




![The influence of γ O and Na on the formation of calcium silicate … · 2007. 2. 27. · Calcium silicate hydrates in the CaO quartz H 2 O system with C/S = 0.66 187 Luke [26] established](https://static.fdocument.org/doc/165x107/60cc12a7d6f169767d3b011c/the-influence-of-o-and-na-on-the-formation-of-calcium-silicate-2007-2-27.jpg)


γλώσσες
Σελίδες
Νομικός


261 Khomari and Javadi
Int. J. Biosci. 2015
RESEARCH PAPER OPEN ACCESS
Effect of calcium and boron on germination of differentially
aged oilseed rape seeds under salinity
Saeid Khomari*, Ahmad Javadi
Department of Agronomy and Plant Breeding, Faculty of Agricultural Science, University of
Mohaghegh Ardabili, Ardabil, Iran
Key words: Boron, Brassica napus, Calcium, Salinity, Seed vigor.
http://dx.doi.org/10.12692/ijb/6.5.261-272 Article published on March 14, 2015
Abstract
A soilless culture experiment was carried out at the glasshouse conditions, in order to elucidate the beneficial
effect of seed vigor and the possible alleviatory influence of boron and calcium on the salt tolerance capability of
Rapeseed. Artificial aging of rapeseed seeds significantly reduced the seedling growth especially under more
saline conditions. In spite of promotive effects of application of both boron and calcium on rapeseed
germinability and seedling growth, there was a more significant enhancement on emergence rate of seedlings
from high vigor seeds. Supplementary concentrations of boron and calcium more significantly reduced the Na+
content and increased the K+ amount of the seedlings under 100 mM NaCl. Finally, it can be concluded that
boron and calcium would improve the salt tolerance capability of rapeseed seedlings from vigorous seeds, at least
in the early seedling development.
* Corresponding Author: Saeid Khomari [email protected]
International Journal of Biosciences | IJB |
ISSN: 2220-6655 (Print), 2222-5234 (Online)
http://www.innspub.net
Vol. 6, No. 5, p. 261-272, 2015

262 Khomari and Javadi
Int. J. Biosci. 2015
Introduction
Rapeseed or canola (Brassica napus L.) is today the
world’s third-leading source of both vegetable oil and
oil extraction meal (Friedt and Snowdon 2009).
Today salinity remains a major abiotic stress that
adversely affects crop productivity and quality (Boyer
1982; Yamaguchi and Blumwald 2005). Salt as NaCl
is the predominant salt causing salinization (Munns
and Tester 2008) and is so abundant on our planet
that it is a major constraint to field crops production
especially rapeseed, jeopardizing the capacity of
agriculture to sustain the burgeoning human
population increase (Flowers 2004). It is estimated
that 20% of all cultivated land and nearly half of
irrigated land is salt-affected, greatly reducing yield
well below the genetic potential (Munns 2002;
Flowers 2004).
Seed germination and subsequent seedling
development are important and vulnerable stages in
the life cycle of crops and determine successful
seedling establishment in the field (Ellis and Roberts
1981; Rehman et al. 1999; De Figueiredo et al. 2003).
There are three main negative effects of high salt
concentrations that influence seed germination and
seedling growth: water deficit, ion toxicity associated
with excessive Cl− and Na+, and interference with
nutrition, leading to nutrient imbalance (Silberbush
and Ben-Asher 2001). Nutrient imbalance can
develop in salt-stressed plants in different ways. It
may result from the effect of salinity on nutrient
availability, uptake, transport, or partitioning within
the plant or may be caused by physiological
inactivation of a given nutrient resulting in an
increase in the plant’s internal requirement for that
essential element (Grattan and Grieve 1999). Hence,
investigating the interaction of NaCl with nutrients
that have key roles in plant development, such as
boron and calcium, and evaluating the effect of seed
and seedling vigor on this interaction are important
to optimize the growth of rapeseed under salt stress.
The presence of Ca2+ as the dominant cation in
agricultural soils generally ensures that the absolute
Ca2+ level is not a primary growth-limiting factor. As
salinity increases, the requirement of plants for Ca2+
increases (Gerard 1971). The interaction between B
and Ca2+ in the nutrition of both mono- and
dicotyledonous plants has long been recognized
(Marsh and Shive 1941). The plant cell wall was
suggested as the place where the B role influenced Ca
metabolism (Yamaguchi et al. 1986). From soilless
culture experiments conducted in a greenhouse,
researchers found that a balanced nutrition of B and
Ca was able to counter-act the deficiency of these
nutrients in salt-stressed plants, leading to a huge
increase in salinity tolerance of symbiotic pea plants
(El-Hamdaoui et al. 2003; Bonilla et al. 2004).
Despite the common occurrence of high boron and
high salinity in many parts of the world, little
research has been done to study the interaction of the
two (Grattan and Grieve 1999). Different factors may
influence alleviatory effects of nutrient elements on
salt tolerance of crop plants. Many reports are
concentrated on genotypic differences on this respect
(Marsh and Shive 1941; Ehret et al. 1990; El-
Hamdaoui et al. 2003; Bonilla et al. 2004). However,
it is not known whether there is any linkage between
high salt tolerance in rapeseed and responsibility to
supplemental boron and calcium nutrition. Therefore,
the main purpose of this study is to determine the
possible effectiveness of high vigor seeds and
supplemental nutrition of calcium and boron on
salinity tolerance of rapeseed.
Materials and methods
Seed preparation and treatments
Seeds of a winter rapeseed (Brassica napus L. var.
oleifera cv. Okapi; Agriculture Organization, Iran)
were divided into two sub-samples. A subsample was
kept as control or high vigor seed lot (V1). The other
sub-sample with about 16% moisture content was
artificially deteriorated at 40°C for 72 hours (V2). As a
result, the two seed lots with 98% (V1) and 88% (V2)
viabilities were provided. For salinity treatments,
NaCl was added at a concentration of 0, 50 mM or
100 mM. Calcium was added as Ca(NO3)2.4H2O and
Boron as H3BO3 in Hoagland-Arnon nutrient
solution. The concentrations of Ca (5 mM and 15 mM)
and B (40 μM, 240 μM and 480 μM) from the
different treatments are summarized in Table 1. The

263 Khomari and Javadi
Int. J. Biosci. 2015
pH was always checked and adjusted around 6.7.
Previously germinated rapeseed seedlings were
transferred to plastic rectangular pots and cultivated
on Perlite (without release of any Ca, B or salt) with
Hoagland-Arnon medium, as the nutrient solution.
Plants were maintained in the glasshouse conditions.
Irrigation with nutrient solutions was repeated every
4 days. In order to ensure that no increase in
concentration of different ions in the pots occurs,
electro-conductivity of output solution was controlled
after irrigation.
Sampling and measurements
The number of emerged seedlings (full expansion of
cotyledons) from each plot was counted every 24
hours. The rate of emergence was calculated by Ellis
and Roberts (1981) equation. At 25th day of
experiment, seedling length, fresh and dry weight and
leaf number were measured. Twenty seedlings of each
treatment were randomly selected and dried at 80 °C
for 24 h. After milling to ensure homogeneity, 0.5 g of
each sample was electric furnace-ashed (2 h at 550
°C) and acid-digested (10 ml of 1 N HCl). The
concentrations of Na+, K+ and Ca2+ were measured by
flame photometer. The chlorophyll content of
seedling leaves was determined according to Arnon
(1949).
A factorial soilless culture experiment on the basis of
randomized complete block design with three
replications was conducted to evaluate salt tolerance
of rapeseed seedlings, affected by seed vigor level and
supplemental nutrition of calcium and boron. All the
measurements were repeated at least two times and
data was statistically analysed by the two-way
ANOVA. The means were compared in accordance
with the Duncan’s multiple range test at p< 0.05.
Results
The percentage of seedling emergence was
significantly (P≤ 0.01) reduced with increased levels
of seed deterioration and salinity (figure 1).
Deteriorated seeds had the minimum emergence of
seedlings in 100 mM NaCl with about 50% reduction
compared to control. The inhibitory effect of NaCl on
the establishment of seedlings from poor vigor seeds
was greater than those from high vigor ones. Different
concentrations of boron and calcium had no
significant effect on seedling emergence in any
concentrations of salt. Rapeseed seedlings slowly
emerged, as different NaCl concentrations were
added into the germination medium (table 2). The
third level of salinity illustrated the minimum value of
seedling emergence rate with about 25% decrease
compared with non-saline condition. Moreover, the
rate of emergence was significantly decreased in the
deteriorated seeds (figure 2). The presence of
increasing concentration of B or Ca (+6B+1Ca,
+12B+1Ca and +1B+3Ca supplements) but not both
significantly improved the emergence rate in non-
deteriorated vigorous seeds in comparison with
deteriorated ones. These observations suggest that
the vigor of the seed lot has influenced the advantage
of B-Ca2+supplementation in the seedling growth
medium.
The length and dry weight of seedlings produced from
deteriorated poor vigor seeds were about 5 cm and
35% less than those of high vigor ones, respectively
(table 3). Increasing concentrations of boron
(+12B+1Ca) and calcium (+1B+3Ca) negligibly but no
significantly increased the length and dry weight of
seedlings compared to base concentrations of these
elements in the nutrient solution. On the other hand,
the levels of 3Ca combined with high boron
concentrations (+6B and +12B) reduced rapeseed
seedling growth. The length and dry weight of
rapeseed seedling was significantly reduced when the
salt concentration increased, in a way that 100 mM of
NaCl decreased these parameters approximately 30%
and 33% respectively, compared with the control. The
results indicated that as the salinity increased, the leaf
number of seedlings was significantly decreased
(figure 3 and 4). The number of leaves of seedlings
obtained from deteriorated seed lot was more
decreased when NaCl concentration increased,
compared to those from non-deteriorated vigorous
seeds. The mean leaf number of seedlings from high
and low vigor seed lots were statistically the same in
the salt-free medium (figure 3). The presence of
264 Khomari and Javadi
Int. J. Biosci. 2015
increasing concentration of B or Ca (+6B+1Ca,
+12B+1Ca and +1B+3Ca supplements) but not both
significantly increased the number of leaves of
seedlings only when no salt was applied to the
nutrient solution. Although, boron and calcium were
added into the perlite as nutrient supplements, the
mean leaf number of seedlings was statistically the
same under NaCl imposed salinity stress (figure 4).
These observations suggest that the B-Ca
supplementation did not overcome the negative effect
of salinity on leaf number of rapeseed seedlings.
Table 1. Boron and calcium treatments used in this experiment.
5 mM Ca2+ 15 mM Ca2+
40 μM B +B+Ca +B+3Ca
240 μM B +6B+Ca +6B+3Ca
480 μM B +12B+Ca +12B+3Ca
Table 2. Seedling emergence rate at different salinity levels.
Salinity Seedling emergence rate (d-1)
S0 (control) 0.463a
S50 0.321b
S100 0.217c
Different letters indicate the significant difference at p<0.05.
Table 3. Length, dry weight and chlorophyll a content of oilseed rape seedling affected by different levels of seed
vigour, B-Ca combinations and salinity.
Seedling length (cm) Seedling dry weight (mg) chlorophyll a
(mg.g-1 FW)
Seed vigour V1 (control) 15.0a 51.8a 159.1a
V2 10.0b 34.2b 138.0b
B-Ca combinations +1B+1Ca (control) 11.8b 40.7b 144.7b
+6B+1Ca 13.1a 45.1a 150.6a
+12B+1Ca 13.1a 45.4a 151.8a
+1B+3Ca 13.2a 45.4a 152.4a
+6B+3Ca 11.8b 40.7b 145.7b
+12B+3Ca 11.9b 40.8b 146.0b
Salinity S0 (control) 14.8a 51.6a 198.8a
S50 12.6b 43.0b 144.9b
S100 10.2c 34.4c 101.9c
Different letters indicate the significant difference at p<0.05.
The sodium content of the seedlings from low vigor
seeds was about 14% more than those from vigorous
seed lot (Table 4). As typically occurs, increasing NaCl
concentration in the nutrient solution increased Na+
level in rapeseed seedlings (Figure 5). Surprisingly,
adding supplementary concentrations of +6B and
+12B or +3Ca to the nutrient solution more
significantly reduced the sodium content of the
seedlings under different strengths (50 and 100 mM
NaCl) of salinity. However, +6B+3Ca and +12B+3Ca
combinations led to a minor decline in Na+ content of
seedling which was not statistically significant. More
salinity and seed deterioration reduced the amount of
potassium in rapeseed seedlings (Figure 6); in a way
that the highest K+ content was observed in the
seedlings obtained from vigorous seeds under non-
saline conditions (168.24 ppm), and the lowest one
was observed in the seedlings from poor vigor seeds
in 100mM salinity (133.04 ppm). Adding further
concentrations of boron and calcium led to increased
potassium level of seedlings only in 50 and 100mM
salinity (Figure 7). In salinity treatments, seedlings
grown in +6B, +12B and 3Ca supplements contained
the maximum amounts of K+ as compared to the
265 Khomari and Javadi
Int. J. Biosci. 2015
control. Supplemental concentrations of boron (+6B
and +12B) in combination with +3Ca concentration
were not significantly different from the control in
terms of this attribute. Artificial aging of rapeseed
seeds negligibly but significantly decreased calcium
content of the seedlings (Table 4). The more saline
medium also led to a significant reduction in the
amount of calcium in seedlings (Figure 8). Adding
supplementary calcium with all concentrations of
boron (+1B+3Ca, +6B+3Ca, +12B+3Ca) in different
NaCl levels increased Ca2+ content in rapeseed
seedlings. On the other hand, boron had no
statistically significant effect on calcium content of
seedlings.
Table 4. The amounts of Na+ and Ca2+ in oilseed rape seedlings affected by different levels of seed vigour.
Seed vigour Na+ (ppm) Ca2+ (ppm)
V1 (control) 155.7b 35.8a
V2 180.3a 34.3b
Different letters indicate the significant difference at p<0.05.
Table 5. The chlorophyll a content of seedlings affected by different levels of seed vigour, B-Ca2+ combinations
and salinity.
chlorophyll a (mg.g-1 FW)
159.1a V1 (control) Seed vigour
138.0b V2
144.7b +1B+1Ca (control) B-Ca Combinations
150.6a +6B+1Ca
151.8a +12B+1Ca
152.4a +1B+3Ca
145.7b +6B+3Ca
146.0b +12B+3Ca
198.8a S0 (control) Salinity
144.9b S50
101.9c S100
Different letters indicate the significant difference at p<0.05.
Seed vigor reduction led to a decline in the amount of
chlorophyll a and b in rapeseed seedlings (Table 3
and Figure 9). Different concentrations of boron and
calcium increased the amount of chlorophyll a, as the
seedlings grown in +6B+1Ca, +12B+1Ca, and
+1B+3Ca combinations had the highest leaf
greenness. The salinity stress decreased chlorophyll a
in the leaves, as 100mM NaCl diminished chlorophyll
a level in leaves by approximately 50% (Table 3).
Seedlings from aged poor vigor seeds in 100mM
salinity had the minimum level (about 25 mg.g-1 FW)
of chlorophyll b (Figure 9). In general, salt stress
caused the loss of chlorophyll b of rapeseed seedlings
by more than two to three folds in comparison with
control (no salt). Additional concentrations of boron
and calcium in 50 and 100mM salinity reduced the
negative effects of salt stress on chlorophyll b of
leaves (Figure 10). Combined treatments of +6B+3Ca
and +12B+3Ca were not significantly different from
the control in terms of the amount of chlorophyll b.
Therefore, it can be concluded that the exogenous
application of B and Ca supplements would be more
effective when the concentration of salt in seedling
growth medium is high. Also, concomitant increase of
B and Ca concentrations had no significant advantage
on rapeseed germinability and seedling development
under salinity stress.
266 Khomari and Javadi
Int. J. Biosci. 2015
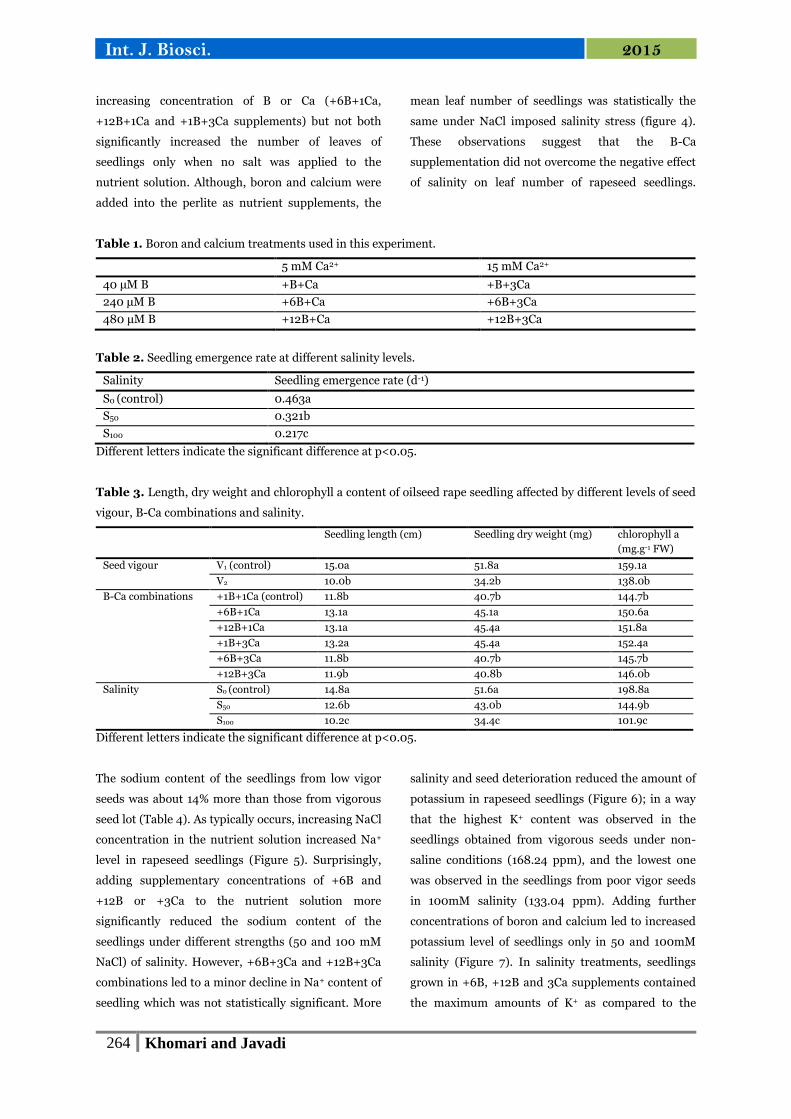
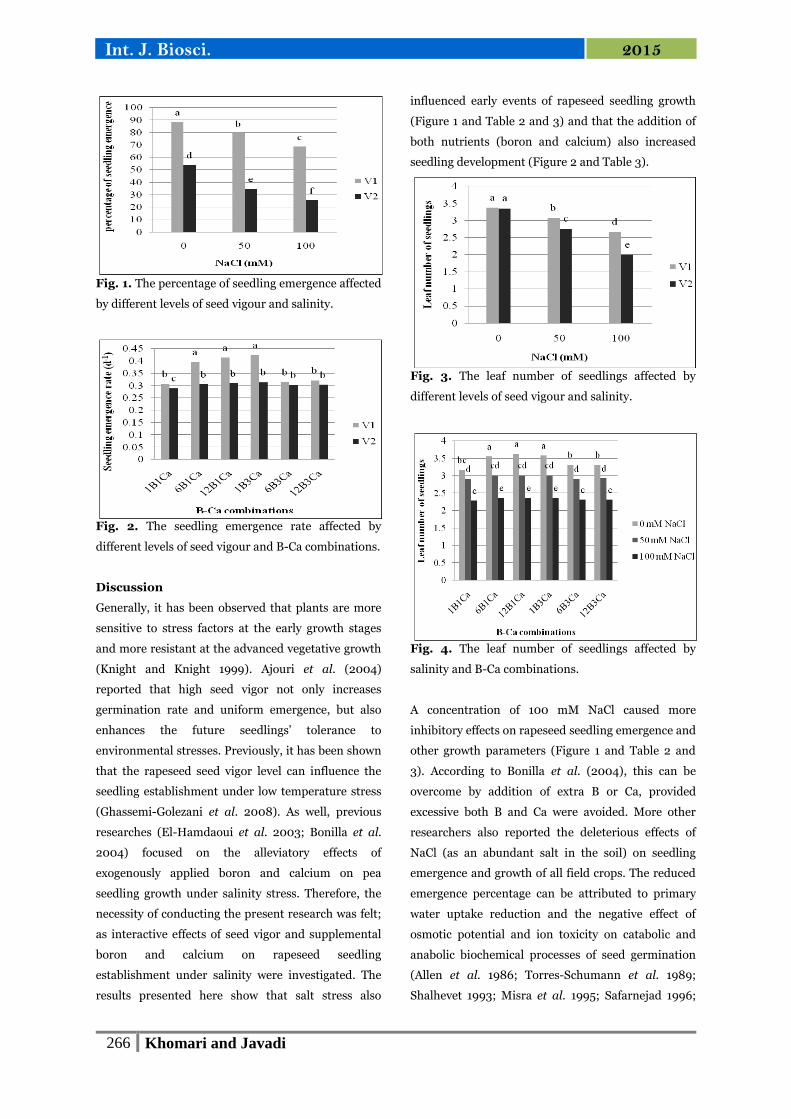
Fig. 1. The percentage of seedling emergence affected
by different levels of seed vigour and salinity.
Fig. 2. The seedling emergence rate affected by
different levels of seed vigour and B-Ca combinations.
Discussion
Generally, it has been observed that plants are more
sensitive to stress factors at the early growth stages
and more resistant at the advanced vegetative growth
(Knight and Knight 1999). Ajouri et al. (2004)
reported that high seed vigor not only increases
germination rate and uniform emergence, but also
enhances the future seedlings’ tolerance to
environmental stresses. Previously, it has been shown
that the rapeseed seed vigor level can influence the
seedling establishment under low temperature stress
(Ghassemi-Golezani et al. 2008). As well, previous
researches (El-Hamdaoui et al. 2003; Bonilla et al.
2004) focused on the alleviatory effects of
exogenously applied boron and calcium on pea
seedling growth under salinity stress. Therefore, the
necessity of conducting the present research was felt;
as interactive effects of seed vigor and supplemental
boron and calcium on rapeseed seedling
establishment under salinity were investigated. The
results presented here show that salt stress also
influenced early events of rapeseed seedling growth
(Figure 1 and Table 2 and 3) and that the addition of
both nutrients (boron and calcium) also increased
seedling development (Figure 2 and Table 3).
Fig. 3. The leaf number of seedlings affected by
different levels of seed vigour and salinity.
Fig. 4. The leaf number of seedlings affected by
salinity and B-Ca combinations.
A concentration of 100 mM NaCl caused more
inhibitory effects on rapeseed seedling emergence and
other growth parameters (Figure 1 and Table 2 and
3). According to Bonilla et al. (2004), this can be
overcome by addition of extra B or Ca, provided
excessive both B and Ca were avoided. More other
researchers also reported the deleterious effects of
NaCl (as an abundant salt in the soil) on seedling
emergence and growth of all field crops. The reduced
emergence percentage can be attributed to primary
water uptake reduction and the negative effect of
osmotic potential and ion toxicity on catabolic and
anabolic biochemical processes of seed germination
(Allen et al. 1986; Torres-Schumann et al. 1989;
Shalhevet 1993; Misra et al. 1995; Safarnejad 1996;
267 Khomari and Javadi
Int. J. Biosci. 2015
Peñuelas et al. 1997). Comparison of our results with
findings of Bonilla et al. (2004) on pea suggested that
the prescribed alleviatory effect of boron and calcium
on seedling establishment and subsequent
development under salt stress is species specific.
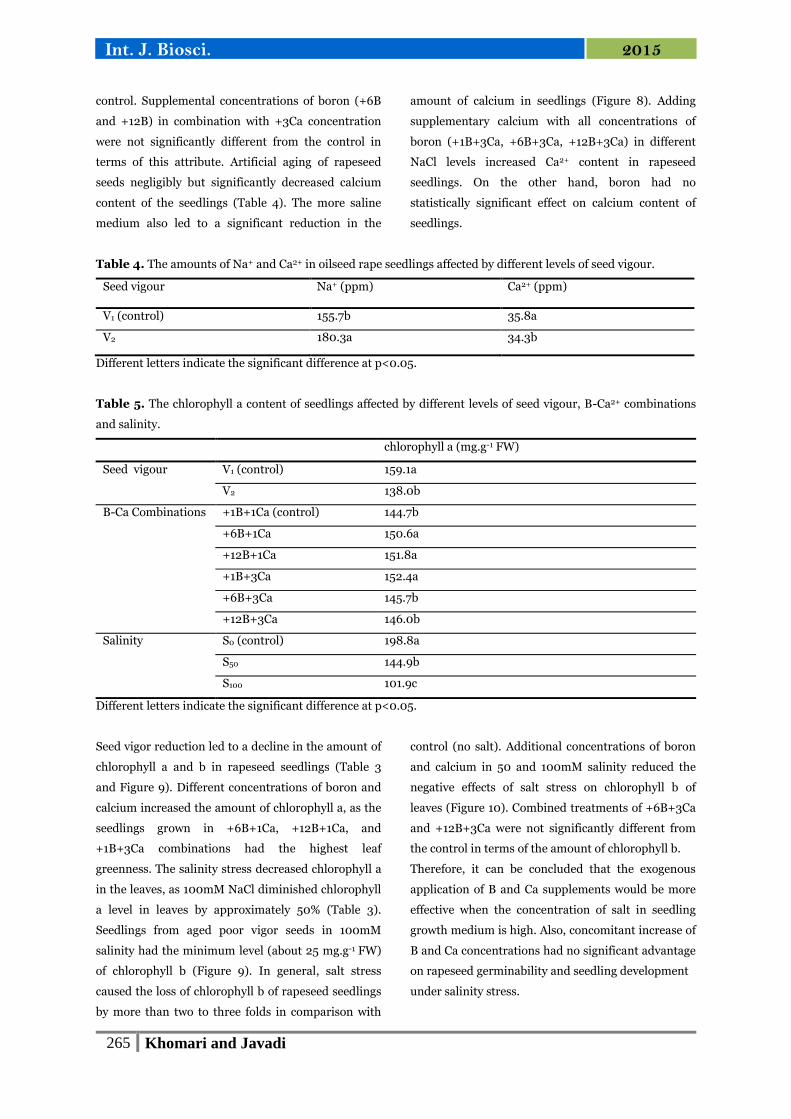
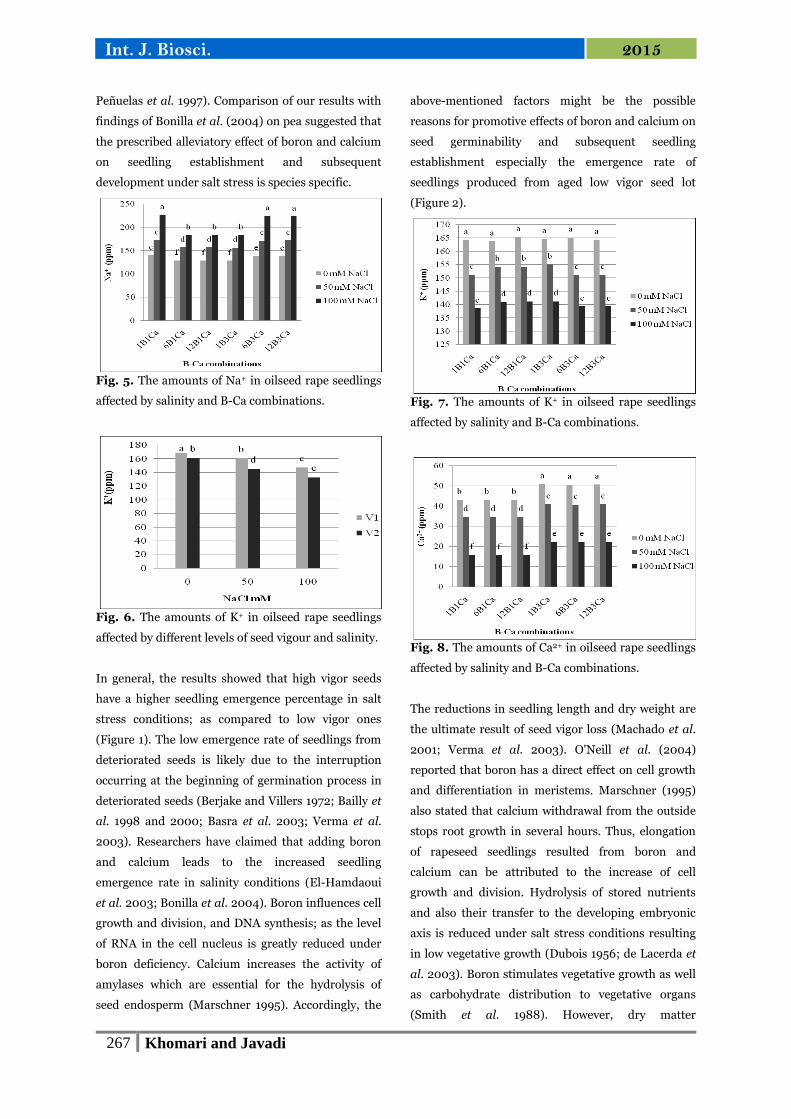
Fig. 5. The amounts of Na+ in oilseed rape seedlings
affected by salinity and B-Ca combinations.
Fig. 6. The amounts of K+ in oilseed rape seedlings
affected by different levels of seed vigour and salinity.
In general, the results showed that high vigor seeds
have a higher seedling emergence percentage in salt
stress conditions; as compared to low vigor ones
(Figure 1). The low emergence rate of seedlings from
deteriorated seeds is likely due to the interruption
occurring at the beginning of germination process in
deteriorated seeds (Berjake and Villers 1972; Bailly et
al. 1998 and 2000; Basra et al. 2003; Verma et al.
2003). Researchers have claimed that adding boron
and calcium leads to the increased seedling
emergence rate in salinity conditions (El-Hamdaoui
et al. 2003; Bonilla et al. 2004). Boron influences cell
growth and division, and DNA synthesis; as the level
of RNA in the cell nucleus is greatly reduced under
boron deficiency. Calcium increases the activity of
amylases which are essential for the hydrolysis of
seed endosperm (Marschner 1995). Accordingly, the
above-mentioned factors might be the possible
reasons for promotive effects of boron and calcium on
seed germinability and subsequent seedling
establishment especially the emergence rate of
seedlings produced from aged low vigor seed lot
(Figure 2).
Fig. 7. The amounts of K+ in oilseed rape seedlings
affected by salinity and B-Ca combinations.
Fig. 8. The amounts of Ca2+ in oilseed rape seedlings
affected by salinity and B-Ca combinations.
The reductions in seedling length and dry weight are
the ultimate result of seed vigor loss (Machado et al.
2001; Verma et al. 2003). O'Neill et al. (2004)
reported that boron has a direct effect on cell growth
and differentiation in meristems. Marschner (1995)
also stated that calcium withdrawal from the outside
stops root growth in several hours. Thus, elongation
of rapeseed seedlings resulted from boron and
calcium can be attributed to the increase of cell
growth and division. Hydrolysis of stored nutrients
and also their transfer to the developing embryonic
axis is reduced under salt stress conditions resulting
in low vegetative growth (Dubois 1956; de Lacerda et
al. 2003). Boron stimulates vegetative growth as well
as carbohydrate distribution to vegetative organs
(Smith et al. 1988). However, dry matter
268 Khomari and Javadi
Int. J. Biosci. 2015
accumulation in rapeseed seedlings can be the result
of stimulation of this process by boron. On the other
hand, affecting cell division and growth, calcium leads
to increased seedling dry weight (Marschner 1995).
Since the presence of high concentrations of both
boron and calcium has not had an improving effect on
rapeseed seedling growth (Table 3), we may state that
there is a rather antagonistic relationship between
boron and calcium in these concentrations.
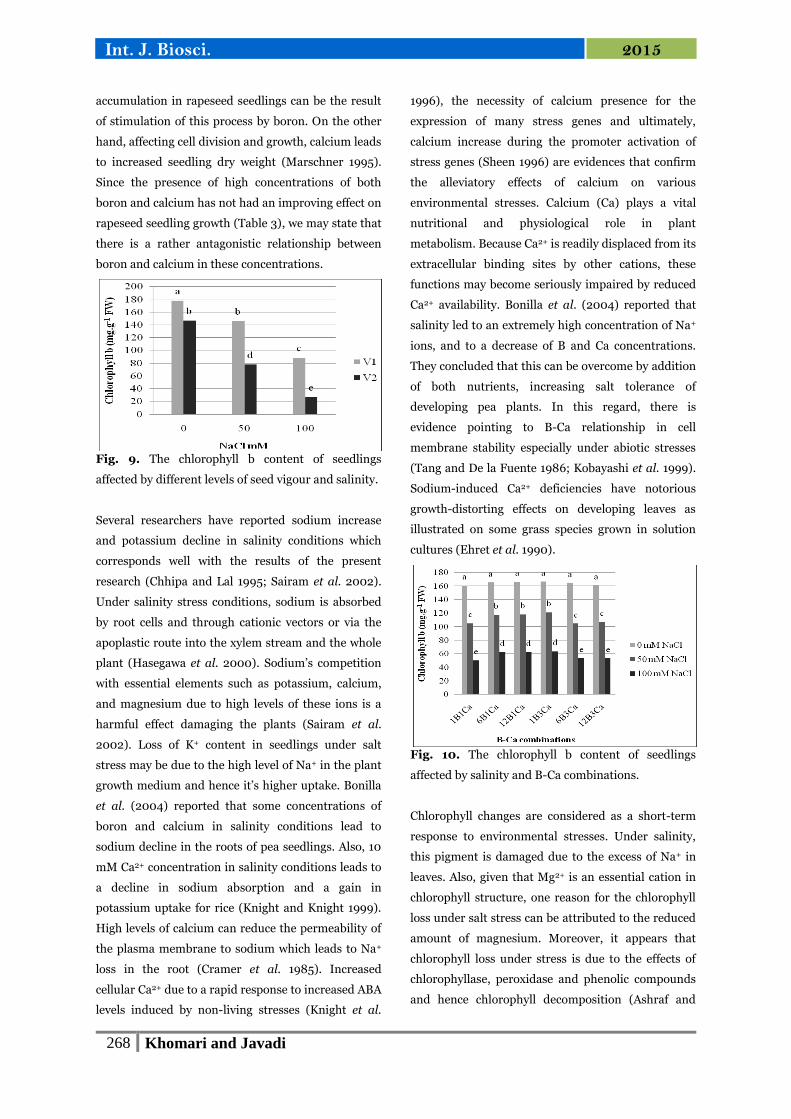
Fig. 9. The chlorophyll b content of seedlings
affected by different levels of seed vigour and salinity.
Several researchers have reported sodium increase
and potassium decline in salinity conditions which
corresponds well with the results of the present
research (Chhipa and Lal 1995; Sairam et al. 2002).
Under salinity stress conditions, sodium is absorbed
by root cells and through cationic vectors or via the
apoplastic route into the xylem stream and the whole
plant (Hasegawa et al. 2000). Sodium’s competition
with essential elements such as potassium, calcium,
and magnesium due to high levels of these ions is a
harmful effect damaging the plants (Sairam et al.
2002). Loss of K+ content in seedlings under salt
stress may be due to the high level of Na+ in the plant
growth medium and hence it’s higher uptake. Bonilla
et al. (2004) reported that some concentrations of
boron and calcium in salinity conditions lead to
sodium decline in the roots of pea seedlings. Also, 10
mM Ca2+ concentration in salinity conditions leads to
a decline in sodium absorption and a gain in
potassium uptake for rice (Knight and Knight 1999).
High levels of calcium can reduce the permeability of
the plasma membrane to sodium which leads to Na+
loss in the root (Cramer et al. 1985). Increased
cellular Ca2+ due to a rapid response to increased ABA
levels induced by non-living stresses (Knight et al.
1996), the necessity of calcium presence for the
expression of many stress genes and ultimately,
calcium increase during the promoter activation of
stress genes (Sheen 1996) are evidences that confirm
the alleviatory effects of calcium on various
environmental stresses. Calcium (Ca) plays a vital
nutritional and physiological role in plant
metabolism. Because Ca2+ is readily displaced from its
extracellular binding sites by other cations, these
functions may become seriously impaired by reduced
Ca2+ availability. Bonilla et al. (2004) reported that
salinity led to an extremely high concentration of Na+
ions, and to a decrease of B and Ca concentrations.
They concluded that this can be overcome by addition
of both nutrients, increasing salt tolerance of
developing pea plants. In this regard, there is
evidence pointing to B-Ca relationship in cell
membrane stability especially under abiotic stresses
(Tang and De la Fuente 1986; Kobayashi et al. 1999).
Sodium-induced Ca2+ deficiencies have notorious
growth-distorting effects on developing leaves as
illustrated on some grass species grown in solution
cultures (Ehret et al. 1990).
Fig. 10. The chlorophyll b content of seedlings
affected by salinity and B-Ca combinations.
Chlorophyll changes are considered as a short-term
response to environmental stresses. Under salinity,
this pigment is damaged due to the excess of Na+ in
leaves. Also, given that Mg2+ is an essential cation in
chlorophyll structure, one reason for the chlorophyll
loss under salt stress can be attributed to the reduced
amount of magnesium. Moreover, it appears that
chlorophyll loss under stress is due to the effects of
chlorophyllase, peroxidase and phenolic compounds
and hence chlorophyll decomposition (Ashraf and

269 Khomari and Javadi
Int. J. Biosci. 2015
Harris 2004). Grewal et al. (1998) stated that the
increased availability of zinc and boron significantly
increases the chlorophyll content of young leaves of
rapeseed. Boron stimulates vegetative growth as well
as carbohydrate transfer to vegetative organs to be
used for chlorophyll production (Smith et al. 1988).
Ghassemi-Golezani et al. (2008) reported that the
amount of chlorophyll a and b in the leaves of
rapeseed seedlings is declined with increased seed
deterioration.
Overall the results indicate that a balanced B-Ca
relationship also increases salt tolerance during the
early stages of rapeseed seedling development and
growth. Nevertheless, besides genetic approaches
searching for tolerant cultivars, concomitantly
studying both the seed vigor status and the
nutritional relationship between B and Ca for other
crops can lead to applied approaches to increase salt-
tolerance and field crop production.
Acknowledgements
The authors wish to thank the Research and post-
graduation Committee of University of Mohaghegh
Ardabili for a research grant in support of this
project..
References
Ajouri A, Asgedom H, Becker M. 2004. Seed
priming enhances germination and seedling growth of
barley under conditions of P and Zn deficiency.
Journal of Plant Nutrition and Soil Science 167, 630-
636.
http://dx.doi.org/10.1002/jpln.200420425
Allen S, Dobrenz A, Bartels P. 1986.
Physiological response of salt-tolerant and non-
tolerant alfalfa to salinity during germination. Crop
Science 26, 1004-1008.
Arnon DI. 1949. Copper enzymes in isolated
chloroplasts. Polyphenoloxidase in Beta vulgaris.
Plant Physiology 24, 1-15.
Ashraf M, Harris P. 2004. Potential biochemical
indicators of salinity tolerance in plants. Plant Science
166, 3-16.
http://dx.doi.org/10.1016/j.plantsci.2003.10.024
Bailly C, Benamar A, Corbineau F, Come D.
1998. Free radical scavenging as affected by
accelerated ageing and subsequent priming in
sunflower seeds. Physiologia Plantarum 104, 646-
652.
http://dx.doi.org/10.1034/j.13993054.1998.1040418.
x
Bailly C, Benamar A, Corbineau F, Côme D.
2000. Antioxidant systems in sunflower (Helianthus
annuus L.) seeds as affected by priming. Seed Science
Research 10, 35-42.
http://dx.doi.org/10.1017/S0960258500000040
Basra S, Ahma Basra S, Ahmad N, Khan M,
Iqbal N, Cheema M. 2003. Assessment of
cottonseed deterioration during accelerated ageing.
Seed science and technology 31, 531-540.
http://dx.doi.org/10.15258/sst.2003.31.3.02
Berjak P, Villiers T. 1972. Ageing in plant embryos.
III. Acceleration of senescence following artificial
ageing treatment. New Phytologist 45, 513-518.
Bonilla I, El-Hamdaoui A, Bolaños L. 2004.
Boron and calcium increase Pisum sativum seed
germination and seedling development under salt
stress. Plant and Soil 267(1-2), 97-107.
http://dx.doi.org/10.1007/s11104-005-4689-7
Boyer JS. 1982. Plant productivity and environment.
Science 218, 443-448.
Chhipa B, Lal P. 1995. Na/K ratios as the basis of
salt tolerance in wheat. Crop and Pasture Science 46,
533-539.
http://dx.doi.org/10.1071/AR9950533
Cramer GR, Läuchli A, Polito VS. 1985.
Displacement of Ca2+ by Na+ from the Plasmalemma
of Root Cells A Primary Response to Salt Stress? Plant

270 Khomari and Javadi
Int. J. Biosci. 2015
Physiology 79, 207-211.
De Figueiredo e Albuquerque M, de Carvalho
N. 2003. Effect of the type of environmental stress on
the emergence of sunflower (Helianthus annuus L.),
soybean (Glycine max L.) and maize (Zea mays L.)
seeds with different levels of vigor. Seed Science and
Technology 31(2), 465–479.
De Lacerda CF, Cambraia J, Oliva MA, Ruiz
HA, Prisco JTN. 2003. Solute accumulation and
distribution during shoot and leaf development in two
sorghum genotypes under salt stress. Environmental
and Experimental Botany 49(2), 107–120.
http://dx.doi.org/10.1016/S0098-8472(02)00064-3
Dubois M, Gilles KA, Hamilton JK, Rebers Pt,
Smith F. 1956. Colorimetric method for
determination of sugars and related substances.
Analytical Chemistry 28(3), 350–356.
Ehret D, Redmann R, Harvey B, Cipywnyk A.
1990. Salinity-induced calcium deficiencies in wheat
and barley. Plant and Soil 128, 143-151.
http://dx.doi.org/10.1007/BF00011103
El-Hamdaoui A, Redondo-Nieto M, Torralba
B, Rivilla R, Bonilla I, Bolaños L. 2003.
Influence of boron and calcium on the tolerance to
salinity of nitrogen-fixing pea plants. Plant and Soil
251(1), 93–103.
DOI:10.1023/A:1022980330718
Ellis R, Roberts E. 1981. The quantification of
ageing and survival in orthodox seeds. Seed Science
and Technology 9, 373–409.
Flowers T. 2004. Improving crop salt tolerance.
Journal of Experimental Botany 55, 307-319.
http://dx.doi.org/10.1093/jxb/erh003
Friedt W, Snowdon R. 2009. Oilseed rape, In:
Vollmann J, Rajcan I, eds. Handbook of plant
breeding oil crops, Springer, Dordrecht. 91-126 p.
Gerard C. 1971. Influence of osmotic potential,
temperature, and calcium on growth of plant roots.
Agronomy Journal 63, 555-558.
Ghassemi-Golezani K, Khomari S, Valizadeh
M, Alyari H. 2008. Changes in chlorophyll content
and fluorescence of leaves of winter rapeseed affected
by seedling vigor and cold acclimation duration.
Journal of Food Agricultur Environ 6, 196-199.
Grattan SR, Grieve CM. 1999. Mineral nutrient
acquisition and response by plants grown in saline
environments, In: Pessarakli M, ed, Handbook of
plant and crop stress. Marcel Dekker, Inc., New York.
203-229 p.
González AMC, García EA, Pineda JP, Arreola
JA, Aguilar LAV, León MTC. 2008. Effect of
calcium, boron and molybdenum on plant growth and
bract pigmentation in poinsettia. Revista Fitotecnia
Mexicana 31.
Grewal HS, Graham RD, Stangoulis J. 1998.
Zinc‐boron interaction effects in oilseed rape. Journal
of plant nutrition 21, 2231-2243.
http://dx.doi.org/10.1080/01904169809365557
Hasegawa PM, Bressan RA, Zhu JK, Bohnert
HJ. 2000. Plant cellular and molecular responses to
high salinity. Annual review of plant biology 51, 463-
499.
http://dx.doi.org/10.1146/annurev.arplant.51.1.463
Kafi M, Borzoui A, Salehi M, Kamandi A,
Ma’soumi A, Nabati J. 2009. Physiology of
environmental stresses in plants. Mashhad University
Jihad, 1st ed., 502 p.
Khajeh-Hosseini M, Powell A, Bingham I.
2003. The interaction between salinity stress and
seed vigour during germination of soyabean seeds.
Seed Science and technology 31, 715-725.
http://dx.doi.org/10.15258/sst.2003.31.3.20
Knight H, Knight MR. 1999. Calcium signaling in

271 Khomari and Javadi
Int. J. Biosci. 2015
plants responding to stress. In: Smallwood MF, Alvert
CM, Bowles DG, ed Plant Responses to Environment
Stress, Scientific Publishers. 1-8 p.
Knight H, Trewavas AJ, Knight MR. 1996. Cold
calcium signaling in Arabidopsis involves two cellular
pools and a change in calcium signature after
acclimation. The Plant Cell Online 8, 489-503.
Kobayashi M, Nakagawa H, Asaka T, Matoh T.
1999. Borate-rhamnogalacturonan II bonding
reinforced by Ca2+ retains pectic polysaccharides in
higher-plant cell walls. Plant Physiology 119, 199-
204.
Machado N, Custodio C, Takaki M. 2001.
Evaluation of naturally and artificially aged seeds of
Phaseolus vulgaris L. Seed Science and Technology
29(1), 137-149.
Marsh R, Shive J. 1941. Boron as a factor in the
calcium metabolism of the corn plant. Soil Science 51:
141-152.
Marschner H. 1996. Mineral nutrition of higher
plants. Wiley Online Library.
Misra N, Dwivedi U. 1995. Carbohydrate
metabolism during seed germination and seedling
growth in green gram under saline stress. Plant
physiology and biochemistry 33, 33-38.
Munns R. 2002. Comparative physiology of salt and
water stress. Plant, cell and environment 25, 239-
250.
http://dx.doi.org/10.1046/j.00168025.2001.00808.x
Munns R, Tester M. 2008. Mechanisms of salinity
tolerance. Annual Review of Plant Biology 59, 651-
681.
http://dx.doi.org/10.1146/annurev.arplant.59.03260
7.092911
O'Neill MA, Ishii T, Albersheim P, Darvill AG.
2004. Rhamnogalacturonan II: structure and
function of a borate cross-linked cell wall pectic
polysaccharide. Annual Review of Plant Biology 55,
109–139.
http://dx.doi.org/10.1146/annurev.arplant.55.031903
.141750
Peñuelas J, Isla R, Filella I, Araus JL. 1997.
Visible and near-infrared reflectance assessment of
salinity effects on barley. Crop Science 37(1), 198-
202.
Priestley DA. 1986. Seed aging: implications for
seed storage and persistence in the soil, Comstock
Associates.
Rehman S, Harris P, Bourne W. 1999. Effect of
artificial ageing on the germination, ion leakage and
salinity tolerance of Acacia tortilis and A. coriacea
seeds. Seed Science and Technology 27(1), 141-149.
Safarnejad A, Collin H, Bruce K, McNeilly T.
1996. Characterization of alfalfa (Medicago sativa L.)
following in vitro selection for salt tolerance.
Euphytica 92(1&2), 55–61.
http://dx.doi.org/10.1007/BF00022828
Sairam RK, Rao KV, Srivastava G. 2002.
Differential response of wheat genotypes to long term
salinity stress in relation to oxidative stress,
antioxidant activity and osmolyte concentration.
Plant Science 163, 1037-1046.
http://dx.doi.org/10.1016/S0168-9452(02)00278-9
Shalhevet J. 1993: Plant adaptation to
environmental stress. In: Fowden L, Mansfield T,
Stoddard J, ed. Plant under salt and water stress,
Chapman and Hall. New York. 133–155 p.
Sheen J. 1996. Ca2+-dependent protein kinases and
stress signal transduction in plants. Science 274,
1900-1902.
http://dx.doi.org/10.1126/science.274.5294.1900
Silberbush M, Ben-Asher J. 2001. Simulation
study of nutrient uptake by plants from soilless

272 Khomari and Javadi
Int. J. Biosci. 2015
cultures as affected by salinity buildup and
transpiration. Plant and soil 233, 59-69.
http://dx.doi.org/10.1023/A:1010382321883
Smith C, Wright G, Woodroofe M. 1988. The
effect of irrigation and nitrogen fertilizer on rapeseed
(Brassica napus L.) production in South-Eastern
Australia. Irrigation Science 9, 15-25.
http://dx.doi.org/10.1007/BF00292140
Tang PM, Dela Fuente RK. 1986. The transport of
indole-3-acetic acid in boron-and calcium-deficient
sunflower hypocotyl segments. Plant physiology 81,
646-650.
Torres-Schumann S, Godoy J, Pintor-Toro J,
Moreno F, Rodrigo R, Garcia-Herdugo G. 1989.
NaCl effects on tomato seed germination, cell activity
and ion allocation. Journal of Plant Physiology
135(2), 228–232.
http://dx.doi.org/10.1016/S0176-1617(89)80182-8
Verma S, Verma U, Tomer R. 2003. Studies on
seed quality parameters in deteriorating seeds in
Brassica (Brassica campestris L.). Seed Science and
Technology 31(2), 389–396.
http://dx.doi.org/10.15258/sst.2003.31.2.15
Yamaguchi T, Blumwald E. 2005. Developing
salt-tolerant crop plants: challenges and
opportunities. Trends in Plant Science 10, 615-620.
http://dx.doi.org/10.1016/j.tplants.2005.10.002
Yamauchi T, Hara T, Sonoda Y. 1986. Effects of
boron deficiency and calcium supply on the calcium
metabolism in tomato plant. Plant and Soil 93, 223-
230.
http://dx.doi.org/10.1007/BF02374224
Top Related