







γλώσσες
Σελίδες
Νομικός


BMB reports
824 BMB reports http://bmbreports.org
*Corresponding author. Tel: 82-51-510-2268; Fax: 82-51-514-1778; E-mail: [email protected] DOI 10.5483/BMBRep.2010.43.12.824
Received 4 October 2010, Accepted 13 November 2010
Keywords: β-carboline, Melanogenesis, MITF, p38 MAPK, Tyrosinase
Beta-carboline alkaloids harmaline and harmalol induce melanogenesis through p38 mitogen-activated protein kinase in B16F10 mouse melanoma cellsSun Young Park1,2, Young Hun Kim1, Young Hee Kim2, Geuntae Park3 & Sang-Joon Lee4,*1Bio-IT Fusion Technology Research Institute, 2Department of Molecular Biology, 3Institute for Research & Industry Cooperation, 4Department of Microbiology, Pusan National University, Busan, Korea
Melanin synthesis is regulated by melanocyte specific en-zymes and related transcription factors. β-carboline alkaloids including harmaline and harmalol are widely distributed in the environment including several plant families and alcoholic beverages. Presently, melanin content and tyrosinase activity were increased in melanoma cells by harmaline and harmalol in concentration- and time-dependent manners. Increased pro-tein levels of tyrosinase, tyrosinase-related protein-1 (TRP-1), and TRP-2 were also evident. In addition, immunofluorescence and Western blot analyses revealed harmaline and harmalol increased cAMP response element binding protein phosphor-ylation and microphthalmia-associated transcription factor ex-pression. In addition to studying the signaling that leads to melanogenesis, roles of the p38 MAPK pathways by the har-maline and harmalol were investigated. Harmaline and harma-lol induced time-dependent phosphorylation of p38 MAPK. Harmaline and harmalol stimulated melanin synthesis and ty-rosinase activity, as well as expression of tyrosinase and TRP-1 and TRP-2 indicating that these harmaline and harmalol in-duce melanogenesis through p38 MAPK signaling. [BMB re-ports 2010; 43(12): 824-829]
INTRODUCTION
The synthesis of melanin pigments or melanogenesis has many important physiological functions that include photoprotection of the human skin from ultraviolet (UV) irradiation (1). Me-lanogenesis is a complex pathway involving melanin syn-thesis, melanin transport, and melanosome release (2). Mela-nin synthesis is stimulated by various effects such as α-melano-cyte-stimulating hormone (α-MSH); cyclic AMP (cAMP) elevat-
ing agents including forskolin, glycyrrhizin, and isobutyl-methylxanthine; UV-B radiation; and the placental total lipid fraction (3-5). In addition, melanin synthesis occurs in melano-cytes and melanoma cells through an enzymatic process cata-lyzed by tryrosinase, tyrosinase related protein-1 (TRP-1), and tyrosinase related protein-2 (TRP-2), which converts tyrosine to 3,4-dihydroxyphenylalanine (DOPA) and catalyzes the oxida-tion of DOPA into DOPAquinone (6). Dopaquinone is con-verted to dopachrome that is in turn converted to dihyrox-yindole or dihydroxyindole-2-carbxylic acid (DHICA) to form eumelanin. The cascade of enzymatic reactions in melanin synthesis is related in tyrosinase, TRP-1 (dopachrome tauto-merase) and TRP-2 (DHICA oxidase) (7).
Microphthalmia-associated transcription factor (MITF) is the most important transcription factor in the regulation of ty-rosinase and expression of the genes for TRP-1 and TRP-2, as tyrosinase, TRP-1, and TRP-2 harbor the MITF binding site, thereby leading to the regulation of activation of melanocyte differentiation (8). cAMP response element binding protein (CREB) is also one of the major transcription factors of MITF and, thus, plays a central role in melanogenesis (9). The CREB binding site is present in the MITF promoter region; as a con-sequence, CREB binds to and activates the MITF promoter, which leads to the indirect activation of melanogenesis (10).
Mitogen-activated protein kinases (MAPKs) are a highly con-served family of protein serine/threonine kinases that include extracellularly responsive kinases (ERK1/2), c-jun N-terminal or stress-regulated protein kinases (JNK/SAPK), and p38 MAPKs. They are involved in a diversity of cellular activities and have an important regulatory role in melanogenesis (11). p38 MAPK activation is related to an increase in melanin synthesis and is involved in the expression of melanogenesis related molecules (12). In addition, p38 MAPK activation is involved in α-MSH- induced melanogeneis, such as activation of MITF expression and activation of tyrosinase transcription (13), whereas the ex-tracellular signal-regulated kinase (ERK)1/2 and c-Jun NH2-terminal kinase/stress-activated protein kinase (JNK/SAPK) pathways are related with the down-regulation of melanogenesis (14). Accor-dingly, ERK signaling inactivation or p38 MAPK signaling acti-vation stimulates melanogenesis by increasing MITF expre-

Melanogenesis induction of harmaline and harmalolSun Young Park, et al.
825http://bmbreports.org BMB reports
Fig. 1. Effect of harmaline and harmalol on melanin synthesis, ty-rosinase activity and melanogenesis related proteins in B16F10 cells. (A) B16 cells treated for 72 h with harmaline (1-10 μM) and harmalol (5-20 μM) were analyzed for their melanin content. The melanin content was corrected based on the amount of pro-tein concentration. α-MSH (1 μM) served as the positive control for 72 h. Cells treated with harmaline (5 μM) and harmalol (20 μM) for determined times were analyzed for their melanin con-tent (B) and tyrosinase activity (C). α-MSH (1 μM) was the pos-itive control for 72 h. The cellular tyrosinase activity was de-termined and is reported as a percentage reported relative to that in the control cells. Values are means ± SE of three independent experiments. *P < 0.05 versus untreated control. In situ tyro-sinase activity determined by incubation of cells in L-DOPA. Imageswere captured under identical conditions using phase contrast mi-croscopy and are representative of three independent experiments (D). (E) Cells were incubated for determined times with harmaline and harmalol. The cellular extracts were then subjected to Westernblot analysis using tyrosinase, TRP-1, and TRP-2 antibody. Equal protein loading was confirmed by α-tubulin expression. Numbers at the bottom are expressed as relative intensity of band (fold of control) estimated by using ImageQuant TL software.
ssion and tyrosinase activity.β-carbolines are component of some medicinal plants, such
as Passiflora edulis, Passiflora incarnate and Peganum harmala. These plants have been used for anti-jaundice, anti-lumbago and anti-inflammation agent in oriental medicine. Recently, the mechanism responsible for traditional herbal medicines variable effects has not been fully clarified yet. β-carboline al-kaloids are widely distributed in environments including sev-eral plant families, well-cooked foods, tobacco smoke, and al-coholic beverages (15). Additionally, they are endogenous in mammalian tissues. β-carboline alkaloids have a reported wide range of pharmacological, neurophysiologic, and biochemical actions (16, 17); are both mutagenic and carcinogenic (18); and act as a neurotoxins in some neurodegenerative diseases. In contrast, the β-carboline alkaloids harmaline and harmalol (Fig. 1A) inhibit monoamine oxidase and do not induce neuro-toxicity (19). In addition, β-carbolines possess effective anti-oxidant and radical scavenging properties, protect from oxida-tive neuronal damage, and inhibit the ion-mediated cytotoxic effect of 1-methyl-4-phenylpyridinium (MMP+) and cytotoxic effects on tumor cells (17). Furthermore, plants containing β-carboline alkaloids have long been used as traditional medi-cines for the treatment of various diseases including cancer, jaundice, malaria, and asthma (19, 20). Recently, it was re-ported that β-carbolines induce apoptosis by caspase-8 activa-tion in carcinoma cells and function as an anti-inflammatory compound that suppresses lipopolysaccharide (LPS)-induced inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) through the nuclear factor (NF)-kappaB and I-kappaB kinase (IKK) signaling pathways (21, 22). However, it is not known whether β-carbolines directly contribute to melano-genesis dysfunction. Therefore, using mouse B16F10 melano-ma cells, we attempted to determine the effects and mecha-nisms of the β-carbolines harmaline and harmalol on melano-genesis, specifically whether these β-carbolines are affected by melanogenesis, and characterized the signal transduction path-way involved.
RESULTS
Harmaline and harmalol increase cellular melanin content in B16F10 melanoma cellsCells were treated with various concentrations of harmaline and harmalol for 72 h. Cellular melanin content was assessed by determination of intracellular melanin, expressed as a per-centage related to the total protein. As shown in Fig. 1A, har-maline and harmalol treatment increased the melanin content, consistent with enhanced melanin synthesis in response to se-rial concentrations of harmaline and harmalol. Of note, aug-mentation of intracellular melanin content by harmaline ap-peared more sustained than that evoked by harmalol. The cell growth effect of harmaline and harmalol were investigated in cells treated with various concentrations (1-20 μM) of harma-line and harmalol for 72 h (data not shown). Cell growth arrest
was induced by 10 μM harmaline and 20 μM harmalol, while treatment with either β-carboline alkaloid did not result in cell necrosis and detachment from the culture plates (data not shown).
Harmaline and harmalol induce tyrosinase activity and melanogenesis-related proteinsTo verify if the effect of harmaline and harmalol on melanin synthesis and tyrosinase activity occurred in a time-dependent

Melanogenesis induction of harmaline and harmalolSun Young Park, et al.
826 BMB reports http://bmbreports.org
Fig. 2. Effects of harmaline and harmalol on MITF and phos-pho-CREB expression. B16F10 cells were grown on glass cover slips and then treated with harmaline (5 μM) and harmalol (20 μM). α-MSH (1 μM) served as the positive control for 24 h. At 24 h, the cells were fixed and analyzed by immunofluorescence labeling with a rabbit monoclonal anti-MITF and anti-phospho- CREB antibody, followed by incubation with Alex Fluor 488-con-jugated goat anti-rabbit IgG. Nuclei were labeled with DAPI. Original magnification, ×100. (A). Cellular protein extracts were analyzed by Western blotting with anti-MITF and anti-phospho- CREB (B). Equal protein loading was confirmed by α-tubulin and CREB expression. Numbers at the bottom are expressed as rela-tive intensity of band (fold of control) estimated by using Image Quant TL software.
manner, cells were exposed to 5 μM harmaline, 20 μM harma-lol, or, as a positive control, 1 μM α-MSH. Tyrosinase is a key enzyme in melanin bisosynthesis, in which melanin is formed from the tyrosinase-catalyzed oxidation of L-tyrosinase. Thus, melanogenesis is regulated by the activity of tyrosinase, TRP-1, and TRP-2. An increase of cellular tyrosinase activity and mel-anin synthesis was evident in cells exposed to harmaline and harmalol, consistent with harmaline and harmalol accumu-lation of melanin biosynthesis by B16F10 cells via enhance-ment of tyrosinase activity. In order to clarify further the mech-anism of tyrosinase activation by harmaline and harmalol, the levels of melanogenesis related proteins including tyrosinase, TRP-1, and TRP-2 were determined in B16F10 cells exposed to 5 μM harmaline and 20 μM harmalol by Western blot ana-lysis. Tyrosinase, TRP-1, and TRP-2 expression were enhanced in harmaline- and harmalol-treated cells (Fig. 1E). The results were consistent with a harmaline- and harmalol-induced mela-nin synthesis mediated by the activation of the expressions of tyrosinase, TRP-1, and TRP-2.
Harmaline and harmalol increase MITF expression and CREB activation MITF plays an important role in melanogenesis as the tran-scription factor that regulates tyrosinase, TRP-1, and TRP-2 gene expression (7). The mechanism by which MITF regulates melanogenesis involves the p38 MAPK signaling pathway (12), which also induces the activation of melanogenesis by α-MSH (4). Furthermore, phosphorylation of CREB binds the CRE con-sensus sequence of the MITF promoter to activate the ex-pression of MITF (14). Since earlier experiments in this study demonstrated that harmaline and harmalol increased melanin synthesis and melanogenesis related gene expression, we fur-ther hypothesized that these harmaline and harmalol could af-fect the expression of MITF and CREB activation, which tran-scriptionally activate melanogenesis. To investigate this hy-pothesis, the effect of harmaline and harmalol on MITF ex-pression and CREB activation was examined in B16F10 cells by immunofluorescence and Western blotting analyses. Immu-nofluorescence studies with monoclonal anti-MITF and an-ti-phospho-CREB antibodies detected increased fluoresecence intensity in cells treated with harmaline and harmalol com-pared with untreated cells (Fig. 2A). Western blotting analysis using the antibodies detected increased MITF expression and CREB activation in harmaline and harmalol treated cells (Fig. 2B). Stimulation of MITF expression and CREB activation by harmaline and harmalol reached its maximum after 24 h of treatment.
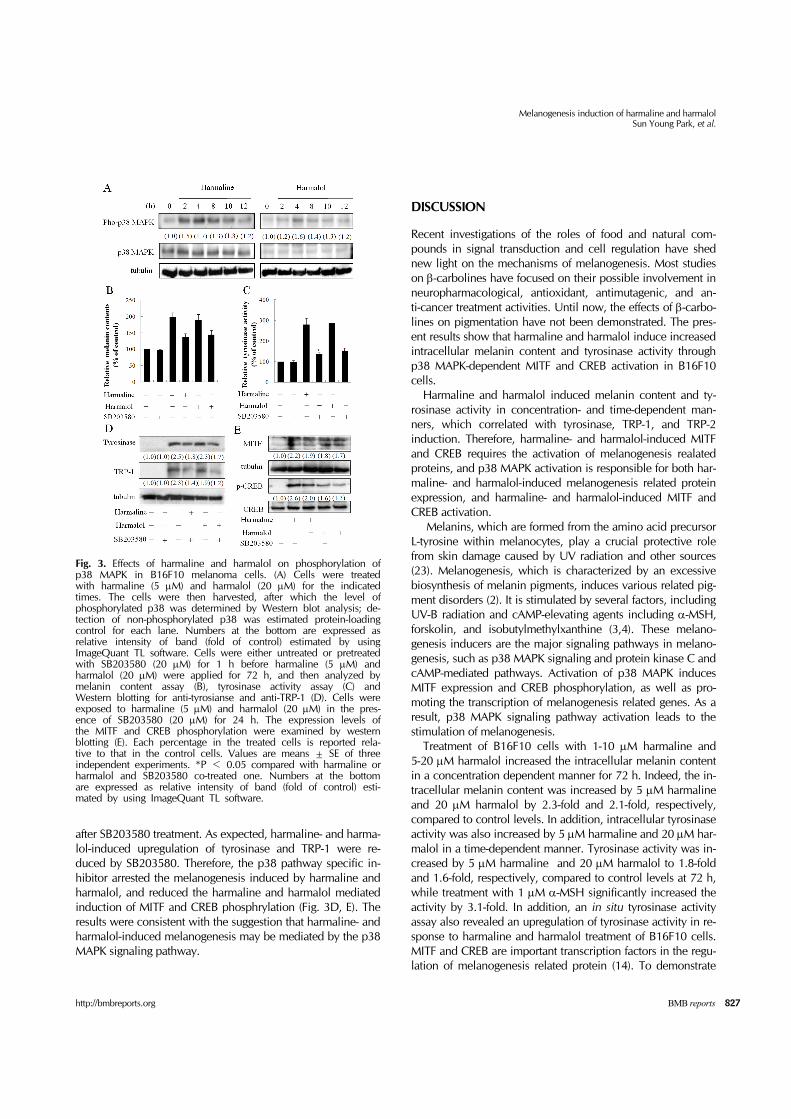
Harmaline and harmalol induces phosphorylation of p38 MAPK in B16F10 melanoma cells p38 MAPK induces melanogenesis (12). In order to demon-strate the mechanisms of harmaline and harmalol-induced melanogenesis, p38 MAPK phosphorylation was evaluated us-ing a Western blotting assay with B16F10 cells treated for vari-
ous times with harmaline or harmalol. p38 MAPK was phos-phorylated in a time-dependent fashion (Fig. 3A) with p38 MAPK activation induced 4 h after the treatments, indicating that the activation of melanogenesis by harmaline and harma-lol is related to the activation of p38 MAPK. To directly dem-onstrate the involvement of p38 MAPK signaling in harmaline- and harmalol-induced melanogenesis, B16F10 cells were treat-ed with the p38 MAPK inhibitor SB203580 for 1 h before har-maline and harmalol treatment, and the cells were treated with harmaline and harmalol for 72 h. Melanin synthesis and ty-rosinase activity were induced in harmaline- and harmalol- treated cells; these inductions were abrogated by SB203580 (Fig. 3B, C). Melanogenesis related protein was also assessed
Melanogenesis induction of harmaline and harmalolSun Young Park, et al.
827http://bmbreports.org BMB reports
Fig. 3. Effects of harmaline and harmalol on phosphorylation of p38 MAPK in B16F10 melanoma cells. (A) Cells were treated with harmaline (5 μM) and harmalol (20 μM) for the indicated times. The cells were then harvested, after which the level of phosphorylated p38 was determined by Western blot analysis; de-tection of non-phosphorylated p38 was estimated protein-loading control for each lane. Numbers at the bottom are expressed as relative intensity of band (fold of control) estimated by using ImageQuant TL software. Cells were either untreated or pretreated with SB203580 (20 μM) for 1 h before harmaline (5 μM) and harmalol (20 μM) were applied for 72 h, and then analyzed by melanin content assay (B), tyrosinase activity assay (C) and Western blotting for anti-tyrosianse and anti-TRP-1 (D). Cells were exposed to harmaline (5 μM) and harmalol (20 μM) in the pres-ence of SB203580 (20 μM) for 24 h. The expression levels of the MITF and CREB phosphorylation were examined by western blotting (E). Each percentage in the treated cells is reported rela-tive to that in the control cells. Values are means ± SE of three independent experiments. *P < 0.05 compared with harmaline or harmalol and SB203580 co-treated one. Numbers at the bottom are expressed as relative intensity of band (fold of control) esti-mated by using ImageQuant TL software.
after SB203580 treatment. As expected, harmaline- and harma-lol-induced upregulation of tyrosinase and TRP-1 were re-duced by SB203580. Therefore, the p38 pathway specific in-hibitor arrested the melanogenesis induced by harmaline and harmalol, and reduced the harmaline and harmalol mediated induction of MITF and CREB phosphrylation (Fig. 3D, E). The results were consistent with the suggestion that harmaline- and harmalol-induced melanogenesis may be mediated by the p38 MAPK signaling pathway.
DISCUSSION
Recent investigations of the roles of food and natural com-pounds in signal transduction and cell regulation have shed new light on the mechanisms of melanogenesis. Most studies on β-carbolines have focused on their possible involvement in neuropharmacological, antioxidant, antimutagenic, and an-ti-cancer treatment activities. Until now, the effects of β-carbo-lines on pigmentation have not been demonstrated. The pres-ent results show that harmaline and harmalol induce increased intracellular melanin content and tyrosinase activity through p38 MAPK-dependent MITF and CREB activation in B16F10 cells.
Harmaline and harmalol induced melanin content and ty-rosinase activity in concentration- and time-dependent man-ners, which correlated with tyrosinase, TRP-1, and TRP-2 induction. Therefore, harmaline- and harmalol-induced MITF and CREB requires the activation of melanogenesis realated proteins, and p38 MAPK activation is responsible for both har-maline- and harmalol-induced melanogenesis related protein expression, and harmaline- and harmalol-induced MITF and CREB activation.
Melanins, which are formed from the amino acid precursor L-tyrosine within melanocytes, play a crucial protective role from skin damage caused by UV radiation and other sources (23). Melanogenesis, which is characterized by an excessive biosynthesis of melanin pigments, induces various related pig-ment disorders (2). It is stimulated by several factors, including UV-B radiation and cAMP-elevating agents including α-MSH, forskolin, and isobutylmethylxanthine (3,4). These melano-genesis inducers are the major signaling pathways in melano-genesis, such as p38 MAPK signaling and protein kinase C and cAMP-mediated pathways. Activation of p38 MAPK induces MITF expression and CREB phosphorylation, as well as pro-moting the transcription of melanogenesis related genes. As a result, p38 MAPK signaling pathway activation leads to the stimulation of melanogenesis.
Treatment of B16F10 cells with 1-10 μM harmaline and 5-20 μM harmalol increased the intracellular melanin content in a concentration dependent manner for 72 h. Indeed, the in-tracellular melanin content was increased by 5 μM harmaline and 20 μM harmalol by 2.3-fold and 2.1-fold, respectively, compared to control levels. In addition, intracellular tyrosinase activity was also increased by 5 μM harmaline and 20 μM har-malol in a time-dependent manner. Tyrosinase activity was in-creased by 5 μM harmaline and 20 μM harmalol to 1.8-fold and 1.6-fold, respectively, compared to control levels at 72 h, while treatment with 1 μM α-MSH significantly increased the activity by 3.1-fold. In addition, an in situ tyrosinase activity assay also revealed an upregulation of tyrosinase activity in re-sponse to harmaline and harmalol treatment of B16F10 cells. MITF and CREB are important transcription factors in the regu-lation of melanogenesis related protein (14). To demonstrate

Melanogenesis induction of harmaline and harmalolSun Young Park, et al.
828 BMB reports http://bmbreports.org
the mechanism of harmaline- and harmalol-induced melano-genesis, B16F10 cells were treated with 5 μM harmaline and 20 μM harmalol for various times. As shown in Fig. 3, treat-ment of B16F10 cells with harmaline and harmalol induced MITF expression and CREB phosphorylation. cAMP is an im-portant factor in the melanogenesis-related signal transduction pathways, regulating the activation of protein kinase A follow-ing CREB phosphorylation. We measured the intracellular cAMP levels in B16F10 cells and observed no increase when cells were treated with harmaline and harmalol (data not shown). These results indicate that harmaline and harmalol do not stimulate melanogenesis via cAMP levels.
Activation of p38 MAPK stimulates melanogenesis (12). We confirmed that the increased synthesis of melanin induced by harmaline and harmalol is mediated by p38 MAPK activation; measurement of the level of p38 MAPK phosphorylation vari-ous lengths of times revealed that p38 MAPK phosphorylation increased when cells were treated with harmaline or harmalol. As both compounds activate the p38 MAPK signaling path-way, we expected, and demonstrated, the suppressive activity of harmaline- and harmalol-induced melanogenesis by SB 203580, a selective inhibitor of p38 MAPK. The melanin con-tent and tyrosinase activity of cells treated with harmaline and harmalol decreased following treatment with SB23580. In ad-dition, this treatment decreased expression of harmaline- and harmalol-induced melanogenesis related proteins.
In summary, the β-carbolines harmaline and harmalol in-duce cellular melanin biosynthesis and tyrosinase activity in B16F10 cells, up-regulating CREB phosphorylation and ex-pression of MITF, tyrosinase, TRP-1 and TRP-2, and phosphor-ylation of p38 MAPK. These consistent results suggest that β-cabolines might be useful for treatment of hypopigmentation- related disorders such as vitiligo.
MATERIALS AND METHODS
MaterialsHarmaline and harmalol, α-MSH, L-DOPA, 3-(4,5-dimetylthiazol- 2-yl)-2,5- diphenyl tetrazolium bromide (MTT) and all other re-agents were purchased from Sigma-Aldrich (St. Louis. MO, USA). SB203580 was purchased from A.G. Scientific (San Diego, CA, USA). Antibodies recognizing phospho-p38, phos-pho-CREB, and CREB were obtained from Cell Signaling Tech-nology (Beverley, MA, USA). Antibodies to p38, tyrosinase, TRP1, TRP2, and MITF were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell cultureCells of the B16-F10 murine melanoma cell line obtained from the American Type Culture Collection (ATCC; Rockville, MD, USA) were grown as monolayers in Dulbecco's modified Eagle's medium (DMEM; GIBCO BRL, Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; GIBCO BRL). The cells were incubated at 37oC in a hu-
midified atmosphere composed of 5% CO2 and 95% air. To avoid changes in cell characteristics produced by extended cell culture periods, cells were used between passages 15 and 25. Each cell suspension was split every 2 days to maintain ex-ponential growth.
Cell viability assayThe cells were incubated in wells of a 24-well plate at a den-sity of 4 × 104 cells/well. MTT solution (50 μg/ml) was added to each well. The plates were then incubated for an additional 3 h at 37oC in a 5% CO2 atmosphere, after which the super-natant was removed and the formazan crystals that had formed in the viable cells were solubilized with dimethylsulfoxide (DMSO). The absorbance of each well was then read at 570 nm using an enzyme-linked immunosorbant assay (ELISA) reader (Wallace, Boston, MA, USA).
Determination of melanin contentData are expressed as mean ± standard error (SE). Each experi-ment was repeated at least three times. Statistical analysis was performed with SPSS, version 16.0 software to determine sig-nificant differences. We used either one- or two-way ANOVA followed by Dunn’s post hoc tests for analyses. Values of *P < 0.05 and **P < 0.01 were considered statistically significant.
B16 F10 cell tyrosinase activity assayTyrosinase activity was determined by measuring the rate of dopachrome formation of L-DOPA. Cells were treated with harmaline and harmalol for 72 h, after which the cells were washed in ice-cold PBS and lysed in PBS containing 1% (w/v) Triton X-100. The tyrosinase substrate, L-DOPA (2 mg/ml), was prepared in the same phosphate lysis buffer. Each extract was placed in wells of a 96-well plate and the enzymatic assay was commenced by adding L-DOPA. After incubation, dopach-rome formation was assayed by measuring absorbance at 405 nm using a microplate reader. The value of each measurement was expressed as percentage change from the control. In-situ L-DOPA reactivity of B16F10 cells was assessed using cultures fixed in 3.5% paraformaldehyde in PBS for 10 min at room temperature, after which they were permeabilized with 100% methanol for 10 min. Cells were incubated in L-DOPA (2 mg/ml) for 4 h at 37oC prior to photography using an Axiovert 40CFL inverted microscope (Carl Zeiss, Jena, Germany) equip-ped with an InFinity CAPTURE application version 4.6.0 digi-tal video camera system (Lumenera, Ottawa, Ontario, Canada).
Western blot analysisCells were harvested in ice-cold lysis buffer consisting of 1% Triton X-100, 1% deoxycholate, and 0.1% sodium dodecyl sulfate (SDS). The protein content of the cell lysates was then determined using Bradford reagent (Bio-Rad; Hercules, CA, USA). Protein in each sample (50 μg total protein) were re-solved by 7.5% SDS-polyacrylamide gel electrophoresis (SDS-

Melanogenesis induction of harmaline and harmalolSun Young Park, et al.
829http://bmbreports.org BMB reports
PAGE), transferred to a polyvinylidene difluoride (PVDF) mem-brane and exposed to the appropriate antibodies. The proteins were visualized by the enhanced chemiluminescence de-tection system (Amersham Biosciences, Piscataway, NJ, USA) using horseradish peroxidase-conjugated anti-rabbit or an-ti-mouse secondary antibodies. Images were acquired using an ImageQuant 350 analyzer (Amersham Biosciences) and densi-tometry was performed using ImageQuant TL software (Amer-sham Biosciences).
Immunofluorescence confocal microscopyB16F10 cells cultured directly on glass cover-slips were fixed with 3.5% paraformaldehyde in PBS for 10 min at room tem-perature, after which they were permeabilized with 100% methanol for 10 min. To evaluate MITF expression and CREB phosphorylation, the cells were treated with a 1:1,000 dilu-tion in PBS of polyclonal antibody against MITF or phosph- CREB overnight. Next, the cells were extensively washed with PBS and incubated with a 1:500 dilution in PBS of secondary fluorescein isothiocynate (FITC)-conjugated donkey anti-rabbit IgG antibody for 1 h at room temperature. Finally, the nuclei were stained with 1 g/ml of 4',6-diamidino-2-phenylindole (DAPI) and then analyzed by confocal microscopy using a Zeiss LSM 510 Meta apparatus.
Statistical analysisThe data is expressed as mean ± standard error (SE). Each ex-periment was repeated at least three times. Statistical analysis was performed with SPSS, version 16.0 software to determine significant differences. We used either one- or two-way ANOVA followed by Dunn’s post hoc tests for analyses. Values of *P < 0.05 was considered statistically significant
Conflict of interestThe authors have declared that no conflict of interest exists.
REFERENCES
1. Eller, M. E., Yaar, M. and Gilchrest, B. A. (1994) DNA damage and melanogenesis. Nature 372, 413-414.
2. Boissy, R. (2003) Melanosome transfer to and trnslocation in the keratinocyte. Exp. Derm. 12, 5-12.
3. Halaban, R., Pomerantz, S. H., Marshall, S. and Lerner, A. (1981) Tyrosinase activity and abundance in Cloudman melanoma cells. Biochem. Biophys. Acta. 230, 383-387.
4. Hunt, G., Todd, C., Creswell, J. E. and Thody, A. J. (1994) Alpha-melanocyte stimulating hormone and its analogue Nle4DPhe7 alpha-MSH affect morphology, tyrosinase ac-tivity and melanogenesis in cultured human melanocytes. J. Cell Sci. 107, 205-211.
5. Wong, G. and Pawelek, J. (1975) Melanocyte-stimulating hormone promotes activation of pre-existing tyrosinase molecules in Cloudman S91 melanoma cells. Nature 255, 644-646.
6. Hearing, V. J. and Jimenez, M. (1989) Analysis of mam-
malian pigmentation at the molecular level. Pigment Cell Res. 2, 75-85.
7. Hearing, V. J. and Tsukamoto, K. (1991) Enzymatic con-trol of pigmentation in mammals. FASEB J. 5, 2902-2909.
8. Yasumoto, K., Yokoyama, K., Takahashi, K., Tomita, Y. and Shibahara, S. (1997) Functional analysis of microphthalmia- associated transcription factor in pigment cell-specific transcription of the human tyrosinase family genes. J. Biol. Chem. 272, 503-509.
9. Buscà, R. and Ballotti, R. (2000) Cyclic AMP a key mes-senger in the regulation of skin pigmentation. Pigment Cell Res. 13, 60-69.
10. Englaro, W., Rezzonico, R., Durand-Clément, M., Lalle-mand, D., Ortonne, J. P. and Ballotti, R. (1995) Mitogen- activated protein kinase pathway and AP-1 are activated during cAMP-induced melanogenesis in B-16 melanoma cells. J. Biol. Chem. 270, 24315-24320.
11. Seger, R. and Krebs, E. G. (1982) The MAPK signaling cascade. FASEB J. 9, 726-735.
12. Singh, S. K., Sarkar, C., Mallick, S., Saha, B., Bera, R. and Bhadra, R. (2005) Human placental lipid induces melano-genesis through p38 MAPK in B16F10 mouse melanoma. Pigment Cell Research 18, 113-121.
13. Corre, S. and Galibert, M. D. (2005) Upstream stimulating factors: highly versatile stress-responsive transcription fac-tors. Pigment Cell Research 18, 337-348.
14. Bu, J., Ma, P. C., Chen, Z. Q., Zhou, W. Q., Fu, Y. J., Li, L. J. and Li, C. R. (2008) Inhibition of MITF and tyrosinase by paeonol-stimulated JNK/SAPK to reduction of phos-phorylated CREB. Am. J. Chin. Med. 36, 245-263.
15. Allen, J. R. and Holmested, B. R. (1980) The simple β-car-boline alkaloids. Phytochemistry 19, 1573-1582.
16. Kim, H. H., Jang, Y. Y., Han, E. S. and Lee, C. S. (1999) Differential antioxidant effects of ambroxol, rutin, gluta-thione and harmaline. J. Appl. Pharmacol. 7, 112-120.
17. Lee, C. S., Han, E. S., Jang, Y. Y., Han, J. H., Ha, H. W. and Kim, D. E. (2000) Protective effect of harmalol and harmaline on MPTP neurotoxicity in the mouse and dop-amine-induced damage of brain mitochondria and PC12 cell. J. Neurochem. 75, 521-531.
18. Wehner, F. C., Thiel, P. G. and Van Rensburg, S. J. (1979) Mutagenicity of alkaloids in the Salmonella/microsome systems. Mutat. Res. 66,187-190.
19. Buckholtz, N. S. and Boggan, W. O. (1977) Monoamine oxidase inhibition in brain and liver produced by b-carbo-line: structure-activity relationships and substrate speci-ficity. Biochem. Pharmacol. 26, 1991-1996.
20. Dymock, W., Warden, C. J. and Hooper, D. (1976) Phar-macopia indica. Vol 1, pp. 252-253, Harmard National Foundation of Pakistan, Pakistan.
21. Abe, A. and Yamada, H. (2009) Harmol induces apoptosis by caspase-8 activation independently of Fas/Fas ligand interaction in human lung carcinoma H596 cells. Antican-cer Drugs 20, 373-381.
22. Yoon, J. W., Kang, J. K. and Lee. K. R. (2005) β-carboline alkaloid suppresses NF-κB transcriptional activity through inhibition of IKK signaling pathway. J. Toxicol. Environ. Health A. 68, 2005-2017.
23. Friedmann, P. S. and Gilchrest, B. A. (1987) Ultraviolet ra-diation directly induces pigment production by cultured human melanocytes. J. Cell Physiol. 133, 88-94.
Top Related