






γλώσσες
Σελίδες
Νομικός


E-Mail [email protected]
Protein Membrane-Enclosed Organelles
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
Bacterial Intracellular Sulfur Globules: Structure and Function
James S. Maki
Department of Biological Sciences, Marquette University, Milwaukee, Wisc. , USA
Introduction
Sulfur is an important bioelement in the global eco-system. On the cellular level, sulfur is required for impor-tant amino acids, cysteine and methionine and is found in the vitamins thiamine pyrophosphate, coenzyme A and biotin, as well as in sulfolipids like α-lipoic acid [Gottschalk, 1986]. Inorganic sulfur compounds have many oxidation states [Dahl et al., 2008a; Steudel, 1989] and numerous transformations of sulfur in the environ-ment are mediated by special groups of prokaryotes, members of both Bacteria and Archaea. In its reduced forms, sulfur compounds are important electron donors and energy sources for those microbes that can utilize them. Chemolithotrophic prokaryotes use the energy for both respiration and fixation of CO 2 into organic carbon. Alternatively, the anaerobic photolithoautotrophic bac-teria use the energy primarily for CO 2 fixation [Dahl et al., 2008a; Ghosh and Dam, 2009]. In the process of oxi-dizing reduced sulfur compounds like sulfides and thio-sulfate into sulfate, a number of bacteria transiently form sulfur globules both extracellularly and intracellularly [Dahl and Prange, 2006]. In this review, the focus will be on the bacteria that form intracellular sulfur globules, intracellular meaning being found inside the cell wall, and what is currently known about globule structure, formation and degeneration.
Key Words
Sulfur oxidation · Intracellular sulfur globules
Abstract
Bacteria that oxidize reduced sulfur compounds like H 2 S of-ten transiently store sulfur in protein membrane-bounded intracellular sulfur globules; intracellular in this case mean-ing found inside the cell wall. The cultured bacteria that form these globules are primarily phylogenetically classi-fied in the Proteobacteria and are chemotrophic or photo-autotrophic. The current model organism is the purple sul-fur bacterium Allochromatium vinosum . Research on this bacterium has provided the groundwork for understanding the protein membranes and the sulfur contents of globules. In addition, it has demonstrated the importance of different genes (e.g. sulfur oxidizing, sox ) in their formation and in the final oxidation of sulfur in the globules to sulfate (e.g. dis-similatory sulfite reductase, dsr ). Pursuing the characteris-tics of other intracellular sulfur globule-forming bacteria through genomics, transcriptomics and proteomics will eventually lead to a complete picture of their formation and breakdown. There will be commonality to some of the ge-netic, physiological and morphological characteristics in-volved in intracellular sulfur globules of different bacteria, but there will likely be some surprises as well.
Copyright © 2013 S. Karger AG, Basel
Published online: August 5, 2013
Dr. James S. Maki Marquette University Department of Biological Sciences PO Box 1881, Milwaukee, WI 53201-1881 (USA) E-Mail james.maki @ marquette.edu
© 2013 S. Karger AG, Basel1464–1801/13/0235–0270$38.00/0
www.karger.com/mmb
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Bacterial Intracellular Sulfur Globules J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
271
The early microbiological study of intracellular sulfur globules, their structure and contents involves the re-search of some of the most well-known microbiologists in the history of the discipline [e.g. van Niel, 1931; Wino-gradsky, 1889]. Those interested in the early research on sulfur bacteria are referred to discussions by Waksman [1922], Waksman and Joffe [1922], Shively [2006], Dahl and Prange [2006], Trüper [2008], and papers referenced therein. The focus of this review will be on more recent information.
Phylogenetic Distribution of Genera with
Intracellular Sulfur Globules
The ability to form intracellular sulfur globules is pres-ent in a wide variety of cultured genera largely found in the α-, β-, γ- and ε-Proteobacteria ( fig. 1 ). Magnetotactic bacteria vary in their ability to form intracellular sulfur globules [Bazylinski et al., 2004; Bazylinski and Williams, 2006; Keim et al., 2005]. Those that do are primarily in the α-Proteobacteria ( fig. 1 ) and have been demonstrated to form globules using sulfide and/or thiosulfate [Bazy-linski and Williams 2006; Williams et al., 2006]. These genera include Magnetococcus , which branches out early from the other α-Proteobacteria [Bazylinski and Wil-liams, 2006] and the newly described genera Magnetovi-brio [Bazylinski et al., 2004, 2012] in which sulfur globules are formed when cells are grown on sulfide, and Magne-
tospira [Williams et al., 2012] in which they form when cells are grown on thiosulfate. However, the ability of magnetotactic bacteria to form globules is not restricted to the α-Proteobacteria. Recently, a magnetotactic bacte-rium that forms intracellular sulfur globules was discov-ered that phylogenetically belongs in the γ-Proteobacteria [Lefevre et al., 2012].
Members of two genera, Thermothrix and Macromo-nas , in the β-Proteobacteria have been observed to form globules. While one species of the genus Thermothrix ,
ThiorhodococcusAllochromatium
ThiocapsaChromatiumThiocystis
MarichromatiumIsochromatium
ThioflavicoccusThiococcus GammaThiohalocapsaHalochromatium
ThioalkalivibrioAchromatium
ThiothrixBeggiatoa
ThiomargaritaThioploca
MacromonasThermothrix
MagnetospiraMagnetovibrio Alpha
Beta
MagnetococcusAquifex
Fig. 1. Phylogenetic tree based on nearest neighbor analysis of 16S rRNA genes of the different genera, all in the Proteobacteria, in which intracellular sulfur globules have been reported. The tree is rooted with the genus Aquifex .
Fig. 2. Thiothrix spp. found on an aquatic macrophyte in the flume of a freshwater hydrothermal vent in Yellowstone Lake, Wyo., USA. Arrows show intracellular sulfur globules. Details about the habitat can be found in Konkol et al. [2010]. Scale bar = 1.0 μm [unpubl. micrograph, J.S. Maki].
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Maki
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
272
T. thiopara , deposits sulfur globules extracellularly [Cald-well et al., 1976], the other, T. azorensis , forms globules intracellularly when there is incomplete oxidation of thiosulfate and the pH is >7.0 [Odintsova et al., 1996]. In the genus Macromonas , the species M. bipunctata , forms sulfur globules in the periplasm when cells are grown in the presence of sulfides but not thiosulfate [Dubinina et al., 2005].
By far the majority of genera in which cultured bacte-ria produce intracellular sulfur globules are found in the γ-Proteobacteria ( fig. 1 ). This includes bacteria in genera like Thiothrix (e.g., fig. 2) [Bland and Staley, 1978; Konkol et al., 2010; Larkin, 1980], Beggiatoa [Strohl et al., 1981], Thiomargarita [Schulz et al., 1999] and Thioploca [Jør-gensen and Gallardo, 1999; Maier and Murray, 1965], some of the largest microorganisms found in nature. It also includes the purple sulfur bacteria in the family Chromatiaceae, but not the Ectothiorhodospiraceae which form extracellular sulfur globules [Frigaard and Dahl, 2009]. Endosymbiotic bacteria [Cavanaugh et al., 1981] found in invertebrates associated with hydrother-mal vents and sulfide gradients have also been indicated to be members of the γ-Proteobacteria [Distel, 1998; Stewart and Cava naugh, 2006].
Additional genera to those presented in figure 1 have been shown to produce internal sulfur globules. Primary among these is the genus Thiovulum . Members of this ge-nus have resisted attempts to get them in pure culture, but 16S rRNA gene analysis places them in the ε-Pro teo-bacteria and a genome analysis has been completed [Mar-shall et al., 2012]. Although no complete 16S rRNA se-quence has been reported for the genus Thiobacterium , a recent publication [Grünke et al., 2010] using fluorescence in situ hybridization indicated that this genus also belongs in the γ-Proteobacteria. However, not all sulfur globule-producing bacteria are in the Proteobacteria. Two genera, Thermonanerobacter and Thermoanaerobacterium in the Firmicutes have also been reported to produce internal sulfur globules [Lee et al., 2007]. A large spirillum, Titano-spirillum velox , that had internal sulfur globules has also been described [Guerrero et al., 1999]; however, the name has not been validated, no culture has been submitted to a collection and no 16S rRNA gene sequence has been sub-mitted to the Ribosomal Database Project or GenBank.
Although most of the described bacteria that construct intracellular sulfur globules are in the Proteobacteria, the fact that the vast majority of microbes in the world have yet to be isolated leads to the speculation that this ability could be more phylogenetically widespread than is cur-rently apparent.
Globule Protein Membranes
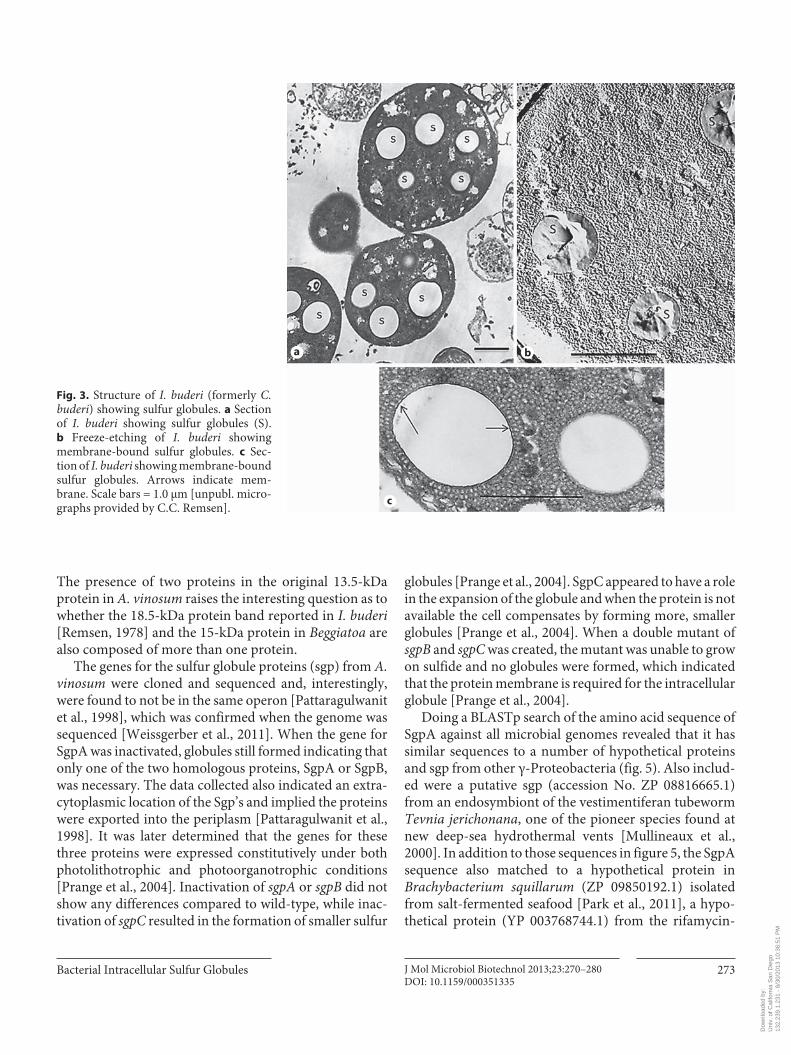
In light micrographs sulfur globules appear as translu-cent or clear areas ( fig. 2 ), while in electron micrographs sulfur globules appear as clear empty areas within thin sec-tions of cells ( fig. 3 a). Electron microscopy can be a bit of an art form and revealing cellular fine structures is some-times difficult depending on the processes of fixation, de-hydration and embedding. However, it has been reported from a variety of bacteria that intracellular sulfur globules are bounded by a protein membrane, e.g. Thiothrix [Bland and Staley, 1978], purple sulfur bacteria [Nicolson and Schmidt, 1971; Remsen, 1978, 1982; Remsen and Trüper, 1973], Beggiatoa [Strohl et al., 1981] and Thiovulum [Wir-sen and Jannasch, 1978]. This protein membrane can be observed on globules using freeze-etching, when it is in the correct plane ( fig. 3 b), and in thin sections using transmis-sion electron microscopy ( fig. 3 c). The proteins encasing the sulfur globules from most of these bacteria have yet to be characterized.
The vast majority of what is known about the protein membranes of sulfur globules comes from investigations involving purple sulfur bacteria in the family Chromatia-ceae. Initially, using centrifugation to isolate globules, ex-traction with chloroform to remove sulfur and retain the protein membrane, Schmidt et al. [1971] reported the pres-ence of a 13.5-kDa protein from the globule membrane from Allochromatium vinosum [formerly Chromatium vinosum , see Imhoff et al., 1998] . Similarly, the presence of a single 18.5-kDa protein from the sulfur globule in Iso-chromatium buderi [formerly C. buderi , Imhoff et al., 1998] was discovered [Gonye, Schroeder and Remsen, unpubl. data; reported by Remsen, 1978]. Outside of the Chroma-tiaceae, a 13-kDa, and more particularly, a 15-kDa protein were evident in sulfur inclusions from Beggiatoa when cells were grown in a medium with sulfide [Schmidt et al., 1986].
In the case of A. vinosum , subsequent analysis and sep-aration of the proteins using HPLC revealed that the 13.5-kDa protein was actually composed of two slightly small-er homologous proteins roughly 10.6 kDa in size (called SgpA and SgpB) [Brune, 1995]. In addition, a third pro-tein, 8.5 kDa (SgpC), was also found associated with the globules [Brune, 1995]. These three proteins make up the membrane of the sulfur globule ( fig. 4 ). Two proteins were also reported from the sulfur globules of Thiocapsa roseopersicina , the larger one approximately 10.6 kDa and the smaller one 8.7 kDa, and both homologous to the large and small proteins of A. vinosum , respectively [Brune, 1995]. These proteins were found to be similar to structural proteins in protein databases [Brune, 1995].
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM
Bacterial Intracellular Sulfur Globules J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
273
The presence of two proteins in the original 13.5-kDa protein in A. vinosum raises the interesting question as to whether the 18.5-kDa protein band reported in I. buderi [Remsen, 1978] and the 15-kDa protein in Beggiatoa are also composed of more than one protein.
The genes for the sulfur globule proteins (sgp) from A. vinosum were cloned and sequenced and, interestingly, were found to not be in the same operon [Pattaragulwanit et al., 1998], which was confirmed when the genome was sequenced [Weissgerber et al., 2011]. When the gene for SgpA was inactivated, globules still formed indicating that only one of the two homologous proteins, SgpA or SgpB, was necessary. The data collected also indicated an extra-cytoplasmic location of the Sgp’s and implied the proteins were exported into the periplasm [Pattaragulwanit et al., 1998]. It was later determined that the genes for these three proteins were expressed constitutively under both photolithotrophic and photoorganotrophic conditions [Prange et al., 2004]. Inactivation of sgpA or sgpB did not show any differences compared to wild-type, while inac-tivation of sgpC resulted in the formation of smaller sulfur
globules [Prange et al., 2004]. SgpC appeared to have a role in the expansion of the globule and when the protein is not available the cell compensates by forming more, smaller globules [Prange et al., 2004]. When a double mutant of sgpB and sgpC was created, the mutant was unable to grow on sulfide and no globules were formed, which indicated that the protein membrane is required for the intracellular globule [Prange et al., 2004].
Doing a BLASTp search of the amino acid sequence of SgpA against all microbial genomes revealed that it has similar sequences to a number of hypothetical proteins and sgp from other γ-Proteobacteria ( fig. 5 ). Also includ-ed were a putative sgp (accession No. ZP 08816665.1) from an endosymbiont of the vestimentiferan tubeworm Tevnia jerichonana , one of the pioneer species found at new deep-sea hydrothermal vents [Mullineaux et al., 2000]. In addition to those sequences in figure 5 , the SgpA sequence also matched to a hypothetical protein in Brachybacterium squillarum (ZP 09850192.1) isolated from salt-fermented seafood [Park et al., 2011], a hypo-thetical protein (YP 003768744.1) from the rifamycin-
a b
c
Fig. 3. Structure of I. buderi (formerly C. buderi ) showing sulfur globules. a Section of I. buderi showing sulfur globules (S). b Freeze-etching of I. buderi showing membrane-bound sulfur globules. c Sec-tion of I. buderi showing membrane-bound sulfur globules. Arrows indicate mem-brane. Scale bars = 1.0 μm [unpubl. micro-graphs provided by C.C. Remsen].
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Maki
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
274
producing actinomycete Amycolatopsis mediterranei , an integral membrane protein (ZP 07979829.1) from Strep-tomyces spp. and a hypothetical protein (ZP 21109042.1) from Streptomyces viridochromogenes .
A similar search with SgpB also revealed matches to a wide variety of hypothetical proteins and sgp from other microbes also found in the γ-Proteobacteria ( fig. 6 ). It also matched a hypothetical protein (ZP 02532721) found in an endosymbiont from the giant tubeworm Riftia pa-chyptila ( fig. 6 ) called Candidatus Endoriftia persephone [Robidart et al., 2008]. The SgpC amino acid sequence also matched to hypothetical proteins and sgp in other γ-Proteobacteria ( fig. 7 ).
Globule Sulfur Content
Even though both light and electron microscopy show globules associated with bacteria that oxidize reduced sulfur compounds, and that when the source of reduced sulfur disappears the globules reduce in size [Winograd-sky, 1889], the question remained as to whether the glob-ules were actually completely filled with sulfur [Steudel, 1989]. The process of dehydration during electron mi-
croscopy removes the sulfur from the globule [Remsen and Trüper, 1973]. Elemental sulfur (S 0 ) forms polymer-ic chains and rings, with cyclooctasulfur (S 8 ) being ther-modynamically the most stable form [Steudel, 1989].
X-ray diffraction was used to analyze sulfur from sul-fur globules in Thiovulum [LaRiviere, 1963] and photo-synthetic sulfur bacteria [Hageage et al., 1970; Trüper and Hathaway, 1967]. Freshly isolated, wet sulfur globules had diffraction patterns similar to that of liquid sulfur while after drying they converted into the cyclooctasulfur form [Hageage et al., 1970]. Analyses of dense inclusion bodies in air-dried filaments of Beggiatoa alba with ener-gy-dispersive X-ray microanalysis showed that the inclu-sions were composed of sulfur [Lawry et al., 1981] but could not designate the form of the sulfur, whether poly-meric chain or cyclooctasulfur. Energy-dispersive X-ray has also been used to identify sulfur-containing globules in magnetotactic bacteria [Keim et al., 2005].
One technique that has been effective in analysis of sulfur globules is a form of X-ray spectroscopy called X-ray absorption near-edge structure (XANES) spectrosco-py. For a description of XANES, the reader is referred to the review by Prange et al. [2008] and references therein. When purple sulfur bacteria were grown photoautotro-phically on sulfide the XANES spectra showed the sulfur in the globules with a structure of long sulfur chains (R-S n -R) with unidentified organic residues at one or both ends [Prange et al., 1999]. No sulfur rings were detected. When globules in Beggiatoa and Thioploca were com-pared to purple sulfur bacteria, the sulfur in the former were found to be dominated by cyclooctasulfur, while the purple sulfur bacteria contained sulfur chains [Prange et al., 2002]. However, using the XANES methodology and examining A. vinosum globules, George et al. [2008] re-ported the presence of cyclooctasulfur (S 8 ), not chains. These two research groups used different detection strat-egies which ultimately provided dissimilar results and conclusions. Further discussions on the controversy can be found in George et al. [2008] and Prange et al. [2008]. Other methods like Raman spectroscopy have also shown the sulfur globules in Thioploca and Beggiatoa to contain cyclooctasulfur (S 8 ) [Pasteris et al., 2001].
Storage/Formation
How does the bacterium go about forming the globule and storing sulfur? For A. vinosum , it is known that the proteins making up the membrane are essential for this to occur [Prange et al., 2004]. Members of the Chroma-
SgpB
SgpB
SgpBSgpB
SgpB
SgpB
SgpC
SgpC
SgpC
SgpC
SgpC
SgpC
SgpA
SgpA
SgpA
SgpA
SgpA
SgpA
S0
Fig. 4. Schematic diagram of sulfur globule membrane proteins identified from A. vinosum . This photosynthetic purple sulfur bac-terium forms transient globules in the periplasm and requires the membrane proteins. SgpA and SgpB are interchangeable and only one of these two proteins is necessary. SgpC is required for expan-sion of the globule. See text for details.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Bacterial Intracellular Sulfur Globules J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
275
tiaceae, C. okenii and A. warmingii , were observed using cinematography to form globules at any location within the cell [Hermann and Truper, unpubl. data; reported by Pattaragulwanit et al., 1998; Trüper, 2008]. The model or-ganism A. vinosum can form globules from sulfur (S 0 ), sulfides and thiosulfate ( fig. 8 ).
When using sulfur (S 0 ) to form globules, it was shown that A. vinosum required direct cell contact with the sul-fur and strongly preferred only polymeric sulfur (sulfur chain), not cyclooctasulfur [Franz et al., 2007]. Similarly, it was shown that when T. roseopersicina and two species of Halorhodospira also formed sulfur globules from ele-mental sulfur, the former intracellularly and the latter extracellularly, they also preferred uptake of chain-like polymers [Franz et al., 2009b]. Although cell contact with the sulfur is essential the mobilization of the sulfur does not appear to involve excretion of soluble sulfur-contain-ing substances [Franz et al., 2009a].
When sulfides were the source of sulfur for the sulfur globules in A. vinosum two primary enzymes were pos-sibly involved in the oxidation: a periplasmic flavocyto-chrome c, and a membrane-bound sulfide:quinone oxi-doreductase [Dahl and Prange, 2006; Frigaard and Dahl, 2009]. By constructing a flavocytochrome c-deficient mutant, it was shown that there was no effect on sulfide oxidation compared to the wild-type cells and pointed to an enhanced role for the sulfide:quinone oxidoreductase [Reinartz et al., 1998]. However, under certain growth conditions as yet unidentified, the flavocytochrome c may play a role [see discussion in Frigaard and Dahl, 2009].
The sulfur-oxidizing enzyme system (Sox) has been indicated to be important in the oxidation of thiosulfate to sulfate [Friedrich et al., 2005; Ghosh and Dam, 2009]. Bacteria that form intracellular sulfur globules do not possess all the genes necessary [Friedrich et al., 2005;
Fig. 5. Amino acid sequence of the SgpA from A. vinosum and the results of BLAST search of microbial genomes for similar protein sequences. Underlined amino ac-ids correspond to the sequence determined by Brune [1995] from the purified sgp. GenBank accession numbers are in paren-theses at the end of each sequence. hp = Hy-pothetical protein; psgp = putative sgp.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Maki
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
276
Hensen et al., 2006]. In A. vinosum , two independent loci were discovered which contained five sox genes, soxBXA and soxYZ , but lacked soxCD [Hensen et al., 2006]. Sul-fane sulfur binds to SoxY and the sulfur dehydrogenase encoded by soxCD is important for further oxidation.
Without soxCD , A. vinosum transfers the sulfane to the sulfur globule in the periplasm [Hensen et al., 2006]. In the absence of reduced sulfur compounds the sox genes are expressed at a low constitutive level [Grimm et al., 2011].
Fig. 6. Amino acid sequence of the SgpB from A. vinosum and the results of BLAST search of microbial genomes for similar protein sequences. Underlined amino ac-ids correspond to the sequence determined by Brune [1995] from the purified sgp. GenBank accession numbers are in paren-theses at the end of each sequence. hp = Hy-pothetical protein.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Bacterial Intracellular Sulfur Globules J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
277
Degeneration of Globules
The presence of the intracellular sulfur globules in these bacteria is transient and the sulfur deposited within the globules is eventually further oxidized to sulfate. As the sulfur is oxidized, the globules decrease in size. Intrin-sically involved with this oxidation are the dissimilatory sulfite reductase ( dsr ) genes. In A. vinosum , fifteen open reading frames were identified as dsr genes and designat-
ed as dsrABEFHCMKLJOPNRS [Dahl et al., 2005; Pott and Dahl, 1998] ( fig. 8 ). In the absence of reduced sulfur compounds the expression of most dsr genes was at a low basal level, but the expression was enhanced in the pres-ence of sulfide [Grimm et al., 2010]. Sulfate production was found to begin concurrently when the expression of the dsr genes was upregulated [Grimm et al., 2010].
The Dsr proteins are a combination of cytoplasmic and membrane-bound proteins. Carrier molecules,
SgpB
SgpB
SgpBSgpB
SgpB
SgpB
SgpC
SgpC
SgpC
SgpC
SgpC
SgpC
SgpA
SgpA
SgpA
SgpA
SgpA
SgpA
S0
S0
H2S
S2O32–
dsrABEFHCMKLJOPNRS
Flavocytochrome cSulfide:quinone oxidoreductase
SoxXABSoxXY
Fig. 7. Amino acid sequence of the SgpC from A. vinosum and the results of BLAST search of microbial genomes for similar protein sequences. Underlined amino ac-ids correspond to the sequence determined by Brune [1995] from the purified sgp. GenBank accession numbers are in paren-theses at the end of each sequence. hp = Hy-pothetical protein.
Fig. 8. Schematic diagram of the formation of sulfur globules in A. vinosum . This pho-tosynthetic purple sulfur bacterium forms transient sulfur globules from elemental sulfur (S 0 ) [Franz et al., 2007], sulfides (e.g. H 2 S) through flavocytochrome c (the phys-iological role is unresolved in A. vinosum ) and/or sulfide:quinone oxidoreductase [Dahl et al., 2008a], and thiosulfate (S 2 O 3 2– ) through SoxXAB and SoxXY [Grimm et al., 2008; Hensen et al., 2006]. The sulfur in the globules is ultimately oxi-dized to sulfate and the dsr proteins are es-sential in this process.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Maki
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
278
proposed as low-molecular-weight organic persulfides, transfer the sulfur from the globules to the Dsr proteins in A. vinosum [Frigaard and Dahl, 2009; Stockdreher et al., 2012]. Many of the dsr genes have been demonstrated to be essential to the conversion of globule sulfur to sul-fate [Dahl et al., 2008b; Pott and Dahl, 1998; Sander et al., 2006]. The proteins DsrEFH and DsrC are cytoplasmic and act as a relay system to transfer sulfur from a persul-fated carrier to DsrAB which is a dissimilatory sulfite re-ductase [Cort et al., 2008; Dahl et al., 2008b; Stockdreher et al., 2012]. DsrMJKOP is a transmembrane complex, all components of which have been demonstrated to be es-sential for the oxidation of globule sulfur in A. vinosum [Sander et al., 2006]. The DsrMJKOP is believed to be able to transfer electrons into the photosynthetic elec-tron transport [Grein et al., 2010; Weissgerber et al., 2011]. DsrS is suggested to be involved in posttranscrip-tional control of the dsr operon [Grimm et al., 2011]. Details of the role of dsrABEFHCMKLJOPNRS in the ox-idation of sulfur in sulfur globules to sulfate in A. vino-sum can be found in the references listed above and in reviews by Frigaard and Dahl [2009] and Ghosh and Dam [2009].
Due to the groundbreaking research on the model pur-ple sulfur bacterium A. vinosum by C. Dahl, her students and colleagues, there is an established, though in places incomplete, pathway for understanding the structure,
formation and degeneration of transient intracellular sul-fur globules. However, the development of other model organisms to investigate these pathways is also impor-tant. Fortunately, this is the era of genomics, transcrip-tomics and proteomics, which means that the similarities of the pathway in A. vinosum compared with other intra-cellular sulfur globule-forming bacteria are easier to de-tect and could point the direction to any differences. For example, genomic analysis revealed sox genes and dsr genes similar to A. vinosum in the genomes of Beggiatoa [Mussmann et al., 2007] and the uncultured Candidatus Ruthia magnifica [Newton et al., 2007], and transcrip-tome analysis of the endosymbiont of Solemya velum also showed a relationship to A. vinosum [Stewart et al., 2011]. Despite the fact that there will undoubtedly be common-ality between the physiological pathways in all the bacte-ria that form intracellular sulfur globules, there may be some interesting surprises.
Acknowledgements
The author is indebted to Prof. Emeritus Charles C. Remsen for providing unpublished data, electron micrographs, for stimulating discussions and comments on an earlier draft of the manuscript. The author also thanks Prof. Dennis Bazylinski for information on magnetotactic bacteria and Mr. Prince Mathai for his assistance in producing the phylogenetic tree.
References
Bazylinski DA, Dean AJ, Williams TJ, Long LK, Middleton SL, Dubbels BL: Chemolithoau-trophy in the marine, magnetotactic bacterial strains MV-1 and MV-2. Arch Microbiol 2004; 182: 373–387.
Bazylinski DA, Williams TJ: Ecophysiology of magnetotactic bacteria; in Schüler D (ed): Magnetoreception and Magnetosomes in Bacteria: Microbiology Monographs (3). Ber-lin, Springer, 2006, pp 37–75.
Bazylinski DA, Williams TJ, Lefevre CT, Trubit-syn D, Fang J, Beveridge TJ, Moskowitz BM, Ward B, Schübbe S, Dubbels BL, Simpson B: Magnetovibrio blakemorei , gen. nov. sp. nov., a new magnetotactic bacterium ( Alphaproteo-bacteria: Rhodospirillaceae ) isolated from a salt marsh. Int J Syst Evol Microbiol 2012, E-pub ahead of print.
Bland JA, Staley JT: Observations on the biology of Thiothrix . Arch Microbiol 1978; 117: 79–87.
Brune DC: Isolation and characterization of the sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina . Arch Microbiol 1995; 163: 391–399.
Caldwell DE, Caldwell SJ, Laycock P: Thermo-thrix thiopara gen. et sp. nov. A facultatively anaerobic facultative chemolithotroph living at neutral pH and high temperature. Can J Microbiol 1976; 22: 1509–1517.
Cavanaugh CC, Gardiner SL, Jones ML, Jannasch HW, Waterbury JB: Prokaryotic cells in the hydrothermal vent tube worm Riftia pachyp-tila Jones: possible chemoautotrophic symbi-onts. Science 1981; 213: 340–342.
Cort JR, Selan U, Schulte A, Grimm F, Kennedy MA, Dahl C: Allochromatium vinosum DsrC: solution-state NMR structure, redox proper-ties, and interaction with DsrEFH, a protein essential for purple sulfur bacterial sulfur oxi-dation. J Mol Biol 2008; 382: 692–707.
Dahl C, Engels S, Pott-Sperling AS, Schulte A, Sander J, Lübbe Y, Deuster O, Brune DC: Novel genes of the dsr gene cluster and evi-dence for close interaction of Dsr proteins during sulfur oxidation in the phototrophic sulfur bacterium Allochromatium vinosum . J Bacteriol 2005; 187: 1392–1404.
Dahl C, Friedrich C, Kletzin A: Sulfur oxidation in prokaryotes; in Encyclopedia of Life Sci-ences. Chichester, John Wiley & Sons, 2008a, pp 1–11.
Dahl C, Prange A: Bacterial sulfur globules: oc-currence, structure and metabolism; in Shive-ly JM (ed): Inclusions in Prokaryotes: Micro-bial Monographs. Berlin, Springer, 2006, pp 21–51.
Dahl C, Schulte A, Stockdreher Y, Hong C, Grimm F, Sander J, Kim R, Kim S-H, Shin DH: Structural and molecular genetic insight into a widespread sulfur oxidation pathway. J Mol Biol 2008b;384: 1287–1300.
Distel DL: Evolution of chemoautotrophic endo-symbioses in bivalves. BioScience 1998; 48: 277–286.
Dubinina GA, Rainey FA, Kuenen JG: Genus VII: Macromonas Utermöhl and Koppe in Koppe 1924, 632 AL ; in Brenner DJ, Krieg NR, Staley JT, Garrity G (eds): Bergey’s Manual of Sys-tematic Bacteriology, ed 2. New York, Spring-er, 2005, vol 2, part C, pp 721–724.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Bacterial Intracellular Sulfur Globules J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
279
Franz B, Gehrke T, Lichtenberg H, Hormes J, Dahl C, Prange A: Unexpected extracellular and in-tracellular species during growth of Allochro-matium vinosum with reduced sulfur com-pounds. Microbiology 2009a;155: 2766–2774.
Franz B, Lichtenberg H, Dahl C, Hormes J, Prange A: Utilization of ‘elemental’ sulfur by differ-ent phototrophic sulfur bacteria ( Chromatia-ceae, Ectothiorhodospiraceae ): a sulfur K-edge XANES spectroscopy study. J Physics 2009b;Conf Ser 190: 012200.
Franz B, Lichtenberg H, Hormes J, Modrow H, Dahl C, Prange A: Utilization of solid ‘ele-mental’ sulfur by the phototrophic purple sul-fur bacterium Allochromatium vinosum : a sulfur K-edge X-ray absorption spectroscopy study. Microbiology 2007; 153: 1268–1274.
Friedrich CG, Bardischewsky F, Rother D, Quent-meier A, Fischer J: Prokaryotic sulfur oxida-tion. Curr Opin Microbiol 2005; 8253–8259.
Frigaard N-U, Dahl C: Sulfur metabolism in pho-totrophic sulfur bacteria. Adv Microbial Physiol 2009; 54: 103–200.
George GN, Gnida M, Bazylinski DA, Prince RC, Pickering IJ: X-ray absorption spectroscopy as a probe of microbial sulfur biochemistry: the nature of bacterial sulfur globules revisit-ed. J Bacteriol 2008; 190: 6376–6383.
Ghosh W, Dam B: Biochemistry and molecular biology of lithotropic sulfur oxidation by tax-onomically and ecologically diverse bacteria and archaea. FEMS Microbiol Rev 2009; 33: 999–1043.
Gottschalk G: Bacterial Metabolism, ed 2. New York, Springer, 1986.
Grein F, Pereira IAC, Dahl C: Biochemical char-acterization of individual components of the Allochromatium vinosum DsrMKJOP trans-membrane complex aids understanding of complex function in vivo. J Bacteriol 2010; 192: 6369–6377.
Grimm F, Dobler N, Dahl C: Regulation of dsr genes encoding proteins responsible for the oxidation of stored sulfur in Allochromatium vinosum . Microbiology 2010; 156: 764–773.
Grimm F, Franz B, Dahl C: Thiosulfate and sulfur oxidation in purple sulfur bacteria; in Dahl C, Friedrich CG (eds): Microbial Sulfur Metabo-lism. Berlin, Springer, 2008, pp 101–116.
Grimm F, Franz B, Dahl C: Regulation of dissim-ilatory sulfur oxidation in the purple sulfur bacterium Allochromatium vinosum . Front Microbiol 2011; 2: 51.
Grünke S, Lichtschlag A, de Beer D, Kuypers M, Lösekann-Behrens T, Ramette A, Boetius A: Novel observations of Thiobacterium , a sul-fur-storing Gammaproteobacterium produc-ing gelatinous mats. ISME Journal 2010; 4: 1031–1043.
Guerrero R, Haselton A, Solé M, Wier A, Margu-lis L: Titanospirillum velox : a huge, speedy, sulfur-storing spirillum from Ebro Delta mi-crobial mats. Proc Natl Acad Sci USA 1999; 96: 11584–11588.
Hageage GJ, Jr, Eanes ED, Gherna RL: X-ray dif-fraction studies of the sulfur globules accu-mulated by Chromatium species. J Bacteriol 1970; 101: 464–469.
Hensen D, Sperling D, Trüper HG, Brune DC, Dahl C: Thiosulphate oxidation in the photo-trophic sulphur bacterium Allochromatium vinosum. Mol Microbiol 2006; 62: 794–810.
Imhoff JF, Süling J, Petri R: Phylogenetic rela-tionships among the Chromatiaceae , their taxonomic reclassification and description of the new genera Allochromatium, Halochro-matium, Isochromatium, Marichromatium, Thiococcus, Thiohalocapsa and Thermop-chromatium . Int J Syst Bacteriol 1998; 48: 1129–1143.
Jørgensen BB, Gallardo VA: Thioploca spp.: fila-mentous sulfur bacteria with nitrate vacuoles. FEMS Microbiol Ecol 1999; 28: 301–313.
Keim CN, Solorzano G, Farina M, Lins U: Intra-cellular inclusions of uncultured magnetotat-ic bacteria. Int Microbiol 2005; 8: 111–117.
Konkol NR, Bruckner JC, Aguilar C, Lovalvo D, Maki JS: Dominance of epiphytic filamentous Thiothrix spp. on an aquatic macrophyte in a hydrothermal vent flume in Sedge Bay, Yel-lowstone Lake, Wyoming. Microb Ecol 2010; 60: 528–538.
La Rivière JWM: Cultivation and properties of Thiovulum majus Hinze; in Oppenheimer CH (ed): Marine Microbiology. Springfield, CC Thomas Publisher, 1963, pp 61–72.
Larkin JM: Isolation of Thiothrix in pure culture and observation of a filamentous epiphyte on Thiothrix . Curr Microbiol 1980; 4: 155–158.
Lawry NH, Jani V, Jensen TE: Identification of the sulfur inclusion body in Beggiatoa alba B18LD by energy-dispersive x-ray microanal-ysis. Curr Microbiol 1981; 6: 71–74.
Lee Y-J, Prange A, Lichtenberg H, Rohde M, Dashti M, Wiegel J: In situ analysis of sulfur species in sulfur globules produced from thio-sulfate by Thermoanaerobacter sulfurigignens and Thermoanaerobacterium thermosulfuri-genes . J Bacteriol 2007; 189: 7525–7529.
Lefevre CT, Viloria N, Schmidt ML, Posfai M, Frankel RB, Bazylinski DA: Novel magnetite-producing magnetotactic bacteria belonging to the Gammaproteobacteria . ISME J 2012; 6: 440–450.
Maier S, Murray RGE: The fine structure of Thio-ploca ingrica and a comparison with Beggia-toa. Can J Microbiol 1965; 11: 645–655.
Marshall IPG, Blainey PC, Spormann AM, Quake SR: A single-cell genome for Thiovulum sp. Appl Environ Microbiol 2012; 78: 8555–8563.
Mullineaux LS, Fisher CR, Peterson CH, Schaeffer SW: Tubeworm succession at hydrothermal vents: use of biogenic cues to reduce habitat selection error? Oecologia 2000; 123: 275–284.
Mussmann M, Hu FZ, Richter M, de Beer D, Preisler A, Jørgensen BB, Huntemann M, Glöckner FO, Amann R, Koopman WJH, Lasken RS, Janto B, Hogg J, Stoodley P, Boissy R, Ehrlich GD: Insights into the genome of large sulfur bacteria revealed by analysis of single filaments. PloS Biology 2007; 5:e230.
Newton ILG, Woyke, Auchtung TA, Dilly GF, Dutton RJ, Fisher MC, Fontanez KM, Lau E, Stewart FJ, Richardson PM, Barry KW, Saun-ders E, Detter JC, Wu D, Eisen JA, Cananaugh CM: The Calyptogena magnifica chemoauto-trophic symbiont genome. Science 2007; 315: 998–1000.
Nicolson GL, Schmidt GL: Structure of the Chro-matium sulfur particle and its protein mem-brane. J Bacteriol 1971; 105: 1142–1148.
Odintsova EV, Jannasch HW, Mamone JA, Lang-worthy TA: Thermothrix azorensis sp. nov., an obligately chemoautotrophic, sulfur-oxi-dizing, thermophilic bacterium. Int J Syst Bacteriol 1996; 46: 422–428.
Park S-K, Kim M-S, Jung M-J, Nam Y-D, Park E-J, Roh SW, Bae J-W: Brachybacterium squillarum sp. nov., isolated from salt-fer-mented seafood. Int J Syst Evol Microbiol 2011; 61: 1118–1122.
Pasteris JD, Freeman JJ, Goffredi SK, Buck KR: Raman spectroscopic and laser scanning con-focal microscopic analysis of sulfur in living sulfur-precipitating marine bacteria. Chem Geol 2001; 180: 3–18.
Pattaragulwanit K, Brune DC, Trüper HG, Dahl C: Molecular genetic evidence for extracyto-plasmic localization of sulfur globules in Chromatium vinosum . Arch Microbiol 1998; 169: 434–444.
Pott AS, Dahl C: Sirohaem sulfite reducatase and other proteins encoded by genes of the dsr lo-cus of Chromatium vinosum are involved in the oxidation of intracellular sulfur. Microbi-ology 1998; 144: 1881–1894.
Prange A, Arzberger I, Engemann C, Modrow H, Schumann O, Trüper HG, Steudel R, Dahl C, Hormes J: In situ analysis of sulfur in the sul-fur globules of phototrophic sulfur bacteria by x-ray absorption near edge spectroscopy. Bio-chim Biophys Acta 1999; 1428: 446–454.
Prange A, Chauvistré R, Modrow H, Hormes J, Trüper HG, Dahl C: Quantitative speciation of sulfur in bacterial sulfur globules: x-ray ab-sorption spectroscopy reveals at least three different species of sulfur. Microbiology 2002; 148: 267–276.
Prange A, Engelhardt H, Trüper HG, Dahl C: The role of the sulfur globule proteins of Allochro-matium vinosum : mutagenesis of the sulfur globule protein genes and expression studies by real-time RT-PCR. Arch Microbiol 2004; 182: 163–174.
Prange A, Hormes J, Modrow H: X-ray absorp-tion spectroscopy as tool for the detection and identification of sulfur compounds in photo-trophic organisms; in Hell R, Dahl C, Knaff D, Leustek T (eds): Sulfur Metabolism in Photo-trophic Organisms: Advances in Photosyn-thesis and Respiration. Dordrecht, Springer, 2008, pp 461–482.
Reinartz M, Tschäpe J, Brüser T, Trüper HG, Dahl C: Sulfide oxidation in the phototrophic sulfur bacterium Chromatium vinosum. Arch Microbiol 1998; 170: 59–68.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM

Maki
J Mol Microbiol Biotechnol 2013;23:270–280 DOI: 10.1159/000351335
280
Remsen CC: Comparative subcellular architec-ture of photosynthetic bacteria; in Clayton RK, Sistrom WR (eds): The Photosynthetic Bacteria. New York, Plenum Press, 1978, pp 31–60.
Remsen CC: Structural attributes of membranous organelles in bacteria. Int Rev Cytol 1982; 76: 195–223.
Remsen CC, Trüper HG: The fine structure of Chromatium buderi . Arch Mikrobiol 1973; 90: 269–280.
Robidart JC, Bench SR, Feldman RA, Novora-dovsky A, Podell SB, Gaasterland T, Allen EE, Felbeck H: Metabolic versatility of the Riftia pachyptila endosymbiont revealed through metagenomics. Environ Microbiol 2008; 10: 727–737.
Sander J, Engels-Schwarzlose S, Dahl C: Impor-tance of the DsrMKJOP complex for sulfur oxidation in Allochromatium vinosum and phylogenetic analysis of related complexes in other prokaryotes. Arch Microbiol 2006; 186: 357–366.
Schmidt GL, Nicolson GL, Kamen MD: Compo-sition of the sulfur particle of Chromatium vinosum Strain D. J Bacteriol 1971; 105: 1137–1141.
Schmidt TM, Vinci VA, Strohl WR: Protein syn-thesis by Beggiatoa alba B18LD in the pres-ence and absence of sulfide. Arch Microbiol 1986; 144: 158–162.
Schulz HN, Brinkhoff T, Ferdelman TG, Mariné MH, Teske A, Jørgensen BB: Dense popula-tions of a giant sulfur bacterium in Namibian shelf sediments. Science 1999; 284: 493–495.
Shively JM: Prokaryote inclusions: descriptions and discoveries; in Shively JM (ed): Inclusions in Prokaryotes. Berlin, Springer, 2006, pp 3–20.
Steudel R: On the nature of the ‘elemental sulfur’ (S 0 ) produced by sulfur-oxidizing bacteria – a model for S 0 globules; in Schlegel HG, Bowien B (eds): Autotrophic Bacteria. Madison, Sci-ence Tech, 1989, pp 289–303.
Stewart FJ, Cavanaugh CM: Bacterial endosym-bioses in Solemya (Mollusca: Bivalvia) – mod-el systems for studies of symbiont-host adap-tation. Ant van Leeuwen 2006; 90: 343–360.
Stewart FJ, Dmytrenko O, DeLong EF, Cava-naugh CM: Metatranscriptomic analysis of sulfur oxidation genes in the endosymbiont of Solemya velum . Front Microbiol 2011; 2: 134.
Stockdreher Y, Venceslau SS, Josten M, Sahl H-G, Pereira IAC, Dahl C: Cytoplasmic sulfur-transferases in the purple sulfur bacteriumAllochromatium vinosum: evidence for sulfur transfer from DsrEFH to DsrC. PLoS ONE 2012;7:e40785.
Strohl WR, Geffers I, Larkin JM: Structure of the sulfur inclusion envelopes from four Beggia-toas . Curr Microbiol 1981; 6: 75–79.
Trüper HG: Sulfur and light? History and ‘thiol-ogy’ of the phototrophic sulfur bacteria; in Dahl C, Freidrich CG (eds): Microbial Sulfur Metabolism. Berlin, Springer, 2008, pp 87–100.
Trüper HG, Hathaway JC: Orthorhombic sulphur formed by photosynthetic sulphur bacteria. Nature 1967; 215: 435–436.
van Niel CB: On the morphology and physiology of the purple and green sulphur bacteria. Arch Mikrobiol 1931; 3: 1–114.
Waksman SA: Microorganisms concerned in the oxidation of sulfur in the soil. I. Introductory. J Bacteriol 1922; 7: 231–238.
Waksman SA, Joffe JS: Microorganisms con-cerned in the oxidation of sulfur in the soil. II. Thiobacillus thiooxidans , a new sulfur-oxidiz-ing organism isolated from the soil. J Bacte-riol 1922; 7: 239–256.
Weissgerber T, Zigann R, Bruce D, Chang Y-J, Detter JC, Han C, Hauser L, Jeffries CD, Land M, Munk AC, Tapia R, Dahl C: Complete ge-nome sequence of Allochromatium vinosum DSM 180 T . Stand Genomic Sci 2011; 5: 311–330.
Williams TJ, Lefevre CT, Zhao W, Beveridge TJ, Bazylinski DA: Magnetospira thiophila gen. nov., sp. nov., a marine magnetotactic bacte-rium that represents a novel lineage within the Rhodospirillaceae ( Alphaproteobacteria ). Int J Syst Evol Microbiol 2012; 62: 2443–2450.
Williams TJ, Zhang CL, Scott JH, Bazylinski DA: Evidence for autotrophy via the reverse tricar-boxylic acid cycle in the marine magnetotac-tic coccus strain MC-1. Appl Environ Micro-biol 2006; 72: 1322–1329.
Winogradsky S: Recherches physiologiques sur les sulfobactéries. Ann Inst Pasteur 1889; 3: 49–60 (English translation: Physiological studies on the sulfur bacteria; in Brock TD (ed): Milestones in Microbiology. Washing-ton, American Society for Microbiology, 1975, pp 227–231).
Wirsen CO, Jannasch HW: Physiological and morphological observations on Thiovulum sp. J Bacteriol 1978; 136: 765–774.
Dow
nloa
ded
by:
Uni
v. o
f Cal
iforn
ia S
an D
iego
13
2.23
9.1.
231
- 8/
30/2
013
10:3
6:51
PM
Top Related