. No. 08-25-00001)from Solis BioDyne by Rotor-Gene Q (Qiagen, USA). Rat glyceralde-hyde-3-phosphate dehydrogenase (Gapdh)was used as an internal ref-erence control and a housekeeping gene. Primer Express 3.0 (AppliedBiosystems, USA) was used to design our primers depending on the nu-cleotide sequence of the gene which obtained from the Gene Bank(Table 1). Thermal cycling conditions included an activation step at95 °C for 15 min then 40 cycles of 95 °C for 15 s, 60 °C for 20 s and72 °C for 20 s. Data acquisition was performed during the extensionstep. Melting curve analysis was performed to confirm the identityand specificity of the PCR products. Rotor-Gene Q (Qiagen, USA) wasused to collect data automatically and analyze the value of ThresholdCycle (Ct). RatNLRP3 and TXNIPmRNA relative expressionswere deter-mined by using the 2−ΔΔCt method.
2.8. Histopathological examination of renal tissue
One longitudinal half of the right kidney was fixed in 10% formalinand embedded in paraffin. Five micrometer-thick sections were cutand stained with Mayer's hematoxylin and eosin (H&E) for cell struc-ture examination by light microscope [25]. Renal sections were exam-ined in a blinded manner and images were obtained with a digitalcamera-aided computer system (Nikon digital camera, Japan).
Additional sections were used for immunohistochemical evaluationof fibronectin as a marker of glomerular fibrosis, superoxide dismutase(SOD) as an antioxidant marker and phosphorylated extracellular sig-nal-regulated kinases (p-ERK), phosphorylated p38mitogen-activatedprotein kinases (p-p38MAPK), phosphorylated c-Jun NH2-terminal ki-nase (p-JNK), TXNIP and NLRP3 asmarkers for inflammation (SOD anti-body from Sigma, St Louis, MO, p-ERK, p-p38MAPK and p-JNKantibodies from Cell Signaling, Beverly, MA and fibronectin, TXNIP andNLRP3 antibodies from Santa Cruz Biotechnology, Santa Cruz, CA), andimages were obtained with confocal microscopy (LSM 510; Carl Zeiss,Thornwood, NY). To quantify immunohistochemical staining, we usedImageJ (National Institutes of Health, Bethesda, MD).
2.9. Statistical analysis
The results are presented as mean ± standard error of the mean(SEM). Differences between groups were analyzed by Graph PadPrism 5 software (San Diego, CA, USA) using one-way analysis of vari-ance (ANOVA) followed by Tukey's post hoc analysis and two-tailed t-test when appropriate; p b 0.05 was considered statistically significant.
3. Results
3.1. Effects of Pip and CEP on metabolic parameters
STZ injected rats showed 5 fold increase in blood glucose level whencompared to normal control rats (p b 0.001). Treatment of STZ-diabeticrats with Pip, CEP or their combination significantly decreased bloodglucose levels by 47, 58 and 60%, respectively (Table 2). Also, asshown in Table 2, the weight of the kidney and the ratio of kidneyweight to body weight were significantly increased in diabetic rats ascompared to the normal group. After treatment with Pip, CEP or theircombination both the weight of the kidney and the ratio of kidneyweight to body weight were markedly reduced. In addition, serum cre-atinine level in STZ-diabetic rats was significantly higher than the nor-mal control group (p b 0.001). This was effectively reduced aftertreatment with Pip, CEP or their combination (p b 0.001) (Table 2).
3.2. Effects of Pip and CEP on renal injury
Creatinine clearance in diabetic rats was significantly lower than thenormal control group (p b 0.001) and this change was significantly im-proved after treatment with Pip, CEP or their combination (p b 0.001)
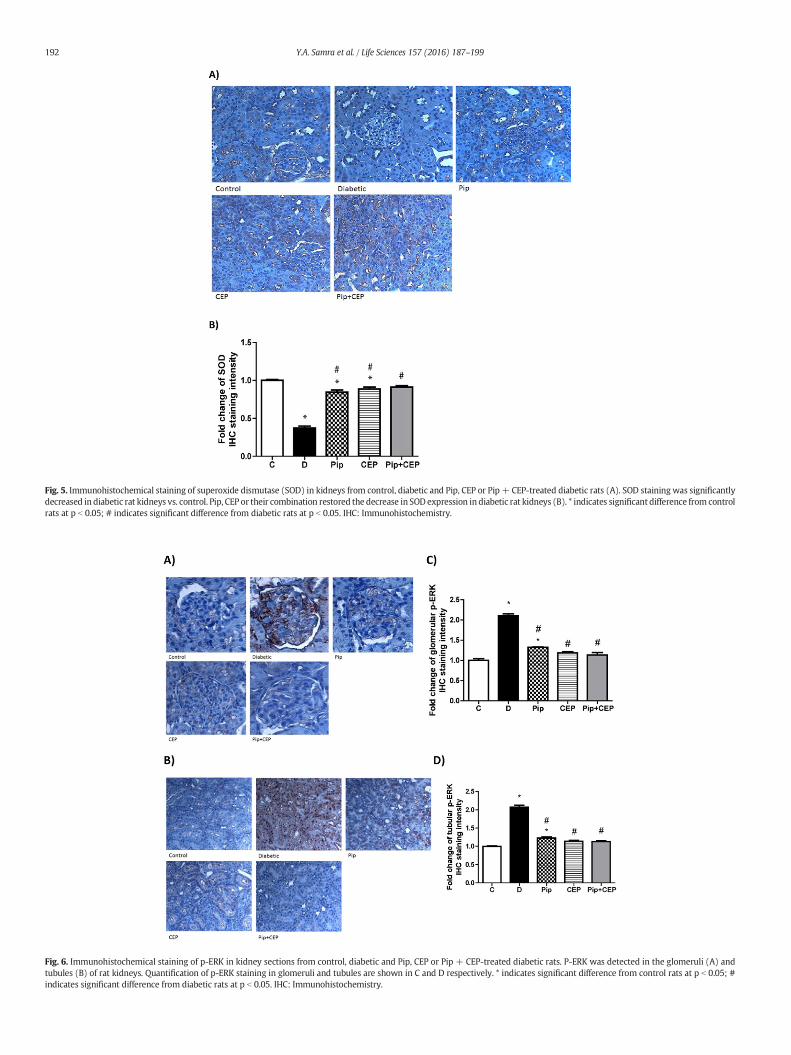
Fig. 5. Immunohistochemical staining of superoxide dismutase (SOD) in kidneys from control, diabetic and Pip, CEP or Pip + CEP-treated diabetic rats (A). SOD staining was significantlydecreased in diabetic rat kidneys vs. control. Pip, CEP or their combination restored the decrease in SOD expression in diabetic rat kidneys (B). * indicates significant difference from controlrats at p b 0.05; # indicates significant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
Fig. 6. Immunohistochemical staining of p-ERK in kidney sections from control, diabetic and Pip, CEP or Pip + CEP-treated diabetic rats. P-ERK was detected in the glomeruli (A) andtubules (B) of rat kidneys. Quantification of p-ERK staining in glomeruli and tubules are shown in C and D respectively. * indicates significant difference from control rats at p b 0.05; #indicates significant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
192 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
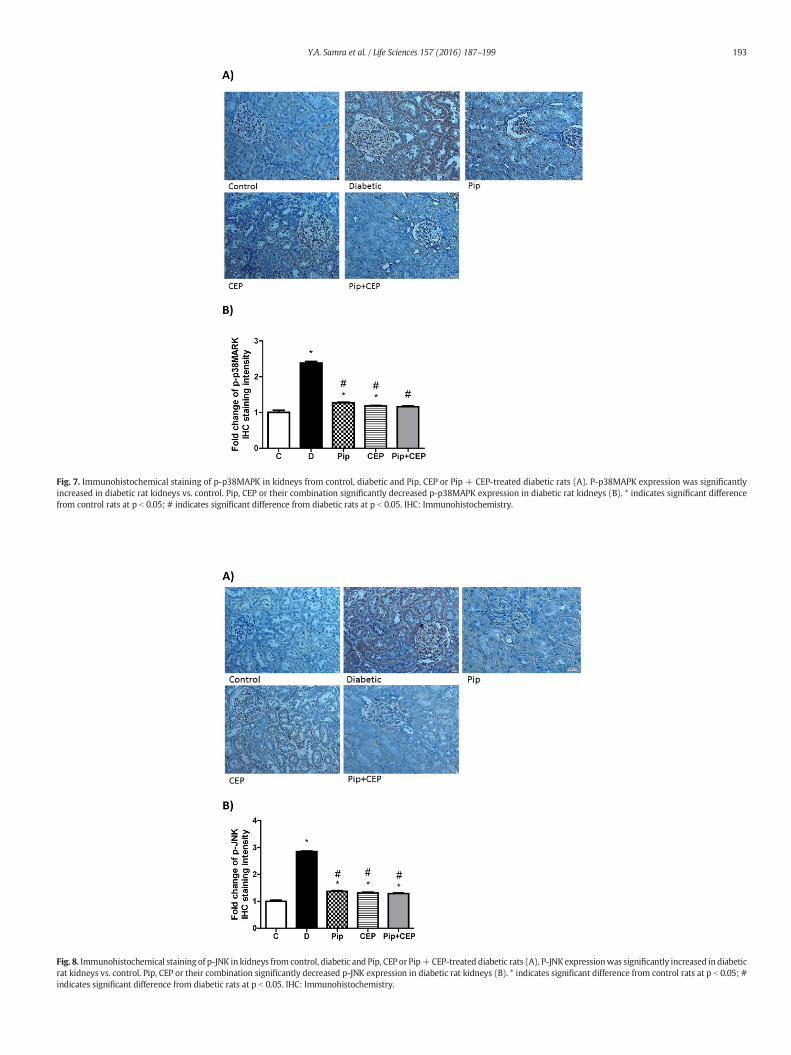
Fig. 7. Immunohistochemical staining of p-p38MAPK in kidneys from control, diabetic and Pip, CEP or Pip + CEP-treated diabetic rats (A). P-p38MAPK expression was significantlyincreased in diabetic rat kidneys vs. control. Pip, CEP or their combination significantly decreased p-p38MAPK expression in diabetic rat kidneys (B). * indicates significant differencefrom control rats at p b 0.05; # indicates significant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
Fig. 8. Immunohistochemical stainingof p-JNK in kidneys from control, diabetic and Pip, CEP or Pip+CEP-treated diabetic rats (A). P-JNK expressionwas significantly increased in diabeticrat kidneys vs. control. Pip, CEP or their combination significantly decreased p-JNK expression in diabetic rat kidneys (B). * indicates significant difference from control rats at p b 0.05; #indicates significant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
193Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
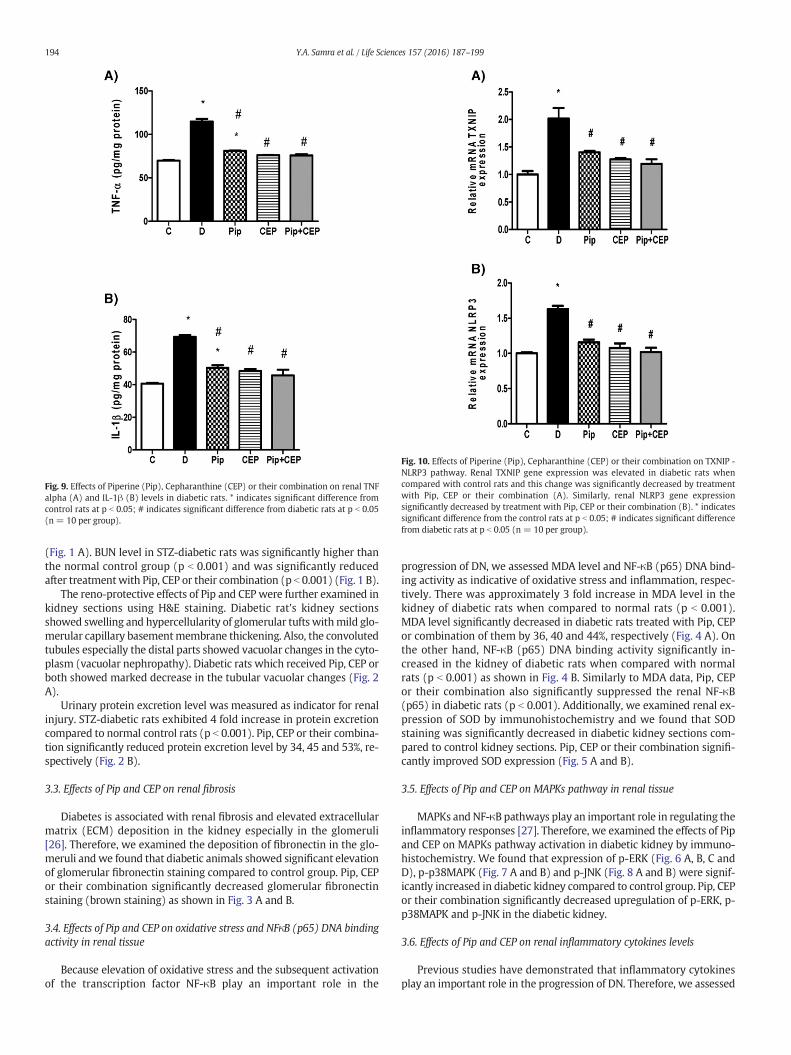
Fig. 9. Effects of Piperine (Pip), Cepharanthine (CEP) or their combination on renal TNFalpha (A) and IL-1β (B) levels in diabetic rats. * indicates significant difference fromcontrol rats at p b 0.05; # indicates significant difference from diabetic rats at p b 0.05(n = 10 per group).
Fig. 10. Effects of Piperine (Pip), Cepharanthine (CEP) or their combination on TXNIP -NLRP3 pathway. Renal TXNIP gene expression was elevated in diabetic rats whencompared with control rats and this change was significantly decreased by treatmentwith Pip, CEP or their combination (A). Similarly, renal NLRP3 gene expressionsignificantly decreased by treatment with Pip, CEP or their combination (B). * indicatessignificant difference from the control rats at p b 0.05; # indicates significant differencefrom diabetic rats at p b 0.05 (n = 10 per group).
194 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
(Fig. 1 A). BUN level in STZ-diabetic rats was significantly higher thanthe normal control group (p b 0.001) and was significantly reducedafter treatmentwith Pip, CEP or their combination (p b 0.001) (Fig. 1 B).
The reno-protective effects of Pip and CEP were further examined inkidney sections using H&E staining. Diabetic rat's kidney sectionsshowed swelling and hypercellularity of glomerular tufts withmild glo-merular capillary basementmembrane thickening. Also, the convolutedtubules especially the distal parts showed vacuolar changes in the cyto-plasm (vacuolar nephropathy). Diabetic rats which received Pip, CEP orboth showed marked decrease in the tubular vacuolar changes (Fig. 2A).
Urinary protein excretion level was measured as indicator for renalinjury. STZ-diabetic rats exhibited 4 fold increase in protein excretioncompared to normal control rats (p b 0.001). Pip, CEP or their combina-tion significantly reduced protein excretion level by 34, 45 and 53%, re-spectively (Fig. 2 B).
3.3. Effects of Pip and CEP on renal fibrosis
Diabetes is associated with renal fibrosis and elevated extracellularmatrix (ECM) deposition in the kidney especially in the glomeruli[26]. Therefore, we examined the deposition of fibronectin in the glo-meruli andwe found that diabetic animals showed significant elevationof glomerular fibronectin staining compared to control group. Pip, CEPor their combination significantly decreased glomerular fibronectinstaining (brown staining) as shown in Fig. 3 A and B.
3.4. Effects of Pip and CEP on oxidative stress and NFκB (p65) DNA bindingactivity in renal tissue
Because elevation of oxidative stress and the subsequent activationof the transcription factor NF-κB play an important role in the
progression of DN, we assessed MDA level and NF-κB (p65) DNA bind-ing activity as indicative of oxidative stress and inflammation, respec-tively. There was approximately 3 fold increase in MDA level in thekidney of diabetic rats when compared to normal rats (p b 0.001).MDA level significantly decreased in diabetic rats treated with Pip, CEPor combination of them by 36, 40 and 44%, respectively (Fig. 4 A). Onthe other hand, NF-κB (p65) DNA binding activity significantly in-creased in the kidney of diabetic rats when compared with normalrats (p b 0.001) as shown in Fig. 4 B. Similarly to MDA data, Pip, CEPor their combination also significantly suppressed the renal NF-κB(p65) in diabetic rats (p b 0.001). Additionally, we examined renal ex-pression of SOD by immunohistochemistry and we found that SODstaining was significantly decreased in diabetic kidney sections com-pared to control kidney sections. Pip, CEP or their combination signifi-cantly improved SOD expression (Fig. 5 A and B).
3.5. Effects of Pip and CEP on MAPKs pathway in renal tissue
MAPKs and NF-κB pathways play an important role in regulating theinflammatory responses [27]. Therefore, we examined the effects of Pipand CEP on MAPKs pathway activation in diabetic kidney by immuno-histochemistry. We found that expression of p-ERK (Fig. 6 A, B, C andD), p-p38MAPK (Fig. 7 A and B) and p-JNK (Fig. 8 A and B) were signif-icantly increased in diabetic kidney compared to control group. Pip, CEPor their combination significantly decreased upregulation of p-ERK, p-p38MAPK and p-JNK in the diabetic kidney.
3.6. Effects of Pip and CEP on renal inflammatory cytokines levels
Previous studies have demonstrated that inflammatory cytokinesplay an important role in the progression of DN. Therefore, we assessed
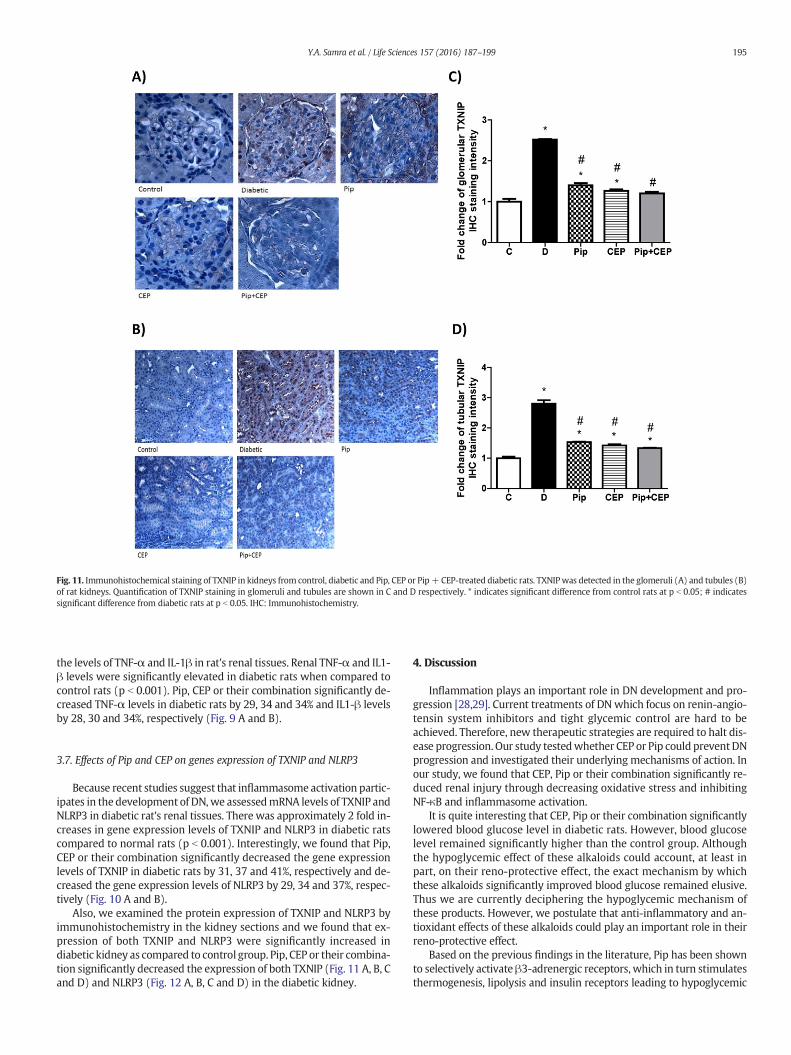
Fig. 11. Immunohistochemical staining of TXNIP in kidneys from control, diabetic and Pip, CEP or Pip+ CEP-treated diabetic rats. TXNIPwas detected in the glomeruli (A) and tubules (B)of rat kidneys. Quantification of TXNIP staining in glomeruli and tubules are shown in C and D respectively. * indicates significant difference from control rats at p b 0.05; # indicatessignificant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
195Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
the levels of TNF-α and IL-1β in rat's renal tissues. Renal TNF-α and IL1-β levels were significantly elevated in diabetic rats when compared tocontrol rats (p b 0.001). Pip, CEP or their combination significantly de-creased TNF-α levels in diabetic rats by 29, 34 and 34% and IL1-β levelsby 28, 30 and 34%, respectively (Fig. 9 A and B).
3.7. Effects of Pip and CEP on genes expression of TXNIP and NLRP3
Because recent studies suggest that inflammasome activation partic-ipates in the development of DN,we assessedmRNA levels of TXNIP andNLRP3 in diabetic rat's renal tissues. There was approximately 2 fold in-creases in gene expression levels of TXNIP and NLRP3 in diabetic ratscompared to normal rats (p b 0.001). Interestingly, we found that Pip,CEP or their combination significantly decreased the gene expressionlevels of TXNIP in diabetic rats by 31, 37 and 41%, respectively and de-creased the gene expression levels of NLRP3 by 29, 34 and 37%, respec-tively (Fig. 10 A and B).
Also, we examined the protein expression of TXNIP and NLRP3 byimmunohistochemistry in the kidney sections and we found that ex-pression of both TXNIP and NLRP3 were significantly increased indiabetic kidney as compared to control group. Pip, CEP or their combina-tion significantly decreased the expression of both TXNIP (Fig. 11 A, B, Cand D) and NLRP3 (Fig. 12 A, B, C and D) in the diabetic kidney.
4. Discussion
Inflammation plays an important role in DN development and pro-gression [28,29]. Current treatments of DNwhich focus on renin-angio-tensin system inhibitors and tight glycemic control are hard to beachieved. Therefore, new therapeutic strategies are required to halt dis-ease progression. Our study testedwhether CEP or Pip could preventDNprogression and investigated their underlying mechanisms of action. Inour study, we found that CEP, Pip or their combination significantly re-duced renal injury through decreasing oxidative stress and inhibitingNF-κB and inflammasome activation.
It is quite interesting that CEP, Pip or their combination significantlylowered blood glucose level in diabetic rats. However, blood glucoselevel remained significantly higher than the control group. Althoughthe hypoglycemic effect of these alkaloids could account, at least inpart, on their reno-protective effect, the exact mechanism by whichthese alkaloids significantly improved blood glucose remained elusive.Thus we are currently deciphering the hypoglycemic mechanism ofthese products. However, we postulate that anti-inflammatory and an-tioxidant effects of these alkaloids could play an important role in theirreno-protective effect.
Based on the previous findings in the literature, Pip has been shownto selectively activate β3-adrenergic receptors, which in turn stimulatesthermogenesis, lipolysis and insulin receptors leading to hypoglycemic
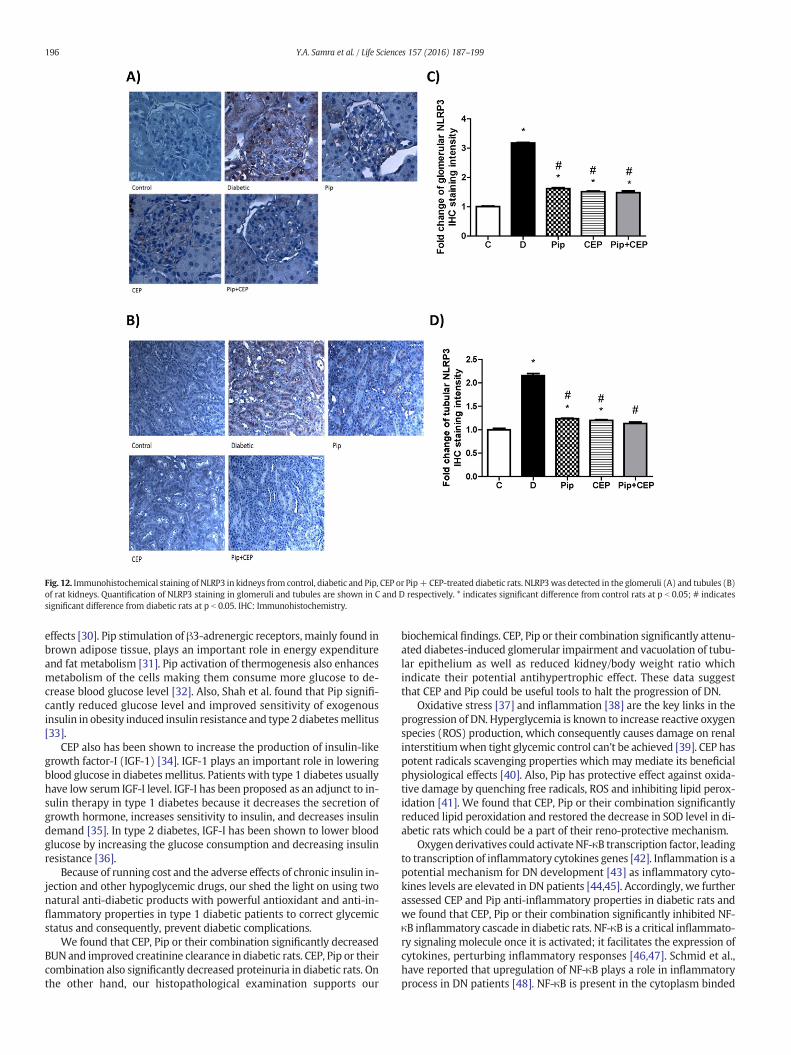
Fig. 12. Immunohistochemical staining of NLRP3 in kidneys from control, diabetic and Pip, CEP or Pip+CEP-treated diabetic rats. NLRP3was detected in the glomeruli (A) and tubules (B)of rat kidneys. Quantification of NLRP3 staining in glomeruli and tubules are shown in C and D respectively. * indicates significant difference from control rats at p b 0.05; # indicatessignificant difference from diabetic rats at p b 0.05. IHC: Immunohistochemistry.
196 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
effects [30]. Pip stimulation of β3-adrenergic receptors, mainly found inbrown adipose tissue, plays an important role in energy expenditureand fat metabolism [31]. Pip activation of thermogenesis also enhancesmetabolism of the cells making them consume more glucose to de-crease blood glucose level [32]. Also, Shah et al. found that Pip signifi-cantly reduced glucose level and improved sensitivity of exogenousinsulin in obesity induced insulin resistance and type 2 diabetesmellitus[33].
CEP also has been shown to increase the production of insulin-likegrowth factor-I (IGF-1) [34]. IGF-1 plays an important role in loweringblood glucose in diabetes mellitus. Patients with type 1 diabetes usuallyhave low serum IGF-I level. IGF-I has been proposed as an adjunct to in-sulin therapy in type 1 diabetes because it decreases the secretion ofgrowth hormone, increases sensitivity to insulin, and decreases insulindemand [35]. In type 2 diabetes, IGF-I has been shown to lower bloodglucose by increasing the glucose consumption and decreasing insulinresistance [36].
Because of running cost and the adverse effects of chronic insulin in-jection and other hypoglycemic drugs, our shed the light on using twonatural anti-diabetic products with powerful antioxidant and anti-in-flammatory properties in type 1 diabetic patients to correct glycemicstatus and consequently, prevent diabetic complications.
We found that CEP, Pip or their combination significantly decreasedBUN and improved creatinine clearance in diabetic rats. CEP, Pip or theircombination also significantly decreased proteinuria in diabetic rats. Onthe other hand, our histopathological examination supports our
biochemical findings. CEP, Pip or their combination significantly attenu-ated diabetes-induced glomerular impairment and vacuolation of tubu-lar epithelium as well as reduced kidney/body weight ratio whichindicate their potential antihypertrophic effect. These data suggestthat CEP and Pip could be useful tools to halt the progression of DN.
Oxidative stress [37] and inflammation [38] are the key links in theprogression of DN. Hyperglycemia is known to increase reactive oxygenspecies (ROS) production, which consequently causes damage on renalinterstitiumwhen tight glycemic control can't be achieved [39]. CEP haspotent radicals scavenging properties which may mediate its beneficialphysiological effects [40]. Also, Pip has protective effect against oxida-tive damage by quenching free radicals, ROS and inhibiting lipid perox-idation [41]. We found that CEP, Pip or their combination significantlyreduced lipid peroxidation and restored the decrease in SOD level in di-abetic rats which could be a part of their reno-protective mechanism.
Oxygenderivatives could activate NF-κB transcription factor, leadingto transcription of inflammatory cytokines genes [42]. Inflammation is apotential mechanism for DN development [43] as inflammatory cyto-kines levels are elevated in DN patients [44,45]. Accordingly, we furtherassessed CEP and Pip anti-inflammatory properties in diabetic rats andwe found that CEP, Pip or their combination significantly inhibited NF-κB inflammatory cascade in diabetic rats. NF-κB is a critical inflammato-ry signaling molecule once it is activated; it facilitates the expression ofcytokines, perturbing inflammatory responses [46,47]. Schmid et al.,have reported that upregulation of NF-κB plays a role in inflammatoryprocess in DN patients [48]. NF-κB is present in the cytoplasm binded

Fig. 13. The proposed reno-protective mechanism of Piperine and Cepharanthine through TXNIP – NLRP3 pathway.
197Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
to its inhibitor IkB. After activation by physiological and non-physiolog-ical stimuli, such as IL-1β, TNF-α, or LPS, IkB dissociates fromNF-κB andundergoes degradation. ThenNF-κB is translocated into the nucleus andphosphorylated on serine 276 in its p65 subunit. P-NF-κB subsequentlybinds with DNA and encourages proinflammatory cytokines transcrip-tion, such as TNF-α and IL-1β [49]. Therefore, measurement of theDNA binding activity of NF-κB p65 subunit is an effective tool for deter-mining NF-κB activation.
On the other hand, TNF-α is a multifunctional regulatory cytokineinvolved in the inflammatory response in DN [50]. TNF-α also inhibitsinsulin signaling pathways [51] and impairs peripheral glucose uptake[52]. Our results showed that NF-κB activation as well as TNF-α andIL-1β concentrations significantly increased in diabetic rats comparedwith normal rats. Interestingly,we found that CEP, Pip or their combina-tion significantly inhibited NF-κB activation and reduced TNF-α and IL-1β levels in diabetic rats. These results suggest that the beneficial effectsof CEP and Pip inDNare at least in part through inhibition of renal NF-κBmediated inflammatory response.
ERK, p38MAPK and JNK are signal transduction factors which trans-mit signals from environmental stimuli to the nucleus [53]. It has beenproved that ERK, p38MAPK and JNK are activated by high glucose levelsin DNmodels [54,55]. Moreover, p-ERK, p-p38MAPK and p-JNK partici-pate in NF-κB activation which regulates the gene expression of
proinflammatory cytokines [56,57]. Our results showed that CEP, Pipor their combination significantly inhibited MAPKs pathway activationin diabetic kidney which might be a part of their reno-protectivemechanism.
DN is associated with glomerular damage which has been linked toinflammation as during diabetes activated inflammatory cells enhancecytokines release which leads to fibrosis and matrix deposition [58,59]. Therefore, wemeasured fibronectin expression asmarker for fibro-sis and we found that CEP, Pip or their combination significantly de-creased fibronectin deposition in the glomeruli of diabetic ratssuggesting that decreased inflammation might confer protective mech-anism against renal fibrosis.
Inflammasome (NLRP3) is a novel interesting inflammatory path-way which leads to a rapid inflammatory response by elevating IL-1βand IL-18 levels [9]. After gathering of the NLRP3, IL-1β cleavages toits active form. Activation of IL-1β occurs within the inflammasomecomplex then mature IL-1β is released into the extracellular space byexocytosis or loss of membrane integrity [60]. Recent studies suggestthat inflammasome and IL-1β/IL-18 axis play an important role ingout, rheumatoid arthritis, atherosclerosis, diabetes mellitus, and oxa-late crystal nephropathy diseases [60]. The role of IL-1β and IL-18 inrenal disease is well known but the importance of the inflammasomein their activation has not been investigated. In our study, NLRP3 level

198 Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
in diabetic kidney was elevated with the progression of DN togetherwith increase in renal IL-1β level. We interestingly found that CEP, Pipor their combination significantly decreased NLRP3 and IL-1β levels indiabetic rats suggesting that decreased inflammasome activationmight inhibit DN progression.
TXNIP is known to negatively regulate TRX function in many tissues[61]. TRX is a thiol oxidoreductase which exerts antioxidant propertiesthrough scavenging of ROS and regulating the activity of redox-sensitivetranscription factors [62]. It was reported that high glucose could in-crease TXNIP which is subsequently involved in oxidative stress in dia-betic rats [63].
TXNIP may also play an important role in the pathogenesis of DN byenhancing hyperglycemia-induced IL-1β production [64]. TXNIP can di-rectly activate NF-κB and its downstream inflammatory cytokines [65].In addition, it was reported that TXNIP binds with NLRP3 and mediatesNLRP3 activation [66]. Therefore, wemeasured TXNIP level in DN to fur-ther explore themechanismof NLRP3 activation. In our study, we foundthat TXNIP level significantly increased in diabetic kidneys comparedwith normal kidneys. We also found that CEP, Pip or their combinationsignificantly decreased TXNIP level in diabetic rats. We postulate thatCEP and Pip anti-inflammatory effect could be mediated through de-creasing TXNIP level, which consequently decrease NLRP3 activationand production of inflammatory cytokines.
It is worthy to mention that the combination of CEP and Pip didn'taugment the renal protection. We postulate that this might be due tothe ability of CEP or Pip alone to normalize free radicals and inflamma-tory cytokines levels to almost the same levels in control rats and ac-cordingly their combination failed to add more reno-protective effects.
Fig. 13 indicates our proposedDNprotectivemechanisms of CEP andPip in STZ-inducedDN in rats. CEP and Pip protected the kidney fromdi-abetic inflammatory complications by inhibiting the activation of NF-κBwhich leads to decrease the levels of TNF-α and IL-1β inflammatory cy-tokines. Also, both CEP and Pip have the ability to decrease the expres-sion of TXNIP which consequently leads to inhibition of NLRP3activation and IL-1β production.
5. Conclusion
Despite the antihyperglycemic effects of CEP and Pip in diabetic rats.We believe that antioxidant and anti-inflammatory effects of CEP andPip might contribute to their potential reno-protective mechanisms indiabetic rats especially their ability to reduce NF-κB and inflammasomeactivation.
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgement
The authors are extremely grateful to Dr.Wageha Kandeel professorof pathology at Faculty of Medicine, Mansoura University, Mansoura,Egypt for her pathological work. Also, the authors are extremely gratefulto Dr. Ahmed El-Marakby assistant professor at Department of Oral Bi-ology, Pharmacology & Toxicology and Vascular Biology Center, DentalCollege of Georgia, Augusta University, Augusta, GA, USA for his writingassistance.
References
[1] D. Luis-Rodrı'guez, A. Martı'nez-Castelao, J.L. Go'rriz, F. De-A' lvaro, J.F. Navarro-Gonza'lez, Pathophysiological role and therapeutic implications of inflammation indiabetic nephropathy, World J. Diabetes 3 (2012) 7–18, http://dx.doi.org/10.4239/wjd.v3.i1.7.
[2] X. Zhou, B. Wang, L. Zhu, S. Hao, A novel improved therapy strategy for diabetic ne-phropathy: targeting AGEs, Organogenesis 8 (2012) 18–21, http://dx.doi.org/10.4161/org.19332.
[3] P. Palsamy, S. Subramanian, Resveratrol protects diabetic kidney by attenuating hy-perglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling, Biochim. Biophys. Acta 2011 (1812) 719–731, http://dx.doi.org/10.1016/j.bbadis.2011.03.008.
[4] K. Gariani, S. de Seigneux, A. Pechere-Bertschi, J. Philippe, P.Y. Martin, Diabetic ne-phropathy: an update, Rev. Med. Suisse 8 (2012) 473–479.
[5] M. Svensson, G. Sundkvist, H.J. Arnqvist, E. Björk, G. Blohmé, J. Bolinder, M.Henricsson, L. Nyström, O. Torffvit, I. Waernbaum, J. Ostman, J.W. Eriksson, Diabetesincidence study in Sweden (DISS). Signs of nephropathy may occur early in youngadults with diabetes despite modern diabetes management: results from the na-tionwide population-based diabetes incidence study in Sweden (DISS), DiabetesCare 26 (2003) 2903–2909.
[6] S. Mariathasan, K. Newton, D.M. Monack, D. Vucic, D.M. French, W.P. Lee, M. Roose-Girma, S. Erickson, V.M. Dixit, Differential activation of the inflammasome by cas-pase-1 adaptors ASC and Ipaf, Nature 430 (2004) 213–218.
[7] V. Pétrilli, C. Dostert, D.A. Muruve, J. Tschopp, The inflammasome: a danger sensingcomplex triggering innate immunity, Curr. Opin. Immunol. 19 (2007) 615–622.
[8] P. Gao, X.F. Meng, H. Su, F.F. He, S. Chen, H. Tang, X.J. Tian, D. Fan, Y.M. Wang, J.S. Liu,Z.H. Zhu, C. Zhang, Thioredoxin-interacting protein mediates NALP3 inflammasomeactivation in podocytes during diabetic nephropathy, Biochim. Biophys. Acta 2014(1843) 2448–2460, http://dx.doi.org/10.1016/j.bbamcr.2014.07.001.
[9] C.M. Turner, N. Arulkumaran, M. Singer, R.J. Unwin, F.W. Tam, Is the inflammasomea potential therapeutic target in renal disease? BMC Nephrol. 15 (2014) 21, http://dx.doi.org/10.1186/1471-2369-15-21.
[10] P. Patwari, L.J. Higgins, W.A. Chutkow, J. Yoshioka, R.T. Lee, The interaction ofthioredoxin with Txnip. Evidence for formation of a mixed disulfide by disulfide ex-change, J. Biol. Chem. 281 (2006) 21884–21891.
[11] P.C. Schulze, J. Yoshioka, T. Takahashi, Z. He, G.L. King, R.T. Lee, Hyperglycemia pro-motes oxidative stress through inhibition of thioredoxin function by thioredoxin-interacting protein, J. Biol. Chem. 279 (2004) 30369–30374.
[12] C.Y. Tan, Q. Weier, Y. Zhang, A.J. Cox, D.J. Kelly, R.G. Langham, Thioredoxin-interacting protein: a potential therapeutic target for treatment of progressive fibro-sis in diabetic nephropathy, Nephron 129 (2015) 109–127, http://dx.doi.org/10.1159/000368238.
[13] C.M. Oslowski, T. Hara, B. O'Sullivan-Murphy, K. Kanekura, S. Lu, M. Hara, S. Ishigaki,L.J. Zhu, E. Hayashi, S.T. Hui, D. Greiner, R.J. Kaufman, R. Bortell, F. Urano,Thioredoxin-interacting protein mediates ER stress-induced beta cell death throughinitiation of the inflammasome, Cell Metab. 16 (2012) 265–273, http://dx.doi.org/10.1016/j.cmet.2012.07.005.
[14] K. Srinivasan, Black pepper and its pungent principle-piperine: a review of diversephysiological effects, Crit. Rev. Food Sci. Nutr. 47 (2007) 735–748.
[15] E.B. Lee, K.H. Shin,W.S. Woo, Pharmacological study on piperine, Arch. Pharm. Res. 7(1984) 127–132, http://dx.doi.org/10.1007/BF02856625.
[16] N. Pathak, S. Khandelwal, Cytoprotective and immunomodulating properties of piper-ine on murine splenocytes: an in vitro study, Eur. J. Pharmacol. 576 (2007) 160–170.
[17] J.S. Bang, H. Oh da, H.M. Choi, B.J. Sur, S.J. Lim, J.Y. Kim, H.I. Yang, M.C. Yoo, D.H.Hahm, K.S. Kim, Anti-inflammatory and antiarthritic effects of piperine in human in-terleukin 1β-stimulated fibroblast-like synoviocytes and in rat arthritis models, Ar-thritis Res. Ther. 11 (2009) R49, http://dx.doi.org/10.1186/ar2662.
[18] S. Umar, A.H. Golam Sarwar, K. Umar, N. Ahmad, M. Sajad, S. Ahmad, C.K. Katiyar,H.A. Khan, Piperine ameliorates oxidative stress, inflammation and histological out-come in collagen induced arthritis, Cell. Immunol. 284 (2013) 51–59, http://dx.doi.org/10.1016/j.cellimm.2013.07.004.
[19] S.S. Bun, M. Laget, A. Chea, H. Bun, E. Ollivier, R. Elias, Cytotoxic activity of alkaloidsisolated from Stephania rotunda [corrected], Phytother. Res. 23 (2009) 587–590,http://dx.doi.org/10.1002/ptr.2617.
[20] K. Kudo, S. Hagiwara, A. Hasegawa, J. Kusaka, H. Koga, T. Noguchi, Cepharanthine ex-erts anti-inflammatory effects via NF-kappaB inhibition in a LPS-induced rat modelof systemic inflammation, J. Surg. Res. 171 (2011) 199–204, http://dx.doi.org/10.1016/j.jss.2010.01.007.
[21] N.M. Elsherbiny, K.H. Abd El Galil, M.M. Gabr, M.M. Al-Gayyar, L.A. Eissa, M.M. El-Shishtawy, Reno- protective effect of NECA in diabetic nephropathy: implicationof IL-18 and ICAM-1, Eur. Cytokine Netw. 23 (2012) 78–86, http://dx.doi.org/10.1684/ecn.2012.0309.
[22] V. Murunikkara, S.J. Pragasam, G. Kodandaraman, E.P. Sabina, M. Rasool, Anti-inflam-matory effect of piperine in adjuvant-induced arthritic rats–a biochemical approach, In-flammation 35 (2012) 1348–1356, http://dx.doi.org/10.1007/s10753-012-9448-3.
[23] E.M. de Cavanagh, F. Inserra, L. Ferder, C.G. Fraga, Enalapril and captopril enhanceglutathione-dependent antioxidant defenses in mouse tissues, Am. J. Phys. Regul.Integr. Comp. Phys. 278 (2000) R572–R577.
[24] F. Bagheri, A. Gol, S. Dabiri, A. Javadi, Preventive effect of garlic juice on renal reper-fusion injury, Iran. J. Kidney Dis. 5 (2011) 194–200.
[25] D.J. Kelly, A.J. Edgley, Y. Zhang, K. Thai, S.M. Tan, A.J. Cox, A. Advani, K.A. Connelly, C.I.Whiteside, R.E. Gilbert, Protein kinase C-beta inhibition attenuates the progressionof nephropathy in non-diabetic kidney disease, Nephrol. Dial. Transplant. 24(2009) 1782–1790, http://dx.doi.org/10.1093/ndt/gfn729.
[26] H.B. Lee, M.R. Yu, Y. Yang, Z. Jiang, H. Ha, Reactive oxygen species-regulated signal-ing pathways in diabetic nephropathy, J. Am. Soc. Nephrol. 14 (2003) S241–S245.
[27] Y. Liu, E.G. Shepherd, L.D. Nelin, MAPK phosphatases–regulating the immune re-sponse, Nat. Rev. Immunol. 7 (2007) 202–212.
[28] J.F. Navarro, C. Mora, Role of inflammation in diabetic complications, Nephrol. Dial.Transplant. 20 (2005) 2601–2604, http://dx.doi.org/10.1093/ndt/gfi155.
[29] P.P. Wolkow, M.A. Niewczas, B. Perkins, L.H. Ficociello, B. Lipinski, J.H. Warram, A.S.Krolewski, Association of urinary inflammatory markers and renal decline inmicroalbuminuric type 1 diabetics, J. Am. Soc. Nephrol. 19 (2008) 789–797, http://dx.doi.org/10.1681/ASN.2007050556.

199Y.A. Samra et al. / Life Sciences 157 (2016) 187–199
[30] H.P. Rang, M.M. Dale, H.M. Ritter, P. Gardner, Pharmacology, fifth ed. Churchill Liv-ingstone, Edinburgh, UK, 2003 163.
[31] J. Klein, M. Fasshauer, M. Benito, C.R. Kahn, Insulin and the beta3-adrenoceptor dif-ferentially regulate uncoupling protein-1 expression, Mol. Endocrinol. 14 (2000)764–773.
[32] H.L. Sharma, K.K. Sharma, Principles of Pharmacology, first ed. Paras Medical Pub-lishers, New Delhi, 2008 653.
[33] S. Shah, G. Shah, M.B. Patel, S. Singh, Effect of piperine in obesity induced insu-lin resistance and type_II diabetes mellitus in rats, J. Nat. Remedies 10 (2010)116–122.
[34] S. Inui, S. Itami, Induction of insulin-like growth factor-I by cepharanthine from der-mal papilla cells: a novel potential pathway for hair growth stimulation, J. Dermatol.40 (2013) 1054–1055, http://dx.doi.org/10.1111/1346-8138.
[35] D.B. Dunger, T.D. Cheetham, E.C. Crowne, Insulin-like growth factors (IGFs) and IGF-I treatment in the adolescent with insulin-dependent diabetes mellitus, Metabolism44 (1995) 119–123.
[36] P.D. Zenobi, S.E. Jaeggi-Groisman, W.F. Riesen, M.E. Røder, E.R. Froesch, Insulin-likegrowth factor-I improves glucose and lipid metabolism in type 2 diabetes mellitus,J. Clin. Invest. 90 (1992) 2234–2241.
[37] D.K. Singh, P. Winocour, K. Farrington, Oxidative stress in early diabetic nephropa-thy: fueling the fire, Nat. Rev. Endocrinol. 7 (2011) 176–184, http://dx.doi.org/10.1038/nrendo.2010.212.
[38] J.F. Navarro-Gonza'lez, C. Mora-Ferna'ndez, M. Muros de Fuentes, J. Garcı'a-Pe'rez, Inflammatory molecules and pathways in the pathogenesis of diabeticnephropathy, Nat. Rev. Nephrol. 7 (2011) 327–340, http://dx.doi.org/10.1038/nrneph.2011.51.
[39] J. Ahmad, Management of diabetic nephropathy: recent progress and future per-spective, Diabetol. Metab. Syndr. (2015), http://dx.doi.org/10.1016/j.dsx.2015.02.008 pii:(S1871-4021(15) 00021-1).
[40] D. Halicka, M. Ita, T. Tanaka, A. Kurose, Z. Darzynkiewicz, Biscoclaurine alkaloidcepharanthine protects DNA in TK6 lymphoblastoid cells from constitutive oxida-tive damage, Pharmacol. Rep. 60 (2008) 93–100.
[41] R. Mittal, R.L. Gupta, In vitro antioxidant activity of piperine, Methods Find. Exp. Clin.Pharmacol. 22 (2000) 271–274.
[42] M. Meyer, R. Schreck, P.A. Baeuerle, H2O2 and antioxidants have opposite effects onactivation of NF-kappa B and AP-1 in intact cells: AP-1 as secondary antioxidant-re-sponsive factor, EMBO J. 12 (1993) 2005–2015.
[43] A.K. Lim, G.H. Tesch, Inflammation in diabetic nephropathy, Mediat. Inflamm. 2012(2012) 146154, http://dx.doi.org/10.1155/2012/146154.
[44] J.C. Pickup, G.D. Chusney, S.M. Thomas, D. Burt, Plasma interleukin-6, tumour necro-sis factor α and blood cytokine production in type 2 diabetes, Life Sci. 67 (2000)291–300.
[45] G. Bruno, F. Merletti, A. Biggeri, G. Bargero, S. Ferrero, G. Pagano, P. Cavallo Perin,C.M. Study, Progression to overt nephropathy in type 2 diabetes: the CasaleMonferrato study, Diabetes Care 26 (2003) 2150–2155.
[46] J. Feng, C. Guo, Y. Zhu, L. Pang, Z. Yang, Y. Zou, X. Zheng, Baicalin down regulates theexpression of TLR4 and NFkB-p65 in colon tissue in mice with colitis induced bydextran sulfate sodium, Int. J. Clin. Exp. Med. 7 (2014) 4063–4072.
[47] N.F. Abo El-Magd, A. El-Karef, M.M. El-Shishtawy, L.A. Eissa, Hepatoprotective effectsof glycyrrhizin and omega-3 fatty acids on nuclear factor-kappa B pathway inthioacetamide-induced fibrosis in rats, Egypt. J. Basic Appl. Sci. 2 (2015) 65–74,http://dx.doi.org/10.1016/j.ejbas.2014.12.005.
[48] H. Schmid, A. Boucherot, Y. Yasuda, A. Henger, B. Brunner, F. Eichinger, A. Nitsche, E.Kiss, M. Bleich, H.J. Grِne, P.J. Nelson, D. Schlِndorff, C.D. Cohen, M. Kretzler, EuropeanRenal cDNA Bank (ERCB) Consortium, Modular activation of nuclear factor-kappaBtranscriptional programs in human diabetic nephropathy, Diabetes 55 (2006)2993–3003.
[49] V. Soetikno, F.R. Sari, P.T. Veeraveedu, R.A. Thandavarayan, M. Harima, V.Sukumaran, A.P. Lakshmanan, K. Suzuki, H. Kawachi, K. Watanabe, Curcumin ame-liorates macrophage infiltration by inhibiting NF-κB activation and proinflammato-ry cytokines in streptozotocin induced-diabetic nephropathy, Nutr. Metab. (Lond.) 8(2011) 35, http://dx.doi.org/10.1186/1743-7075-8-35.
[50] C. Popa, M.G. Netea, P.L. van Riel, J.W. van der Meer, A.F. Stalenhoef, The role ofTNFalpha in chronic inflammatory conditions, intermediary metabolism, and car-diovascular risk, J. Lipid Res. 48 (2007) 751–762, http://dx.doi.org/10.1194/jlr.R600021-JLR200.
[51] G.S. Hotamisligil, P. Peraldi, A. Budavari, R. Ellis, M.F. White, B.M. Spiegelman, IRS-1-mediated inhibition of insulin receptor tyrosine kinase activity in TNF alpha- andobesity induced insulin resistance, Science 271 (1996) 665–668.
[52] C.H. Lang, C. Dobrescu, G.J. Bagby, Tumor necrosis factor impairs insulin action onperipheral glucose disposal and hepatic glucose output, Endocrinology 130 (1992)43–52.
[53] A.J. Whitmarsh, R.J. Davis, Transcription factor AP-1 regulation by mitogen-acti-vated protein kinase signal transduction pathways, J. Mol. Med. (Berl) 74 (1996)589–607.
[54] M. Haneda, S. Araki, M. Togawa, T. Sugimoto, M. Isono, R. Kikkawa, Activation of mi-togen-activated protein kinase cascade in diabetic glomeruli and mesangial cellscultured under high glucose conditions, Kidney Int. Suppl. 60 (1997) S66–S69.
[55] A.P. Lakshmanan, R.A. Thandavarayan, K. Watanabe, F.R. Sari, H. Meilei, V.V.Giridharan, V. Sukumaran, V. Soetikno, S. Arumugam, K. Suzuki, M. Kodama, Modu-lation of AT-1R/MAPK cascade by an olmesartan treatment attenuates diabetic ne-phropathy in streptozotocin-induced diabetic mice, Mol. Cell. Endocrinol. 348(2012) 104–111, http://dx.doi.org/10.1016/j.mce.2011.07.041.
[56] M. Goebeler, R. Gillitzer, K. Kilian, K. Utzel, E.B. Brِcker, U.R. Rapp, S. Ludwig, Multiplesignaling pathways regulate NF-kappaB-dependent transcription of the monocytechemoattractant protein-1 gene in primary endothelial cells, Blood 97 (2001)46–55.
[57] Y. Pan, X. Zhang, Y. Wang, L. Cai, L. Ren, L. Tang, J. Wang, Y. Zhao, Y. Wang, Q. Liu, X.Li, G. Liang, Targeting JNK by a new curcumin analog to inhibit NF-kB-mediated ex-pression of cell adhesion molecules attenuates renal macrophage infiltration and in-jury in diabetic mice, PLoS One 8 (2013) e79084, http://dx.doi.org/10.1371/journal.pone.0079084.
[58] A. Pradhan, M. Umezu, M. Fukagawa, Heme-oxygenase upregulation amelioratesangiotensin II-induced tubulointerstitial injury and salt-sensitive hypertension,Am. J. Nephrol. 26 (2006) 552–561.
[59] G.H. Tesch, Macrophages and diabetic nephropathy, Semin. Nephrol. 30 (2010)290–301, http://dx.doi.org/10.1016/j.semnephrol.2010.03.007.
[60] A. MacKenzie, H.L. Wilson, E. Kiss-Toth, S.K. Dower, R.A. North, A. Surprenant, Rapidsecretion of interleukin-1beta by microvesicle shedding, Immunity 15 (2001)825–835.
[61] Y. Nishinaka, H. Masutani, S. Oka, Y. Matsuo, Y. Yamaguchi, K. Nishio, Y. Ishii, J.Yodoi, Importin alpha1 (Rch1) mediates nuclear translocation of thioredoxin-bind-ing protein-2/vitamin D(3)-up-regulated protein 1, J. Biol. Chem. 279 (2004)37559–37565.
[62] S. Fang, Y. Jin, H. Zheng, J. Yan, Y. Cui, H. Bi, H. Jia, H. Zhang, Y. Wang, L. Na, X. Gao, H.Zhou, High glucose condition upregulated Txnip expression level in rat mesangialcells through ROS/MEK/MAPK pathway, Mol. Cell. Biochem. 347 (2011) 175–182,http://dx.doi.org/10.1007/s11010-010-0626-z.
[63] W. Qi, X. Chen, R.E. Gilbert, Y. Zhang, M. Waltham, M. Schache, D.J. Kelly, C.A.Pollock, High glucose-induced thioredoxin interacting protein in renal proximal tu-bule cells is independent of transforming growth factor-beta1, Am. J. Pathol. 171(2007) 744–754.
[64] A. Advani, R.E. Gilbert, K. Thai, R.M. Gow, R.G. Langham, A.J. Cox, K.A. Connelly, Y.Zhang, A.M. Herzenberg, P.K. Christensen, C.A. Pollock, W. Qi, S.M. Tan, H.H.Parving, D.J. Kelly, Expression, localization, and function of the thioredoxin systemin diabetic nephropathy, J. Am. Soc. Nephrol. 20 (2009) 730–741, http://dx.doi.org/10.1681/ASN.2008020142.
[65] I.N. Mohamed, S.S. Hafez, A. Fairaq, A. Ergul, J.D. Imig, A.B. El-Remessy, Thioredoxininteracting protein is required for endothelial NLRP3 inflammasome activation andcell death in a rat model of high-fat diet, Diabetologia 57 (2014) 413–423, http://dx.doi.org/10.1007/s00125-013-3101-z.
[66] R. Zhou, A. Tardivel, B. Thorens, I. Choi, J. Tschopp, Thioredoxin-interacting proteinlinks oxidative stress to inflammasome activation, Nat. Immunol. 11 (2010)136–140, http://dx.doi.org/10.1038/ni.1831.