







γλώσσες
Σελίδες
Νομικός


ION CHANNELS, RECEPTORS AND TRANSPORTERS
BK channel activation by tungstate requires the β1 subunitextracellular loop residues essential to modulate voltage sensorfunction and channel gating
Ana I. Fernández-Mariño & Miguel A. Valverde &
José M. Fernández-Fernández
Received: 23 July 2013 /Revised: 8 October 2013 /Accepted: 8 October 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Tungstate, a compound with antidiabetic,antiobesity, and antihypertensive properties, activates thelarge-conductance voltage- and Ca2+-dependent K+ (BK)channel containing either β1 or β4 subunits. The BK activationby tungstate is Mg2+-dependent and promotes arterialvasodilation, but only in precontracted mouse arteriesexpressing β1. In this study, we further explored how the β1
subunit participates in tungstate activation of BK channels.Activation of heterologously expressed human BKαβ1
channels in inside-out patches is fully dependent on the Mg2+
sensitivity of the BK α channel subunit even at high (10 μM)cytosolic Ca2+ concentration. Alanine mutagenesis of β1
extracellular residues Y74 or S104, which destabilize the activevoltage sensor, greatly decreased the tungstate-induced left-shift of the BKαβ1 G–V curves in either the absence orpresence of physiologically relevant cytosolic Ca2+ levels(10 μM). The weakened tungstate activation of theBKαβ1Y74A and BKαβ1S104A mutant channels was not relatedto decreased Mg2+ sensitivity. These results, together withpreviously published reports, support the idea that the putativebinding site for tungstate-mediated BK channel activation islocated in the pore-forming α channel subunit, around theMg2+ binding site. The role of β1 in tungstate-induced channelactivation seems to rely on its interaction with the BKα subunitto modulate channel activity. Loop residues that are essentialfor the regulation of voltage sensor activation and gating of theBK channel are also relevant for BK activation by tungstate.
Keywords BKchannel . Tungstate .KCNMB1(β1) subunit .
Extracellular loop
Introduction
The large conductance voltage- and Ca2+-activated K+ (BK)channel is widely expressed in mammalian tissues involved ina variety of physiological processes, including the regulationof smooth muscle tone [5]. Tetramers of the pore-forming αsubunit, encoded by a single gene (SLO (KCNMA1 )),constitute the core of the BK channel, which is activatedallosterically by membrane depolarization and binding ofintracellular Ca2+ and Mg2+. Thus, the energy contributionsprovided by voltage, Ca2+, and Mg2+ binding are additive ingating the channel, suggesting that these stimuli operatethrough independent pathways [13, 34]. In the tetramericBK channel, the voltage sensor domain (VSD) of each αsubunit is formed by segments S1 through S4 and the largecytoplasmic C terminus comprises two regulators of K+
conductance (RCK) domains that contribute to the formationof a gating ring at the intracellular membrane surface (Fig. 1a).Structural and functional evidences suggest that thephysiological regulation of BK channel by Ca2+ is achievedthrough two high-affinity Ca2+ binding sites located in bothRCK1 [44], and RCK2, involving a stretch of negativelycharged aspartates, termed the "Ca2+ bowl" [6, 46, 47]. Topromote channel activation, Mg2+ binds to a low-affinity,divalent cation binding site that consists of amino acid sidechains from both the VSD (residues D99 and N172) and theRCK1 domain (residues E374 and E399) of the BKα channelsubunit [45].
In some mammal tissues, BK channels also containmodulatory β subunits (β1–4) [39]. The β1 subunit isprimarily found in smooth muscle cells, where it increases
Electronic supplementary material The online version of this article(doi:10.1007/s00424-013-1379-9) contains supplementary material,which is available to authorized users.
A. I. Fernández-Mariño :M. A. Valverde :J. M. Fernández-Fernández (*)Laboratori de Fisiologia Molecular i Canalopaties, Departament deCiències Experimentals i de la Salut, Universitat Pompeu Fabra,C/ Dr. Aiguader 88, Barcelona 08003, Spaine-mail: [email protected]
Pflugers Arch - Eur J PhysiolDOI 10.1007/s00424-013-1379-9
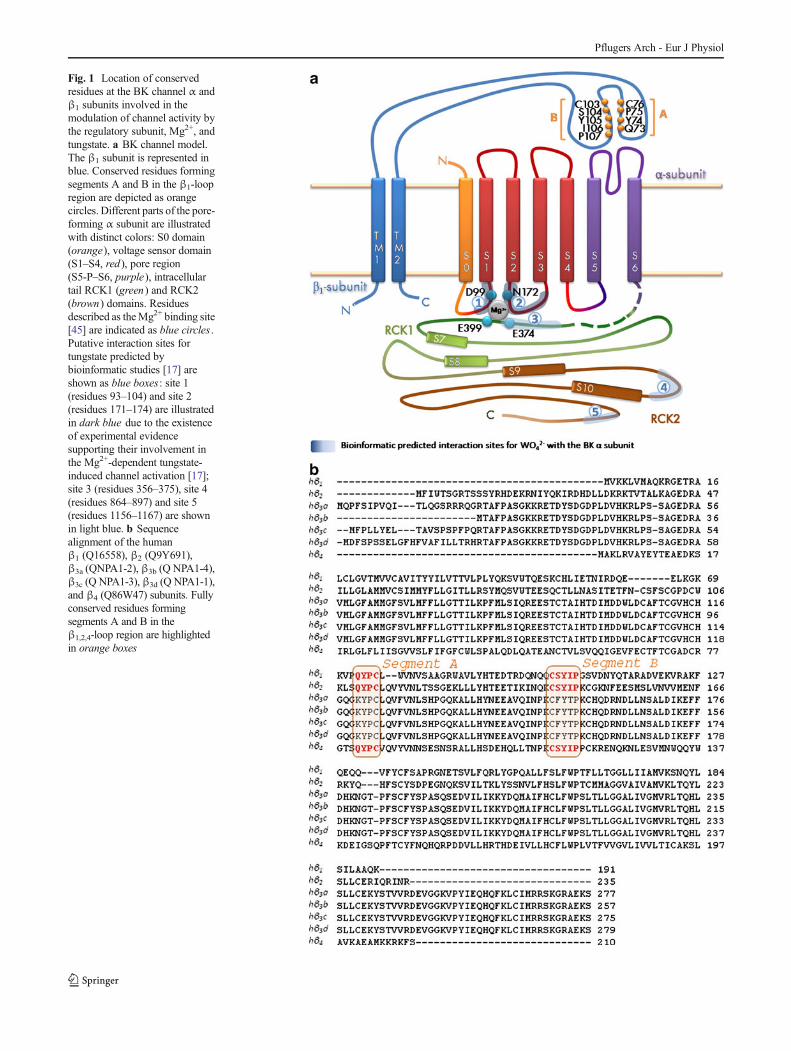
Fig. 1 Location of conservedresidues at the BK channel α andβ1 subunits involved in themodulation of channel activity bythe regulatory subunit, Mg2+, andtungstate. a BK channel model.The β1 subunit is represented inblue. Conserved residues formingsegments A and B in the β1-loopregion are depicted as orangecircles. Different parts of the pore-forming α subunit are illustratedwith distinct colors: S0 domain(orange), voltage sensor domain(S1–S4, red), pore region(S5-P–S6, purple), intracellulartail RCK1 (green) and RCK2(brown) domains. Residuesdescribed as theMg2+ binding site[45] are indicated as blue circles .Putative interaction sites fortungstate predicted bybioinformatic studies [17] areshown as blue boxes: site 1(residues 93–104) and site 2(residues 171–174) are illustratedin dark blue due to the existenceof experimental evidencesupporting their involvement inthe Mg2+-dependent tungstate-induced channel activation [17];site 3 (residues 356–375), site 4(residues 864–897) and site 5(residues 1156–1167) are shownin light blue. b Sequencealignment of the humanβ1 (Q16558), β2 (Q9Y691),β3a (QNPA1-2), β3b (Q NPA1-4),β3c (Q NPA1-3), β3d (Q NPA1-1),and β4 (Q86W47) subunits. Fullyconserved residues formingsegments A and B in theβ1,2,4-loop region are highlightedin orange boxes
Pflugers Arch - Eur J Physiol

the apparent voltage and Ca2+ sensitivity of the BK channel αsubunit by stabilizing the voltage sensor in the activeconfiguration [3, 11, 12]. Mutagenesis studies have shownthat the large extracellular loop connecting the two putativeTM segments of the β1 subunit is, at least in part, responsiblefor its regulatory action on the voltage sensor activation andgating of the BK channel. We and others have reported thatamino acid substitutions in this loop either tune up (E65K)[16] or down (Y74A, S104A, Y105A and I106A) [18] thepositive β1-induced modulation of BK channel activity.
In smooth muscle, BK channels are activated by local andtransient Ca2+ increases ("Ca2+ sparks") caused by Ca2+
release from intracellular stores. The efflux of K+ throughactivated BK channels is sufficient to hyperpolarize themembrane potential, limiting membrane depolarization, Ca2+
entry via voltage-operated Ca2+ channels, and smooth musclecontraction [20]. Due to its action on BK channel function, thepresence of the regulatory β1 subunit makes this negativefeedback mechanism more efficient [7]. Accordingly,expression of the β1 subunit is relevant to smooth musclecontraction in airways [31] and the regulation of arterial toneand blood pressure in both animal hypertension models and inhumans [1, 2, 7, 16, 30, 33, 37]. For this reason, the BKαβ1
channel has been considered an attractive target for thedevelopment of therapies for smooth muscle diseases suchas asthma [31] and arterial hypertension [16, 22, 33, 38].
Tungstate, a compound with antidiabetic [4] andantiobesity properties [10], also reduces blood pressure inexperimental animal models of both hypertension [35, 36]and metabolic syndrome [27]. This antihypertensive effectof tungstate has been associated with its action on both theendothelial xanthine oxidase [35] and the BK channel at thevascular smooth muscle cells [17]. We recently reported thattungstate activates BK channels containing either theβ1 or theβ4 (but not β2 or β3) subunits. Furthermore, tungstateactivation of the vascularβ1-containing BK channel promotesvasodilation of pre-contracted blood vessels and requires theMg2+ sensitivity of the channel [17]. Based on these resultsand the analysis of available structural data on tungstate–protein complexes, we proposed a putative tungstate bindingsite located at the human BK channel α subunit that mightinvolve residues around those of the VSD and the RCK1domain composing the Mg2+ binding site [17, 45]. Thepresent study further explores the role of the β1 subunit inthe activation of BK channels by tungstate.
Materials and methods
cDNA constructs and cell transfection
Human β1 subunit (KCNMB1) of the BK channel cloned intopcDNA3was a gift fromLigia Toro (University of California–
Los Angeles, USA). Human α (hSlo) (KCNMA1) of the BKchannel cloned into pcDNA3was supplied byRamón Latorre,(Centro de Neurociencias, Valparaíso, Chile). D99A mutationin the human BK channel α (hSlo) subunit was introducedusing the QuikChangeR IIXL Site-Directed Mutagenesis Kit(Stratagene, La Jolla, CA, USA), following manufacturer'sinstructions. Primers for the hSloD99A mutant were 5′-CCTCCGTGAAGGCCTGGGCGGGGGTGATGATATC-3′( forward) and 5 ′ -GATATCATCACCCCCGCCCAGGCCTTCACGGAGG-3′ (reverse). Q73A, Y74A, andS104A mutations in the human BK channel β1 subunit wereintroduced by site-directed mutagenesis (GenScriptCorporation, Piscatway, NJ, USA). All cDNA clones used inthis study were sequenced in full to confirm their integrity.
HEK293 cells were transfected using polyethylenimine(PEI) ExGen 500 (Fermentas Inc., Hanover, MD, USA),following the manufacturer's instructions (7 PEI equivalentsper 3.3 μg of cDNAs expressing the wild-type [WT] or D99ABK channel human α subunit with either the WTor one of themutant [Q73A, Y74A, S104A] human β1 subunits [1:2 ratio],together with the transfection reporter pEGFPN1).
Electrophysiology
Inside-out ionic currents were recorded in macropatches fromEGFP-positive cells, 2–3 days after transfection [17].Borosilicate glass patch pipettes had 2–3 MΩ resistance andwere filled with a solution containing (in mM): 140 KCl, 1.2MgCl2, 0.15 CaCl2, 5 EGTA, 10 HEPES (300 mOsm/l,pH 7.35). In experiments performed at 0 Mg2+, the pipettesolution contained (in mM): 140 KCl, 0.15 CaCl2, 5 EGTA,10 HEPES (300 mOsm/l, pH 7.25). The 0 Ca2+ solution(nominal 0 Ca2+) bathing the cytoplasmic face of the patchmembrane contained (in mM): 140 KCl, 5 EGTA, 10 HEPES(0 Mg2+ condition). When necessary, the cytosolic levels ofMg2+ were increased by adding 0.7, 5, 30, or 100 mMMg2+ tothe 0 Ca2+ cytosolic solution, as appropriate. All these bathsolutions were adjusted to 300 mOsm/l and pH 7.25.When needed, the intracellular free Ca2+ concentration(calculated using EqCal; Biosoft, Cambridge, UnitedKingdom) was adjusted to 10 μM by adding the appropriatecombination of CaCl2 and EGTA to the 0.7 mM Mg2+ bathsolution.
For current-activation studies at the 0 Ca2+ condition,membrane macropatches were clamped at 0 mV, pulsed for150ms from −100 to +200mVin 10-mV steps, and repolarizedto −80 mV for 20 ms. When using 10 μM cytosolic Ca2+, themacropatches were also clamped at 0 mV, but pulsed for150 ms from −180 to +140 mV (or +200 mV) in 10-mV stepsand repolarized to −100 mV for 50 ms. Experiments wereperformed at room temperature (22–26 °C).
Relative conductance was determined by measuring tailcurrent amplitudes. For each patch, the conductance–voltage
Pflugers Arch - Eur J Physiol

(G–V) relationship was fitted with the following Boltzmannequation:
G=Gmax ¼ 1= 1þ exp�− V−V 1=2 act
� �=kact
� ��;
where G is the value of the instantaneous tail current at eachtest voltage, Gmax is the maximum obtained tail current, V isthe test voltage applied to the membrane, V1/2 act is the voltagefor half-maximal current activation (which is a convenientparameter to study the effect of BK channels modulatorsbecause it is directly related to the energy required to openthe channel), and kact (an index of the minimum number ofelementary charges that move through the electric field to gatethe channel) is the slope factor of the Boltzmann term. G–Vcurves are represented as averages from a set of patches at theindicated experimental condition.
Recordings were obtained with a D-1600 Darmstadtamplifier (List Medical, Germany). Currents were acquiredat 10 kHz and low-pass-filtered at 1 kHz. The pClamp8software (Axon Instruments, Foster City, CA, USA) was usedfor pulse generation, data acquisition and subsequent analysis.
Statistics
Data are presented as the means ± SEM. Statistical testsincluded Student paired t test, Mann–Whitney test, or one-way analysis of variance (ANOVA) followed by a Dunnettpost-hoc test, as appropriate. Differences were consideredsignificant if P <0.05.
Results
Tungstate-induced activation of the BKαβ1 channelat physiological cytosolic Ca2+ levels dependson BK α Mg2+ sensitivity
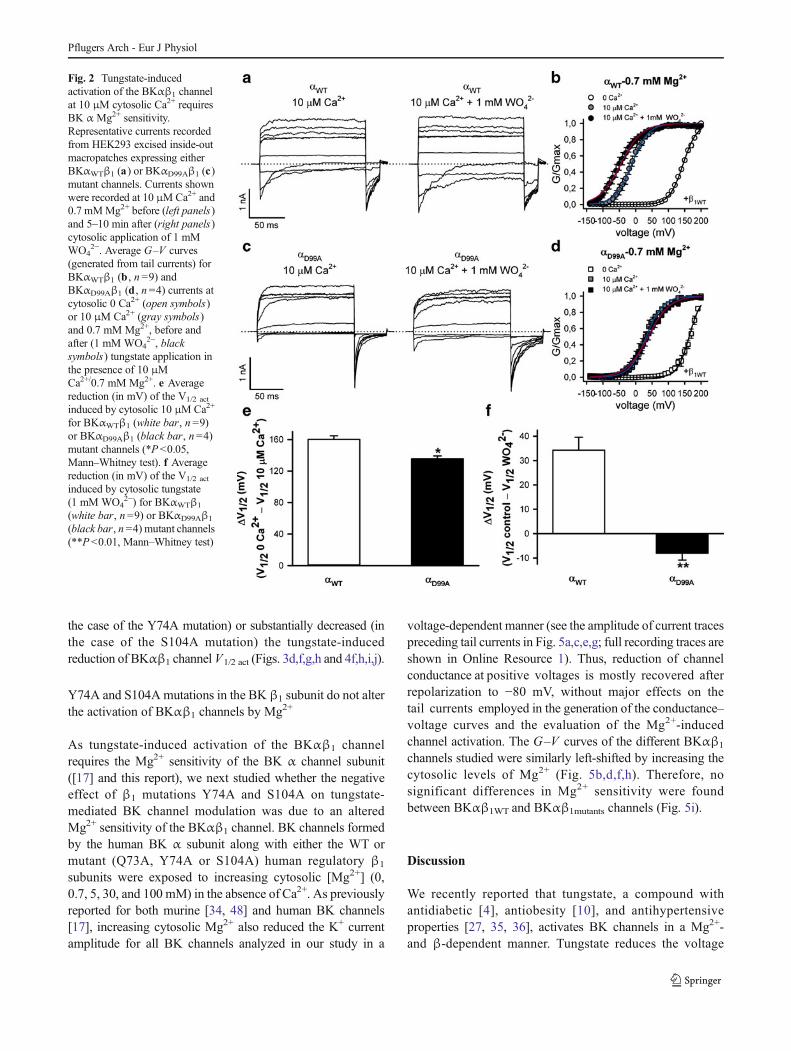
We first reassessed the effect of tungstate on BKαβ1 V1/2 act athigh (10 μM) cytosolic Ca2+, which is within the range ofCa2+ levels delivered by "Ca2+ sparks" during smooth musclecontraction [28]. It is precisely at these intracellular Ca2+
concentrations that the modulatory effect of the β1 subuniton BK channel function acquires physiological relevance [25,28]. As previously reported [17], the tungstate-induceddecrease of the V1/2 act for heterologously expressed humanBKαβ1 channels is significantly higher in the presence of10 μM cytosolic Ca2+ (34 mV) (Fig. 2a,b,f) than in theabsence of cytosolic Ca2+ (20 mV; P <0.01, Mann–Whitneytest) (Fig. 4a,b,i,j). We previously reported that the Mg2+
sensitivity of the BKαβ1 channel is required for thetungstate-induced left-shift of BKαβ1 V1/2 act in the absenceof cytosolic Ca2+ [17]. In order to test whether cytosolic Mg2+
is also required for the larger tungstate-induced activation of
BKαβ1 channels at cytosolic 10 μM Ca2+, we tested thetungstate effect on human BKαβ1 channels containing analanine substitution in the highly conserved D99 residue ofthe α subunit VSD contributing to Mg2+ coordination(Fig. 1a). D99A mutation has been reported to abolishMg2+-induced activation of mSlo [45] and human BKαβ1
[17] channels and to prevent the activation of BKαβ1
channels by tungstate in the absence of cytosolic Ca2+ [17].We found that at 10 μM cytosolic Ca2+ tungstate also failed toreduce the V1/2 act for heterologously expressed BKαD99Aβ1
mutant channels (Fig. 2c,d,f).
β1 Extracellular loop residues modulating voltage sensorfunction and channel gating affect activation of BKαβ1
channels by tungstate
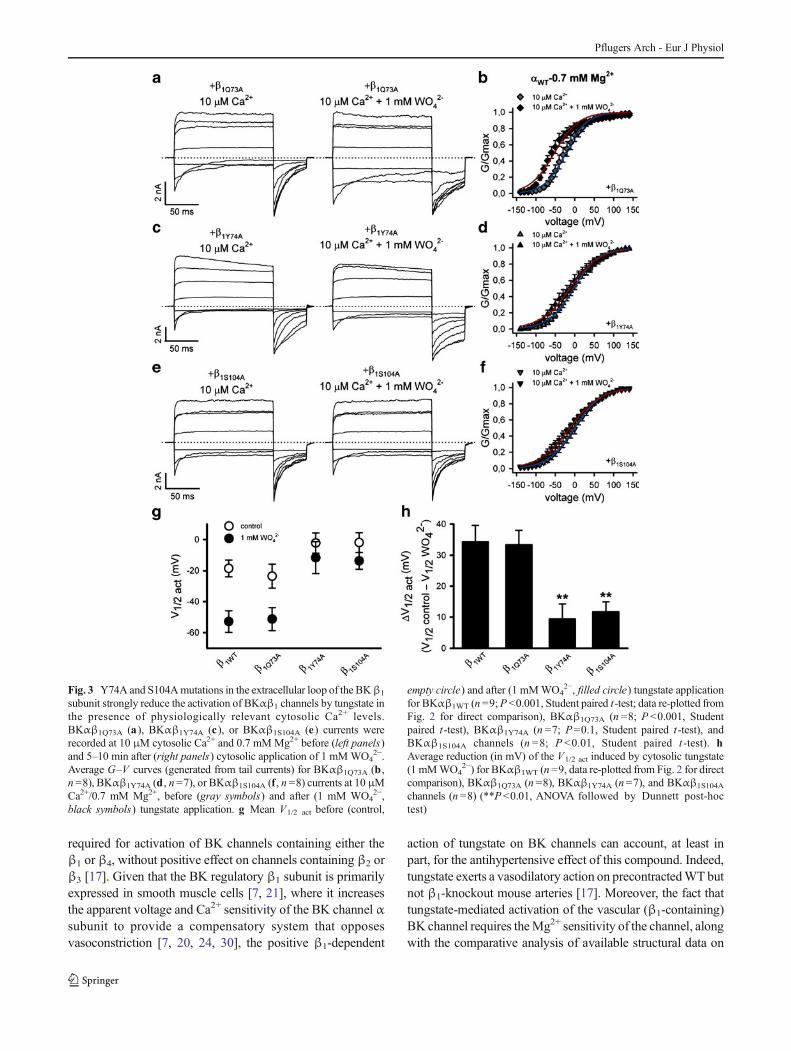
Alanine-scanning mutagenesis studies have shown thatresidues located in two segments (named A and B) of thelarge extracellular loop connecting the two TM segments ofthe β1 subunit (Fig. 1) are involved in the positive regulatoryaction on the voltage sensor activation of the BK channel.Thus, mutation Y74A at segment A (which is formed by thefour residues 73QYPC76) and mutations S104A, Y105A andI106A at segment B (delimited by the five residues103CSYIP107) impair the β1-induced stabilization of theopen-channel voltage sensor activation, either for Ca2+-bound(in the presence of physiological cytosolic Ca2+ levels in theμM range) or Ca2+-unbound BK channels [18] (for location ofthe abovementioned residues in the BK β1-extracellular loop,see Fig. 1). As tungstate promotes BK channel activation in aβ-dependent manner (it only activates BK channelscontaining either the β1 or the β4 subunits) [17], we evaluatedwhether residues Y74 and S104, respectively located atsegment A and B of the β1-extracellular loop, also play a rolein the tungstate-mediated BK channel activation. For thatpurpose, we mutated Y74 and S104 of the human BK channelβ1 subunit to alanine residues. The effects of these mutationson the tungstate-mediated activation of heterologouslyexpressed BKαβ1 channels were analyzed in the presence of10 μM cytosolic Ca2+ (Fig. 3) and in the absence of cytosolicCa2+ (nominal 0 Ca2+) (Fig. 4). The Q73A mutation (locatedat segment A of the β1 extracellular loop; Fig. 1a) was alsointroduced as an additional control that does not impair theβ1-induced positive modulation of BK channel activity [18].The functional analysis of these mutations was carried out inthe presence of cytosolic 0.7 mM Mg2+ (given the Mg2+
dependence of the tungstate-induced BK channel activation).At both experimental cytosolic Ca2+ concentrations (0 and10 μM Ca2+), tungstate left-shifted the G–V curves ofBKαβ1WT (Figs. 2b and 4b) and BKαβ1Q73A channels(Figs. 3b and 4d), inducing a significant decrease of thechannel V1/2 act (Figs. 3g,h and 4i,j). However, the presenceof mutations Y74A or S104A in the β1 subunit abolished (in
Pflugers Arch - Eur J Physiol
the case of the Y74A mutation) or substantially decreased (inthe case of the S104A mutation) the tungstate-inducedreduction of BKαβ1 channel V1/2 act (Figs. 3d,f,g,h and 4f,h,i,j).
Y74A and S104Amutations in the BK β1 subunit do not alterthe activation of BKαβ1 channels by Mg2+
As tungstate-induced activation of the BKαβ1 channelrequires the Mg2+ sensitivity of the BK α channel subunit([17] and this report), we next studied whether the negativeeffect of β1 mutations Y74A and S104A on tungstate-mediated BK channel modulation was due to an alteredMg2+ sensitivity of the BKαβ1 channel. BK channels formedby the human BK α subunit along with either the WT ormutant (Q73A, Y74A or S104A) human regulatory β1
subunits were exposed to increasing cytosolic [Mg2+] (0,0.7, 5, 30, and 100 mM) in the absence of Ca2+. As previouslyreported for both murine [34, 48] and human BK channels[17], increasing cytosolic Mg2+ also reduced the K+ currentamplitude for all BK channels analyzed in our study in a
voltage-dependent manner (see the amplitude of current tracespreceding tail currents in Fig. 5a,c,e,g; full recording traces areshown in Online Resource 1). Thus, reduction of channelconductance at positive voltages is mostly recovered afterrepolarization to −80 mV, without major effects on thetail currents employed in the generation of the conductance–voltage curves and the evaluation of the Mg2+-inducedchannel activation. The G–V curves of the different BKαβ1
channels studied were similarly left-shifted by increasing thecytosolic levels of Mg2+ (Fig. 5b,d,f,h). Therefore, nosignificant differences in Mg2+ sensitivity were foundbetween BKαβ1WT and BKαβ1mutants channels (Fig. 5i).
Discussion
We recently reported that tungstate, a compound withantidiabetic [4], antiobesity [10], and antihypertensiveproperties [27, 35, 36], activates BK channels in a Mg2+-and β-dependent manner. Tungstate reduces the voltage
Fig. 2 Tungstate-inducedactivation of the BKαβ1 channelat 10 μM cytosolic Ca2+ requiresBK α Mg2+ sensitivity.Representative currents recordedfrom HEK293 excised inside-outmacropatches expressing eitherBKαWTβ1 (a) or BKαD99Aβ1 (c)mutant channels. Currents shownwere recorded at 10 μMCa2+ and0.7 mMMg2+ before (left panels)and 5–10 min after (right panels)cytosolic application of 1 mMWO4
2−. Average G–V curves(generated from tail currents) forBKαWTβ1 (b , n =9) andBKαD99Aβ1 (d , n =4) currents atcytosolic 0 Ca2+ (open symbols)or 10 μM Ca2+ (gray symbols)and 0.7 mM Mg2+, before andafter (1 mM WO4
2−, blacksymbols) tungstate application inthe presence of 10 μMCa2+/0.7 mM Mg2+. e Averagereduction (in mV) of the V1/2 act
induced by cytosolic 10 μM Ca2+
for BKαWTβ1 (white bar, n =9)or BKαD99Aβ1 (black bar, n =4)mutant channels (*P<0.05,Mann–Whitney test). f Averagereduction (in mV) of the V1/2 act
induced by cytosolic tungstate(1 mM WO4
2−) for BKαWTβ1
(white bar, n =9) or BKαD99Aβ1
(black bar, n=4) mutant channels(**P <0.01, Mann–Whitney test)
Pflugers Arch - Eur J Physiol
required for activation of BK channels containing either theβ1 or β4, without positive effect on channels containing β2 orβ3 [17]. Given that the BK regulatory β1 subunit is primarilyexpressed in smooth muscle cells [7, 21], where it increasesthe apparent voltage and Ca2+ sensitivity of the BK channel αsubunit to provide a compensatory system that opposesvasoconstriction [7, 20, 24, 30], the positive β1-dependent
action of tungstate on BK channels can account, at least inpart, for the antihypertensive effect of this compound. Indeed,tungstate exerts a vasodilatory action on precontractedWT butnot β1-knockout mouse arteries [17]. Moreover, the fact thattungstate-mediated activation of the vascular (β1-containing)BK channel requires theMg2+ sensitivity of the channel, alongwith the comparative analysis of available structural data on
Fig. 3 Y74A and S104Amutations in the extracellular loop of the BKβ1
subunit strongly reduce the activation of BKαβ1 channels by tungstate inthe presence of physiologically relevant cytosolic Ca2+ levels.BKαβ1Q73A (a), BKαβ1Y74A (c), or BKαβ1S104A (e) currents wererecorded at 10 μM cytosolic Ca2+ and 0.7 mMMg2+ before (left panels)and 5–10 min after (right panels) cytosolic application of 1 mMWO4
2−.Average G–V curves (generated from tail currents) for BKαβ1Q73A (b ,n =8), BKαβ1Y74A (d , n =7), or BKαβ1S104A (f , n =8) currents at 10 μMCa2+/0.7 mM Mg2+, before (gray symbols) and after (1 mM WO4
2−,black symbols) tungstate application. g Mean V1/2 act before (control,
empty circle) and after (1 mMWO42−, filled circle) tungstate application
for BKαβ1WT (n =9;P <0.001, Student paired t-test; data re-plotted fromFig. 2 for direct comparison), BKαβ1Q73A (n =8; P <0.001, Studentpaired t-test), BKαβ1Y74A (n =7; P=0.1, Student paired t-test), andBKαβ1S104A channels (n =8; P <0.01, Student paired t -test). hAverage reduction (in mV) of the V1/2 act induced by cytosolic tungstate(1 mMWO4
2−) for BKαβ1WT (n =9, data re-plotted from Fig. 2 for directcomparison), BKαβ1Q73A (n =8), BKαβ1Y74A (n=7), and BKαβ1S104A
channels (n =8) (**P<0.01, ANOVA followed by Dunnett post-hoctest)
Pflugers Arch - Eur J Physiol
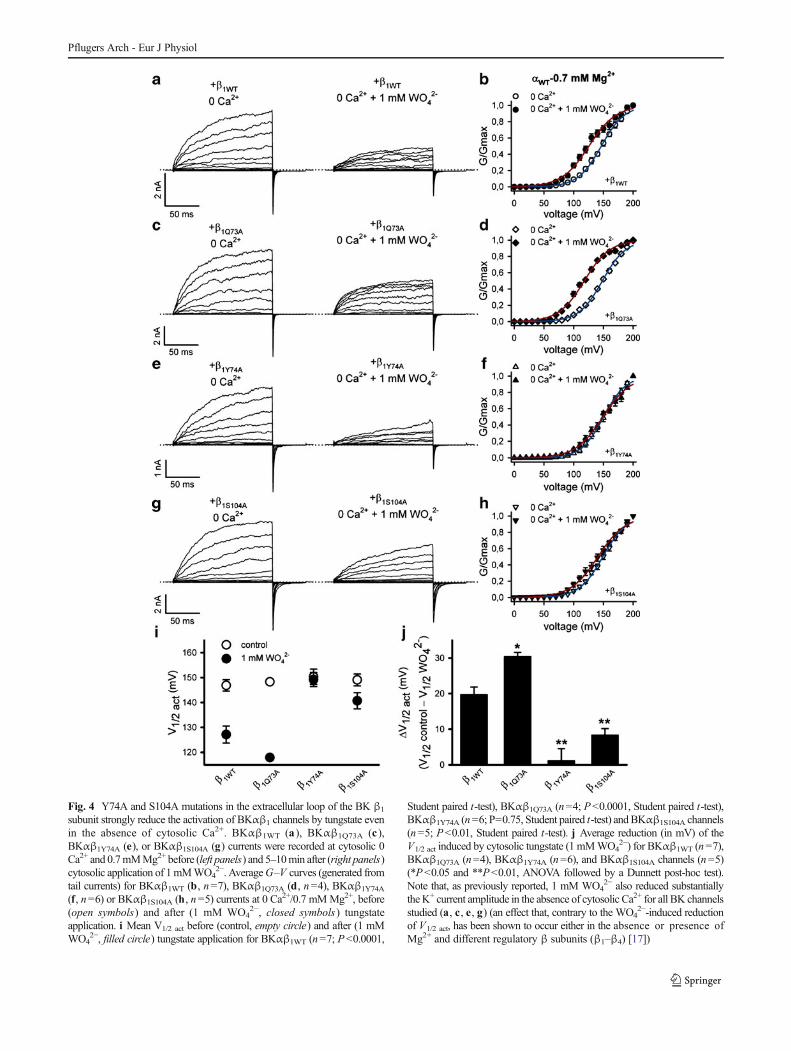
Fig. 4 Y74A and S104A mutations in the extracellular loop of the BK β1
subunit strongly reduce the activation of BKαβ1 channels by tungstate evenin the absence of cytosolic Ca2+. BKαβ1WT (a ), BKαβ1Q73A (c ),BKαβ1Y74A (e), or BKαβ1S104A (g) currents were recorded at cytosolic 0Ca2+ and 0.7mMMg2+ before (left panels) and 5–10min after (right panels)cytosolic application of 1mMWO4
2−. AverageG–V curves (generated fromtail currents) for BKαβ1WT (b, n=7), BKαβ1Q73A (d, n=4), BKαβ1Y74A
(f, n=6) or BKαβ1S104A (h, n=5) currents at 0 Ca2+/0.7 mMMg2+, before(open symbols) and after (1 mM WO4
2−, closed symbols) tungstateapplication. i Mean V1/2 act before (control, empty circle) and after (1 mMWO4
2−, filled circle) tungstate application for BKαβ1WT (n=7; P<0.0001,
Student paired t-test), BKαβ1Q73A (n=4; P<0.0001, Student paired t-test),BKαβ1Y74A (n=6; P=0.75, Student paired t-test) andBKαβ1S104A channels(n=5; P<0.01, Student paired t-test). j Average reduction (in mV) of theV1/2 act induced by cytosolic tungstate (1 mMWO4
2−) for BKαβ1WT (n=7),BKαβ1Q73A (n=4), BKαβ1Y74A (n=6), and BKαβ1S104A channels (n=5)(*P<0.05 and **P<0.01, ANOVA followed by a Dunnett post-hoc test).Note that, as previously reported, 1 mM WO4
2− also reduced substantiallytheK+ current amplitude in the absence of cytosolic Ca2+ for all BK channelsstudied (a, c, e, g) (an effect that, contrary to the WO4
2−-induced reductionof V1/2 act, has been shown to occur either in the absence or presence ofMg2+ and different regulatory β subunits (β1–β4) [17])
Pflugers Arch - Eur J Physiol
tungstate–protein complexes, allowed us to suggest that theputative tungstate binding site may consist of aspartate–lysine/lysine–aspartate (DK/KD) motifs placed at the BK channel αsubunit, close to amino acidic residues of the VSD and theRCK1 domain delimiting the binding site for Mg2+ [17]. Herewe report that residues in segments A and B of the largeextracellular loop, which promote β1-dependent stabilizationof the BK channel voltage sensor in its active state, also play akey role in channel activation by tungstate.
It has been shown that single alanine substitutions ofseveral residues placed either at segment A (Y74) or segmentB (S104A, Y105A, I106E) of the β1 extracellular loop reducethe left-shift of the BK G–V curves produced by theregulatory β1 subunit at high (60 μM) cytosolic Ca2+ in anonadditive way. In addition, at least Y74A, Y105A, or I106Amutations destabilize open-channel voltage sensor activationeven in the absence of Ca2+ binding to the BK channel [18].We have now observed that β1 mutations Y74A or S104A
Pflugers Arch - Eur J Physiol

abolished or substantially decreased, respectively, the left-shift of the BKαβ1 G–V curves induced by tungstate,therefore severely impairing channel activation by thiscompound. This effect was observed both in the presence of10 μM cytosolic Ca2+ (which is within the physiologicalwindow of Ca2+ levels that are transiently and locally reached("Ca2+ sparks") in the cytosol of smooth muscle cells duringcontraction) or in the absence of cytosolic Ca2+ (nominal 0Ca2+). In contrast, mutation Q73A in the β1 subunit, whichdoes not alter the left shift of BKG–V curves at high (60 μM)cytosolic Ca2+ [18], did not prevent the tungstate-inducedactivation of BKαβ1 channels. We have also observed thatthe activation of BKαβ1 channels by tungstate in the presenceof 10 μM cytosolic Ca2+ remains fully dependent on the Mg2+
sensitivity of the BK α channel subunit, as the activation isprevented by the D99Amutation in its VSD that abrogates BKchannel activation by Mg2+. These results are similar to thosewe obtained in the absence of cytosolic Ca2+ [17].Furthermore, the impaired activation by tungstate ofBKαβ1Y74A and BKαβ1S104A mutant channels is not due toany decrease in their Mg2+ sensitivity, which remains similarto that of the BKαβ1WT channel. Altogether, our findingssupport the idea that the putative binding site for tungstate
that mediates BK channel activation is located at the BK αchannel subunit, around those residues of the VSD and theRCK1 domain that coordinate the binding of Mg2+. Still,residues in segments A and B of the β1 extracellular loopare also required for a proper tungstate-induced activation ofBK channels. The role of these residues can be explained byan indirect action related to their involvement in maintaining aparticular three-dimensional structure of the BK α and/or theBKβ1 channel subunits that allows the binding of tungstate tothe α VSD and RCK1 regions and the subsequent channelactivation. An alternative explanation suggested by severalauthors [9, 18, 19, 29] is direct participation of these β1 aminoacids in the modulation of BK channel activity due to theirclose proximity to the external mouth, and possibly theselectivity filter and gate, of the channel. In this sense, it isknown that residues placed either at segment A (Y74) or B(S104A, Y105A, I106E) of the β1 extracellular loopmodulatenot only the activation of the voltage sensor but also theintrinsic gating of the BK channel, pointing to a possible roleof these particular β1 regions in the coupling of the activatedvoltage sensor and the channel gate [18]. By affecting thisprocess, mutations in segments A and B of theβ1 extracellularloop might, in turn, impair the intramolecular transductionmechanism by which tungstate interaction with the BK αsubunit activates the channel.
The relevance of the β1 extracellular loop in themodulation of BK channel activity has been reported bymultiple authors. On one hand, it has been suggested thatsteroids activate BK channels through direct binding to anexternal site available in the presence of the regulatory β1
subunit, although the exact site of interaction remains to beelucidated. Thus, 17β-estradiol (in the μM range) seems toincrease the activity of BKαβ1 channels by direct interactionwith the β extracellular loop, as channel activation is alsotriggered by estradiol conjugated to a membrane-impenetrablecarrier protein [40]. Similar results have been obtained usingthe xenoestrogen tamoxifen [15]. On the other hand, humanβ1 polymorphisms placed at the extracellular loop have bothfunctional and clinical relevance. The β1 E65K variant hasbeen reported to increase the voltage and Ca2+ sensitivity ofthe BK channel [16] and the R140W variant to decreasechannel openings [31]. Consistent with such gain (inducedby E65K) and loss (produced by R140W) of channel function,along with the key role of the BK channel in the control ofsmooth muscle tone both in the vasculature [20] and in theairways [32], E65K has been found to protect againstmoderate-to-severe diastolic hypertension and "combinedcardiovascular disease" (myocardial infarction and stroke)[16, 33] and R140W has been linked to reduced pulmonaryfunction [31].
Although not all regulatory β subunits change thebiophysical properties of the BK channel in a similar way(reviewed in [39]), gating current measurements show that the
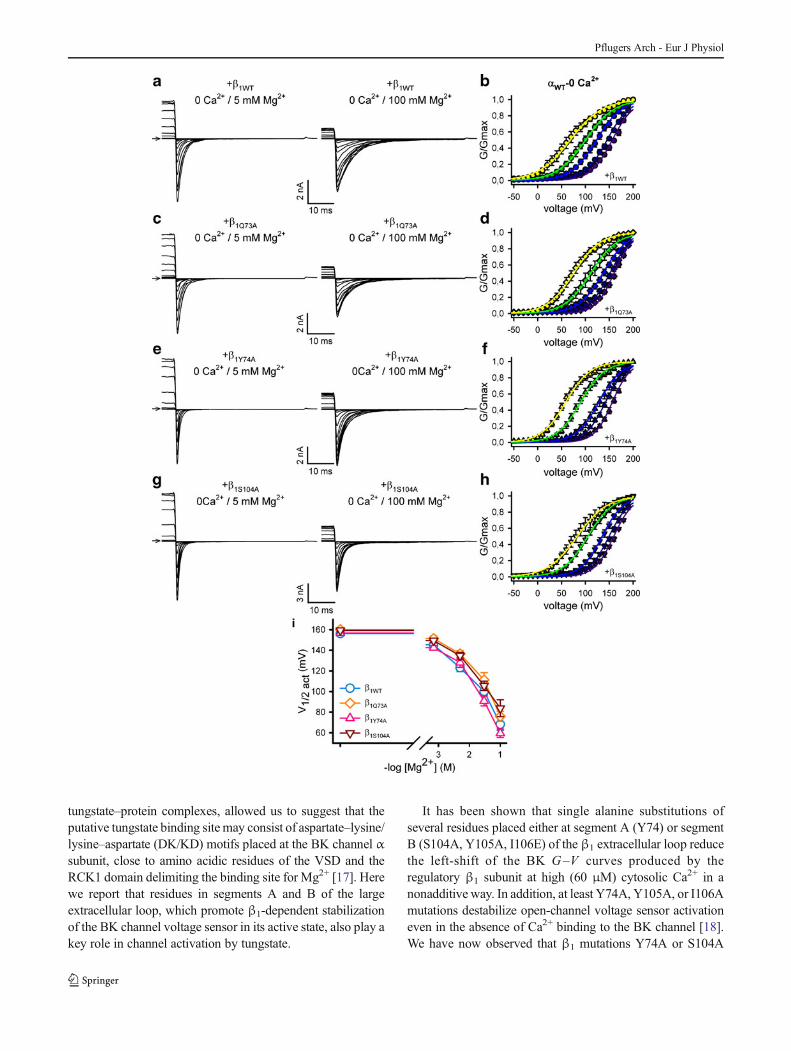
�Fig. 5 Y74A and S104A mutations in the extracellular loop of the BKβ1 subunit have no significant effect on the activation of BKαβ1 channelsby Mg2+. In order to focus on BK channel activation induced by Mg2+,only a magnified view of representative tail currents recorded fromHEK293 excised inside-out macropatches expressing BKαβ1WT (a),BKαβ1Q73A (c ), BKαβ1Y74A (e ) or BKαβ1S104A (g ) channels isillustrated (full recording traces are shown in Online Resource 1).Currents shown were recorded from each patch at cytosolic 0 Ca2+ and5 mM Mg2+ (left panels) or 0 Ca2+ and 100 mM Mg2+ (right panels).Average G–V curves (generated from tail currents) for BKαβ1WT
(b , n =7), BKαβ1Q73A (d , n =6), BKαβ1Y74A (f , n =5) orBKαβ1S104A (h , n =8) currents in the presence of increasingconcentrations of cytosolic Mg2+ (0 Mg2+, purple symbols ;0.7 mM Mg2+, dark blue symbols ; 5 mM Mg2+, blue symbols ;30 mM Mg2+, green symbols ; and 100 mM Mg2+, yellowsymbols ). i Mean V1/2 act for BKαβ1WT (empty circle , n =7),BKαβ1Q73A (empty diamond , n =6), BKαβ1Y74A (empty uprighttriangle , n =5) and BKαβ1S104A (empty inverted triangle , n =8)channels, at different cytosolic Mg2+ levels. For each BKαβ1 channel,differences in V1/2 act between the different Mg2+ conditions were testedby one-way analysis of variance (ANOVA) followed by a Bonferronipost-hoc test (0.7 mM Mg2+ vs. 0 Mg2+: BKαβ1WT channels, P>0.05;BKαβ1Q73A channels, P >0.05; BKαβ1Y74A channels, P >0.05; andBKαβ1S104A channels, P >0.05/5 mM Mg2+ vs. 0.7 mM Mg2+:BKαβ1WT channels, P <0.001; BKαβ1Q73A channels, P <0.05;BKαβ1Y74A channels, P <0.05; and BKαβ1S104A channels, P <0.05/30 mM Mg2+ vs. 5 mM Mg2+: BKαβ1WT channels, P <0.01;BKαβ1Q73A channels, P <0.001; BKαβ1Y74A channels, P<0.001; andBKαβ1S104A channels, P <0.001/100 mM Mg2+ vs. 30 mM Mg2+:BKαβ1WT channels, P <0.001; BKαβ1Q73A channels, P <0.001;BKαβ1Y74A channels, P <0.001; and BKαβ1S104A channels, P<0.01).One-way ANOVA followed by a Dunnett post-hoc test found nosignificant differences between BKαβ1WT and BKαβ1mutants channelsfor the V1/2 act at each cytosolic Mg2+ level tested
Pflugers Arch - Eur J Physiol

stabilization of the active configuration of the voltage sensorinduced by β1 is a trait shared with β2 and β4, but not β3,subunits [11]. Thus, both β2 and β4 subunits induce areduction in the standard free energy difference between theactive and resting states of the voltage sensor, albeit to a lesserextent than that produced by the β1 subunit [11]. Furthermore,segments A and B of the β1 extracellular loop, containing keyresidues for the modulation of voltage sensor function andgating of the BK channel, are the longest sequences ofconsecutive residues that are identical in both β2 and β4
subunits [18]. Therefore, this high degree of conservationamong β1, β2 and β4 subunits of segments A and B at theextracellular loop may underlie both the stabilization of theactive voltage sensor induced by these three regulatory βsubunits and the β-dependent effect of tungstate on BKchannels. Regarding the latter, tungstate promotes theactivation not only of BKαβ1 but also of BKαβ4 channels.The fact that we did not observe a positive effect of tungstateon BKαβ2 channels activity might be due to the specificmodulatory actions that the β2 subunit (but not β1 or β4)has on BK channels. Indeed, β2 induces fast and completeinactivation of the BK channel mediated by the interaction ofits N terminus with a receptor site in the open α subunit[41–43]. In addition, β2 produces an instantaneous outwardrectification that involves the β2 external loop [8].
Whether tungstate is a better BK channel modulator thanother known BK channel openers still remains unclear.Certainly, a variety of small synthetic molecules and naturalproduct compounds have been identified as selective BKchannel activators. However, little is known about their site(s)of interaction and most have serious safety and efficacyproblems for clinical use, such as low potency, inadequateselectivity, or poor membrane permeability [14, 23, 26].
In summary, this study provides new insights into themolecular mechanism by which tungstate favors the activityof BK channels. Our results support the idea that tungstateactivates BK channels by interacting with the pore-forming αchannel subunit in a region involving the Mg2+ binding site,thus explaining the Mg2+ dependence of the tungstate action.Our data also suggest that the β dependence of BK channelactivation by tungstate is related to how the different βsubunits interact with the BK α channel subunit to modulateits activity, with the essential involvement of residues locatedat the β extracellular loop that have an important role in theregulation of voltage sensor activation and gating of the BKchannel. Finally, we believe that a better understanding of themechanisms used by BK channel modulators may help toimprove the design of efficient and safe BK opening drugs.
Acknowledgments We thank Dr. R. Latorre and Dr. L. Toro forproviding the cDNAS for the human BK channel α and β1 subunits,respectively. This work was supported by grants from the SpanishMinistry of Economy and Competitiveness (SAF2012-31089,
SAF2012-38140), FEDER Funds, Fondo de Investigación Sanitaria(Red HERACLES RD12/0042/0014), and Generalitat de Catalunya(grant SGR09-1369). MAV is the recipient of an ICREA AcademiaAward (Generalitat de Catalunya).
Conflict of interest The authors declare that they have no conflict ofinterest.
References
1. Amberg GC, Bonev AD, RossowCF, NelsonMT, Santana LF (2003)Modulation of the molecular composition of large conductance, Ca2+
activated K+ channels in vascular smooth muscle duringhypertension. J Clin Invest 112:717–724
2. Amberg GC, Santana LF (2003) Downregulation of the BK channelβ1 subunit in genetic hypertension. Circ Res 93:965–971
3. Bao L, Cox DH (2005) Gating and ionic currents reveal how theBKCa channel's Ca2+ sensitivity is enhanced by its β1 subunit. J GenPhysiol 126:393–412
4. Barberá A, Gomis RR, Prats N, Rodríguez-Gil JE, Domingo M,Gomis R, Guinovart JJ (2001) Tungstate is an effective antidiabeticagent in streptozotocin-induced diabetic rats: a long-term study.Diabetologia 44:507–513
5. Berkefeld H, Fakler B, Schulte U (2010) Ca2+-activated K+ channels:from protein complexes to function. Physiol Rev 90:1437–1459
6. Bian S, Favre I, Moczydlowski E (2001) Ca2+-binding activity of aCOOH-terminal fragment of the Drosophila BK channel involved inCa2+-dependent activation. Proc Natl Acad Sci U S A 98:4776–4781
7. Brenner R, Perez GJ, Bonev AD, Eckman DM, Kosek JC, Wiler SW,Patterson AJ, Nelson MT, Aldrich RW (2000) Vasoregulation by the β1
subunit of the calcium-activated potassium channel. Nature 407:870–8768. Chen M, Gan G, Wu Y, Wang L, Wu Y, Ding J (2008) Lysine-rich
extracellular rings formed by hβ2 subunits confer the outwardrectification of BK channels. PLoS One 3:e2114
9. Chen X, Aldrich RW (2011) Charge substitution for a deep-poreresidue reveals structural dynamics during BK channel gating. JGen Physiol 138:137–154
10. Claret M, Corominola H, Canals I, Saura J, Barceló-Batllori S,Guinovart JJ, Gomis R (2005) Tungstate decreases weight gain andadiposity in obese rats through increased thermogenesis and lipidoxidation. Endocrinology 146:4362–4369
11. Contreras GF, Neely A, Alvarez O, Gonzalez C, Latorre R (2012)Modulation of BK channel voltage gating by different auxiliary βsubunits. Proc Natl Acad Sci U S A 109:18991–18996
12. Cox DH, Aldrich RW (2000) Role of the β1 subunit in large-conductance Ca2+-activated K+ channel gating energetics.Mechanisms of enhanced Ca2+ sensitivity. J Gen Physiol 116:411–432
13. Cui J, Aldrich RW (2000) Allosteric linkage between voltage andCa2+-dependent activation of BK-type mslo1 K+ channels.Biochemistry 39:15612–15619
14. Cui YM, Yasutomi E, Otani Y, Ido K, Yoshinaga T, Sawada K,Ohwada T (2010) Design, synthesis, and characterization of BKchannel openers based on oximation of abietane diterpenederivatives. Bioorg Med Chem 18:8642–8659
15. Dick GM, Sanders KM (2001) (Xeno)estrogen sensitivity of smoothmuscle BK channels conferred by the regulatory β1 subunit: a studyof β1 knockout mice. J Biol Chem 276:44835–44840
16. Fernández-Fernández JM, Tomás M, Vázquez E, Orio P, Latorre R,Sentí M, Marrugat J, Valverde MA (2004) Gain-of-function mutationin the KCNMB1 potassium channel subunit is associated with lowprevalence of diastolic hypertension. J Clin Invest 113:1032–1039
17. Fernández-Mariño AI, Porras-González C, González-Rodríguez P,Selent J, Pastor M, Ureña J, Castellano A, Valverde MA, Fernández-
Pflugers Arch - Eur J Physiol

Fernández JM (2012) Tungstate activates BK channels in a βsubunit- and Mg2+-dependent manner: relevance for arterialvasodilatation. Cardiovasc Res 95:29–38
18. Gruslova A, Semenov I, Wang B (2012) An extracellular domain ofthe accessory β1 subunit is required for modulating BK channelvoltage sensor and gate. J Gen Physiol 139:57–67
19. Hanner M, Vianna-Jorge R, Kamassah A, Schmalhofer WA, KnausHG, Kaczorowski GJ, Garcia ML (1998) The β subunit of the highconductance calcium-activated potassium channel. Identification ofresidues involved in charybdotoxin binding. J Biol Chem 273:16289–16296
20. Jaggar JH, Porter VA, Lederer WJ, Nelson MT (2000) Calciumsparks in smoothmuscle. Am J Physiol Cell Physiol 278:C235–C256
21. Jiang Z, Wallner M, Meera P, Toro L (1999) Human and rodentMaxiK channel β-subunit genes: cloning and characterization.Genomics 55:57–67
22. Kohler R (2012) Heavy metal to lower the pressure? Cardiovasc Res95:3–4
23. Kohler R, Kaistha BP, Wulff H (2010) Vascular KCa-channels astherapeutic targets in hypertension and restenosis disease. ExpertOpin Ther Targets 14:143–155
24. McManus OB, Helms LM, Pallanck L, Ganetzky B, Swanson R,Leonard RJ (1995) Functional role of the β subunit of highconductance calcium-activated potassium channels. Neuron 14:645–650
25. Meera P, Wallner M, Jiang Z, Toro L (1996) A calcium switch for thefunctional coupling between α (hslo) and β subunits (KV, Caβ) ofmaxi K channels. FEBS Lett 385:127–128
26. Nardi A, Olesen SP (2008) BK channel modulators: a comprehensiveoverview. Curr Med Chem 15:1126–1146
27. Peredó HA, Zabalza M, Mayer MA, Carranza A, Puyó AM (2010)Sodium tungstate and vanadyl sulfate effects on blood pressure andvascular prostanoids production in fructose-overloaded rats. Clin ExpHypertens 32:453–457
28. Pérez GJ, Bonev AD, Nelson MT (2001) Micromolar Ca2+ fromsparks activates Ca2+-sensitive K+ channels in rat cerebral arterysmooth muscle. Am J Physiol Cell Physiol 281:C1769–C1775
29. Piskorowski RA, Aldrich RW (2006) Relationship between poreoccupancy and gating in BK potassium channels. J Gen Physiol127:557–576
30. Pluger S, Faulhaber J, Furstenau M, Lohn M, Waldschutz R, GollaschM, Haller H, Luft FC, Ehmke H, Pongs O (2000) Mice with disruptedBK channel β1 subunit gene feature abnormal Ca2+ spark/STOCcoupling and elevated blood pressure. Circ Res 87:E53–E60
31. Seibold MA, Wang B, Eng C, Kumar G, Beckman KB, Sen S,Choudhry S, Meade K, LeNoir M, Watson HG, Thyne S, WilliamsLK, Kumar R, Weiss KB, Grammer LC, Avila PC, Schleimer RP,Burchard EG, Brenner R (2008) An African-specific functionalpolymorphism in KCNMB1 shows sex-specific association withasthma severity. Hum Mol Genet 17:2681–2690
32. Semenov I, Wang B, Herlihy JT, Brenner R (2006) BK channel β1-subunit regulation of calcium handling and constriction in trachealsmooth muscle. Am J Physiol Lung Cell Mol Physiol 291:L802–L810
33. Sentí M, Fernández-Fernández JM, Tomás M, Vázquez E, Elosua R,Marrugat J, Valverde MA (2005) Protective effect of the KCNMB1
E65K genetic polymorphism against diastolic hypertension in agingwomen and its relevance to cardiovascular risk. Circ Res 97:1360–1365
34. Shi J, Cui J (2001) Intracellular Mg2+ enhances the function of BK-type Ca2+-activated K+ channels. J Gen Physiol 118:589–606
35. Suzuki H, Delano FA, Parks DA, Jamshidi N, Granger DN, Ishii H,Suematsu M, Zweifach BW, Schmid-Schonbein GW (1998)Xanthine oxidase activity associated with arterial blood pressure inspontaneously hypertensive rats. Proc Natl Acad Sci U S A 95:4754–4759
36. Swei A, Lacy F, Delano FA, Parks DA, Schmid-Schonbein GW(1999) A mechanism of oxygen free radical production in the Dahlhypertensive rat. Microcirculation 6:179–187
37. Tanaka Y, Meera P, Song M, Knaus HG, Toro L (1997) Molecularconstituents of maxi KCa channels in human coronary smoothmuscle: predominant α + β subunit complexes. J Physiol (Lond)502(Pt 3):545–557
38. Tomás M, Vázquez E, Fernández-Fernández JM, Subirana I, Plata C,Heras M, Vila J, Marrugat J, Valverde MA, Sentí M (2008) Geneticvariation in the KCNMA1 potassium channelα subunit as risk factorfor severe essential hypertension and myocardial infarction. JHypertens 26:2147–2153
39. Torres YP, Morera FJ, Carvacho I, Latorre R (2007) A marriage ofconvenience: β-subunits and voltage-dependent K+ channels. J BiolChem 282:24485–24489
40. ValverdeMA, Rojas P, Amigo J, Cosmelli D, Orio P, BahamondeMI,Mann GE, Vergara C, Latorre R (1999) Acute activation of Maxi-Kchannels (hSlo) by estradiol binding to the β subunit. Science 285:1929–1931
41. WallnerM,Meera P, Toro L (1999)Molecular basis of fast inactivationin voltage and Ca2+-activated K+ channels: a transmembrane beta-subunit homolog. Proc Natl Acad Sci U S A 96:4137–4142
42. Xia XM, Ding JP, Lingle CJ (2003) Inactivation of BK channels bythe NH2 terminus of the β2 auxiliary subunit: an essential role of aterminal peptide segment of three hydrophobic residues. J GenPhysiol 121:125–148
43. Xia XM, Ding JP, Lingle CJ (1999) Molecular basis for theinactivation of Ca2+- and voltage-dependent BK channels in adrenalchromaffin cells and rat insulinoma tumor cells. J Neurosci 19:5255–5264
44. Xia XM, Zeng X, Lingle CJ (2002)Multiple regulatory sites in large-conductance calcium-activated potassium channels. Nature 418:880–884
45. Yang H, Shi J, Zhang G, Yang J, Delaloye K, Cui J (2008) Activationof Slo1 BK channels by Mg2+ coordinated between the voltagesensor and RCK1 domains. Nat Struct Mol Biol 15:1152–1159
46. Yuan P, Leonetti MD, Hsiung Y, MacKinnon R (2011) Openstructure of the Ca2+ gating ring in the high-conductanceCa2+-activated K+ channel. Nature 481:94–97
47. Yuan P, Leonetti MD, Pico AR, Hsiung Y, MacKinnon R (2010)Structure of the human BK channel Ca2+-activation apparatus at 3.0A resolution. Science 329:182–186
48. Zhang X, Solaro CR, Lingle CJ (2001) Allosteric regulation of BKchannel gating byCa2+ andMg2+ through a nonselective, low affinitydivalent cation site. J Gen Physiol 118:607–636
Pflugers Arch - Eur J Physiol
Top Related