University of Zurich - zora.uzh.ch fileExpression and putative functions of GDF-15, a member of the...
59
University of Zurich Zurich Open Repository and Archive Winterthurerstr. 190 CH-8057 Zurich http://www.zora.uzh.ch Year: 2008 Expression and putative functions of GDF-15, a member of the TGF-β superfamily, in human glioma and glioblastoma cell lines Strelau, J; Schmeer, C; Peterziel, H; Sackmann, T; Herold-Mende, C; Steinert, H; Weller, M; Unsicker, K Strelau, J; Schmeer, C; Peterziel, H; Sackmann, T; Herold-Mende, C; Steinert, H; Weller, M; Unsicker, K (2008). Expression and putative functions of GDF-15, a member of the TGF-β superfamily, in human glioma and glioblastoma cell lines. Cancer Letters, 270(1):30-39. Postprint available at: http://www.zora.uzh.ch Posted at the Zurich Open Repository and Archive, University of Zurich. http://www.zora.uzh.ch Originally published at: Cancer Letters 2008, 270(1):30-39.
Transcript of University of Zurich - zora.uzh.ch fileExpression and putative functions of GDF-15, a member of the...

University of ZurichZurich Open Repository and Archive
Winterthurerstr. 190
CH-8057 Zurich
http://www.zora.uzh.ch
Year: 2008
Expression and putative functions of GDF-15, a member of theTGF-β superfamily, in human glioma and glioblastoma cell lines
Strelau, J; Schmeer, C; Peterziel, H; Sackmann, T; Herold-Mende, C; Steinert, H;Weller, M; Unsicker, K
Strelau, J; Schmeer, C; Peterziel, H; Sackmann, T; Herold-Mende, C; Steinert, H; Weller, M; Unsicker, K (2008).Expression and putative functions of GDF-15, a member of the TGF-β superfamily, in human glioma andglioblastoma cell lines. Cancer Letters, 270(1):30-39.Postprint available at:http://www.zora.uzh.ch
Posted at the Zurich Open Repository and Archive, University of Zurich.http://www.zora.uzh.ch
Originally published at:Cancer Letters 2008, 270(1):30-39.
Strelau, J; Schmeer, C; Peterziel, H; Sackmann, T; Herold-Mende, C; Steinert, H; Weller, M; Unsicker, K (2008).Expression and putative functions of GDF-15, a member of the TGF-β superfamily, in human glioma andglioblastoma cell lines. Cancer Letters, 270(1):30-39.Postprint available at:http://www.zora.uzh.ch
Posted at the Zurich Open Repository and Archive, University of Zurich.http://www.zora.uzh.ch
Originally published at:Cancer Letters 2008, 270(1):30-39.

Expression and putative functions of GDF-15, a member of theTGF-β superfamily, in human glioma and glioblastoma cell lines
Abstract
Recent studies have demonstrated growth-inhibiting effects of growth differentiation factor-15(GDF-15) on different cancer cell lines invitro and on tumor growth in vivo. Here, we present dataconcerning expression of GDF-15 in glioblastoma. We found low levels of GDF-15 transcripts inprimary glioblastoma. Thus, GDF-15 expression might be exploited as a useful indicator fordistinguishing primary from other glial derived tumors. In contrast to the documented proapoptotic andanti-tumorigenic activities of GDF-15 in several cancer cell lines, our data suggest that GDF-15 does notdecrease proliferation of glioblastoma cell lines, while its effects on invasiveness are not consistent.

Elsevier Editorial System(tm) for Cancer Letters
Manuscript Draft
Manuscript Number: CAN-D-07-01136R1
Title: EXPRESSION AND PUTATIVE FUNCTIONS OF GDF-15, A MEMBER OF THE TGF-ß
SUPERFAMILY, IN HUMAN GLIOMA AND GLIOBLASTOMA CELL LINES
Article Type: Original Research Paper
Section/Category:
Keywords: GDF-15; TGF-ß superfamily; glioblastomas; tumor
Corresponding Author: Dr. Jens Strelau, Ph.D.
Corresponding Author's Institution:
First Author: Jens Strelau, Dr.
Order of Authors: Jens Strelau, Dr.; Corina Schmeer, Dr.; Heike Peterziel, Dr.; Tina Paech; Christel Herold-
Mende; Hans H Steiner, Prof. Dr. ; Michael Weller, Prof. Dr.; Klaus Unsicker, Prof. Dr.
Manuscript Region of Origin:
Response to Reviewers: Response to Reviewers:
Reviewer 1
1. First this subtitle "GDF-15 is differentially expressed in cultures of primary and secondary glioblastoma" is
misleading.
We changed the subtitle into “Low expression of GDF-15 in cultures of primary glioblastomas” and present
in the subsequent chapter data on diffuse astrocytomas grade II, anaplastic astrocytomas grade III,
secondary glioblastomas grade IV, primary glioblastomas grade IV, oligodendroglioblastomas grade II and
III, and oligoastrocytomas grade II and III.

We also changed the first subtitle of the Results part into “GDF-15 immunoreactivity in de novo glioblastoma,
high grade astrocytoma and secondary glioblastoma” and subdivided figure 1 into A and B for the sake of
clarity of the immunohistochemical results.
2. No details are given about how these cultures were derived, maintained or what passage numbers were
used in the present studies.
Details are now provided in the Materials and Methods section on how the cultures were derived.
Furthermore, data on GDF-15 expression in oligodendrogliomas grade II and III and oligoastrocytomas
grade II and III were added to figure 2 and Results. We also incorporated a table comprising tumor type,
passage numbers, age and sex of each specimen (Tab.1).
4. Did the authors conduct similar studies on the primary cell lines used in Figure 2?
Figure 2 shows real-time PCR results with cDNA of 59 different primary cell cultures with low passage
numbers provided by the Molecular Biology Laboratories, Department of Neurosurgery, University of
Heidelberg. Since cell cultures with similarly low passage numbers were not available we did not perform
invasion-, proliferation-, and TUNEL-assays on primary cell lines.
5. How do the authors explain the differences in responsiveness of the different cell lines?
Our studies show that the GDF-15 antiserum decreases proliferation in 8 of 9 cell cultures. Exogenous GDF-
15 increased cell division in 4 cell lines, while the remaining 5 cell lines did not respond. Several possibilities
may be conceived to explain this result. First, cell lines unresponsive to exogenous GDF-15, but responsive
to the anti-GDF-15 might already be maximally stimulated by the endogenous GDF-15. Moreover, non-
responsive cell lines might require, in addition to GDF-15, a synergistically acting factor, which was not
available at the required amounts. Alternatively, GDF-15 receptors in non-responsive cell lines might already
be maximally saturated with endogenous GDF-15. A similar explanation might account for the lack of
response to exogenous GDF-15 in the invasion experiments. It should be noted, however, that both U87MG
and, to a lesser extent, LN-428 and LN-229, showed a tendency to respond to GDF-15.
It has been shown that, dependent on tumor type and origin (for review see Eling at al., 2006; Bauskin et al.,
2006), GDF-15 can exhibit either tumorigenic or anti-tumorigenic activity. The cell lines investigated in our
studies are derived from tumors of different stages and origin, and it is therefore conceivable that GDF-15
may regulate in either a positive or negative manner, depending on molecular and cellular contexts.
Antagonistic effects have also been shown for TGF � and its effects on different glioma cell lines (see also
Jennings et al., 1991).
Although previous studies suggested that GDF-15 can regulate invasiveness in cancer cell lines, results of
these former studies were based on a single cell line, without comparing the results with other cell lines

derived from the same cancer types. It has been shown, for example, that GDF-15 significantly increases
invasiveness of SNU-216, a human gastric cancer cell line (Lee et al., 2003). Evidently, this does not prove
that all available gastric cancer cell lines would respond in the same way. Our study compares GDF-15
dependent invasion in 8 different cancer cell lines, which may differ in a variety of subtle respects.
6. Lastly, no effect on apoptosis was noted. How do the authors explain this result, which is different from
previously published studies?
Most in vitro studies suggest GDF-15- induces apoptosis in cancer cell lines. However, data from the
literature provide high expression of GDF-15 in many tumors, a finding that is hard to be reconciled with the
proposed tumor suppressor activity of GDF-15. Previous studies from our laboratory clearly demonstrate
that GDF-15 can protect cerebellar granule cells from apoptotic cell death in vitro (Subramaniam et al.,
2003), i.e. GDF-15 may function as an anti-apoptotic factor. To our knowledge, none of the previously
published studies has shown pro-apoptotic effects of GDF-15 in glioma cultures.
7. The conclusions of this study are overstated.
Conclusions have been changed
Reviewer 2
1. How are primary and secondary GBM differentiated (clinically or molecular).
Primary and secondary GBM were differentiated as described earlier [22] In brief, tumors were
histopathologically classified according to the classification of the WHO. Tumors were characterized as
secondary GBM in patients with previous identified low grade (WHO II and WHO III) glioblastomas in
accordance with Kleihues.
2. Why was no fresh material used for qPCR?
We have now included fresh material provided by the Molecular Biology Laboratories, Department of
Neurosurgery, University of Heidelberg. We got 9 frozen tissue samples of primary GBM and used the
samples for qPCR and immunohistochemistry. The results of the qPCR with fresh primary GBM confirmed
data obtained by IHC (low GDF-15 expression in 8 out of 9 tumors, (see also the figure on the last page of
our revision notes). Three of the primary cultures used for qPCR correspond to fresh tissue samples (Tab.1).
Since one secondary GBM, two anaplastic astrocytoma and one oligodendroblastoma were provided as
frozen sections on coverslips we could not perform qPCR and used the material directly for

immunohistochemistry. However, primary cells derived from these tumors revealed high amounts of GDF-15
mRNA (Tab.1), although we were not able to show any ir in the oligodendroglioma grade III.
3. What is a low-passage culture and how was the immunohisto / qPCR correlation?
We added a list with tumor number, passage number, sex and age of the total material used in this study
(Tab. 1).
4. Table 2 should be presented earlier in the results section.
The correlation of p53 and GDF-15 is now mentioned in the Results section.
5. Discussion should be shortened.
We now shortened the discussion.
Literature:
Asne R. Bauskin et al., Role of Macrophage Inhibitory Cytokine-1 in Tumorigenesis and Diagnosis of Cancer.
Cancer Res. 2006; 66: (10). May 15, 4983-4986
Thomas E. Eling et al., NSAID Activated Gene (NAG-1), a Modulator of Tumorigenesis. J Biochem. and Mol.
Biol. 2006; 39: (6). Nov. 649-655
Mark T. Jennings et al., TGF-ß1 and TGF-ß2 are potential growth regulators for low-grade and malignant
gliomas in vitro: Evidence in support of an autocrine hypothesis. Int. J. Cancer 1991; 49, 129-139)

1
Cancer Letters
EXPRESSION AND PUTATIVE FUNCTIONS OF GDF-15, A MEMBER
OF THE TGF-ß SUPERFAMILY, IN HUMAN GLIOMA AND
GLIOBLASTOMA CELL LINES
Jens Strelau 1, Corina Schmeer 1, Heike Peterziel 1, Tina Paech 1, Christel Herold-
Mende 2, Hans Steiner 3, Michael Weller 4 and Klaus Unsicker 1
1Department of Neuroanatomy and Interdisciplinary Center for Neurosciences (IZN)
University of Heidelberg, Germany2 Molecular Biology Laboratories, Department of Neurosurgery, University of Heidelberg,
Germany 3 Department of Neurosurgery, Nürnberg Süd, Nürnberg, Germany4 Department of Neurology, University Hospital Zurich, Switzerland.
Address for correspondence:
Dr. Jens Strelau
Neuroanatomy & Interdisciplinary Center for Neurosciences (IZN),
University of Heidelberg,
INF 307
D-69120 Heidelberg, Germany
Fax: 49-6221-54 56 04
e-mail: [email protected]
Manuscript

2
Abstract
Recent studies have demonstrated growth-inhibiting effects of Growth differentiation factor-
15 (GDF-15) on different cancer cell lines in vitro and on tumor growth in vivo. Here, we
present data concerning expression of GDF-15 in glioblastoma. We found low levels of GDF-
15 transcripts in primary glioblastoma. Thus, GDF-15 expression might be exploited as a
useful indicator for distinguishing primary from other glial derived tumors. In contrast to the
documented proapoptotic and anti-tumorigenic activities of GDF-15 in several cancer cell
lines, our data suggest that GDF-15 does not decrease proliferation of glioblastoma cell lines,
while its effects on invasiveness are not consistent.
Key words: GDF-15, TGF- superfamily, glioblastomas, tumor

3
Introduction
Growth differentiation factor-15 (GDF-15), also known as macrophage inhibitory cytokine-1
[1], prostate-derived factor [2], placental bone morphogenetic protein [3], and nonsteroidal
anti-inflammatory drug-activated protein-1 [4], is a divergent member of the TGF-ß
superfamily and widely expressed in epithelial tissues and in brain [5, 6, 7].
Most recently, several reports have provided evidence for functional links between GDF-15
and factors involved in tumorigenesis. The tumor suppressor gene product p53, for instance,
strongly induces GDF-15, indicating for the first time a functional connection between p53
and the TGF-ß superfamily [8-12]. Nonsteroidal anti-inflammatory drugs (NSAIDs) that
inhibit tumor development can also increase GDF-15 expression [13]. Other studies have
documented increased levels of GDF-15 in serum and in tumors of patients with metastatic
cancer [14-18]. Perhaps most excitingly, GDF-15 has been reported to exert proapoptotic and
anti-tumorigenic functions on human colorectal, prostate, breast, and mammary cancer cells
in vitro and on colon and glioblastoma tumors in vivo [4, 8, 19-21]. Together, these findings
strongly suggest a putative role of GDF-15 in tumor development and/or progression and
underscore the importance of further exploration of an involvement of GDF-15 in
tumorigenesis.
We report here the expression of GDF-15 in human glioblastoma tissues, glioblastoma
primary cultures, and glioblastoma cell lines. Our analyses reveal that GDF-15 may be
implicated in the regulation of proliferation and invasiveness of human glioblastoma cells.

4
Materials and Methods
Samples and Cell culture
Tissue specimens were obtained intraoperatively from central parts of the tumors. Primary
and secondary GBM were histopathologically classified according to the classification of the
WHO. Tumors were characterized as secondary GBM in patients with previous identified low
grade (WHO II and WHO III) glioblastomas. Primary cultures of glioma tissues (Tab. 1) were
established as described earlier [22]. The human glioma cell lines A172, D247MG, LN-18,
LN-229, LN-308, LN-319, and LN-428 (see Tab. 2) were maintained in DMEM with 4.5 g
glucose/L supplemented with 10% fetal bovine serum (FBS), penicillin and streptomycin.
Cultures were maintained in a humidified 5% CO2 atmosphere at 37oC.
GDF-15 immunohistochemistry
Samples of primary and secondary glioblastomas were obtained from surgical cases in the
Department of Neurosurgery, University of Heidelberg, Germany. Snap frozen samples were
embedded in Tissue-Tek (Sakura, Ramsey, MI, USA) and directly cut into 16µm slices. For
immunohistochemistry, sections were fixed in 4% PFA for 1 hour. Non-specific peroxidase
activity was blocked by incubating sections in 1% hydrogen peroxide in PBS for 10 min at
room temperature (RT). Non-specific antibody binding was blocked using blocking buffer
(5% goat serum and 0.1% Triton X-100 in PBS) for 1 hour at RT. After washing, coverslips
were incubated with rabbit anti-mouse GDF-15 polyclonal antibodies (affinity-purified and
diluted 1:200) or with PBS overnight at 4oC. Slides were subsequently washed 3 times for 5
min in PBS and incubated 30 min at RT with biotinylated goat anti-rabbit IgG (Vectastain
Elite ABC Kit, Vector Laboratories, Burlingame, CA, USA). All subsequent steps were
performed according to the manufacturer’s instructions. Slides were counterstained with
hematoxylin, coverslipped, and photographed using a Zeiss Axioplan 2 equipped with

5
AxioVision 3.1 software. Immunofluorescence was performed as described [23]. Primary
antibodies used were affinity purified polyclonal anti-GDF-15, monoclonal anti-CD68 (BD
Biosciences, San Jose, CA, USA), and monoclonal anti-GFAP (Sigma, Chicago, IL, USA).
Secondary antibodies were anti-mouse IgG Cy2 and anti-rabbit IgG Cy3 (Jackson
ImmunoResearch, West Baltimore Pike West Grove, PA, USA)
Gel electrophoresis and immunoblot analyses
Protein extracts were prepared by homogenizing the cells in electrophoresis sample buffer,
and the protein content was determined using densitometry [24]. 25µg protein extract per lane
were loaded on SDS-polyacrylamide gels and transferred to nitrocellulose membranes
(Hybond ECL, Amersham Pharmacia, Göttingen, Germany) by electroblotting. The
membranes were incubated with purified polyclonal rabbit anti-rat GDF-15/MIC-1 antibody
for 16 h at 4 °C. Bound antibody was detected with a peroxidase-conjugated secondary
antibody and the ECL western blotting substrate system (Amersham Pharmacia) according to
the manufacturer´s manual. Samples of purified recombinant GDF-15 were visualized with
Coomassie blue and quantified by densitometry comparison [24] with defined concentrations
of protein standards.
ELISA
For the determination of secreted GDF-15 from the glioma cell lines, 25.000 cells/well were
seeded in 48 well plates. The cell culture supernatants were collected after 24 hours and the
amount of GDF-15 determined by sandwich ELISA using a monoclonal anti-human GDF-15
capture antibody (R&D Systems, MAB957) and a biotinylated goat anti-human GDF-15
detection antibody (R&D Systems, BAF940) according to the manufacturer’s protocol.
Recombinant human GDF-15 (R&D Systems) diluted in medium (15 pg/ml to 250 pg/ml)

6
was used as a standard for the determination of the total amount of GDF-15 secreted by the
cells. The number of cells per well after 14 hours was determined by direct counting. The
experiment was performed three times with every cell line tested in triplicate per experiment.
Total RNA extraction and cDNA synthesis
Samples of tumor tissue (100 – 200 mg each) and cultured glioma cells were lysed in
peqGOLD TriFast reagent (Peqlab, Erlangen, Germany), and phenol-chloroform extraction of
total RNA was performed. Prior to reverse transcription RNA samples were treated with RQ1
DNase (Promega). cDNA synthesis was then performed using M-MLV Reverse Transcriptase
(Promega). Additionally, total RNA was extracted from primary (low-passage) cultures
established from glioma tissue of different malignancy grades.
Real-time quantitative RT-PCR analysis
Relative quantitation of human GDF-15 and GAPDH transcripts in cDNA samples was
performed on an ABI PRISM 7000 Sequence Detection System using the corresponding
TaqMan Assays-on-Demand Gene Expression Products (Applied Biosystems) and following
the manufacturer’s protocol. Each sample was analyzed in triplicate. GAPDH expression was
considered as an internal control to which GDF-15 expression was normalized.
Cell proliferation assays
Cells were seeded onto 96-well plates at a density of 104 (for U373MG: 5 x 103) cells/well.
On the next day, cultures were treated with rabbit anti-mouse GDF-15 polyclonal serum (5
L/ml), pre-immune serum (5 l/ml) as a control, or human recombinant GDF-15 (20 ng/ml)
(R&D). 24 hours later, cell proliferation was measured using 5-Bromo-2`-deoxy-uridine

7
labeling and the Detection Kit III (Roche Applied Science), following the manufacturer’s
instructions.
In vitro cell invasion assays
The ability of cells to migrate through Matrigel-coated filters was determined using the 24-
well plate BD BioCoat Matrigel Invasion Chamber (BD Biosciences, Bedford, MA, USA).
Briefly, cells were seeded at a density of 2.5x104 cells in 500 L DMEM onto Matrigel
inserts. Rabbit anti-mouse GDF-15 polyclonal serum (5 l/m), pre-immune serum (5 l/ml, as
a control), or human recombinant GDF-15 (20 ng/ml) were added to the inserts. Wells of the
24-well plates contained DMEM supplemented with 5% FBS as a chemoattractant. After
incubation for 24 h at 37oC in 5% CO2 the cells that had not invaded the filters were removed
with cotton swabs, and cells that had migrated to the lower surface of the filter were fixed in
methanol. Cells were then stained with Toluidine Blue (Merck, Darmstadt, Germany) and
counted in 32 fields (x200) per filter, covering the periphery and the center of the membrane.
All experiments were performed in triplicate.
TUNEL
Cells were washed and fixed with 4% PFA, permeabilized with 0.2% Triton X-100, and
processed for TUNEL (Promega). 5 µg/ml DAPI was added to stain cell nuclei.
Photomicrographs from 4-6 different fields in each coverslip were captured under green
channels and merged using adobe photoshop 5.5. Typically, about 1000 cells were analyzed
for the number of TUNEL-positive glioblastoma cells.

8
Statistical analysis
Data were expressed as means ± SEM. All experiments were performed in triplicates or
duplicates and repeated at least three times. Statistical analyses among groups were performed
using the t test (Origin 6.1). P-values are as follows: * P < 0.05, ** P < 0.01, and *** P <
0.001.

9
Results
GDF-15 immunoreactivity in de novo glioblastoma, high grade astrocytoma and
secondary glioblastoma
We first investigated the possible presence of GDF-15 protein in surgically removed human
primary and secondary glioblastomas using immunohistochemistry. Only one out of the nine
investigated primary glioblastomas grade IV showed moderate GDF-15 immunoreactivity (ir)
(Figure 1A; left panel). GDF-15 staining was localized to tumor cells, but absent from blood
vessels. The right panel shows the control experiment where the GDF-15 antibody was
omitted. In contrast to the largely GDF-15 negative primary glioblastomas we found
prominent staining for GDF-15 ir in 2 anaplastic astrocytomas grade III (AAG III) and one
secondary glioblastoma. The upper left panel of Figure 1B exemplifies GDF-15 ir in one of
the AAG III. Notably, the stroma and perivascular spaces of the AAG III were
immunoreactive for GDF-15. An oligodendroglioma grade III (OG III) was negative for
GDF-15 ir. In samples of tumor tissues, where GDF-15 ir was detectable, there was no
costaining of GDF-15 with glial fibrillary acidic protein (GFAP; Figure 1B; bottom left
panel). Similarly, GDF-15 immunoreactive cells in AAG III (Figure 1B; bottom right panel)
and secondary glioblastomas did not show co-labeling for the microglial marker CD68.
Together, these data suggest the presence of GDF-15 protein in GFAP- and CD68-negative
cells, preferentially in AAG III and secondary glioblastoma.
Lowest expression of GDF-15 in cultures of primary glioblastomas
Diffuse astrocytomas grade II (DAG II), anaplastic astrocytomas grade III (AAG III), and
secondary glioblastomas grade IV (SGG IV) represent distinct stages in the evolution of
secondary glioblastomas, whereas primary glioblastomas grade IV (PGG IV) develop along a
different genetic pathway [25]. Other brain tumours distinguishable from those mentioned

10
above due to their distinct genetical alteration, mainly characterized by a combined loss of
chromosomes 1p and 19q, are oligodendroglioblastomas and oligoastrocytomas grade II and
III (OGG II, OGG III; OAG II, OAG III) [26,27]. Figure 2 shows the results of quantitative
real-time PCR determinations of GDF-15 mRNA expressed in cultures derived from 8 DAG
II, 8 AAG III, 8 SGG IV, 8 PGG IV, 5 OGG II, 5 OGG III, 5 OAG II, and 5 OAG III. High
and largely comparable expression levels were found in DAG II, AAG III, and SGG IV
(Figure 2). In contrast, PGG IV showed significantly lower levels of GDF-15 expression. This
difference was even more pronounced when GDF-15 expression in PGG IV was compared to
the average expression levels of cell cultures derived from oligodendroglioblastomas grade II
(OGG II) and anaplastic oligodendroglioblastomas grade III (OGG III). The most frequent
type of mixed glioblastomas, OAG II, also showed a significantly higher GDF-15 level
compared to PGG IV. Thus, differences in GDF-15 expression might be exploited as a useful
indicator for distinguishing primary from other glial derived tumors.
GDF-15 expression does not correlate with spontaneous proliferation or the p53 status
of glioma cell lines
In an attempt to elucidate putative functions of GDF-15 on glioblastoma growth we
investigated a panel of 9 human glioma cell lines (LN-319, U87MG, A172, D-247MG, LN-
229, LN-308, LN-428, LN-18 and U373MG). We first compared the expression levels of
endogenous GDF-15 mRNA in relation to the spontaneous proliferation rate for each human
cell line. Figure 3A shows the wide range of mRNA expression levels in these cell lines, from
very high rates in A172 and D-247MG cells to up to 15-fold lower levels in LN-18 and
U87MG cells. Similar differences were found in terms of GDF-15 protein in cell lysates and
supernatants of selected cell lines analysed by Western blot (Figure 3B) and ELISA assay

11
(Figure 3C). Rates of proliferation of the 9 cell lines varied up to 3-fold, but failed to
correlate, neither in a positive or negative fashion, with levels of GDF-15 mRNA expression.
Since several studies have reported a strong p53 dependent induction of GDF-15 in a large
variety of cancers [4, 8-10], we additionally compared levels of GDF-15 expression with the
p53 status of the cell lines. Highest GDF-15 mRNA expression was found in 3 cell lines with
wt p53 status. However, p53 wt U87MG and mutant/p53 LN-18 cells showed lowest levels of
GDF-15 transcripts (Fig. 3A, Tab. 2) suggesting no significant correlation between the p53
status and the level of GDF-15 expression.
GDF-15 modulates proliferation of glioma cell lines
We next analysed, whether endogenous GDF-15 has an effect on the proliferation of
glioblastoma cells using a blocking antibody to GDF-15. As shown in Figure 4A, the antibody
(5µl/ml) significantly attenuated the mitogenic activity of 8 of the 9 human glioblastoma cell
lines. LN-18 cells showed no response to the antibody treatment in terms of proliferation.
Having shown that, with one exception, all glioblastoma cell lines employed in this study
responded to endogenous GDF-15, we next investigated their responsiveness to exogenously
applied GDF-15. As shown in Figure 4A, addition of recombinant GDF-15 (10ng/ml) to cell
cultures promoted proliferation of A172, D-247MG, LN-18, LN-319, but had no effect on
proliferation of LN-229, LN-308, LN-428, U373MG, and U373MG cells.
Taken together, these data suggest that endogenous GDF-15 is implicated in promoting
growth of most glioma cell lines studied, and that exogenous GDF-15 exerts growth
promoting effects on a more restricted set of glioma cell lines.

12
GDF-15 modulates invasiveness of distinct glioma cell lines
Invasiveness is a key feature of glioblastoma cells [28]. Figure 4B reveals that neutralization
of endogenous GDF-15 significantly suppressed invasiveness of LN-308 and U373MG
human glioma cells, promoted invasiveness of LN-428 cells, and did not affect invasiveness
of the other cell lines studied. Exogenous GDF-15 did not interfere with invasiveness of any
glioma cell line investigated.
Together, these data indicate that endogenous, but not exogenous GDF-15 affects
invasiveness of several but not all glioma cell lines studied.
GDF-15 does not affect apoptosis in glioma cell lines
Previous studies have identified GDF-15 as an important mediator of DNA damage in various
cancer cell lines, explaining its largely anti-tumorigenic and proapoptotic effects [4, 8, 14]. To
explore whether GDF-15 affects apoptosis in glioblastoma cell lines we quantified the number
of TUNEL positive cells in cultures previously treated with recombinant GDF-15 or GDF-15
antiserum (Table 3). Comparisons with control cultures showed no difference in numbers of
TUNEL positive cells suggesting that neither exogenous GDF-15 nor the inhibition of
endogenous GDF-15 have an impact on apoptotic cell death in the glioblastoma cell lines
investigated.

13
Discussion
The present study provides evidence for expression of GDF-15 in select human glioblastoma
tissues, glioblastoma primary cultures, and glioblastoma cell lines. While GDF-15
immunoreactivity was prominent in 3 of the 4 investigated secondary glioblastomas, only one
of nine primary glioblastomas showed staining for GDF-15. In support of the notion that
primary and secondary glioblastomas may differ with regard to their GDF-15 protein
expression, quantitative real-time RT-PCR carried out on cultured primary and secondary
glioblastoma cells revealed higher levels of GDF-15 transcripts in secondary as compared to
primary glioblastoma cells. With regard to secondary glioblastoma cell cultures we found an
overall high expression in all WHO grades, but no differences that could be correlated to the
degree of malignancy. This suggests that GDF-15 is unlikely to hold a key position in terms
of promoting progression of glioblastoma from a less to a more malignant phenotype
It is well established that the genetic pathways of primary and secondary glioblastomas
leading to glioma as the common phenotypic endpoint are different and show little overlap
[29]. Thus, higher expression levels of GDF-15 mRNA in secondary than primary
glioblastomas might be regulated by one or several of the known mutated, down- or
upregulated signalling pathways. In this context, p53 mutations were among the first genetic
alterations identified in glioma tumors. p53 mutations are less common in primary
glioblastoma (< 10%), while secondary glioblastomas have a high incidence of p53 mutations
(>65%). In the light of studies that have reported a strong p53 dependent induction of GDF-15
in a large variety of cancers [4, 8-10], we had expected to detect lower levels of GDF-15
expression in secondary glioblastoma because of increased probability for p53 mutations. One
explanation for this somewhat unexpected result might be a possible p53 independent
regulation of GDF-15 gene expression. p53 independent upregulation of GDF-15 has
previously been shown as a result of DNA damage, induced by genotoxic agents, irradiation

14
[8], or treatments with nonsteroidal anti-inflammatory drugs [4]. Thus, it is likely that GDF-
15 can be regulated through signals and pathways not involving p53.
Immunocytochemistry carried out on primary and secondary glioblastoma tissues revealed
clear differences in the localization of GDF-15 protein. In the primary glioblastoma GDF-15
immunoreactivity was exclusively localized in tumor cells, but absent from blood vessels and
perivascular areas. In contrast, we found prominent GDF-15 immunoreactivity in the ECM of
perivascular spaces of 2 anaplastic astrocytoma grade III (AAG III) and one secondary
glioblastoma grade IV (SGG IV), whereas tumor cells and microglia were GDF-15-negative.
Thus, it appears that GDF-15 of secondary glioblastomas is primarily located in the ECM.
Stromal ECM stores for GDF-15 in prostate tumors have previously been shown by Bauskin
and co-workers [30]. Interestingly, increased stromal stores of GDF-15 in prostate tumors
were associated with a significantly greater chance of remaining disease free. Whether
stromal storage of proGDF-15 in the ECM has a biological significance in secondary
glioblastomas remains to be elucidated.
To address putative roles of GDF-15 for glioblastoma cell properties, we investigated GDF-
15 expression and functions in a panel of 9 human glioblastoma cell lines with different p53
backgrounds. We found only weak GDF-15 mRNA expression in cell lines showing p53
mutations or deletions. Two of the four cell lines with wild type p53 status showed a higher
GDF-15 expression rate. This difference was further reflected in the amount of GDF-15
protein levels. Whether or not the increased expression depends on p53 remains to be
investigated.
Previous studies have identified GDF-15 as an important downstream mediator of DNA
damage signalling in various cancer cell lines and anti-tumorigenic and proapoptotic effects
of GDF-15 have been reported [4, 8, 16]. Moreover, recent reports demonstrated that
transfection of GDF-15 cDNA into human colorectal carcinoma and glioblastoma cells

15
decreased tumor growth in vivo and in vitro [4, 19]. In the present study we were unable to
demonstrate that recombinant GDF-15 activates apoptosis and/or suppresses proliferation of
glioblastoma cell lines. Our analyses rather revealed that exogenously added GDF-15 has no
effect or can stimulate proliferation of glioblastoma cell lines. More importantly, neutralizing
antibodies against GDF-15 attenuated the mitogenic activity of 8 (out of nine) cell lines
suggesting that blocking antibodies or Fab fragments of GDF-15 might be developed into a
therapeutic tool for treating glioblastoma. Together, the present and previous sets of data do
not allow to assign unequivocal promoting or inhibitory roles to GDF-15 in the regulation of
cell proliferation.
GDF-15 ambiguously affected glioma cell invasiveness in this study. Other studies carried out
with other types of cancer cells have shown that recombinant GDF-15 can promote
invasiveness of gastric cancer cells in a dose dependent manner [31]. Similarly, loss of cell
adhesion has been described in prostate cancer cells upon GDF-15 treatment [16]. Given the
diversity of effects exerted by GDF-15 in relation to invasiveness depending on cancer cell
type, it is conceivable that GDF-15, as other members of the TGF-ß superfamily, may
function in a crucially context-dependent manner
In conclusion, we have provided evidence for distinct levels and sites of expression of GDF-
15, a distant member of the TGF-ß superfamily, in primary and secondary glioblastoma. In
contrast to the documented proapoptotic and anti-tumorigenic activities of GDF-15 in several
cancer cell lines, our data suggest that GDF-15 does not decrease proliferation of
glioblastoma cell lines, while its effects on invasiveness are inconsistent. Mechanisms that
determine the distinct directions of the effects of GDF-15 may be sought in its context-
dependent actions with other growth factors.

16
Acknowledgements
This work was supported by grants of the Deutsche Forschungsgemeinschaft (STR 616/3-4,
KU 34/23-1) and the DFG Graduate College 791 at Heidelberg University. “Neural
Developmental and Degenerative Processes; Basic Research and Clinical Implications”.

17
References
[1] M.R. Bootcov, A.R. Bauskin, S.M. Valenzuela, et al., MIC-1, a novel macrophage inhibitory cytokine, is a
divergent member of the TGF-beta superfamily, Proc. Natl. Acad. Sci. USA 94 (1997) 11514-11519.
[2] V.M. Paralkar, A.L. Vail, W.A. Grasser, et al., Cloning and characterization of a novel member of the
transforming growth factor-beta/bone morphogenetic protein family, J. Biol. Chem. 273 (1998) 13760-
13767.
[3] R. Hromas, M. Hufford, J. Sutton, D. Xu, Y. L, L. Lu, PLAB, a novel placental bone morphogenetic protein,
Biochem. Biophy, Acta 1354 (1997) 40-44
[4] S.J. Baek, K.S. Kim, J.B. Nixon, L.C. Wilson, T.E. Eling, Cyclooxygenase inhibitors regulate the expression
of a TGF-beta superfamily member that has proapoptotic and antitumorigenic activities, Mol. Pharmacol.
59 (2001) 901-908.
[5] M. Böttner, C. Suter-Crazzolara, A. Schober, K. Unsicker, Expression of a novel member of the TGF-beta
superfamily, growth/differentiation factor-15/macrophage-inhibiting cytokine-1 (GDF-15/MIC-1) in adult
rat tissues, Cell Tissue Res. 297 (1999) 103-110.
[6] J. Strelau, A. Sullivan, M. Böttner, et al., Growth/differentiation factor-15/macrophage inhibitory cytokine-1
is a novel trophic factor for midbrain dopaminergic neurons in vivo, J. Neurosci. 20 (2000) 8597-8603.
[7] M. Böttner, K. Krieglstein, K. Unsicker, The transforming growth factor-betas: structure, signaling, and roles
in nervous system development and functions, J. Neurochem. 75 (2000) 2227-2240.
[8] P.-X. Li, J. Wong, A. Ayed, et al., Placental transforming growth factor- is a downstream mediator of the
growth arrest and apoptotic response of tumor cells to DNA damage and p53 overexpression, J. Biol.
Chem. 275 (2000) 20127-20135.
[9] K. Kannan, N. Amariglio, G. Rechavi, D. Givol, Profile of gene expression regulated by induced p53:
connection to the TGF- family, FEBS Lett. 470 (2000) 77-82.
[10] M. Tan, Y. Wang, K. Guan, Y. Sun, PTGF-, a type transforming growth factor (TGF-) superfamily
member, is a p53 target gene that inhibits tumor cell growth via TGF- signaling pathway, Proc. Natl.
Acad. Sci. USA 97 (2000) 109-114.
[11] J. Wong, P.X. Li, H.J. Klamut, A novel p53 transcriptional repressor element (p53TRE) and the
asymmetrical contribution of two p53 binding sites modulate the response of the placental transforming
growth factor-beta promoter to p53, J. Biol. Chem. 277 (2002) 26699-26707.

18
[12] F.G. Bottone Jr., S.J. Baek, J.B. Nixon, T.E. Eling, Diallyl disulfide (DADS) induces the antitumorigenic
NSAID-activated gene (NAG-1) by a p53-dependent mechanism in human colorectal HCT 116 cells, J.
Nutr. 132 (2002) 773-778.
[13] S.J. Baek, L.C. Wilson, C.-H. Lee, T.E. Eling, Dual function of nonsteroidal anti-inflammatory drugs
(NSAIDs): inhibition of cyclooxygenase and induction of NSAID-activated gene, J. Pharmacol. Exp.
Ther. 301 (2002) 1126-1131.
[14] P. Buckhaults, C. Rago, B. St Croix, et al., Secreted and cell surface genes expressed in benign and
malignant colorectal tumors, Cancer Res. 61 (2001) 6996-7001.
[15] R. Thomas, L.D. True, P.H. Lange, R.L.Vessella, Placental bone morphogenetic protein (PLAB) gene
expression in normal, pre-malignant and malignant human prostate: relation to tumor development and
progression, Int. J. Cancer 93 (2001) 47-52.
[16] T. Liu, A.R. Bauskin, J. Zaunders, et al., Macrophage inhibitory cytokine 1 reduces cell adhesion and
induces apoptosis in prostate cancer cells, Cancer Res. 63 (2003) 5034-5040.
[17] J.B. Welsh, L.M. Sapinoso, S.G. Kern, et al., Large-scale delineation of secreted protein biomarkers
overexpressed in cancer tissue and serum, Proc. Natl. Acad. Sci. USA 100 (2003) 3410-3415.
[18] D. Brown, R.L. Ward, P. Buckhaults, et al., MIC-1 serum level and genotype: associations with progress
and prognosis of colorectal carcinoma, Clin. Cancer Res. 9 (2003) 2642-2650.
[19] M. Albertoni, P.H. Shaw, M. Nozaki, et al., Anoxia induces macrophage inhibitory cytokine-1 (MIC-1) in
glioblastoma cells independently of p53 and HIF-1, Oncogene 21 (2002) 4212-4219.
[20] R. Graichen, D. Liu, Y. Sun, K.O. Lee, P.E. Lobie, Autocrine human growth hormone inhibits placental
transforming growth factor-beta gene transcription to prevent apoptosis and allow cell cycle progression
of human mammary carcinoma cells, J. Biol. Chem. 277 (2002) 26662-26672.
[21] M. Shim, T.E. Eling, Protein kinase C-dependent regulation of NAG-1/placental bone morphogenic
protein/MIC-1 expression in LNCaP prostate carcinoma cells, J. Biol. Chem. 280 (2005) 18636-18642.
[22] S. Karcher, H.H. Steiner, R. Ahmadi,et al., Different angiogenic phenotypes in primary and secondary
glioblastomas, Int. J. Cancer 118 (2006) 2182-2189.
[23] A. Schober, M. Böttner, J. Strelau, et al., Expression of growth differentiation factor-15/ macrophage
inhibitory cytokine-1 (GDF-15/MIC-1) in the perinatal, adult, and injured rat brain, J. Comp. Neurol. 439
(2001) 32-45.

19
[24] A.W. Henkel, S.C. Bieger, Quantification of proteins dissolved in an electrophoresis sample buffer, Anal.
Biochem. 223 (1994) 329-331.
[25] M. Gupta, A. Djalilvand, D.J. Brat. Clarifying the diffuse gliomas: an update on the morphologic features
and markers that discriminate oligodendroglioma from astrocytoma, Am J Clin Pathol. 124 (2005) 755-
768.
[26] C. Hartmann, W. Mueller, A.von Deimling. Pathology and molecular genetics of oligodendroglial tumors. J
Mol Med. 82 (2004) 638-655.
[27] H. Ohgaki, Genetic pathways to glioblastomas, Neuropathology 25 (2005) 1-7.
[28] T. Demuth, M.E. Berens, Molecular mechanisms of glioma cell migration and invasion, J. Neurooncol. 70
(2004) 217-228.
[29] P. Kleihues, W.K Cavenee, (Eds.) Pathology and genetics of tumours of the nervous system. IARC Press
Lyon, 2000, pp9-54.
[30] A.R. Bauskin, D.A. Brown, S. Junankar, et al., The propeptide mediates formation of stromal stores of
PROMIC-1: role in determining prostate cancer outcome, Cancer Res. 65 (2005) 2330-2336.
[31] D.H. Lee, Y. Yang, S.J. Lee, et al., Macrophage inhibitory cytokine-1 induces the invasiveness of gastric
cancer cells by up-regulating the urokinase-type plasminogen activator system, Cancer Res. 63 (2003)
4648-4655.

20
Figure Legends
Fig. 1A. Presence of GDF-15 in human primary and secondary glioblastoma tissue samples.
Left panel, peroxidase immunostaining of GDF-15 in a primary glioblastoma counterstained
with hematoxylin. Tumor cells are GDF-15 positive (T), whereas no staining was detectable
in the stroma of the tumor (S). Right panel, in the negative control (without 1st antibody), no
unspecific binding was observed. Fig. 1B. Fluorescence immunostainings of GDF-15, CD68
and GFAP in serial sections of an anaplastic astrocytoma grade III. Upper left panel
represents an overlay of GDF-15 and DAPI staining. Notably, GDF-15 is present in the
perivascular spaces of the stroma. Upper right panel: negative control using GDF-15 pre-
immune serum and DAPI staining. Bottom left panel: GFAP positive tumor cells are negative
for GDF-15. Bottom right panel: CD68 positive microglia do not show GDF-15
immunoreactivity.
Fig. 2. GDF-15 expression in primary cultures from human brain tumors of different origin
and malignancy grades was analyzed by quantitative RT-PCR. GDF-15 expression was
normalized against GAPDH and ß-Actin. PGG IV showed a significantly lower GDF-15
expression compared to SGG IV or OAG II (p<0.05). Comparison of PGG IV with OGG II or
OGG III revealed a significant difference of p<0.01. DAG II: diffuse astrocytomas grade II;
AAG III: anaplastic astrocytomas grade III; SGG IV: secondary glioblastomas grade IV; PGG
IV: primary glioblastomas grade IV: oligodendroglioblastomas grade II and III; OGG II and
III: oligoastrozytomas grade II and III; OAG II and III. Bars represent mean ± SD.
Fig. 3. No correlation exists between GDF-15 expression and rates of proliferation in
glioblastoma cell lines. (A) The upper figure shows GDF-15 expression levels in nine human
glioblastoma cell lines (quantitative RT-PCR), the lower figure shows proliferation rates of

21
the same cell lines, measured by BrdU incorporation. Bars represent ± SD. (B) Immunoblots
of cell lysates with purified GDF-15 antiserum confirm the highest expression of GDF-15
protein in D-247MG and A-172 cells. (C) GDF-15 protein concentrations per 10.000 cells in
supernatants of D-247MG, LN-308 and U87MG using an ELISA assay.
Fig. 4. Effects of GDF-15 anti-serum and recombinant human GDF-15 on the rates of
proliferation and invasive capacities of glioblastoma cell lines. (A) Proliferation assays
performed on ten glioblastoma cell lines show that the growth of most cell lines is inhibited
by GDF-15 anti-serum and slightly stimulated by recombinant GDF-15. (B) Invasion assays
indicate that recombinant GDF-15 does not affect invasiveness of glioblastoma cell lines.
However, GDF-15 anti-serum does significantly affect the invasive capacity of three of the
glioblastoma cell lines. * p<0.05; ** p<0.01; *** p<0.001. Bars represent ± SD.

1
Cancer Letters
EXPRESSION AND PUTATIVE FUNCTIONS OF GDF-15, A MEMBER
OF THE TGF-ß SUPERFAMILY, IN HUMAN GLIOMA AND
GLIOBLASTOMA CELL LINES
Jens Strelau 1, Corina Schmeer 1, Heike Peterziel 1, Tina Paech 1, Christel Herold-
Mende 2, Hans Steiner 3, Michael Weller 4 and Klaus Unsicker 1
1Department of Neuroanatomy and Interdisciplinary Center for Neurosciences (IZN)
University of Heidelberg, Germany2 Molecular Biology Laboratories, Department of Neurosurgery, University of Heidelberg,
Germany 3 Department of Neurosurgery, Nürnberg Süd, Nürnberg, Germany4 Department of Neurology, University Hospital Zurich, Switzerland.
Address for correspondence:
Dr. Jens Strelau
Neuroanatomy & Interdisciplinary Center for Neurosciences (IZN),
University of Heidelberg,
INF 307
D-69120 Heidelberg, Germany
Fax: 49-6221-54 56 04
e-mail: [email protected]
* Manuscript with track changes

2
Abstract
Recent studies have demonstrated growth-inhibiting effects of Growth differentiation factor-
15 (GDF-15) on different cancer cell lines in vitro and on tumor growth in vivo. Here, we
present data concerning expression of GDF-15 in glioblastoma. We found low levels of GDF-
15 transcripts in primary glioblastoma. Thus, GDF-15 expression might be exploited as a
useful indicator for distinguishing primary from other glial derived tumors. In contrast to the
documented proapoptotic and anti-tumorigenic activities of GDF-15 in several cancer cell
lines, our data suggest that GDF-15 does not decrease proliferation of glioblastoma cell lines,
while its effects on invasiveness are not consistent.
Key words: GDF-15, TGF- superfamily, glioblastomas, tumor

3
Introduction
Growth differentiation factor-15 (GDF-15), also known as macrophage inhibitory cytokine-1
[1], prostate-derived factor [2], placental bone morphogenetic protein [3], and nonsteroidal
anti-inflammatory drug-activated protein-1 [4], is a divergent member of the TGF-ß
superfamily and widely expressed in epithelial tissues and in brain [5, 6, 7].
Most recently, several reports have provided evidence for functional links between GDF-15
and factors involved in tumorigenesis. The tumor suppressor gene product p53, for instance,
strongly induces GDF-15, indicating for the first time a functional connection between p53
and the TGF-ß superfamily [8-12]. Nonsteroidal anti-inflammatory drugs (NSAIDs) that
inhibit tumor development can also increase GDF-15 expression [13]. Other studies have
documented increased levels of GDF-15 in serum and in tumors of patients with metastatic
cancer [14-18]. Perhaps most excitingly, GDF-15 has been reported to exert proapoptotic and
anti-tumorigenic functions on human colorectal, prostate, breast, and mammary cancer cells
in vitro and on colon and glioblastoma tumors in vivo [4, 8, 19-21]. Together, these findings
strongly suggest a putative role of GDF-15 in tumor development and/or progression and
underscore the importance of further exploration of an involvement of GDF-15 in
tumorigenesis.
We report here the expression of GDF-15 in human glioblastoma tissues, glioblastoma
primary cultures, and glioblastoma cell lines. Our analyses reveal that GDF-15 may be
implicated in the regulation of proliferation and invasiveness of human glioblastoma cells.

4
Materials and Methods
Samples and Cell culture
Tissue specimens were obtained intraoperatively from central parts of the tumors. Primary
and secondary GBM were histopathologically classified according to the classification of the
WHO. Tumors were characterized as secondary GBM in patients with previous identified low
grade (WHO II and WHO III) glioblastomas. Primary cultures of glioma tissues (Tab. 1)
were established as described earlier [22]. The human glioma cell lines A172, D247MG, LN-
18, LN-229, LN-308, LN-319, and LN-428 (see Tab. 2) were maintained in DMEM with 4.5
g glucose/L supplemented with 10% fetal bovine serum (FBS), penicillin and streptomycin.
Cultures were maintained in a humidified 5% CO2 atmosphere at 37oC.
GDF-15 immunohistochemistry
Samples of primary and secondary glioblastomas were obtained from surgical cases in the
Department of Neurosurgery, University of Heidelberg, Germany. Snap frozen samples were
embedded in Tissue-Tek (Sakura, Ramsey, MI, USA) and directly cut into 16µm slices. For
immunohistochemistry, sections were fixed in 4% PFA for 1 hour. Non-specific peroxidase
activity was blocked by incubating sections in 1% hydrogen peroxide in PBS for 10 min at
room temperature (RT). Non-specific antibody binding was blocked using blocking buffer
(5% goat serum and 0.1% Triton X-100 in PBS) for 1 hour at RT. After washing, coverslips
were incubated with rabbit anti-mouse GDF-15 polyclonal antibodies (affinity-purified and
diluted 1:200) or with PBS overnight at 4oC. Slides were subsequently washed 3 times for 5
min in PBS and incubated 30 min at RT with biotinylated goat anti-rabbit IgG (Vectastain
Elite ABC Kit, Vector Laboratories, Burlingame, CA, USA). All subsequent steps were
performed according to the manufacturer’s instructions. Slides were counterstained with
hematoxylin, coverslipped, and photographed using a Zeiss Axioplan 2 equipped with

5
AxioVision 3.1 software. Immunofluorescence was performed as described [23]. Primary
antibodies used were affinity purified polyclonal anti-GDF-15, monoclonal anti-CD68 (BD
Biosciences, San Jose, CA, USA), and monoclonal anti-GFAP (Sigma, Chicago, IL, USA).
Secondary antibodies were anti-mouse IgG Cy2 and anti-rabbit IgG Cy3 (Jackson
ImmunoResearch, West Baltimore Pike West Grove, PA, USA)
Gel electrophoresis and immunoblot analyses
Protein extracts were prepared by homogenizing the cells in electrophoresis sample buffer,
and the protein content was determined using densitometry [24]. 25µg protein extract per lane
were loaded on SDS-polyacrylamide gels and transferred to nitrocellulose membranes
(Hybond ECL, Amersham Pharmacia, Göttingen, Germany) by electroblotting. The
membranes were incubated with purified polyclonal rabbit anti-rat GDF-15/MIC-1 antibody
for 16 h at 4 °C. Bound antibody was detected with a peroxidase-conjugated secondary
antibody and the ECL western blotting substrate system (Amersham Pharmacia) according to
the manufacturer´s manual. Samples of purified recombinant GDF-15 were visualized with
Coomassie blue and quantified by densitometry comparison [24] with defined concentrations
of protein standards.
ELISA
For the determination of secreted GDF-15 from the glioma cell lines, 25.000 cells/well were
seeded in 48 well plates. The cell culture supernatants were collected after 24 hours and the
amount of GDF-15 determined by sandwich ELISA using a monoclonal anti-human GDF-15
capture antibody (R&D Systems, MAB957) and a biotinylated goat anti-human GDF-15
detection antibody (R&D Systems, BAF940) according to the manufacturer’s protocol.
Recombinant human GDF-15 (R&D Systems) diluted in medium (15 pg/ml to 250 pg/ml)

6
was used as a standard for the determination of the total amount of GDF-15 secreted by the
cells. The number of cells per well after 14 hours was determined by direct counting. The
experiment was performed three times with every cell line tested in triplicate per experiment.
Total RNA extraction and cDNA synthesis
Samples of tumor tissue (100 – 200 mg each) and cultured glioma cells were lysed in
peqGOLD TriFast reagent (Peqlab, Erlangen, Germany), and phenol-chloroform extraction of
total RNA was performed. Prior to reverse transcription RNA samples were treated with RQ1
DNase (Promega). cDNA synthesis was then performed using M-MLV Reverse Transcriptase
(Promega). Additionally, total RNA was extracted from primary (low-passage) cultures
established from glioma tissue of different malignancy grades.
Real-time quantitative RT-PCR analysis
Relative quantitation of human GDF-15 and GAPDH transcripts in cDNA samples was
performed on an ABI PRISM 7000 Sequence Detection System using the corresponding
TaqMan Assays-on-Demand Gene Expression Products (Applied Biosystems) and following
the manufacturer’s protocol. Each sample was analyzed in triplicate. GAPDH expression was
considered as an internal control to which GDF-15 expression was normalized.
Cell proliferation assays
Cells were seeded onto 96-well plates at a density of 104 (for U373MG: 5 x 103) cells/well.
On the next day, cultures were treated with rabbit anti-mouse GDF-15 polyclonal serum (5
L/ml), pre-immune serum (5 l/ml) as a control, or human recombinant GDF-15 (20 ng/ml)
(R&D). 24 hours later, cell proliferation was measured using 5-Bromo-2`-deoxy-uridine

7
labeling and the Detection Kit III (Roche Applied Science), following the manufacturer’s
instructions.
In vitro cell invasion assays
The ability of cells to migrate through Matrigel-coated filters was determined using the 24-
well plate BD BioCoat Matrigel Invasion Chamber (BD Biosciences, Bedford, MA, USA).
Briefly, cells were seeded at a density of 2.5x104 cells in 500 L DMEM onto Matrigel
inserts. Rabbit anti-mouse GDF-15 polyclonal serum (5 l/m), pre-immune serum (5 l/ml, as
a control), or human recombinant GDF-15 (20 ng/ml) were added to the inserts. Wells of the
24-well plates contained DMEM supplemented with 5% FBS as a chemoattractant. After
incubation for 24 h at 37oC in 5% CO2 the cells that had not invaded the filters were removed
with cotton swabs, and cells that had migrated to the lower surface of the filter were fixed in
methanol. Cells were then stained with Toluidine Blue (Merck, Darmstadt, Germany) and
counted in 32 fields (x200) per filter, covering the periphery and the center of the membrane.
All experiments were performed in triplicate.
TUNEL
Cells were washed and fixed with 4% PFA, permeabilized with 0.2% Triton X-100, and
processed for TUNEL (Promega). 5 µg/ml DAPI was added to stain cell nuclei.
Photomicrographs from 4-6 different fields in each coverslip were captured under green
channels and merged using adobe photoshop 5.5. Typically, about 1000 cells were analyzed
for the number of TUNEL-positive glioblastoma cells.

8
Statistical analysis
Data were expressed as means ± SEM. All experiments were performed in triplicates or
duplicates and repeated at least three times. Statistical analyses among groups were performed
using the t test (Origin 6.1). P-values are as follows: * P < 0.05, ** P < 0.01, and *** P <
0.001.

9
Results
GDF-15 immunoreactivity in de novo glioblastoma, high grade astrocytoma and
secondary glioblastoma
We first investigated the possible presence of GDF-15 protein in surgically removed human
primary and secondary glioblastomas using immunohistochemistry. Only one out of the nine
investigated primary glioblastomas grade IV showed moderate GDF-15 immunoreactivity (ir)
(Figure 1A; left panel). GDF-15 staining was localized to tumor cells, but absent from blood
vessels. The right panel shows the control experiment where the GDF-15 antibody was
omitted. In contrast to the largely GDF-15 negative primary glioblastomas we found
prominent staining for GDF-15 ir in 2 anaplastic astrocytomas grade III (AAG III) and one
secondary glioblastoma. The upper left panel of Figure 1B exemplifies GDF-15 ir in one of
the AAG III. Notably, the stroma and perivascular spaces of the AAG III were
immunoreactive for GDF-15. An oligodendroglioma grade III (OG III) was negative for
GDF-15 ir. In samples of tumor tissues, where GDF-15 ir was detectable, there was no
costaining of GDF-15 with glial fibrillary acidic protein (GFAP; Figure 1B; bottom left
panel). Similarly, GDF-15 immunoreactive cells in AAG III (Figure 1B; bottom right panel)
and secondary glioblastomas did not show co-labeling for the microglial marker CD68.
Together, these data suggest the presence of GDF-15 protein in GFAP- and CD68-negative
cells, preferentially in AAG III and secondary glioblastoma.
Lowest expression of GDF-15 in cultures of primary glioblastomas
Diffuse astrocytomas grade II (DAG II), anaplastic astrocytomas grade III (AAG III), and
secondary glioblastomas grade IV (SGG IV) represent distinct stages in the evolution of
secondary glioblastomas, whereas primary glioblastomas grade IV (PGG IV) develop along a
different genetic pathway [25]. Other brain tumours distinguishable from those mentioned

10
above due to their distinct genetical alteration, mainly characterized by a combined loss of
chromosomes 1p and 19q, are oligodendroglioblastomas and oligoastrocytomas grade II and
III (OGG II, OGG III; OAG II, OAG III) [26,27]. Figure 2 shows the results of quantitative
real-time PCR determinations of GDF-15 mRNA expressed in cultures derived from 10 DAG
II, 10 AAG III, 10 SGG IV, 10 PGG IV, 5 OGG II, 5 OGG III, 5 OAG II, and 5 OAG III.
High and largely comparable expression levels were found in DAG II, AAG III, and SGG IV
(Figure 2). In contrast, PGG IV showed significantly lower levels of GDF-15 expression. This
difference was even more pronounced when GDF-15 expression in PGG IV was compared to
the average expression levels of cell cultures derived from oligodendroglioblastomas grade II
(OGG II) and anaplastic oligodendroglioblastomas grade III (OGG III). The most frequent
type of mixed glioblastomas, OAG II, also showed a significantly higher GDF-15 level
compared to PGG IV. Thus, differences in GDF-15 expression might be exploited as a useful
indicator for distinguishing primary from other glial derived tumors.
GDF-15 expression does not correlate with spontaneous proliferation or the p53 status
of glioma cell lines
In an attempt to elucidate putative functions of GDF-15 on glioblastoma growth we
investigated a panel of 9 human glioma cell lines (LN-319, U87MG, A172, D-247MG, LN-
229, LN-308, LN-428, LN-18 and U373MG). We first compared the expression levels of
endogenous GDF-15 mRNA in relation to the spontaneous proliferation rate for each human
cell line. Figure 3A shows the wide range of mRNA expression levels in these cell lines, from
very high rates in A172 and D-247MG cells to up to 15-fold lower levels in LN-18 and
U87MG cells. Similar differences were found in terms of GDF-15 protein in cell lysates and
supernatants of selected cell lines analysed by Western blot (Figure 3B) and ELISA assay

11
(Figure 3C). Rates of proliferation of the 9 cell lines varied up to 3-fold, but failed to
correlate, neither in a positive or negative fashion, with levels of GDF-15 mRNA expression.
Since several studies have reported a strong p53 dependent induction of GDF-15 in a large
variety of cancers [4, 8-10], we additionally compared levels of GDF-15 expression with the
p53 status of the cell lines. Highest GDF-15 mRNA expression was found in 3 cell lines with
wt p53 status. However, p53 wt U87MG and mutant/p53 LN-18 cells showed lowest levels of
GDF-15 transcripts (Fig. 3A, Tab. 2) suggesting no significant correlation between the p53
status and the level of GDF-15 expression.
GDF-15 modulates proliferation of glioma cell lines
(deleted text)
We next analysed, whether endogenous GDF-15 has an effect on the proliferation of
glioblastoma cells using a blocking antibody to GDF-15. As shown in Figure 4A, the antibody
(5µl/ml) significantly attenuated the mitogenic activity of 8 of the 9 human glioblastoma cell
lines. LN-18 cells showed no response to the antibody treatment in terms of proliferation.
Having shown that, with one exception, all glioblastoma cell lines employed in this study
responded to endogenous GDF-15, we next investigated their responsiveness to exogenously
applied GDF-15. As shown in Figure 4A, addition of recombinant GDF-15 (10ng/ml) to cell
cultures promoted proliferation of A172, D-247MG, LN-18, LN-319, but had no effect on
proliferation of LN-229, LN-308, LN-428, U373MG, and U373MG cells.
Taken together, these data suggest that endogenous GDF-15 is implicated in promoting
growth of most glioma cell lines studied, and that exogenous GDF-15 exerts growth
promoting effects on a more restricted set of glioma cell lines.

12
GDF-15 modulates invasiveness of distinct glioma cell lines
Invasiveness is a key feature of glioblastoma cells [28]. Figure 4B reveals that neutralization
of endogenous GDF-15 significantly suppressed invasiveness of LN-308 and U373MG
human glioma cells, promoted invasiveness of LN-428 cells, and did not affect invasiveness
of the other cell lines studied. Exogenous GDF-15 did not interfere with invasiveness of any
glioma cell line investigated.
Together, these data indicate that endogenous, but not exogenous GDF-15 affects
invasiveness of several but not all glioma cell lines studied.
GDF-15 does not affect apoptosis in glioma cell lines
Previous studies have identified GDF-15 as an important mediator of DNA damage in various
cancer cell lines, explaining its largely anti-tumorigenic and proapoptotic effects [4, 8, 14]. To
explore whether GDF-15 affects apoptosis in glioblastoma cell lines we quantified the number
of TUNEL positive cells in cultures previously treated with recombinant GDF-15 or GDF-15
antiserum (Table 3). Comparisons with control cultures showed no difference in numbers of
TUNEL positive cells suggesting that neither exogenous GDF-15 nor the inhibition of
endogenous GDF-15 have an impact on apoptotic cell death in the glioblastoma cell lines
investigated.

13
Discussion
The present study provides evidence for expression of GDF-15 in select human glioblastoma
tissues, glioblastoma primary cultures, and glioblastoma cell lines. While GDF-15
immunoreactivity was prominent in 3 of the 4 investigated secondary glioblastomas, only one
of nine primary glioblastomas showed staining for GDF-15. In support of the notion that
primary and secondary glioblastomas may differ with regard to their GDF-15 protein
expression, quantitative real-time RT-PCR carried out on cultured primary and secondary
glioblastoma cells revealed higher levels of GDF-15 transcripts in secondary as compared to
primary glioblastoma cells. With regard to secondary glioblastoma cell cultures we found an
overall high expression in all WHO grades, but no differences that could be correlated to the
degree of malignancy. This suggests that GDF-15 is unlikely to hold a key position in terms
of promoting progression of glioblastoma from a less to a more malignant phenotype
It is well established that the genetic pathways of primary and secondary glioblastomas
leading to glioma as the common phenotypic endpoint are different and show little overlap
[29]. Thus, higher expression levels of GDF-15 mRNA in secondary than primary
glioblastomas might be regulated by one or several of the known mutated, down- or
upregulated signalling pathways. In this context, p53 mutations were among the first genetic
alterations identified in glioma tumors. p53 mutations are less common in primary
glioblastoma (< 10%), while secondary glioblastomas have a high incidence of p53 mutations
(>65%). In the light of studies that have reported a strong p53 dependent induction of GDF-15
in a large variety of cancers [4, 8-10], we had expected to detect lower levels of GDF-15
expression in secondary glioblastoma because of increased probability for p53 mutations. One
explanation for this somewhat unexpected result might be a possible p53 independent
regulation of GDF-15 gene expression. p53 independent upregulation of GDF-15 has
previously been shown as a result of DNA damage, induced by genotoxic agents, irradiation

14
[8], or treatments with nonsteroidal anti-inflammatory drugs [4]. (deleted text) Thus, it is
likely that GDF-15 can be regulated through signals and pathways not involving p53.
Immunocytochemistry carried out on primary and secondary glioblastoma tissues revealed
clear differences in the localization of GDF-15 protein. In the primary glioblastoma GDF-15
immunoreactivity was exclusively localized in tumor cells, but absent from blood vessels and
perivascular areas. In contrast, we found prominent GDF-15 immunoreactivity in the ECM of
perivascular spaces of 2 anaplastic astrocytoma grade III (AAG III) and one secondary
glioblastoma grade IV (SGG IV), whereas tumor cells and microglia were GDF-15-negative.
Thus, it appears that GDF-15 of secondary glioblastomas is primarily located in the ECM.
Stromal ECM stores for GDF-15 in prostate tumors have previously been shown by Bauskin
and co-workers [30]. Interestingly, increased stromal stores of GDF-15 in prostate tumors
were associated with a significantly greater chance of remaining disease free. Whether
stromal storage of proGDF-15 in the ECM has a biological significance in secondary
glioblastomas remains to be elucidated.
To address putative roles of GDF-15 for glioblastoma cell properties, we investigated GDF-
15 expression and functions in a panel of 9 human glioblastoma cell lines with different p53
backgrounds. We found only weak GDF-15 mRNA expression in cell lines showing p53
mutations or deletions. Two of the four cell lines with wild type p53 status showed a higher
GDF-15 expression rate. This difference was further reflected in the amount of GDF-15
protein levels. Whether or not the increased expression depends on p53 remains to be
investigated.
Previous studies have identified GDF-15 as an important downstream mediator of DNA
damage signalling in various cancer cell lines and anti-tumorigenic and proapoptotic effects
of GDF-15 have been reported [4, 8, 16]. Moreover, recent reports demonstrated that
transfection of GDF-15 cDNA into human colorectal carcinoma and glioblastoma cells

15
decreased tumor growth in vivo and in vitro [4, 19]. In the present study we were unable to
demonstrate that recombinant GDF-15 activates apoptosis and/or suppresses proliferation of
glioblastoma cell lines. Our analyses rather revealed that exogenously added GDF-15 has no
effect or can stimulate proliferation of glioblastoma cell lines. More importantly, neutralizing
antibodies against GDF-15 attenuated the mitogenic activity of 8 (out of nine) cell lines
suggesting that blocking antibodies or Fab fragments of GDF-15 might be developed into a
therapeutic tool for treating glioblastoma. Together, the present and previous sets of data do
not allow to assign unequivocal promoting or inhibitory roles to GDF-15 in the regulation of
cell proliferation.
GDF-15 ambiguously affected glioma cell invasiveness in this study. Other studies carried out
with other types of cancer cells have shown that recombinant GDF-15 can promote
invasiveness of gastric cancer cells in a dose dependent manner [31]. Similarly, loss of cell
adhesion has been described in prostate cancer cells upon GDF-15 treatment [16]. Given the
diversity of effects exerted by GDF-15 in relation to invasiveness depending on cancer cell
type, it is conceivable that GDF-15, as other members of the TGF-ß superfamily, may
function in a crucially context-dependent manner
In conclusion, we have provided evidence for distinct levels and sites of expression of GDF-
15, a distant member of the TGF-ß superfamily, in primary and secondary glioblastoma. In
contrast to the documented proapoptotic and anti-tumorigenic activities of GDF-15 in several
cancer cell lines, our data suggest that GDF-15 does not decrease proliferation of
glioblastoma cell lines, while its effects on invasiveness are inconsistent. Mechanisms that
determine the distinct directions of the effects of GDF-15 may be sought in its context-
dependent actions with other growth factors.

16
Acknowledgements
This work was supported by grants of the Deutsche Forschungsgemeinschaft (STR 616/3-4,
KU 34/23-1) and the DFG Graduate College 791 at Heidelberg University. “Neural
Developmental and Degenerative Processes; Basic Research and Clinical Implications”.

17
References
[1] M.R. Bootcov, A.R. Bauskin, S.M. Valenzuela, et al., MIC-1, a novel macrophage inhibitory cytokine, is a
divergent member of the TGF-beta superfamily, Proc. Natl. Acad. Sci. USA 94 (1997) 11514-11519.
[2] V.M. Paralkar, A.L. Vail, W.A. Grasser, et al., Cloning and characterization of a novel member of the
transforming growth factor-beta/bone morphogenetic protein family, J. Biol. Chem. 273 (1998) 13760-
13767.
[3] R. Hromas, M. Hufford, J. Sutton, D. Xu, Y. L, L. Lu, PLAB, a novel placental bone morphogenetic protein,
Biochem. Biophy, Acta 1354 (1997) 40-44
[4] S.J. Baek, K.S. Kim, J.B. Nixon, L.C. Wilson, T.E. Eling, Cyclooxygenase inhibitors regulate the expression
of a TGF-beta superfamily member that has proapoptotic and antitumorigenic activities, Mol. Pharmacol.
59 (2001) 901-908.
[5] M. Böttner, C. Suter-Crazzolara, A. Schober, K. Unsicker, Expression of a novel member of the TGF-beta
superfamily, growth/differentiation factor-15/macrophage-inhibiting cytokine-1 (GDF-15/MIC-1) in adult
rat tissues, Cell Tissue Res. 297 (1999) 103-110.
[6] J. Strelau, A. Sullivan, M. Böttner, et al., Growth/differentiation factor-15/macrophage inhibitory cytokine-1
is a novel trophic factor for midbrain dopaminergic neurons in vivo, J. Neurosci. 20 (2000) 8597-8603.
[7] M. Böttner, K. Krieglstein, K. Unsicker, The transforming growth factor-betas: structure, signaling, and roles
in nervous system development and functions, J. Neurochem. 75 (2000) 2227-2240.
[8] P.-X. Li, J. Wong, A. Ayed, et al., Placental transforming growth factor- is a downstream mediator of the
growth arrest and apoptotic response of tumor cells to DNA damage and p53 overexpression, J. Biol.
Chem. 275 (2000) 20127-20135.
[9] K. Kannan, N. Amariglio, G. Rechavi, D. Givol, Profile of gene expression regulated by induced p53:
connection to the TGF- family, FEBS Lett. 470 (2000) 77-82.
[10] M. Tan, Y. Wang, K. Guan, Y. Sun, PTGF-, a type transforming growth factor (TGF-) superfamily
member, is a p53 target gene that inhibits tumor cell growth via TGF- signaling pathway, Proc. Natl.
Acad. Sci. USA 97 (2000) 109-114.
[11] J. Wong, P.X. Li, H.J. Klamut, A novel p53 transcriptional repressor element (p53TRE) and the
asymmetrical contribution of two p53 binding sites modulate the response of the placental transforming
growth factor-beta promoter to p53, J. Biol. Chem. 277 (2002) 26699-26707.

18
[12] F.G. Bottone Jr., S.J. Baek, J.B. Nixon, T.E. Eling, Diallyl disulfide (DADS) induces the antitumorigenic
NSAID-activated gene (NAG-1) by a p53-dependent mechanism in human colorectal HCT 116 cells, J.
Nutr. 132 (2002) 773-778.
[13] S.J. Baek, L.C. Wilson, C.-H. Lee, T.E. Eling, Dual function of nonsteroidal anti-inflammatory drugs
(NSAIDs): inhibition of cyclooxygenase and induction of NSAID-activated gene, J. Pharmacol. Exp.
Ther. 301 (2002) 1126-1131.
[14] P. Buckhaults, C. Rago, B. St Croix, et al., Secreted and cell surface genes expressed in benign and
malignant colorectal tumors, Cancer Res. 61 (2001) 6996-7001.
[15] R. Thomas, L.D. True, P.H. Lange, R.L.Vessella, Placental bone morphogenetic protein (PLAB) gene
expression in normal, pre-malignant and malignant human prostate: relation to tumor development and
progression, Int. J. Cancer 93 (2001) 47-52.
[16] T. Liu, A.R. Bauskin, J. Zaunders, et al., Macrophage inhibitory cytokine 1 reduces cell adhesion and
induces apoptosis in prostate cancer cells, Cancer Res. 63 (2003) 5034-5040.
[17] J.B. Welsh, L.M. Sapinoso, S.G. Kern, et al., Large-scale delineation of secreted protein biomarkers
overexpressed in cancer tissue and serum, Proc. Natl. Acad. Sci. USA 100 (2003) 3410-3415.
[18] D. Brown, R.L. Ward, P. Buckhaults, et al., MIC-1 serum level and genotype: associations with progress
and prognosis of colorectal carcinoma, Clin. Cancer Res. 9 (2003) 2642-2650.
[19] M. Albertoni, P.H. Shaw, M. Nozaki, et al., Anoxia induces macrophage inhibitory cytokine-1 (MIC-1) in
glioblastoma cells independently of p53 and HIF-1, Oncogene 21 (2002) 4212-4219.
[20] R. Graichen, D. Liu, Y. Sun, K.O. Lee, P.E. Lobie, Autocrine human growth hormone inhibits placental
transforming growth factor-beta gene transcription to prevent apoptosis and allow cell cycle progression
of human mammary carcinoma cells, J. Biol. Chem. 277 (2002) 26662-26672.
[21] M. Shim, T.E. Eling, Protein kinase C-dependent regulation of NAG-1/placental bone morphogenic
protein/MIC-1 expression in LNCaP prostate carcinoma cells, J. Biol. Chem. 280 (2005) 18636-18642.
[22] S. Karcher, H.H. Steiner, R. Ahmadi,et al., Different angiogenic phenotypes in primary and secondary
glioblastomas, Int. J. Cancer 118 (2006) 2182-2189.
[23] A. Schober, M. Böttner, J. Strelau, et al., Expression of growth differentiation factor-15/ macrophage
inhibitory cytokine-1 (GDF-15/MIC-1) in the perinatal, adult, and injured rat brain, J. Comp. Neurol. 439
(2001) 32-45.

19
[24] A.W. Henkel, S.C. Bieger, Quantification of proteins dissolved in an electrophoresis sample buffer, Anal.
Biochem. 223 (1994) 329-331.
[25] M. Gupta, A. Djalilvand, D.J. Brat. Clarifying the diffuse gliomas: an update on the morphologic features
and markers that discriminate oligodendroglioma from astrocytoma, Am J Clin Pathol. 124 (2005) 755-
768.
[26] C. Hartmann, W. Mueller, A.von Deimling. Pathology and molecular genetics of oligodendroglial tumors. J
Mol Med. 82 (2004) 638-655.
[27] H. Ohgaki, Genetic pathways to glioblastomas, Neuropathology 25 (2005) 1-7.
[28] T. Demuth, M.E. Berens, Molecular mechanisms of glioma cell migration and invasion, J. Neurooncol. 70
(2004) 217-228.
[29] P. Kleihues, W.K Cavenee, (Eds.) Pathology and genetics of tumours of the nervous system. IARC Press
Lyon, 2000, pp9-54.
[30] A.R. Bauskin, D.A. Brown, S. Junankar, et al., The propeptide mediates formation of stromal stores of
PROMIC-1: role in determining prostate cancer outcome, Cancer Res. 65 (2005) 2330-2336.
[31] D.H. Lee, Y. Yang, S.J. Lee, et al., Macrophage inhibitory cytokine-1 induces the invasiveness of gastric
cancer cells by up-regulating the urokinase-type plasminogen activator system, Cancer Res. 63 (2003)
4648-4655.

20
Figure Legends
Fig. 1A. Presence of GDF-15 in human primary and secondary glioblastoma tissue samples.
Left panel, peroxidase immunostaining of GDF-15 in a primary glioblastoma counterstained
with hematoxylin. Tumor cells are GDF-15 positive (T), whereas no staining was detectable
in the stroma of the tumor (S). Right panel, in the negative control (without 1st antibody), no
unspecific binding was observed. Fig. 1B. Fluorescence immunostainings of GDF-15, CD68
and GFAP in serial sections of an anaplastic astrocytoma grade III. Upper left panel
represents an overlay of GDF-15 and DAPI staining. Notably, GDF-15 is present in the
perivascular spaces of the stroma. Upper right panel: negative control using GDF-15 pre-
immune serum and DAPI staining. Bottom left panel: GFAP positive tumor cells are negative
for GDF-15. Bottom right panel: CD68 positive microglia do not show GDF-15
immunoreactivity.
Fig. 2. GDF-15 expression in primary cultures from human brain tumors of different origin
and malignancy grades was analyzed by quantitative RT-PCR. GDF-15 expression was
normalized against GAPDH and ß-Actin. PGG IV showed a significantly lower GDF-15
expression compared to SGG IV or OAG II (p<0.05). Comparison of PGG IV with OGG II or
OGG III revealed a significant difference of p<0.01. DAG II: diffuse astrocytomas grade II;
AAG III: anaplastic astrocytomas grade III; SGG IV: secondary glioblastomas grade IV; PGG
IV: primary glioblastomas grade IV: oligodendroglioblastomas grade II and III; OGG II and
III: oligoastrozytomas grade II and III; OAG II and III. Bars represent mean ± SD.
Fig. 3. No correlation exists between GDF-15 expression and rates of proliferation in
glioblastoma cell lines. (A) The upper figure shows GDF-15 expression levels in nine human
glioblastoma cell lines (quantitative RT-PCR), the lower figure shows proliferation rates of

21
the same cell lines, measured by BrdU incorporation. Bars represent ± SD. (B) Immunoblots
of cell lysates with purified GDF-15 antiserum confirm the highest expression of GDF-15
protein in D-247MG and A-172 cells. (C) GDF-15 protein concentrations per 10.000 cells in
supernatants of D-247MG, LN-308 and U87MG using an ELISA assay.
Fig. 4. Effects of GDF-15 anti-serum and recombinant human GDF-15 on the rates of
proliferation and invasive capacities of glioblastoma cell lines. (A) Proliferation assays
performed on ten glioblastoma cell lines show that the growth of most cell lines is inhibited
by GDF-15 anti-serum and slightly stimulated by recombinant GDF-15. (B) Invasion assays
indicate that recombinant GDF-15 does not affect invasiveness of glioblastoma cell lines.
However, GDF-15 anti-serum does significantly affect the invasive capacity of three of the
glioblastoma cell lines. * p<0.05; ** p<0.01; *** p<0.001. Bars represent ± SD.

Dear Editor:
thank you very much for sending us the constructive criticisms of two reviewers on our manuscript. We have dealt with their remarks in every respect (see below) and now submit a carefully revised version of the paper.
Response to Reviewers:
Reviewer 1
1. First this subtitle "GDF-15 is differentially expressed in cultures of primary and secondary glioblastoma" is misleading.We changed the subtitle into “Low expression of GDF-15 in cultures of primary glioblastomas” and present in the subsequent chapter data on diffuse astrocytomas grade II, anaplastic astrocytomas grade III, secondary glioblastomas grade IV, primary glioblastomas grade IV, oligodendroglioblastomas grade II and III, and oligoastrocytomas grade II and III.We also changed the first subtitle of the Results part into “GDF-15 immunoreactivity in de novo glioblastoma, high grade astrocytoma and secondary glioblastoma” and subdivided figure 1 into A and B for the sake of clarity of the immunohistochemical results.
2. No details are given about how these cultures were derived, maintained or what passage numbers were used in the present studies.Details are now provided in the Materials and Methods section on how the cultures were derived. Furthermore, data on GDF-15 expression in oligodendrogliomas grade II and III and oligoastrocytomas grade II and III were added to figure 2 and Results. We also incorporated a table comprising tumor type, passage numbers, age and sex of each specimen (Tab.1).
4. Did the authors conduct similar studies on the primary cell lines used in Figure 2?Figure 2 shows real-time PCR results with cDNA of 59 different primary cell cultures with low passage numbers provided by the Molecular Biology Laboratories, Department of Neurosurgery, University of Heidelberg. Since cell cultures with similarly low passage numbers were not available we did not perform invasion-, proliferation-, and TUNEL-assays on primary cell lines.
5. How do the authors explain the differences in responsiveness of the different cell lines?Our studies show that the GDF-15 antiserum decreases proliferation in 8 of 9 cell cultures. Exogenous GDF-15 increased cell division in 4 cell lines, while the remaining 5 cell lines did not respond. Several possibilities may be conceived to explain this result. First, cell lines unresponsive to exogenous GDF-15, but responsive to the anti-GDF-15 might already be maximally stimulated by the endogenous GDF-15. Moreover, non-responsive cell lines might require, in addition to GDF-15, a synergistically acting factor, which was not available at the required amounts. Alternatively, GDF-15 receptors in non-responsive cell lines might already be maximally saturated with endogenous GDF-15. A similar explanation might account for the lack of response to exogenous GDF-15 in the invasion experiments. It should be noted, however, that both U87MG and, to a lesser extent, LN-428 and LN-229, showed a tendency to respond to GDF-15.
It has been shown that, dependent on tumor type and origin (for review see Eling at al., 2006; Bauskin et al., 2006), GDF-15 can exhibit either tumorigenic or anti-tumorigenic activity. The cell lines investigated in our studies are derived from tumors of different stages and origin,and it is therefore conceivable that GDF-15 may regulate in either a positive or negative
* Revision notes

manner, depending on molecular and cellular contexts. Antagonistic effects have also been shown for TGF and its effects on different glioma cell lines (see also Jennings et al., 1991).
Although previous studies suggested that GDF-15 can regulate invasiveness in cancer cell lines, results of these former studies were based on a single cell line, without comparing the results with other cell lines derived from the same cancer types. It has been shown, for example, that GDF-15 significantly increases invasiveness of SNU-216, a human gastric cancer cell line (Lee et al., 2003). Evidently, this does not prove that all available gastric cancer cell lines would respond in the same way. Our study compares GDF-15 dependent invasion in 8 different cancer cell lines, which may differ in a variety of subtle respects.
6. Lastly, no effect on apoptosis was noted. How do the authors explain this result, which is different from previously published studies?Most in vitro studies suggest GDF-15- induces apoptosis in cancer cell lines. However, data from the literature provide high expression of GDF-15 in many tumors, a finding that is hard to be reconciled with the proposed tumor suppressor activity of GDF-15. Previous studies from our laboratory clearly demonstrate that GDF-15 can protect cerebellar granule cells from apoptotic cell death in vitro (Subramaniam et al., 2003), i.e. GDF-15 may function as an anti-apoptotic factor. To our knowledge, none of the previously published studies has shown pro-apoptotic effects of GDF-15 in glioma cultures.
7. The conclusions of this study are overstated.Conclusions have been changed
Reviewer 2
1. How are primary and secondary GBM differentiated (clinically or molecular). Primary and secondary GBM were differentiated as described earlier [22] In brief, tumors were histopathologically classified according to the classification of the WHO. Tumors were characterized as secondary GBM in patients with previous identified low grade (WHO II and WHO III) glioblastomas in accordance with Kleihues.
2. Why was no fresh material used for qPCR?We have now included fresh material provided by the Molecular Biology Laboratories, Department of Neurosurgery, University of Heidelberg. We got 9 frozen tissue samples ofprimary GBM and used the samples for qPCR and immunohistochemistry. The results of the qPCR with fresh primary GBM confirmed data obtained by IHC (low GDF-15 expression in 8 out of 9 tumors, (see also the figure on the last page of our response). Three of the primary cultures used for qPCR correspond to fresh tissue samples (Tab.1). Since one secondary GBM, two anaplastic astrocytoma and one oligodendroblastoma were provided as frozen sections on coverslips we could not perform qPCR and used the material directly for immunohistochemistry. However, primary cells derived from these tumors revealed high amounts of GDF-15 mRNA (Tab.1), although we were not able to show any ir in the oligodendroglioma grade III.
3. What is a low-passage culture and how was the immunohisto / qPCR correlation?
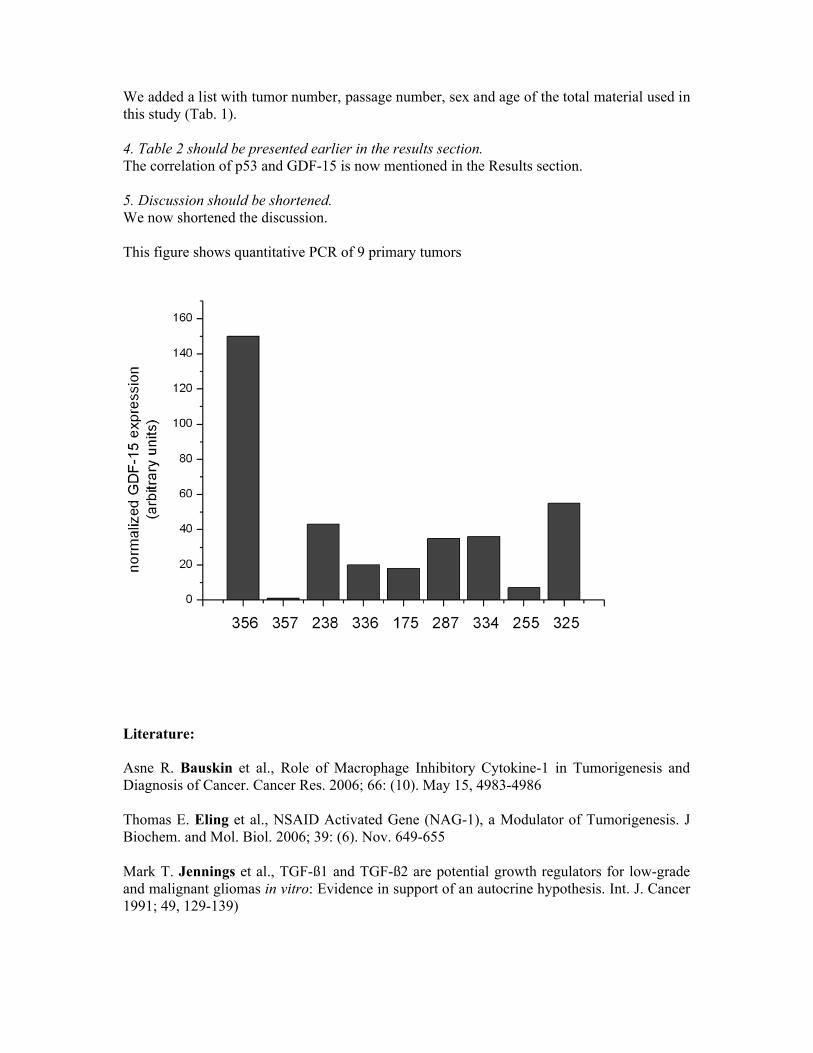
We added a list with tumor number, passage number, sex and age of the total material used in this study (Tab. 1).
4. Table 2 should be presented earlier in the results section.The correlation of p53 and GDF-15 is now mentioned in the Results section.
5. Discussion should be shortened.We now shortened the discussion.
This figure shows quantitative PCR of 9 primary tumors
Literature:
Asne R. Bauskin et al., Role of Macrophage Inhibitory Cytokine-1 in Tumorigenesis and Diagnosis of Cancer. Cancer Res. 2006; 66: (10). May 15, 4983-4986
Thomas E. Eling et al., NSAID Activated Gene (NAG-1), a Modulator of Tumorigenesis. J Biochem. and Mol. Biol. 2006; 39: (6). Nov. 649-655
Mark T. Jennings et al., TGF-ß1 and TGF-ß2 are potential growth regulators for low-grade and malignant gliomas in vitro: Evidence in support of an autocrine hypothesis. Int. J. Cancer 1991; 49, 129-139)
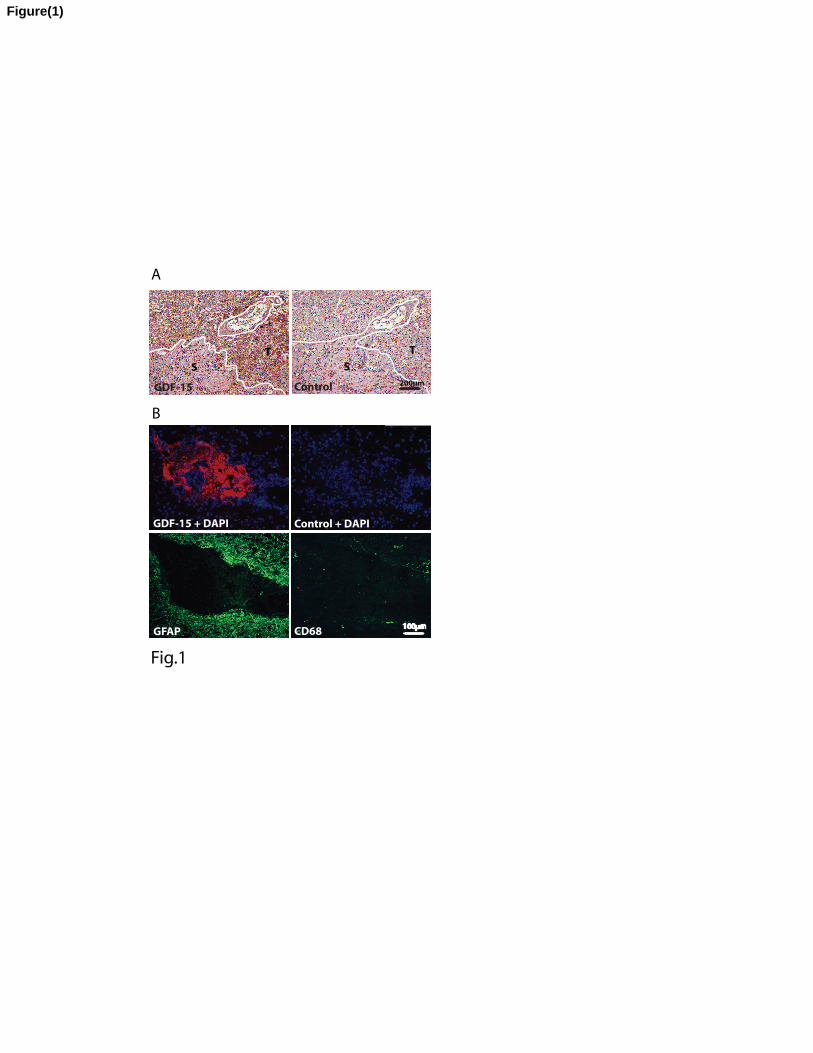
T
S
GFAP CD68
DAPI
Fig.1
GDF-15 + DAPI Control + DAPI
T
S
ControlGDF-15200µm
100µm
A
B
Figure(1)
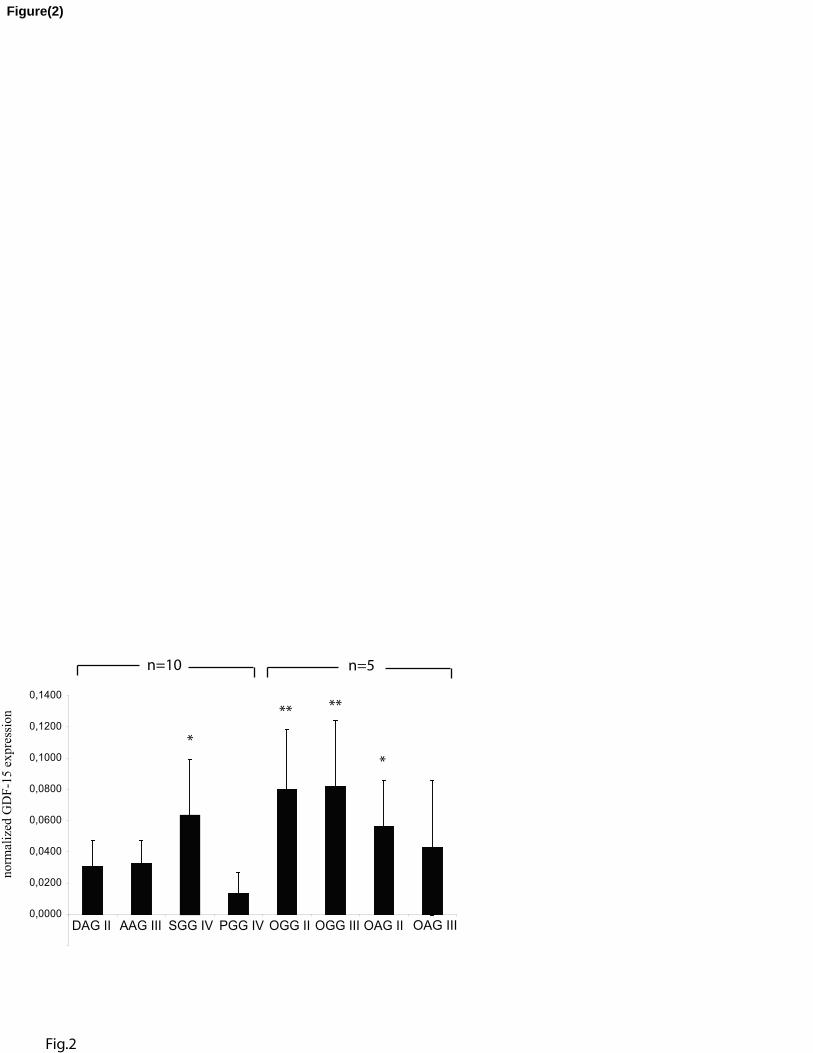
0,0000
0,0200
0,0400
0,0600
0,0800
0,1000
0,1200
0,1400
DAG II AAG III SGG IV PGG IV OGG II OGG III OAG II
*
** **
*
Fig.2
n=5n=10
OAG III
norm
aliz
ed G
DF
-15 e
xpre
ssio
nFigure(2)
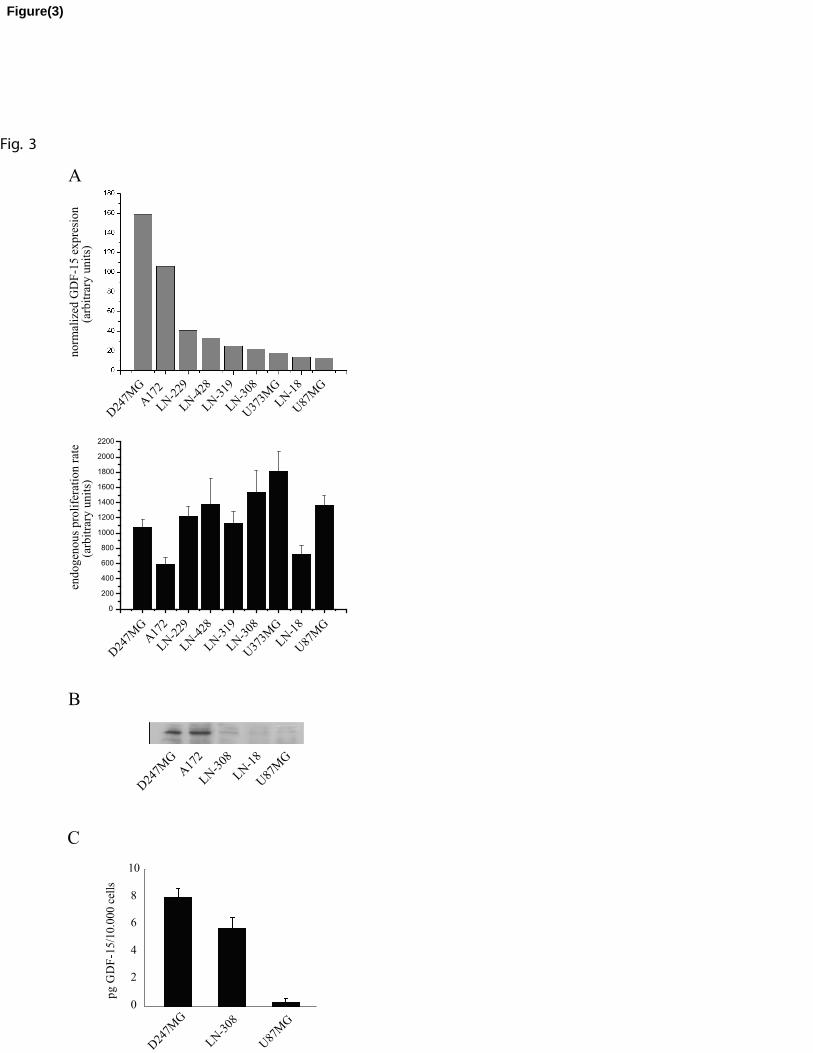
B
D24
7MG
A17
2
LN
-308
LN
-18
U87
MG
D24
7MG
A17
2
LN-2
29
LN-4
28
LN-3
19
LN
-308
U37
3MG
LN
-18
U87
MG
A
norm
aliz
ed G
DF
-15 e
xpre
sion
(arb
itra
ry u
nit
s)
0
200
400
600
800
1000
1200
1400
1600
1800
2000
2200
endogen
ous
pro
life
rati
on r
ate
(arb
itra
ry u
nit
s)
D24
7MG
A17
2
LN-2
29
LN-4
28
LN-3
19
LN
-308
U37
3MG
LN
-18
U87
MG
Fig. 3
C
D24
7MG
LN
-308
U87
MG
0
2
4
6
8
10
pg
GD
F-1
5/1
0.0
00 c
ells
Figure(3)
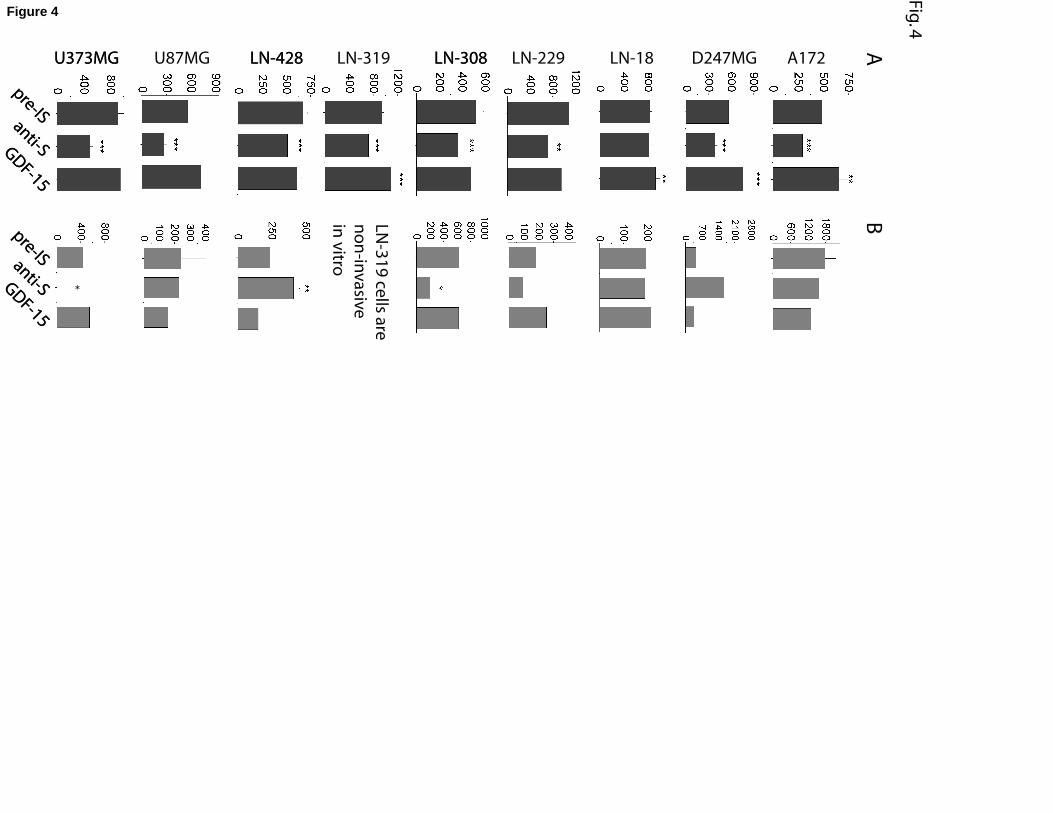
LN-18 D247MG
AB
LN-308
LN
-31
9 ce
lls are
no
n-in
va
sive
in v
itro
LN-319LN-428U373MG
pre-IS
anti-SGD
F-15
pre-ISanti-SGD
F-15
LNT-229 A172LN-308LN-428U87MGU373MG
pre-IS
anti-SGD
F-15
pre-ISanti-SGD
F-15
LN-229
Fig
. 4
*
Figure 4
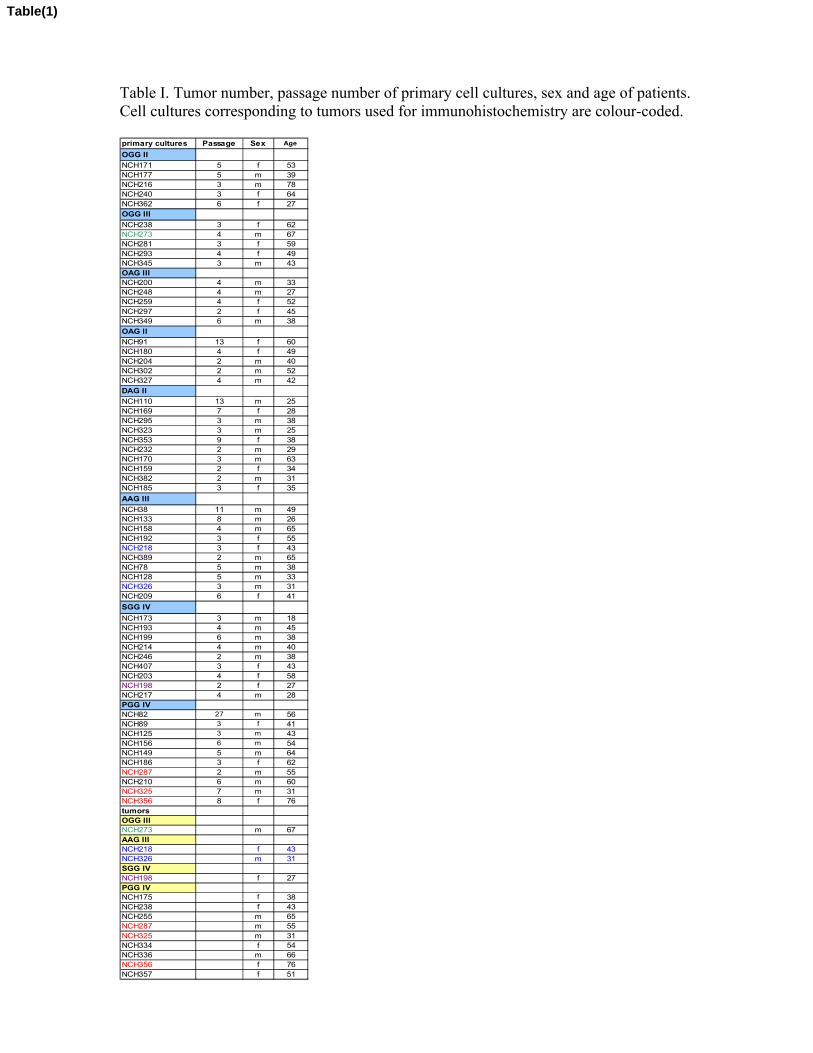
Table I. Tumor number, passage number of primary cell cultures, sex and age of patients. Cell cultures corresponding to tumors used for immunohistochemistry are colour-coded.
primary cultures Passage Sex Age
OGG II
NCH171 5 f 53NCH177 5 m 39NCH216 3 m 78NCH240 3 f 64NCH362 6 f 27OGG III
NCH238 3 f 62NCH273 4 m 67NCH281 3 f 59NCH293 4 f 49NCH345 3 m 43OAG IIINCH200 4 m 33NCH248 4 m 27NCH259 4 f 52NCH297 2 f 45NCH349 6 m 38
OAG II
NCH91 13 f 60NCH180 4 f 49NCH204 2 m 40NCH302 2 m 52NCH327 4 m 42
DAG II
NCH110 13 m 25NCH169 7 f 28NCH295 3 m 38NCH323 3 m 25NCH353 9 f 38NCH232 2 m 29NCH170 3 m 63NCH159 2 f 34NCH382 2 m 31NCH185 3 f 35
AAG III
NCH38 11 m 49NCH133 8 m 26NCH158 4 m 65NCH192 3 f 55NCH218 3 f 43NCH389 2 m 65NCH78 5 m 38NCH128 5 m 33NCH326 3 m 31NCH209 6 f 41
SGG IV
NCH173 3 m 18NCH193 4 m 45NCH199 6 m 38NCH214 4 m 40NCH246 2 m 38NCH407 3 f 43NCH203 4 f 58NCH198 2 f 27NCH217 4 m 28PGG IVNCH82 27 m 56NCH89 3 f 41NCH125 3 m 43NCH156 6 m 54NCH149 5 m 64NCH186 3 f 62NCH287 2 m 55NCH210 6 m 60NCH325 7 m 31NCH356 8 f 76tumorsOGG IIINCH273 m 67AAG IIINCH218 f 43NCH326 m 31SGG IVNCH198 f 27PGG IVNCH175 f 38NCH238 f 43NCH255 m 65NCH287 m 55NCH325 m 31NCH334 f 54NCH336 m 66NCH356 f 76NCH357 f 51
Table(1)


Table II. Origin and p53 status of the nine human glioblastoma cell lines used in our study.
cell line origin p53 status
A172 GB wt
D247MG gliosarcoma wt
LN-18 de novo GB mutant / wt
LN-229 de novo GB wt
LN-308 de novo GB null
LN-319 de novo AA mutant
LN-428 de novo GB mutant
U87MG de novo GB wt
U373MG de novo GB mutant
GB: glioblastoma; AA: anaplastic astrocytoma.
Table(2)

Table III. Number of TUNEL-positive cells / 1000 cells in ten glioblastoma cell lines treated
as indicated. Neither GDF-15 anti-serum nor recombinant human (rh)GDF-15 affect apoptosis
in the glioblastoma cell lines under study.
cell line pre-immuneserum
GDF-15 anti-
serum
rhGDF-15
A172 1 1 <1
D247 1 1 <1
LN-18 1 2 2
LN-229 1 2 1
LN-308 2 3 1
LN-319 4 5 4
LN-428 3 3 2
U87MG 3 4 4
U373MG 4 4 6
Table(3)




![University of Groningen Exploring chemical versatility within · PDF filefor aromatic hydrocarbons.[1-3] 4-OT is a member of the tautomerase superfamily, a group of homologous proteins](https://static.fdocument.org/doc/165x107/5aa239597f8b9ac67a8cd428/university-of-groningen-exploring-chemical-versatility-within-aromatic-hydrocarbons1-3.jpg)