







γλώσσες
Σελίδες
Νομικός


Carlos Eduardo Cardoso Fedeli
Expressão de um gene codificador de cisteína proteinase de Leishmania (Leishmania) amazonensis em sistema bacteriano e
avaliação das respostas imunes induzidas pela proteína recombinante em modelo murino
Tese apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina para a obtenção do título de Mestre em Ciências.
São Paulo 2007

Carlos Eduardo Cardoso Fedeli
Expressão de um gene codificador de cisteína proteinase de Leishmania (Leishmania) amazonensis em sistema bacteriano e
avaliação das respostas imunes induzidas pela proteína recombinante em modelo murino
Orientadora: Profa. Dra. Clara Lúcia Barbiéri Mestriner
Tese apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina para a obtenção do título de Mestre em Ciências.
São Paulo 2007

Fedeli, Carlos Eduardo Cardoso Expressão de um gene codificador de cisteína
proteinase de Leishmania (Leishmania) amazonensis em sistema bacteriano e avaliação das respostas imunes induzidas pela proteína recombinante em modelo murino/Carlos Eduardo Cardoso Fedeli. -- São Paulo, 2007.
xv, 71f.
Tese (Mestrado) – Universidade Federal de São Paulo. Escola Paulista de Medicina. Programa de Pós-Graduação em Microbiologia e Imunologia.
Título em inglês: Expression in bacteria of a gene
encoding a cysteine proteinase from Leishmania (Leishmania) amazonensis and evaluation of the immune responses induced by the recombinant protein in a murine model.
1. Leishmania (L.) amazonensis. 2. leishmaniose cutânea.
3. cisteína proteinase. 4. resposta imune.

Trabalho realizado na Disciplina de Parasitologia do Departamento de Microbiologia, Imunologia e Parasitologia da Universidade Federal de São Paulo – Escola Paulista de Medicina, com auxílios financeiros concedidos pela Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) e Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP).

Agradecimentos
À minha orientadora Profa. Dra. Clara Lúcia Barbiéri Mestriner pelo apoio,
dedicação, paciência e por ter aberto um novo mundo para mim. Agradeço também
por transmitir seu conhecimento tornando-me um profissional melhor.
À Profa. Dra. Ieda Maria Longo Maugéri, à Juliana S. Mussalem e à Carla
Cristina Squaiella pela ajuda imprescindível nos experimentos de citometria de fluxo.
À Profa. Dra. Márcia Regina Machado dos Santos e à Luciana Girotto Gentil
pelo auxílio nos experimentos de subclonagem e expressão do gene Lacys24.
Aos amigos do laboratório Érico, Josie, Paulo, Simone e Suzana que sempre
me ajudaram na realização desse trabalho. Agradeço pelo apoio, confiança e pelas
risadas que demos juntos.
Aos meus pais Marcelo e Lígia que sempre fizeram de tudo para que eu e
meus irmãos nos tornássemos acima de tudo pessoas de bem e por sempre nos
terem incentivado ao estudo.
Aos meus queridos irmãos Marcelo, Cláudia, Teresa, Luís e Cecília que
sempre me ajudaram e me aconselharam nos momentos mais difíceis de minha
vida.
Aos meus sobrinhos Bruna, Mateo, Marcela, Alexandre, Felipe, Gabriel e
Teresa.
Ao meu avô Carlos. Aos meus avós Guilhermo, Maria e Tósca, in memorian.
Aos meus tios, Otávio, Carlito, Teresa, Vera, Mariângela, e Luís.
A todos os meus primos.

Aos meus cunhados Maurício, Shirlei e Paulinha que se tornaram meus
irmãos também.
Aos Professores da Disciplina de Parasitologia da Universidade Federal de
São Paulo, Dra. Nobuko Yoshida, Dr. Renato Arruda Mortara, Dr. José Franco da
Silveira, Dr. Maurício Martins Rodrigues e Dr. Michel P. Rabinovitch.
A todos os amigos da Disciplina de Parasitologia Fernando, Ketna, Kendi,
Solange, Miriam, Cláudio, Cecília, Emanuelle, Vanessa, Mauro, Daniele, Charles,
Silene, Renata Baida, Renata Torres, Luciana, Esteban, Roberto, Fábio, Marjorie,
Fanny, Adilson, Sandrinha, Cidinha, Dona Fátima, Mércia e Regiane.
Aos meus irmãos de escolha João, Bruno, Sandro, Alexandre, Andrey,
Juliana, Tita e Rogério, que sempre fizeram parte da minha vida.
Aos meus amigos Mônica, João e Thays por terem me deixado escrever estes
agradecimentos em horário de serviço.
À minha sogra Vera e ao meu cunhado Maurício.
Aos animais de laboratório que contribuem para o desenvolvimento da
pesquisa.
À CAPES pelo apoio financeiro para a realização desse estudo.
Para encerrar agradeço especialmente a Deus, à minha esposa Renata e ao
meu filho Pedro.

Abreviaturas
ACF Adjuvante completo de Freund
AcMo Anticorpo monoclonal
AM Amastigota
CM Controle: células + meio
DAB Diaminobenzidina
DMSO Dimetilsulfóxido
DNA Ácido desoxirribonucléico
DO Densidade óptica
EDTA Ácido etileno diamino tetracético
ELISA Ensaio imunoenzimático
g Grama
g Aceleração da gravidade (9,8 m/s2)
h Hora
IFN-γ Interferon-gama
IL-4 Interleucina-4
IL-10 Interleucina-10
IPTG Isopropil-µ-D-tiogalactopiranosídeo
kDa Kilodalton
l Litro
LB Luria Bertani
LBamp LB contendo ampicilina 100µg/mL
LPS Lipopolissacarídeo
min Minuto
M Molar
mA Miliamper
mg Miligrama
MHC Complexo principal de histocompatibilidade
ml Mililitro
mM Milimolar

mV Minivolt
ng Nanograma
OPD O-fenilenodiamina
P. acnes Propionibacterium acnes
PAGE-SDS Eletroforese em gel de poliacrilamida contendo SDS
PBS Salina em tampão fosfato 10 mM, pH 7,2
PCR Reação da polimerase em cadeia
rpm Rotações por minuto
RNA Ácido ribonucléico
SDS Dodecil sulfato de sódio
seg Segundo
TEMED N, N, N’, N’ - tetrametilenodiamina
Th1 Linfócito T helper 1
Th2 Linfócito T helper 2
TNF-α Fator de necrose tumoral-alfa
Tris Tris [hidroximetilaminometano]
U Unidade
UV Luz ultravioleta
v Volume
V Volt
µg Micrograma
µl Microlitro
µM Micromolar

Sumário
Resumo.................................................................................................................. 001
Abstract...................................................................................................................004 Introdução...............................................................................................................007 1. Aspectos gerais....................................................................................................008
2. Respostas imunes nas leishmanioses.................................................................010
3. Respostas imunes à L. (L.) amazonensis em modelo murino..............................012
4. Antígenos protetores nas leishmanioses..............................................................014
5. Cisteína proteinases de Leishmania....................................................................016
Objetivos.................................................................................................................020 Materiais e Métodos...............................................................................................022 1. Obtenção de formas amastigotas de L. (L.) amazonensis...................................023
2. Animais de experimentação.................................................................................023
3. Extração de ácidos nucléicos...............................................................................024
4. Amplificação da região codificadora do clone Lacys24 de L. (L.) amazonensis por
PCR..........................................................................................................................024
5. Bactérias e plasmídios.........................................................................................025
5.1. Bactérias.................................................................................................025
5.2. Plasmídios..............................................................................................026
6. Clonagem gene Lacys24 no vetor Topo 2.1 e subclonagem no vetor de expressão
pHIS..........................................................................................................................026
6.1. Preparação das bactérias competentes.................................................027
6.2. Transformação das bactérias competentes com o DNA plasmidial.......027
6.3. Purificação do DNA plasmidial................................................................027
7. Seqüenciamento dos clones recombinantes Topo 2.1/Lacys24......................... 028
7.1. Preparação do gel para o sistema ABI/Prisma 377................................029
8. Expressão e purificação da proteína recombinante 029
8.1. Análise de proteínas por eletroforese em gel de poliacrilamida
com SDS (PAGE-SDS)............................................................................030
9. Reatividade da proteína recombinante com anticorpos anti-cisteína proteinase.030
9.1. Produção de anticorpos policlonais anti-rLacys24.................................030
9.2. ELISA......................................................................................................031

9.3. “Western blotting”....................................................................................031
10. Avaliação da resposta imune estimulada pela rLacys24....................................032
10.1. Imunização dos animais com a rLacys24.............................................032
10.2. Análise das populações de linfócitos estimuladas pela rLacys24........032
10.3. Ensaio de citotoxicidade pelo método do cristal violeta.......................033
10.4. Ensaios por “Enzyme-linked immunospot” (ELISPOT).........................034
11. Imunização ativa.................................................................................................035
11.1. Imunização de camundongos BALB/c com a rLacys24 e desafio dos
animais com a L. (L.) amazonensis...............................................................035
11.2. Imunização de camundongos BALB/c com a rLacys24, a rLdccys1 ou os
dois antígenos recombinantes.......................................................................035
12. Análise estatística...............................................................................................036
Resultados..............................................................................................................037 1. Seqüenciamento do clone Lacys24 no vetor Topo 2.1........................................038
2. Subclonagem do gene Lacys24 de L. (L.) amazonensis em vetor de expressão e
análise da proteína recombinante............................................................................042
3. Respostas imunes celulares induzidas pela rLacys24.........................................045
4. Ensaio de citotoxicidade com linfócitos de camundongos imunizados com a
rLacys24...................................................................................................................048
5. Imunização ativa de camundongos BALB/c com a rLacys24...............................049
6. Imunização ativa com a rLacys24 e a rLdccys1 de L. (L.) chagasi......................051
Discussão................................................................................................................056 Referências Bibliográficas....................................................................................064

Resumo
Resumo
1

Resumo
O enfoque do nosso trabalho foi avaliar a antigenicidade de uma cisteína
proteinase recombinante de Leishmania (Leishmania) amazonensis e a capacidade
protetora desse antígeno contra a infecção homóloga em camundongos BALB/c.
Um fragmento de 500 pb do gene Lacys24, codificador de uma isoforma de
cisteína proteinase de L. (L.) amazonensis previamente clonado em nosso
laboratório, foi subclonado e expresso no vetor bacteriano pHis, originando uma
proteína recombinante de 24 kDa, rLacys24. Por experimentos de “Western blotting”
foi demonstrado que anticorpos produzidos contra essa proteína reconhecem uma
banda de 30 kDa do extrato de amastigotas de L. (L.) amazonensis, indicando que a
rLacys24 corresponde a uma isoforma de cisteína proteinase de 30 kDa
abundantemente expressa nesses parasitas.
A antigenicidade da rLacys24 foi avaliada pela análise das populações de
linfócitos dos camundongos BALB/c previamente imunizados com a proteína
recombinante mais Propionibacterium acnes ou adjuvante completo de Freund
(ACF) como adjuvantes. A análise por citometria de fluxo dos linfócitos provindos
dos linfonodos inguinais e poplíteos dos animais imunizados com a rLacys24 + ACF
por via subcutânea mostrou um aumento significante da expressão de linfócitos
CD8+ comparada à dos controles que receberam apenas ACF. Quanto à CD4+, não
houve diferença na expressão dessa população de linfócitos entre os controles e os
animais imunizados com a rLacys24. Por outro lado, foi baixa a expressão de
linfócitos CD4+ e CD8+ no baço dos camundongos imunizados com a rLacys24 + P.
acnes pela via intraperitoneal.
O ensaio de citotoxicidade dos linfócitos dos camundongos imunizados com a
rLacys24 + ACF foi realizado utilizando-se como células-alvo macrófagos peritoneais
de camundongos BALB/c infectados com L. (L.) amazonensis. A lise dos macrófagos
infectados na presença dos linfócitos dos animais imunizados com a rLacys24 foi
significantemente maior que a observada quando as células-alvo foram incubadas
com os linfócitos dos animais controle.
A imunização ativa dos camundongos BALB/c com a rLacys24 + ACF resultou
em pequeno, porém significante decréscimo das lesões das patas dos animais após
o desafio com a L. (L.) amazonensis, enquanto que nos animais imunizados com a
rLacys24 + P. acnes as lesões apresentaram aumento gradativo até três meses
após o desafio.
2

Resumo
3
No outro esquema de imunização avaliado, os animais foram imunizados com
a cisteína proteinase de amastigotas de L. (L.) chagasi, rLdccys, além da rLacys24,
e P. acnes como adjuvante e desafiados com a L. (L.) amazonensis. A avaliação das
respostas imunes dos animais imunizados foi realizada por citometira de fluxo e pela
detecção por ELISPOT de IFN-γ, IL-4 e IL-10 nas culturas dos linfócitos dos animais
três meses após o desafio. Em todos os animais imunizados houve predomínio da
expressão de linfócitos CD4+, enquanto que a dos linfócitos CD8+ foi baixa (em torno
de 8-10%). IFN-γ foi secretado pelos linfócitos isolados de todos os animais
estudados, não tendo havido diferenças significantes na secreção dessa linfocina
entre os animais imunizados com os antígenos recombinantes e os controles que
receberam apenas a P. acnes. Enquanto IL-10 não foi detectada nas culturas de
linfócitos de nenhum dos animais imunizados, IL-4 foi secretada pelos linfócitos de
todos os animais estudados. Entretanto, a expressão da IL-4 foi significantemente
menor nas culturas de linfócitos dos animais imunizados com os antígenos
recombinantes comparada à dos controles. Não foram observadas diferenças do
tamanho das lesões das patas dos camundongos imunizados com os antígenos
recombinantes em relação aos controles, sendo esses resultados confirmados
quando a infecção dos animais foi avaliada pelo crescimento em meio axênico dos
parasitas isolados das lesões.
Apesar da reduzida proteção conferida pela rLacys24, os dados do presente
trabalho sugerem que esse antígeno pode ser útil em novos esquemas de
imunização que induzam respostas mediadas por linfócitos CD4+ e CD8+ mais
eficazes contra a infecção pela L. (L.) amazonensis.

Abstract
4

Abstract
Our work focused on the antigenicity of a recombinant cysteine proteinase
from Leishmania (Leishmania) amazonensis and on the protective role of this antigen
in the infection of BALB/c mice with the parasite.
A 500 bp fragment from the Lacys24 gene, encoding an isoform of cysteine
proteinase from L. (L.) amazonensis previously cloned in our laboratory, was
subcloned and expressed in the pHis vector, resulting in a recombinant protein of 24
kDa, rLacys24. In Western blots of L. (L.) amazonensis extracts, antibodies directed
to the rLacys24 antigen recognized a 30 kDa band identified as an isoform of
cysteine proteinase abundantly expressed in these parasites.
The antigenicity of the rLacys24 was evaluated by analysis of the T
lymphocyte profile in BALB/c mice previously immunized with this recombinant
protein plus either Propionibacterium acnes or Freund’s complete adjuvant (CFA).
Lymphocytes isolated from the popliteal and inguinal lymph nodes of animals
immunized with rLacys24 plus CFA by the subcutaneous route were analysed by
fluorescence-activated cell sorter (FACS). A significantly higher expression of CD8+
lymphocytes was observed in animals immunized with rLacys24 plus CFA compared
to the controls which received CFA alone. In contrast, a low expression of CD4+
lymphocytes was observed in these animals. On the other hand, the expression of
CD4+ and CD8+ lymphocytes there was lower in spleens from animals immunized
with rLacys24 plus P. acnes by intraperitoneal route.
The cytotoxicity of lymphocytes isolated from mice immunized with rLacys24
plus CFA was assyed by use of L. (L.) amazonensis-infected macrophages as the
target cells. The cytotoxicity of lymphocytes from rLacys24-immunized mice was
significantly higher than that observed when the target cells were incubated with
lymphocytes from control animals.
Active immunization of BALB/c mice with rLacys24 plus CFA resulted in a low
but significant decrease of foot lesions after challenge with L. (L.) amazonensis in
comparison with the lesion size in control mice. In contrast, there was a similar
increase of foot lesions among mice immunized with rLacys24 plus P. acnes or with
P. acnes alone.
In another immunization protocol, animals were immunized with a recombinant
cysteine proteinase from L. (L.) chagasi, rLdccys1, besides rLacys24, plus P. acnes
as the adjuvant. Immune responses were evaluated by FACS and ELISPOT for
detection of IFN-γ, IL-4 and IL-10 in lymphocyte cultures from animals three months
5

Abstract
6
after challenge with L. (L.) amazonensis. All of the immunized animals showed a
predominance of T CD4+ lymphocytes and a low expression of CD8+ (8-10%). IFN-γ
was secreted by lymphocytes isolated from all immunized animals and there were no
significant differences of IFN-γ secretion among animals immunized with the
recombinant antigens or with P. acnes alone. IL-10 was not detected in lymphocyte
cultures from the immunized animals. However, IL-4 expression was significantly
lower in lymphocyte cultures from animals immunized with the recombinant antigens
compared to the controls. There were no significant differences in foot lesion size
from BALB/c mice immunized with the recombinant antigens compared to the
controls; these results were confirmed by estimates of parasites isolated from the foot
lesions.
In spite of the low protection conferred by the rLacys24, our data suggest that
this recombinant antigen may be useful in new immunization schedules aiming to
elicit more efficient CD4+ and CD8+ protective responses against L. (L.) amazonensis
infection.

Introdução
7

Introdução
1. Aspectos gerais As leishmanioses compreendem um grupo de parasitoses causadas por várias
espécies de protozoários do gênero Leishmania. Estima-se que existam atualmente 12
milhões de casos de leishmanioses, com mortalidade anual de aproximadamente
60.000. Segundo a Organização Mundial de Saúde as leishmanioses ocorrem em 88
países e sua notificação é compulsória em apenas 30 deles. Anualmente surgem 2
milhões de novos casos e 350 milhões de pessoas encontram-se sob o risco de
adquirir alguma forma da doença (WHO, 2001). A coinfecção Leishmania/HIV agrava
ainda mais esse quadro, considerando-se o número de pessoas HIV positivas
(Desjeux, 2004).
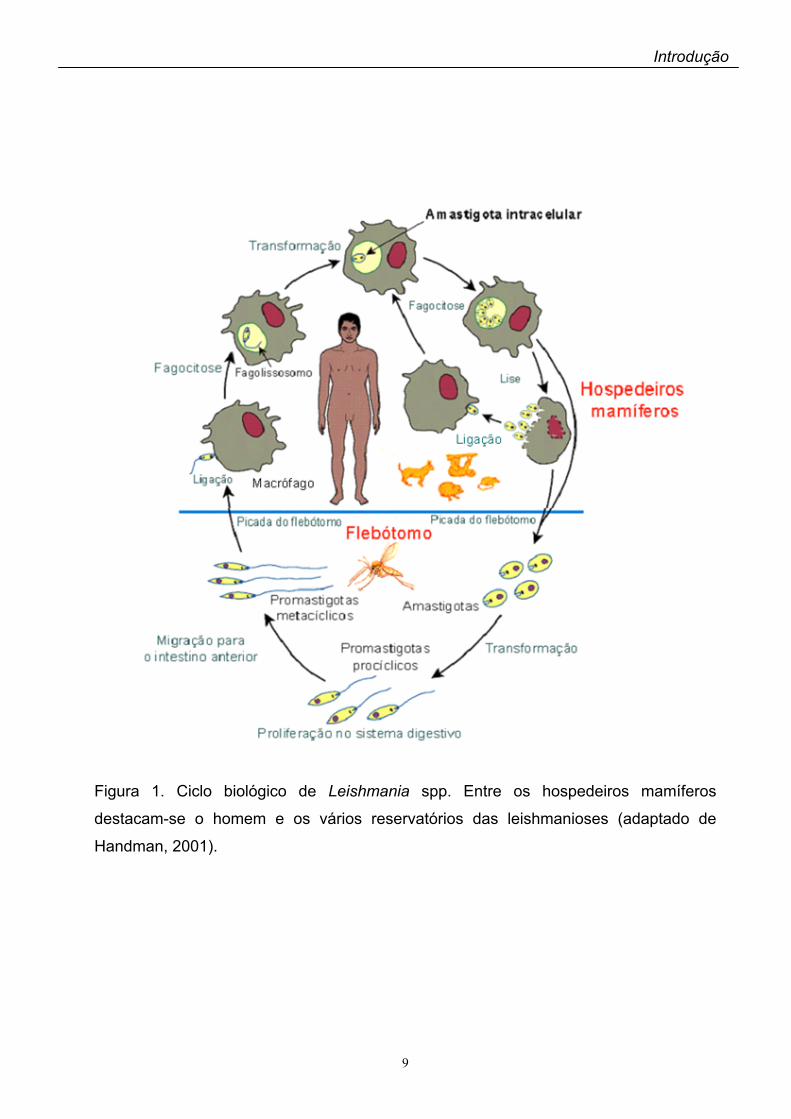
O gênero Leishmania pertence à ordem Kinetoplastida, família
Trypanosomatidae, e compreende parasitas digenéticos que se desenvolvem
alternadamente em hospedeiros vertebrados mamíferos e insetos vetores. Nos
hospedeiros mamíferos, entre eles o homem, os parasitas assumem a forma
amastigota, arredondada e imóvel, que se multiplica obrigatoriamente dentro de células
do sistema monocítico fagocitário em um vacúolo parasitóforo. À medida que os
amastigotas se multiplicam por divisão binária os macrófagos se rompem liberando os
parasitas que são fogocitados por outros macrófagos. Os vetores das leishmanioses
são representados por insetos dípteros da sub-família Phlebotominae (gênero
Lutzomya nas Américas e Phlebotomus no Velho Mundo). Nos flebotomíneos os
parasitas vivem no trato digestivo onde as formas amastigotas, ingeridas durante o
repasto sangüíneo em um hospedeiro mamífero infectado, diferenciam-se em
promastigotas, formas alongadas, flageladas e móveis. Os promastigotas são
inoculados pela fêmea do vetor juntamente com a saliva na pele dos mamíferos
durante a picada, completando o ciclo. Além do homem, vários mamíferos são
suscetíveis à infecção por Leishmania. Entre esses alguns representam reservatórios
importantes das leishmanioses, principalmente os roedores silvestres, edentados (tatu,
tamanduá, preguiça), marsupiais (gambá) e canídeos. O ciclo biológico de Leishmania
está representado na Figura 1.
8
Introdução
Figura 1. Ciclo biológico de Leishmania spp. Entre os hospedeiros mamíferos
destacam-se o homem e os vários reservatórios das leishmanioses (adaptado de
Handman, 2001).
9

Introdução
Existem mais de 20 espécies de Leishmania capazes de infectar o homem e sua
diferenciação se baseia em características extrínsecas (quadro clínico da doença,
distribuição geográfica, epidemiologia) e intrínsecas (tamanho, forma e estrutura
molecular dos parasitas).
As várias formas clínicas das leishmanioses dependem da espécie do parasita,
carga genética do hospedeiro e imunidade adquirida durante o desenvolvimento da
doença. As formas cutâneas, caracterizadas por lesões de pele, são causadas por
espécies pertencentes aos complexos Leishmania (Leishmania) mexicana e
Leishmania (Viannia) braziliensis nas Américas [L. (L.) amazonensis, L. (V.) guyanensis
e L. (V.) braziliensis no Brasil] e ao complexo L. (L.) tropica no Velho Mundo [L. (L.)
tropica e L. (L.) major, entre outras]. A forma cutânea difusa, que apresenta lesões
crônicas e disseminadas, no Brasil é causada pela L. (L.) amazonensis, enquanto que
o agente da leishmaniose mucocutânea, forma invasiva e mutilante, é a L. (V.)
braziliensis em nosso país. A forma visceral, também conhecida como calazar, é
causada por espécies do complexo Leishmania (Leishmania) donovani [L. (L.)
donovani e L. (L.) infantum no Velho Mundo e L. (L.) chagasi nas Américas].
Os últimos dados epidemiológicos registrados pela Organização Mundial da
Saúde revelam que os países em que se encontram mais de 90% dos casos de
leishmanioses cutâneas são Irã, Arábia Saudita, Afeganistão, Síria, Argélia, Brasil e
Peru e que 90% dos casos de leishmaniose visceral estão concentrados em
Bangladesh, Brasil, Índia, Nepal e Sudão.
O grande número de casos existente e o fato de representarem parasitoses cuja
incidência está aumentando tornam as leishmanioses um grande desafio de saúde
pública e justificam o investimento que tem sido aplicado para o estudo dessas
parasitoses.
2. Respostas imunes nas leishmanioses
De forma geral, as respostas protetoras contra as leishmanioses estão
associadas à imunidade clássica mediada por células, enquanto que a suscetibilidade é
relacionada a uma forte resposta humoral na ausência de resposta celular. A resposta
imune nas leishmanioses é dependente da espécie do parasita e do genótipo do
hospedeiro e várias linhagens de camundongos são utilizadas nos estudos de
10

Introdução
imunidade à Leishmania. O modelo murino utilizado como o protótipo de suscetibilidade
ou resistência à Leishmania foi estabelecido com a espécie Leishmania (L.) major.
Nesse modelo, observou-se que camundongos resistentes desenvolvem resposta
mediada por linfócitos T CD4+ da subpopulação Th1, produtores de IFN-γ, IL-2 e TNF-α
(Heinzel et al, 1989; Wang et al, 1994). O IFN-γ ativa macrófagos que, por sua vez,
passam a produzir óxido nítrico (NO) e produtos reativos de oxigênio (H2O2 e O2-) que
são os principais compostos envolvidos com a morte dos amastigotas intracelulares
(Murray et al., 1982; Liew et al., 1990). Por outro lado, em camundongos suscetíveis a
resposta é mediada por linfócitos T da subpopulação Th2, produtores de IL-4, IL-5, IL-
10 e IL-13, resultando na inativação de macrófagos (Sadick et al, 1990; Chatelain et al,
1992; Kopf et al, 1996).
Em L. (L.) major, os estudos sugerem que o fenótipo de suscetibilidade, com a
geração de células de perfil Th2, não é derivado do fenômeno de produção precoce de
IL-4 em resposta a antígenos parasitários, mas sim da falha em produzir ou responder
à IL-12 (revisado por McMahon-Pratt & Alexander, 2004). Considerando o ambiente de
citocinas gerado durante a resposta imune, a IL-12 parece ser primordial na indução do
fenótipo resistente em camundongos. A depleção de IL-12, por meio de técnicas
genéticas ou de neutralização por anticorpos, mostra que os camundongos resistentes
adquirem um fenótipo suscetível, enquanto que os animais suscetíveis quando tratados
com IL-12 desenvolvem resistência à infecção por L. (L.) major. No ambiente da
resposta imune inata a IL-12, produzida principalmente por células dendríticas e às
vezes com o auxílio da IL-18, ativa células NK (“Natural Killer”) a produzirem IFN-γ que
influencia não somente a resposta inata, mas também a resposta Th1. O IFN-γ medeia
a proteção induzindo a expressão da iNOS2 (óxido nítrico sintase-2 indutível) e a
produção de NO pelos macrófagos (revisado por Scott et al., 2004).
A IL-10 produzida por linfócitos Th2 está intimamente relacionada à
suscetibilidade. Em experimentos com camundongos BALB/c suscetíveis à infecção
por L. (L.) major foi demonstrado que aqueles deficientes em IL-10 eram capazes de
controlar a doença, desenvolvendo lesões relativamente pequenas. Quando foi
estudado o mecanismo de resposta de macrófagos à infecção por L. (L.) major in vitro,
observou-se que os amastigotas opsonizados por IgG murino eram fagocitados pelos
macrófagos via receptor Fcγ com produção de grandes quantidades de IL-10, indicando
que a sobrevivência e multiplicação dos amastigotas seriam facilitadas pela
11

Introdução
desativação dos mecanismos microbicidas do macrófago sob o efeito da IL-10 (Kane &
Mosser, 2001).
Considerando que a IL-10 é tão importante quanto a IL-4, resta saber se são
apenas os linfócitos T CD4+ de perfil Th2 que estão produzindo essa citocina ou se
outras populações de linfócitos T também contribuem para essa produção, culminando
na suscetibilidade gerada em camundongos. Uma possibilidade para a fonte de IL-10
são os linfócitos T CD4+CD25+, denominados células reguladoras (Treg), que ocorrem
naturalmente em modelos suscetíveis e resistentes à infecção por L. (L.) major. Essas
células constituem 5-10% das células do sangue periférico de seres humanos e
camundongos e expressam constitutivamente a cadeia α do receptor da IL-2 (CD25).
Em camundongos BALB/c suscetíveis à infecção por L. (L.) major essas células
reguladoras agem prevenindo uma resposta exarcebada do tipo Th2, enquanto que nos
camundongos C57BL/6, resistentes, elas controlam a resposta imune protetora Th1,
permitindo a persistência parasitária e ao mesmo tempo mantendo a resposta imune
efetora de memória (Belkaid, 2003).
3. Respostas imunes à L. (L.) amazonensis em modelo murino
A L. (L.) amazonensis é responsável por uma forma cutânea de leishmaniose,
porém as respostas imunes desencadeadas contra essa espécie apresentam um
padrão bem diferente do descrito para L. (L.) major. Ao contrário dos camundongos
infectados com L. (L.) major, em que ocorre uma diferenciação polarizada a Th1 e Th2,
respectivamente, em animais resistentes e suscetíveis, os camundongos suscetíveis à
L. (L.) amazonensis não apresentam uma resposta Th2 exacerbada (Afonso e Scott,
1993; Soong et al.,1996) e são capazes de gerar respostas do tipo Th1 (Afonso e Scott,
1993; Ji et al., 2002; Soong et al., 1997). Foi também determinado que a deleção
dirigida dos genes codificadores da IL-4 ou IL-10 em camundongos C57BL/6 infectados
com L. (L.) amazonensis não afeta o desenvolvimento das lesões e as cargas
parasitárias desses animais (Jones et al., 2000, Jones et al., 2002). Recentemente, foi
observada a expressão diminuída da cadeia β2 do receptor da IL-12 dos linfócitos
CD4+ (IL-12R β2) em camundongos C3H infectados com L. (L.) amazonensis, assim
como em linfócitos CD4+ de camundongos C57BL/6 depletados de IL-4. Esses
resultados sugerem que um mecanismo independente de IL-4 é responsável pela
resposta reduzida a IL-12 e a conseqüente resposta Th1 prejudicada nos animais
12

Introdução
infectados com L. (L.) amazonensis (Jones et al., 2000). Entretanto, o tratamento dos
camundongos com IFN-γ (Barral-Neto et al., 1996) ou IL-12 (Jones et al., 2000) não
teve efeito significante sobre o curso da infecção com L. (L.) amazonensis nesses
animais. Uma vez que essas duas citocinas induzem a expressão da cadeia IL-12R β2,
esses dados sugerem que os mecanismos imunes responsáveis pela suscetibilidade
dos camundongos à L. (L.) amazonensis são complexos e requerem mais investigação.
Mais recentemente foi avaliado o possível envolvimento das células CD4+CD25+
regulatórias (Treg) na infecção e progressão da leishmaniose causada pela L. (L.)
amazonensis. Nesse estudo foi observado um aumento das células Treg juntamente
com a expressão aumentada de FoxP3, TGF-β e IL-10R nas lesões das patas e
linfonodos regionais de camundongos C57BL/6 infectados. Recipientes singênicos, nos
quais foram transferidas as células Treg de camundongos C57BL/6 infectados,
apresentaram redução significante das lesões após a infecção com L. (L.)
amazonensis. Esses resultados sugerem que as células Treg podem limitar a
progressão da doença causada por esse parasita, porém esse efeito benéfico foi
transitório e correlacionado com a ativação diminuída de células T CD4+CD25-
produtoras de IFN-γ (Ji et al., 2005). Esses dados mostram a importância de um
balanço fino entre as células Treg e as T efetoras na suscetibilidade do hospedeiro à L.
(L.) amazonensis.
A participação de linfócitos CD8+ como células produtoras de IFN- γ nas
respostas imunes à infecção por L. (L.) amazonensis também foi demonstrada em
camundongos C57BL/6 (Chan, 1983). A caracterização de linhagens de linfócitos T
CD8+ específicas ao antígeno GP46/M-2, capazes de reconhecer macrófagos
infectados com L. (L.) amazonensis, demonstrou o envolvimento desses linfócitos nas
respostas a esse antígeno utilizado para a imunização protetora de camundongos CBA
contra a infecção homóloga (Champsi e McMahon-Pratt, 1988; Kima et al., 1997).
Proteção significante contra a L. (L.) amazonensis foi observada em camundongos
C57BL/6 imunizados com o antígeno P8 associado à membrana da L. (L.) pifanoi,
tendo sido demonstrada a participação de linfócitos CD4+ e CD8+ nas respostas
protetoras desses animais imunizados. Nesse trabalho foi demonstrada a produção de
perforina, além de IFN-γ, pelos linfócitos CD8+ no sítio da infecção, sendo ambas as
moléculas efetoras na proteção dos animais vacinados (Colmenares et al., 2003).
13

Introdução
4. Antígenos protetores nas leishmanioses
Vários antígenos têm sido descritos quanto à sua capacidade de induzir
respostas imunes protetoras e resultados satisfatórios de imunização com antígenos
nativos e recombinantes, assim como com os genes codificadores de alguns desses
antígenos, têm sido alcançados quando fatores tais como a dose do antígeno, a via de
imunização e os adjuvantes são bem estabelecidos.
O antígeno LACK possui um peso molecular de 36 kDa e é expresso nas formas
amastigota e promastigota, sendo altamente conservado entre as várias espécies de
Leishmania (Mougneau et al., 1995). A imunização com o DNA do LACK foi capaz de
induzir imunidade prolongada por levar à produção de IFN-γ por linfócitos T CD8+,
sendo essa imunidade abolida após o tratamento dos animais imunizados com anti-
CD8 (Gurunathan et al., 2000). A administração intranasal com o gene LACK induziu
imunidade protetora de longa duração contra a infecção por L. (L.) amazonensis em
camundongos BALB/c (Pinto et al., 2004). Mais recentemente a imunização intranasal
como gene LACK complexado a microesferas de quitosana reduziu o
desenvolvimemnto das lesões e a carga parasitária de camundongos BALB/c
desafiados com a L. (L.) amazonensis (Costa-Souza et al., 2007).
A imunização com a forma nativa ou recombinante da gp63, a metaloprotease
de superfície presente em promastigotas de todas as espécies de Leishmania, foi
capaz de conferir maior proteção contra a infecção por L. (L.) amazonensis quando
comparada à infecção por L. (L.) major (Handman, 2001) e a imunização com um
peptídeo sintético da gp63 administrado subcutaneamente foi eficaz no controle da
infecção por L. (L.) major (Spitzer et al., 1999). A imunização de camundongos BALB/c
com bacilo Calmette-Guerin (BCG) (Connel et al, 1993) ou Salmonella (AroA-) (Yang et
al, 1990) expressando a gp63 de L. (L.) major induziu proteção contra o desafio
homólogo ou na infecção por L. (L.) mexicana. Resultados semelhantes foram obtidos
com a gp63 recombinante administrada com IL-12 como adjuvante em animais
desafiados com L. (L.) mexicana (Aebischer et al, 2000). A imunização de
camundongos por injeção intramuscular de DNA da gp63 também conferiu proteção
significante contra o desafio com L. (L.) major e L. (L.) mexicana, caracterizada como
Th1 (Xu e Liew, 1995; Walker et al, 1998; Dumonteil et al, 2003; Ahmed et al, 2004).
O antígeno PSA-2 (antígeno 2 de superfície do parasita) é expresso nas várias
espécies de Leishmania, exceto na L. (V.) braziliensis. Há evidências de que a
14

Introdução
imunização com o PSA-2 de L. (L.) donovani protege camundongos contra a infecção
por L. (L.) major (Handman et al., 1995). Camundongos imunizados com o DNA
codificador do PSA-2 e desafiados com L. (L.) major desenvolvem lesões menores e
com menos parasitas quando comparadas às de camundongos não imunizados,
gerando uma população definida de células T CD4+ secretoras de citocinas do perfil
Th1 (Ahmed et al., 2004).
Antígenos nucleares têm se mostrado bastante eficazes no controle das
leishmanioses cutâneas. Vacinas de DNA utilizando genes que codificam as histonas
H2A, H2B, H3 e H4 de L. (L.) infantum contra a infecção por L. (L.) major foram
testadas. Os animais imunizados desenvolveram uma forte resposta Th1, com o
aumento da produção de IFN-γ, assim como de anticorpos do isótipo IgG2a, e tanto as
células T CD4+ como as T CD8+ contribuem para a proteção contra a L. (L.) major
(Iborra et al., 2004). Outro antígeno que gerou bons resultados de proteção foi a P4
nuclease. Camundongos da linhagem BALB/c foram imunizados com o DNA cujas
seqüências codificam a P4 nuclease + IL-12 ou HSP/70 como adjuvante e
posteriormente desafiados contra L. (L.) major e L. (L.) amazonensis. Os animais
imunizados com a P4 nuclease + IL-12 e desafiados com a L. (L.) amazonensis
apresentaram proteção, ao contrário dos animais imunizados com a P4 nuclease +
HSP/70. Por outro lado, os animais imunizados com a P4 nuclease + HSP/70
desenvolveram proteção frente ao desafio com a L. (L.) major (Campbell et al., 2003).
O antígeno A2 de L. (L.) donovani foi identificado como uma família de proteínas
específica da forma amastigota e cujos genes são conservados entre as espécies dos
complexos L. (L.) donovani e L. (L.) mexicana. Camundongos BALB/c imunizados com
a proteína A2 recombinante (rA2) ou com o gene A2 foram protegidos contra a infecção
por L. (L.) donovani (Ghosh et al., 2001; Ghosh et al., 2002). Mais recentemente foi
demonstrada também proteção significante em camundongos BALB/c contra o desafio
com L. (L.) amazonensis pela imunização desses animais com a rA2 mais rIL-12
(Coelho et al., 2003). A imunização com o complexo ligante de fucose-manose (FML)
purificado de L. (L.) donovani (uma nucleosídeo hidrolase de 36 kDa denominada
NH36) foi capaz de retardar significantemente o desenvolvimento das lesões por L. (L.)
mexicana em camundongos BALB/c (Aguilar-Be et al., 2005)
15

Introdução
5. Cisteína proteinases de Leishmania
Nos últimos anos o estudo de enzimas proteolíticas de parasitas tem aumentado
consideravelmente pelo fato de essas moléculas constituírem importantes fatores de
virulência diretamente relacionados com a interação parasita-hospedeiro e
representarem alvos promissores para a imuno e quimioterapia (McKerrow, 1989;
Selzer et al, 1999, Das et al, 2001; Sajid e McKerrow, 2002). Entre os protozoários, as
principais enzimas proteolíticas caracterizadas pertencem ao grupo das cisteína
proteinases, sendo a maioria delas membros da superfamília da papaína. Essa
superfamília, por sua vez, é subdividida em duas famílias, a das proteases do tipo
catepsina e as do tipo calpaína. As proteases do tipo catepsina são ainda sub-divididas
nas sub-famílias das catepsinas dos tipos B e L. Essas guardam grandes semelhanças
entre si e apresentam alguns motivos na seqüência de aminoácidos que as
diferenciam, sendo ambas sintetizadas como zimogênios inativos que são
posteriormente processados pela remoção da pré e pró-região para que se tornem
enzimas ativas (Eakin et al, 1992).
Em Leishmania há predominância de cisteína proteinases do tipo catepsina L
que são expressas em grande quantidade nas formas amastigotas e estão localizadas
preferencialmente em lisossomos diferenciados denominados megassomos, descritos
primeiramente em espécies do complexo L. (L.) mexicana (Pupkins et al., 1986) e mais
recentemente em L. (L.) chagasi (Alberio et al., 2004).
Em L. (L.) mexicana a maior atividade proteolítica detectada é a das cisteína
proteinases expressas pelos genes cpb, arranjados em 19 cópias repetidas em
"tandem". Os genes cpb3 a cpb18 codificam proteínas que possuem uma extensão C-
terminal (CTE), sendo expressos predominantemente nas formas amastigotas,
enquanto que a cisteína proteinase codificada pelo gene cpa não possui CTE (Mottram
et al, 1997; Brooks et al., 2000). Outras cisteína proteinases bem caracterizadas em
Leishmania são as codificadas pelos genes Lpcys1 e Lpcys2 em L. (L.) pifanoi
(Duboise et al., 1994) e as CPb e CPa de L. (L.) major (Sakanari et al., 1997). Nas
espécies visceralizantes são expressas as cisteína proteinases do tipo catepsina L de
L. (L.) chagasi codificadas pelos genes Ldccys1 e Ldccys2 e que apresentam
seqüências correspondentes em L. (L.) infantum (cpa e cpb) e L. (L.) donovani
(Lddcys1 e Lddcys2) (Omara-Opyene e Gedamu, 1997; Mundodi et al, 2002).
16

Introdução
Embora não esteja clara a função das cisteína proteinases em Leishmania, seu
papel tem sido intensamente estudado e sabe-se que elas têm grande participação na
virulência do parasita. A análise dessas enzimas em cepas avirulentas e virulentas de
promastigotas de L. (L.) amazonensis mostrou que elas são preferencialmente
expressas nas cepas virulentas (Soares et al, 2003). Camundongos suscetíveis
infectados com L. (L.) mexicana em que o gene da cisteína proteinase cpb foi
nocauteado apresentaram lesões consideravelmente menores comparadas às dos
animais infectados com parasitas selvagens e parasitas nulos para os genes cpa e cpb
foram incapazes de desenvolver lesões nesses animais. Interessantemente, a
detecção das linfocinas nesses animais mostrou que houve mudança de uma resposta
predominantemente Th2 para Th1, com produção de IFN-γ e IL-2 (Alexander et al,
1998). A análise funcional das catepsinas do tipo B de L. (L.) donovani e L. (L.) chagasi
mostrou que elas são capazes de clivar a forma latente do TGF-β1, convertendo-o à
forma ativa inibidora do macrófago (Somanna et al, 2002). Esses resultados indicam
que essas enzimas não só agem como fatores de virulência, mas também como
moduladoras da resposta imune do hospedeiro.
As cisteínas proteinases têm representado também um grande alvo para o
diagnóstico das leishmanioses visceral e cutânea e o estudo de vacinas. As CPA e
CPB de L. (L.) infantum foram utilizadas no Irã para o diagnóstico por ELISA da
leishmaniose visceral humana e canina (Rafati et al, 2003). Em relação à
antigenicidade para respostas imunes celulares foi demonstrada a capacidade de as
cisteína proteinases induzirem resposta do tipo Th1 após a morte dos amastigotas de
L. (L.) mexicana no interior de macrófagos peritoneais de camundongos e a
apresentação de epítopos dos parasitas na sua superfície (Wolfram et al, 1995).
Camundongos BALB/c imunizados com a cisteína proteinase de 24 kDa de L. (L.)
major, CPa, desenvolveram resistência à infecção homóloga. Esses camundongos
produziram altos níveis de IFN-γ, gerando uma resposta imune Th1 com conseqüente
resolução da doença (Rafati et al., 2000). Proteção duradoura contra a infecção
homóloga também foi observada em camundongos BALB/c imunizados com uma
mistura dos genes cpa e cpb de L. (L.) major, assim como com as cisteína proteinases
recombinantes (rCPb e rCPa) (Rafati et al., 2001). A análise da resposta imune nesses
animais mostrou que eles apresentaram predomínio de linfócitos Th1 com produção de
IFN-γ. Mais recentemente, a imunização de camundongos BALB/c com uma mistura
17

Introdução
dos genes cpa e cpb codificadores de cisteína proteinases de L. (L.) infantum, seguida
por um reforço com as proteínas recombinantes rCPa e rCPb juntamente com CpG
ODN e Montanide como adjuvantes, resultou em proteção contra a leishmaniose
visceral nos animais imunizados mediada por linfócitos Th1 (Rafati et al., 2006). Em
humanos foi demonstrado que as cisteínas proteinases CPa e CPb de L. (L.) major
induziram alta produção de IFN-γ e IL-2 e baixa secreção de IL-10 em culturas de
linfócitos periféricos de pacientes com leishmaniose cutânea causada por L. (V.)
guyanensis (Pascalis et al, 2003).
Nosso laboratório tem desenvolvido estudos sobre antígenos de formas
amastigotas de L. (L.) chagasi e de L. (L.) amazonensis e a capacidade de esses
antígenos desencadearem respostas imunes protetoras em modelo murino. Dentro
dessa linha, foram identificados alguns antígenos que induzem respostas
linfoproliferativas em camundongos BALB/c previamente imunizados com Leishmania.
Entre eles um, de massa molecular aparente de 30 kDa (p30) presente em amastigotas
de L. (L.) amazonensis e L. (L.) chagasi, induz preferencialmente linfócitos T CD4+ da
subpopulação Th1 e significante proteção contra a infecção homóloga (Beyrodt et al,
1997; Pinto et al, 2000). A caracterização bioquímica desse antígeno em L. (L.)
amazonensis foi feita após a sua purificação em coluna de imunoafinidade com um
anticorpo monoclonal dirigido à p30 (AcMo 2E5D3). O antígeno purificado foi submetido
à PAGE-SDS em gel copolimerizado com gelatina e utilizando-se o inibidor E-64
demonstrou-se que a p30 de L. (L.) amazonensis apresenta forte atividade de cisteína
proteinase (Beyrodt et al., 1997). Visando à clonagem do gene p30 de L. (L.)
amazonensis, foram obtidos vários clones de 500 pb e a análise da seqüência desses
clones mostrou que alguns apresentam alto grau de identidade com as seqüências de
genes de cisteína proteinases de Leishmania previamente descritos. A subclonagem
desses clones em vetor de expressão mostrou que algumas das proteínas
recombinantes geradas são capazes de desencadear respostas linfoproliferativas
parcialmente protetoras em camundongos BALB/c contra a infecção por L. (L.)
amazonensis (Beyrodt et al., 1996; Beyrodt, 1998; Oliveira, 2001). O racional do
presente trabalho foi estudar um desses clones, Lacys24, em cuja seqüência foram
identificados três prováveis epítopos de ligação ao MHC de classe II e um de classe I.
Esse estudo envolveu a subclonagem do clone Lacys24 em vetor bacteriano e a
avaliação das respostas imunes e o grau de proteção desencadeados pela proteína
18

Introdução
19
recombinante (rLacys24) em camundongos BALB/c após o desafio com L. (L.)
amazonensis.

Objetivos
20

21
Os principais objetivos do nosso trabalho podem ser assim definidos: 1) Subclonagem do gene Lacys24 codificador de uma isoforma de cisteína
proteinase de 30 kDa de amastigotas de Leishmania (Leishmania) amazonensis em
vetor de expressão bacteriano e purificação da proteína recombinante (rLacys24).
2) Determinação da antigenicidade da rLacys24 para respostas imunes celulares em
camundongos BALB/c e caracterização das populações de linfócitos envolvidas
nessas respostas.
3) Ensaios de imunização de camundongos BALB/c com a rLacys24 e avaliação das
respostas imunes e do grau de proteção gerados após a infecção dos animais pela
L. (L.) amazonensis.

Materiais e Métodos
20

Materiais e Métodos
1. Obtenção dos amastigotas de L. (L.) amazonensis
A cepa MHOM/BR/71973/M2269 de Leishmania (L.) amazonensis utilizada foi
proveniente do Instituto Evandro Chagas (Belém, PA, Brasil) e gentilmente cedida
pelo Dr. Jeffrey J. Shaw.
Amastigotas de L. (L.) amazonensis foram isolados de lesões de patas de
hamsters previamente infectados (5x107 parasitas/pata). Os granulomas obtidos
após 8 semanas de infecção foram retirados, colocados em PBS, raspados com
bisturi e o homogenado centrifugado a 2.500 x g por 10 min. O sobrenadante obtido
foi centrifugado a 1.400 x g durante 10 min e ao precipitado resultante adicionou-se
uma solução de ACK (NH4Cl 155 mM, KHCO3 10 mM, EDTA 0,1 mM) para a lise das
hemácias e após 15 min foi realizada uma nova centrifugação a 1.400 x g. Os
amastigotas foram ressuspensos em PBS e mantidos sob agitação à temperatura
ambiente durante 4 horas na presença de antibiótico. Após esse período a
suspensão foi centrifugada a 1.400 x g por 10 min, o precipitado final foi
ressuspenso em PBS e os amastigotas foram contados em câmara de Neubauer
(Barbiéri et al., 1990).
2. Animais de experimentação
Para a manutenção dos amastigotas de L. (L.) amazonensis foram utilizados
hamsters de 8 a 10 semanas de idade obtidos do Centro de Desenvolvimento de
Modelos Experimentais para Medicina e Biologia (CEDEME) da Universidade
Federal de São Paulo - Escola Paulista de Medicina.
As linhagens de camundongos isogênicos BALB/c utilizados nos
experimentos, com idade entre 6 e 8 semanas, também foram cedidos pelo
CEDEME.
Todos os procedimentos foram aprovados pelo Comitê de Ética da
Universidade Federal de São Paulo – Escola Paulista de Medicina.
23

Materiais e Métodos
3. Extração de ácidos nucléicos
Os amastigotas de L. (L.) amazonensis (1x108) foram incubados em 150µl de
tampão de lise (Tris-HCL 50 mM, pH 8,0, EDTA 62,5 mM, pH 9,0, Triton X-100 4%)
por 5 min a 37°C e o DNA genômico extraído com 150 µl de fenol-clorofórmio (1:1
v/v). A amostra foi centrifugada a 11.000 x g por 5 min e o DNA precipitado
acrescentando-se 2 v de etanol absoluto à fase aquosa. A amostra foi novamente
centrifugada a 11.000 x g por 10 min e o precipitado lavado com etanol 70% e
ressuspenso em 50 µl de TE (Tris-HCL 10 mM, pH 8,0, EDTA 0,1 M) contendo
RNAse 0,6 µg/µl. Após incubação de 30 min a 37°C, o DNA foi precipitado com 3 v
de etanol absoluto e acetato de sódio 0,3 M, lavado com etanol 70% e ressuspenso
em 50 µl de TE (Medina-Acosta e Cross, 1993)
4. Amplificação da região codificadora do clone Lacys24 de L. (L.)
amazonensis por PCR
A região codificadora do fragmento de 500 pb do gene de cisteína proteinase
de L. (L.) amazonensis (Lacys24) foi amplificada utilizando-se “primers” baseados
nas seqüências determinadas em estudos anteriores em nosso laboratório (Beyrodt,
1998; Lasakosvitsch et al., 2003). Os “primers sense” (5’– ATG GAT GGG GAT GGA
AGG GGA ATGT–3’) e “antisense” (5’-GGG AAT TGG ATG AGT TTT TTG AGG
ATGG-3’), contendo seqüências para sítios de restrição HindIII e EcoRI, foram
utilizados para a clonagem dirigida. Para a reação foram utilizados 100 ng de DNA
genômico de L. (L.) amazonensis, 100 pmol de cada “primer”, 0,2 mM da mistura de
dNTPs, 1U de Taq DNA polimerase e tampão para PCR (Tris-HCL 10 mM, pH 9.0,
KCL 50 mM, MgCl2 2 mM) em um volume final de 50 µl. As condições de
amplificação foram: 30 ciclos a 94°C por 1min, a 51°C por 30 seg e a 72°C por 30
seg. As amostras foram analisadas em gel de agarose a 1%.
Para a preparação do gel a agarose foi fundida em tampão TBE (Tris-HCL
0,01 M, EDTA 0,02 mM, ácido bórico 0,086 M) e à mistura foi acrescentado brometo
de etídio 0,5 µg/ml. As amostras foram aplicadas no gel após serem
24

Materiais e Métodos
homogeneizadas em tampão de amostra para DNA (glicerol 5%, xileno cianol 0,04%,
azul de bromofenol 0,04%, EDTA 2mM) e a eletroforese realizada a 80 V em tampão
TBE. As bandas de DNA foram visualizadas sob luz UV e o fragmento de interesse
recortado do gel e purificado segundo o protocolo do kit Wizard (Promega).
5. Bactérias e plasmídios 5.1. Bactérias
As diferentes cepas de Escherichia coli foram crescidas e mantidas a 4°C em
placas de Petri contendo meio LB sólido (bactotriptona 1%, extrato de levedura
0,5%, NaCl 0,5%, bacto-ágar 1,5%) e as colônias isoladas foram repicadas
mensalmente. As cepas utilizadas como hospedeiras dos plasmídios foram:
E. coli DH5-α
Genótipo: F¯ φ80d/acZ∆M15 ∆(lacZYA¯ argF)U169 endA1 recA1 hdR17 (rk
¯mk+) deoR thi-1 supE44 λ¯ gyrA96 relA1 (Hanahan, 1985).
Condições de crescimento: uma colônia foi isolada da placa de Petri e
inoculada em 10 ml de meio LB líquido (bactotriptona 1%, extrato de levedura 0,5%,
NaCl 0,5%). A cultura foi incubada durante a noite a 37°C sob agitação.
Função: manutenção dos plasmídios recombinantes Topo 2.1 e pHIS.
E. coli BL21DE3
Genótipo: BF¯ dcm ompT hsdS(ra¯ ma¯) gal λ(DE3) (Stratagene)
Condições de crescimento: uma colônia foi isolada da placa de Petri e
inoculada em 10 ml de meio LB líquido. A cultura foi incubada durante a noite a 37°C
sob agitação.
Função: expressão da proteína recombinante.
25

Materiais e Métodos
5.2. Plasmídios
Topo 2.1 Genótipo: promotor tac, gene de resistência a ampicilina
Condições de crescimento: bactérias E. coli DH-α e BL21DE3 foram
transformadas com o plasmídio e plaqueadas em meio LB sólido contendo
ampicilina 100µg/ml e incubadas a 37°C duranta a noite.
Função: vetor de seqüenciamento.
pHIS paralelo 3 Genótipo: promotor T7, operador lac, 6xHis amino-terminal, sítio múltiplo de
clonagem (variação do pHIS paralelo 1).
Condições de crescimento: bactérias E. coli BL21DE3 foram transformadas
com o plasmídio e plaqueadas em meio LB sólido contendo ampicilina 100 µg/ml e
incubadas a 37°C durante a noite.
Função: vetor bacteriano de expressão induzida.
6. Clonagem do gene Lacys24 no vetor Topo 2.1 e subclonagem no vetor de expressão pHIS
O produto de PCR purificado e o plasmídio Topo 2.1 foram digeridos com as
enzimas de restrição EcoR1 e HindIII para a clonagem dirigida. A ligação foi
26

Materiais e Métodos
realizada em uma reação contendo inserto:vetor (3:1 em massa de DNA), 1 U de T4
ligase e tampão de ligação (Invitrogem) por 18 h a 22°C.
Para a subclonagem o inserto foi digerido do vetor Topo 2.1 com as mesmas
enzimas de restrição e ligado ao vetor pHIS sob as mesmas condições.
6.1. Preparação das bactérias competentes
As bactérias competentes (E. coli DH5-α e BL21D3) foram preparadas
inoculando-se uma colônia em 3 ml de meio LB durante a noite a 37°C sob agitação.
Após esse período os 3 ml da cultura foram adicionados a 247 ml de meio LB e uma
nova incubação foi realizada a 37°C até a cultura bacteriana atingir uma densidade
óptica de 0,35-06. Em seguida as células foram incubadas no gelo por 10-60
minutos e centrifugadas a 3.000 rpm por 15 min a 4°C, o meio da cultura foi
descartado e as bactérias ressuspensas em 1/3 do volume em FB (KCL 100 mM,
CaCl2 50 mM, glicerol 10% e acetato de potássio 10 mM). Após incubação no gelo
por 10-60 min as células foram centrifugadas e ressuspensas em 20 ml de FB.
6.2. Transformação das bactérias competentes com o DNA plasmidial
Para cada transformação 200 ng do DNA plasmidial e 100 µl da suspensão
das bactérias competentes foram misturados em um tubo de vidro e incubados por 1
hora no gelo. A seguir as células foram submetidas a um choque térmico de 42°C
por 3 minutos e reincubadas em gelo por 10 minutos. Em seguida adicionaram-se
400 µl de meio LB, incubando-se a 37°C por 1 hora sob agitação. Após esse período
o volume total da mistura foi semeado em placas de LB sólido contendo ampicilina
(100 µg/ml) e incubadas em estufa a 37°C durante a noite.
6.3. Purificação do DNA plasmidial
As colônias de bactérias contendo o DNA plasmidial foram isoladas da placa
de LB sólido e cultivadas em 3 ml de LB líquido contendo ampicilina 100 µg/ml por
18 h a 37°C sob agitação. Após o crescimento as células foram centrifugadas a
4.000 x g por 10 min e ressuspensas em 100 µl da solução A (Tris-HCI 25 mM,
27

Materiais e Métodos
EDTA 10 mM e glicose 50 mM) contendo 2 µg/ml de RNAse. Em seguida foram
adicionados 200 µl da solução B (NaOH 0,2 mM e SDS 1%) incubando-se por 5 min.
Após a incubação foram acrescentados 200 µl da solução C (KC2H3O2 3 M e ácido
acético glacial 6 M). As amostras foram mantidas no gelo por 25 min e centrifugadas
a 13°C a 12.000 x g por 8 min e ao sobrenadante foram acrescentados 400 µl de
isopropanol. Após outra centrifugação a 12.000 x g por 8 min, o precipitado foi
lavado com 500 µl de etanol 70% gelado e ressupenso em 25 µl de água Mili Q.
7. Seqüenciamento dos clones recombinantes Topo 2.1/Lacys24
Os plasmídios recombinantes Topo 2.1/Lacys24 foram submetidos à digestão
com as enzimas de restrição EcoRI e BamHI para verificar a presença do inserto. A
digestão foi realizada em uma reação contendo 200 ng de DNA e 10 U das
respectivas enzimas em tampão próprio a 37oC por 1 h. Após a análise das
amostras em gel de agarose 0,8% e confirmação da presença do inserto, cada clone
foi submetido a uma reação de PCR utilizando-se o "ABI Prism Big Dye Terminator
Cycle Sequencing Ready Reaction Kit". Para cada reação de seqüenciamento foram
utilizados 2 µL da solução “Big Dye terminator”, 1 µl (1,6 pmol) dos oligonucleotídeos
específicos T7: 5’ – TAA TAC GAC TCA CTA TAG GG – 3’ (senso) e U19: - GTT
TTC CCA GTC ACG AC-3’ (antisenso) e 400 ng do DNA a ser seqüenciado. As
condições de amplificação foram: 96°C por 5 min seguidos de 25 ciclos de 96°C por
20 seg, 50°C por 10 seg e 60°C por 4 min. Após esse período os tubos foram
mantidos a 4°C até o DNA ser precipitado acrescentando-se 100 µl de etanol 80%
gelado, seguindo-se uma centrifugação a 12.500 x g por 20 min a 4°C. Os
sobrenadantes foram novamente descartados e aos precipitados acrescentaram-se
200 µl de etanol 70% gelado, centrifugando-se novamente a 12.500 x g por 10 min a
4°C. Os precipitados foram secos a 37°C por 60 min. Acrescentaram-se em cada
tubo 2,5 µl de tampão de corrida (“blue dextran” 50 mg/ml/EDTA 25 mM, pH 8,0, e
formamida deioniozada na proporção de 1:5). Antes da aplicação no gel de
seqüenciamento as amostras foram fervidas por 2 min a 90°C e o seqüenciamento
foi realizado no sistema ABI prism 377 – Perkin-Elmer.
28

Materiais e Métodos
7.1. Preparação do gel para o sistema ABI/Prisma 377
Para o volume final de 25 ml de gel de poliacrilamida foram utilizados 9 g de
uréia, 2,5 ml de "Long Ranger gel solution" (FMC Bio Products), 2,5 ml de TBE 10x
e 13 ml de água Milli Q. A mistura foi mantida sob agitação por 20 min e em seguida
foram acrescentados 17,5 µl de TEMED e 125 µl de persulfato de amônio 10%. O
gel foi aplicado entre placas de vidro e a eletroforese realizada a 4.500 V por 7 h. As
seqüências foram obtidas por análise do "software" do sistema ABI/Prisma 377 e
comparadas às de genes codificadores de cisteína proteinases de outras espécies
de Leishmania utilizando o programa BLAST (http://www.ncbi.nlm.nih.gov/BLAST/).
8. Expressão e purificação da proteína recombinante
As colônias de bactérias contendo o plasmídio recombinante pHIS/Lacys24
foram isoladas da placa de LB sólido e cultivadas em 3 ml de LB líquido contendo
ampicilina 100 µg/ml por 18 h a 37ºC sob agitação. Após o crescimento, as bactérias
foram transferidas para 25 ml do mesmo meio e novamente crescidas durante a
noite a 37ºC. Para a expressão da proteína recombinante, a 500 ml de meio LBamp
foram acrescentados 25 ml da cultura de bactérias. A suspensão foi mantida sob
agitação a 37oC até a DO600nm= 0,6 e a expressão da proteína foi induzida com IPTG
0,2 mM durante 3 h nas mesmas condições. Após a incubação as bactérias foram
centrifugadas a 4.000 x g por 10 min e ressuspensas em 20 ml de tampão de
sonicação (NaH2PO4 0,1 M, Tris-HCl 10 mM, pH 8,0, PMSF 2 mM, iodoacetamida 2
mM) contendo 0,75 mg/ml de lisozima. A amostra foi mantida sob agitação por 30
min à temperatura ambiente, sonicada em 3 ciclos de 30 segundos e centrifugada a
12.000 x g por 30 min. O precipitado foi ressuspenso e lavado 3 vezes com 20 ml de
CHAPS 1% em Tris-HCl 10 mM, pH 8,0, e solubilizado com 20 ml de tampão de
solubilização (uréia 8 M em NaH2PO4 0,1 M, Tris-HCl 10 mM, pH 8,0).
Para a purificação da proteína recombinante, 5 ml de resina contendo níquel
(Ni-NTA Superflow agarose matrix - QIAGEN) foram acrescentados à suspensão de
bactérias solubilizada como descrito, mantendo-se sob agitação por 1 h. A mistura
foi transferida para coluna de vidro (Bio-Rad) e a resina lavada com 50 ml de tampão
de lavagem (NaH2PO4 0,1 M, Tris-HCl 10 mM, uréia 4 M, pH 6,3). A proteína
29

Materiais e Métodos
recombinante ligada à resina foi eluída com 20 ml de tampão de eluição (NaH2PO4
0,1 M, Tris-HCl 10 mM, uréia 4 M, imidazol 0,1 M, pH 6,3) e coletada em frações de
2 ml. As frações foram analisadas em gel de poliacrilamida a 12% e aquelas que
continham a proteína recombinante, visualizada no gel com massa molecular
aparente de 24 kDa, foram misturadas e dialisadas por 36 h a 4oC contra Tris-HCl 10
mM, pH 8,0, contendo uréia 0,5 M (Skeiky et al, 2002).
8.1. Análise de proteínas por eletroforese em gel de poliacrilamida com SDS (PAGE-SDS)
A análise eletroforética de proteínas foi feita em gel de poliacrilamida a 12%
utilizando-se o sistema Mini-Protean III (Bio-Rad). Os extratos dos amastigotas de L.
(L.) amazonensis e das bactérias recombinantes foram homogeneizados em tampão
de amostra e fervidos por 5 min. O equivalente a 25 µg de proteínas totais de cada
extrato foi adicionado por canal do gel e a corrida realizada a 200 V por 45 min. Após
a corrida os géis foram corados por 30 min com azul de Coomassie R-250 0,1% em
metanol:ácido acético:água (5:4:1) (v/v/v) e descorados com solução de
metanol:ácido acético:água (5:4:1) (v/v/v).
9. Reatividade da proteína recombinante com anticorpos anti-cisteína proteinase 9.1. Produção de anticorpos policlonais anti-rLacys24
Para a produção de anticorpos policlonais, camundongos BALB/c foram
imunizados com 25 µg da proteína recombinante rLacys24 emulsificada em
adjuvante completo de Freund (ACF) 1:1 por via intraperitonial. Após uma semana
os animais receberam duas doses de 25 µg do antígeno, com um intervalo de uma
semana entre as doses, com adjuvante incompleto de Freund por via intraperitoneal.
Uma semana após a terceira dose o sangue dos animais foi coletado e o título de
anticorpos do soro foi testado por ELISA. Os animais que apresentaram títulos altos
de anticorpos foram submetidos à punção cardíaca para a coleta do sangue e os
soros foram misturados, divididos em alíquotas e armazenados a –20ºC.
30

Materiais e Métodos
9.2. ELISA
Uma placa de 96 poços (high binding Costar) foi sensibilizada com 100
ng/poço da rLacys24 diluída em tampão Na2CO3/NaHCO3 0,05 M, pH 9,6. A placa foi
incubada durante a noite a 4°C, bloqueda com PBS Molico 5% por 1 h à temperatura
ambiente e os soros foram testados em diluições seriadas de 1:50 a 1:3.200 em PBS
Molico 5%. A placa foi incubada por 1h à temperatura ambiente, lavada 3 vezes com
PBS-Tween 20 0,05% e incubada novamente com anticorpo anti-IgG de
camundongo conjugado com peroxidase (Bio-Rad) 1:500 por 1 h à temperatura
ambiente. Após 3 lavagens a reação foi revelada adicionando-se 100 µl/poço de
tampão de revelação (Na2HPO4 0,05 M, ácido cítrico 0,05 M, pH 4,5) contendo
ortofenilenodiamina (OPD) 0,5 mg/ml e H2O2 0,03% por 15 min. A reação foi
interrompida adicionando-se 50 µl por poço de H2SO4 4 N e a absorbância medida a
492 nm em microleitor de ELISA Labsystem Multiskan MS.
9.3. “Western blotting’’
Os experimentos de “Western Blotting’’ foram feitos como descrito por Towbin
et al (1979). Extratos de amastigotas de L. (L.) amazonensis foram submetidos à
PAGE-SDS como descrito anteriormente. Após a separação eletroforética as
proteínas foram transferidas para a membrana de nitrocelulose (NC) (Hybond ECLTM
Pharmacia) por eletroforese a 250 mA por 2 h no sistema “Mini Trans-Blot” (Bio-Rad)
em tampão de transferência (Tris-HCl 2,5 mM, glicina 0,192 M, metanol 20%). A
visualização das proteínas e marcadores de massa molecular foi feita por coloração
da NC com Ponceau-S 0,1% em ácido acético 10% e descoloração em água. Em
seguida as membranas foram bloqueadas com PBS-Molico 5% por 1 h e incubadas
com o anticorpo monoclonal anti-cisteína proteinase de L. (L.) amazonensis (AcMo
2E5D3) (1:100) ou com o anticorpo policlonal anti-rLacys24 (1:100). Após 2 horas de
incubação com os anticorpos, as membranas foram lavadas com PBS-Tween 0,05%
e incubadas por 1 h com anticorpo anti-IgG de camundongo conjugado com
peroxidase (Bio-Rad) (1:500). Após a lavagem das membranas, ela foram reveladas
com diaminobenzidina (DAB) 0,15 mg/ml e H2O2 0,005% em PBS.
31

Materiais e Métodos
10. Avaliação da resposta imune estimulada pela rLacys24 10.1. Imunização dos animais com a rLacys24 Camundongos BALB/c, com 6-8 semanas de idade, foram imunizados com 25
µg da rLacys24. Os adjuvantes testados foram Propionibacterium acnes (suspensão
de bactérias autoclavadas correspondente a 150 µg de proteína por dose) ou
adjuvante completo de Freund (ACF) na proporção 1:1. Quando ACF foi utilizado a
via de imunização foi a subcutânea na pata dos animais. Com a P. acnes os animais
foram imunizados pela via intraperitoneal, recebendo 4 doses semanais da rLacys24
mais P. acnes.
10.2. Análise das populações de linfócitos estimuladas pela rLacys24
As populações celulares estimuladas após a imunização dos camundongos
BALB/c com a rLacys24 foram analisadas por citometria de fluxo. Foram utilizados
os anticorpos monoclonais anti-CD3 conjugado à aloficocianina (APC) (Pharmingen),
anti-CD4 conjugado à ficoeritrina (PE) (Pharmingen) e anti-CD-8 conjugado à
proteína clorofila peridinina (PerCP) (Pharmingen). Para as marcações foram
utilizados 1x106 linfócitos por tubo obtidos dos linfonodos inguinais e poplíteos dos
camundongos BALB/c imunizados com a rLacys24 mais ACF e 1x106 esplenócitos
dos camundongos imunizados com a rLacys24 mais P. acnes. Os linfócitos foram
fixados em 500 µl de paraformaldeído 1% em PBS por 30 minutos a 4ºC, lavados
duas vezes com 500 µl de PBS e ressuspensos em 100 µl de PBS. A cada tubo foi
acrescentado 1 µl do anticorpo de interesse, incubando-se as células por 1 hora a
4ºC, protegidas da luz. Em seguida as células foram lavadas duas vezes com 500 µl
de PBS e novamente fixadas com 500 µl de paraformaldeído 1% em PBS por 30
minutos a 4ºC. Após mais duas lavagens com 500 µl de PBS, as células foram
ressuspensas em 1 ml de PBS e a fluorescência foi analisada por citometria de fluxo
(FACSCAN – Cell Sorter Becton Dickinson).
32

Materiais e Métodos
10.3. Ensaio de citotoxicidade pelo método do cristal violeta
Nesses ensaios foram utilizadas como células-alvo macrófagos peritoneais de
camundongos BALB/c infectados com L. (L.) amazonensis. Para a cultura de
macrófagos as células foram isoladas de camundongos BALB/c após lavado do
peritônio com PBS. A amostra foi diluída em solução de cristal violeta 1 mg/ml em
ácido acético 30% para a contagem diferencial de células e os macrófagos foram
cultivados em placas de 96 orifícios à concentração de 1x105 por orifício, mantidas
por 30 min à temperatura ambiente, lavadas com PBS para a retirada das células
não aderentes e incubadas em meio RPMI (Gibco) com soro fetal bovino 10% em
estufa a 37oC com 5% de CO2 por 24 horas. A seguir os macrófagos foram
infectados na proporção de 2:1 (amastigota:macrófago) durante 72 horas em estufa
contendo 5% de CO2 a 37ºC. Os linfócitos isolados dos linfonodos inguinais e
poplíteos dos camundongos BALB/c previamente imunizados com a rLacys24 mais
ACF foram adicionados às placas contendo os macrófagos infectados com L. (L.)
amazonensis na proporção de 200 linfócitos:1 macrófago. As células foram
incubadas por 4 horas em estufa contendo 5% de CO2 a 37ºC. A seguir as células
foram lavadas 4 vezes com PBS, fixadas por 10 minutos com metanol 100%,
lavadas com PBS e coradas com uma solução de cristal violeta a 0,2% em etanol
2% por 30 minutos. Após 3 lavagens com PBS, procedeu-se à eluição adicionando-
se 150 µl de metanol 50% por orifício e após 10 minutos foi feita a leitura a 540 nm
em microleitor de ELISA Labsystem Multiskan MS. Os controles do ensaio foram:
lise total dos macrófagos infectados com água ou HCl 0,1 N; macrófagos infectados
sem incubação com os linfócitos (controle); macrófagos infectados incubados com
os linfócitos dos animais imunizados apenas com ACF; coloração dos orifícios
vazios, sem células para avaliar o possível ”background” da placa.
A porcentagem de lise foi determinada pela seguinte fórmula:
% de lise= (absorbância do controle – absorbância do grupo experimental)/absorbância do controle X 100
33

Materiais e Métodos
10.4. Ensaios por “Enzyme-linked immunospot” (ELISPOT)
A técnica é baseada no ensaio de ELISA de captura e detecta e enumera os
linfócitos secretores de uma determinada linfocina pela utilização dos anticorpos de
captura e detecção e revelação pela reação enzimática (Czerkinsky et al., 1983). Ela
foi utilizada para avaliar a secreção de IFN-γ, IL-4 e IL-10 pelos linfócitos dos
camundongos BALB/c imunizados com a rLacys24 ou rLdccys1 ou ambos os
antígenos mais P. acnes e desafiados com a L. (L.) amazonensis.
Foram utilizados os kits para ELISPOT da BD Biosciences. Placas de 96
poços próprias para ELISPOT estéreis foram incubadas com o anticorpo monoclonal
anti-IFN-γ ou anti-IL-4 ou anti-IL-10 à concentração de 5 µg/ml em PBS estéril e
incubadas durante a noite a 4ºC. A seguir as placas foram lavadas com 200 µl por
poço de RPMI + soro fetal bovino 10% (R10) estéril e mantidas nesse meio por 2
horas à temperatura ambiente. A seguir o meio foi descartado e adicionaram-se os
linfócitos isolados dos linfonodos inguinais e poplíteos dos camundongos BALB/c
imunizados e desafiados. Foram utilizados 5x105 linfócitos por poço em meio
completo (RPMI 1649 contendo NaHCO3 20 mM, HEPES 10 mM, penicilina 100
U/ml, estreptomicina 100 µg/ml, L-glutamina 2 mM, β-mercaptoetanol 50 µM,
piruvato de sódio 5 mM, mistura de aminoácidos não essenciais (Sigma) 100 µM e
soro humano normal 2% previamente inativado por 1 h a 56oC) na presença de cada
antígeno recombinante ou de ambos à concentração de 2,5 µg/ml ou do extrato total
de amastigotas de L. (L.) amazonensis (AM) (correspondente a 1x107
amastigotas/poço), incubando-se as placas em estufa a 37ºC contendo 5% de CO2
durante 3 dias. A seguir as células foram aspiradas, os poços foram lavados 2 vezes
com água destilada e 3 vezes com PBS contendo Tween 20 0,05% (tampão de
lavagem), adicionando-se os anticorpos de detecção anti-IFN-γ ou anti-IL-4 ou anti-
IL-10 biotinilado à concentração de 2 µg/ml. Após 2 horas à temperatura ambiente,
os anticorpos foram descartados e os poços lavados 3 vezes com tampão de
lavagem, adicionou-se o conjugado enzimático (estreptoavidina-peroxidase) 2,5
µg/ml em PBS + 10% de soro fetal bovino, incubando-se por 1 hora à temperatura
ambiente. A seguir os poços foram lavados com PBS e adicionou-se a solução do
substrato (3-amino-9-etilcarbazol 0,33 mg/ml + dimetilformamida 3% + H2O2 0,015%
em tampão acetato de sódio 0,1 M, pH 5,0), incubando-se por 5 a 30 minutos à
34

Materiais e Métodos
temperatura ambiente. Em seguida interrompia-se a reação adicionando-se água
destilada aos poços. Após a secagem, os “spots” foram enumerados por contagem
em microscópio estereoscópico. Os resultados são expressos como o número de
“spots”/106 linfócitos. 11. Imunização ativa
11.1. Imunização de camundongos BALB/c com a rLacys24 e desafio dos animais com a L. (L.) amazonensis
Os animais foram imunizados com 25 µg da rLacys24 e P. acnes ou ACF
como adjuvantes, conforme descrito no item 10.1. Duas semanas após a imunização
os animais foram desafiados pela inoculação nas patas de 2,5x105 amastigotas de L.
(L.) amazonensis. A infecção foi avaliada pela medida semanal das patas com
paquímetro até 3 meses após o desafio.
11.2. Imunização de camundongos BALB/c com a rLacys24, a rLdccys1 ou os dois antígenos recombinantes
Nesses experimentos os camundongos BALB/c foram imunizados com 25 µg
da rLacys24, 25 µg da rLdccys1 ou 25 µg da rLacys24 + 25 µg da rLdccys1 e P.
acnes como adjuvante (correspondente a 150 µg de proteína por dose) por via
subcutânea na base da cauda. Os animais receberam 4 doses, com intervalo de
uma semana entre cada uma, e 14 dias após a última dose foram desafiados com
2,5x105 amastigotas de L. (L.) amazonensis na pata traseira direita. O grupo controle
foi imunizado apenas com 4 doses da P. acnes. Após 3 meses do desafio, foram
avaliadas as populações de linfócitos dos animais por citometria de fluxo e a
produção de IFN-γ, IL-4 e IL-10 por ELISPOT, como descrito nos itens 10.2 e 10.4,
respectivamente. A infecção dos animais foi avaliada pela medida semanal das
patas com paquímetro até três meses após o desafio. Após os três meses de
infecção e o sacrifício dos animais, os amastigotas das lesões foram isolados e
cultivados a 23 ºC durante 7 dias em meio 199 (Sigma) contendo 10% de soro fetal
bovino previamente inativado por 1 hora a 56 ºC. A contagem dos promastigotas
35

Materiais e Métodos
36
resultantes da diferenciação dos amastigotas nesse meio foi o outro parâmetro
utilizado para a avaliação da carga parasitária dos animais.
12. Análise estatística
A significância dos resultados foi avaliada pela análise estatística de todos os
experimentos pelos testes ANOVA e Turkey HSD disponíveis na URL
(http://faculty.vassar.Edu/lowry/VassarStats.html).

Resultados
37

Resultados
1. Seqüenciamento do clone Lacys24 no vetor Topo 2.1
Visando à clonagem de um fragmento de 500 pb de amastigotas de L. (L.)
amazonensis, Lacys24, que foi previamente demonstrado apresentar alta identidade
com a de genes codificadores de cisteína proteinases de Leishmania descritos, o clone
Lacys24 foi amplificado por PCR utilizando-se DNA genômico de amastigotas de L. (L.)
amazonensis e “primers” contendo seqüências para sítios de restrição HindIII e EcoRI
construídos a partir da seqüência desse clone obtida em estudos anteriores em nosso
laboratório. Dessa amplificação foram obtidos vários fragmentos de 500 pb (Figura 2).
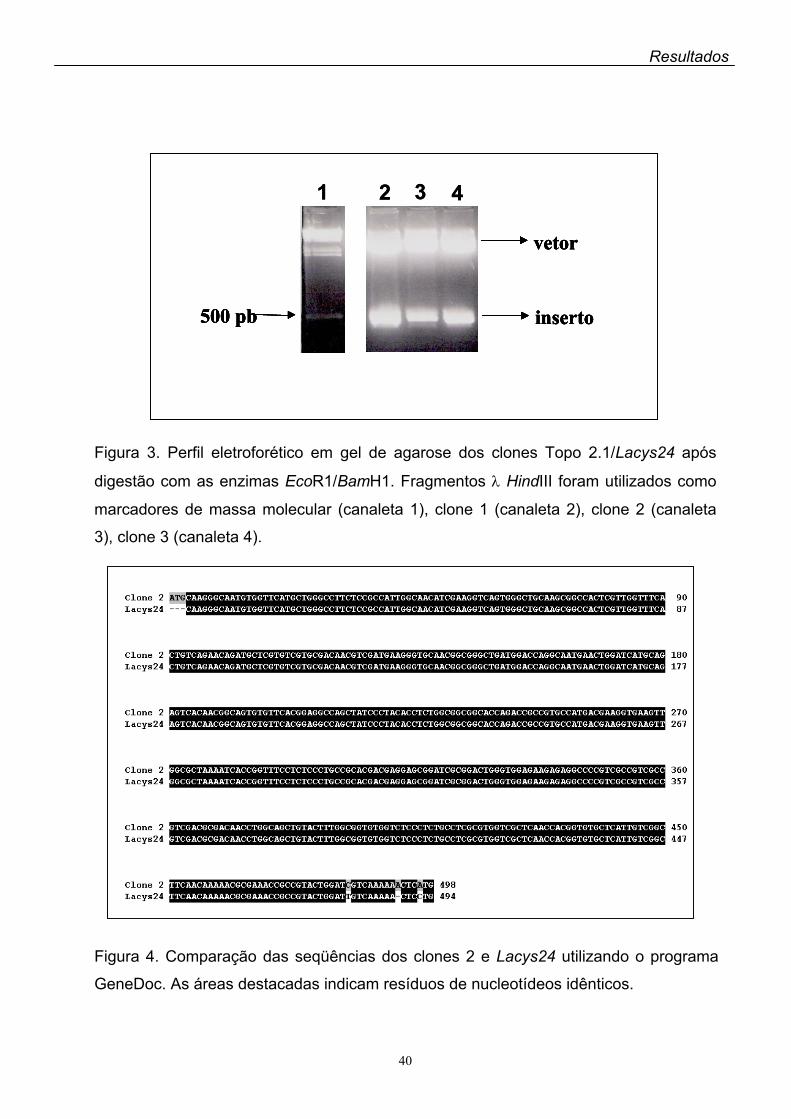
Esse fragmento de 500 pb foi clonado em vetor Topo 2.1 e três dos clones obtidos
foram submetidos à digestão enzimática por HindIII e EcoRI e separação eletroforética
(Figura 3). Os três clones que liberaram o inserto de 500 pb foram amplificados por
PCR e seqüenciados. A Figura 4 mostra a análise comparativa da seqüência de
nucleotídeos de um desses clones (clone 2) com a do clone Lacys24 que foi
anteriormente clonado em pMOS em nosso laboratório e que apresenta alto grau de
identidade com as seqüências de genes codificadores de cisteína proteinases
previamente descritas no gênero Leishmania (Oliveira, 2001). A comparação da
seqüência do clone 2 com a do Lacys24 mostrou um alto grau de similaridade entre
essas seqüências, havendo diferenças apenas em 3 bases no final do inserto (Figura
4).
Na Figura 5 são mostrados os possíveis epítopos de linfócitos T na seqüência
de aminoácidos do clone Lacys24 (Oliveira, 2001). Essa análise foi feita utilizando o
banco de dados para motivos de ligação ao MHC obtido via Internet. Foram analisados
os motivos de ligação específica ao MHC de classe I para o haplótipo H-2d expresso
nos camundongos BALB/c empregados em nossos estudos. Como mostra a Figura 5,
um provável motivo de ligação ao MHC classe I está presente no clone Lacys24. Os
motivos de ligação ao MHC de classe II desse clone foram analisados pelo programa
“Dnastar-Protean” em conjunto com os métodos AMPHI (Margalit et al., 1987) e
Motivos de MHC (Sette et al., 1989). Os epítopos determinados pelo programa AMPHI
apresentam-se como motivos de ligação ao MHC de classe II para os haplótipos IAd e
IEd. Foram localizadas três regiões que representam prováveis epítopos de linfócitos T
CD4+.
38

Resultados
500 pb
1 2 3 4 5 6
500 pb
1 2 3 4 5 6
Figura 2. Perfil eletroforético em gel de agarose dos clones de 500 pb amplificados por
PCR (3, 4 e 5). Fragmentos γDNA/Hind III e φX174 RF DNA/Hae III foram utilizados
como marcadores de massa molecular e são mostrados nas canaletas 1 e 2. Controle
da reação de PCR (em ausência do DNA genômico) (6).
39
Resultados
vetor
inserto500 pb
vetor
inserto500 pb
1 2 3 4
vetor
inserto500 pb
vetor
inserto500 pb
1 2 3 4
Figura 3. Perfil eletroforético em gel de agarose dos clones Topo 2.1/Lacys24 após
digestão com as enzimas EcoR1/BamH1. Fragmentos λ HindIII foram utilizados como
marcadores de massa molecular (canaleta 1), clone 1 (canaleta 2), clone 2 (canaleta
3), clone 3 (canaleta 4).
Figura 4. Comparação das seqüências dos clones 2 e Lacys24 utilizando o programa
GeneDoc. As áreas destacadas indicam resíduos de nucleotídeos idênticos.
40

Resultados
Figura 5. Identificação dos prováveis epítopos de células T e motivos de ligação ao
MHC do clone pMOS/Lacys24. As regiões destacadas representam os epítopos para
célula T identificados pelo padrão de anfipaticidade da molécula. Os motivos de ligação
ao MHC de classe I e II estão destacados em vermelho e azul, respectivamente
(adaptado de Oliveira, 2001).
41

Resultados
2. Subclonagem do clone Lacys24 de L. (L.) amazonensis em vetor de expressão e análise da proteína recombinante
Para a obtenção da proteína recombinante o fragmento de 500 pb do clone
Lacys24 foi digerido do plasmídio Topo 2.1 nos sítios de restrição EcoRI/HindIII e
subclonado no vetor de expressão pHIS paralelo 3 em fase com 6 resíduos de histidina
na porção N-terminal da proteína. A expressão do clone Lacys24 em bactérias
competentes BL21DE3 transformadas com o plasmídio Lacys24/pHIS resultou em uma
proteína recombinante de aproximadamente 24 kDa que foi denominada rLacys24. A
sonicação das bactérias seguida de centrifugação mostrou que a rLacys24 estava
contida em corpúsculos de inclusão insolúveis. Assim, a proteína recombinante foi
solubilizada em uréia 8 M e purificada por cromatografia de afinidade em coluna de
níquel. A expressão e purificação da rLacys24 foram avaliadas por PAGE-SDS (Figura
6).
42
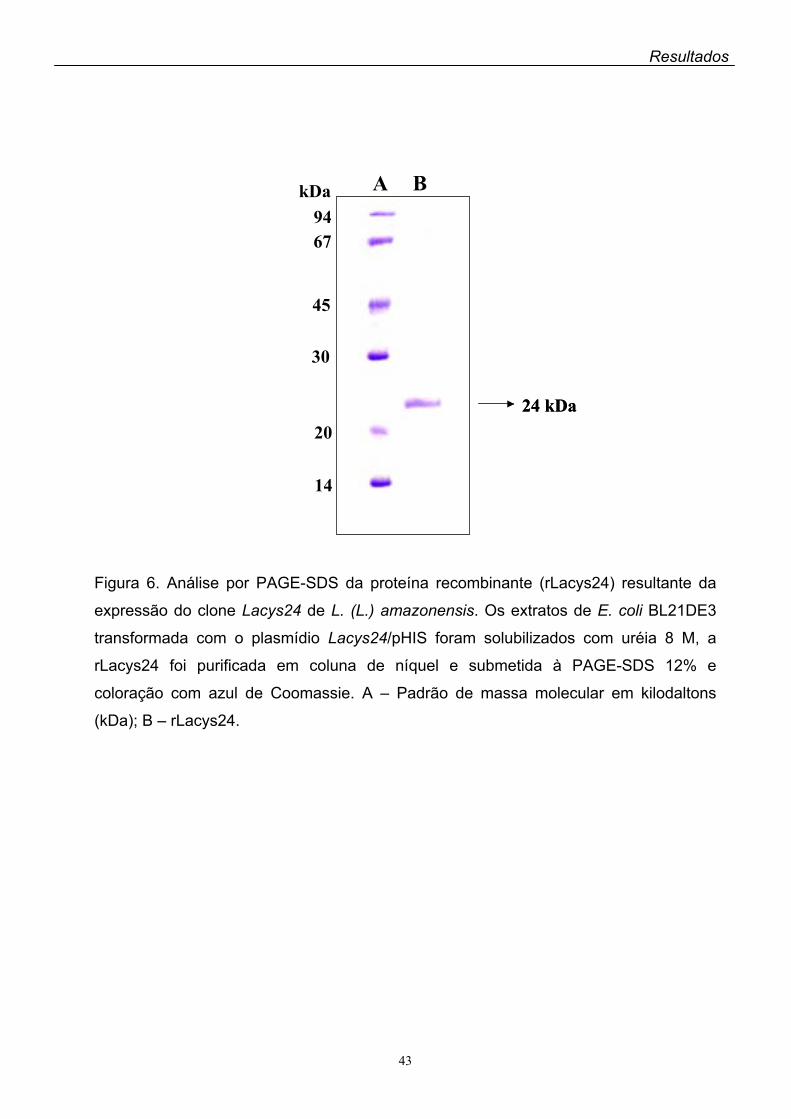
Resultados
24 kDa
9467
45
30
20
14
kDa A B
24 kDa
9467
45
30
20
14
kDa A B
Figura 6. Análise por PAGE-SDS da proteína recombinante (rLacys24) resultante da
expressão do clone Lacys24 de L. (L.) amazonensis. Os extratos de E. coli BL21DE3
transformada com o plasmídio Lacys24/pHIS foram solubilizados com uréia 8 M, a
rLacys24 foi purificada em coluna de níquel e submetida à PAGE-SDS 12% e
coloração com azul de Coomassie. A – Padrão de massa molecular em kilodaltons
(kDa); B – rLacys24.
43

Resultados
Para verificar se a proteína rLacys24 corresponde a uma das isoformas de
cisteína proteinase de 30 kDa abundantemente expressas em amastigotas de L. (L.)
amazonensis, a rLacys24 foi utilizada para a imunização de camundongos BALB/c e
obtenção de soro policlonal monoespecífico contra a proteína recombinante (anti-
rLacys24). O extrato de amastigotas de L. (L.) amazonensis foi submetido à PAGE-
SDS e ”Western blotting”, utilizando-se o anticorpo monoclonal 2E5D3 produzido contra
uma das isoformas de 30 kDa de L. (L.) amazonensis e o soro policlonal anti-rLacys24.
A Figura 7 mostra que, semelhante à reatividade observada com o anticorpo
monoclonal, o soro anti-rLacys24 reagiu com uma proteína de 30 kDa do extrato do
parasita.
A B C DA B C D
Figura 7: Análise por “Western blotting” da rLacys24. Os extratos de amastigotas de L.
(L.) amazonensis foram submetidos à PAGE-SDS, transferidos para membrana de
nitrocelulose e incubados com soro de camundongo normal (B), o AcMo 2E5D3 (C) e
soro de camundongo anti-rLacys24 (D). O padrão de massa molecular aparente está
representado em kilodaltons (kDa) em A.
44

Resultados
3. Respostas imunes celulares induzidas pela rLacys24
Após a obtenção e purificação da proteína recombinante, foi determinada a
capacidade de a rLacys24 induzir respostas imunes celulares em camundongos
BALB/c. Assim, os animais foram primeiramente imunizados com a rLacys24 e ACF.
Quinze dias após a imunização subcutânea com 25 µg da rLacys24 mais ACF, os
linfonodos poplíteos e inguinais desses animais foram isolados para a análise das
populações de linfócitos T por citometria de fluxo. Os resultados dessa análise são
mostrados na Figura 8. Pode-se observar que nos animais controle imunizados apenas
com ACF a população dos linfócitos T CD4+ e T CD8+ foi de 25% e 11%,
respectivamente, enquanto que os camundongos imunizados com a rLacys24
apresentaram 23% de linfócitos T CD4+ e aumento significante da expressão de T
CD8+ (19%).
A resposta imune desencadeada pela rLacys24 foi também testada utilizando-se
outro adjuvante, P. acnes, para a imunização dos animais. Nesses experimentos os
camundongos receberam 4 doses da P. acnes mais rLacys24 por via intraperitoneal,
com intervalo de 1 semana entre as doses, e 14 dias após a última imunização foi
avaliado o perfil da resposta imune celular por citometria de fluxo. Os resultados
desses experimentos são mostrados na Figura 9. Como pode ser observado, os
animais do grupo controle imunizados apenas com a P. acnes apresentaram 14% e
5,8% de linfócitos CD4+ e CD8+, respectivamente, e nos imunizados com a rLacys24
essas porcentagens foram 15% de CD4+ e 5,5% de CD8+.
45
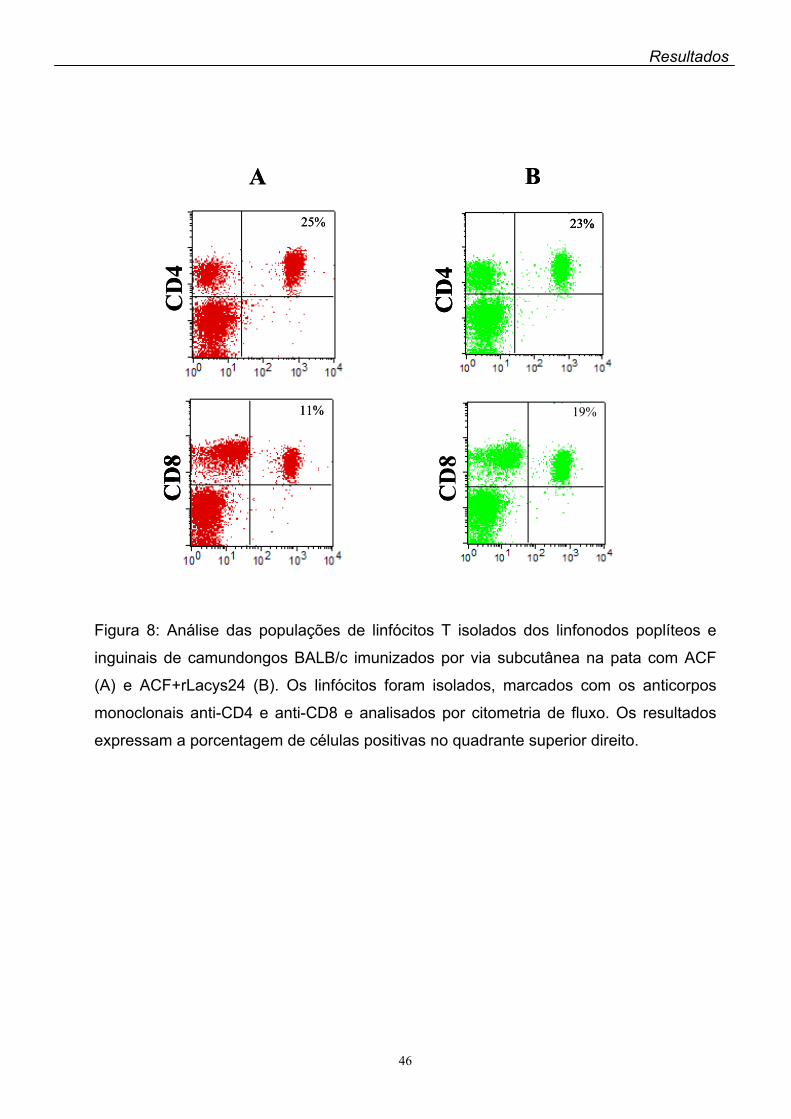
Resultados
A BC
D4
25%
CD
8
11%
CD
4
23%
CD
819%
A BC
D4
25%
CD
4
25%
CD
4
25%
CD
8
11%
CD
8
11%
CD
8
11%
CD
4
23%
CD
4
23%
CD
4
23%
CD
4
23%
CD
819%
CD
819%
Figura 8: Análise das populações de linfócitos T isolados dos linfonodos poplíteos e
inguinais de camundongos BALB/c imunizados por via subcutânea na pata com ACF
(A) e ACF+rLacys24 (B). Os linfócitos foram isolados, marcados com os anticorpos
monoclonais anti-CD4 e anti-CD8 e analisados por citometria de fluxo. Os resultados
expressam a porcentagem de células positivas no quadrante superior direito.
46
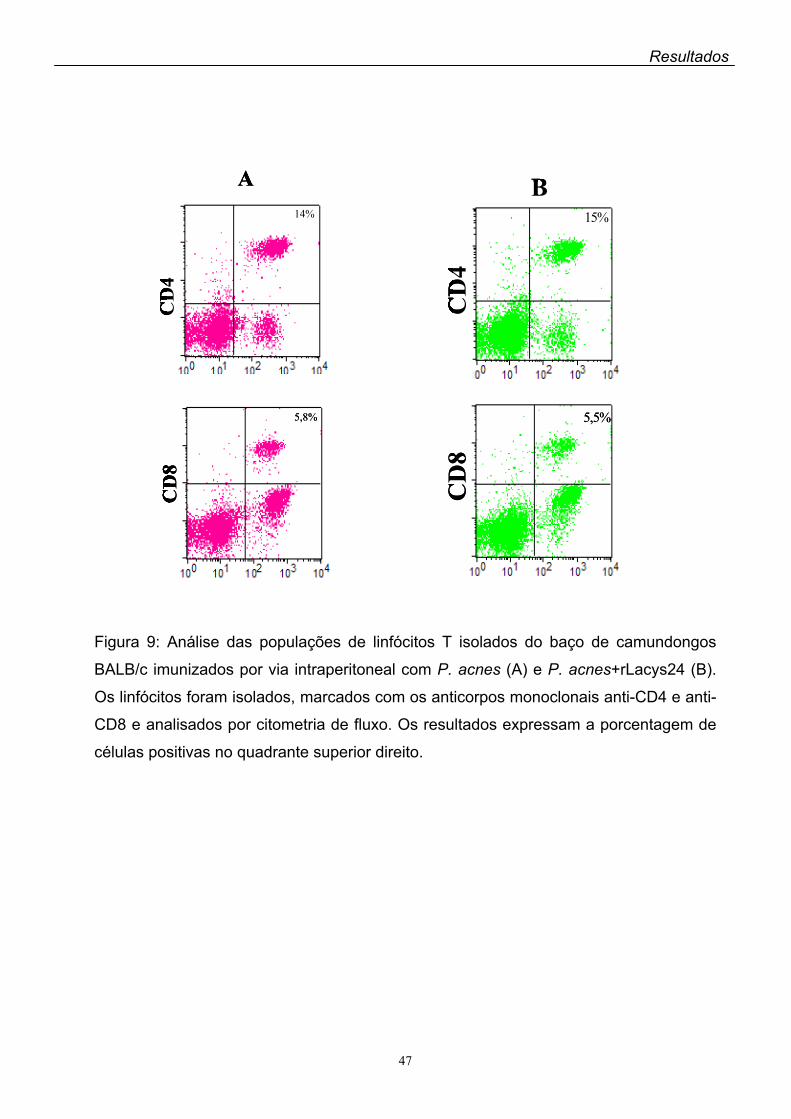
Resultados
CD
8
5,8%
14%
CD
4C
D8
5,8%
CD
8
5,8%
14%
CD
4A
CD
4
15%
CD
8
5,5%
CD
4
15%
CD
8
5,5%
CD
8
5,5%
B
CD
4
15%
CD
8
5,5%
CD
8
5,5%
CD
4
15%
CD
8
5,5%
CD
8
5,5%
BC
D8
5,8%
14%
CD
4C
D8
5,8%
CD
8
5,8%
14%
CD
4A
CD
8
5,8%
14%
CD
4C
D8
5,8%
CD
8
5,8%
14%
CD
4A
CD
4
15%
CD
8
5,5%
CD
4
15%
CD
8
5,5%
CD
8
5,5%
B
CD
4
15%
CD
8
5,5%
CD
8
5,5%
CD
4
15%
CD
8
5,5%
CD
8
5,5%
B
Figura 9: Análise das populações de linfócitos T isolados do baço de camundongos
BALB/c imunizados por via intraperitoneal com P. acnes (A) e P. acnes+rLacys24 (B).
Os linfócitos foram isolados, marcados com os anticorpos monoclonais anti-CD4 e anti-
CD8 e analisados por citometria de fluxo. Os resultados expressam a porcentagem de
células positivas no quadrante superior direito.
47
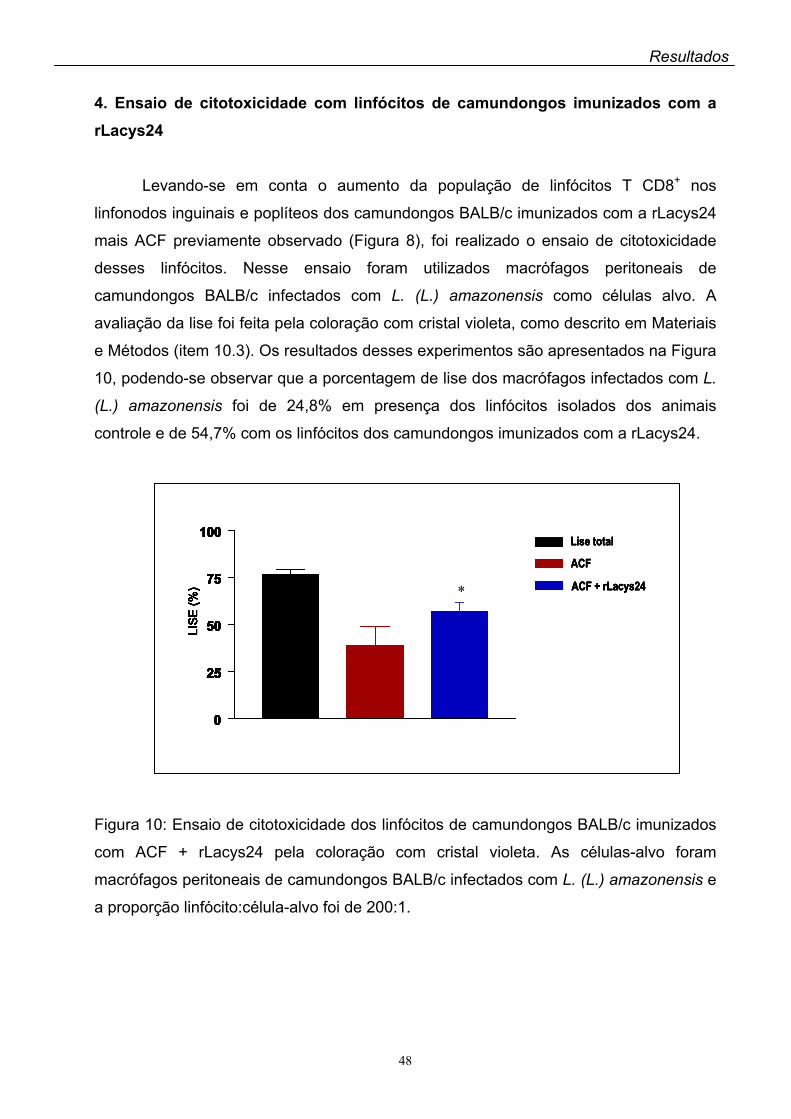
Resultados
4. Ensaio de citotoxicidade com linfócitos de camundongos imunizados com a rLacys24
Levando-se em conta o aumento da população de linfócitos T CD8+ nos
linfonodos inguinais e poplíteos dos camundongos BALB/c imunizados com a rLacys24
mais ACF previamente observado (Figura 8), foi realizado o ensaio de citotoxicidade
desses linfócitos. Nesse ensaio foram utilizados macrófagos peritoneais de
camundongos BALB/c infectados com L. (L.) amazonensis como células alvo. A
avaliação da lise foi feita pela coloração com cristal violeta, como descrito em Materiais
e Métodos (item 10.3). Os resultados desses experimentos são apresentados na Figura
10, podendo-se observar que a porcentagem de lise dos macrófagos infectados com L.
(L.) amazonensis foi de 24,8% em presença dos linfócitos isolados dos animais
controle e de 54,7% com os linfócitos dos camundongos imunizados com a rLacys24.
0
25
50
75
100
LIS
E (%
) *
0
25
50
75
100
LIS
E (%
)
0
25
50
75
100
LIS
E (%
) *
ACF
ACF + rLacys24
Lise total
0
25
50
75
100
LIS
E (%
) *
0
25
50
75
100
LIS
E (%
)
0
25
50
75
100
LIS
E (%
) *
ACF
ACF + rLacys24
Lise total
0
25
50
75
100
LIS
E (%
) *
0
25
50
75
100
LIS
E (%
)
0
25
50
75
100
LIS
E (%
) *
ACF
ACF + rLacys24
Lise total
0
25
50
75
100
LIS
E (%
) *
0
25
50
75
100
LIS
E (%
)
0
25
50
75
100
LIS
E (%
) *
ACF
ACF + rLacys24
Lise total
Figura 10: Ensaio de citotoxicidade dos linfócitos de camundongos BALB/c imunizados
com ACF + rLacys24 pela coloração com cristal violeta. As células-alvo foram
macrófagos peritoneais de camundongos BALB/c infectados com L. (L.) amazonensis e
a proporção linfócito:célula-alvo foi de 200:1.
48
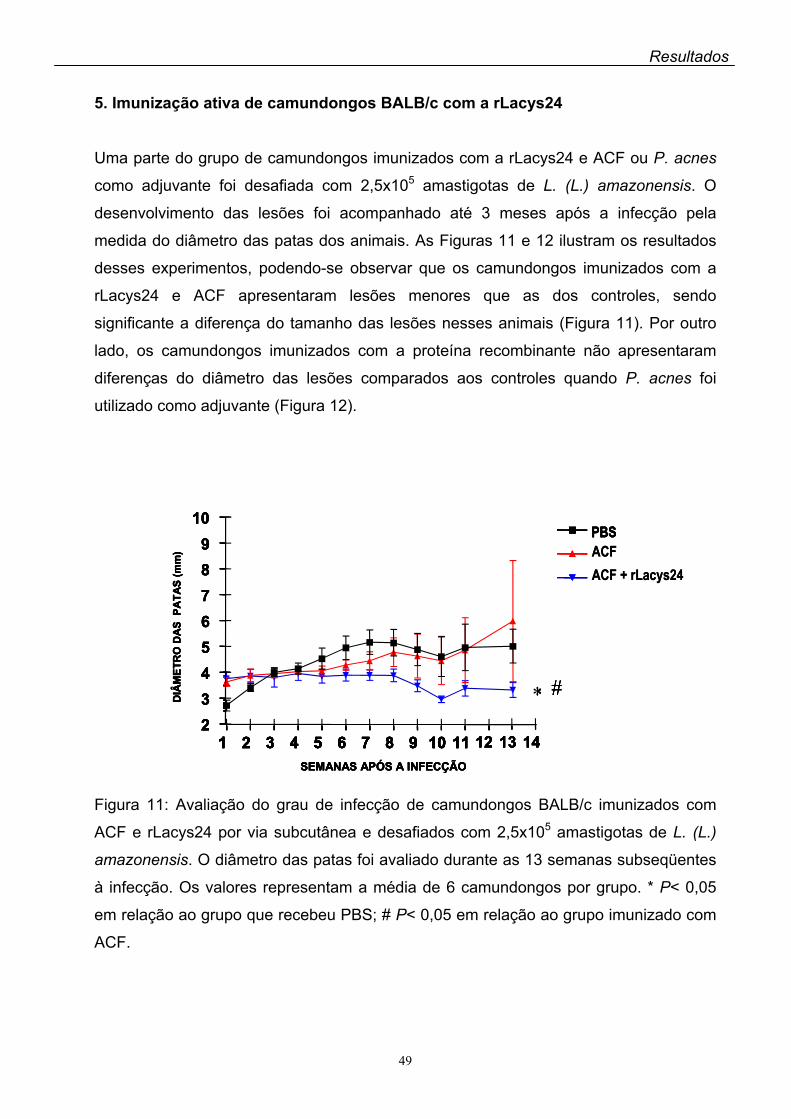
Resultados
5. Imunização ativa de camundongos BALB/c com a rLacys24
Uma parte do grupo de camundongos imunizados com a rLacys24 e ACF ou P. acnes
como adjuvante foi desafiada com 2,5x105 amastigotas de L. (L.) amazonensis. O
desenvolvimento das lesões foi acompanhado até 3 meses após a infecção pela
medida do diâmetro das patas dos animais. As Figuras 11 e 12 ilustram os resultados
desses experimentos, podendo-se observar que os camundongos imunizados com a
rLacys24 e ACF apresentaram lesões menores que as dos controles, sendo
significante a diferença do tamanho das lesões nesses animais (Figura 11). Por outro
lado, os camundongos imunizados com a proteína recombinante não apresentaram
diferenças do diâmetro das lesões comparados aos controles quando P. acnes foi
utilizado como adjuvante (Figura 12).
1 2 3 4 5 6 7 8 9 10 11 12 13 1423456789
10PBS
SEMANAS APÓS A INFECÇÃO
DIÂ
MET
RO
DA
S P
ATA
S (m
m)
* #
1 2 3 4 5 6 7 8 9 10 111 2 3 4 5 6 7 8 9 10 11 12 13 1423456789
10PBS
SEMANAS APÓS A INFECÇÃO
DIÂ
MET
RO
DA
S P
ATA
S (m
m)
* #
ACF + rLacys24ACF
1 2 3 4 5 6 7 8 9 10 111 2 3 4 5 6 7 8 9 10 11 12 13 1423456789
10PBS
SEMANAS APÓS A INFECÇÃO
DIÂ
MET
RO
DA
S P
ATA
S (m
m)
* #
1 2 3 4 5 6 7 8 9 10 111 2 3 4 5 6 7 8 9 10 11 12 13 1423456789
10PBS
SEMANAS APÓS A INFECÇÃO
DIÂ
MET
RO
DA
S P
ATA
S (m
m)
* #
ACF + rLacys24ACF
Figura 11: Avaliação do grau de infecção de camundongos BALB/c imunizados com
ACF e rLacys24 por via subcutânea e desafiados com 2,5x105 amastigotas de L. (L.)
amazonensis. O diâmetro das patas foi avaliado durante as 13 semanas subseqüentes
à infecção. Os valores representam a média de 6 camundongos por grupo. * P< 0,05
em relação ao grupo que recebeu PBS; # P< 0,05 em relação ao grupo imunizado com
ACF.
49
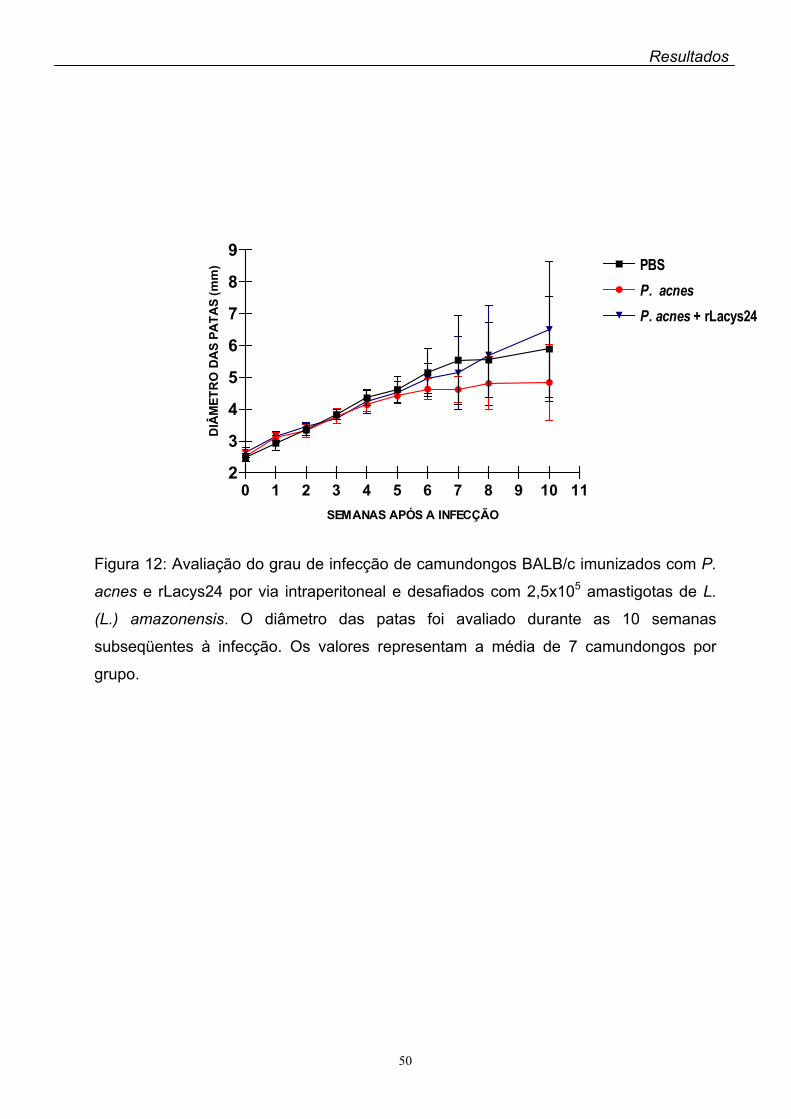
Resultados
0 1 2 3 4 5 6 7 8 9 10 112
3
4
5
6
7
8
9PBSP. acnesP. acnes + rLacys24
SEMANAS APÓS A INFECÇÃO
DIÂ
MET
RO
DA
S PA
TAS
(mm
)
Figura 12: Avaliação do grau de infecção de camundongos BALB/c imunizados com P.
acnes e rLacys24 por via intraperitoneal e desafiados com 2,5x105 amastigotas de L.
(L.) amazonensis. O diâmetro das patas foi avaliado durante as 10 semanas
subseqüentes à infecção. Os valores representam a média de 7 camundongos por
grupo.
50

Resultados
6. Imunização ativa com a rLacys24 e a rLdccys1 de L. (L.) chagasi
Os experimentos de imunização com a rLacys24 que resultaram em decréscimo
da infecção homóloga quando se utilizou ACF como adjuvante (Fig. 11) levaram-nos a
testar outro antígeno nos experimentos de imunização ativa, visando a um maior grau
de proteção. Assim, a rLdccys1, uma cisteína proteinase recombinante de L. (L.)
chagasi, foi também utilizada para a imunização dos animais. Essa alternativa baseou-
se nos dados anteriores de nosso laboratório que mostraram que a rLdccys1 é capaz
de induzir respostas medidas por linfócitos CD4+ Th1 e alto grau de proteção de
camundongos BALB/c e hamsters contra a infecção por L. (L.) chagasi (Ferreira, 2006).
Nesses experimentos camundongos BALB/c foram imunizados com 4 doses de
rLdccys1 e P. acnes como adjuvante por via subcutânea e 14 dias após a última dose
foram desafiados com 2,5x105 amastigotas de L. (L.) amazonensis. Um grupo de
animais foi imunizado com a rLacys24, outro com a rLdccys1 e outro com a rLdccys1
mais a rLacys24, além do grupo controle que recebeu apenas o adjuvante. A carga
parasitária desses animais foi acompanhada pela medida das lesões das patas durante
3 meses após o desafio com L. (L.) amazonensis e por cultivo dos parasitas isolados
dos linfonodos inguinais e poplíteos em meio axênico após o sacrifício dos animais. A
resposta imune desses animais foi avaliada pela análise das populações de linfócitos
inguinais e poplíteos por citometria de fluxo e pela secreção de linfocinas por ELISPOT
nas culturas desses linfócitos reestimulados in vitro com os antígenos utilizados para a
imunização. As Figuras 13 e 14 representam os resultados desses experimentos.
Pode-se observar que as respostas imunes dos animais foi predominantemente CD4+ e
não houve diferença significante da porcentagem dessas células entre os grupos
estudados. A expressão de linfócitos CD8+ foi baixa em todos os grupos (em torno de
8-10%), porém houve um aumento significante da porcentagem dessas células nos
animais imunizados com os antígenos comparada à apresentada pelo grupo controle
(Figura 13). Na Figura 14 são mostrados os resultados de análise das linfocinas pelos
ensaios de ELISPOT. Em todos os grupos estudados houve produção de IFN-γ, não
tendo havido diferenças significantes de secreção dessa linfocina pelos linfócitos dos
animais controle, imunizados apenas com P. acnes, ou mesmo pelos linfócitos
reestimulados in vitro pelos antígenos recombinantes utilizados (Figura 14A). Em
relação à IL-4, pôde-se observar a produção significante dessa linfocina pelos linfócitos
de todos os animais. Entretanto, o nível secretado pelos linfócitos dos animais
51

Resultados
imunizados com os antígenos recombinantes foi significantemente mais baixo que o
dos animais controle, sendo esse decréscimo mais acentuado nos camundongos
imunizados com a rLdccys1 (Figura 14B). A IL-10 também foi analisada, porém não foi
detectada nas culturas de linfócitos de nenhum dos grupos de animais estudados.
A análise da infecção dos animais durante 3 meses após o desafio com a L. (L.)
amazonensis mostrou que não houve diferença significante do tamanho das lesões das
patas dos animais imunizados com os antígenos recombinantes em relação aos
controles (Figura 15 A). Também não foram observadas diferenças em nenhum dos
grupos estudados quando se avaliou a infecção dos animais pelo número de parasitas
presentes nos linfonodos inguinais e poplíteos (Figura 15B).
52
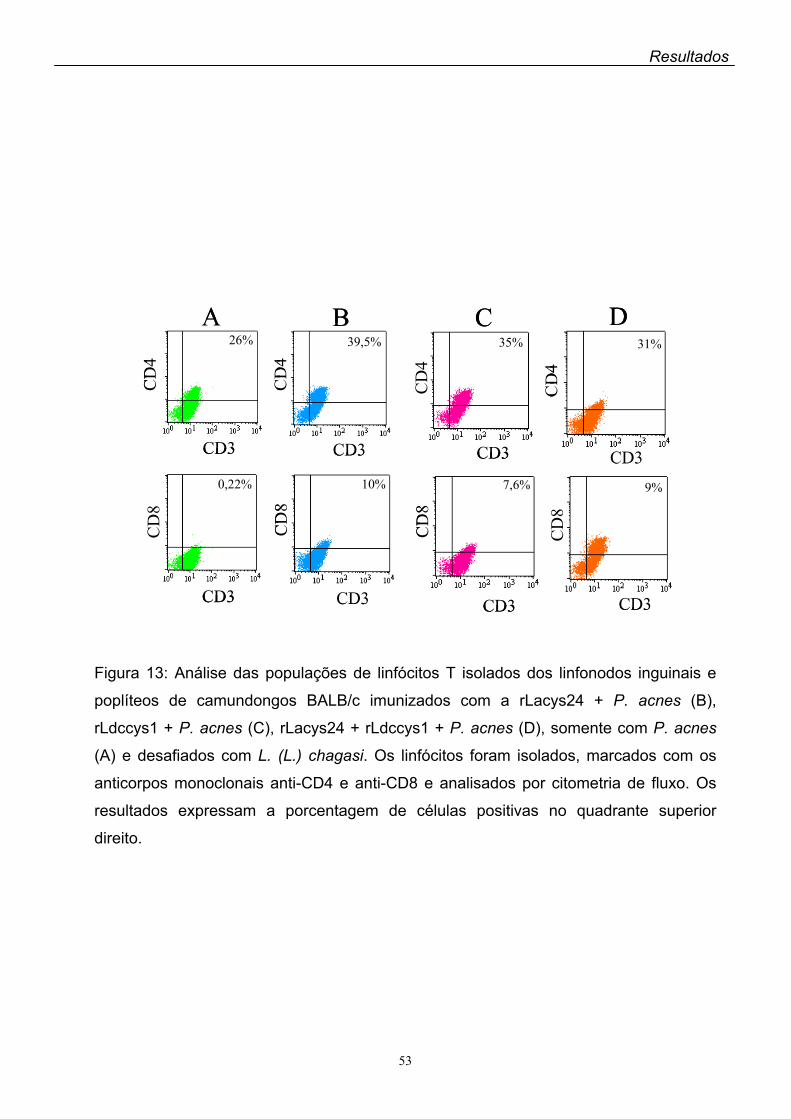
Resultados
CD3
CD
4
CD3
CD
8
26%
0,22%
CD
4
CD3
CD
8
CD3
39,5%
10%
CD
4
CD3
CD
8
CD3
35%
7,6%
A B C
CD3
CD
8
CD3
CD
4
9%
31%
D
CD3
CD
4
CD3
CD
8
26%
0,22%
CD
4
CD3
CD
8
CD3
39,5%
10%
CD
4
CD3
CD
8
CD3
35%
7,6%
A B C
CD3
CD
8
CD3
CD
4
9%
31%
D
Figura 13: Análise das populações de linfócitos T isolados dos linfonodos inguinais e
poplíteos de camundongos BALB/c imunizados com a rLacys24 + P. acnes (B),
rLdccys1 + P. acnes (C), rLacys24 + rLdccys1 + P. acnes (D), somente com P. acnes
(A) e desafiados com L. (L.) chagasi. Os linfócitos foram isolados, marcados com os
anticorpos monoclonais anti-CD4 e anti-CD8 e analisados por citometria de fluxo. Os
resultados expressam a porcentagem de células positivas no quadrante superior
direito.
53
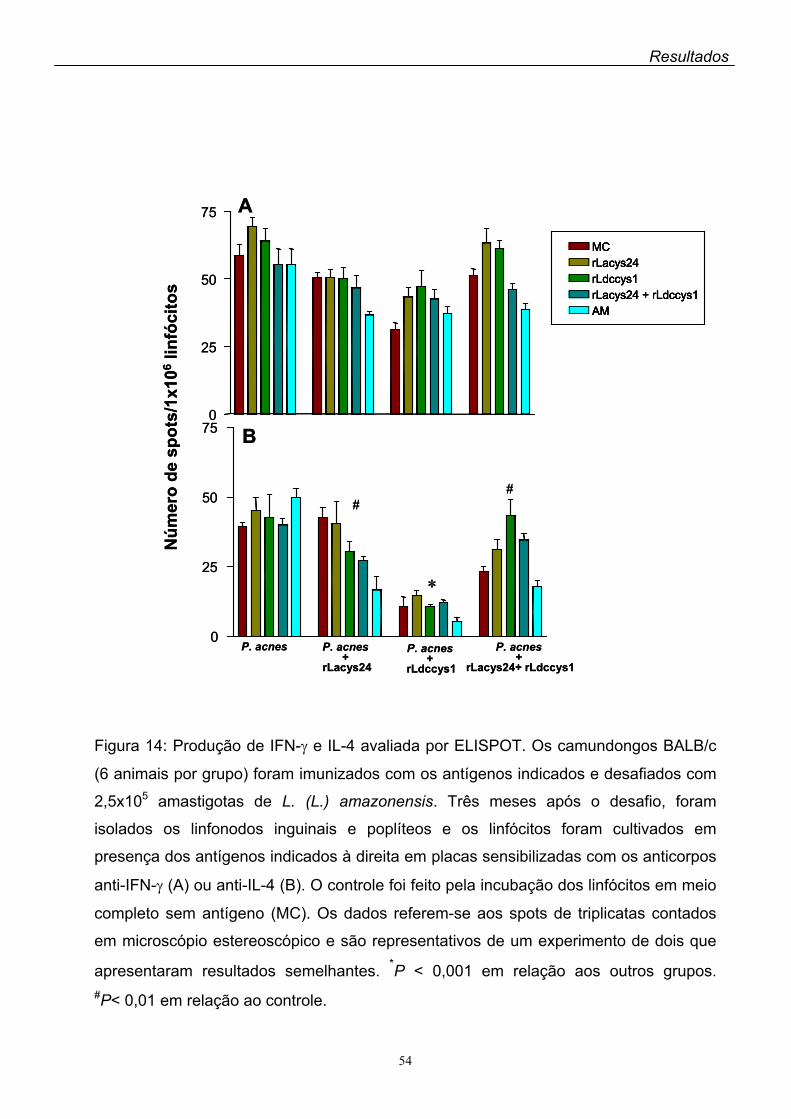
Resultados
0
25
50
750
25
50
75
P. acnes +
rLacys24P. acnes
+rLdccys1
P. acnes+
rLacys24+ rLdccys1
P. acnes
Núm
ero
desp
ots/
1x10
6lin
fóci
tos
A
B
MCrLacys24rLdccys1rLacys24 + rLdccys1AM
MCrLacys24rLdccys1rLacys24 + rLdccys1AM
*
##
0
25
50
750
25
50
75
P. acnes +
rLacys24P. acnes
+rLdccys1
P. acnes+
rLacys24+ rLdccys1
P. acnes
Núm
ero
desp
ots/
1x10
6lin
fóci
tos
A
B
MCrLacys24rLdccys1rLacys24 + rLdccys1AM
MCrLacys24rLdccys1rLacys24 + rLdccys1AM
*
##
Figura 14: Produção de IFN-γ e IL-4 avaliada por ELISPOT. Os camundongos BALB/c
(6 animais por grupo) foram imunizados com os antígenos indicados e desafiados com
2,5x105 amastigotas de L. (L.) amazonensis. Três meses após o desafio, foram
isolados os linfonodos inguinais e poplíteos e os linfócitos foram cultivados em
presença dos antígenos indicados à direita em placas sensibilizadas com os anticorpos
anti-IFN-γ (A) ou anti-IL-4 (B). O controle foi feito pela incubação dos linfócitos em meio
completo sem antígeno (MC). Os dados referem-se aos spots de triplicatas contados
em microscópio estereoscópico e são representativos de um experimento de dois que
apresentaram resultados semelhantes. *P < 0,001 em relação aos outros grupos. #P< 0,01 em relação ao controle.
54
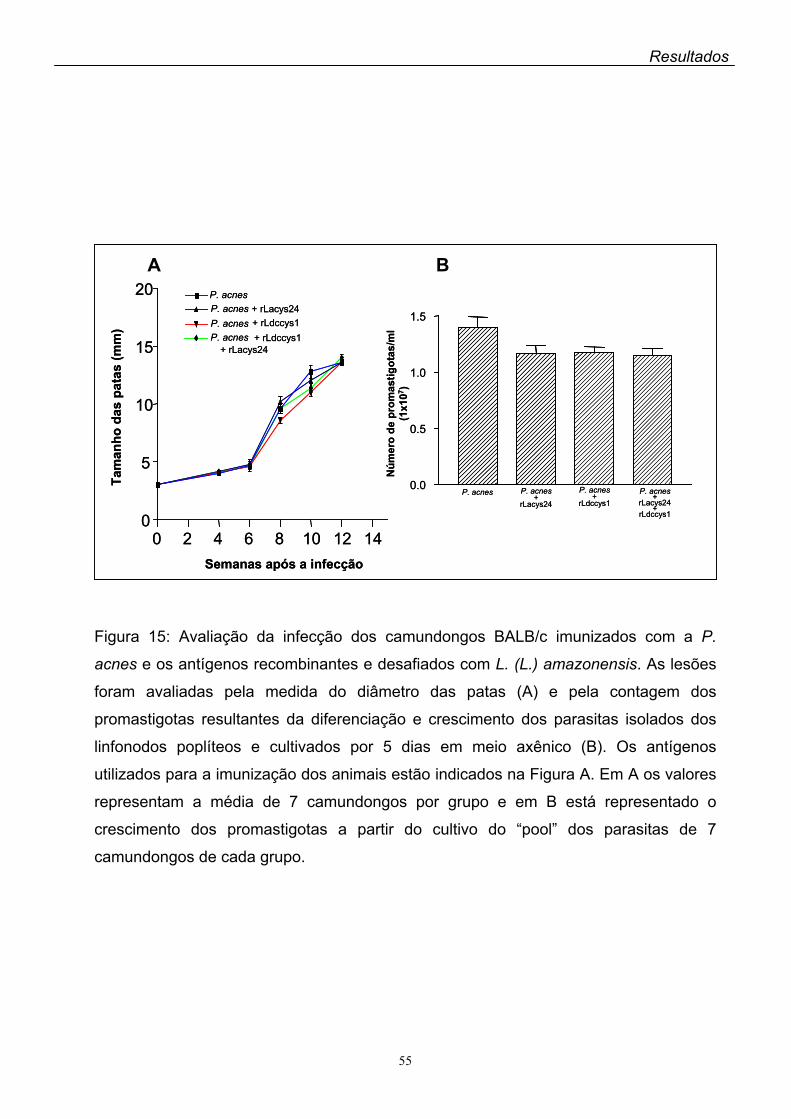
Resultados
55
A B
Semanas após a infecção
Tam
anho
das
pat
as (m
m)
0 2 4 6 8 10 12 140
5
10
15
20 P. acnesP. acnes + rLacys24P. acnes + rLdccys1P. acnes
+ rLacys24 + rLdccys1
Semanas após a infecção
Tam
anho
das
pat
as (m
m)
0 2 4 6 8 10 12 140
5
10
15
20 P. acnesP. acnes + rLacys24P. acnes + rLdccys1P. acnes
+ rLacys24 + rLdccys1
Núm
ero
depr
omas
tigot
as/m
l(1
x107 )
0.0
0.5
1.0
1.5
P. acnes P. acnes +
rLdccys1P. acnes
+ rLacys24
P. acnes +
rLacys24+
rLdccys1
Núm
ero
depr
omas
tigot
as/m
l(1
x107 )
0.0
0.5
1.0
1.5
P. acnes P. acnes +
rLdccys1P. acnes
+ rLacys24
P. acnes +
rLacys24+
rLdccys1
Figura 15: Avaliação da infecção dos camundongos BALB/c imunizados com a P.
acnes e os antígenos recombinantes e desafiados com L. (L.) amazonensis. As lesões
foram avaliadas pela medida do diâmetro das patas (A) e pela contagem dos
promastigotas resultantes da diferenciação e crescimento dos parasitas isolados dos
linfonodos poplíteos e cultivados por 5 dias em meio axênico (B). Os antígenos
utilizados para a imunização dos animais estão indicados na Figura A. Em A os valores
representam a média de 7 camundongos por grupo e em B está representado o
crescimento dos promastigotas a partir do cultivo do “pool” dos parasitas de 7
camundongos de cada grupo.

Discussão
56

Discussão
A abordagem do presente trabalho foi avaliar a capacidade de uma cisteína
proteinase recombinante de L. (L.) amazonensis induzir respostas imunes protetoras
contra a infecção homóloga em modelo murino. A maior parte dos estudos de
cisteína proteinases de Leishmania enfoca a participação dessas enzimas como
fatores de virulência desses protozoários aos macrófagos e na sobrevivência e
crescimento desses parasitas nos hospedeiros vertebrados (Mottram et al., 1996;
Mottram et al., 1998). Por outro lado, são bem mais escassos os trabalhos que
analisam a implicação de cisteína proteinases de Leishmania em respostas imunes
celulares. Enquanto duas cisteína proteinases caracterizadas em amastigotas de L.
(L.) infantum, CPA e CPB, foram utilizadas para a imunização protetora de
camundongos BALB/c e cães contra a leishmaniose visceral (Rafati et al., 2000;
Rafati et al., 2005), a utilização de cisteína proteinases das espécies cutâneas de
Leishmania como imunógenos tem sido pouco explorada. Demonstrou-se que a CPa
de L. (L.) major induziu a produção de IFN-γ e proteção de camundongos BALB/c
contra a infecção homóloga (Rafati et al., 2000) e mais recentemente proteção
duradoura foi observada pela imunização dos camundongos com a mistura dos
genes cpa e cpb, assim como com as cisteína proteinases recombinantes (rCPa e
rCPb) (Rafati et al., 2006). No modelo L. (L.) amazonensis não existem relatos da
literatura sobre a utilização de cisteína proteinases na indução de respostas imunes,
tendo sido pioneiro o trabalho em que a forma nativa de uma cisteína proteinase de
30 kDa de amastigotas de L. (L. ) amazonensis conferiu imunidade protetora em
camundongos BALB/c contra o desafio homólogo (Beyrodt et al., 1997).
No presente trabalho inicialmente um fragmento de 500 pb, codificador de
cisteína proteinase de amastigotas de L. (L.) amazonensis originado previamente em
nosso laboratório, foi subclonado em vetor de expressão bacteriano. Esse clone,
denominado Lacys24 e anteriormente clonado no vetor pMOS, apresentou
identidade superior a 90% com os genes codificadores de cisteína proteinases de L.
(L.) pifanoi e L. (L.) mexicana denominados, respectivamente, Lpcys1 e Lcpa,
enquanto que um menor grau de identidade, porém significante, com os genes
Ldccys2 de L. (L.) chagasi e cpa de L. (L.) major (Oliveira, 2001). A análise
comparativa da seqüência de nucleotídeos do clone Lacys24 anteriormente obtido
com a seqüência de um dos três clones resultantes da amplificação do Lacys24 por
PCR, realizada no presente trabalho, mostrou um alto grau de similaridade entre
essas seqüências (Figura 4). A localização dos prováveis epítopos de células T do
57

Discussão
clone Lacys24 pelo programa AMPHI mostrou três motivos de ligação ao MHC de
classe II e um ao MHC de classe I na seqüência desse clone (Figura 5). Esses
resultados indicam a possibilidade de participação de linfócitos T CD4+ e CD8+ nas
respostas induzidas pela proteína codificada pelo clone Lacys24, levando-nos a
subcloná-lo no vetor de expressão bacteriano pHIS paralelo 3. A expressão do clone
Lacys24 nesse vetor originou uma proteína de tamanho esperado de 24 kDa
(rLacys24) cuja pureza foi demonstrada por PAGE-SDS após a passagem do extrato
bacteriano em coluna de níquel (Figura 6). Por “Western blotting” verificou-se que o
soro policlonal de camundongo anti-rLacys24 reagiu com uma proteína de 30 kDa do
extrato de amastigotas de L. (L.) amazonensis (Figura 7). Esse resultado indica que
a rLacys24 corresponde a uma das isoformas de cisteína proteinase de 30 kDa
abundantemente expressas em amastigotas de L. (L.) amazonensis. Por outro lado,
a rLacys24 não foi reconhecida pelo anticorpo monoclonal 2E5D3 produzido contra
uma isoforma de 30 kDa de amastigotas de L. (L.) amazonensis (Beyrodt et al.,
1997). A não reatividade da rLacys24 com esse anticorpo pode indicar que o clone
Lacys24 codifica uma isoforma diferente da reativa com o MoAc 2E5D3. A outra
possibilidade é que o epítopo reconhecido por esse anticorpo pode ser codificado
por uma região do gene não presente no fragmento de 500 pb amplificado para a
obtenção do clone Lacys24, pois a clonagem e análise da seqüência completa do
gene Lacys24 mostrou que ele apresenta 1,08 kb (Lasakosvitsch et al., 2003).
A antigenicidade da rLacys24 foi primeiramente avaliada pela sua capacidade
de induzir respostas proliferativas de linfócitos CD4+ e CD8+ em camundongos
BALB/c utilizando-se ACF ou P. acnes como adjuvantes para a imunização dos
animais com a proteína recombinante. Os resultados desses experimentos
permitiram concluir que a imunização subcutânea com a rLacys24 + ACF levou ao
aumento da população T CD8+, enquanto que não houve diferença na expressão
dos linfócitos T CD4+ nesses animais comparada à dos controles imunizados apenas
com o adjuvante (Figura 8). Por outro lado, a imunização com a rLacys24 + P. acnes
por via intraperitoneal não levou ao aumento da expressão de CD4+ ou CD8+ no
baço dos camundongos imunizados (Figura 9).
O aumento da população de linfócitos T CD8+ nos linfonodos poplíteos e
inguinais dos camundongos BALB/c levou-nos a testar a possível atividade citotóxica
desses linfócitos sobre macrófagos infectados com L. (L.) amazonensis. Observou-
se que a atividade citotóxica dos linfócitos dos animais imunizados com a rLacys24
58

Discussão
foi significantemente maior que a exercida pelos linfócitos dos controles que
receberam apenas o ACF (Figura 10). Esses resultados mostram que os linfócitos T
CD8+ induzidos pela rLacys24 reconhecem epítopos dos antígenos de L. (L.)
amazonensis naturalmente processados e apresentados pelos macrófagos
infectados. Apesar de escassos, alguns exemplos de parasitas que residem em
compartimentos vacuolares de macrófagos e são capazes de acessar a via classe I
de apresentação aos linfócitos têm sido mostrados com Chlamydia trachomatis e
Toxoplasma gondii (Beatty and Stephens, 1994; Denkers et al., 1993). Com
Leishmania foi demonstrado que macrófagos infectados com L. (L.) major, L. (L.)
infantum e L. (L.) amazonensis podem apresentar antígenos dos parasitas via MHC
classe I aos linfócitos CD8+ (Conceição-Silva et al., 1994; Pinelli et al., 1995, 1995;
Kima et al., 1997).
O possível poder protetor induzido nos camundongos BALB/c contra a
infecção homóloga foi analisado pelo desafio com L. (L.) amazonensis dos animais
imunizados com a rLacys24 juntamente com ACF ou P. acnes. Os resultados desses
experimentos mostraram que enquanto os animais imunizados com a rLacys24 + P.
acnes não apresentaram diferenças no diâmetro das lesões das patas durante a
infecção com L. (L.) amazonensis, as lesões dos camundongos imunizados com a
rLacys24 +ACF foram menores que as dos controles que receberam apenas o ACF
(Figura 11). Esses resultados sugerem que a proteção parcial contra a L. (L.)
amazonensis exibida por esses animais pode estar relacionada à atividade citotóxica
dos linfócitos CD8+ observada in vitro pela lise dos macrófagos infectados com L.
(L.) amazonensis (Figura 10). Embora a maior parte dos trabalhos de imunização
contra o desafio com L. (L.) amazonensis mostre o envolvimento de linfócitos CD4+
Th1 produtores de IFN-γ na proteção dos animais imunizados (Campbell et al., 2003;
Coelho et al., 2003; Campbell et al., 2004; Pinto et al., 2004; Aguilar-Be et al., 2005;
Vasquez e Soong, 2006), a participação de linfócitos T CD8+ com produção de
perforina, além de IFN-γ, foi demonstrada em camundongos BALB/c imunizados com
o antígeno P8 de L. (L.) pifanoi e protegidos contra o desafio com L. (L.)
amazonensis (Colmenares et al., 2003).
Os resultados de proteção parcial dos camundongos pela rLacys24
encorajaram-nos a testar outro antígeno, a cisteína proteinase recombinante de
amastigotas de L. (L.) chagasi, rLdccys1, visando à maior proteção dos animais
contra o desafio com a L. (L.) amazonensis. Nesses experimentos a P. acnes foi
59

Discussão
utilizada como adjuvante para a imunização e o desafio com a L. (L.) amazonensis.
Os animais imunizados com a rLacys24, a rLdccys1 ou os dois antígenos
recombinantes associados, não apresentaram diferenças do tamanho das lesões
comparados aos controles que receberam apenas o adjuvante (Figura 15A). Esses
resultados foram confirmados pelo outro parâmetro utilizado para a avaliação da
infecção, o crescimento dos promastigotas resultantes da diferenciação dos
amastigotas isolados das lesões em meio axênico (Figura 15B).
A análise das respostas imunes por citometria de fluxo mostrou o predomínio
de linfócitos CD4+ nos animais imunizados com os antígenos recombinantes e
desafiados (Figura 13) e não houve diferença significante da porcentagem dessas
células nos controles que receberam apenas a P. acnes. Resultados semelhantes
também foram observados em relação à produção de IFN-γ, pois houve secreção
dessa linfocina pelos linfócitos de todos os animais analisados (Figura 14). Não
houve também diferenças quanto à secreção de IFN-γ quando os linfócitos foram
reestimulados ou não in vitro com os antígenos recombinantes. Isso provavelmente
se deve à forte estimulação antigênica dos animais pela L. (L.) amazonensis, pois a
análise das respostas imunes foi feita três meses após o desafio e, como foi
observado, o número de parasitas nas lesões das patas era muito grande. É
importante lembrar também que nas infecções de camundongos pela L. (L.)
amazonensis existe uma infiltração muito grande de parasitas nos linfonodos
regionais.
Os resultados de indução de respostas imunes mediadas por linfócitos CD4+
com a produção de IFN-γ nos controles que receberam apenas o adjuvante
corroboram os dados da literatura que mostram que a P. acnes, empregada no
tratamento de pacientes com imunossupressão, induz a síntese de citocinas pró-
inflamatórias, como IL-1α, IL-6, TNF-α e principalmente IL-12 e IL-18 que levam à
liberação de IFN-γ (Bomford, 1980; Matsui et al., 1997). P. acnes foi utilizada como
adjuvante na imunização gênica de camundongos BALB/c contra a infecção com
Trypanosoma cruzi, induzindo forte resposta mediada por linfócitos CD4+ Th1 e
significante redução da parasitemia dos animais imunizados (Mussalem et al., 2006).
Na leishmaniose experimental, a P. acnes foi utilizada em experimentos de
imunização com vários antígenos de Leishmania em animais suscetíveis, resultando
em imunidade protetora (Scott et al., 1987; Jaffe et al., 1990; White and McMahon-
60

Discussão
Pratt, 1990; Scott et al., 1990; Soong et al., 1995; Ghosh et al., 2002; Colmenares et
al., 2003). Em nosso laboratório, utilizou-se a P. acnes como adjuvante para a
imunização de camundongos BALB/c com uma cisteína proteinase nativa e
recombinante de L. (L.) chagasi (rLdccys1) que resultou em forte proteção contra o
desafio homólogo mediada por respostas Th1 (Pinto et al, 2000; Ferreira, 2006;
Ferreira et al., 2007). No presente trabalho, não houve aumento das respostas
imunes induzidas pelos antígenos recombinantes nos animais imunizados em
relação às desencadeadas pela P. acnes. Quanto à rLdccys1, esses resultados
diferem dos observados quando esse antígeno foi utilizado para a imunização dos
camundongos BALB/c e desafio com a L. (L.) chagasi. Vários fatores devem ser
considerados que levaram à discrepância das respostas imunes à rLddcys1
observada. Um deles é a via de imunização, pois nos experimentos com a L. (L.)
chagasi os camundongos foram imunizados pela via intraperitoneal. No presente
trabalho a via subcutânea foi a escolhida visando privilegiar a resposta nos
linfonodos regionais, já que o desafio foi com uma espécie cutânea de Leishmania.
O outro fator, e provavelmente o mais relevante, refere-se à suscetibilidade dos
camundongos BALB/c às duas espécies de Leishmania utilizadas. Enquanto nesses
animais a infecção pela L. (L.) chagasi evolui para uma doença subclínica, há
progressão acentuada das lesões cutâneas pela L. (L.) amazonensis, com uma
infiltração muito grande de amastigotas nessas lesões.
A única diferença entre as respostas imunes dos controles e as dos animais
imunizados com os antígenos recombinantes foi a produção de IL-4. Essa linfocina
foi detectada nas culturas de linfócitos de todos os animais, porém a sua secreção
foi significantemente menor nos camundongos imunizados com os antígenos, sendo
a menor expressão observada nas culturas dos linfócitos dos camundongos
imunizados com a rLdccys1 (Figura 14B). Com base nesses achados, poderia ser
aventado que no grupo que recebeu apenas a P. acnes a IL-4 estaria exercendo um
controle negativo sobre a possível ação leishmanicida dos macrófagos por IFN-γ.
Entretanto, nos animais imunizados com a rLacys24 ou a rLdccys1 a secreção de IL-
4 foi significantemente reduzida e esses animais também não apresentaram
diferenças do tamanho das lesões. Esses resultados corroboram as evidências da
literatura que demonstraram que as lesões crônicas características da infecção por
L. (L.) amazonensis em animais suscetíveis são independentes da expressão de IL-4
(Afonso & Scott, 1993; Jones et al., 2000). Em nossos experimentos foi também
61

Discussão
avaliada a secreção de IL-10, porém essa linfocina não foi detectada nas culturas
dos linfócitos. Assim como a IL-4, a IL-10 não pode ser correlacionada com a
persistência da infecção pela L. (L.) amazonensis nos animais imunizados, tendo
sido observado que não há cura das lesões em camundongos C57BL/6 deficientes
em IL-10 (Jones et al., 2002). Todos esses achados indicam que, como algumas
evidências apontam, enquanto a dicotomia Th1/Th2 é correlacionada com a
suscetibilidade/resistência à L. (L.) major em modelo murino, a suscetibilidade à L.
(L.) amazonensis não está associada a respostas polarizadas para Th2 (Afonso and
Scott, 1993; Soong et al., 1997). Além disso, a resistência não pode ser
correlacionada ao predomínio de respostas Th1, pois embora a ativação dos
macrófagos por IFN-γ exerça um efeito leishmanicida sobre a L. (L.) amazonensis,
esse efeito pode ser modulado pela inibição da síntese de óxido nítrico pelos
amastigotas nos macrófagos infectados (Balestieri et al., 2002). Apesar de esses
autores terem observado que a ativação por IFN-γ é capaz de reverter essa inibição,
parece ocorrer também uma modulação negativa do efeito leishmanicida do óxido
nítrico pela L. (L.) amazonensis nos macrófagos infectados. Isso pode ser observado
quando macrófagos infectados são ativados com IFN-γ e LPS com produção de
níveis significantes de óxido nítrico que, no entanto, é acompanhada pela destruição
de apenas 20-25% dos amastigotas intracelulares (comunicação pessoal).
Outro fator importante a ser considerado na infecção por L. (L.) amazonensis
é a forma do parasita utilizada para o desafio dos animais. A indução de respostas
mediadas por Th1 pela transferência endovenosa de uma linhagem celular secretora
de IFN-γ e TNF-α não foi capaz de controlar a infecção de camundongos C57BL/6
por amastigotas de L. (L.) amazonensis, enquanto que reduziu mais de 1.0000
vezes a carga parasitária dos animais desafiados com as formas promastigotas.
Ensaios com macrófagos infectados in vitro com L. (L.) amazonensis e ativados com
IFN-γ corroboraram esses achados, pois a atividade leishmanicida dos macrófagos
tratados foi observada somente quando eles eram infectados com os promastigotas
(Qi et al., 2004). Em nossos ensaios os animais foram desafiados com amastigotas
recém-isolados das lesões e isso pode ter limitado a resolução da infecção nos
animais imunizados mesmo na presença de IFN-γ. Isso indica que novos esquemas
de imunização com a rLacys24 e a rLdccys1 devem ser testados visando à indução
62

Discussão
63
de outros fatores que possam exercer uma ação sinergística com o IFN-γ que
resultem em um efeito leishmanicida mais eficaz.
É importante ressaltar ainda que nos animais imunizados com os antígenos
recombinantes não houve expressão significante de linfócitos CD8+ (Figura 13) que,
como foi mostrado nesse trabalho (Figuras 8, 10 e 11) e apoiado por alguns dados
da literatura (Colmenares et al., 2003), exercem um papel importante na redução da
carga parasitária dos animais infectados com L. (L.) amazonesnis.
Um outro ponto a ser considerado é que o efeito protetor parcial obtido pela
imunização com a rLacys24 e ACF como adjuvante mostrada no presente trabalho
foi menor do que o previamente obtido pela imunização com a cisteína proteinase de
30 kDa nativa de L. (L.) amazonensis (Beyrodt et al., 1997). Essa diferença pode
estar relacionada ao próprio antígeno utilizado, pois como anteriormente
especificado, as cisteína proteinases de 30 kDa dos amastigotas de L. (L.)
amazonensis são expressas como isoformas e muito provavelmente a Lacys24
representa uma isoforma diferente da anteriormente utilizada para a imunização dos
animais.
Em conclusão, apesar da reduzida proteção conferida pela rLacys24
observada no presente trabalho, os dados obtidos sugerem que esse antígeno pode
ser utilizado em novas estratégias de imunização que induzam respostas mediadas
por linfócitos CD4+ e CD8+ mais eficazes contra a infecção pela L. (L.) amazonensis.

Referências Bibliográficas
64

Referências Bibliográficas
AEBISCHER T, WOLFRAM M, PATZER SI, ILG T, WIESE M, OVERATH P. Subunit vaccination
of mice against New World cutaneous leishmaniasis: comparison of three proteins
expressed in amastigotes and six adjuvants. Infect Immun. 2000; 68: 1328-1336.
AFONSO LCC, SCOTT P. Immune responses associated with susceptibility of C57BL/10
mice to Leishmania amazonensis. Infect Immun. 1993; 61: 2952-2959.
AGUILAR-BE I, ZARDO RS, PARAGUAI DE SOUZA E, BORJA-CABRERA GP, ROSADO-VALLADO
M, MUT-MARTIN M, GARCÍA-MISS MR, PALATNIK DE SOUSA CB, DUMONTEIL E. Cross-
protective efficacy of a prophylactic Leishmania donovani DNA vaccine against visceral
and cutaneous murine leishmaniasis. Infect Immun. 2005; 73: 812-819.
AHMED SBHA, BAHLOUL C, ROBBANA C, ASKRI S, DELLAGI K. A comparative evaluation of
different DNA vaccine candidates against experimental murine leishmaniasis due to L.
major. Vaccine. 2004; 22: 1631-1639.
ALBERIO SO, DIAS SS, FARIA FP, MORTARA RA, BARBIÉRI CL, HAAPALAINEN EF.
Ultrastructural and cytochemical identification of megasome in Leishmania (Leishmania)
chagasi. Parasitol Res. 2004; 92: 246-254.
ALEXANDER J, COOMBS GH, MOTTRAM JC. Leishmania mexicana cysteine proteinase-
deficient mutants have attenuated virulence for mice and potentiate a Th1 response. J Immunol. 1998; 161: 6794-6801.
BALESTIERI FMP, QUEIROZ ARP, SCAVONE C, COSTA VMA, BARRAL-NETO M,
ABRAHAMSOHN I. Leishmania (L.) amazonensis-induced inhibition of nitric oxide
synthesis in host macrophages. Microbes Infect. 2002; 4: 23-29.
BARBIÉRI CL, DOINE AI, FREYMÜLLER E. Lysosomal depletion in macrophages from
spleen and foot lesions of Leishmania-infected hamster. Exp Parasitol. 1990; 71: 218-
228.
BARRAL-NETO M, VON SOHSTEN RL, TEIXEIRA M, SANTOS WL, POMPEU ML, MOREIRA RA,
OLIVEIRA JT, CAVADA BS, FALCOFF E, BARRAL A. In vivo protective effect of the lectin
65

Referências Bibliográficas
from Canavalia brasiliensis on BALB/c mice infected by Leishmania amazonensis. Acta Trop. 1996; 60: 237-250.
BEATTY PR, STEPHENS RS. CD8+ T lymphocyte-mediated lysis of Chlamydia-infected L
cells using an endogenous antigen pathway. J Immunol. 1994; 153: 4588-4595.
BELKAID Y. The role of CD24+ CD25+ regulatory T cells in Leishmania infection. Expert. Opin. Biol. Ther. 2003; 3: 1-11.
BEYRODT CGP, PINTO AR, FREYMÜLLER E, BARBIÉRI CL. Characterization of an antigen
from Leishmania amazonensis amastigotes able to elicit protective responses in a
murine model. Infect Immun. 1997; 65: 2052-2059.
BEYRODT CGP. Caracterização molecular de um antígeno de 30 kDa de Leishmania
(Leishmania) amazonensis envolvido na resposta imune celular em modelo murino.
[Tese de Doutorado] São Paulo: Universidade Federal de São Paulo – Escola Paulista
de Medicina; 1998.
BEYRODT, CGP, SILVEIRA, JF, BARBIÉRI, CL. “Characterization of na antigen from
Leishamania (L.) amazonensis amastigotes which elicits protective responses in mice”.
Mem Inst Oswaldo Cruz. 1996; 91: 37-38.
BOMFORD R. The comparative selectivity of adjuvants for humoral and cell-mediated
immunity. II. Effect on delayed-type hypersensitivity in the mouse and guinea pig, and
cell-mediated immunity to tumour antigens in the mouse of Freund's incomplete and
complete adjuvants, alhydrogel, Corynebacterium parvum, Bordetella pertussis,
muramyl dipeptide and saponin. Clin Exp Immunol. 1980; 39: 435–441.
BROOKS DR, TETLEY L, COOMBS GH, MOTTRAM JC. Processing and trafficking of cysteine
proteases in Leishmania mexicana. J Cell Sci. 2000; 113: 4035.
CAMPBELL K, DIAO H, JI J, SOONG L. DNA immunization with the gene encoding P4
nuclease of Leishmania amazonensis protects mice against cutaneous leishmaniasis.
Infect Immun. 2003; 71: 6270-6278.
66

Referências Bibliográficas
CAMPBELL K, POPOV V, SOONG L. Identification and Molecular Characterization of a
Gene Encoding a Protective Leishmania amazonensis Trp-Asp (WD) Protein. Infect Immun. 2004; 72: 2194-2202.
CHAMPSI J, MCMAHON-PRATT D. Membrane glycoprotein M-2 protects against
Leishmania amazonensis infection. Infect Immun. 1988; 56: 3272-3279.
CHAN MM. T cell response in murine Leishmania mexicana amazonensis infection:
production of interferon-g by CD8+ cells. Eur J Immunol. 1993; 23: 1181-1184.
CHATELAIN R, VARKILA K, COFFMAN RL. IL-4 induces a Th2 response in Leishmania
major-infected mice. J Immunol. 1992; 148: 1182-1187.
COELHO EAF, TAVARES CAP, CARVALHO FAA, CHAVES KF, TEIXEIRA KN, RODRIGUES RC,
CHAREST H, MATLASHEWSKI G, GAZZINELLI RT, FERNANDES AP. Immune responses
induced by the Leishmania (Leishmania) donovani A2 antigen, but not by the LACK
antigen, are protective against experimental Leishmania (Leishmania) amazonensis
infection. Infect Immun. 2003; 71: 3988-3994.
COLMENARES M, KIMA PE, SAMOFF E, SOONG L, MCMAHOM-PRATT D. Perforin and
Gamma Interferon are critical CD8+ T-cell-mediated responses in vaccine-induced
immunity against Leishmania amazonensis infection. Infect Immun. 2003; 71: 3172-
3182.
CONCEIÇÃO-SILVA F, PERLAZA BL, LOUIS JA, ROMERO P. Leishmania major infection in
mice primes for specific major histocompatibility complex class I restricted CD8+
cytotoxic T cell responses. Eur J Immunol. 1994; 24: 2813-2817.
CONNEL ND, MEDINA-COSTA E, MCMASTER R, BLOOM BR, RUSSEL DG. Effective
immunization against cutaneous leishmaniasis with recombinant bacille Calmette-
Guérin expressing the Leishmania surface proteinase gp63. Proc Natl Acad Sci. 1993;
90: 11473-11477.
67

Referências Bibliográficas
COSTA-SOUZA BLS, PINTO EF, DELMÓRIO P, RÉ MI, ROSSI-BERGMAN B. “Development of
delivery system in glyceraldehyde-crosslinked microparticles of chitosan for intranasal
vaccination with antileishmanial LACK DNA”. XXXIV Annual Meeting on Basic Research in Chagas’ Disease, XXIII Annual Meeting of the Brazilian Society of Protozoology., 2007; pg. 126.
CZERKINSKY C, NILSSON L, NYGREN H, OUCHTERLONY O, TARKOWSKI A. A solid-phase
enzyme-linked immunospot (ELISPOT) assay for enumeration of specific antibody-
secreting cells. J Immunol Methods. 1983; 65: 109-121.
DAS L, DATTA N, BANDYOPADHYAY S, DAS PK. Successful therapy of lethal murine
visceral leishmaniasis with cystatin involves up-regulation of nitric oxide and a favorable
T cell response. J Immunol. 2001; 166: 4020-4028.
DENKERS EY, SHER A, GAZZINELLI RT. CD8+ T-cell interactions with Toxoplasma gondii:
implications for processing of antigens for class-I-restricted recognition. Res Immunol. 1993; 144: 51-57.
DESJEUX P. Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis. 2004; 27: 305-318.
DUBOISE SM, VANNIER-SANTOS MA, COSTA-PINTO D, RIVAS L, PAN AA, TRAUB-CZECO Y,
SOUZA W, MCMAHON-PRATT D. The biosynthesis, processing and immunolocalization of
Leishmania pifanoi amastigote cysteine proteinase. Mol Biochem Parasitol. 1994; 68:
119-132.
DUMONTEIL E, JESUS RSM, JAVIER EO, ROSARIO GMM. DNA vaccines induce parcial
protection against Leishmania mexicana. Vaccine. 2003; 21: 2161-2168.
EAKIN AE, MILLS AA, HARTH G, MCKERROW JH, CRAIK CS. The sequence, organization
and expression of the major cysteine proteinase (cruzain) from Trypanosoma cruzi. J Biol Chem. 1992; 267: 7411-7420.
68

Referências Bibliográficas
FERREIRA JHL, GENTIL LG, DIAS SS, FEDELI CEC, KATZ S, BARBIÉRI CL. Immunization
with the cysteine proteinase Ldccys1 gene from Leishmania (Leishmania) chagasi and
the recombinant Ldccys1 protein elicits protective immune responses in a murine model
of visceral leishmaniasis. Vaccine. 2007; in press.
Ferreira JHL. Utilização do gene codificador da cisteína proteinase de 30 kDa de
Leishmania (Leishmania) chagasi (Ldccys1) e da proteína recombinante (rLdccys1) em
novos esquemas de imunização em modelo murino. [Tese de Mestrado] São Paulo:
Universidade Federal de São Paulo – Escola Paulista de Medicina; 2006.
GHOSH A, LABRECQUE S, MATLASHEWSKI G. Protection against Leishmania donovani
infection by DNA vaccination: increased DNA vaccination efficiency through inhibiting
the cellular p53 response. Vaccine. 2001; 19: 3169-3178.
GHOSH A, ZHANG WW, MATLASHEWSKI G. Immunization with A2 protein results in a
mixed Th1/Th2 and a humoral response which protects mice against Leishmania
donovani infections. Vaccine. 2002; 20: 59-66.
GURUNATHAN S, STOBIE L, PRUSSIN C, SACKS DL, GLAINCHENHAUS N, FOWELL DJ,
LOCKSLEY RM, CHANG JT, WU CY, SEDER RA. Requeriment for the maintenance of Th1
immunity in vivo following DNA vaccination: a potential immunoregulatory role for CD8+
T cells. J Immun. 2000; 165: 915-924.
HANAHAN D. Techiniques for transformation of E. coli in DNA cloning: a practical
approach (DM glover, Ed.); 1985; vol. 1, pp 109-175.
HANDMAN E, SYMONS FM, BALDWIN TM, CURTIS JM, SCHEERLINCK JPY. Protective
vaccination with promastigote surface antigen 2 from Leishmania major is mediated by
Th1 type of immune response. Infect Immun. 1995; 63: 4261-4267.
HANDMAN E. Leishmaniasis: current status of vaccine development. Clin Microbiol Review. 2001; 14: 229-243.
69

Referências Bibliográficas
HEINZEL FP, SADICK MD, HOLADAY BJ, COFFMAN RL, LOCKSLEY RM. Reciprocal
expression of interferon gamma or interleukin 4 during the resolution or progression of
murine leishmaniasis. Evidence for expansion of distinct helper T cell subsets. J Exp Med. 1989; 169: 59-72.
IBORRA S, SOTO M, CARRIÓN J, ALONSO C, REQUENA JM. Vaccination with a plasmid DNA
cocktail encoding the nucleosomal histones of Leishmania confers protection against
murine cutaneous leishmaniasis. Vaccine. 2004; 22: 3865-3876.
INGRID AGUILAR-BE I, ZARDO RS, PARAGUAI DE SOUZA E, BORJA-CABRERA GP, ROSADO-
VALLADO M, MUT-MARTIN M, GARCÍA-MISS MR, PALATNIK DE SOUSA CB, DUMONTEIL E.
Cross-protective efficacy of a prophylactic Leishmania donovani DNA vaccine against
visceral and cutaneous murine leishmaniasis. Infect Immun. 2005; 73: 812-819.
JI J, MASTERSON J, SUN J, SOONG L. CD4+CD25+ regulatory T cells restrain pathogenic
responses during Leishmania amazonensis infection. J Immunol. 2005; 174: 7147-
7153.
JI J, SUN J, QI H, SOONG L. Analysis of T helper cell responses during infection with
Leishmania amazonesnis. Am J Trop Med Hyg. 2002; 66: 338-345.
JONES DE, ACKERMANN MR, WILLE U, HUNTER CA, SCOTT P. Early enhanced Th1
response after Leishmania amazonensis infection of C57BL/6 interleukin-10-deficient
mice does not lead to resolution of infection. Infect Immun. 2002; 70: 2151-2158.
JONES DE, BUXBAUM LU, SCOTT P. IL-4-independent inhibition of IL-12 responsiveness
during Leishmania amazonensis infection. J Immunol. 2000; 165: 364-372.
KANE MM, MOSSER DM. The role of IL-10 in promoting disease progression in
leishmaniasis. J. Immunol. 2001; 166: 1141-1147.
KIMA PE, RUDDLE NH, MCMAHON-PRATT D. Prresentation via the class I pathway by
Leishmania amazonensis-infected macrophages of an endogenous leishmanial antigen
to CD8+ T cells. J Immunol. 1997; 159: 1828-1834.
70

Referências Bibliográficas
KOPF M, BROMBACHER F, KOHLER G, KIENZLE G, WIDMANN KH, LEFRANG K, HUMBORG C,
LEDERMANN B, SOLBACH W. IL-4-deficient Balb/c mice resist infection with Leishmania
major. J Exp Med. 1996; 184: 1127-1136.
LASAKOSVITSCH F, GENTIL LG, SANTOS MRM, SILVEIRA JF, BARBIÉRI CL. Cloning and
characterisation of a cysteine proteinase gene expressed in amastigotes of Leishmania
(L.) amazonensis. Int J Parasitol. 2003; 33: 445-454.
LIEW FY, LI Y, MILLOTT S. Tumor necrosis factor-alpha synergizes with IFN-gamma in
mediating killing of Leishmania major through the induction of nitric oxide. J. Immunol. 1990; 145: 4306-4310.
MARGALIT H, SPOUGE JL, CORNETTE JL, CEASE KB, DELISI C, BERZOFSKY JA. Prediction of
immunodominat helper T cell antigenic sites from the primary sequence. J Immunol. 1987; 138: 2213-2229.
MATSUI K, YOSHIMOTO T, TSUTSUI H, HYODO Y, HAYASHI N, HIROISHI K, KAWADA N,
OKAMURA H, NAKANISHI K, HIGASHINO K. Propionibacterium acnes treatment diminishes
CD4+NK1.1+ T cells in the liver by induction of IL-12 and IL-18 production from Kupffer
cells. J Immunol. 1997; 159: 97-106.
MCKERROW JH. Parasite proteases. Exp Parasitol. 1989; 68: 111-115.
MCMAHON-PRATT D, ALEXANDER J. Does the Leishmania major paradigm of
pathogenesis and protection hold for new world cutaneous leishmaniasis or the visceral
disease? Immunol Rev. 2004; 201: 206-221.
MEDINA-ACOSTA E, CROSS GA. Rapid isolation of DNA from trypanosomatid protozoa
using simple "mini-prep" procedure. Mol Biochem Parasitol. 1993; 59: 327-329.
MOTTRAM JC, BROOKS DR, COOMBS GH. Roles of cysteine proteinases of trypanosomes
and Leishmania in host-parasite interactions. Curr Opin Microbiol. 1998; 1: 455-460.
71

Referências Bibliográficas
MOTTRAM JC, FRAME MJ, BROOKS DR, TETLEY L, HUTCHISON JE, SOUZA AE, COOMBS GH.
The multiple cpb cysteine proteinase genes of Leishmania mexicana encode
isoenzymes that differ in their stage regulation and substrate preferences. J Biol Chem.
1997; 272: 14285-14293.
MOTTRAM JC, SOUZA AE, HUTCHISON JE, CARTER R, FRAME MJ, COOMBS GH. Evidence
from disruption of lmcpb gene array of Leishmania mexicana that cysteine proteinases
are virulence factors. Proc Natl Acad Sci USA. 1996; 93: 6008-6013.
Mougneau E, Altere F, Wakil AE, Zheng S, Coppola T, Wang ZE, Waldmann R,
Locksley RM, Glaichenhaus N. Expression cloning of a protective Leishmania antigen.
Science. 1995; 268: 563-566.
MUNDODI V, SOMANNA A, GEDAMU L. Genomic organization and functional expression of
diferentially regulated cysteine proteinase genes of Leishmania donovani complex.
Gene. 2002; 282: 257-265.
MURRAY HW, MASUR H, KEITHLY JS. Cell-mediated immune response in experimental
visceral leishmaniasis. II. Oxygen-dependent killing of intracellular Leishmania donovani
amastigotes. J immunol. 1982; 129: 351.
MUSSALEM JS, VASCONCELOS JRC, SQUAIELLA CC, ANANIAS RZ, BRAGA EG, RODRIGUES
MM, LONGO-MAUGÉRI IM. Adjuvant effect of the Propionibacterium acnes and its purified
soluble polysaccharide on the immunization with plasmidial DNA containing a
Trypanosoma cruzi gene. Microbiol Immunol. 2006; 50: 253-263.
OLIVEIRA TR. Expressão em bactérias de dois genes de cisteína proteinases de
amastigotas de Leishmania (L.) amazonensis: implicação das proteínas recombinantes
em respostas linfoproliferativas em modelo murino. [Tese de Mestrado] São Paulo:
Universidade Federal de São Paulo – Escola Paulista de Medicina; 2001.
OMARA-OPYENE AL, GEDAMU L. Molecular cloning, characterization and overexpression
of two distinct cysteine protease cDNAs from Leishmania donovani chagasi. Mol Biochem Parasitol. 1997; 90: 247-267.
72

Referências Bibliográficas
PASCALIS H, LAVERGNE A, BOURREAU E, PRÉVOT-LINGUET G, KARIMINIA A, PRADINAUD R,
RAFATI S, LAUNOIS P. Th1 cell development induced by cysteine proteinases A and B in
localized cutaneous leishmaniasis due to Leishmania guyanensis. Infect Immun. 2003;
71: 2924-2926.
PINELLI E, GONZALO RM, BOOG CJP, RUTTEN VPMG, GEBHARD D, DEL REAL G,
RUITENBERG EJ. Leishmania infantum-specific T cell lines derived from asymptomatic
dogs that lyse infected macrophages in a major histocompatibility. Eur J Immunol. 1995; 25: 1594-1600.
PINTO AR, BEYRODT CGP, LOPES RAML, BARBIÉRI CL. Identification of a 30 kDa antigen
from Leishmania (L.) chagasi amastigotes implicated in protective cellular responses in
a murine model. Intern J Parasitol. 2000; 30: 599-607.
PINTO EF, PINHEIRO RO, RAYOL A, LARRAGA V, ROSSI-BERGMANN B. Intranasal
vaccination against cutaneous leishmaniasis with a particulated leishmanial antigen or
DNA encoding LACK. Infect Immun. 2004; 72: 4521-4527.
PUPKINS ME, TETLEY L, COOMBS GH. Leishmania mexicana: amastigotes hydrolases in
unusual lysosomes. Exp Parasitol. 1986; 62: 29.
QI H, JI J, WANASEN N, SOONG L. Enhanced replication of Leishmania amazonensis
amastigotes in gamma interferon-stimulated murine macrophages: implications for the
pathogenesis of cutaneous leishmaniasis. Infect Immun. 2004; 72: 988-995.
RAFATI S, BABA AA, BAKHSHAYESH M, VAFA M. Vaccination of BALB/c mice with
Leishmania major amastigote-specific cysteine proteinase. Clin Exp Immunol. 2000;
120: 134-138.
RAFATI S, NAKHAEE A, TAHERI T, GHASHGHAII A, SALMANIAN AH, JIMENEZ M, MOHEBALI M,
MASINA S, FASEL N. Expression of cysteine proteinase type I and II of Leishmania
infantum and their recognition by sera during canine and human visceral leishmaniasis.
Exp Parasitol. 2003; 103: 143-151.
73

Referências Bibliográficas
RAFATI S, SALMANIAN AH, TAHERI T, VAFA M, FASEL N. A protective cocktail vaccine
against murine cutaneous leishmaniasis with DNA encoding cysteine proteinases of
Leishmania major. Vaccine 2001; 19: 3369-3375.
RAFATI, S., NAKHAEE, A., TAHERI, T., TASLIMI, Y., DARABI, H., ERAVANI, D., SANOS, S.,
KAYE, P., TAGHIKHANI, M., JAMSHIDI, S, AND RAD, M.A. Protective vaccination against
experimental canine visceral leishmaniasis using a combination of DNA and protein
immunization with cysteine proteinases type I and II of L. infantum. Vaccine 2005; 23:
3716-3725.
Rafati, S., Zahedifard, F., and Nazgouee, F. Prime-boost vaccination using cysteine
proteinases type I and II of Leishmania infantum confers protective immunity in murine
visceral leishmaniasis. Vaccine 2006; 24: 2169-2175.
SADICK MD, HEINZEL FP, HOLADAY BJ, PU RT, DAWKINS RS, LOCKSLEY RM. Cure of
murine leishmaniasis with anti-interleukin 4 monoclonal antibody. Evidence for a T cell-
dependent, interferon gamma-independent mechanism. J Exp Med. 1990; 171: 115-
127.
SAJID M, MCKERROW JH. Cysteine proteases of parasitic organisms. Mol Biochem Parasitol. 2002; 120: 1-21.
SAKANARI JA, NADLER SA, CHAN VJ, ENGEL JC, LEPTAK C, BOUVIER J. Leishmania major:
comparison of the cathepsin L- and B-like cysteine proteinase genes with those of other
trypanosomatids. Exp Parasitol. 1997; 85: 63-76.
SCOTT P, ARTIS D, UZONNA J, ZAPH C. The development of effector and memory T cells
in cutaneous leishmaniasis: the implications for vaccine development. Immunol Rev.
2004; 201: 318-338.
SCOTT P, CASPAR P, SHER A. protection against Leishmania major in BALB/c mice by
adoptive transfer of a T cell clone recognizing a low molecular weight antigen released
by promastigotes. J Immunol. 1990; 144: 1075-1079.
74

Referências Bibliográficas
SCOTT P, PEARCE E, NATOVITZ P, SHER A. Vaccination against cutaneous leishmaniasis
in a murine model. II. Immunologic properties of protective and nonprotective
subfractions of soluble promastigote extract. J Immunol. 1987; 39: 3118-3125.
SELZER PM, PINGEL S, HSIEH I, UGELE B, CHAN VJ, ENGEL JC, BOGYO M, RUSSEL DG,
SAKANARI JA, MCKERROW JH. Cysteine protease inhibitors as chemotherapy: lessons
from a parasite target. Proc Natl Acad Sci. 1999; 96: 11015-11022.
SETTE A, BUUS S, APPELLA E, SMITH JA, CHESNUT R, MILES C, COLON SM, GREY H.
Prediction of major histocompatibility complex binding regions of protein antigens by
sequence pattern analysis. Proc Natl Acad Sci USA. 1989; 86: 3296-3300.
SKEIKY YAW, COLER RN, BRANNON M, STROMBERG E, GREESON K, CRANE RT, CAMPOS-
NETO A, REED SG. Protective efficacy of a tandemly linked, multi-subunit recombinant
leishmanial vaccine (Leish-111f) formulated in MPL® adjuvant. Vaccine. 2002; 20:
3292-3303.
SOARES RMA, SANTOS ALS, BONALDO MC, ANDRADE AFB, ALVIANO CS, ANGLUSTER J,
GOLDENBERG S. Leishmania (Leishmania) amazonensis: differential expression of
proteinases and cell-surface polypeptides in avirulent and virulent promastigotes. Exp Parasitol. 2003; 104: 104-112.
SOMANNA A, MUNDODI V, GEDAMU L. Functional analysis of cathepsin B-like cysteine
proteases from Leishmania donovani complex. J Biol Chem. 2002; 277: 25305-25312.
SOONG L, CHANG CH, SUN J, LONGLEY JR BJ, RUDDLE NH, FLAVELL RA, MCMAHON-PRATT
D. Role of CD4+ T cells in pathogenesis associated with Leishmania amazonensis
infection. J Immunol. 1997; 158: 5374-5383.
SOONG L, DUBOISE SM, KIMA P, MCMAHON-PRATT D. Leishmania pifanoi amastigote
antigens protect mice against cutaneous leishmaniasis. Infect Immun. 1995; 63: 3559-
3566.
SPITZER N, JARDIM A, LIPPERT D, OLAFSON RW. Long-term protection of mice against
Leishmania major with a synthetic peptide vaccine. Vaccine. 1999; 17: 1298-1300.
75

Referências Bibliográficas
TOWBIN H, STAEHELIN T, BORDON J. Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979; 76: 4350-4354.
VASQUEZ RE, SOONG L. CXCL10/gamma interferon-inducible protein 10-mediated
protection against Leishmania amazonensis infection in mice. Infect Immun. 2006; 74:
6769-6777.
WALKER PS, SCHARTON-KERSTEN T, ROWTON ED, HENGGE U, BOULOC A, UDEY MC,
VOGEL JC. Genetic immunization with glycoprotein 63 cDNA results in a helper T cell
type 1 immune response and protection in a murine model of leishmaniasis. Hum Gene Ther. 1998; 9: 1899-1907.
WANG ZE, ZHENG S, CORRY DB, DALTON DK, SEDER RA, REINER SL, LOCKSLEY RM.
Interferon gamma-independent effects of interleukin 12 administered during acute or
established infection due to Leishmania major. Proc Natl Acad Sci USA. 1994; 91:
12932-12936.
WHITE AC, MCMAHON-PRATT D. Prophylactic immunization against experimental
Leishmania donovani infection by use of a purified protein vaccine. J Infect Dis. 1990;
161: 1313-1314.
WHO, WORLD HEALTH REPORT. 2001. Disponível em URL:
http://www.who.int/tdr/diseases/leish/diseaseinfo.htm.
WOLFRAM M, ILG T, MOTTRAM JC, OVERATH P. Antigen presentation by Leishmania
mexicana-infected macrophages: activation of helper T cells specific for amastigote
cysteine proteinases requires intracellular killing of the parasites. Eur J Immunol. 1995;
25: 1094-1100.
XU D, LIEW FY. Protection against leishmaniasis by injection of DNA encoding a major
surface glycoprotein, gp63, of Leishmania major. Immunology. 1995; 84: 173-176.
76

Referências Bibliográficas
77
YANG DM, FAIRWEATHER N, BUTTON LL, MCMASTER WR, KAHL LP, LIEW FY. Oral
Salmonella typhimurium (AroA-) vaccine expressing a major leishmanial surface protein
(gp63) preferentially induces T helper 1 cells and protective immunity against
leishmaniasis. J Immunol. 1990; 145: 2281-2285.
Top Related