
2) multiple stress bleaching: Temperature plus bacterial...
7
*800 ml of natural seawater *3 coral branches *Stirring continuously Coral species: Montipora digitata 䜶䝎䝁䝰䝃䞁䝂 䠄䛴䛾䝪䝖䝹䛻䠏ᮏ䠅 Circulating water: natural seawater with a flow rate of 20 ml min -1 ᐇ㦂ᾏỈ䛾ὶ㏿䠖⮬↛ᾏỈ ᐇ㦂䛾ὶ㏿ ẖศ䠎䠌䡉䡈 Incubation cases: 27Υ , 27Υ+bact., 32Υ, 32Υ+bact. 3 replicates of each) 㣫⫱᮲௳䠖 27Υ䚸27Υ䠇䝞䜽䝔䝸䜰䚸 32Υ 䚸32Υ䠇䝞䜽䝔䝸䜰䛾䠐᮲௳ Sampling on day 0, 4 and 8 ヨᩱ᥇᪥䠖䠌᪥┠䚸䠐᪥┠䚸䠔᪥┠ Illumination: metal halide lamp(~280μmol m -2 s -1 ) with 12:12h light:dark cycle 䝯䝍䝹䝝䝷䜲䝗䝷䞁䝥 䠍䠎㛫(䠅䞉䠍䠎㛫䠄ᬯ䠅 䝃䞁䝂䛾ⓑ䛾䝯䜹䝙䝈䝮䛾ゎ 㟼ᒸᏛ 㕥ᮌ Ḱ䞉䜹䝃䝺䝖 䝧䜰䝖䝸䝇 Before incubation 27Υ Υ 27Υ + Bacteria 32Υ 32Υ + Bacteria Aspect of coral branches after 8 days incubation 0 1 2 3 4 5 6 Day 0 Day 8 cells (x10 5 ) cm -2 Zooxanthellae density After 8 days of incubation 2) multiple stress bleaching: Temperature plus bacterial infection The combined effects of high temperature and bacterial infection enhance bleaching 」䝇䝖䝺䝇䛻䜘䜛ⓑ䠖 ᗘ䠇䝞䜽䝔䝸䜰ឤᰁ 〓⸴ᐦᗘ ⣽⬊ 㧗Ỉ 䛸䝞䜽䝔䝸䜰ឤ ᰁ䛾」ⓗ䛺ᙳ㡪䛜ⓑ 䜢㐍䛩䜛. 㧗Ỉ ୗ䛷䝃䞁䝂䛾⏕ ⌮ⓗᶵ⬟䛜ᙅ䛟䛺䜚䚸䛭 䛣䛻䝃䞁䝂䛜ᨺฟ䛩䜛 ᭷ᶵ≀䜔䜰䞁䝰䝙䜰䜢 ⏝䛧䛶䝞䜽䝔䝸䜰䛜ቑ Ṫ䛧䚸ឤᰁ䛧䚸ⓑ䜢 㐍䛩䜛䠄᪂䛯䛺ド䠅 8᪥㛫䛾㣫⫱ᚋ䛾䝃䞁䝂䛾ᵝᏊ 32Υ + Bacteria䛾᮲௳䛜ᐇ㦂ᮇ㛫୰䛻ⓑ䛜ㄆ䜑䜙䜜䛯 資料2
Transcript of 2) multiple stress bleaching: Temperature plus bacterial...

*800 ml of natural seawater*3 coral branches*Stirring continuously
Coral species: Montipora digitata
Circulating water: natural seawater with a flow rate of 20 ml min-1
Incubation cases: 27 , 27 +bact., 32 , 32 +bact. 3 replicates of each) 27 27 32 32
Sampling on day 0, 4 and 8
Illumination: metal halide lamp(~280μmol m-2 s-1) with 12:12h light:dark cycle(
Before incubation 27 27 + Bacteria 32 32 + Bacteria
Aspect of coral branches after 8 days incubation
0
1
2
3
4
5
6
Day 0 Day 8
cells
(x10
5 ) c
m-2
Zooxanthellae density
After 8 days of incubation
2) multiple stress bleaching: Temperature plus bacterial infectionThe combined effects of high temperature and bacterial infection enhance bleaching
.
8
32 + Bacteria
資料2
27
32
0 1000 2000 3000 4000 5000 6000
(x1000 cells/cm2)
27
32
99.3%
97.8%
82.1% 17.9%
2.2%
0.7%
52.5%
15.7%
47.5%
84.3%
27
32
99.3%
97.8%
82.1% 17.9%
2.2%
0.7%
52.5%
15.7%
47.5%
84.3%
Chlorophyll a Unknown chlorophyll PeridininP686
27
32
0
500
1000
1500
2000
2500
3000
3500
4000
Day 0 Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7
Cells
h-1
cm-2
Control (normal temperature, 27 )
0
500
1000
1500
2000
2500
3000
3500
4000
Day 0 Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7
Cells
h-1
cm-2
High temperature (32 )
Number of expelled zooxanthellae during 8 days under thermal stress
400x
1000x
1000x
400x
1000x
1000x
8
27
32
0.1%
P686
Chlorophyll a
P686
P686CPPaE (Goericke et al., 2000)
• a
a••• a
• aa a
CPPaE

( )( )
Normal condition,Less expulsion of zoox
Thermal stress,More expulsion of zoox
Bleaching
Previous concept
Normal condition,Less expulsion of zoox.
Thermal stress,More expulsionof zoox.
Bleaching
Previous concept
Bleaching
Normal condition,More expulsion of abnormalZooxanthellae
Thermal, UV and oxidative stress,Degradation of zooxanthellae and accumulation of abnormal cell inside the coral tissue before visible bleaching Visible bleaching
Our hypothesis
1) Temperature stress bleaching
.
高水温条件における造礁サンゴの白化プロセスの解明:従来説の誤りを検証
カサレト ベアトリス・鈴木 利幸・鈴木 款(創造科学技術大学院)
エダコモンサンゴを用いた室内実験を行い、高温ストレス下のサンゴ組織内と周囲の海水中の褐虫藻を分
析した結果、サンゴの白化現象はこれまで知られていたものとは大きく異なるものであることが明らかになった。
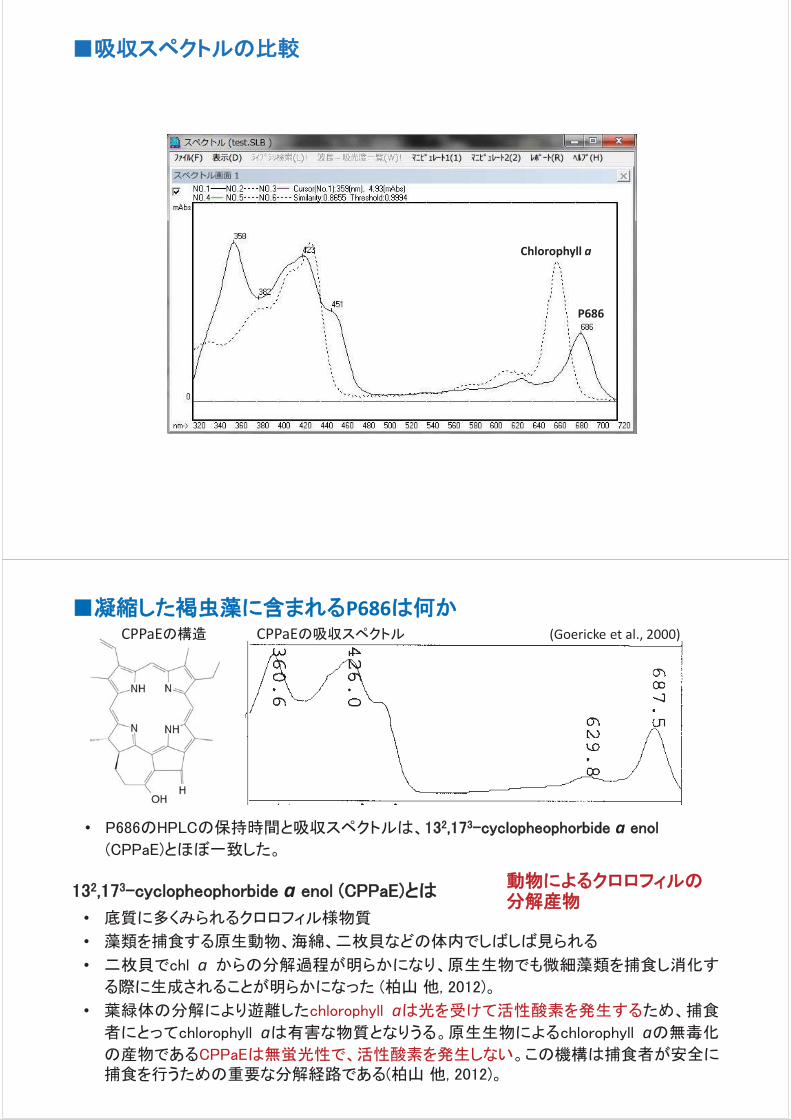
褐虫藻には正常な丸い形態の物だけではなく、葉緑体が分解され色の
抜けたもの(Bleached)や細胞自身が凝縮して小さくなったもの(Shrunken)
が観察された。正常な褐虫藻は紫外線を当てると赤い自家蛍光を発する
が、凝縮した褐虫藻は蛍光を持たなかった。これは光合成色素であるクロ
ロフィルを持っていないことを表している。この凝縮した細胞の色素組成を
分析すると、クロロフィルのかわりにシクロエノールと呼ばれる色素を持っ
ていることが明らかになった。シクロエノールはクロロフィルの分解により生
じるものであるため、凝縮した細胞は何らかの分解により生じたものである
と言える。
これまでサンゴの白化は高水温で褐虫藻が外へ逃げてしまうことで
起こると言われていた。しかし実際に飼育実験を行った結果、この考え
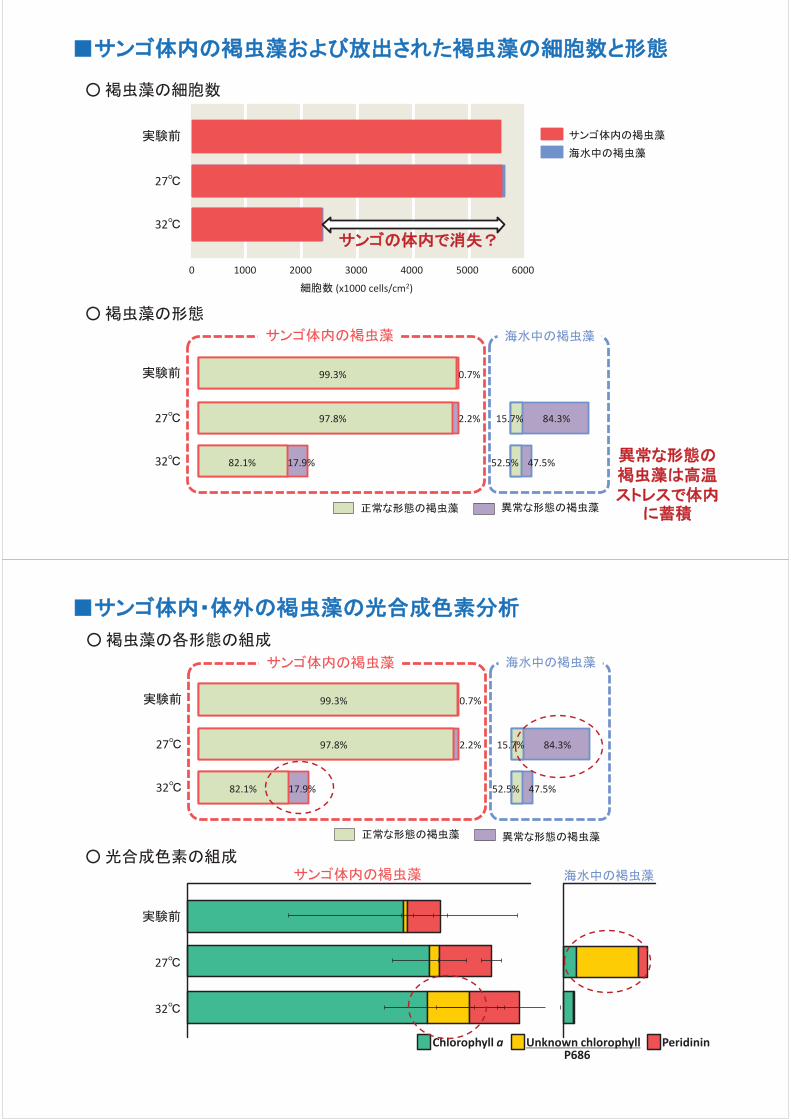
は誤りであることが明らかになった。サンゴ組織内の褐虫藻とサンゴか
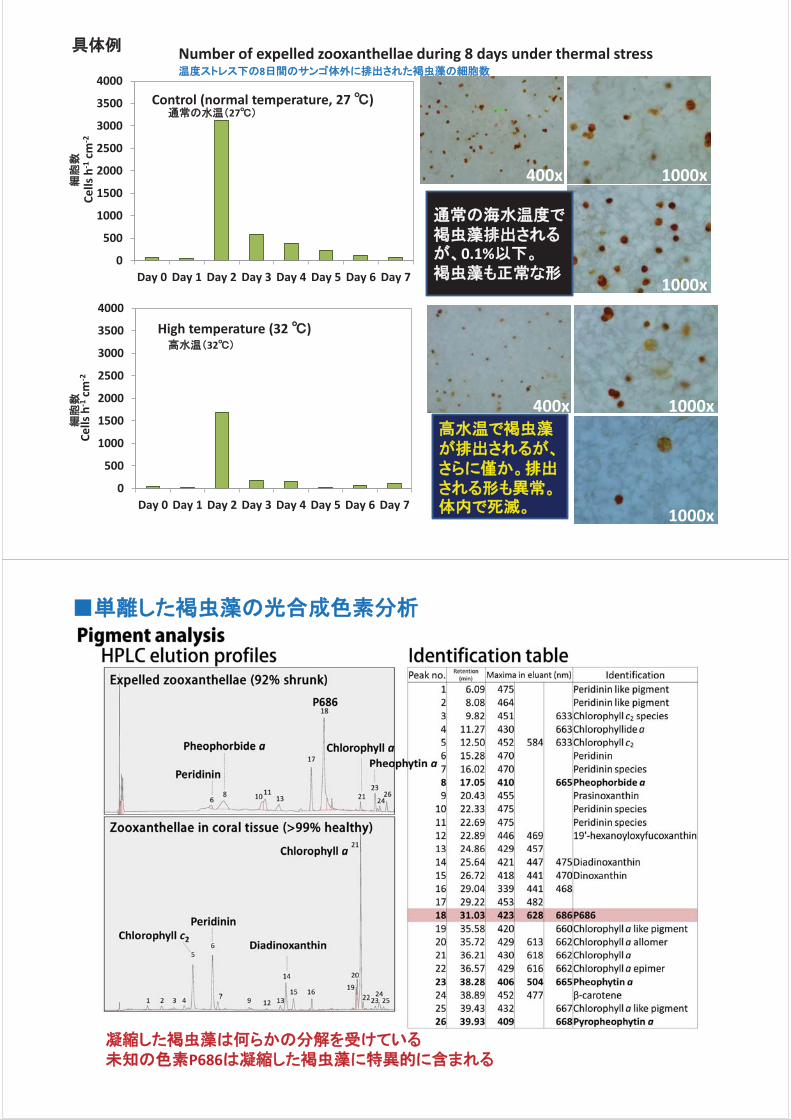
ら外へ放出された褐虫藻の数を計測した結果、通常の水温(27℃)では
褐虫藻の放出がみられるが、高水温(32℃)ではほとんど放出がないこと
が明らかになった。また 27℃で放出された褐虫藻の数は 1日あたりおよ
そ 1万~2万細胞であるが、サンゴの組織には 1cm2あたり 600万細胞
の褐虫藻が存在し、常に分裂して増殖するため 27℃の飼育では褐虫
藻の放出が起こってもサンゴ組織内の細胞数に変化がみられない。さ
らに放出された褐虫藻の大半は凝縮した形態の細胞であった(体内で
は 1%以下の存在率)。これらのことから、サンゴからの褐虫藻の放出
は、健康なサンゴの体内のメンテナンスによるものであり、放出が原因で
白化が起こるわけではないと考えられる。
高水温条件では褐虫藻の放出はほとんど無い
のに、なぜサンゴから褐虫藻がいなくなってしまう
のか。サンゴ組織内の凝縮した褐虫藻の数を数え
てみると、高温ストレスを受けたサンゴでは放出が
減る代わりに組織内の凝縮した褐虫藻の割合が
大幅に増加していた(約 20%。健康なサンゴでは 1%
以下)。従って白化現象は、サンゴから褐虫藻が逃
げることで起こるのではなく、サンゴ体内で褐虫藻
が分解されることで起こることが明らかになった。
27℃
32℃

サンゴは褐虫藻に栄養塩類を供給し、褐虫藻
はサンゴに光合成で得られた有機物を供給する共
生関係を築いている。それなのになぜサンゴは自
ら褐虫藻を分解してしまうのか。褐虫藻は高水温
条件下で葉緑体が損傷し再生できなくなることが
知られている。損傷した葉緑体からは、クロロフィ
ルが遊離する。クロロフィルは光合成を行うために
重要な色素で、葉緑体内に配置されているときは
光エネルギーを正常に光合成機関に受け渡す
が、遊離したクロロフィルにはエネルギーの受け渡
し先がないために光エネルギーを電子として放出
する。放出された電子は水中の酸素と反応し、活性酸素を発生させる。活性酸素は酸化力が強く周囲の細胞
(サンゴ)を傷付けてしまう。すなわち高水温条件下で葉緑体の損傷した褐虫藻は、光合成を行わないだけでな
く有害な活性酸素を発生する、サンゴにとって危険な存在である。ところがクロロフィルの分解産物であるシク
ロエノールは、光を受けても電子を放出せず、活性酸素を発生させない。サンゴは損傷した褐虫藻を分解する
ことで凝縮させ、酸化ストレスを軽減し、残った褐虫藻による光合成産物や触手による捕食によって水温が正
常化するのを待つことで生き延びようとするのだろう。自ら褐虫藻を分解するメカニズムは、サンゴの高温スト
レスに対する防御応答であると考えられる。これまでサンゴは白化の被害者であると考えられてきたが、この一
連のメカニズムから、白化現象はサンゴにとっての生存戦略の一部であるのかもしれない。
Ramphul C, Casareto BE, Suzuki T, Yoshinaga K, Yeemin T, Suzuki Y (2015): Abundance of virus-like particles and its
links to phytoplankton, bacteria and nutrients cycling in coastal coral ecosystem. Eco-Engineering, 27 (3), 81-90
Miyata W, Suzuki T, Casareto BE, Suzuki Y, Shioi Y (2015): A survey of photosynthetic pigments from surface to oligotrophic
deep seawater in Suruga Bay, Japan. Procedia Chemistry 14, 444-454. doi: 10.1016/j.proche.2015.03.060
Suzuki T, Casarteto BE, Shioi Y, Ishikawa Y, Suzuki Y (2015): Finding of 132, 173-cyclopheophorbide a enol as a
degradation product of Chlorophyll in shrunk zooxanthellae of the coral Montipora digitata. Journal of Phycology,
51 (1) 37-45 DOI: 10.1111/jpy.12253
Higuchi T, Agostini S, Casareto BE, Yoshinaga K, Suzuki T, Nakano Y , Fujimura H, Suzuki Y (2013): Bacterial
enhancement of bleaching and physiological impacts on the coral Montipora digitata. Journal of Experimental
marine Biology and Ecology, 440, 54–60
Weil E, Irikawa A, Casareto BE, Suzuki Y (2012): Extended geographic distribution of several Indo-Pacific coral reef
diseases. Diseases of Aquatic Organisms, vol. 98, no. 2, pp. 163–170
Agostini S, Suzuki Y, Higuchi T, Casareto BE, Yoshinaga K, Nakano Y, Fujimura H. (2012): Biological and chemical
characteristics of the coral gastric cavity. Coral Reefs, vol. 31, no. 1, pp. 147–156
(国際サンゴ礁学会から 2012最優秀論文賞受賞)















![EXT-T24-D201 LCD Temperature Controller - …V1.2_22_9_2017].pdf · EXT-T24-D201 LCD Temperature Controller ... LCD temperature controller EXT-T24-D201 provides the foundation for](https://static.fdocument.org/doc/165x107/5a80a5287f8b9a0c748c8809/ext-t24-d201-lcd-temperature-controller-v122292017pdfext-t24-d201-lcd.jpg)


