α-Adrenergic receptors in human myometrium during pregnancy
6
Volume 154 Number 3 occurring in 46,XX individuals. Clin Obstet Gynecol 1972;15:157. 3. Simpson jL, Elias S, Malinak RL, Buttram VC. Heritable aspects of endometriosis. I. Genetic studies. AM j OSSTET GYNECOL 1980;137:327. 4. Malinak RL, Buttram VC, Elias S, Simpson jL. Her- itable aspects of endometriosis. II. Clinical character- istics of familial endometriosis. AM j OSSTET GYNECOL 1980; 137:332. 5. Kistner RW. Current concepts in endometriosis: an in- ternational review. j Reprod Med 1977; 19:299. 6. Dmowski WP, Radwanska E. Current concepts on pa- thology, histogenesis and etiology of endometriosis. Acta Obstet Gynecol Scand Suppl 1984;123:29. 7. Weed jC, Arquembourg PC. Endometriosis: can it pro- duce an autoimmune response resulting in infertility? Clin Obstet Gynecol 1980;23:885. 8. Badawy SZA, Cuenca V, Stitzel A, jacobs RDB, Tomar RH. Autoimmune phenomena in infertile patients with endometriosis. Obstet Gynecol 1984;63:271. Familial tendencies to endometriosis 9. Coulam CB, Ryan RJ. Premature menopause. l. Etiology. AM j OSSTET GYNECOL 1979; 133:639. 10. Shands KN, Schlech WF, Hargrett NT, Dan BB, Schmid GP, BennettjV. Toxic shock syndrome: case-control stud- ies at the Centers for Disease Control. Ann Intern Med 1982;96:895. II. Halgerson SD, Foster LR. Toxic shock syndrome in ore- gon: epidemiologic findings. Ann Intern Med 1982; 96:909. 12. Osterholm MT, Davis jP, Gibson RW, Forfang jC, Stolz Sj, VergerontjM. Toxic shock syndrome: relation to ca- tamenial products, personal health and hygiene, and sex- ual practices. Ann Intern Med 1982;96:954. 13. Shands KN, Schmid GP, Dan BB, et al. Toxic shock syn- drome in menstruating women: association with tampon use and staphyloccus au reus and clinical features in 52 cases. N Engl j Med 1980;303: 1436. 14. Houston DE. Evidence for the risk of pelvic endometriosis by age, race and socioeconomic status. Epidemiol Rev 1984;6:167. a-Adrenergic receptors in human myometrium during pregnancy Goran Berg, M.D., Rolf G. G. Andersson, Ph.D., and Gunnar Ryden, M.D. LinkOping, Sweden The distribution and mechanisms of Ct-adrenergic receptors have been studied in myometrial preparations from women delivered by cesarean section at term. With the use of the radioligand tritiated dihydroergocryptine, the number of Ct-adrenergic receptors was 210 fmol/mg of protein in the uterine fundus and 195 fmol/mg of protein in the lower uterine segment. Competition experiments showed that 60% of the Ct-adrenergic receptors had properties as Ct,-adrenergic receptors and 40% as Ct 2 -adrenergic receptors. In vitro tension studies verified the existence of physiologically active Ct,- and Ct 2 -adrenergic receptors in the myometrial preparations. Ct2-Adrenergic receptor stimulation resulted in lowered levels of intracellular cyclic adenosine monophosphate. The intracellular cyclic adenosine monophosphate was further reduced by additional Ct,-adrenergic receptor stimulation, probably secondary to an activation of calcium-calmodulin-dependent phosphodiesterase. (AM J OBSTET GYNECOL 1986;154:601-6.) Key words: a-Adrenergic receptors, myometrium, labor, cyclic adenosine mono phosphate There is considerable experimental evidence that the human myometrium contains both 13- and a-adrenergic receptors. I. 2 The mechanism of action of l3-adrenergic receptors in the myometrium has been thoroughly studied.' However, the distribution of a-adrenergic re- ceptors in human myometrium has just recently been investigated and only in nonpregnant women. 2 The From the Department of Obstetrics and Gynecology and the Depart- ment of Pharmacology, Linkoping University. This study was financially supported by Ostergotlands ltins landsting, the Expressen Perinatal Research Foundation, the Swedish Medical Society, Tore Nilsson's Fund, and the Medical Research Council (Grant 04P-6155). Received for publication june 7, 1985; revised November 21, 1985; accepted November 25, 1985. Reprint requests: Goran Berg, M.D., Department of Obstetrics and Gynecology, Linkoping Unversity, S-581 85 Linkoping, Sweden. aim of this study was, therefore, to evaluate the a-ad- renergic receptor distribution and function in myo- metrium from pregnant women and to study in a conventional in vitro system the effect of different pharmacologic a-adrenergic receptor blockers on spon- taneously contracting myometrium and in receptor- binding assays in order to classify the subtypes of a- adrenergic receptors and to look for possible new means to improve today's tocolytic treatment. Material and methods Patients. Myometrial biopsy specimens were taken during cesarean section from the lower uterine seg- ment and/or the uterine fundus. The reason for ce- sarean section was related to obstetrics in all cases. The gestational ages were between 37 and 41 weeks and 601
Transcript of α-Adrenergic receptors in human myometrium during pregnancy

Volume 154 Number 3
occurring in 46,XX individuals. Clin Obstet Gynecol 1972;15:157.
3. Simpson jL, Elias S, Malinak RL, Buttram VC. Heritable aspects of endometriosis. I. Genetic studies. AM j OSSTET GYNECOL 1980;137:327.
4. Malinak RL, Buttram VC, Elias S, Simpson jL. Heritable aspects of endometriosis. II. Clinical characteristics of familial endometriosis. AM j OSSTET GYNECOL 1980; 137:332.
5. Kistner RW. Current concepts in endometriosis: an international review. j Reprod Med 1977; 19:299.
6. Dmowski WP, Radwanska E. Current concepts on pathology, histogenesis and etiology of endometriosis. Acta Obstet Gynecol Scand Suppl 1984;123:29.
7. Weed jC, Arquembourg PC. Endometriosis: can it produce an autoimmune response resulting in infertility? Clin Obstet Gynecol 1980;23:885.
8. Badawy SZA, Cuenca V, Stitzel A, jacobs RDB, Tomar RH. Autoimmune phenomena in infertile patients with endometriosis. Obstet Gynecol 1984;63:271.
Familial tendencies to endometriosis
9. Coulam CB, Ryan RJ. Premature menopause. l. Etiology. AM j OSSTET GYNECOL 1979; 133:639.
10. Shands KN, Schlech WF, Hargrett NT, Dan BB, Schmid GP, BennettjV. Toxic shock syndrome: case-control studies at the Centers for Disease Control. Ann Intern Med 1982;96:895.
II. Halgerson SD, Foster LR. Toxic shock syndrome in oregon: epidemiologic findings. Ann Intern Med 1982; 96:909.
12. Osterholm MT, Davis jP, Gibson RW, Forfang jC, Stolz Sj, VergerontjM. Toxic shock syndrome: relation to catamenial products, personal health and hygiene, and sexual practices. Ann Intern Med 1982;96:954.
13. Shands KN, Schmid GP, Dan BB, et al. Toxic shock syndrome in menstruating women: association with tampon use and staphyloccus au reus and clinical features in 52 cases. N Engl j Med 1980;303: 1436.
14. Houston DE. Evidence for the risk of pelvic endometriosis by age, race and socioeconomic status. Epidemiol Rev 1984;6:167.
a-Adrenergic receptors in human myometrium during
pregnancy
Goran Berg, M.D., Rolf G. G. Andersson, Ph.D., and Gunnar Ryden, M.D.
LinkOping, Sweden
The distribution and mechanisms of Ct-adrenergic receptors have been studied in myometrial preparations from women delivered by cesarean section at term. With the use of the radioligand tritiated dihydroergocryptine, the number of Ct-adrenergic receptors was 210 fmol/mg of protein in the uterine
fundus and 195 fmol/mg of protein in the lower uterine segment. Competition experiments showed that 60% of the Ct-adrenergic receptors had properties as Ct,-adrenergic receptors and 40% as Ct2-adrenergic
receptors. In vitro tension studies verified the existence of physiologically active Ct,- and Ct2-adrenergic receptors in the myometrial preparations. Ct2-Adrenergic receptor stimulation resulted in lowered levels of
intracellular cyclic adenosine monophosphate. The intracellular cyclic adenosine monophosphate was further reduced by additional Ct,-adrenergic receptor stimulation, probably secondary to an activation of calcium-calmodulin-dependent phosphodiesterase. (AM J OBSTET GYNECOL 1986;154:601-6.)
Key words: a-Adrenergic receptors, myometrium, labor, cyclic adenosine mono phosphate
There is considerable experimental evidence that the human myometrium contains both 13- and a-adrenergic receptors. I. 2 The mechanism of action of l3-adrenergic receptors in the myometrium has been thoroughly studied.' However, the distribution of a-adrenergic receptors in human myometrium has just recently been investigated and only in nonpregnant women. 2 The
From the Department of Obstetrics and Gynecology and the Department of Pharmacology, Linkoping University.
This study was financially supported by Ostergotlands ltins landsting, the Expressen Perinatal Research Foundation, the Swedish Medical Society, Tore Nilsson's Fund, and the Medical Research Council (Grant 04P-6155).
Received for publication june 7, 1985; revised November 21, 1985; accepted November 25, 1985.
Reprint requests: Goran Berg, M.D., Department of Obstetrics and Gynecology, Linkoping Unversity, S-581 85 Linkoping, Sweden.
aim of this study was, therefore, to evaluate the a-adrenergic receptor distribution and function in myometrium from pregnant women and to study in a conventional in vitro system the effect of different pharmacologic a-adrenergic receptor blockers on spontaneously contracting myometrium and in receptorbinding assays in order to classify the subtypes of aadrenergic receptors and to look for possible new means to improve today's tocolytic treatment.
Material and methods
Patients. Myometrial biopsy specimens were taken during cesarean section from the lower uterine segment and/or the uterine fundus. The reason for cesarean section was related to obstetrics in all cases. The gestational ages were between 37 and 41 weeks and
601

602 Berg, Andersson, and Ryden
active labor had not started in any patients at the time of operative delivery. About two thirds of the women were anesthesized with an epidural block, and the remainder received the conventional intravenous anesthetics thiopental sodium and leptanale. The myometrial preparations were immediately immersed in Ringer's solution and transported to the laboratory. Biopsy specimens from a total of 48 women were studied.
Studies of contractility in vitro. The myometrial preparations were mounted in special holders for registration of isometric tension by FT03 transducers on a Grass polygraph. The myometrial strips were suspended in a Krebs buffer solution at 37° C with the following composition: sodium chloride, 118.8 mmol/L; potassium chloride, 4.7 mmollL; calcium chloride, 2.2 mmollL; magnesium chloride, 1.2 mmollL; sodium bicarbonate, 23.8 mmollL; potassium phosphate, 1.2 mmol/L; glucose, 5.5 mmol/L. The solution was aerated with a gas mixture containing 95% oxygen and 5% carbon dioxide. All myometrial preparations were either untreated or pretreated with (1) phentolamine (ai-and a,-adrenergic receptor blocker), (2) yohimbine (a,-adrenergic receptor blocker), and (3) pralOsin (ai-adrenergic receptor blocker). All preparations were then treated with increasing concentrations of norepinephrine and the tension was measured.
Preparation of myometrial membranes. The myometrial tissue was cut into small pieces and homogenized in 10 volumes of buffer solution (sucrose, 0.25 mollL; Tris, 5 mmollL; magnesium chloride, 1 mmollL; pH 7.4) with the use of a Polytron PT 10. During the preparation procedure, the myometrial tissue was kept in ice-cold buffer solution. The homogenate was filtered through a cheese-cloth and thereafter centrifugated at 400 x g for 10 minutes at 4° C. The supernatant was centrifugated at 28,000 x g for 10 minutes. The resulting pellets were washed and recentrifuged twice at 28,000 x g. The washed pellets were suspended in three volumes of incubation buffer (Tris, 50 mmollL; magnesium chloride, 10 mmollL; pH 7.5) and used for the determination of tritiated dihydroergocryptine binding.
Binding of tritiated dihydroergocryptine. Binding of tritiated dihydroergocryptine to human myometrial membranes was assayed by filtration of the preparations on glass fiber filters.' Membrane proteins were incubated with the indicated concentrations of tritiated dihydroergocryptine and incubation buffer in a total volume of 0.15 ml for 20 minutes at 37° C. At the end of incubation the samples were diluted in 2 ml of ice
cold incubation buffer, followed by rapid vacuum filtration through Whatman glass fiber filters on a Millipore Sampling Manifold. The filters were then rapidly washed six times with 3 ml of the ice-cold in-
March, 1986 Am J Obstet Gynecol
cubation buffer. After drying, radioactivity bound to the filters was determined on a Packard liquid scintillation spectrometer. Specific binding was determined by subtracting nonspecific binding from total binding. In all figures and tables, bound tritiated dihydroergocryptine refers to specific binding. Nonspecific binding was determined in the presence of phentolamine, 10 fJ..mollL. Protein was determined according to Lowry et al. 4
Determination of cyclic adenosine monophosphate in myometrial tissue. Stimulation with norepinephrine (1 fJ..moIlL) of the myometrial strips was performed in an organ bath with or without prazosin (1 fJ..moIlL) or yohimbine (1 fJ..moIlL) and in the same manner with myometrial preparations pretreated with propranolol (1 fJ..moIlL). After treatment with norepinephrine for 3 minutes in the organ bath, the strips were dismounted and frozen in Frigen 12 and solid carbon dioxide at -70° C.
The frozen preparations were homogenized in perchloric acid and the nucleotides were separated on columns of Ag-1-X-8 (200 to 400 mesh, formate form). After elution with formic acid the eluates were lyophilized. The cyclic adenosine monophosphate content was determined according to Gilman.5
Determination of phosphodiesterase activity. The activity of phosphodiesterase was determined by the method described by Thompson and Appleman6 with minor modifications. After 30,000 x g centrifugation of the myometrial homogenate for 10 minutes, the supernatant was used for analysis. The assay was performed in a solution containing Tris hydrochloride, 40 mmollL, pH 7.5; magnesium chloride, 5 mmollL; 20 fJ..g of 5-nucleotidase (snake venom), and tritiated cyclic adenosine monophosphate, 1 fJ..moIlL, containing more than 100,000 cpm, in a total volume of 0.1 ml. After 3 to 5 minutes at 37° C the reaction was terminated by heating the reaction mixture at 90° C for 1 minute, and 0.4 ml of distilled water was then added to the incubation tube. The entire contents of the tubes were transferred to prewashed AG 1 x 8 columns (C 1 form, 0.5 by 1.0 cm) followed by addition of 1 ml of distilled water for sampling of fractions containing the tritiated adenosine. The activity was measured in a liquid scintillation counter. Calcium-dependent phosphodiesterase activity was measured in six strips. Ionized calcium was added in increasing concentrations to an enzyme preparation dialyzed in Tris buffer containing ethyleneglycol-bis(l3-amino-ethyl ether)n,n'tetraacetic acid (EGTA), and phosphodiesterase activity was measured
as described above. In 12 myometrial preparations trifluoperazine was added in increasing concentrations (0.1 fJ..mollL to 1 mmol/L of trifluoperazine) (see Fig.
5, Aand B).
Volume 154 Number 3
~ .. .2
J 'Ii
I .. .!
101 [nond"RlIiIll] M
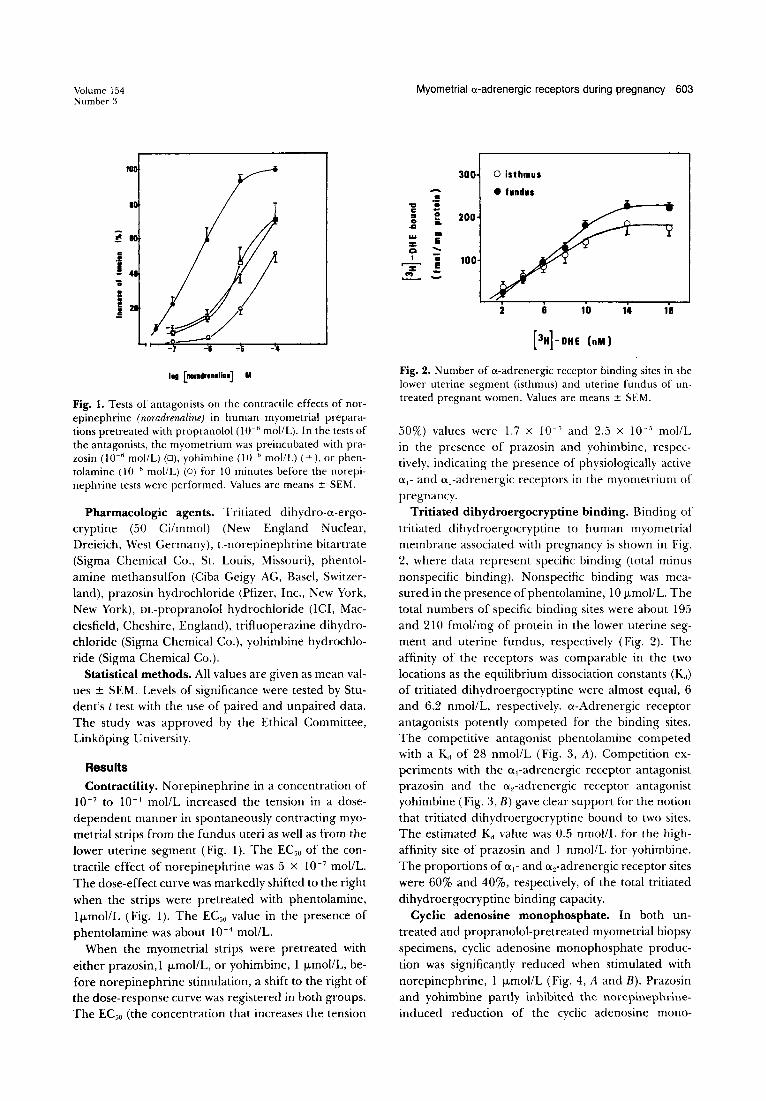
Fig.!. Tests of antagonists on the contractile effects of norepinephrine (noradrenaline) in human myometrial preparations pretreated with propranolol (10- 6 moI/L). In the tests of the antagonists, the myometrium was pre incubated with prazosin (10- 6 mollL) (0), yohimbine (10- 6 mollL) (+), or phentolamine (10- 6 mollL) (0) for 10 minutes before the norepinephrine tests were performed. Values are means ± SEM.
Pharmacologic agents. Tritiated dihydro-a-ergocryptine (50 Ci/mmol) (New England Nuclear, Dreieich, West Germany), L-norepinephrine bitartrate (Sigma Chemical Co., St. Louis, Missouri), phentolamine methansulfon (Ciba Geigy AG, Basel, Switzerland), prazosin hydrochloride (Pfizer, Inc., New York, New York), DL-propranolol hydrochloride (ICI, Macclesfield, Cheshire, England), trifluoperazine dihydrochloride (Sigma Chemical Co.), yohimbine hydrochloride (Sigma Chemical Co.).
Statistical methods. All values are given as mean values ± SEM. Levels of significance were tested by Student's t test with the use of paired and unpaired data. The study was approved by the Ethical Committee, Link6ping University.
Results Contractility. Norepinephrine in a concentration of
10- 7 to 10-4 mol/L increased the tension in a dosedependent manner in spontaneously contracting myometrial strips from the fundus uteri as well as from the lower uterine segment (Fig. 1). The ECoo of the contractile effect of norepinephrine was 5 X 10- 7 mollL. The dose-effect curve was markedly shifted to the right when the strips were pretreated with phentolamine, Ij.LmollL (Fig. 1). The ECoo value in the presence of phentolamine was about 10- 4 mollL.
When the myometrial strips were pretreated with either prazosin, 1 j.Lmol/L, or yohimbine, 1 j.LmollL, before norepinephrine stimulation, a shift to the right of the dose-response curve was registered in both groups. The EC50 (the concentration that increases the tension
Myometrial a-adrenergic receptors during pregnancy 603
300 o isthmus
- • fundus .. .. . ~ .. ,. f! 200 0 .. ... ... .. = E ell ...... I 'i 100
'Z" E .!:!..- -....
2 • 10 14 18
Fig. 2. Number of a-adrenergic receptor binding sites in the lower uterine segment (isthmus) and uterine fundus of untreated pregnant women. Values are means ± SEM.
50%) values were 1.7 X 10- 5 and 2.5 x 10-' mollL in the presence of prazosin and yohimbine, respectively, indicating the presence of physiologically active 0.,- and a,-adrenergic receptors in the myometrium of pregnancy.
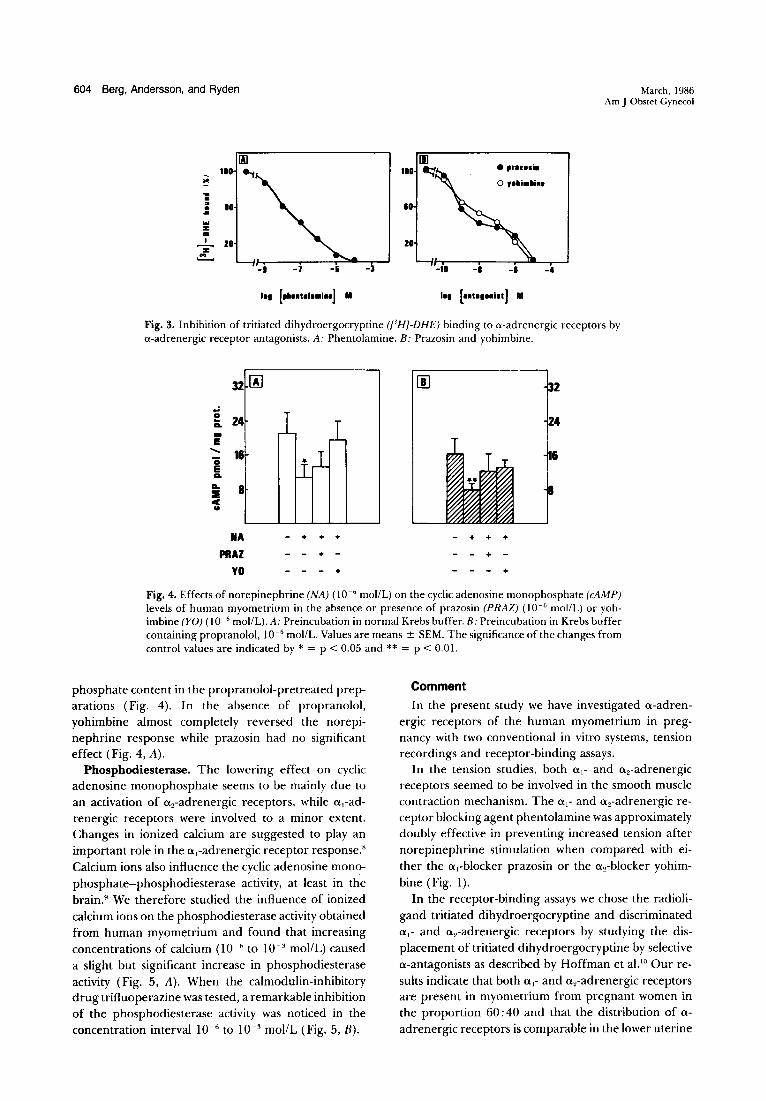
Tritiated dihydroergocryptine binding. Binding of tritiated dihydroergocryptine to human myometrial membrane associated with pregnancy is shown in Fig. 2, where data represent specific binding (total minus nonspecific binding). Nonspecific binding was measured in the presence of phentolamine, 10 j.LmollL. The total numbers of specific binding sites were about 195 and 210 fmol/mg of protein in the lower uterine segment and uterine fundus, respectively (Fig. 2). The affinity of the receptors was comparable in the two locations as the equilibrium dissociation constants (K,,) of tritiated dihydroergocryptine were almost equal, 6 and 6.2 nmollL, respectively. a-Adrenergic receptor antagonists potently competed for the binding sites. The competitive antagonist phentolamine competed with a K" of 28 nmol/L (Fig. 3, A). Competition experiments with the ai-adrenergic receptor antagonist prazosin and the a,-adrenergic receptor antagonist yohimbine (Fig. 3, B) gave clear support for the notion that tritiated dihydroergocryptine bound to two sites. The estimated Kd value was 0.5 nmollL for the highaffinity site of prazosin and 1 nmol/L for yohimbine. The proportions of 0.,- and a 2-adrenergic receptor sites were 60% and 40%, respectively, of the total tritiated dihydroergocryptine binding capacity.
Cyclic adenosine monophosphate. In both untreated and propranolol-pretreated myometrial biopsy specimens, cyclic adenosine monophosphate production was significantly reduced when stimulated with norepinephrine, 1 j.LmollL (Fig. 4, A and B). Prazosin and yohimbine partly inhibited the norepinephrineinduced reduction of the cyclic adenosine mono-
604 Berg, Andersson, and Ryden
III 100 ..., .. .. .. • 10 • .. ...
z co ,
20 '"Z .!!......
2
-I -& -3 -4
I.. [ •• " ••• il'] M
March, 1986 Am J Obstet Gynecol
Fig. 3. Inhibition of tritiated dihydroergocryptine ([3 HJ-DHE) binding to a-adrenergic receptors by a-adrenergic receptor antagonists. A: Phentolamine. B: Prazosin and yohimbine.
32 [A] 00 .. ~ 24 IL -E
...... 16 .. E IL
a. 8 ::Ii c u
NA - + + + - + + +
PHAZ + - - - + -YO - + +
Fig. 4. Effects of norepinephrine (NA) (10- 6 mollL) on the cyclic adenosine monophosphate (cAMP) levels of human myometrium in the absence or presence of prazosin (PRAZ) (10- 6 mollL) or yohimbine (YO) (10- 6 moIlL). A: Preincubation in normal Krebs buffer. B: Preincubation in Krebs buffer containing propranolol, 10- 6 mollL. Values are means ± SEM. The significance of the changes from control values are indicated by * = P < 0.05 and ** = p < 0.01.
phosphate content in the propranolol-pretreated preparations (Fig. 4). In the absence of propranolol, yohimbine almost completely reversed the norepinephrine response while prazosin had no significant effect (Fig. 4, A).
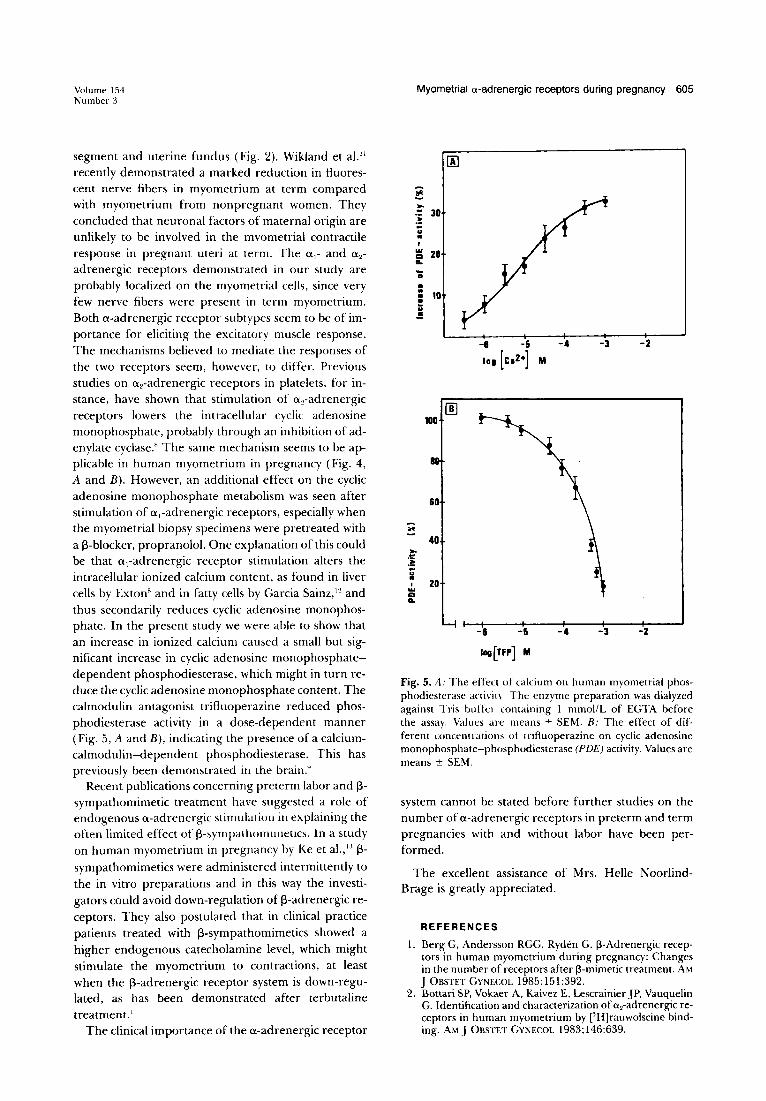
Phosphodiesterase. The lowering effect on cyclic adenosine monophosphate seems to be mainly due to an activation of a 2-adrenergic receptors, while a,-adrenergic receptors were involved to a minor extent. Changes in ionized calcium are suggested to play an important role in the a,-adrenergic receptor response.8
Calcium ions also influence the cyclic adenosine monophosphate-phosphodiesterase activity, at least in the brain.9 We therefore studied the influence of ionized calcium ions on the phosphodiesterase activity obtained from human myometrium and found that increasing concentrations of calcium (10- 6 to 10- 3 mollL) caused a slight but significant increase in phosphodiesterase activity (Fig. 5, A). When the calmodulin-inhibitory drug trifluoperazine was tested, a remarkable inhibition of the phosphodiesterase activity was noticed in the concentration interval 10- 6 to 10- 3 moUL (Fig. 5, B).
Comment
In the present study we have investigated a-adrenergic receptors of the human myometrium in pregnancy with two conventional in vitro systems, tension recordings and receptor-binding assays.
In the tension studies, both a,- and a 2-adrenergic receptors seemed to be involved in the smooth muscle contraction mechanism. The a,- and a 2-adrenergic receptor blocking agent phentolamine was approximately doubly effective in preventing increased tension after norepinephrine stimulation when compared with either the a,-blocker prazosin or the a 2-blocker yohimbine (Fig. 1).
In the receptor-binding assays we chose the radioligand tritiated dihydroergocryptine and discriminated a,- and a 2-adrenergic receptors by studying the displacement of tritiated dihydroergocryptine by selective a-antagonists as described by Hoffman et al.lO Our results indicate that both a,- and a 2-adrenergic receptors are present in myometrium from pregnant women in the proportion 60: 40 and that the distribution of a
adrenergic receptors is comparable in the lower uterine
Volume 154 Number 3
segment and uterine fundus (Fig. 2). Wikland et al." recently demonstrated a marked reduction in fluorescent nerve fibers in myometrium at term compared with myometrium from nonpregnant women. They concluded that neuronal factors of maternal origin are unlikely to be involved in the mvometrial contractile response in pregnant uteri at term. The a l - and a 2 -
adrenergic receptors demonstrated in our study are probably localized on the myometrial cells, since very few nerve fibers were present in term myometrium. Both a-adrenergic receptor subtypes seem to be of importance for eliciting the excitatory muscle response. The mechanisms believed to mediate the responses of the two receptors seem, however, to differ. Previous studies on a 2-adrenergic receptors in platelets, for instance, have shown that stimulation of a 2-adrenergic receptors lowers the intracellular cyclic adenosine monophosphate, probably through an inhibition of adenylate cyclase." The same mechanism seems to be applicable in human myometrium in pregnancy (Fig. 4, A and B). However, an additional effect on the cyclic adenosine monophosphate metabolism was seen after stimulation of ai-adrenergic receptors, especially when the myometrial biopsy specimens were pretreated with a i3-blocker, propranolol. One explanation of this could be that ai-adrenergic receptor stimulation alters the intracellular ionized calcium content, as found in liver cells by Exton" and in fatty cells by Garcia Sainz,12 and thus secondarily reduces cyclic adenosine monophosphate. In the present study we were able to show that an increase in ionized calcium caused a small but significant increase in cyclic adenosine monophosphatedependent phosphodiesterase, which might in turn reduce the cyclic adenosine monophosphate content. The calmodulin antagonist trifluoperazine reduced phosphodiesterase activity in a dose-dependent manner (Fig. 5, A and B), indicating the presence of a calciumcalmodulin-dependent phosphodiesterase. This has previously been demonstrated in the brain."
Recent publications concerning preterm labor and 13-sympathomimetic treatment have suggested a role of endogenous a-adrenergic stimuhuioll ill explaining the often limited effect of i3-sympathollllllletics. In a study on human myometrium in pregnancy by Ke et al.,l:l 13-sympathomimetics were administered intermittently to the in vitro preparations and in this way the investigators could avoid down-regulation of i3-adrenergic receptors. They also postulated that in clinical practice patients treated with i3-sympathomimetics showed a higher endogenous catecholamine level, which might stimulate the myometrium to contractions, at least when the i3-adrenergic receptor system is down-regulated, as has been demonstrated after terbutaline treatment. I
The clinical importance of the a-adrenergic receptor
Myometrial a-adrenergic receptors during pregnancy 605
60
.. 40
~
:i -u • 20 I ... Q Go
-6 -5 -4 -3 -2
Fig. 5. A: The effect of calcium on human myometrial phosphodiesterase aClivit\ The enzyme preparation was dialyzed against Tris buller containing I mmollL of EGTA before the assay. Values are means ± SEM. B: The effect of different concentrations of trifluoperazine on cyclic adenosine monophosphate-phosphodiesterase (PDE) activity. Values are means ± SEM.
system cannot be stated before further studies on the number of a-adrenergic receptors in preterm and term pregnanCies with and without labor have been performed.
The excellent assistance of Mrs. Helle NoorlindBrage is greatly appreciated.
REFERENCES
I. Berg G, Andersson RGG, Ryden G. I3-Adrenergic receptors in human myometrium during pregnancy: Changes in the number of receptors after l3-mimetic treatment. AM J OBSTET GYNECOL 1985;151:392.
2. Bottari SP, Vokaer A, Kaivez E, Lescrainier JP, Vauquelin G. Identification and characterization of a 2-adrenergic receptors in human myometrium by ['H]rauwolscine binding. AM J OBSTET GYNECOL 1983; 146:639.

Berg, Andersson, and Ryden
3. Huszar G, Roberts JM. Biochemistry and pharmacology of the myometrium and labor: Regulation at the cellular and molecular levels. AM J OBSTET GV"ECOI. I 9S2; 142:225.
4. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Bioi Chern 1951; 193:265.
5. Gilman AG. A protein binding assay for adenosine 3'5' cyclic monophosphate. Proc Nat! Acad Sci USA 1970; 67:305.
6. Thompson WJ, Appleman MM. Multiple cyclic nucleotide phosphodiesterase activities from rat brain. Biochemistry 1971;10:311.
7. Williams LT, Mullikin D, Lefkowitz RJ. Identiffication of alpha-adrenergic receptors in uterine smooth muscle membranes by "H-dihydroergocryptine binding. J Bioi Chern 1976;251:6915.
S. Exton J. Molecular mechanism involved in alpha-adrenergic responses. Mol Cell Endocrinol I 9S1 ;23:233.
March, 1986 Am J Obstet Gynecol
9. Kakiuchi S, Yamazaki R. Calcium dependent phosphodiesterase activity and its activating factor (PAS) from brain. Biochem Biophys Res Commun 1970;41: II 04.
10. Hoffman BB, DeLean A, Wood CL, Schocken DD, Lefkowitz RJ. Alpha adrenergic receptor sUbtypes: quantitative assessment by ligand binding. Life Sci 1979; 24: 1739.
II. Wikland M, Lindblom B, Dahlstrom A, Haglid KG. Structural and functional evidence for the denervation of human myometrium during pregnancy. Obstet Gynecol 19S4;64:503.
12. Garcia-SainzJA, Hoffman BB, Li S-Y, Lefkowitz RJ, Fain JF. Role of alpha-I adrenoceptors in the turnover of phosphatidylinositol and of alpha-2 adrenoceptors in the regulation of cyclic AMP accumulation in hamster adipocytes. Life Sci 19S0;27:953.
13. Ke R, Vohra M, Caspar R. Prolonged inhibition of human myometrial contractility by intermittent isoproterenol. AM J OBSTET GVNECOL 19S4;149:S41.
Corticotropin-releasing factor can stimulate gonadotropin
secretion by human fetal pituitaries in superfusion
Zeev Blumenfeld,* M.D., Robert W. Kuhn, Ph.D., and Robert B. Jaffe, M.D.
San Francisco, California
In previous studies, corticotropin-releasing factor was found to elicit a rise in circulating adrenocorticotropic
hormone in human subjects and laboratory animals, but no stimulatory effect of corticotropin-releasing factor on other pituitary hormones was detected. Since stress may be associated with luteinizing hormone
changes as well as with those of corticotropin-releasing factor and adrenocorticotropic hormone, we quantified gonadotropin responses to corticotropin-releasing factor and arginine vasopressin in 11 human fetal pituitaries with use of both superfusions and static incubations. Exposure to corticotropin-releasing factor brought about a significant increase in adrenocorticotropic hormone and gonadotropin concentrations in the effluent media by means of the superfusion system. Similar concentrations of corticotropin-releasing factor significantly increased adrenocorticotropic hormone secretion into the medium by dispersed fetal pituitary cells cultured on an extracellular matrix but failed to increase luteinizing hormone and follicle-stimulating hormone secretion. Exposure to 3 mmol/L 8-bromo-cyclic adenosine
monophosphate caused an increase in all three peptides, both in superfusion and static incubations. Doseresponse studies showed that the effect on gonadotropin secretion occurred at concentrations of 8-bromocyclic adenosine monophosphate two orders of magnitude lower than those affecting adrenocorticotropic hormone secretion. The purity of corticotropin-releasing factor and arginine vasopressin used in these studies was confirmed by high-performance liquid chromatography. These in vitro results are consistent with a paracrine effect of corticotropes acting on gonadotropes. The combination of static incubation and superfusion has proved useful in elucidating the effects of different secretagogues on pituitary cells. (AM J OSSTET GVNECOL 1986;154:606-12.)
Key words: Fetal pituitary, gonadotropin
From the Reproductive Endocrinology Center, Department of Obstetrics, Gynecology and Reproductive Sciences, University of California, San Francisco.
Supported in part by National Institutes of Health Grants HD 08478 and HD 11979.
Received for publication june 9,1985; revised November 7,1985; accepted November 25, 1985.
Reprint requests: Robert B. jaffe, M.D., Department of Obstetrics, Gynecology and Reproductive Sciences, Room M -1490, University of California, San Francisco, CA 94143.
*Recipient of American Physician Fellowship.
606
Shortly after the elucidation of the structure of corticotropin-releasing factor by Vale et aI., I there were several studies in which its effects on the secretion of adrenocorticotropic hormone, growth hormone, thyroid-stimulating hormone, luteinizing hormone, and prolactin in laboratory animals2
." and in normal humans4
,5 were assessed. Whereas corticotropin-releasing factor was found to elicit a rise in circulating adreno-