
PATRONES DE RECURRENCIA: VARIABILIDAD … · TRM: Transmembrana μg: Microgramos ... 5.2. ANÁLISIS...
88
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS FACULTAD DE CIENCIAS BIOLÓGICAS UNIDAD DE POSGRADO Patrones de recurrencia y resistencia asociadas a la variabilidad genética de Plasmodium vivax durante la malaria asintomática en la localidad de Mazán-Iquitos TESIS Para optar el Grado Académico de Magister en Biología Molecular AUTOR Edward Valencia Ayala Lima – Perú 2012
Transcript of PATRONES DE RECURRENCIA: VARIABILIDAD … · TRM: Transmembrana μg: Microgramos ... 5.2. ANÁLISIS...

UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS
FACULTAD DE CIENCIAS BIOLÓGICAS
UNIDAD DE POSGRADO
Patrones de recurrencia y resistencia asociadas a la
variabilidad genética de Plasmodium vivax durante la
malaria asintomática en la localidad de Mazán-Iquitos
TESIS
Para optar el Grado Académico de Magister en Biología Molecular
AUTOR
Edward Valencia Ayala
Lima – Perú
2012

LISTA DE ABREVIACIONES
aa: Aminoácidos
Alu I: Enzima de restricción Arthrobacter luteus
ATP: Adenosina de Tri-Fosfato
COI: Complejidad de la infección
CQR: Cloroquino-resistencia
Ct: Ciclo de threshold
DIRESA: Dirección Regional de Salud
DNA: Ácido desoxiribonucleotido
dNTPs: Desoxinucleótidos
EDTA: Ácido etilendiaminotetraacético
E/K: Cambio de Glutamina por Lisina
F/L: Cambio de Fenilalanina por Leucina
GG: Gota gruesa
Hd: Haplotipo diferente
Hha I: Enzima de restricción Haemophilus haemolyticus
Hhe: Hipnozoito heterólogo
Hho: Hipnozoito homólogo
Hi: Haplotipo idéntico
HRM: High resolution melting
IAP: Índice parasitario anual
IC50: Coeficiente de inhibición media
IPTG: Isopropiltio–β–D–galactósido
KD: Kilodaltons
LB: Medio Luria-Bertani
L/L: Sin cambio de aminoácido Leucina
OD: Densidad óptica
MDR1: Proteína multidrogo resistente
MgCl2: Cloruro de magnesio

MINSA: Ministerio de Salud
MSP3-α: Proteína se superficie del merozoito
M/T: Cambio de Metionina por Treonina
ng: Nanogramos
nM: Nanomolar
NBD: Nucleotide Binding Dominion
OMS: Organización Mundial de la Salud
ORF: Open Read Frame
pb: Pares de bases
pfmdr1: Gen mutidrogoresistente de Plasmodium falciparum
PLD: Nivel de la diversidad poblacional
pvald1: Gen aldolasa de Plasmodium vivax
pvmdr1: Gen mutidrogoresistente de Plasmodium vivax
pvmsp3-α: Gen de la proteína de superficie del merozoito
qPCR: Reacción en Cadena de la Polimerasa Cuantitativo
RENACE: Red Nacional de Control Epidemiológico
RFLP: Patrones de longitud de fragmentos de restricción
RNA: Ácido ribonucleico
RQ: Cuantificación Relativa
SE: Semana Epidemiológica
SNPs: Polimorfismos de nucleótidos únicos.
SVE: Servicio de Vigilancia Epidemiológica
Taq: Thermus acuaticus
TMDs: Dominios transmembrana
TRM: Transmembrana
μg: Microgramos
μl: Microlitros
μM: Micromolar
UV: Ultravioleta
X-Gal: 5–bromo–4–cloro–3–indolil–β–D–galactósido.
Y/F: Cambio de Tirosina por Fenilalanina

ÍNDICE
TÍTULO
AGRADECIMIENTO
DEDICATORIA
RESUMEN
ABSTRACT
I. INTRODUCCIÓN 1
II. ANTECEDENTES 3
2.1. GENERALIDADES 3
2.1.1. ETIOLOGÍA 3
2.1.2. CICLO BIOLÓGICO 4
2.1.3. EPIDEMIOLOGÍA 5
2.1.4. SINTOMATOLOGÍA 6
2.1.4.1. Malaria Asintomática 7
2.1.5. DIAGNÓSTICO 8
2.1.6. TRATAMIENTO 9
2.2. RECURRENCIA DE LA MALARIA POR P. vivax 10
2.2.1. RELAPSO O REINFECCIÓN 10
2.2.1.1. Análisis Polimórfico del gen pvmsp3-α 12
2.2.2. RESISTENCIA 13
2.2.2.1. Análisis Polimórfico del gen pvmdr1 14
III. OBJETIVOS 16
IV. HIPÓTESIS 16
V. MATERIALES Y MÉTODOS 17
5.1. POBLACIÓN DE ESTUDIO 17
5.2. ANÁLISIS DE DATOS 18
5.3. MÉTODOS DE INVESTIGACIÓN 19
5.3.1. EXTRACCIÓN DE DNA 19
5.3.2. DIAGNÓSTICO MOLECULAR DE Plasmodium 19
5.3.2.1. Nested PCR 19

5.3.2.1.1. Primera PCR 19
5.3.2.1.2. Segunda PCR 20
5.3.3. GENOTIPIFICACIÓN MOLECULAR DE P. vivax 20
5.3.3.1. Nested PCR 20
5.3.3.1.1. Primera PCR 21
5.3.3.1.2. Segunda PCR 21
5.3.3.2. Digestión enzimática 21
5.3.4. ANÁLISIS DE RESISTENCIA DE P. vivax 22
5.3.4.1. PCR Convencional 22
5.3.4.2. Secuenciamiento 23
5.3.4.3. Número de copias 24
5.3.4.4. Clonamiento 25
VI. RESULTADOS 28
6.1. DETECCIÓN DE LA MALARIA DURANTE LA VIGILANCIA ACTIVA 28
6.2. VARIABILIDAD POLIMÓRFICA DEL GEN pvmsp3-α 30
6.3. PATRONES DE RECURRENCIA 36
6.4. MECANISMOS DE RESISTENCIA 39
VII. DISCUSIÓN 47
7.1. DETECCIÓN DE LA MALARIA DURANTE LA VIGILANCIA ACTIVA 47
7.2. VARIABILIDAD POLIMÓRFICA DEL GEN pvmsp3-α 48
7.3. PATRONES DE RECURRENCIA 50
7.4. MECANISMOS DE RESISTENCIA 52
VIII. CONCLUSIONES 55
IX. RECOMENDACIONES 56
X. BIBLIOGRAFÍA 57
XI. ANEXOS 67

DEDICATORIA
A mis padres Saturnino Valencia y Juana Ayala, por ser el pilar fundamental en todo lo que
soy, porque creyeron en mí y porque me sacaron adelante, dándome ejemplos dignos de
superación y entrega, porque en gran parte gracias a ustedes, hoy puedo ver alcanzada mí
meta.
A mi esposa Miriam y a mi hijo Leonardo, quienes han sido mi inspiración y motivación
para los esfuerzos que he hecho en mi vida para superarme en mi formación personal y
quienes al final han soportado cada situación difícil que pasamos debido a esos esfuerzos.
Dios los bendiga por eso.
A mis hermanos, Carlos, Nikita, John y Engels, a mis sobrinos Alexandra, Ródrigo e Italo
por estar conmigo y apoyarme siempre, los quiero mucho. Y a todos aquellos que
participaron directa o indirectamente en la elaboración de esta tesis.
¡Gracias a ustedes!

AGRADECIMIENTO
Al Laboratorio de Enfermedades Infecciosas de la Universidad Peruana Cayetano Heredia
por haberme dado cobijo y por todo lo que aprendí en el, así mismo, por haberme dado su
voto de confianza y por todo el apoyo otorgado a mi persona.
Un agradecimiento especial al Dr. Robert Gilman, al Dr. Joe Vinetz, a la Dra. Manuela
Verastegui y al Dr. Holger Mayta, por su invaluable apoyo y confianza para la realización
de esta tesis. A mi asesora, la Dra. Maritza Calderón Sánchez por su paciencia, apoyo y
confianza en mí como persona y en mi trabajo. Gracias por sus consejos personales y
académicos. Gracias por escucharme.
A mis maestros de la Universidad Nacional Mayor de San Marcos, que ayudaron en mi
formación profesional. A mis jurados Mag. Ruth Garcia de la Guarda, Mag. Débora
Alvarado, Mag. Teresa Barreto, Dr. Abelardo Maturrano, gracias por todo su tiempo
invertido en la revisión de esta tesis.
A mis amigos y colegas: Edith Málaga, Roxana Zamudio, Nancy Chile, Susan Espetia,
Sonia Apaza, Leny Sánchez, Janeth Acosta, Juan Pacori, Jesus Pinto, Manuel Fasabi y Gary
Malpartida, por todo lo que vivimos y por su influencia en mi vida. Gracias por infundirme
sus ánimos y compartir conmigo sus conocimientos. También quiero agradecerles el
haberme hecho participe de su trabajo. Gracias de corazón. A mis amigos, porque sin ellos
no hubiera sido lo mismo. Gracias, eternamente gracias, a todos.

RESUMEN
Plasmodium vivax agente etiológico de la malaria, exhibe una gran variabilidad genética
durante episodios recurrentes de la enfermedad. Esta recurrencia es informada como de
baja prevalencia asociada con la malaria asintomática. Así mismo los episodios recurrentes
(reinfecciones o relapsos) a menudo pueden ser confundidos por resistencia a fármacos
como la cloroquina. Por lo tanto el objetivo principal de este estudio fue relacionar los
patrones de recurrencia y la resistencia con la variabilidad genética de P. vivax. En este
estudio se evaluaron las muestras secuenciales de individuos provenientes de una región
endémica del Perú (Mazán-Iquitos), diagnosticados previamente con malaria, por
microscopía, durante seguimientos activos y sometidos a un régimen de tratamiento
estándar con cloroquina. La genotipificación realizada en base al gen pvmsp3-α, utilizando
el Nested PCR y la digestión enzimática, permitió identificar una alta variabilidad genética
de P. vivax, a partir de la cual, se identificaron los patrones de recurrencia, establecidos
como relapsos, a partir de estadios latentes o hipnozoitos homólogos (con haplotipos
idénticos) y reinfecciones (con haplotipos diferentes). Los rangos de tiempo permitieron
una identificación más precisa, observándose mayores frecuencias de relapsos por
hipnozoitos homólogos antes de los 90 días post-primera evaluación y mayores frecuencias
de reinfecciones después de este periodo. Así mismo las recurrencias en el primer periodo
de tiempo, por haplotipos diferentes, pueden deberse también a hipnozoitos heterólogos.
Complementando el estudio, el análisis de secuenciamiento del gen pvmdr1, permitió
identificar SNPs, codificantes de mutaciones no sinónimas, relacionadas con resistencia a
cloroquina. Estos SNPs, a través del software U-Melt (análisis in sílico), presentaron
variaciones en las temperaturas de fusión. Finalmente los resultados de cuantificación
relativa con qPCR Real Time no mostraron diferencias significativas en el número de
copias del gen pvmdr1.
Palabras clave: Cloroquina, Genotipificación, Haplotipos, Hipnozoito, Malaria
Asintomática, Recurrencia, Variabilidad.

ABSTRACT
Plasmodium vivax etiologic agent of malaria has a large genetic variability during recurrent
episodes of the disease. This recurrence is reported as low prevalence associated with
asymptomatic malaria. Also recurrent episodes (reinfection or relapse) can often be
mistaken for drug resistance as chloroquine. Therefore the main objective of this study was
to correlate the patterns of recurrence and resistance to the genetic variability of P. vivax. In
this study, we evaluated the sequential samples of individuals from an endemic region of
Peru (Mazán-Iquitos), previously diagnosed with malaria microscopy during active follows
and subjected to a standard treatment regimen with chloroquine. Genotyping based on the
pvmsp3-α gene, using Nested PCR and enzymatic digestion, identified high genetic
variability of P. vivax, from which were identified recurrence patterns established as
relapse, from latent stages or homologous hypnozoites (with identical haplotypes) and
reinfections (with different haplotypes). The time ranges allow more accurate identification,
with higher frequency of relapses by homologous hypnozoites before 90 days post-first
evaluation and higher frequencies of reinfection after this period. Also recurrences in the
first period of time, for different haplotypes may also be due to heterologous hypnozoites.
Complementing the study, the sequencing analysis of the gene pvmdr1, identified SNPs,
encoding nonsynonymous mutations related to resistance to chloroquine. These SNPs,
through U-Melt software (in sílico analysis), showed variations in the melting temperatures.
Finally the results of relative cuantification with Real Time qPCR no showed significant
differences in copy number of the pvmdr1 gene.
Keywords: Chloroquine, Genotyping, Haplotypes, Hipnozoite, Recurrence, Asymptomatic
Malaria, Variability.

I. INTRODUCCIÓN
La malaria es una hemoparasitosis aguda, de evolución crónica y
recurrente, producida por protozoarios intracelulares obligados del género
Plasmodium y transmitida por mosquitos hembras del género Anopheles. Dentro
de las principales especies de parásitos tenemos a Plasmodium vivax, el cual
causa la malaria terciaria benigna y ocasiona entre 147 a 436 millones de
infecciones clínicas/año. A diferencia de Plasmodium falciparum, que causa la
malaria terciaria maligna, P. vivax está menos asociado con mortalidad aunque se
relaciona con una considerable morbilidad (Hay et al., 2004; Vargas et al., 2003).
Esta enfermedad parasitaria es un problema de salud global, ampliamente
distribuida en el mundo. Según la OMS, malaria es endémica en 108 países,
siendo el continente africano el más afectado con el 60% de los casos. En
Latinoamérica, Brasil es el país con la más alta prevalencia de malaria, seguido
por Colombia y Perú (WHO, 2005). En nuestro país, P. vivax es la especie
predominante, sin embargo, recientemente varios estudios muestran regiones
endémicas con una baja tasa de prevalencia en la transmisión, lo cual está
relacionado con el alto índice de formas asintomáticas de la enfermedad (Branch et
al., 2005).
Las infecciones asintomáticas son poco conocidas en estas regiones pero a
través de los sistemas de vigilancia activa, se ha demostrado persistencia de la
infección y constantes recurrencias de la parasitemia. Es de considerar además
que esta forma de la enfermedad, donde se ubican los portadores asintomáticos,
tiene un gran significado epidemiológico, pues aunque es invisible al ojo clínico,
contribuyen a mantener, difundir y perpetuar la infección. (Koram, 1993).
P. vivax se caracteriza por ocasionar recurrencias, principalmente por: a)
relapsos, debidos a estadios latentes del mismo parásito en el hígado, conocidos

como hipnozoitos; la parasitemia reaparece semanas después de la cura del
episodio inicial, b) reinfecciones, por la picadura infectante de otro mosquito.
Estas recurrencias originan adaptación inmunológica e incremento de la
inmunidad en los huéspedes humanos, evitando las complicaciones de la
enfermedad, por lo tanto se relacionan con las manifestaciones subclínicas y
asintomáticas (Loyola, 1991; Branch et al., 2005). Estas características propias del
huésped junto con la variabilidad genética de P. vivax son una limitante para el
tratamiento oportuno y efectivo de la enfermedad.
Sumado a esto, la variabilidad genética es un factor que influye
significativamente en el flujo de genes, como aquellos que confieren resistencia a
fármacos. En P. vivax la resistencia es otra forma de recurrencia de la parasitemia
(Baird et al., 2004). Al respecto, actualmente en el Perú la recurrencia por resistencia
está pobremente definida, entendiendo estos mecanismos solo desde el punto de
vista de las variaciones en el estado inmune de los pacientes.
Las medidas establecidas para el control de la malaria tienen como
necesidad primordial conocer las frecuencias reales de las reinfecciones, las
mismas que son confundidas a menudo por los relapsos y el incremento de la
resistencia a fármacos como la cloroquina. En regiones endémicas como el distrito
de Mazán, al sur de Iquitos, en el departamento de Loreto, no existen datos reales
sobre la malaria asintomática, se carece de información sobre la dinámica de las
reinfecciones, relapsos, resistencia a drogas y variabilidad genética de P. vivax;
teniendo como población más expuesta y en mayor riesgo a los individuos no
inmunes, las mujeres embarazadas y los niños.
Por lo tanto, este estudio tiene como principal objetivo: Relacionar los
patrones de recurrencia y la resistencia con la variabilidad genética de P. vivax,
estableciendo técnicas que nos permitan determinar la reinfección, el relapso o la
resistencia. De esta manera se planteó la interrogante sobre: ¿Cuál es la

frecuencia de estos patrones de infección, en individuos asintomáticos
provenientes de una región endémica? y apoyados en herramientas de
epidemiología molecular, como la genotipificación y secuenciamiento, es posible
identificar estas características en P. vivax.
Para lograr estos objetivos se tomaron en cuenta la utilización de varias
técnicas moleculares. Nested-PCR, utilizado para el diagnóstico, en base a la
amplificación del gen que codifica el RNA ribosomal 18S (SSUrRNA). PCR-RFLP,
para la genotipificación, en base al polimorfismo del gen pvmsp3-α que codifica la
proteína de superficie del merozoito (MSP3). Secuenciamiento, para identificar
SNPs en el gen pvmdr1, el cual es ortólogo al gen de resistencia pfmdr1 de P.
falciparum. Así mismo, el uso del software U-melt (análisis in sílico), para
establecer diferencias en las temperaturas de fusión de estos SNPs. Finalmente,
clonamiento y la utilización de técnicas como el qPCR Real Time, para establecer
el número de copias del gen de resistencia.
II. ANTECEDENTES
2.1. GENERALIDADES
2.1.1. ETIOLOGÍA
La malaria es una enfermedad parasitaria causada por protozoarios del
género Plasmodium. Más de 120 especies han sido descritas en mamíferos,
reptiles y aves; sin embargo clásicamente, solo 4 especies son las que causan
infección en humanos, dentro de estos tenemos a P. falciparum, P. vivax, P. ovale
y P. malariae; aunque recientemente P. knowlesi, el cual tiene a los monos

macacos como hospederos naturales, ha sido propuesto como el quinto parásito
causante de malaria en humanos. Estas 5 especies de Plasmodium difieren en
morfología, ciclo de vida y presentación clínica (Singh et al., 2004; White, 2008).
2.1.2. CICLO BIOLÓGICO
En el ciclo biológico de Plasmodium existen 2 huéspedes, uno definitivo (la
hembra del mosquito Anopheles, donde el Plasmodium se reproduce
sexualmente) y otro intermediario (el ser humano, donde se realiza la reproducción
asexual del parásito) (Manguin et al., 1995). El ciclo empieza con la picadura de un
huésped infectado por el mosquito Anopheles, en el cual se inicia la fase sexual o
esporogónica, con una duración de 8 a 16 días, donde los gametocitos ingeridos
maduran a gametos, se fusionan y dan lugar al cigoto, este es el único estadio
diploide en el ciclo de vida de Plasmodium (la meiosis ocurre a pocas horas de la
formación del cigoto, permaneciendo el parásito en las formas haploides durante
semanas o meses) (McKenzie et al., 2001). El cigoto se introduce en el epitelio
intestinal del insecto, donde se desarrollan miles de esporozoitos que migran a las
glándulas salivales, a partir de donde serán inoculadas en el torrente sanguíneo
de un nuevo hospedero, iniciándose la fase asexual o esquizogónica del parásito.
En la fase asexual se observan 2 estadios: 1) exoeritrocítico, los
esporozoitos pasan de la circulación sanguínea a las células hepáticas, donde se
forman esquizontes (esquizogenésis), dando lugar a la formación de merozoitos
que al quedar libres inician esta fase nuevamente, con una duración de 6 a 8 días,
este estadio termina cuando los merozoitos invaden los hematíes, 2) eritrocítico;
los merozoitos en el interior de los hematíes forman el trofozoito joven, luego
adulto, seguido del esquizonte, originándose nuevamente merozoitos que al
romper el hematíe repiten el ciclo y dependiendo de la especie, luego de 2 ó 3
generaciones, algunos merozoitos desarrollan los gametocitos que son la forma
infectante para el mosquito (Aramburu et al., 1999; Hay et al., 2004). Una característica

propia de P. vivax y P. ovale es que estos tienen una forma asexual latente,
conocido como hipnozoito, el cual persiste en el hígado y causa relapsos semanas
después de la eliminación de los estadios sanguíneos que causan la infección
aguda (Hanf et al., 2009).
2.1.3. EPIDEMIOLOGÍA
Según la Organización mundial de la salud (OMS), la malaria es
considerada una de las enfermedades tropicales más importantes, siendo
endémica en África sub-Sahariana, oeste de Asia, América, el Caribe, Nueva
Guinea, Vanuatu, Islas Salomón, India y Pakistán, donde vive la mitad de la
población mundial. A mediados del 2008 unos 3200 millones de personas vivían
en zonas con riesgo de transmisión, en 109 países y territorios (WHO, 2008a). Se
estima que anualmente ocurren entre 350 y 500 millones de casos clínicos y entre
1,5 a 2,7 millones de muertes, la mayoría causados por P. falciparum y P. vivax
(WHO, UNICEF, 2005). Siendo el continente africano el más afectado con alrededor
del 60% de estos casos y más de 80% de muertes (WHO, 2007).
Entre las 4 especies causantes de malaria, P. falciparum y P. vivax tienen la
mayor incidencia, considerando a P. falciparum como la más peligrosa, por su
letalidad, por la dispersión mundial de sus estirpes resistentes a las drogas anti-
maláricas y por su predominio en África. P. vivax es el segundo parásito causante
de malaria mas prevalente en el mundo, predominantemente en Asia, América
Central y Sudamérica (Ousmane et al., 2005). Actualmente P. vivax ocasiona 70-80
millones de infecciones/año, que es un poco más de la mitad de la casuística
global registrada fuera de África, con más del 50% de los casos registrados en
Latinoamérica (WHO, 2009). La habilidad de P. vivax de completar su ciclo
esporogónico a una temperatura mínima de 16°C, comparada con 21°C utilizado
por P. falciparum, ha contribuido a la capacidad de establecerse en distintas
regiones tropicales, subtropicales y templadas (Mendis et al., 2001).

En América, el 38,4% de la población vive en regiones ecológicas propicias
para la transmisión de malaria. En el año 2000, de los 1,14 millones de casos
notificados, 53,6% se presentaron en Brasil, seguido por 9,45% en Colombia,
8,65% en Ecuador, 6,12% en Perú, 4,68% en Guatemala, 3,08% en Honduras,
2,76% en Bolivia, 2,61% en Venezuela, 2,11% en Guyana y 1,15% en Surinam. A
partir del 2005, con el incremento de casos de malaria por P. vivax en la región
amazónica, el Perú ocupa el tercer lugar después de Brasil y Colombia (WHO,
2005).
En el Perú la malaria es endémica y caracterizada por ser cíclica o
estacional. El área de transmisión involucra el 75% del territorio nacional,
distribuido en escenarios epidemiológicos como la cuenca amazónica, valles
occidentales de la costa norte y valles interandinos hasta los 2300 m.s.n.m.
(Aramburu et al., 1999; DRE-SVE, 2007). En nuestro país, P. vivax es el más
importante parásito causante de malaria en términos de su morbilidad y a pesar de
la amplia prevalencia, estudios concernientes a su biología y a las manifestaciones
de la enfermedad son limitados, en parte, debido a que la mayor atención ha sido
dirigida a P. falciparum por ser el causante de la malaria maligna (Ayala et al., 2005;
Volman et al., 2005).
La mayor incidencia de malaria, en el Perú, se encuentra en el norte de la
región amazónica, donde se ubica el departamento de Loreto, siendo el epicentro
de esta enfermedad desde los años 1990. En el 2008, esta región registró el 55%
de los casos notificados, seguido por los departamentos de Madre de Dios, Piura,
Tumbes y Junín, que constituyen un segundo estrato en relación a la carga de
enfermedad (MINSA, 2009). En el 2009, esta región también registró el brote de
mayor magnitud con 25928 casos. En el 2011 a nivel nacional se notificó 23062
casos, 11779 procedieron de Loreto (51 % del total).
Actualmente DIRESA Loreto ha notificado hasta la semana epidemiológica
(SE) 29 del 2012, un total de 14463 casos, de los cuales 85,8% son por P. vivax y
14,2 % por P. falciparum. La provincia de Maynas, una de las siete que conforman
el departamento de Loreto, en el 2008 registró 30% de los casos del país. Esta

provincia, de gran extensión territorial, limita al norte con Colombia y al Oeste con
Ecuador, presenta distritos como Fernando Lores, San Juan Bautista y Mazán que
conforman un importante foco de malaria principalmente por P. vivax. Estas
localidades se ubican próximas a la ciudad de Iquitos, donde la dinámica de
transmisión guarda relación con los desplazamientos de la población (MINSA, 2008).
2.1.4. SINTOMATOLOGÍA
La sintomatología de la malaria consiste en un cuadro clínico clásico de
escalofríos, episodios febriles intensos y sudoración. Al terminar la sudoración el
paciente entra en un período asintomático, hasta el próximo acceso febril. Los
primeros síntomas son poco específicos y similares a los de una infección viral:
dolor de cabeza, debilidad, fatiga, mialgias y artralgias. En casos severos se
presentan coma, acidosis metabólica, anemia severa, hipoglucemia, falla renal,
falla pulmonar y esplenomegalia. En este estado, aun con tratamiento, la letalidad
puede llegar a ser de 15-20% y si no se trata, la malaria complicada es casi
siempre fatal. Sin embargo todos los signos y síntomas varían en función a la
especie de Plasmodium, la carga parasitaria y estado inmune del paciente (Mwangi
et al., 2005; WHO, 2006).
Todas las especies de Plasmodium tienen un componente hemolítico, por lo
que cada vez que una generación de merozoitos destruye el glóbulo rojo se
originan los síntomas. En el caso de P. falciparum, los parásitos se multiplican
rápidamente y pueden infectar más del 30% de los eritrocitos, causando altos
niveles de hemólisis e incremento las complicaciones de la enfermedad; este
parásito invade células de todas las edades mientras que P. vivax y P. ovale
prefieren células jóvenes y P. malariae busca células maduras (Carlton et al., 2003).
Las características únicas de P. vivax y P. ovale en la reactivación de los
hipnozoitos, llevan a la reaparición de nuevos episodios o relapsos, en estos
casos, los síntomas subclínicos como dolor de cabeza, dolor de espalda, nauseas
y malestar general pueden ser muy leves o estar ausentes (OPS, 2006).

2.1.4.1. Malaria Asintomática
La malaria asintomática está relacionada con las constantes infecciones y
los relapsos, que ocasionan incremento de la inmunidad adquirida, con la
consecuente disminución en las complicaciones de la enfermedad.
Frecuentemente en regiones endémicas, tras prolongados periodos de infección,
las personas expuestas tienen un alto grado de inmunidad y la malaria clínica es
poco frecuente, con lo cual se incrementa el número de portadores asintomáticos.
Además varios estudios en P. vivax han señalado que durante los relapsos,
la respuesta inmune es más rápida y amplificada, seguida de elevaciones
adicionales en los niveles de anticuerpos (Loyola, 1991; Branch et al., 2005).
La malaria puede comportarse como una infección asintomática sólo puede
detectarse mediante procedimientos de laboratorio), como una infección subclínica
(que supera el umbral clínico, pero no lo hace de forma típica), como una infección
sintomática o, incluso, como una infección hiperaguda fulminante, de rápida
evolución que produce la muerte. Las formas subclínicas y asintomáticas, donde
se ubican los portadores, tienen gran significado epidemiológico pues aunque son
invisibles al ojo clínico, contribuyen a mantener, difundir y perpetuar la infección
(Koram, 1993).
La frecuencia de malaria asintomática no es conocida en regiones de baja
transmisión. En Perú, los estudios se han limitado a diseños transversales, como
los efectuados en las comunidades al sur de Iquitos, donde, a partir de 998
individuos, se encontraron 13 con P. falciparum y 30 con P. vivax, todos
detectados por microscopía, de éstos, sólo 8 y 19 respectivamente, reportaron
solo fiebre. (Alves et al., 2002; Roshanravan et al., 2003; Branch et al., 2005).
2.1.5. DIAGNÓSTICO
El diagnóstico oportuno y preciso es crítico para el manejo efectivo de la
malaria. Las implicaciones del diagnóstico guardan relación con la diferente

morfología de las especies maláricas, la esquizogonia eritrocítica, la endemicidad,
los niveles de transmisión, la densidad parasitaria, la inmunidad, los signos y
síntomas clínicos, la resistencia a drogas, la malaria recurrente y la parasitemia
persistente, todo un conjunto complejo de elementos que influyen en la
interpretación del diagnóstico (Tangpukdee et al., 2009). La presentación clínica de la
malaria es variable y poco específica, por lo tanto es necesario incorporar técnicas
de laboratorio para su diagnóstico (Perkins y Bell, 2008).
La microscopía convencional, utilizando el examen de gota gruesa o frotis
sanguíneo, se considera el método idóneo para el diagnóstico y control de la
malaria, aún cuando puede presentar menor sensibilidad que las técnicas
moleculares, particularmente ante parasitemias bajas e infecciones mixtas (Gillet et
al., 2009; Tangpukdee et al., 2009). Su umbral de detección se ubica entre 4 y 20
parásitos/μl (Wongsrichanalai et al., 2007), aunque en condiciones de campo, un
umbral entre 50 y 100 parásitos/μl se considera más realista, con un número
recomendado de campos a ser leídos entre 100 y 400 (WHO, 1991).
Por otra parte diferentes estudios han demostrado que los métodos
moleculares detectan 8 veces más infecciones, por Plasmodium spp., que la
microscopía, representando las infecciones mixtas, hasta un tercio de ellas. Las
herramientas de diagnóstico molecular han modificado la interpretación de la
epidemiología malárica, al revelar grandes reservorios de infecciones
asintomáticas (Steenkeste et al., 2009). Además son consideradas muy importantes
por su utilidad para identificar especies cuando las densidades parasitarias son
muy bajas, en las infecciones mixtas o cuando las muestras se han deteriorado.
Esta es una técnica altamente específica y sensible, con un umbral de detección
entre 1 y 10 parásitos/ μl. Las técnicas moleculares también permiten detectar
parásitos resistentes a drogas y analizar el polimorfismo genético en estudios de
relapsos (Johnston et al., 2006; Kain et al., 1993).

2.1.6. TRATAMIENTO
La pauta antimalárica, para P. vivax, contempla el tratamiento de cura
radical recomendado por la OMS: Combinación de Cloroquina (CQ) a razón de 25
mg/kilo de peso corporal, distribuidos en 3 días y Primaquina (PQ) a razón de 0,25
mg/kilo de peso corporal, por día, durante 14 días. La primera actúa como
esquizonticida sanguíneo, elimina trofozoitos y esquizontes que son las formas
asexuales responsables de las manifestaciones clínicas y la segunda actúa como
esquizonticida tisular, elimina los parásitos en desarrollo o latentes (hipnozoitos)
en el hígado, para prevenir los relapsos (WHO, 2006). En el caso de relapsos, al
igual que en el episodio inicial, las pautas para el tratamiento son las mismas. En
caso de un segundo o tercer relapso deberá prescribirse la cloroquina en la misma
dosis y la primaquina al doble, dosificando a 0,5 mg/kg por día por 14 días ó 0,25
mg/kg por día durante 28 días (Álvarez et al., 2006).
2.2. RECURRENCIA DE LA MALARIA POR P. vivax
La reinfección, los relapsos y la resistencia a fármacos son el origen de las
recurrencias. Cuando una parasitemia reaparece después del tratamiento, puede
tratarse de: a) un relapso, por reactivación de estadios latentes en el hígado,
conocidos como hipnozoitos, b) una reinfección, por la picadura infectante de otro
mosquito, que en zonas endémicas es constante o c) una resistencia, originada
por estadios asexuales sanguíneos que sobrevivieron a la terapia y permanecieron
a niveles subpatentes (Baird et al., 2004; Orjuela et al., 2009a).
Diversos ensayos clínicos con drogas antimaláricas estiman alarmantes
rangos de recurrencia de P. vivax, los cuales pueden llegar de 26% a 40 % en 180
días de seguimiento post-tratamiento. Algunos estudios en regiones de la
amazonía rural reportan recurrencias de la parasitemia por haplotipos idénticos de
P. vivax (consistentes con reactivación de hipnozoitos homólogos) y por haplotipos

relacionados pero diferentes, sugiriendo la necesidad de caracterizar un número
mayor de muestras con infección primaria y recurrente para determinar
exactamente la asociación genotípica (Mallika et al., 2007; Baird et al., 2008; Orjuela et
al., 2009b).
2.2.1. RELAPSO O REINFECCIÓN
El manejo y control de P. vivax es complicado por su habilidad de causar
relapsos y por las continuas reinfecciones. Los relapsos presentan variaciones
notables de acuerdo a las diferentes regiones geográficas y climáticas, así las
cepas originarias de regiones tropicales están caracterizadas por una infección
primaria temprana, seguido por un corto periodo latente, de 5 a 10 semanas, antes
de la aparición de frecuentes relapsos durante el siguiente año. En contraste
cepas de regiones templadas están caracterizadas por un periodo variable antes
de la infección primaria, seguido por un largo periodo latente, de 5 a 10 meses,
antes del comienzo de los relapsos. Claros ejemplos son los estudios realizados
en Guyana Francesa, donde se mostraron picos de relapsos en los primeros 3
meses después del episodio inicial, posterior a este periodo la mayoría de
episodios fueron principalmente debidos a reinfecciones. Estas observaciones
mostraron un patrón de cepa tropical, con un corto periodo latente para el inicio de
los relapsos. (Alyson et al., 1996; Hanf et al., 2009).
En varios estudios registran frecuencias para los relapsos que varían desde
11% en la India hasta 51% en Afganistán, con valores intermedios de 30% en
Indonesia (WHO, 2006). En Colombia, un estudio realizado en el año 2004 mostró
que el tiempo en que se presentan la recurrencias (relapsos), en pacientes
sometidos a diferentes esquemas de tratamiento, fluctúan entre 53 a 129 días
después del episodio inicial (Álvarez et al., 2006).
Diferenciar relapso y reinfección en regiones endémicas resulta complicado,
la mayoría de estudios usan técnicas moleculares como la genotipificación para

hacer una distinción entre los tipos de recurrencias, diferenciando los relapsos
(con el mismo genotipo como en la infección inicial) de las reinfecciones (con un
genotipo diferente) (Orjuela et al., 2009b), cabe señalar además que los relapsos
pueden originarse tanto desde la reactivación del mismo clon del parásito,
encontrado en la infección inicial (hipnozoito homólogo), como de otro clon
genéticamente diferente (hipnozoito heterólogo). Otros estudios estiman el relapso
usando rangos de tiempo según el tipo de cepa o basado en la frecuencia de
casos de P. vivax durante la estación de no transmisión. De forma más precisa,
algunos proponen una simple clasificación para la fácil diferenciación entre
relapsos y reinfecciones, se plantean rangos de tiempo a escala individual,
identificando infecciones antes de 90 días post-primer episodio, con una alta
probabilidad que el ataque secundario sea un relapso y después de los 90 días es
más probable que el ataque secundario sea por una reinfección (Chen et al., 2007;
Veron et al., 2009).
2.2.1.1. Análisis Polimórfico del gen pvmsp3-α
La epidemiología molecular contribuye a diferenciar entre relapsos y
reinfecciones, mediante la utilización de técnicas moleculares. El marcador
pvmsp3-α, que codifica para la proteína de superficie del merozoito (MSP3-α), ha
sido extensamente estudiado (Cui et al., 2003). Este gen de copia única
comúnmente utilizado para la genotipificación de P. vivax, codifica una región
altamente polimórfica con un dominio central rico en Alanina que contiene una
región interna compuesta por motivos repetitivos de longitud variable (PlasmoDB
gene ID: PVX_097720). La proteína codificada por este gen, tiene un peso
molecular entre 148 a 150 KD, se encuentra en el estadio eritrocítico del ciclo de
vida del parásito y es considerada como un potencial blanco de vacunas. (Cui et al.,
2003; Mascorro et al., 2005; Imwong et al., 2005).
En países como Irán, Papúa Nueva Guinea y Colombia el polimorfismo del
gen pvmsp3-α ha sido ampliamente estudiado, mediante la utilización de técnicas
como el Nested-PCR y digestión enzimática o RFLP, con enzimas como Alu I y

Hha I (Zakeri et al., 2006; Cole-Tobian et al., 2005; Cristiano et al., 2008). A su vez en
Venezuela y Tailandia se han llevado a cabo técnicas de secuenciamiento génico,
para determinar más extensamente el polimorfismo de este gen (Ord et al., 2005; Cui
et al., 2003).
Tres alelos principales han sido reportados después de realizar el Nested-
PCR, los cuales son caracterizados como alelo A = 1800 a 1900 pb, B = 1500 pb y
C = 1100 a 1200 pb. La diversidad puede ser mayor dentro de estos alelos, si se
utiliza la digestión enzimática combinando las enzimas Hha I y Alu I. La digestión
nos permite obtener patrones de corte enzimático, a partir del producto de
amplificación, que pueden ser compilados y usados para la genotipificación de P.
vivax. Las técnicas tanto de Nested-PCR como de RFLP permiten identificar no
solo haplotipos monoclonales sino también policlonales y las frecuencias de estos
haplotipos permiten definir el nivel de la diversidad poblacional (PLD) y la
complejidad de la infección (COI) (Bruce et al. 1999, Ord et al.,. 2005, Sutton et al., 2009).
Gen polimórfico pvmsp3-α de P. vivax
Cui et al., 2003
2.2.2. RESISTENCIA
La recurrencia, debida a la incompleta eliminación de estadios eritrocíticos
del parásito, ocasionada por el inadecuado tratamiento terapéutico, también puede
ser producida por mecanismos de resistencia. Aunque la resistencia podría ser
común después de un tratamiento inadecuado, también es necesaria la presencia

de parásitos resistentes, con variaciones genéticas en su genoma (Cogswell et al.,
1992). En el control de la malaria por P. vivax, las medidas aplicadas son
confundidas a menudo por factores como el surgimiento de resistencia a drogas,
principalmente a cloroquina. El primer caso de resistencia de P. vivax a cloroquina
fue reportado en 1989 desde Papúa Nueva Guinea, donde la monoterapia con
este fármaco es ahora virtualmente inefectiva. Más recientemente casos
esporádicos han sido reportados desde Myanmar, Sudamérica, Vietnam, y
Turquía (Hay et al., 2004; Price et al., 2007).
Reportes desde Sudamérica también sugieren la presencia de casos
aislados de P. vivax resistente a cloroquina, principalmente en regiones
endémicas como la ciudad de Manaus, en Brasil, donde en el 2000 se identificaron
casos de infección con P. vivax resistente al tratamiento con cloroquina (Alecrim,
2000). En Perú a partir de 1998, debido al peligro de propagación de la resistencia
a cloroquina, el Ministerio de Salud empezó una serie de ensayos de eficacia
terapéutica de este fármaco (Garg et al., 1995; Dua et al., 1996). Sin embargo a pesar
de los reportes clínicos, la prevalencia global de resistencia por P. vivax
permanece pobremente definida, principalmente debido a las variaciones de las
condiciones inmunológicas de cada paciente, por las continuas reinfecciones y por
los relapsos (Nomura et al., 2001; Tjitra et al., 2002; Ratcliff et al., 2007).
2.2.2.1. Análisis Polimórfico del gen pvmdr1
Estudios sobre el perfil de susceptibilidad a drogas han permitido identificar
proteínas transportadoras de membrana, como el MDR1, asociadas con
resistencia a cloroquina (CQR) principalmente en P. falciparum. Así mismo el
secuenciamiento permitió identificar proteínas ortólogas en P. vivax, codificadas
por el gen pvmdr1 (Baird et al., 2004; Babiker et al., 2001).
El gen pvmdr1 esta caracterizado por un único ORF de 4392 bp que
codifica una proteína de 1464 aa. Para determinar la resistencia a drogas fueron
tomadas en consideración 2 regiones de este gen: La primera abarca la posición

homóloga a los sitios polimórficos 86 y 184, la segunda abarca la posición
homóloga a los sitios polimórficos 1034 y 1042 de P. falciparum, que están
relacionadas con el fenotipo CQR.
El análisis de secuenciamiento permitió identificar polimorfismo de
nucleótidos únicos o SNPs en los codones 976 y 1076, localizados en los
segmentos hidrofóbicos X y XI de la proteína MDR. SNPs en estos codones
originan mutaciones no sinónimas como el cambio de aminoácidos Y/F y F/L,
respectivamente. Estos SNPs fueron asociados con CQR tanto in vitro como in
vivo y fueron reportados esporádicamente en pacientes provenientes de las
regiones amazónicas tanto del Perú como de Brasil (Sattabongkot et al., 2004; Fidock
et al., 2000; Brega et al., 2005; Suwanarusk et al., 2007).
Proteína TRM codificada por el gen pvmdr1 de P. vivax
Orjuela et al., 2009a
El análisis completo de la secuencia nucleotídica del gen pvmdr1 demostró
que contiene 24 SNPs y una única secuencia microsatélite conservada.
Notablemente 17 (73%) de los SNPs fueron no-sinónimos; sin embargo a pesar de
esta alta frecuencia de SNPs, ninguno fue encontrado en los motivos conservados
ABC. El motivo ABC es una secuencia aminoacídica corta, presente en el sitio de
fijación a nucleótidos (NBD), el cual junto con los motivos Walker, es responsable
de la hidrólisis de ATP (Brega et al., 2005). El SNP en el codón Y976F fue propuesto

como un marcador temprano de CQR y en estudios de susceptibilidad in vitro fue
encontrado en un 96.1% (123/128) de aislados provenientes de Indonesia
comparados a un 25% (17/69) de aislados provenientes de Tailandia. En
Indonesia, la media geométrica IC50 de susceptibilidad a cloroquina en aislados
con mutación Y976F fue de 283 nM, significativamente mayor que en los aislados
con el genotipo wild type con 44,5 nM (Suwanarusk et al., 2007; Orjuela et al., 2009a).
Por otro lado la CQR también está relacionada con la variación en el
número de copias de genes que codifican proteínas de resistencia como el MDR1.
Estudios efectuados en cepas provenientes de África principalmente en P.
falciparum se basan en técnicas de qPCR Real Time para determinar estas
variaciones (Ferreira et al., 2006). Así mismo los plásmidos clonados (fragmentos de
pvmdr1 y pvald1) utilizados como calibradores permitieron determinar el número
de copias en distintos aislados provenientes de Indonesia y Tailandia (Suwanarusk
et al., 2007).

III. OBJETIVOS
Objetivo general
Relacionar los patrones de recurrencia y la resistencia con la variabilidad
genética de Plasmodium vivax, en individuos con infección asintomática
provenientes de Mazán, región endémica del Perú.
Objetivos específicos
Detectar infecciones recurrentes por P. vivax, en individuos asintomáticos.
Identificar la variabilidad genética de P. vivax, en base al gen pvmsp3-α, en
individuos con infección recurrente.
Identificar los patrones de recurrencia asociados a la variabilidad genética
de P. vivax.
Identificar mutaciones no sinónimas e incremento en el número de copias
del gen pvmdr1, relacionados con resistencia de P. vivax a la cloroquina.
IV. HIPÓTESIS
Los patrones de recurrencia y la resistencia de P. vivax, en individuos con
infección asintomática, están relacionados con la variabilidad genética del
parásito, identificada por el polimorfismo del gen pvmsp3-α y por la
presencia de mutaciones no sinónimas e incremento en el número de
copias del gen pvmdr1.

V. MATERIALES Y MÉTODOS
5.1. POBLACIÓN DE ESTUDIO
El presente estudio se realizó con muestras colectadas, entre Junio del
2006 y Noviembre del 2008, en el distrito de Mazán ubicado en la parte Nororiental
del Perú a 50 km de la ciudad de Iquitos, capital del departamento de Loreto.
Presenta un clima tropical, cálido, húmedo y lluvioso, con una temperatura
promedio de 28ºC y es considerada una zona de alto riesgo para la transmisión de
P. vivax. (Sawyer, 1993).
La población total de individuos, supuestamente sanos, captados para este
estudio fue de 496. La normalización estadística de datos permitió obtener una
muestra significativa de 222 individuos. Para ello se tomaron en cuenta niveles de
confianza al 95% (Zα2 = 1.962), con proporción esperada del 50% (p=0.5; q=1-p) y
precisión estadística en 5% (d=0.05). A partir de este grupo se obtuvieron 444
muestras, colectadas mediante el sistema de vigilancia activa y evaluadas dentro
del contexto de un estudio de eficacia de cloroquina in vivo para el tratamiento de
la malaria por P. vivax.
Los criterios de inclusión fueron: Individuos comprendidos entre 0 a 75 años
de edad, firma de consentimiento informado, residentes en la zona endémica, con
2 muestras secuenciales, previamente diagnosticados por microscopía y
evaluados durante un año de seguimiento activo. Criterios de exclusión fueron:
Individuos con tratamiento antimalárico previo al diagnóstico y con
manifestaciones clínicas o subclínicas de la enfermedad.
Todas las muestras colectadas en tubos de EDTA y en un volumen de 5 ml
de sangre total (punción venosa) fueron trasladadas en cajas provistas con hielo
seco y enviadas al área de Biología Molecular en el Laboratorio de Investigación y

Desarrollo de la Universidad Peruana Cayetano Heredia en Lima–Perú. Este
estudio forma parte del proyecto: “Human reservoirs of Plasmodium vivax
transmission in the Peruvian Amazon”, aprobado por el comité de ética de la
universidad en mención (código SIDISI: 50457). En colaboración con la
Universidad Johns Hopkins en USA, el Instituto de Medicina Tropical “Alexander
Von Humbolt”, la Asociación Benéfica “PRISMA” y el Ministerio de Salud a través
de la Dirección Regional de Salud de Loreto.
5.2. ANÁLISIS DE DATOS
La variabilidad genética de P. vivax fue evaluada mediante la determinación
del nivel de la diversidad poblacional (PLD), definido como la variación genética de
un gen, en un loci antigénico, variable o neutral. Una medida del PLD es la
heterocigosidad esperada (He = 1/(1 − N) (1 – Σp2i) (Sutton et al., 2009), con p igual
a la frecuencia del alelo y N igual al número de alelos detectados, los valores del
He oscilan entre 0 a 1, considerando una alta variabilidad genética cuando los
valores son cercanos a la unidad. La complejidad de la infección (COI) es usada
también para evaluar la variabilidad genética, identificando el mínimo número de
alelos presentes en un loci polimórfico de copia única. Un COI > 1 puede ser
determinado por la observación de múltiples alelos dentro de una única muestra.
Para medir la significancia de las frecuencias de haplotipos encontrados se
utilizó la prueba de Fisher (F). En esta prueba el valor de la razón estadística (F
prueba) fue comparada con el valor de la razón tabular (F tabla). Considerando un
nivel de significancia del 95% (1-α = 0,95) y valorando las frecuencias observadas
como similares (F prueba<F tabla) o variables (F prueba > F tabla). Para el
análisis de relación se utilizó el Chi-cuadrado (X2). Considerando una relación
significativa con una probabilidad del 95%, cuando p < 0,05 (Tabla X2) y altamente
significativa con una probabilidad del 99%, cuando p < 0,01 (Tabla X2) con grados
de libertad mayores a cero (GL ≥ 1).

5.3. MÉTODOS DE INVESTIGACIÓN
El diseño de la investigación planteada es de tipo Experimental.
5.3.1. EXTRACCIÓN DE DNA
Se realizó la extracción de DNA a partir de las muestras de sangre en
EDTA, mediante el protocolo del Kit comercial QIAamp DNA Blod (Qiagen
Sciences, Maryland-USA). El volumen utilizado fue de 200 μl, a partir de lo cual se
obtuvo una concentración entre 20 a 50 ng de DNA, el cual fue conservado
inmediatamente a una temperatura de -20ºC, posteriormente utilizado tanto para el
diagnóstico como para la genotipificación molecular.
5.3.2. DIAGNÓSTICO MOLECULAR DE Plasmodium
5.3.2.1. Nested PCR
La detección de las especies de Plasmodium se realizó mediante el Nested-
PCR, el cual consta de 2 PCR consecutivos, realizados con la finalidad de
aumentar tanto la especificidad como la sensibilidad del análisis. Los
oligonucleótidos y condiciones de reacción y amplificación fueron adaptadas de
Roshanravan et al., 2003.
5.3.2.1.1. Primera PCR: Se amplificó un segmento del gen
que codifica el RNA ribosomal 18S (SSUrRNA) de 1200 pb, específico del género
Plasmodium. Los oligonucleótidos fueron: Plagen1= 5´-
CTTGTTGTTGCCTTAAACTTC-3´ y Plagen2= 5´-
TTAAAATTGTTGCAGTTAAAACG-3´.

La reacción de amplificación se realizó en un volumen final de 15 μl
conteniendo: buffer 1X (10 mM Tris-HCl pH 8,4, 50 mM KCl), 2 mM de MgCl2, 0,2
mM de dNTPs, 0,25 μM de cada oligonucleótido, 0,032 U/μl Taq DNA polimerasa
(Promega) y 50 ng de DNA. Las condiciones fueron: 95ºC por 3 min.; 19 ciclos
cada uno de 94°C por 30 seg., 58°C por 30 seg., 72°C por 1 min.30seg. y 1 ciclo
de extensión final de 72°C por 7 min.
5.3.2.1.2. Segunda PCR: Se amplificó una región interna
específica para P. vivax y para P. falciparum. Los oligonucleótidos fueron: Para P.
vivax: Viv1= 5´-CGCTTCTAGCTTAATCCACATAACTGATAC-3´ y Viv2= 5´-
ACTTCCAAGCCGAAGCAAAGAAAGTCCTTA-3´, que amplificaron 120 pb. Para
P. falciparum: Fal1= 5´-TTAAACTGGTTTGGGAAAACCAAATATATT-3´ y Fal2=
5´-ACACAATGAACTCAATCATGACTACCCGTC-3´, que amplificaron 205 pb.
La reacción de amplificación se realizó en un volumen final de 15 μl
conteniendo: buffer 1X (10 mM Tris-HCl pH 8,4, 50 mM KCl), 2 mM de MgCl2, 0,2
mM de dNTPs, 0,5 μM de cada oligonucleótido, 0,032 U/μl Taq DNA polimerasa
(Promega) y 1 μl de DNA obtenido de la primera amplificación. Las condiciones
fueron: 95ºC por 2 min, 34 ciclos cada uno de 94ºC por 30 seg., 60ºC por 1
min.30seg., 72ºC por 1 min. 30 seg., y finalmente 1 ciclo de extensión de 72ºC por
7 min.
Las reacciones de PCR fueron realizadas en un termociclador PT-100 MJ
Research. Los productos de PCR fueron separados por electroforesis en geles de
agarosa al 2% (Invitrogen, Life technologies) conteniendo 0,05 mg de bromuro de
etidio (Sigma Chemical CO.) y visualizados bajo iluminación UV. Los
oligonucleótidos seleccionados fueron evaluados con: BLASTn
(http://www.ncbi.nlm.nih.gov/BLAST/), CLUSTAL X (Gibson et al., 1994), PRIMER
PREMIER (htpp://www.premierbiosoft.com/primer design/index.html).

5.3.3. GENOTIPIFICACIÓN MOLECULAR DE P. vivax
5.3.3.1. Nested PCR
Para la amplificación de la región polimórfica del gen pvmsp3-α se realizó
un Nested PCR. Los oligonucleótidos y condiciones de reacción y amplificación
fueron adaptadas de Bruce et al., 1999 y Cui et al., 2003.
5.3.3.1.1. Primera PCR: Los oligonucleótidos utilizados
fueron: P1V1= 5´-CAGCAGACACCATTTAAGG-3´ y P2V1= 5´-
CCGTTTGTTGATTAGTTGC-3´. La reacción de amplificación se realizó en un
volumen final de 15 μl, conteniendo: buffer 1X (10 mM Tris-HCl pH 8,4, 50 mM
KCl), 2 mM de MgCl2, 0,15 mM de dNTPs, 0,15 μM de cada oligonucleótido, 0,013
U/μl. Taq DNA polimerasa (Promega) y 50 ng de DNA. Las condiciones fueron:
94ºC por 3 min., 34 ciclos cada uno de 94ºC por 30 seg., 56ºC por 30 seg., 68ºC
por 2 min. 30seg. y 1 ciclo de extensión final de 72ºC por 4 min.
5.3.3.1.2. Segunda PCR: Los oligonucleótidos utilizados
fueron: N1V1= 5´-GACCAGTGTGATACCATTAACC-3´ y N2V1= 5´-
ATACTGGTTCTTCGTCTTCAGG-3´. Estas secuencias amplificaron fragmentos
de 1900 pb y 1500 pb, correspondientes a los alelos A y B respectivamente. Las
reacciones de amplificación fueron similares a las usadas para la primera PCR,
agregando 1 μl de DNA a cada reacción, obtenido de la primera amplificación. Las
condiciones fueron de: 94ºC por 3 min., 29 ciclos cada uno de 94ºC por 30 seg.,
57ºC por 30 seg., 68ºC por 2 min. 30seg. y 1 ciclo de extensión final de 68ºC por 4
min.
Las reacciones de PCR y el análisis de los productos de PCR en geles de
agarosa al 1% se realizaron tal como se describe en la sección 5.3.2.1.2. Los
tamaños de los fragmentos fueron determinados usando un marcador de peso

molecular de 1 Kb (Invitrogen, Life technologies). Todas las muestras fueron
analizadas por duplicado para confirmar la reproducibilidad de los resultados.
.
5.3.3.2. Digestión enzimática
Para determinar el polimorfismo de la longitud de los fragmentos de
restricción (RFLP), que permite establecer los diferentes haplotipos, se utilizaron 2
enzimas de digestión Alu I y Hha I (20000 U/ml. BioLabs Inc. UK) usadas para la
genotipificación de P. vivax (Kosek et al., 2012). Cien nanogramos de DNA
amplificado, proveniente de la segunda PCR, fueron digeridos con: 1X NEBuffer 4
(50 mM de acetato de potasio, 20 mM Tris-acetato, 10 mM Acetato de magnesio, 1
mM ditiotreitol pH 7,9), 0,5U de Alu I y 0,5U de Hha I en un volumen final de 16 μl.
La incubación de la reacción se realizó a 37ºC por 3 horas, posteriormente las
enzimas fueron inactivadas a 65°C por 10 min.
Los fragmentos de DNA obtenidos de la digestión enzimática fueron
separados por electroforesis en geles de agarosa al 2%, tal como se describe en
la sección 5.3.2.1.2. Los tamaños de los fragmentos fueron determinados usando
un marcador de peso molecular de 100 bp (Invitrogen, Life technologies). Los
patrones enzimáticos fueron establecidos en base al número y al tamaño de los
fragmentos producidos con cada enzima, designados como haplotipos PA para
Alu I y PH para Hha I. Las infecciones policlonales o mixtas fueron identificadas
usando 2 criterios: Por el Nested PCR, con la presencia de 2 o más secuencias
alélicas del producto no digerido (Alelos A ó B) y por el RFLP comparando la suma
de los tamaños de los fragmentos del producto digerido con el tamaño del
producto no digerido.

5.3.4. ANÁLISIS DE RESISTENCIA DE P. vivax
5.3.4.1. PCR Convencional
Para identificar la resistencia de P. vivax se realizó inicialmente una PCR
convencional, para amplificar el gen pvmdr1. Posteriormente los productos de
amplificación previamente purificados fueron secuenciados. Los oligonucleótidos y
condiciones de reacción y amplificación fueron adaptadas de Brega et al., 2005.
Los oligonucleótidos utilizados fueron los siguientes: MdrF800= 5´-
ATAGTCATGCCCCAGGATTG-3’ y MdrR800= 5’-
ACGTTTGGTCTGGACAAGTATC-3’, cuyo producto de amplificación fue de 800
pb. La reacción de amplificación se realizó en un volumen final de 20 μl,
conteniendo: buffer 1X (10 mM Tris-HCl (pH 8,4), 50 mM KCl), 2,5 mM de MgCl2,
0,25 mM de dNTPs, 0,5 μM de cada oligonucleótido, 0,03 U/μl. Taq DNA
polimerasa (Promega) y 50 ng de DNA. Las condiciones fueron: 94ºC por 2 min.,
34 ciclos cada uno de 94ºC por 1 min., 62ºC por 1 min., 72°C por 1 min. y 1 ciclo
de extensión final de 72ºC por 5 min.
5.3.4.2. Secuenciamiento
Los productos de amplificación del gen pvmdr1 (sección 5.3.4.1.) fueron
enviados a los laboratorios de la corporación Macrogen en Maryland-USA para su
secuenciamiento, mediante el secuenciador BigDye Terminator 3.1 (Applied
Biosystems). Se envió a secuenciar la región que comprende los SNPs en los
codones Y976F y F1076L del gen pvmdr1, usando los mismos oligonucleótidos
mencionados en la sección anterior.
Estas secuencias fueron comparadas con las secuencias del gen pvmdr1
de la cepa Sal1 de P. vivax (GenBank Acc. No. AY618622) sensible a cloroquina y
proveniente del Salvador, usada como cepa de referencia en este estudio. Las

secuencias obtenidas fueron alineadas y analizadas usando el programa
ClustalW2 disponible libremente en la página web:
http://www.ebi.ac.uk/Tools/msa/clustalw2/, además para investigar la relación
filogenética entre estas secuencias provenientes de diferentes muestras, se utilizó
el programa Clustal C. La presencia de SNPs en estos codones relacionados con
resistencia a cloroquina fue confirmado, identificando los cambios de aminoácidos
en la secuencia proteica a través de un proceso de transducción in sílico,
mediante el software Translate a DNA Sequence disponible libremente en la
página web: http://arbl.cvmbs.colostate.edu/molkit/translate/.
Adicionalmente las secuencias del gen pvmdr1, conteniendo los SNPs en
los codones Y976F y F1076L, fueron analizados in sílico para determinar las
temperaturas de fusión en alta resolución o HRM mediante el software U-Melt
disponible libremente en la página web: http://www.dna.utah.edu/umelt/um.php, el
cual proporciona datos para una fácil discriminación entre los genotipos wild-type y
los genotipos mutantes.
5.3.4.3. Número de copias
Para determinar el número de copias del gen pvmdr1, se utilizó el método
de cuantificación relativa mediante qPCR en tiempo real con fluoróforo SYBR
Green I, mediante el sistema de ∆∆Ct comparativo en el equipo Step One Plus
Real Time 7700. En este sistema de cuantificación el gen de copia única, Aldolasa
(pvald1) codificado por P. vivax (GenBank Acc. No. AF247063), es usado como
gen endógeno (normalizador) para estimar el número de copias del gen pvmdr1
(problema) Las secuencias, así como los oligonucleótidos y las condiciones de
reacción y amplificación fueron adaptadas de Suwanarusk et al., 2007.
Los oligonucleótidos utilizados para amplificar el gen pvmdr1 fueron los
siguientes: mdr1F= 5´-CTGATACAAGTGAGGAAGAACTACG-3´ y mdr1R= 5´-
GTCCACCTGACAACTTAGATGC-3´, cuyo producto de amplificación fue de 151

pb y para amplificar el gen pvald1 se utilizaron: AldF= 5´-
GACAGTGCCACCATCCTTACC-3´ y AldR= 5´-
CCTTCTCAACATTCTCCTTCTTTCC-3´, cuyo producto de amplificación fue de
191 pb.
La reacción de amplificación se realizó en un volumen final de 20 μl,
conteniendo: Master Mix SYBR Green 1X (Invitrogen, Life technologies) y 0,1 μM
de cada oligonucleótido. Las condiciones fueron: 95ºC por 10 min., seguido de 40
ciclos cada uno de 95ºC por 30 seg., 60ºC por 1 min., y 72ºC por 30 seg., los
cambios de fluorescencia se midieron en el canal F1 del instrumento StepOne
Plus Real Time 7700, en el final de la fase de extensión de cada ciclo. Los
ensayos fueron optimizados con una concentración de DNA a partir de 50 ng/μl.
El sistema de cuantificación relativa en tiempo real, requiere la utilización de
fragmentos clonados del gen pvald1 y del gen pvmdr1 de P. vivax. Estos
fragmentos fueron utilizados como calibradores y controles de amplificación en
cada experimento. El software para la cuantificación relativa se basa en la
utilización del sistema de ciclo del threshold (Ct) comparativo dada por la fórmula:
N= 2∆∆Ct, donde ∆∆Ct = (Ct pvald1-Ct pvmdr1)-(Ct pvald1cal-Ctpvmdr1cal). El Ct
pvald1 y Ct pvmdr1 son valores de Ct para el gen pvald1 y pvmdr1
respectivamente, mientras que el Ctcal es una diferencia promedio entre Ct pvald1
y Ct pvmdr1 obtenida de los calibradores conteniendo una única copia de los
fragmentos clonados.
5.3.4.4. Clonamiento
Los calibradores (fragmentos de los genes pvald1 de 191 pb y pvmdr1 de
151 pb) fueron clonados, siguiendo los siguientes procedimientos:

PCR Convencional y ligación: Para el PCR la reacción de amplificación
se realizó en un volumen final de 20 μl, conteniendo: buffer 1X (10 mM Tris-HCl
pH 8,4, 50 mM KCl), 2,5 mM de MgCl2, 0,25 mM de dNTPs, 0,075 μM de cada
oligonucleótido, 0,03 U/μl. Taq DNA polimerasa (Promega) y 50 ng de DNA. Las
condiciones de reacción fueron: 94ºC por 1 min., 34 ciclos cada uno de 94ºC por 1
min., 54ºC por 1 min., 72°C por 1 min. y 1 ciclo de extensión final de 72ºC por 5
min. Los productos de PCR fueron purificados utilizando el kit QIAquick PCR
Purification a través de la columna de Spin DNAquick (Qiagen).
El proceso de ligación de los fragmentos de los genes pvald1 y pvmdr1
(insertos), amplificados previamente, se realizó con el vector pGEM-T easy
(Promega). Este vector es lineal y presenta bases de Timina (T) en los extremos
3´ terminal, que se unen por complementariedad con los extremos poliA 5´
terminal de los productos de amplificación; esta característica facilita la ligación y
evita la recirculización del vector. La ligación se realizó en un volumen final de 10
μl, conteniendo: buffer 1X (60 mM Tris-HCl pH 7,8, 20 mM MgCl2, 2mM ATP),
pGEM-T easy 5 ng/μl, T4 DNA ligasa 10U y una relación de inserto con vector de
3:1.
Preparación de células competentes y transformación: La preparación
de células competentes se inicio con el cultivo de Escherichia coli (E. coli - cepa
NovaBlue) en agar/LB (Difco) e incubación a 37°C durante toda la noche.
Posteriormente colonias de E.coli fueron inoculadas en medio LB (Triptona 1%,
extracto de levadura 0,5% (Difco) y NaCl 0,5% (Merck)) e incubadas a 37°C con
agitación a 250 rpm, durante 2 horas, hasta que el OD (600 nm) sea de 0,35 a
0,45.
Las células con valores óptimos de OD fueron centrifugadas a 4000 rpm por
10 min. a 4°C, el sobrenadante fue descartado y el pellet resuspendido con 20 ml
de 0,1 M CaCl2 y 15% glicerol frio. Después de 15 minutos de incubación en hielo,
se realizó una segunda centrifugación a 3000 rpm por 10 min. a 4°C, el

sobrenadante fue descartado y el pellet resuspendido con 1 ml de 0,1 M CaCl2 y
15% glicerol frio, finalmente las células competentes fueron conservadas a -70°C
en alícuotas de 50 μl.
El proceso de transformación se realizó mediante shock térmico, con 2 μl
del inserto ligado al vector pGEM-T easy y 50 μl de células competentes
previamente descongeladas a 4°C. Las células fueron incubadas en hielo a 4°C
por 30 min. e inmediatamente llevadas a baño maría a 42°C por 50 seg.
Posteriormente volúmenes de 50 μl, 100 μl y 150 μl de células transformadas
fueron sembradas en agar/LB suplementado con 100 μg/ml de carbenicilina, 80
μl/ml de 5-bromo-4-cloro-3-indolil-β-D-galactopiranósido (X-gal) y 0,5 mM de
isopropil-β-D-1-tiogalactopiranósido (IPTG); esparcidas con una densidad menor a
104 células por placa de cultivo e incubadas a 37°C durante toda la noche.
Análisis de recombinantes: La clonación y transformación fue confirmada
por el crecimiento de E. coli en agar/LB suplementado con antibiótico de selección
(carbenicilina 100 mg/ml) y para diferenciar entre células transformadas que
adquirieron o no el inserto, el medio de crecimiento fue enriquecido con X-gal
(sustrato cromogénico) e IPTG. De esta manera las células que captaron los
plásmidos con el operón lac intacto, una vez inducidas por IPTG, expresaron
activamente β-galactosidasa, que al hidrolizar a X-gal coloreo las colonias de azul.
Por otro lado las células que captaron los plásmidos con el operón lac
recombinante, con el inserto del fragmento de interés, sufrieron con la inserción la
inactivación del gen lacZ (uno de los genes que codifica la β-galactosidasa) y por
tanto, no expresaron β-galactosidasa ni degradaron X-gal, dejando las colonias de
color blanco. Dichas colonias blancas también fueron utilizadas para PCR, con los
oligonucleótidos específicos para los insertos, descrito anteriormente en la sección
5.3.4.4.

Purificación del plásmido calibrador: Colonias de células transformadas
que adquirieron el inserto fueron inoculadas en 10 ml de medio LB + Carbenicilina
e incubadas a 37°C con agitación a 250 rpm, durante toda la noche.
Posteriormente las células en medio LB fueron centrifugadas a 3000 rpm por 15
min. a 4°C, el sobrenadante fue descartado y el pellet resuspendido y purificado
mediante kit de purificación de plásmidos EasyPrep (Invitrogen, Life technologies),
siguiendo las instrucciones del fabricante. El DNA plasmídico fue diluido hasta
obtener una concentración equivalente a una copia/μl del fragmento clonado
(pvald1 ó pvmdr1), siendo esta de 3,5 atogramos/μl (3,513 x 10-18 gramos).

VI. RESULTADOS
6.1. DETECCIÓN DE LA MALARIA DURANTE LA VIGILANCIA
ACTIVA
La detección de la malaria por el sistema de vigilancia activa, se realizó a
través del Nested PCR (Fig. 1). Durante la primera evaluación se detectaron: 86%
(191/222) de casos con infección por P. vivax; 0,9% (2/222) de casos con
infección por P. falciparum, 9,5% (21/222) de casos con infección mixta y 3,6%
(8/222) fueron negativos. Durante la segunda evaluación, considerando los casos
con infección primaria por P. vivax, se detectaron recurrencias de la infección por
este parásito en 81% (180/222) de los casos, por P. falciparum 1,8% (4/222), por
infecciones mixtas 2,7% (6/222) y no hubo recurrencias 0,45% (1/222). También
durante la segunda evaluación se detectaron infecciones por P. vivax, en 0,9%
(2/222), 7,2% (16/222) y 2,7% (6/222) de casos, que durante la primera evaluación
fueron detectados como infecciones por P. falciparum, mixtas y negativas
respectivamente. Además se observó que 2,3% (5/222) de casos permanecieron
como infecciones mixtas y que 0,45% (1/222) de infecciones por P. falciparum así
como 0,45% (1/222) de infecciones mixtas detectados en la segunda evaluación,
anteriormente fueron negativos (Tabla 1 y Fig. 2).
Fig. 1. Diagnóstico de la malaria por Nested PCR. Detección de P. vivax, P. falciparum e
infecciones mixtas durante la vigilancia activa. M: marcador 100 pb., P1-P18: pacientes
diagnosticados (P11 con infección mixta, P17 con P. falciparum y P18 negativo), C+: control
positivo.
El diagnóstico por Nested PCR mostró una mayor sensibilidad en
comparación con la microscopía, realizada previamente, a través del método de
gota gruesa (GG). En la primera evaluación, la sensibilidad del Nested PCR fue de
un 99% frente a un 91,6% de la microscopía, en este grupo, solo 24 muestras
fueron diagnosticas como negativas por microscopía, de las cuales 18 fueron
positivas y 6 negativas por Nested PCR, así mismo solo 2 muestras que fueron
positivas por microscopía fueron negativos por Nested PCR, las cuales tuvieron un
conteo menor a 40 parásitos/μl en 50 campos observados y una densidad < 100
parásitos/μl. En la segunda evaluación, la sensibilidad del Nested PCR fue del
100% frente a 87,8% de la microscopía, en este grupo, de 28 muestras negativas
por microscopía, 27 fueron positivos y solo 1 negativa por Nested PCR (Tabla 2).
Tabla 1. Detección de la malaria mediante Nested PCR. Rango de frecuencias de P. vivax, P.
falciparum, infecciones mixtas y casos negativos, durante la vigilancia activa.
PRIMERA EVALUACIÓN
SEGUNDA EVALUACIÓN
INFECCIÓN P. vivax P. falciparum Mixtas Negativos
P. vivax 86,0%
(191/222)
81%
(180/222)
1,8%
(4/222)
2,7%
(6/222)
0,45%
(1/222)
P.
falciparum
0,9%
(2/222)
0,9%
(2/222)
0%
-----
0%
-----
0%
-----
Mixtas 9,5%
(21/222)
7,2%
(16/222)
0%
-----
2,3%
(5/222)
0%
-----
Negativas 3,6%
(8/222)
2,7%
(6/222)
0,45%
(1/222)
0,45%
(1/222)
0%
-----
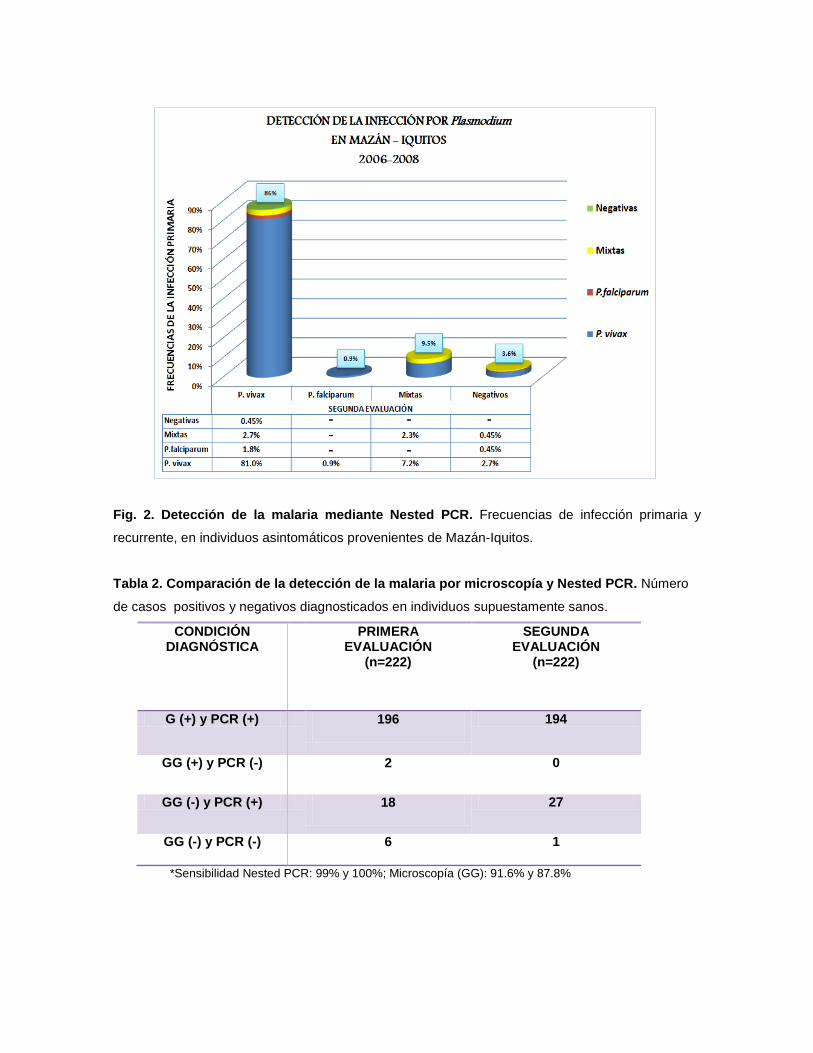
Fig. 2. Detección de la malaria mediante Nested PCR. Frecuencias de infección primaria y
recurrente, en individuos asintomáticos provenientes de Mazán-Iquitos.
Tabla 2. Comparación de la detección de la malaria por microscopía y Nested PCR. Número
de casos positivos y negativos diagnosticados en individuos supuestamente sanos.
CONDICIÓN DIAGNÓSTICA
PRIMERA EVALUACIÓN
(n=222)
SEGUNDA EVALUACIÓN
(n=222)
G (+) y PCR (+) 196 194
GG (+) y PCR (-) 2 0
GG (-) y PCR (+) 18 27
GG (-) y PCR (-) 6 1
*Sensibilidad Nested PCR: 99% y 100%; Microscopía (GG): 91.6% y 87.8%
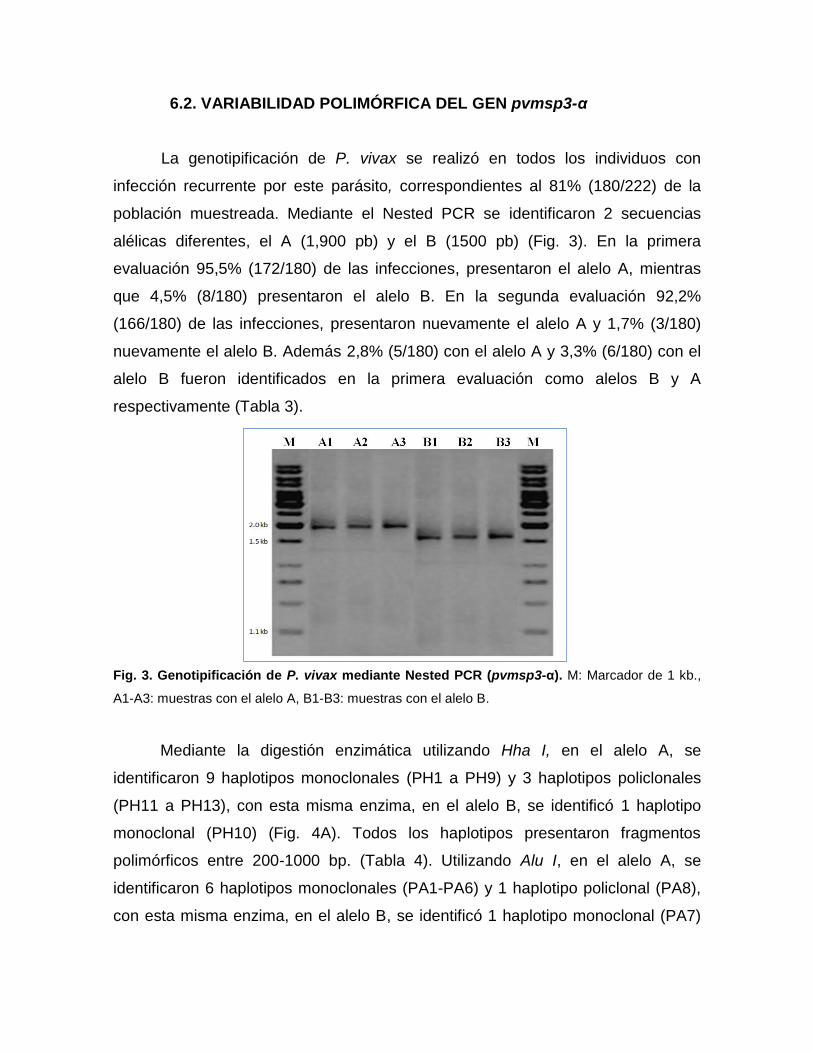
6.2. VARIABILIDAD POLIMÓRFICA DEL GEN pvmsp3-α
La genotipificación de P. vivax se realizó en todos los individuos con
infección recurrente por este parásito, correspondientes al 81% (180/222) de la
población muestreada. Mediante el Nested PCR se identificaron 2 secuencias
alélicas diferentes, el A (1,900 pb) y el B (1500 pb) (Fig. 3). En la primera
evaluación 95,5% (172/180) de las infecciones, presentaron el alelo A, mientras
que 4,5% (8/180) presentaron el alelo B. En la segunda evaluación 92,2%
(166/180) de las infecciones, presentaron nuevamente el alelo A y 1,7% (3/180)
nuevamente el alelo B. Además 2,8% (5/180) con el alelo A y 3,3% (6/180) con el
alelo B fueron identificados en la primera evaluación como alelos B y A
respectivamente (Tabla 3).
Fig. 3. Genotipificación de P. vivax mediante Nested PCR (pvmsp3-α). M: Marcador de 1 kb.,
A1-A3: muestras con el alelo A, B1-B3: muestras con el alelo B.
Mediante la digestión enzimática utilizando Hha I, en el alelo A, se
identificaron 9 haplotipos monoclonales (PH1 a PH9) y 3 haplotipos policlonales
(PH11 a PH13), con esta misma enzima, en el alelo B, se identificó 1 haplotipo
monoclonal (PH10) (Fig. 4A). Todos los haplotipos presentaron fragmentos
polimórficos entre 200-1000 bp. (Tabla 4). Utilizando Alu I, en el alelo A, se
identificaron 6 haplotipos monoclonales (PA1-PA6) y 1 haplotipo policlonal (PA8),
con esta misma enzima, en el alelo B, se identificó 1 haplotipo monoclonal (PA7)
(Fig. 4B). En este caso todos los haplotipos presentaron fragmentos polimórficos
entre 150-550 bp. (Tabla 4).
Tabla 3. Genotipificación de P. vivax en individuos con infección recurrente. Rango de
frecuencias del alelo A (1900 pb) y alelo B (1500 pb).
PRIMERA
EVALUACIÓN
SEGUNDA
EVALUACIÓN
ALELOS A B
A 95,5% (172/180)
92,2%
(166/180)
3,3% (6/180)
B 4,5% (8/180)
2,8%
(5/180)
1,7% (3/180)
Con la combinación de ambas enzimas se logró identificar una mayor
variabilidad genética, con 14 haplotipos monoclonales (PAH1-PAH14) y 7
haplotipos policlonales (PHA15-PHA21) (Tabla 4). Analizando la recurrencia de
estos haplotipos, se identificó a PHA13, PHA2 y PHA6 como los más recurrentes
con 5,5%, 7,2% y 12,2%, respectivamente. Por otra parte los haplotipos PHA8,
PHA10, PHA17 y PHA 19 no fueron identificados en la primera evaluación, por lo
tanto no se pudo evaluar las recurrencias en estos haplotipos. Del mismo modo,
fue posible identificar la frecuencia de haplotipos no recurrentes y policlonales
(presentes como haplotipos diferentes en la segunda evaluación) (Fig. 5).
En el análisis estadístico, el 5,6% (10/180) de los individuos en la primera
evaluación y 3,9% (7/180) en la segunda evaluación, presentaron un COI>1,
correspondientes a infecciones por haplotipos policlonales o infecciones mixtas.
Los valores PLD fueron He=0,88 y 0,87 en ambos periodos de evaluación,
demostrando una alta variabilidad genética. Además la prueba F (α=0,05; Fp<Ft),
indicó que no existe diferencias reales entre las frecuencias de haplotipos
identificados, en ambos periodos de evaluación (Tabla 5).
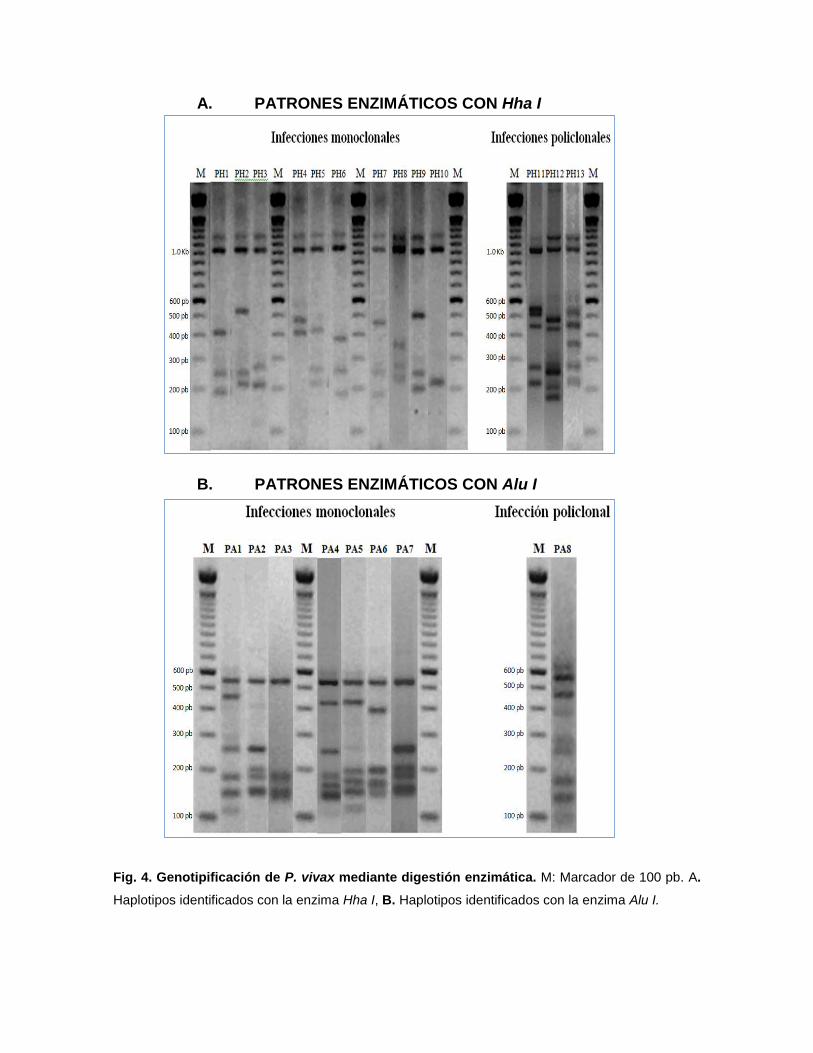
A. PATRONES ENZIMÁTICOS CON Hha I
B. PATRONES ENZIMÁTICOS CON Alu I
Fig. 4. Genotipificación de P. vivax mediante digestión enzimática. M: Marcador de 100 pb. A.
Haplotipos identificados con la enzima Hha I, B. Haplotipos identificados con la enzima Alu I.
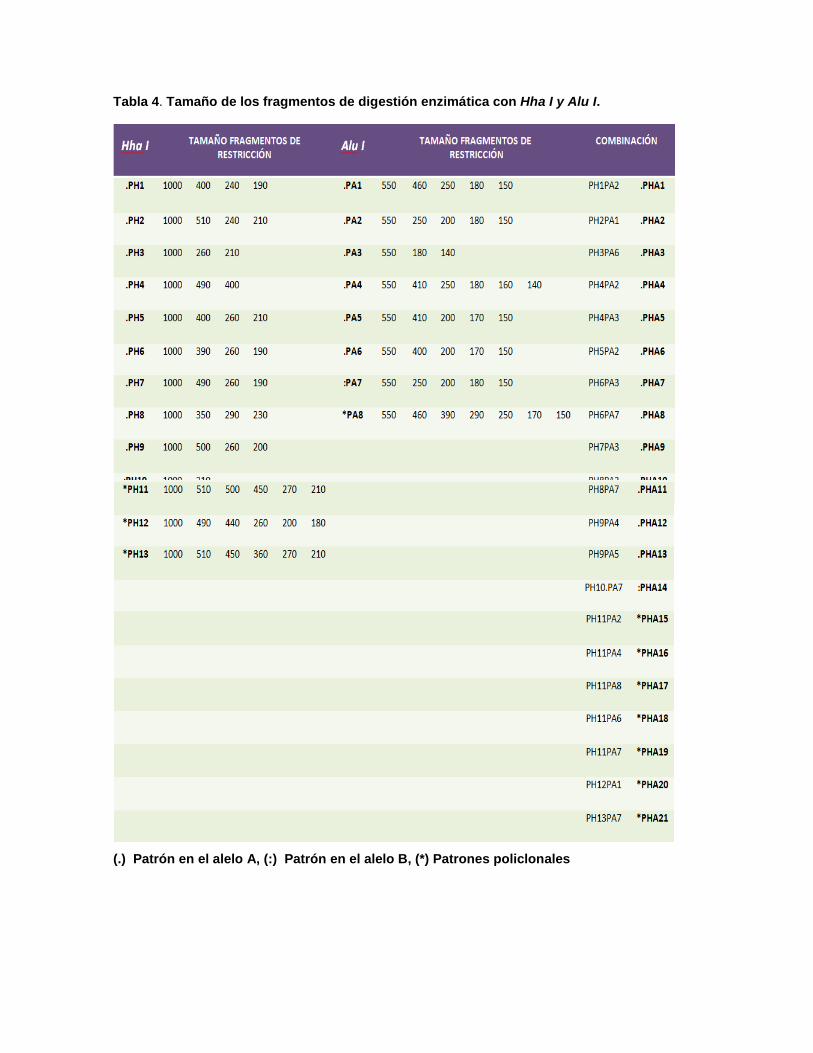
Tabla 4. Tamaño de los fragmentos de digestión enzimática con Hha I y Alu I.
(.) Patrón en el alelo A, (:) Patrón en el alelo B, (*) Patrones policlonales
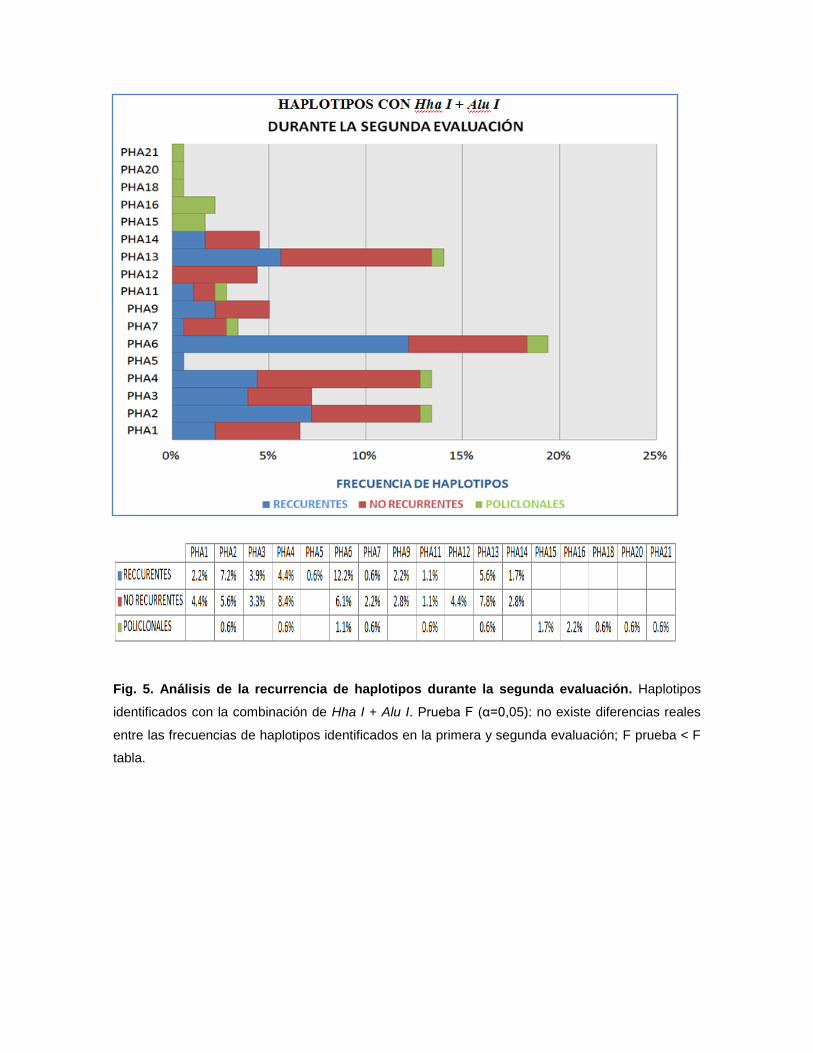
Fig. 5. Análisis de la recurrencia de haplotipos durante la segunda evaluación. Haplotipos
identificados con la combinación de Hha I + Alu I. Prueba F (α=0,05): no existe diferencias reales
entre las frecuencias de haplotipos identificados en la primera y segunda evaluación; F prueba < F
tabla.

Tabla 5. Evaluación de la variabilidad genética de pvmsp3-α, COI: Complejidad de la infección,
PLD: Nivel de la Diversidad poblacional, He: Heterocigosidad esperada, F: Prueba de Fisher.
6.3. PATRONES DE RECURRENCIA
La variabilidad genética permitió identificar recurrencias por haplotipos
monoclonales idénticos (relapsos) a la infección primaria en 41,7% (75/180) y por
haplotipos monoclonales diferentes (reinfecciones) en 48,9% (88/180) de los
casos. Las infecciones con el haplotipo PHA6 tuvieron la mayor frecuencia de
recurrencias por el mismo haplotipo con 12,2% y las infecciones con PHA5 y
PHA7 tuvieron las menores recurrencias por el mismo haplotipo, ambos con 0,6%
(1/180), además la infección con PHA5 presentó recurrencia solo por el mismo
haplotipo. Por otra parte las infecciones con el haplotipo PHA4 tuvieron la mayor
frecuencia de recurrencias, pero por haplotipos diferentes, con 8,4% (15/180) y las
infecciones con PHA 11 tuvieron recurrencias de 1,1% (2/180) tanto por el mismo
haplotipo como por haplotipos diferentes. También las infecciones con el haplotipo
PHA12 presentaron únicamente recurrencias por haplotipos diferentes (Tabla 6 y
Fig. 6).
EVALUACIÓN DE LA VARIABILIDAD GENÉTICA DE pvmsp3-α
PRIMERA EVALUACIÓN
SEGUNDA EVALUACIÓN
ENZIMAS Hha I + Alu I Hha I + Alu I
N° HAPLOTIPOS
21 21
COI>1 5,6% 3,9%
PLD (He) 0,88 0,87
Fisher (F)
Fp < Ft ; α=0,05 (0,0023 < 4,22)
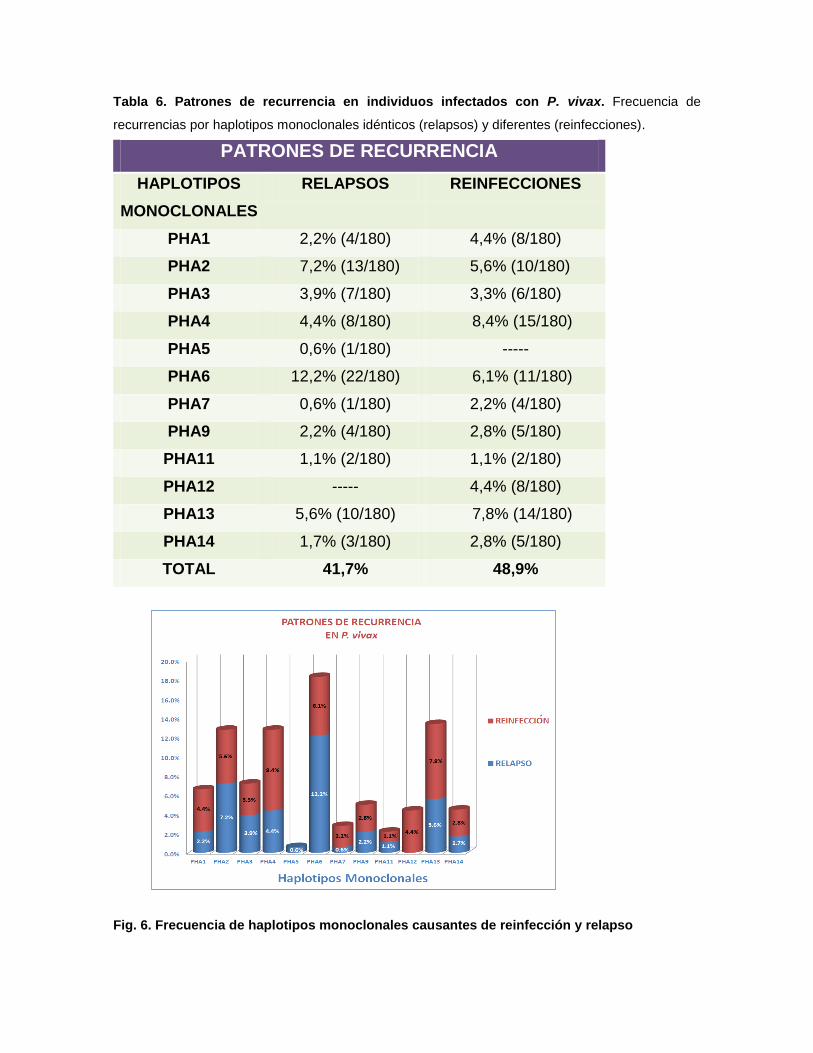
Tabla 6. Patrones de recurrencia en individuos infectados con P. vivax. Frecuencia de
recurrencias por haplotipos monoclonales idénticos (relapsos) y diferentes (reinfecciones).
PATRONES DE RECURRENCIA
HAPLOTIPOS
MONOCLONALES
RELAPSOS REINFECCIONES
PHA1 2,2% (4/180) 4,4% (8/180)
PHA2 7,2% (13/180) 5,6% (10/180)
PHA3 3,9% (7/180) 3,3% (6/180)
PHA4 4,4% (8/180) 8,4% (15/180)
PHA5 0,6% (1/180) -----
PHA6 12,2% (22/180) 6,1% (11/180)
PHA7 0,6% (1/180) 2,2% (4/180)
PHA9 2,2% (4/180) 2,8% (5/180)
PHA11 1,1% (2/180) 1,1% (2/180)
PHA12 ----- 4,4% (8/180)
PHA13 5,6% (10/180) 7,8% (14/180)
PHA14 1,7% (3/180) 2,8% (5/180)
TOTAL 41,7% 48,9%
Fig. 6. Frecuencia de haplotipos monoclonales causantes de reinfección y relapso
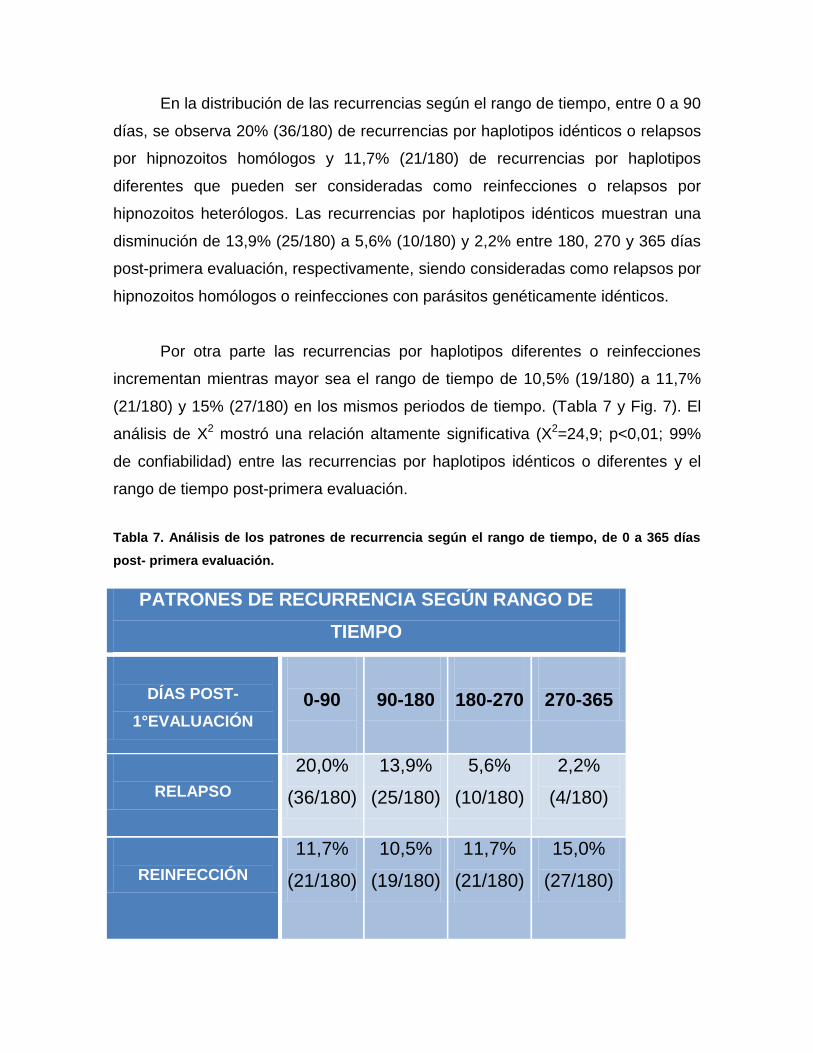
En la distribución de las recurrencias según el rango de tiempo, entre 0 a 90
días, se observa 20% (36/180) de recurrencias por haplotipos idénticos o relapsos
por hipnozoitos homólogos y 11,7% (21/180) de recurrencias por haplotipos
diferentes que pueden ser consideradas como reinfecciones o relapsos por
hipnozoitos heterólogos. Las recurrencias por haplotipos idénticos muestran una
disminución de 13,9% (25/180) a 5,6% (10/180) y 2,2% entre 180, 270 y 365 días
post-primera evaluación, respectivamente, siendo consideradas como relapsos por
hipnozoitos homólogos o reinfecciones con parásitos genéticamente idénticos.
Por otra parte las recurrencias por haplotipos diferentes o reinfecciones
incrementan mientras mayor sea el rango de tiempo de 10,5% (19/180) a 11,7%
(21/180) y 15% (27/180) en los mismos periodos de tiempo. (Tabla 7 y Fig. 7). El
análisis de X2 mostró una relación altamente significativa (X2=24,9; p<0,01; 99%
de confiabilidad) entre las recurrencias por haplotipos idénticos o diferentes y el
rango de tiempo post-primera evaluación.
Tabla 7. Análisis de los patrones de recurrencia según el rango de tiempo, de 0 a 365 días
post- primera evaluación.
PATRONES DE RECURRENCIA SEGÚN RANGO DE
TIEMPO
DÍAS POST-
1°EVALUACIÓN
0-90
90-180
180-270
270-365
RELAPSO
20,0%
(36/180)
13,9%
(25/180)
5,6%
(10/180)
2,2%
(4/180)
REINFECCIÓN
11,7%
(21/180)
10,5%
(19/180)
11,7%
(21/180)
15,0%
(27/180)
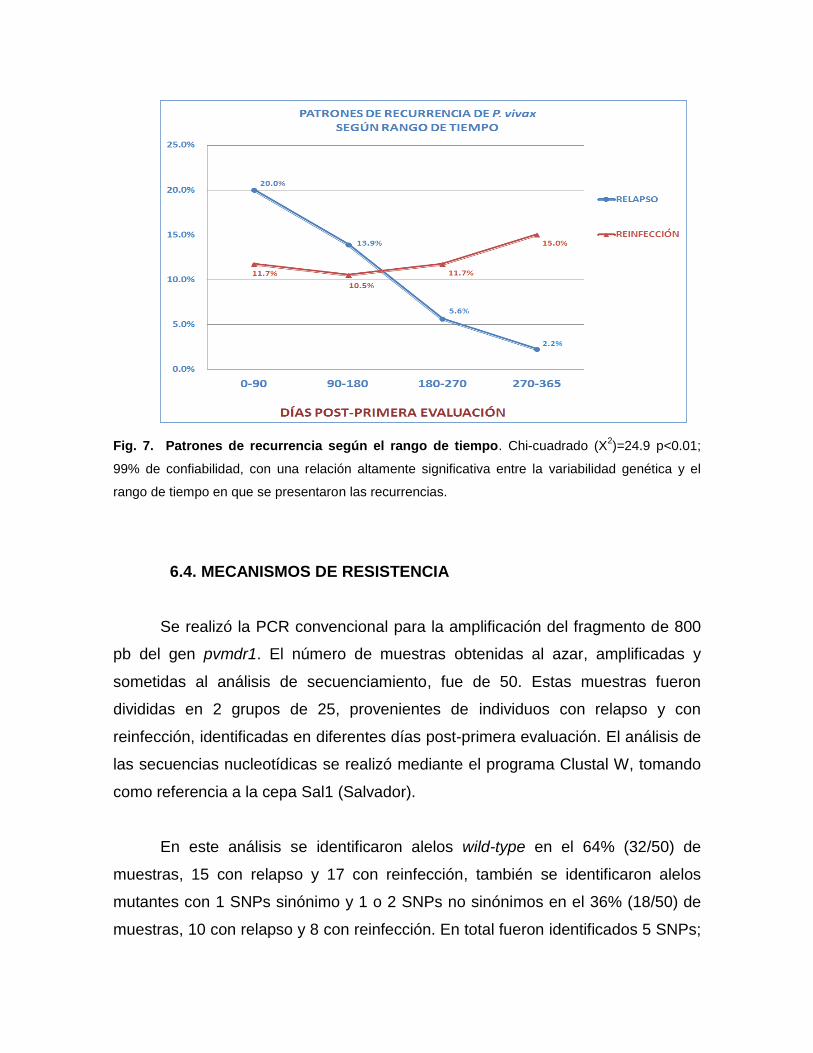
Fig. 7. Patrones de recurrencia según el rango de tiempo. Chi-cuadrado (X2)=24.9 p<0.01;
99% de confiabilidad, con una relación altamente significativa entre la variabilidad genética y el
rango de tiempo en que se presentaron las recurrencias.
6.4. MECANISMOS DE RESISTENCIA
Se realizó la PCR convencional para la amplificación del fragmento de 800
pb del gen pvmdr1. El número de muestras obtenidas al azar, amplificadas y
sometidas al análisis de secuenciamiento, fue de 50. Estas muestras fueron
divididas en 2 grupos de 25, provenientes de individuos con relapso y con
reinfección, identificadas en diferentes días post-primera evaluación. El análisis de
las secuencias nucleotídicas se realizó mediante el programa Clustal W, tomando
como referencia a la cepa Sal1 (Salvador).
En este análisis se identificaron alelos wild-type en el 64% (32/50) de
muestras, 15 con relapso y 17 con reinfección, también se identificaron alelos
mutantes con 1 SNPs sinónimo y 1 o 2 SNPs no sinónimos en el 36% (18/50) de
muestras, 10 con relapso y 8 con reinfección. En total fueron identificados 5 SNPs;

en las posiciones de los codones 958 (ATG/ACG), 976 (TAC/TTC), 1022
(CTA/TTA), 1076 (TTT/CTT) y 1125(GAA/AAA). (Fig. 8).
Las secuencias nucleotídicas fueron sometidas al proceso de transducción
in sílico mediante el software Translate a DNA Sequence, el cual permitió
identificar el cambio de aminoácidos en la proteína MDR1. Mediante este proceso
fueron identificados 4 mutaciones no sinónimas o cambio de aminoácidos en los
codones M958T, Y976F, F1076L y E1125K (Fig. 9) y 1 mutación sinónima, el cual
no originó cambio de aminoácidos, en el codón L1022L.
En el grupo de muestras de individuos con relapso, 12% (3/25) presentaron
SNPs únicos sinónimos en el codón L1022L. Así mismo 12% (3/25) presentaron
SNPs únicos no sinónimos en el codón M958T y 16% (4/25) presentaron SNPs
dobles no sinónimos en el codón Y976F y F1076L, estos últimos están reportados
como posibles marcadores de resistencia a cloroquina por P. vivax (Fig. 10A). En
el grupo de individuos con reinfección, 8% (2/25) presentaron SNPs únicos
sinónimos en el codón L1022L, 24% (6/25) presentaron SNPs únicos no
sinónimos, 4 en el codón M958T y 2 en el codón E1125K, los cuales no están
relacionados con mecanismos de resistencia. (Fig. 10B). Mediante el análisis
estadístico se encontró una relación significativa entre la presencia de SNPs que
originan mutaciones no sinónimas y la ocurrencia de relapsos con X2=3.84;
p<0.05, correspondiente al 95% de confiabilidad.
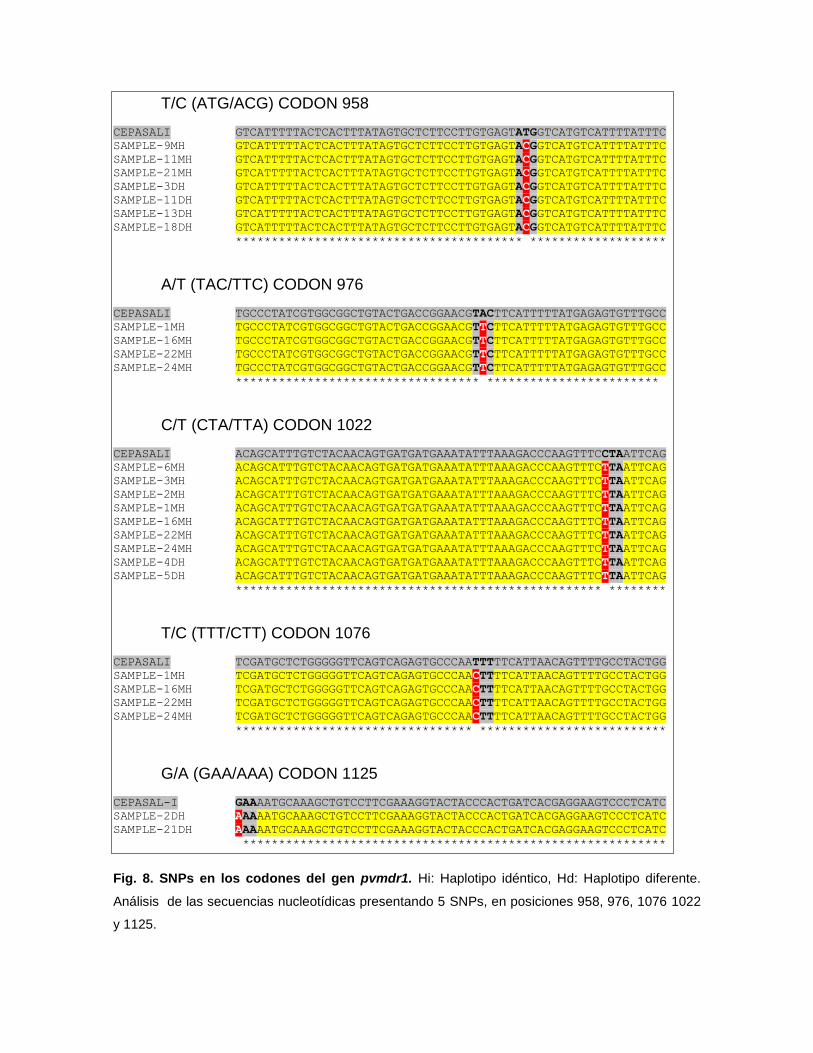
T/C (ATG/ACG) CODON 958
CEPASALI GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTATGGTCATGTCATTTTATTTC
SAMPLE-9MH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-11MH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-21MH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-3DH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-11DH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-13DH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
SAMPLE-18DH GTCATTTTTACTCACTTTATAGTGCTCTTCCTTGTGAGTACGGTCATGTCATTTTATTTC
**************************************** *******************
A/T (TAC/TTC) CODON 976
CEPASALI TGCCCTATCGTGGCGGCTGTACTGACCGGAACGTACTTCATTTTTATGAGAGTGTTTGCC
SAMPLE-1MH TGCCCTATCGTGGCGGCTGTACTGACCGGAACGTTCTTCATTTTTATGAGAGTGTTTGCC
SAMPLE-16MH TGCCCTATCGTGGCGGCTGTACTGACCGGAACGTTCTTCATTTTTATGAGAGTGTTTGCC
SAMPLE-22MH TGCCCTATCGTGGCGGCTGTACTGACCGGAACGTTCTTCATTTTTATGAGAGTGTTTGCC
SAMPLE-24MH TGCCCTATCGTGGCGGCTGTACTGACCGGAACGTTCTTCATTTTTATGAGAGTGTTTGCC
********************************** ************************
C/T (CTA/TTA) CODON 1022
CEPASALI ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCCTAATTCAG
SAMPLE-6MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-3MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-2MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-1MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-16MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-22MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-24MH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-4DH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
SAMPLE-5DH ACAGCATTTGTCTACAACAGTGATGATGAAATATTTAAAGACCCAAGTTTCTTAATTCAG
*************************************************** ********
T/C (TTT/CTT) CODON 1076
CEPASALI TCGATGCTCTGGGGGTTCAGTCAGAGTGCCCAATTTTTCATTAACAGTTTTGCCTACTGG
SAMPLE-1MH TCGATGCTCTGGGGGTTCAGTCAGAGTGCCCAACTTTTCATTAACAGTTTTGCCTACTGG
SAMPLE-16MH TCGATGCTCTGGGGGTTCAGTCAGAGTGCCCAACTTTTCATTAACAGTTTTGCCTACTGG
SAMPLE-22MH TCGATGCTCTGGGGGTTCAGTCAGAGTGCCCAACTTTTCATTAACAGTTTTGCCTACTGG
SAMPLE-24MH TCGATGCTCTGGGGGTTCAGTCAGAGTGCCCAACTTTTCATTAACAGTTTTGCCTACTGG
********************************* **************************
G/A (GAA/AAA) CODON 1125
CEPASAL-I GAAAATGCAAAGCTGTCCTTCGAAAGGTACTACCCACTGATCACGAGGAAGTCCCTCATC
SAMPLE-2DH AAAAATGCAAAGCTGTCCTTCGAAAGGTACTACCCACTGATCACGAGGAAGTCCCTCATC
SAMPLE-21DH AAAAATGCAAAGCTGTCCTTCGAAAGGTACTACCCACTGATCACGAGGAAGTCCCTCATC
***********************************************************
Fig. 8. SNPs en los codones del gen pvmdr1. Hi: Haplotipo idéntico, Hd: Haplotipo diferente.
Análisis de las secuencias nucleotídicas presentando 5 SNPs, en posiciones 958, 976, 1076 1022
y 1125.

MUTACIÓN M958T
CEPASAL-I VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-9MH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-11MH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-21MH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-3DH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-11DH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-13DH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-18DH VIFTHFIVLFLVSTVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
************* **********************************************
MUTACIÓN Y976F
CEPASAL-I VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTYFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-1MH VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTFFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-16MH VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTFFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-22MH VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTFFIFMRVFAIRARIAANKDVEKKRVNQPG
SAMPLE-24MH VIFTHFIVLFLVSMVMSFYFCPIVAAVLTGTFFIFMRVFAIRARIAANKDVEKKRVNQPG
*******************************:****************************
MUTACIÓN F1076L
CEPASAL-I SMLWGFSQSAQFFINSFAYWFGSFLIRRGTIQVDDFMKSLFTFLFTGSYAGKLMSLKGDS
SAMPLE-1MH SMLWGFSQSAQLFINSFAYWFGSFLIRRGTIQVDDFMKSLFTFLFTGSYAGKLMSLKGDS
SAMPLE-16MH SMLWGFSQSAQLFINSFAYWFGSFLIRRGTIQVDDFMKSLFTFLFTGSYAGKLMSLKGDS
SAMPLE-22MH SMLWGFSQSAQLFINSFAYWFGSFLIRRGTIQVDDFMKSLFTFLFTGSYAGKLMSLKGDS
SAMPLE-24MH SMLWGFSQSAQLFINSFAYWFGSFLIRRGTIQVDDFMKSLFTFLFTGSYAGKLMSLKGDS
***********:************************************************
MUTACIÓN E1125K
CEPASAL-I ENAKLSFERYYPLITRKSLIDVRDNGGIKIKNSNDIKG
SAMPLE-2DH KNAKLSFERYYPLITRKSLIDVRDNGGIKIKNSNDIKG
SAMPLE-21DH KNAKLSFERYYPLITRKSLIDVRDNGGIKIKNSNDIKG
:*************************************
Fig. 9. Mutaciones no sinónimas en las secuencias aminoacídicas de la proteína MDR1. Hi:
Haplotipo idéntico, Hd: Haplotipo diferente. Análisis de las mutaciones no sinónimas (marcadas en
azul) originando el cambio de aminoácidos M/T en el codón 958, Y/F en el codón 976, F/L en el
codón 1076 y E/K en el codón 1125.

A. SNPs Y976F, SNPs F1076L B. SNPs E1125K
SNPs M958T SNPs M958T
Wild-type Wild-type
Fig. 10. Distribución de SNPs codificando mutaciones sinónimas y no sinónimas
(Cladograma). A. Distancias filogenéticas en casos con relapso, B. Distancias filogenéticas en
casos con reinfección.
Mediante el análisis in sílico, con el software U-Melt, se identificaron las
temperaturas de fusión en alta resolución (HRM), distinguiendo claramente los
SNPs en alelos mutantes así como los alelos wild-type en los codones 976 y 1076
del gen pvmdr1. Se analizaron fragmentos pequeños de hasta 100 pb
correspondientes a regiones con las mutaciones no sinónimas relacionadas con
resistencia. Se encontraron diferencias en los picos de las temperaturas de fusión
de ambos alelos Y976F y L1076F de hasta 0,5°C. El análisis mostró una
temperatura de 85,6°C para el alelo wild-type Y976 y 85,1°C para el alelo mutante
F976 (Fig. 11A). También se observó una temperatura de 83,5°C para el alelo wil-
type L1076 y 84°C para el alelo mutante F1076. (Fig.11B).
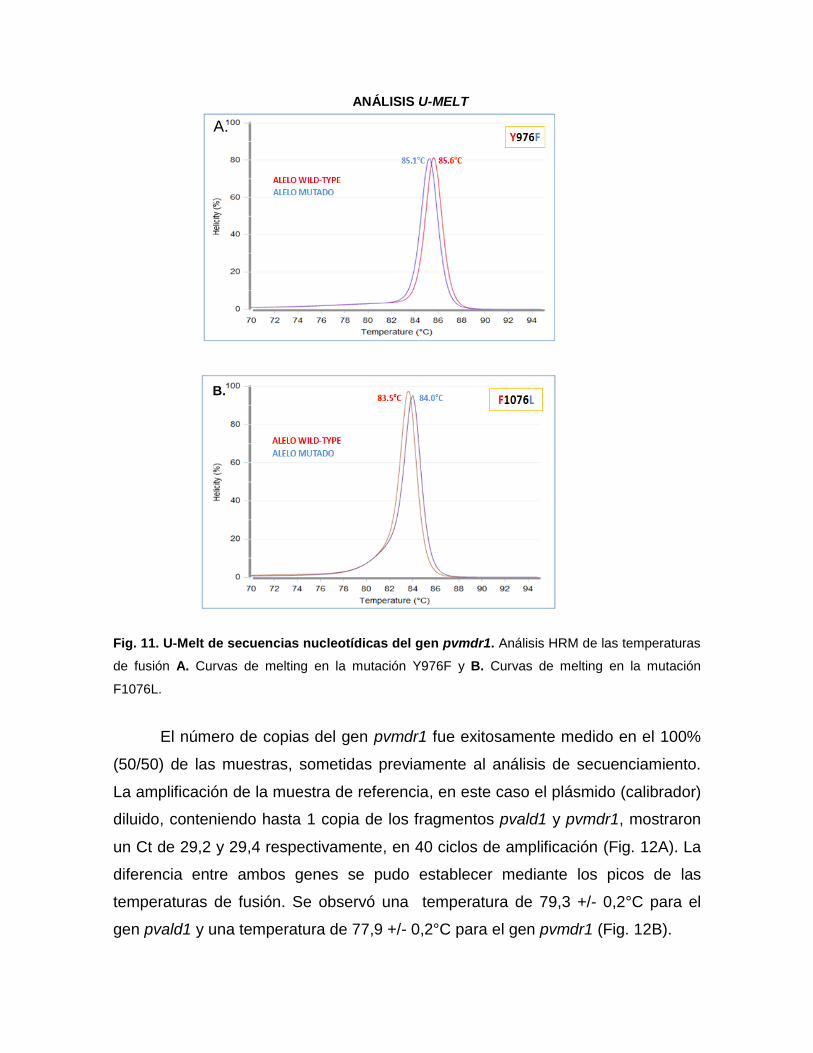
ANÁLISIS U-MELT
A.
B.
Fig. 11. U-Melt de secuencias nucleotídicas del gen pvmdr1. Análisis HRM de las temperaturas
de fusión A. Curvas de melting en la mutación Y976F y B. Curvas de melting en la mutación
F1076L.
El número de copias del gen pvmdr1 fue exitosamente medido en el 100%
(50/50) de las muestras, sometidas previamente al análisis de secuenciamiento.
La amplificación de la muestra de referencia, en este caso el plásmido (calibrador)
diluido, conteniendo hasta 1 copia de los fragmentos pvald1 y pvmdr1, mostraron
un Ct de 29,2 y 29,4 respectivamente, en 40 ciclos de amplificación (Fig. 12A). La
diferencia entre ambos genes se pudo establecer mediante los picos de las
temperaturas de fusión. Se observó una temperatura de 79,3 +/- 0,2°C para el
gen pvald1 y una temperatura de 77,9 +/- 0,2°C para el gen pvmdr1 (Fig. 12B).
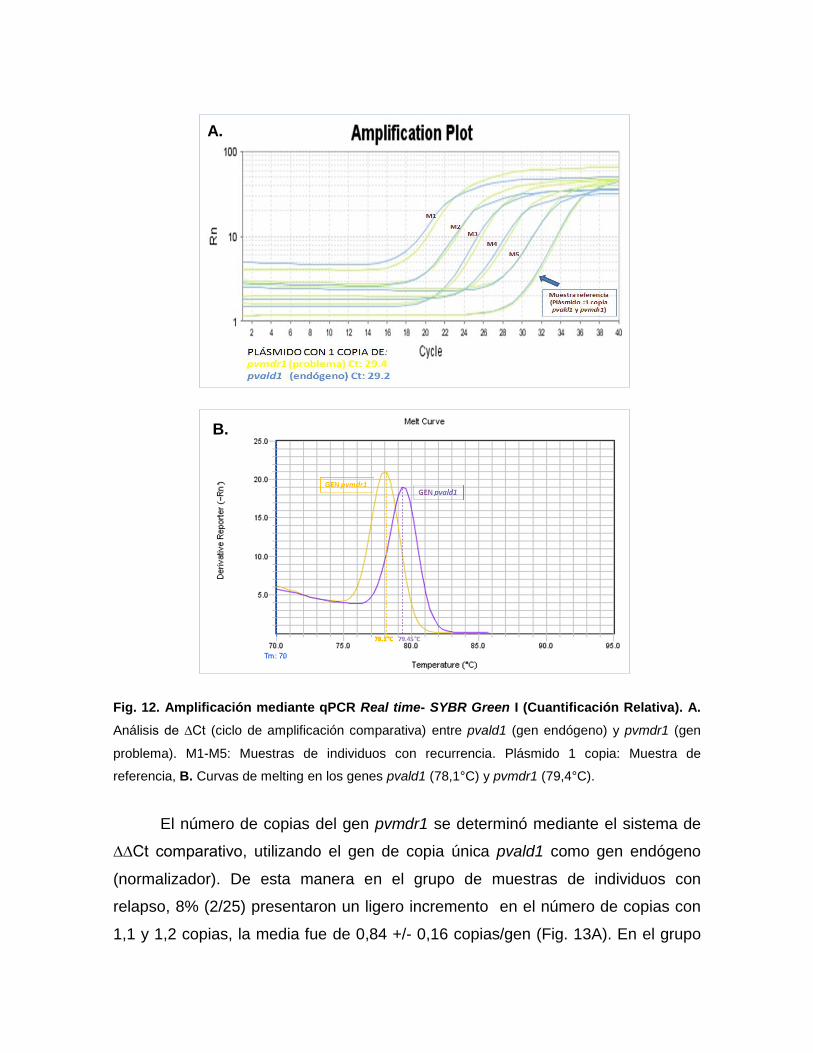
A.
B.
Fig. 12. Amplificación mediante qPCR Real time- SYBR Green I (Cuantificación Relativa). A.
Análisis de ∆Ct (ciclo de amplificación comparativa) entre pvald1 (gen endógeno) y pvmdr1 (gen
problema). M1-M5: Muestras de individuos con recurrencia. Plásmido 1 copia: Muestra de
referencia, B. Curvas de melting en los genes pvald1 (78,1°C) y pvmdr1 (79,4°C).
El número de copias del gen pvmdr1 se determinó mediante el sistema de
∆∆Ct comparativo, utilizando el gen de copia única pvald1 como gen endógeno
(normalizador). De esta manera en el grupo de muestras de individuos con
relapso, 8% (2/25) presentaron un ligero incremento en el número de copias con
1,1 y 1,2 copias, la media fue de 0,84 +/- 0,16 copias/gen (Fig. 13A). En el grupo
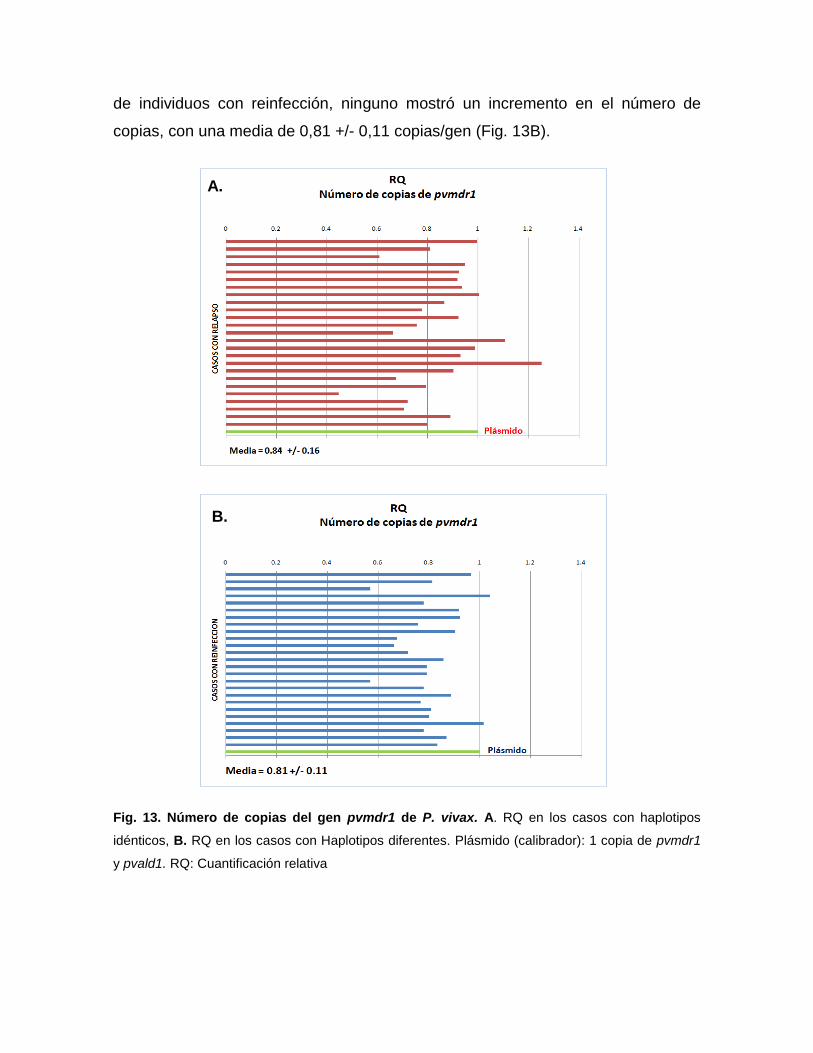
de individuos con reinfección, ninguno mostró un incremento en el número de
copias, con una media de 0,81 +/- 0,11 copias/gen (Fig. 13B).
A.
B.
Fig. 13. Número de copias del gen pvmdr1 de P. vivax. A. RQ en los casos con haplotipos
idénticos, B. RQ en los casos con Haplotipos diferentes. Plásmido (calibrador): 1 copia de pvmdr1
y pvald1. RQ: Cuantificación relativa

VII. DISCUSIÓN
7.1. DETECCIÓN DE LA MALARIA DURANTE LA VIGILANCIA
ACTIVA
Estudios realizados por Mayxay et al., 2001, mencionan que en muchas
regiones endémicas existen problemas y limitaciones, asociadas con la
dependencia al diagnóstico microscópico, para la detección de casos activos y
pasivos de la malaria. Por su parte Siripoon et al., 2002, indican que en el caso de
individuos sintomáticos con relativa alta parasitemia (>500 parásitos asexuales/μl)
el diagnóstico por microscopía puede ser agudo y usado para iniciar un apropiado
tratamiento, sin embargo la agudeza de este método puede disminuir
significativamente en los casos de individuos con bajos niveles de parasitemia,
como en aquellos individuos asintomáticos donde los rangos de parasitemia son a
menudo muy bajos. Una solución a este problema es el uso de técnicas
novedosas como la PCR, planteado por Singh et al., 1999, la cual ofrece alta
sensibilidad y especificidad para la detección del DNA genómico de Plasmodium,
además es efectiva para la determinación de infecciones mixtas. Barker et al.,
1992 y Snounou et al., 1993 indicaron que la PCR fue más sensible y específico
que la examinación microscópica por gota gruesa y frotis sanguíneo,
particularmente en casos con baja parasitemia o infecciones mixtas.Por su parte
Andrade et al., 2010 usaron una variante, el Nested PCR, para el diagnóstico de
Plasmodium, el cual fue utilizado como prueba estándar en diversos estudios.
En Mazán-Iquitos, el uso del Nested PCR para el diagnóstico de
Plasmodium presentó una alta capacidad de discriminación entre las especies,
identificación de infecciones mixtas, además de una alta sensibilidad comparada
con la microscopía. Se detectó tan solo 0,9% (2/222) de muestras negativas para
el Nested PCR y positivas por microscopía, lo cual está relacionado con las bajas
densidades observadas, < 50 parásitos/μl. Estos resultados también son
observados por Scopel et al., 2004 quienes determinaron que el uso de DNA

extraído a partir del frotis sanguíneo presentó una pobre detección de malaria,
particularmente con densidades menores a 20 parásitos/μl.
Los hallazgos en este estudio mostraron individuos asintomáticos con altas
frecuencias de infecciones por P. vivax y bajas por P. falciparum, condiciones
similares a las reportadas en Venezuela por Rodulfo et al., 2007, quienes
indicaron infecciones asintomáticas de 34,8% para P. vivax y 15,2% para P.
falciparum. Otros estudios como de Orjuela et al., 2009 en la amazonía brasileña,
indicaron bajos índices de transmisión tanto en P. vivax como en P. falciparum,
con un alto número de individuos asintomáticos no detectados. Esto es
corroborado por Sojo et al., 2008 y Sutton et al., 2009, por estudios realizados en
la amazonia peruana.
Generalmente los estudios epidemiológicos no consideran las altas
frecuencias de infecciones asintomáticas, por lo que es necesario el empleo de
métodos de vigilancia activa para la detección de las especies de Plasmodium.
Este el principal problema en muchas regiones endémicas del mundo, debido
principalmente a que los individuos desarrollan un incremento de la inmunidad y
resistencia a la enfermedad, después de repetidas infecciones, lo cual minimiza la
intensidad de los síntomas, manteniendo bajos niveles parasitemia por largos
períodos de tiempo. Este hecho es epidemiológicamente importante ya que los
portadores asintomáticos son un reservorio frecuente de la infección. Al respecto,
en Cali-Colombia, Echeverri et al., 2011 reportaron un caso de malaria transmitida
por transfusión a un recién nacido prematuro, por la sangre infectada de un
donante asintomático. La prueba de la gota gruesa confirmó la enfermedad por
Plasmodium vivax en el recién nacido y la muestra del donante se sometió a PCR
y fue positiva para la misma especie.

7.2. VARIABILIDAD POLIMÓRFICA DEL GEN pvmsp3-α
La genotipificación realizada en las muestras de Mazán, se basó en el
polimorfismo del gen pvmsp3-α, varios estudios como los efectuados en Guyana
Francesa por Verón et al., 2009, en Perú por Ayala et al., 2005, en Colombia por
Cristiano et al., 2008 y en Brasil por Ribeiro et al., 2011, también utilizaron este
gen como marcador genético para evaluar la diversidad poblacional de P. vivax.
La genotipificación mediante el Nested-PCR permitió detectar 2 secuencias
alélicas diferentes, de 1900 pb (Alelo A) y de 1500 pb (Alelo B). Adicionalmente,
investigaciones realizadas por Bruce et al., 1999 en Papúa Nueva Guinea y Cui et
al., 2003 en Tailandia, reportaron un tercer alelo de 1100 pb (Alelo C), incluso
investigaciones realizadas por Prajapati et al., 2010 en la India (Chennai),
mencionaron a un cuarto alelo de 500 pb (Alelo D). En este estudio, los resultados
de las frecuencias para estos alelos (A y B) son concordantes con los reportados
por Bruce et al., 1999, quienes indicaron un 70,5% y 6,7% y por Cui et al., 2003,
quienes indicaron un 74,8% y 6,5%, respectivamente. Sin embargo en regiones
como Venezuela Ord et al., 2005, indicaron 9,3% para el alelo A y 21,9% para el
alelo B. Incluso contrastando con la mayoría de investigaciones de esta parte del
mundo, un estudio realizado en el Sur de Irán por Zakeri et al., 2006 mencionaron
al alelo C como el más prevalente. Estos resultados nos muestran un predominio
de determinado tipo alélico, por una determinada región, aunque con una escaza
variabilidad genética de P. vivax, considerando al alelo A como el más prevalente
en muchas regiones endémicas.
Mediante la utilización de técnicas de genotipificación combinadas como la
PCR–RFLP, se observó una mayor variabilidad genética en P. vivax. Se
identificaron 10 haplotipos monoclonales con la enzima Hha I y 7 con Alu I, de los
cuales 6 con Hha I y 3 con Alu I, también fueron encontrados por Prajapati et al.,
2011 en la India, por Cui et al., 2003 en Tailandia y por Zakeri et al., 2006 en Irán y
al menos 3 fueron encontradas por Bruce et al., 1999 en Papúa Nueva Guinea;

sugiriendo que algunas de estas variantes alélicas tienen una distribución global.
Por otra parte también se pudo observar modificaciones en los patrones
enzimáticos del alelo B, identificado por Ribeiro et al., 2011 en Brasil, el cual
presentó diferentes tamaños de fragmentos digeridos comparados con los
encontrados en este estudio. Al igual que en otros estudios también se
identificaron haplotipos policlonales, los cuales representan el 9,5% (5,6% en la
primera evaluación y 3,9% en la segunda evaluación) de los casos,
contrariamente, Ribeiro et al., 2011 indicaron 14% de infecciones policlonales
analizadas solo con la enzima Hha I, por su parte Bruce et al., 1999 y Cui et al.,
2003 también encontraron un mayor número de infecciones policlonales o mixtas.
La variabilidad genética mostrada por P. vivax indica que la diversidad de
este parásito es mantenida por una selección positiva, sin embargo puede haber
otras razones incluyendo algunas de las propiedades biológicas del parásito tales
como su habilidad para ocasionar relapsos, interacción con poblaciones de P.
falciparum co-circulantes o por la migración de personas infectadas desde una
región a otra. Esta posibilidad es de especial atención porque durante los últimos
años, parte de la población rural de la selva peruana ha sido sujeta a constante y
continuo desplazamiento. En este estudio la variabilidad genética fue evaluada
determinando los niveles de significancia de la diversidad poblacional (PLD) en
base a la heterocigosidad esperada (He) y la complejidad de la infección (COI).
Los datos encontrados coinciden con el estudio efectuado por Sutton et al., 2009,
en la amazonía peruana, en donde mencionaron un PLD cercano a la unidad
(He=0,845), pero un 23,6% de infecciones policlonales (COI>1), siendo mayores a
los porcentajes encontrados en este estudio.

7.3. PATRONES DE RECURRENCIA
En este estudio se encontró 81% de recurrencias por P. vivax en un periodo
de 365 días post-primera evaluación, por su parte Orjuela et al., 2009b, reportaron
rangos de recurrencia entre 26% a 40% dentro de 180 días post-tratamiento.
Mediante métodos de genotipificación, Chen et al., 2007 asociaron un solo tipo
alélico al 99 % de las recurrencias y 71% a tipos alélicos similares, en 2 episodios,
con un intervalo de tiempo entre 29 y 343 días, lo cual es concordante con el
periodo de tiempo que se utilizó para evaluar las recurrencias en este estudio,
identificándose 41,7% de recurrencias por haplotipos idénticos o relapsos y 48,9%
de recurrencias por haplotipos diferentes o reinfecciones. Otros estudios como el
de Hanf et al., 2009 en Guyana Francesa, utilizaron rangos de tiempo e indicaron
que el incremento de los relapsos se produjo en los primeros 3 meses después del
episodio inicial, posterior a esto, la mayoría de ataques secundarios fueron
producto de reinfecciones. En regiones endémicas más próximas, como Colombia,
estudios realizados por Álvarez et al., 2006 también indicaron que el tiempo en
que se presentan las recurrencias (relapsos), en pacientes sometidos a diferentes
esquemas de tratamiento, fluctúan entre 53 a 129 días después del episodio
inicial. Estos reportes fueron concordantes con los relapsos que muestra una cepa
tropical y al igual que en este estudio, la mayor frecuencia de recurrencias por
haplotipos idénticos o relapsos se produjeron durante los primeros 3 meses
después de la infección primaria, posterior a este periodo se incrementaron las
frecuencias de recurrencias por haplotipos diferentes o reinfecciones.
Las recurrencias por reinfecciones son comunes en regiones endémicas ya
que los individuos permanecen en la misma zona todo el tiempo y es más
probable que pueden adquirir rápidamente una nueva infección, sin embargo estas
deducciones deben interpretarse con cuidado ya que estos individuos también
pueden tener un relapso causado por activación de hipnozoitos heterólogos. Al
respecto Imwong et al., 2007 reportaron infecciones recurrentes, por parásitos no
relacionados con la infección primaria, en 78% de individuos provenientes de

Tailandia y 63% en India, en estos casos la reactivación de hipnozoitos
heterólogos podría ser la principal causa de las recurrencias. Concordante con
estos datos, la OMS en el 2006 informó frecuencias bajas de relapsos que varían
desde 11% en la India hasta 51% en Afganistán, con valores intermedios de 30%
en Indonesia.
En este estudio, las altas frecuencias de reinfecciones y relapsos pueden
ser relacionados con un posible incremento de la inmunidad adquirida y por
consiguiente la resistencia a las complicaciones de la enfermedad e incremento en
el número de portadores asintomáticos. Esta forma de la malaria fue identificada
por Loyola, 1991 quien mencionó, que en regiones endémicas, tras prolongados
periodos de infección, las personas adquieren un alto grado de inmunidad y la
malaria asintomática es frecuente. Además Branch et al., 2005 señalan que
durante las recurrencias la respuesta inmune es más rápida y amplificada.
Finalmente, al igual que Alyson et al., 1996 concordamos que el manejo y
control de P. vivax es complicado por su habilidad de causar relapsos y por las
continuas reinfecciones en zonas endémicas, por lo cual es necesario una
detección e identificación oportuna del tipo de infección malárica.
7.4. MECANISMOS DE RESISTENCIA
Reportes como el de Baird et al., 1991; Phillips et al., 1996 y Brega et. al.,
2005 relacionaron la CQR con el polimorfismo de nucleótidos únicos o SNPs, en el
gen de resistencia pvmdr1 de P. vivax. En este estudio, por el análisis de
secuenciamiento, se identificó 16% (4/25) de muestras infectadas con un alelo
doble mutante, con mutaciones no sinónimas en los 2 codones Y976F-F1076L
relacionados con resistencia. Estas mutaciones son observadas también por
Suwanarusk et al., 2007 en Indonesia y Tailandia, lugares donde se realizaron
estudios de susceptibilidad a la cloroquina, in vitro. Frecuencias más altas son

reportadas por Brega et. al., 2005, encontrando 26% (6/23) de aislados, 4 desde
Tailandia y 2 desde Indonesia con el alelo Y976F y 61% (14/23) de aislados con el
alelo F1076L, 5 desde Tailandia, 4 de los cuales también tuvieron la mutación en
el codón Y976F, 4 desde Azerbaiyán, 3 desde Turquía y 2 desde Indonesia, los
cuales también tuvieron la mutación en el codón Y976F. Otros estudios como el de
Dua et al., 1996, efectuados en poblaciones que presentan CQR definido como la
India, Papúa Nueva Guinea y Myanmar muestran un incremento de estas
mutaciones, por lo que se podría especular que estos 2 SNPs serían marcadores
moleculares tempranos que indiquen CQR en P. vivax.
Las muestras para el secuenciamiento fueron obtenidas al azar, debido
principalmente a que las mismas no fueron colectadas a través de un seguimiento
estricto para determinar la eficacia o falla del tratamiento, sin embargo todos los
individuos positivos inicialmente a microscopía (primera evaluación), que
representan el 89.1% de la población analizada, fueron sometidos a tratamiento
por 3 días con cloroquina. Al respecto, varios estudios de sensibilidad a este
fármaco, como los efectuados por Trenton et al., 2003 en la región amazónica y en
la costa del pacífico de Perú, recomiendan un seguimiento de 28 días para
determinar la eficacia de cloroquina in vivo.
Otros estudios en Sudamérica como los efectuados en Colombia y Brasil,
por Soto et al., 2001 y Alecrim, 1999 también indican periodos similares de
seguimiento. Incluso estudios efectuados en Madagascar por Barnadas et al.,
2008, indican que los pacientes deben someterse a un régimen de seguimientos
acorde a los lineamientos recomendados por la OMS en el 2001. A pesar de las
limitaciones, en este estudio, cabe resaltar lo mencionado por Brega et al., 2004,
quienes señalan que la identificación de genes potencialmente involucrados en la
resistencia a drogas y sus variaciones moleculares, podrían proveer un esquema
para estudiar la incidencia y propagación de cepas de P. vivax resistentes a
drogas.

Por otra parte un aporte importante de este estudio fue el uso del software
U-Melt, para el análisis in sílico del HRM, el cual permitió establecer diferencias
entre los SNPs de los alelos mutantes con los alelos wild type. Estos datos son
una información útil para poder discriminar fácilmente la presencia de
determinados SNPs que originen mutaciones no sinónimas en la secuencia de
aminoácidos. Ensayos similares utilizando la PCR en Tiempo Real para analizar
HRM fueron implementados por Alonso et al., 2010, la cual permitió diferenciar
fácilmente distintos alelos de un gen, con presencia o ausencia de SNPs en
determinados codones.
En relación a la cuantificación relativa, Brega et al., 2005 indicaron que la
sobreexpresión del gen pvmdr1 de P. vivax ha sido asociada con resistencia a
drogas como cloroquina, estudios similares en P. falciparum como el efectuado
por Ferreira et al., 2006, evaluaron el número de copias del gen pfmdr1 utilizando
como gen endógeno a la Actina, y mostraron un incremento de 3 veces respecto a
la cepa de referencia (cepa 3D7). En este estudio el análisis del número de copias
del gen pvmdr1, utilizando como gen endógeno a la Aldolasa, no mostró ningún
incremento significativo tanto en las muestras de individuos con relapso así como
en los que presentaron reinfección.
La importancia de realizar un diagnóstico preciso reside primeramente en
determinar la especie de Plasmodium que causa la enfermedad, así mismo es
necesario establecer la recurrencia de la infección, a fin de que el paciente reciba
el tratamiento adecuado. Además Bray et al., 2005 mencionaron la importancia de
la variabilidad genética de P. vivax en el flujo de genes como los que confieren
resistencia, Brega et al., 2005 también mencionaron que la resistencia a
cloroquina podría deberse a que la diversidad genética de P. vivax tiende a variar
según el área geográfica, por lo tanto se considera necesario efectuar estudios
similares en diferentes regiones geográficas endémicas para P. vivax.

VIII. CONCLUSIONES
En individuos asintomáticos provenientes de una región endémica como
Mazán, el diagnóstico mediante Nested PCR, a partir de muestras
secuenciales evaluadas durante un año de seguimiento activo, permitió
detectar altas recurrencias de la infección por P. vivax.
La genotipificación, en base al gen pvmsp3-α, utilizando sólo el Nested
PCR no fue suficiente para establecer una clara variabilidad, sin embargo la
posterior digestión de los productos de amplificación, combinando enzimas
como Alu I y Hha I, permitió identificar diversos haplotipos. Los valores de
COI>1 y PLD (He) fueron concordantes con una alta variabilidad genética,
logrando identificarse infecciones mixtas por haplotipos policlonales e
infecciones simples por haplotipos monoclonales.
En individuos con infección recurrente por P. vivax, se identificó que la
recurrencia se debía a una reinfección cuando se observaron haplotipos
diferentes a la infección primaria y a un relapso cuando se observaron
haplotipos idénticos. Además concordante con el comportamiento de cepas
tropicales, los relapsos fueron más frecuentes que las reinfecciones durante
los primeros 90 días después de la infección primaria, posterior a este
periodo las reinfecciones fueron más frecuentes.
Los SNPs dobles mutantes, causantes de mutaciones no sinónimas, en los
codones Y976F y F1076L del gen pvmdr1 de P. vivax, previamente
relacionados con resistencia, fueron identificados solo en 4 individuos con
relapso. Además no se mostró ninguna variación cuantitativa en el gen
pvmdr1, que sea significativa e indique resistencia, tanto en las muestras de
individuos con relapso y con reinfección.

IX. RECOMENDACIONES
En regiones endémicas de baja transmisión de la malaria, se debe tomar en
cuenta la persistencia de infecciones asintomáticas y el incremento de las
recurrencias principalmente debidas a reinfecciones o relapsos.
Para una mejor caracterización de la variabilidad genética de P. vivax, se
requiere analizar un gran número de muestras, así mismo también es
necesario utilizar, en conjunto, otros marcadores genéticos para proveer un
mayor análisis de variabilidad.
Para estudios de evaluación de la variabilidad genética de P. vivax mediante
PCR-RFLP, es necesario utilizar por lo menos 2 enzimas de restricción.
Debido a que infecciones inicialmente clasificadas como simples, basadas en
los resultados de digestión con una sola enzima, posteriormente pueden ser
identificadas como infecciones mixtas con una segunda enzima e incluso la
utilización de una tercera enzima proporcionaría mayor información de la
diversidad genotípica.
Para determinar los patrones de recurrencia, en futuras investigaciones, es
necesario evaluar como mínimo 2 muestras secuenciales por paciente, con
seguimiento terapéutico durante los primeros 28 días hasta 3 meses post-
infección primaria. Se recomienda evaluar los relapsos y las reinfecciones
después de un mes de la infección primaria. La resistencia por el contrario
debe ser evaluada antes de este periodo, por lo general se recomienda
seguimientos a los 0, 7, 14 y 28 días.
En estudios de enfermedades transmisibles como la malaria por P. vivax, no
solo es necesario la utilización de marcadores genéticos de resistencia para
discriminar la falla a tratamiento o relapso desde hipnozoitos homólogos, sino
también aplicar otros procedimientos de inmunoserología, que podrían ayudar
a distinguir entre nuevas infecciones o relapsos.

X. BIBLIOGRAFÍA
1. Alecrim G. C. 2000. Estudio clínico, resistencia e polimorfismo parasitario
na malaria pelo Plasmodium vivax, em Manaus-AM. Ph.D. thesis.
Universidad de Brasília, Brasilia, Brazil.
2. Alonso M., Navarro Y., Barletta F., Martinez M., Gotuzzo L., Bouza E.,
Garcia D., 2010. A novel method for the rapid and prospective identification
of Beijing Mycobacterium tuberculosis strains by high-resolution melting
analysis. University Cayetano Heredia Lima-Peru. Clin Microbiol Infect. 1:
32-35.
3. Alvarez G. 2006. Efficacy of three choroquine-primaquine regimens for
treatment of Plasmodium vivax malaria in Colombia. Am J Trop, Med Hyg.
75(4): 605-609.
4. Alves F. P., Durlacher R. R., Menezes M. J., Krieger H., Silva L. H.,
Camargo E. P., 2002. High prevalence of asymptomatic Plasmodium vivax
and Plasmodium falciparum infections in native Amazonian populations. Am
J Trop. Med Hyg. 66:641-648.
5. Alyson A., Craig and Kevin C. K., 1996. Molecular Analysis of Strains of
Plasmodium vivax from Paired Primary and Relapse Infections. The Journal
of Infectious Diseases. Tropical Disease Unit. Toronto Hospital. 200
Elizabeth St., Toronto, Canada, 174:373-9.
6. Andrade B. B., Reis-Filho A., Souza-Neto S. M., Clarêncio J., Camargo L.
M., Barral A., Barral-Netto M., 2010. Severe Plasmodium vivax malaria
exhibits marked inflammatory imbalance. Fundação Oswaldo Cruz – Brazil.
Malaria Journal. 9:13.
7. Aramburu G. J., Ramal A. C., Witzig R. 1999. Malaria reemergence in the
Peruvian Amazon region. Emerg Infect Dis. 5:209-215.
8. Ayala E., Lescano A. G., Gilman R. H., Calderon M., Pinedo V., Terry H.,
Cabrera L., Vinetz J. 2005. Polymerase Chain Reacction and Molecular
Genotyping To Monitor Parasitological Response To Anti-Malarial

Chemotherapy in the Peruvian Amazon. Am J Trop Med. Hyg. 74(4): 546-
553.
9. Babiker H. A., Pringle S. J., Abdel-Muhsin A., Mackinnon M., Hunt P.,
Walliker D., 2001. High-level chloroquine resistance in Sudanese isolates of
Plasmodium falciparum is associated with mutations in the chloroquine
resistance transporter gene pfcrt and the multidrug resistance gene pfmdr1.
J Infect Dis. 183:1535–1538.
10. Baird J. K., Basri H., Subianto B., Patchen L. C., Hoffman S. L., 1991.
Resistance to chloroquine by Plasmodium vivax in Irian Jaya, Indonesia. Am
J Trop Med Hyg. 44:547–552.
11. Baird J. K. 2004. Chloroquine resistance in Plasmodium vivax. Antimicrob
Agents Chemother. 48:4075–4083.
12. Baird J. K., 2008. Real - world therapies and the problem of vivax malaria. N
Engl J Med. 359: 2601–2603.
13. Barker R. H., Banchongaksorn T., Courval J. M., Suwonkerd W.,
Rimwungtragoon K., Wirth D. F. 1992. A simple method to detect
Plasmodium falciparum directly from blood samples using the polymerase
chain reaction. Am J Trop Med Hyg. 46:416-426.
14. Barnadas C., Ratsimbasoa A., Tichit M., Bouchier C., Jahevitra M., Picot S.,
Menard D. 2008. Plasmodium vivax Resistance to Chloroquine in
Madagascar: Clinical Efficacy and Polymorphisms in pvmdr1 and pvcrt-o
Genes. American Society for Microbiology. Parasitology and Tropical
Medicine.52: 4233–4240
15. Branch O, Casapia W. M., Gamboa D. V., Hernandez J. N., Alava F.F.,
Roncal N., Alvarez E., Perez E. J., Gotuzzo E., 2005. Clustered local
transmission and asymptomatic Plasmodium falciparum and Plasmodium
vivax malaria infections in a recently emerged, hypoendemic Peruvian
Amazon community. Malaria Journal 4:27.
16. Bray P., Deed S., Fox E., Kalkanidis M., Mungthin M., Deady L. W., Tilley L.
2005. Primaquine Synergises the Activity of Chloroquine Against

Chloroquine-Resistant P. falciparum. Biochemical Pharmacology. 70: 1158-
1166.
17. Brega S., Meslin B., Monbrison F., Severini C., Gradoni L., Udomsangpetch
R., Sutanto I., Peyron F., Picot S., 2005. Identification of the Plasmodium
vivax mdr-like gene (pvmdr1) and analysis of single-nucleotide
polymorphisms among isolates from different areas of endemicity. J Infect
Dis. 191:272–277.
18. Bruce M. C., Galinski M. R., Barnwell J. W., Snounou G., Day K. P., 1999.
Polymorphism at the merozoite surface protein-3alpha locus of Plasmodium
vivax: global and local diversity. Am J Trop Med Hyg. 61:518-525.
19. Carlton J., 2003. The Plasmodium vivax genome sequencing project.
Trends Parasitol. 19: 227–231.
20. Chen N., Auliff A., Rieckmann K., Gatton M., Cheng Q., 2007. Relapses of
Plasmodium vivax infection result from clonal hypnozoites activated at
predetermined intervals. J Infect Dis. 195:934-941.
21. Cole-Tobian J. L., Biasor M., King C. L., 2005. High complexity of
Plasmodium vivax infections in Papua New Guinean children. Am J Trop
Med Hyg. 73:626-633.
22. Cogswell F. B., 1992. The Hypnozoite and Relapse in Primate Malaria.
Tulane Regional Primate Research Center. Louisiana- USA. Clinical
microbiology reviews. 1: 26-35.
23. Cristiano F. A., Pérez M. A., Nicholls R. S., Guerra A. P. 2008.
Polymorphism in the Plasmodium vivax msp 3α gene in field samples from
Tierralta, Instituto Nacional de Salud, Bogotá, D.C., Colombia. Mem Inst
Oswaldo Cruz, Rio de Janeiro. 103(5): 493-496.
24. Cui L., Mascorro C. N., Fan Q., Rzomp K. A., Khuntirat B., Zhou G., Chen
H., Yan G., Sattabongkot J., 2003. Genetic diversity and multiple infections
of Plasmodium vivax malaria in Western Thailand. Am J Trop Med Hyg.
68:613-619.
25. Dirección General de epidemiología. Sistema de vigilancia epidemiológica
DRE SVE-2007: Etapas de la vigilancia epidemiológica.

26. Dua V. K., Kar P. K., Sharma V. P., 1996. Chloroquine resistant
Plasmodium vivax malaria in India. Trop Med Int Health. 1:816–819.
27. Echeverri D., Barreto D. K., Osorio L., Cortés A., Martínez E. 2011. Malaria
por Plasmodium vivax transmitida por transfusión de un donante
asintomático a un recién nacido prematuro. Universitario del Valle - Cali
Colombia. Biomédica. 32:8-12
28. Ferreira I. D., Rosario V., Cravo P., 2006. Real-time quantitative PCR with
SYBR Green I detection for estimating copy numbers of nine drug
resistance candidate genes in Plasmodium falciparum. Malaria Journal. 5:1.
29. Fidock D. A., Nomura T., Talley A. K., Cooper R. A., Dzekunov S. M., Ferdig
M. T., Ursos L. M., Roepe P. D., Wellems T. E., 2000. Mutations in the P.
falciparum digestive vacuole transmembrane protein PfCRT and evidence
for their role in chloroquine resistance. Mol Cell. 6:861-871.
30. Garg M., Gopinathan N., Bodhe P., Kshirsagar N. A., 1995. Vivax malaria
resistant to chloroquine: case reports from Bombay. Trans R Soc Trop Med
Hyg. 89: 656–657.
31. Gillet P., Bosselaers K., Cnops L., Bottieau E., Van Esbroeck M., Jacobs J.
2009. Evaluation of the SD FK70 Malaria Ag Plasmodium vivax Rapid
Diagnostic Test in a non-endemic setting. Malaria Journal. 8: 129.
32. Hanf M., Stéphani A., Basurko C., Nacher M., Carme B., 2009.
Determination of the Plasmodium vivax relapse pattern in Camopi, French
Guiana. Malaria Journal. 8:278.
33. Hay S. I., Guerra C. A., Tatem A. J., Noor A. M., Snow R. W., 2004. The
global distribution and population at risk of malaria: past, present, and
future. Lancet Infect Dis. 4:327–36.
34. Imwong M., Pukrittayakamee S., Gruner A. C., Renia L., Letourneur F.,
Looareesuwan S., White N. J., Snounou G., 2005. Practical PCR
genotyping protocols for Plasmodium vivax using Pvcs and Pvmsp1. Malar
J. 4:20.
35. Imwong M., Snounou G., Pukrittayakamee S., Tanomsing N., Kim J. R.,
Nandy A., Guthmann J. P., Nosten F., Carlton J., Looareesuwan S., Nair S.,

Sudimack D., Day N. P., Anderson T. J., White N. J., 2007. Relapses of
Plasmodium vivax infection usually result from activation of heterologous
hypnozoites. J Infect Dis. 195:927-933.
36. Jonhnston S. P., Pieniazek N. J., Xayavong M. V., Slemenda S. B., Wilkins
P. P., daSilva A. J. 2006. PCR as a confirmatory technique for laboratory
diagnosis of malaria. J Clinical Microb. 44:1087-1089.
37. Kain K. C., Brown A. E., Mirabelli L., Webster H. K. 1993. Detection of
Plasmodium vivax by Polymerase Chain Reaction in a Field Study. Journal
of Infectious Diseases. 168: 1323-1326.
38. Koram K.1993. Socioeconomic and environmental risk factors in severe
malaria: a case control study. Unpublished phd thesis. London School of
Hygiene and Tropical Medicine. University of London.
39. Kosek M., Yori P. P., Gilman R. H., Calderon M., Zimic M., Chuquiyauri R.,
Jeri C., Pinedo-Cancino V., Matthias M. A., Llanos-Cuentas A., Vinetz J. M.
2012. High Degree of Plasmodium vivax Diversity in the Peruvian Amazon
Demonstrated by Tandem Repeat Polymorphism Analysis. Am J Trop Med
Hyg. 86(4): 580–586.
40. Loyola E.1991. Seroepidemiologia del Paludismo. Técnica del INDRE. 1:77-
95
41. McKenzie F.E., Ferreira M.U., Baird J.K., Snounou G., Bossert W.H., 2001.
Meiotic recombination, cross-reactivity and persistence in Plasmodium
falciparum. National Institutes of Helth-Evolution. 55(7): 1299–1307.
42. Mallika I., Georges S., Sasithon P., Naowarat T., Jung R. K., Amitab N.,
Jean-Paul G., Francois N., Jane C., Sornchai L., Shalini N., Daniel S.,
Nicholas P. J., Timothy J. C., Anderson J., W., 2007. Relapses of
Plasmodium vivax Infection Usually Result from Activation of Heterologous
Hypnozoites. The Journal of Infectious Diseases. 195:927–933.
43. Manguin S., Roberts D. R., Peyton E. L., Fernandez-Salas I., Barreto M.,
Fernandez L. R., Elgueta S. R., Martinez G. R., Rodriguez M. H., 1995.
Biochemical systematics and population genetic structure of Anopheles

pseudopunctipennis, vector of malaria in Central and South America. Am J
Trop Med Hyg. 53:362-377.
44. Mascorro C. N., Zhao K., Khuntirat B., Sattabongkot J., Yan G., Escalante
A. A., Cui L., 2005. Molecular evolution and intragenic recombination of the
merozoite surface protein MSP-3alpha from the malaria parasite
Plasmodium vivax in Thailand. Parasitology. 131:25-35
45. Mayxay M., Pukritrayakamee S., Chotivanich K., Imwong M., Looareesuwan
S., White N. J. 2001. Identification of cryptic coinfection with Plasmodium
falciparum in patients presenting with vivax malaria. Am J Trop Med Hyg.
65: 588-592.
46. Mendis K., Sina B., Marchesini P., Carter R., 2001. The neglected burden of
Plasmodium vivax malaria. Am J Trop Med Hyg. 64:97-106.
47. Ministerio de Salud. 2009. Dirección General de Epidemiología. Red
Nacional de Control Epidemiológico. RENACE-SE 20.
48. Ministerio de Salud. 2012. Dirección General de Epidemiología. Red
Nacional de Epidemiologia. Dirección Regional de Salud-Loreto SE 29.
49. Mwangi T. W., Mohammed M., Dayo H., Snow R. W., Marsh K. 2005.
Clinical algorithms for malaria diagnosis lack utility among people of
different age groups. Trop med Int Health. 10:530-536.
50. Nomura T., Carlton J. M. R., Baird K., 2001. Evidence for different
mechanisms of chloroquine resistance in 2 Plasmodium species that cause
human malaria. J Infect Dis. 183:1653–1661.
51. Ord R., Olley S., Tami A., Sutherland C. J., 2005. High sequence diversity
and evidence of balancing selection in the Pvmsp3alpha gene of
Plasmodium vivax in the Venezuelan Amazon. Mol Biochem Parasitol. 144:
86–93.
52. Orjuela P. S., Simoes F., Machado A., Francis Y. C., Farias M. R., Gracias
M. C., del Portillo H. A., 2009a. Analysis of Single-Nucleotide
Polymorphisms in the crt-o and mdr1Genes of Plasmodium vivax among
Chloroquine-Resistant Isolates from the Brazilian Amazon Region,
Antimicrobial agents and Chemotherapy. 1: 3561–3564.

53. Orjuela P. S., da Silva N., da Silva M., Urbano F. M., 2009b. Recurrent
Parasitemias and Population Dynamics of Plasmodium vivax
Polymorphisms in Rural Amazonia. Department of Health Sciences, Federal
University of Acre, Rio Branco, Brazil. Am J Trop Med Hyg. 81: 961–968.
54. Organización Panamericana de la Salud. 2006. Plan estratégico regional
contra la malaria en las Américas 2006-2010. Washington, D. C.
55. Ousmane A. K., Sango H. O., Maiga M. S., Mounkoro M., Sangare L., Keita
N., Fane S., Mahamadou I., Traore K., Maiga A. S., Maaiga M. K., Diallo A.,
Krogstad D. J., 2005. Effect of Seasonality on the Prevalence of the Four
Malaria Parasite Species In Northern Mali. Abstrat Book. American Society
of Tropical Medicine and Hygiene 54th Annual Meeting. 73(6): 283.
56. Perkins M. D., Bell D. R., 2008. Working without a blindfold: the critical role
of diagnostic in malaria control. Malaria Journal. 7:S5.
57. Phillips E. J., Keystone J. S., Kain K. C., 1996. Failure of combined
chloroquine and high-dose primaquine therapy for Plasmodium vivax
malaria acquired in Guyana, South America. Clin Infect Dis. 23: 1171–1173.
58. Prajapati S. K., Joshi H., Valecha N., 2010. Plasmodium vivax merozoite
surface protein-3α: a high-resolution marker for genetic diversity studies. J
Vector Borne Dis. 47: 85–90.
59. Ratcliff A., Siswantoro H., Kenangalem E., Wuwung M., Brockman A., 2007
Therapeutic response of multidrug-resistant Plasmodium falciparum and P.
vivax to chloroquine and sulfadoxine-pyrimethamine in southern Papua,
Indonesia. Trans R Soc Trop Med Hyg. 101: 351–359.
60. Ribeiro R. S., Ladeira L., Rezende A. M., Fernandes C. J., Carvalho L. H.,
Ferreira C., 2011. Analysis of the genetic variability of PvMSP-3α among
Plasmodium vivax in Brazilian field isolates. Mem Inst Oswaldo Cruz, Rio de
Janeiro. 6: 27-33.
61. Rodulfo H., De Donato M., Quijada I., Peña A. 2007. High prevalence of
malaria infection in Amazonas state, Venezuela. Rev Inst Med Trop. 49: 79-
85.

62. Roshanravan B., Kari E., Gilman R. H., Cabrera L., Lee E., Metcalfe J.,
Calderon M., Lescano A. G., Montenegro S. H., Calampa C., Vinetz J. M.,
2003. Endemic malaria in the Peruvian Amazon region of Iquitos. Am J Trop
Med Hyg. 69:45-52.
63. Sattabongkot J., Tsuboi T., Zollner G. E., Sirichaisinthop J., Cui L., 2004.
Plasmodium vivax transmission: chances for control? Trends Parasitol.
20:192–198.
64. Sawyer D. 1993. Economic and Social Consequences of Malaria in New
Colonization Projects in Brazil. Soc Sci Med. 37(9): 1131-1136.
65. Scopel K. K., Fontes C. J., Nunes A. C., Horta M. F., Braga E. M. 2004. Low
sensitivity of nested PCR using Plasmodium DNA extracted from stained
thick blood smears: an epidemiological retrospective study among subjects
with low parasitaemia in an endemic area of the Brazilian Amazon region.
Malaria Journal. 31:3-8.
66. Singh B., Bobogare A., Cox-Singh J., Snounou G., Abdullah M. S., Rahman
H. A. 1999. A genus- and species-specific nested polymerase chain reaction
malaria detection assay for epidemiologic studies. Am J Trop Med Hyg. 60:
687-692.
67. Singh B., Lee K. S., Matusop A., Radhakrihsnan A., Shamsul S. G., Cox-
Singh J., Thomas A., Conway D., 2004. A large focus of naturally acquired
Plasmodium knowlesi infections in human beings. Lancet. 363: 1017-1024.
68. Siripoon N., Snounou G., Yamogkul P., Na-Bangchang K., Thaithong S.
2002. Cryptic Plasmodium falciparum parasites in clinical P. vivax blood
samples from Thailand. Trans R Soc Trop Med Hyg. 96:70-71.
69. Snounou G., Viriyakosol S., Jarra W., Thaithong S., Brown K. N. 1993.
Identification of the four human malaria parasite species in field samples by
the polymerase chain reaction and detection of a high prevalence of mixed
infections. Mol Biochem Parasitol. 58:283-292.
70. Sojo M. M., Cáceres J. L., Pizzo N. N., Rojas J., Maldonado A., Pulgar R.
N., Campos E., Terán E., Pérez A., 2008. Malaria recurrente a Plasmodium
vivax. Municipio Cajigal, Estado Sucre, Venezuela. Rev Biomed. p: 3-15.

71. Soto J., Toledo J., Gutierrez P., Luz M., Llinas N., Cedeño N., Dunne M.,
Berman J. 2001. Plasmodium vivax clinically resistant to chloroquine in
Colombia. Am J Trop Med Hyg. 65: 90–93
72. Steenkeste N., Incardona S., Chy S., Duval L., Ekala M. T., Lim P., Hewitt
S., Sochantha T., Socheat D., Rogier C., Mercereau-Puijalon O., Fnadeur
T., Ariey F. 2009. Towards high-throughput molecular detection of
Plasmodium: New approaches and molecular markers. Malaria Journal.
8:86
73. Sutton P. L., Neyra V., Hernandez J. N., Branch O. H., 2009. Plasmodium
falciparum and Plasmodium vivax Infections in the Peruvian Amazon:
Propagation of Complex, Multiple Allele-Type Infections without Super-
Infection. Am J Trop Med Hyg. 81(6): 950–960.
74. Suwanarusk R., Russell B., Chavchich M., Chalfein F., Kenangalem E.,
Kosaisavee V., Prasetyorini B., Piera K. A., Barends M., Tjitra E., Nosten F.,
Cheng Q., Price R. N., 2007. Chloroquine resistant Plasmodium vivax: in
vitro characterization and association with molecular polymorphisms. PLoS
ONE. 2:9.
75. Tangpukdee N., Duangdee C., Wilairatana P., Krudsood S. 2009. Malaria
Diagnosis. A brief Review. Korean J Parasitol. 47:93-102.
76. Tjitra E., Baker J., Suprianto S., Cheng Q., Anstey N. M., 2002. Therapeutic
efficacies of artesunate-sulfadoxine-pyrimethamine and chloroquine-
sulfadoxinepyrimethamine in vivax malaria pilot studies: relationship to
Plasmodium vivax dhfr mutations. Antimicrob. Agents Chemother. 46:
3947–3953.
77. Trenton K., Ruebush I., Zegarra J., Cairo J., Ellen M., Andersen, Green M.,
Dylan R. Pillai, Marquiño W., Huilca M., Arevalo E., Garcia C., Solary L.,
Kain K. C. 2003. Chloroquine-resistant Plasmodium vivax malaria in Peru.
U. S. Naval Medical Research Center Detachment, Lima, Peru. Am J Trop
Med Hyg. 69(5): 548–552.

78. Vargas J., Elgegren J., San Miguel A., Cardoso R. 2003. Malaria en una
Población Urbano Marginal de Iquitos. Revista Peruana de Epidemiología
11(1).
79. Veron V., Legrand E., Yrinesi J., Volney B., Simon S., Carme B., 2009.
Genetic diversity of msp3alpha and msp1_b5 markers of Plasmodium vivax
in French Guiana. Malar J. 8:40.
80. Volman S., Sabeti P. C., Harti D. L., Birren B., Lander E., Wirth D. F., 2005.
A Haplotype Map For Plasmodium falciparum. Abstrat Book. American
Society of Tropical Medicine and Hygiene 54th Annual Meeting 73(6).
81. White N. J., 2008. Plasmodium knowlesi: The Fifth Human Malaria Parasite
Medicine, Mahidol University, Bangkok-Thailand. Clinical Infectious
Diseases. 46:172–3.
82. Wongsrichanalai C., Barcus M. J., Muth S., Sutamihardja A., Wernsdorfer
W.H. 2007. A review of malaria diagnostic tools: microscopy and Rapid
Diagnostic Test (RDT). Am J Trop Med Hyg. 77: 119-127.
83. World Health Organization 1991. Basic Malaria Microscopy. Geneva: WHO
Press.
84. World Health Organization 2005. UNICEF World malaria report. Geneva:
WHO Press.
85. World Health Organization 2006. Who guidelines for the treatment of
malaria. Geneva: WHO Press.
86. World Health Organization 2007. Global Paludismo Programme. Field
application of in vitro assays for the sensitivity of human paludismo
parasites to antipaludismo drugs. Geneva: Who Press.
87. World Health Organization 2008. World malaria report. Geneva: WHO
Press.
88. World Health Organization 2009. Methods for surveillance of antimalarial
drug efficacy. Geneva: WHO Press.
89. Zakeri S., Barjesteh H., Djadid N. D., 2006. Merozoite surface protein-
3alpha is a reliable marker for population genetic analysis of Plasmodium
vivax. Malar J. 5:53.16.
XI. ANEXOS
Fig. A1. Mapa de riesgo de malaria por P. vivax, incidencia acumulada por
1000 habitantes, Loreto 2009
Fig. A2 .Mapa de riesgo de malaria del departamento de Loreto 2012
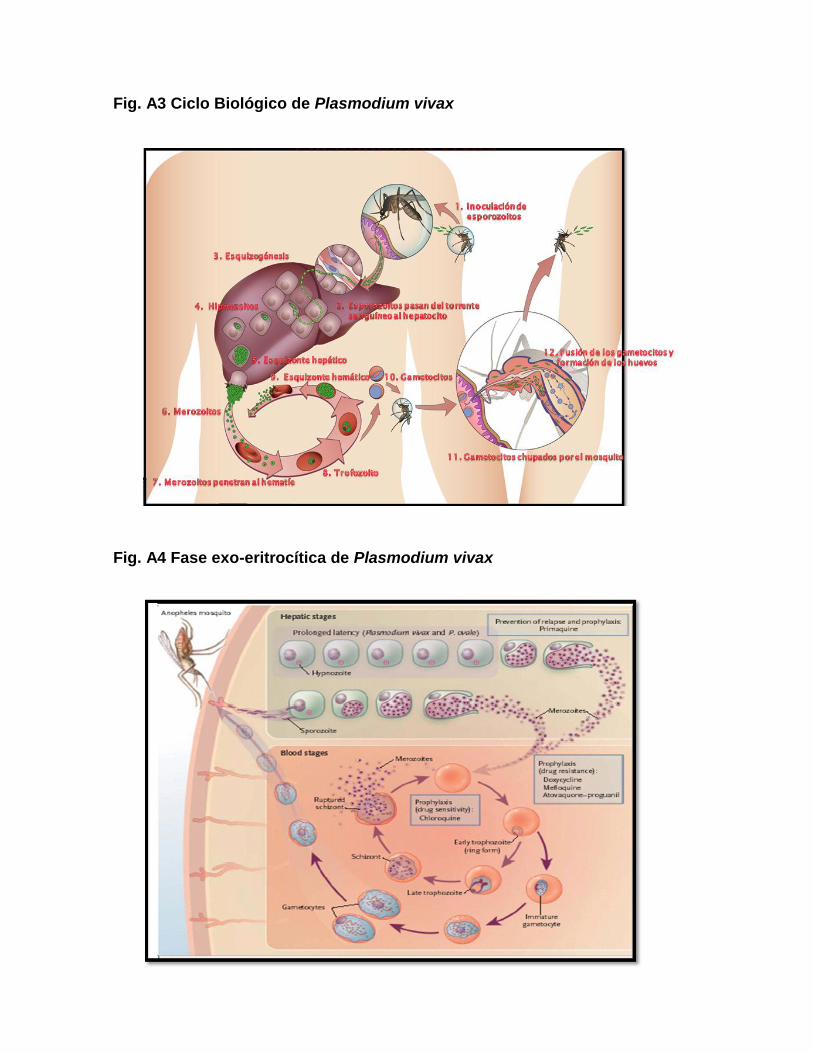
Fig. A3 Ciclo Biológico de Plasmodium vivax
Fig. A4 Fase exo-eritrocítica de Plasmodium vivax
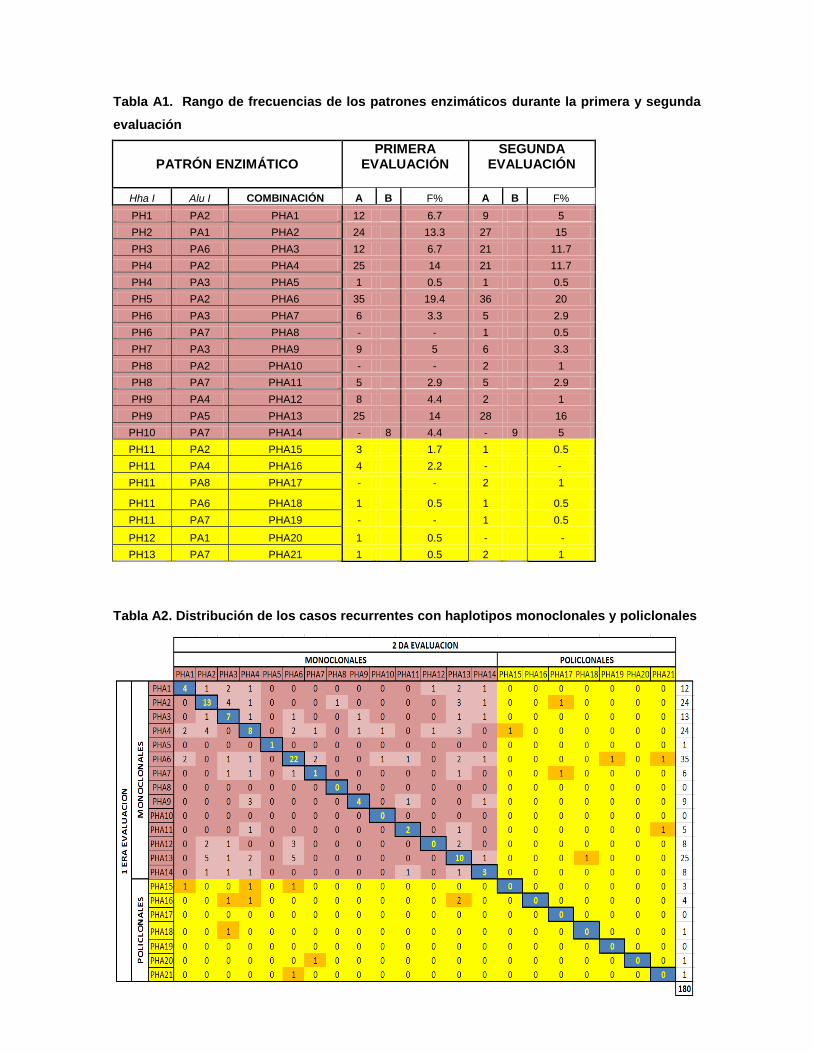
Tabla A1. Rango de frecuencias de los patrones enzimáticos durante la primera y segunda
evaluación
PATRÓN ENZIMÁTICO
PRIMERA EVALUACIÓN
SEGUNDA EVALUACIÓN
Hha I Alu I COMBINACIÓN A B F% A B F%
PH1 PA2 PHA1 12 6.7 9 5
PH2 PA1 PHA2 24 13.3 27 15
PH3 PA6 PHA3 12 6.7 21 11.7
PH4 PA2 PHA4 25 14 21 11.7
PH4 PA3 PHA5 1 0.5 1 0.5
PH5 PA2 PHA6 35 19.4 36 20
PH6 PA3 PHA7 6 3.3 5 2.9
PH6 PA7 PHA8 - - 1 0.5
PH7 PA3 PHA9 9 5 6 3.3
PH8 PA2 PHA10 - - 2 1
PH8 PA7 PHA11 5 2.9 5 2.9
PH9 PA4 PHA12 8 4.4 2 1
PH9 PA5 PHA13 25 14 28 16
PH10 PA7 PHA14 - 8 4.4 - 9 5
PH11 PA2 PHA15 3 1.7 1 0.5
PH11 PA4 PHA16 4 2.2 - -
PH11 PA8 PHA17 - - 2 1
PH11 PA6 PHA18 1 0.5 1 0.5
PH11 PA7 PHA19 - - 1 0.5
PH12 PA1 PHA20 1 0.5 - -
PH13 PA7 PHA21 1 0.5 2 1
Tabla A2. Distribución de los casos recurrentes con haplotipos monoclonales y policlonales
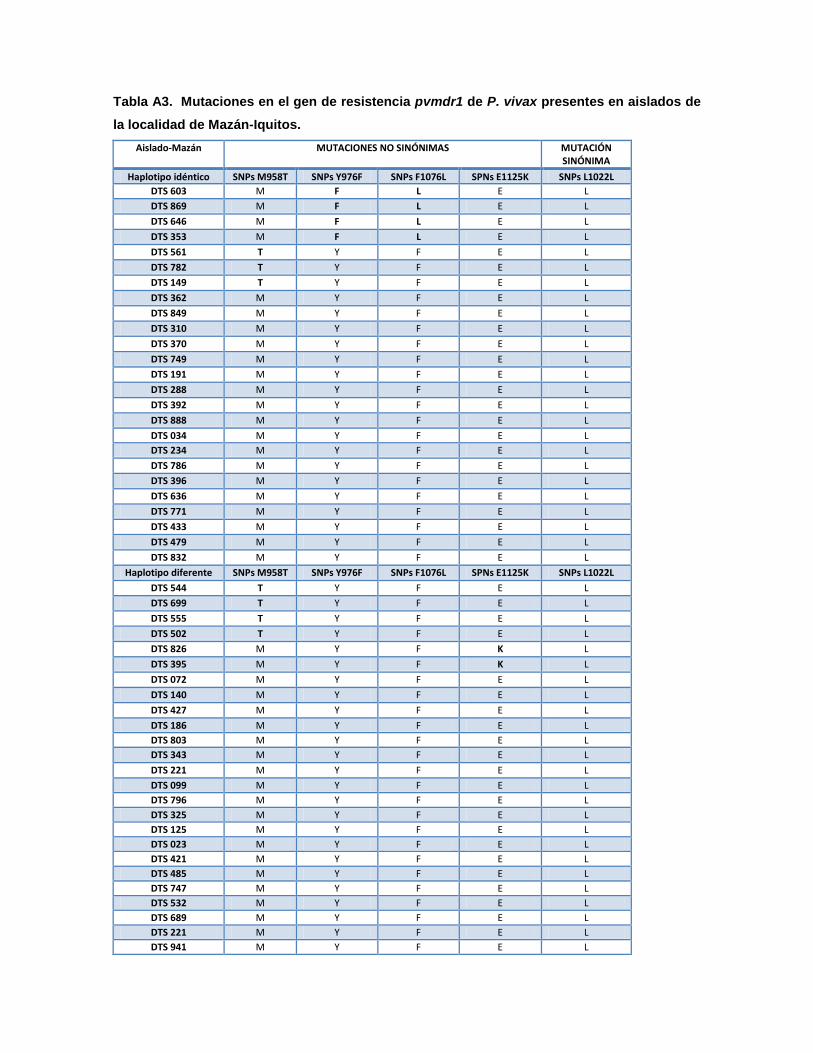
Tabla A3. Mutaciones en el gen de resistencia pvmdr1 de P. vivax presentes en aislados de
la localidad de Mazán-Iquitos.
Aislado-Mazán MUTACIONES NO SINÓNIMAS MUTACIÓN SINÓNIMA
Haplotipo idéntico SNPs M958T SNPs Y976F SNPs F1076L SPNs E1125K SNPs L1022L
DTS 603 M F L E L
DTS 869 M F L E L
DTS 646 M F L E L
DTS 353 M F L E L
DTS 561 T Y F E L
DTS 782 T Y F E L
DTS 149 T Y F E L
DTS 362 M Y F E L
DTS 849 M Y F E L
DTS 310 M Y F E L
DTS 370 M Y F E L
DTS 749 M Y F E L
DTS 191 M Y F E L
DTS 288 M Y F E L
DTS 392 M Y F E L
DTS 888 M Y F E L
DTS 034 M Y F E L
DTS 234 M Y F E L
DTS 786 M Y F E L
DTS 396 M Y F E L
DTS 636 M Y F E L
DTS 771 M Y F E L
DTS 433 M Y F E L
DTS 479 M Y F E L
DTS 832 M Y F E L
Haplotipo diferente SNPs M958T SNPs Y976F SNPs F1076L SPNs E1125K SNPs L1022L
DTS 544 T Y F E L
DTS 699 T Y F E L
DTS 555 T Y F E L
DTS 502 T Y F E L
DTS 826 M Y F K L
DTS 395 M Y F K L
DTS 072 M Y F E L
DTS 140 M Y F E L
DTS 427 M Y F E L
DTS 186 M Y F E L
DTS 803 M Y F E L
DTS 343 M Y F E L
DTS 221 M Y F E L
DTS 099 M Y F E L
DTS 796 M Y F E L
DTS 325 M Y F E L
DTS 125 M Y F E L
DTS 023 M Y F E L
DTS 421 M Y F E L
DTS 485 M Y F E L
DTS 747 M Y F E L
DTS 532 M Y F E L
DTS 689 M Y F E L
DTS 221 M Y F E L
DTS 941 M Y F E L
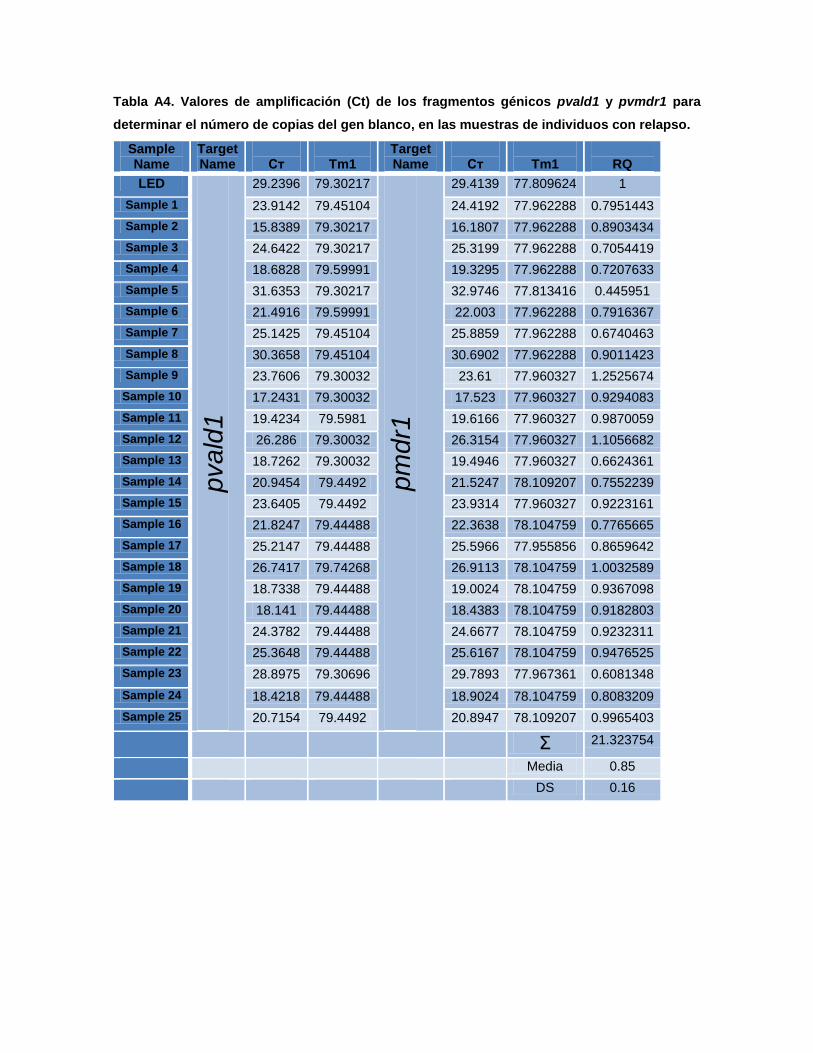
Tabla A4. Valores de amplificación (Ct) de los fragmentos génicos pvald1 y pvmdr1 para
determinar el número de copias del gen blanco, en las muestras de individuos con relapso.
Sample Name
Target Name
Cт
Tm1
Target Name
Cт
Tm1
RQ
LED
pva
ld1
29.2396 79.30217
pm
dr1
29.4139 77.809624 1
Sample 1 23.9142 79.45104 24.4192 77.962288 0.7951443
Sample 2 15.8389 79.30217 16.1807 77.962288 0.8903434
Sample 3 24.6422 79.30217 25.3199 77.962288 0.7054419
Sample 4 18.6828 79.59991 19.3295 77.962288 0.7207633
Sample 5 31.6353 79.30217 32.9746 77.813416 0.445951
Sample 6 21.4916 79.59991 22.003 77.962288 0.7916367
Sample 7 25.1425 79.45104 25.8859 77.962288 0.6740463
Sample 8 30.3658 79.45104 30.6902 77.962288 0.9011423
Sample 9 23.7606 79.30032 23.61 77.960327 1.2525674
Sample 10 17.2431 79.30032 17.523 77.960327 0.9294083
Sample 11 19.4234 79.5981 19.6166 77.960327 0.9870059
Sample 12 26.286 79.30032 26.3154 77.960327 1.1056682
Sample 13 18.7262 79.30032 19.4946 77.960327 0.6624361
Sample 14 20.9454 79.4492 21.5247 78.109207 0.7552239
Sample 15 23.6405 79.4492 23.9314 77.960327 0.9223161
Sample 16 21.8247 79.44488 22.3638 78.104759 0.7765665
Sample 17 25.2147 79.44488 25.5966 77.955856 0.8659642
Sample 18 26.7417 79.74268 26.9113 78.104759 1.0032589
Sample 19 18.7338 79.44488 19.0024 78.104759 0.9367098
Sample 20 18.141 79.44488 18.4383 78.104759 0.9182803
Sample 21 24.3782 79.44488 24.6677 78.104759 0.9232311
Sample 22 25.3648 79.44488 25.6167 78.104759 0.9476525
Sample 23 28.8975 79.30696 29.7893 77.967361 0.6081348
Sample 24 18.4218 79.44488 18.9024 78.104759 0.8083209
Sample 25 20.7154 79.4492 20.8947 78.109207 0.9965403
Σ 21.323754
Media 0.85
DS 0.16
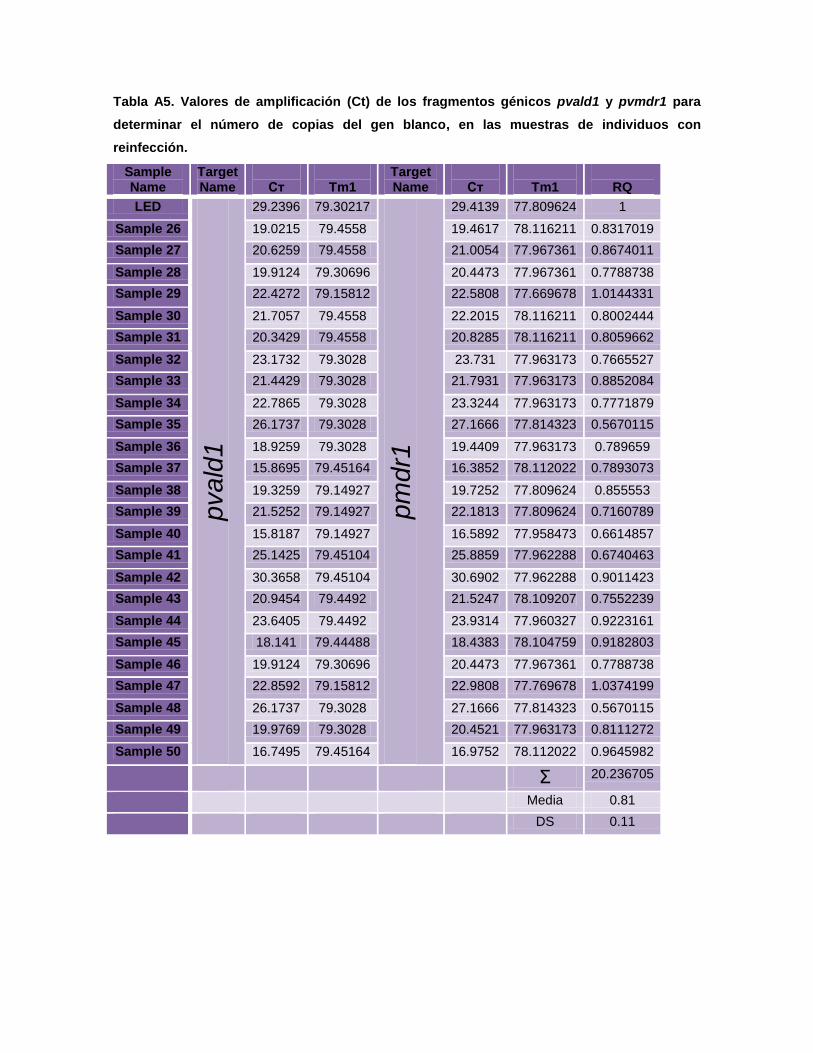
Tabla A5. Valores de amplificación (Ct) de los fragmentos génicos pvald1 y pvmdr1 para
determinar el número de copias del gen blanco, en las muestras de individuos con
reinfección.
Sample Name
Target Name
Cт
Tm1
Target Name
Cт
Tm1
RQ
LED p
va
ld1
29.2396 79.30217
pm
dr1
29.4139 77.809624 1
Sample 26 19.0215 79.4558 19.4617 78.116211 0.8317019
Sample 27 20.6259 79.4558 21.0054 77.967361 0.8674011
Sample 28 19.9124 79.30696 20.4473 77.967361 0.7788738
Sample 29 22.4272 79.15812 22.5808 77.669678 1.0144331
Sample 30 21.7057 79.4558 22.2015 78.116211 0.8002444
Sample 31 20.3429 79.4558 20.8285 78.116211 0.8059662
Sample 32 23.1732 79.3028 23.731 77.963173 0.7665527
Sample 33 21.4429 79.3028 21.7931 77.963173 0.8852084
Sample 34 22.7865 79.3028 23.3244 77.963173 0.7771879
Sample 35 26.1737 79.3028 27.1666 77.814323 0.5670115
Sample 36 18.9259 79.3028 19.4409 77.963173 0.789659
Sample 37 15.8695 79.45164 16.3852 78.112022 0.7893073
Sample 38 19.3259 79.14927 19.7252 77.809624 0.855553
Sample 39 21.5252 79.14927 22.1813 77.809624 0.7160789
Sample 40 15.8187 79.14927 16.5892 77.958473 0.6614857
Sample 41 25.1425 79.45104 25.8859 77.962288 0.6740463
Sample 42 30.3658 79.45104 30.6902 77.962288 0.9011423
Sample 43 20.9454 79.4492 21.5247 78.109207 0.7552239
Sample 44 23.6405 79.4492 23.9314 77.960327 0.9223161
Sample 45 18.141 79.44488 18.4383 78.104759 0.9182803
Sample 46 19.9124 79.30696 20.4473 77.967361 0.7788738
Sample 47 22.8592 79.15812 22.9808 77.769678 1.0374199
Sample 48 26.1737 79.3028 27.1666 77.814323 0.5670115
Sample 49 19.9769 79.3028 20.4521 77.963173 0.8111272
Sample 50 16.7495 79.45164 16.9752 78.112022 0.9645982
Σ 20.236705
Media 0.81
DS 0.11
Tabla A6. Código Genético, tripletes de nucleótidos codificantes de aminoácidos

Formato A1: Consentimiento Informado de Participación de Adultos
CONSENTIMIENTO INFORMADO DE PARTICIPACIÓN-ADULTOS
Título del Proyecto de Investigación: Genotipificación de Plamodium vivax en la Amazonía Peruana (Reservorios
Humanos para la Transmisión de Malaria por Plasmodium vivax en la amazonía Peruana)
Investigador principal: Dr. Joseph Vinetz. Co-Investigadores: Dr. Raul Chuquiyauri, Dr. Robert H. Gilman, Dra. Margaret Koset, Dra. Maritza Calderón
Propósito del Estudio: Este es un estudio conducido por la Universidad Peruana Cayetano Heredia, la Universidad de California en San Diego y el Instituto de Medicina Tropical “Alexander von Humboldt”; y en colaboración con el Ministerio de Salud a través de la Dirección Regional de Salud de Loreto. El objetivo es conocer los perfiles genéticos del Plasmodium vivax causante de malaria y determinar la forma en que se transmite en las zonas de estudio.
Como usted sabe la malaria es una enfermedad muy frecuente en nuestra Amazonia, con importantes trastornos entre las personas que la padecen. Hay dos especies de parásito que causan malaria en Perú: Plasmodium vivax y Plasmodium falciparum. Se esta haciendo un gran esfuerzo para luchar contra ella. Estos esfuerzos en el diagnóstico y prevención requieren de la participación directa de las personas como usted.
Usted esta siendo invitado a tomar parte en este estudio porque su gota gruesa es positiva para Plasmodium vivax. Su participación en este estudio depende completamente de usted.
Procedimientos: Si usted acepta participar en el estudio, un trabajador de salud realizará lo siguiente:
Le hará preguntas básicas sobre usted, su trabajo, su casa y su historial médico y síntomas.
Se tomara una muestra de sangre de 10 ml. Exclusiones: Si usted tiene alguna de las siguientes condiciones no podrá participar en este estudio:
Uso de antibióticos al momento o una semana antes de ingresar al estudio.
Estar recibiendo tratamiento antimalárico ahora o en la última semana.
No haber firmado el consentimiento informado.
Tener malaria por Plasmodium falciparum o malaria mixta.
Tener menos de 5 años de edad. Riesgos e Incomodidades: Su participación en el estudio con lleva un riesgo mínimo. La toma de sangre puede ocasionarle cierta incomodidad. Existe un mínimo riesgo de que aparezca un moretón después de la toma de sangre. Nosotros trataremos de mantener este riesgo tan bajo como sea posible. Este procedimiento se realizará con material estéril. Si decide participar, necesitaremos unos minutos para completar todo el proceso.

Beneficios: Tomando parte en este estudio, ayudará a los médicos a conocer más sobre la malaria en nuestra Amazonia. El beneficio también será para su comunidad, ya que los médicos conocerán mejor sobre como se transmite la malaria en su comunidad y se podrán diseñar mejores técnicas de control. Los análisis de la sangre son completamente gratuitos para usted. Un personal de salud le visitara después de iniciado el tratamiento, de esta manera su condición de salud será estrechamente evaluada. Confidencialidad: Protegemos su confidencialidad. Sus respuestas, resultados y otra información que nos provea, tendrán un código en lugar de su nombre. Guardaremos toda la información bajo llave en nuestro laboratorio de la calle Morona 448, Iquitos, y sólo personal del estudio tendrá acceso. La información que lo identifique no será publicada con los resultados. Nosotros nos reservamos el privilegio, de en el futuro, hacer otras pruebas para otros estudios sobre infecciones en las muestras obtenidas pero su nombre no será relacionado con la muestra. Voluntad a participar: La participación es voluntaria. En el caso de que usted decida participar, tiene opción de retirarse en cualquier momento si así lo desea, sin que esto afecte la atención médica que usted recibe. Preguntas acerca de este estudio: Este formato de consentimiento explica el objetivo del estudio. Por favor léalo con mucho cuidado antes de firmarlo. Puede hacer las preguntas que desee sobre lo que no entienda. Si no tiene preguntas ahora, siempre puede hacerlas en el futuro. Durante este estudio, va a ser informado sobre toda nueva información que pueda afectar a su salud. Si tiene alguna pregunta, o si quiere hablar con alguien porque siente que usted ha sido maltratado, o siente que participar en el estudio ha afectado su salud, por favor comuníquese por teléfono con el Dr. Joseph Vinetz (investigador principal, 065-242226) o al Dr. Raúl Chuquiyauri (coordinador, 065-9610846). Si firma abajo significa que desea participar.
Nombre y firma del participante
Loreto, ____ de __________ del 200
Nombre y firma del testigo
Loreto, ____ de __________ del 200
Firma del testigo
Loreto, ____ de __________ del 200

Formato A2: Consentimiento Informado de Participación – Menores
CONSENTIMIENTO INFORMADO DE PARTICIPACIÓN-MENORES
Título del Proyecto de Investigación: Genotipificación de Plamodium vivax en la Amazonía Peruana (Reservorios
Humanos para la Transmisión de Malaria por Plasmodium vivax en la amazonía Peruana)
Investigador principal: Dr. Joseph Vinetz. Co-Investigadores: Dr. Raul Chuquiyauri, Dr. Robert H. Gilman, Dra. Margaret Koset, Dra. Maritza Calderón
Propósito del Estudio: Este es un estudio conducido por la Universidad Peruana Cayetano Heredia, la Universidad de California San Diego y EL Instituto de Medicina Tropical Alexander von Humboldt; y en colaboración con el Ministerio de Salud a través de la Dirección Regional de Salud de Loreto.
La malaria es una enfermedad muy frecuente en nuestras comunidades, afectando a niños y adultos, con importantes trastornos entre las personas que la padecen. Hay dos especies de parásito que causan malaria en Perú: Plasmodium vivax y Plasmodium falciparum. Desde hace muchos años numerosas instituciones estamos haciendo un gran esfuerzo para luchar contra ella. Estos esfuerzos en el diagnóstico, cuidado de enfermos y prevención requieren la participación directa de las personas afectadas como usted.
Usted esta siendo invitado a tomar parte en este estudio porque su gota gruesa es positiva para Plasmodium vivax. Su participación en este estudio depende completamente de ustedes.
Procedimientos: Si usted esta de acuerdo que su hijo participe, un trabajador de salud realizará lo siguiente:
Le hará preguntas básicas sobre su casa, historial médico y síntomas de su hijo.
Se le tomara a su hijo una muestra de sangre de 05 ml. Exclusiones: Si su hijo tiene alguna de las siguientes condiciones no podrá participar en este estudio:
Uso de antibióticos al momento o una semana antes de ingresar al estudio.
Estar recibiendo tratamiento antimalárico ahora o en la última semana.
No haber firmado el consentimiento informado.
Tener malaria por Plasmodium falciparum o malaria mixta.
Tener menos de 5 años de edad. Riesgos e Incomodidades: La participación en el estudio conlleva un riesgo mínimo. La toma de sangre puede ocasionarle cierta incomodidad. Existe un mínimo riesgo de que aparezca un moretón después de la toma de sangre. Nosotros trataremos de mantener este riesgo tan bajo como sea posible. El procedimiento se realizará con material estéril. Necesitaremos unos minutos de su tiempo. Beneficios:

Con su participación ayudará a los médicos a conocer más sobre la malaria en nuestra Amazonia. Los análisis de la sangre de su hijo son completamente gratuitos. Un personal de salud le visitará después de iniciado el tratamiento, de esta manera la condición de salud de su hijo será estrechamente evaluada. Confidencialidad: Nosotros protegemos su confidencialidad y la de su hijo dentro de los estándares establecidos. Sus respuestas, resultados de pruebas, y otra información que nos provea, serán identificados con un código en lugar de su nombre. Guardaremos toda la información del estudio bajo llave en nuestras oficinas en la calle Morona 448, y sólo el personal del estudio tendrá acceso a la información, a menos que nos dé su permiso. Su nombre, el de su hijo y otra información que lo identifique no serán publicados. Nosotros nos reservamos el privilegio, de en el futuro, hacer otras pruebas para otros estudios sobre infecciones en las muestras obtenidas pero su nombre o el de su hijo no serán relacionados con la muestra. Voluntad a participar: La participación es voluntaria. En el caso de que usted decida hacer participar a su hijo, tiene la opción de retirarlo del estudio en cualquier momento si así lo desea, sin que esto afecte a la calidad o atención médica que usted o su hijo recibe. Preguntas acerca de este estudio: Este formato de consentimiento explica el objetivo del estudio. Por favor léalo con cuidado antes de firmarlo. Puede hacer preguntas sobre lo que no entienda. Si no tiene preguntas ahora, siempre puede hacerlas en el futuro. Durante el estudio, va a ser informado sobre toda nueva información que pueda afectar la salud de su hijo. Si tiene alguna pregunta, o siente que su hijo ha sido maltratado, o siente que participar en el estudio ha afectado su salud, por favor comuníquese por teléfono con el Dr. Joseph Vinetz (investigador principal – 065-242226) o al Dr. Raúl Chuquiyauri (coordinador – 065-9610846). Si Usted firma abajo significa que desea hacer participar a su hijo.
Nombre y firma del padre o tutor responsable
Iquitos, ____ de __________ del 200
Nombre del niño y fecha de nacimiento
Iquitos, ____ de __________ del 200
Nombre y Firma del testigo
Iquitos, ____ de __________ del 200
Firma y nombre del investigador responsable
Iquitos, ____ de __________ del 200


















