ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ …Bu sonuçlara göre L-12 likenaz enzimi,...
103
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ YÜKSEK LİSANS TEZİ Fatma GÖZÜKARA TERMOFİL Bacillus sp. BAKTERİSİNDEN LICHENAZ (β-1,3 VE 1,4 GLUCANASE) ENZİMİ ÜRETİMİ, KARAKTERİZASYONU VE BİYOTEKNOLOJİK KULLANILABİRLİĞİ BİYOTEKNOLOJİ ANABİLİM DALI ADANA, 2009
Transcript of ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ …Bu sonuçlara göre L-12 likenaz enzimi,...

ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
YÜKSEK LİSANS TEZİ Fatma GÖZÜKARA TERMOFİL Bacillus sp. BAKTERİSİNDEN LICHENAZ (β-1,3 VE 1,4 GLUCANASE) ENZİMİ ÜRETİMİ, KARAKTERİZASYONU VE BİYOTEKNOLOJİK KULLANILABİRLİĞİ BİYOTEKNOLOJİ ANABİLİM DALI ADANA, 2009

ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
TERMOFİL Bacillus sp. BAKTERİSİNDEN LICHENAZ (β-1,3 VE 1,4 GLUCANASE) ENZİMİ ÜRETİMİ, KARAKTERİZASYONU VE BİYOTEKNOLOJİK
KULLANILABİLİRLİĞİ
Fatma GÖZÜKARA
YÜKSEK LİSANS TEZİ
BİYOTEKNOLOJİ ANABİLİM DALI
Bu Tez 06 / 08 / 2009 Tarihinde Aşağıdaki Jüri Üyeleri Tarafından Oybirliği İle Kabul Edilmiştir.
İmza............………...
Prof. Dr. Burhan ARIKAN
DANIŞMAN
İmza............………...
Prof.Dr. Mustafa CANLI
ÜYE
İmza............………...
Doç.Dr. Hatice KORKMAZ
GÜVENMEZ
ÜYE
Bu Tez Enstitümüz Biyoteknoloji Anabilim Dalında Hazırlanmıştır. Kod No:
Prof. Dr. Aziz ERTUNÇ
Enstitü Müdürü İmza ve Mühür
Bu Çalışma Ç. Ü. Rektörlüğü Bilimsel Araştırma Projeleri Birimi Tarafından Desteklenmiştir. Proje No: FEF2008YL27
Not: Bu tezde kullanılan özgün ve başka kaynaktan yapılan bildirişlerin, çizelge, şekil ve fotoğrafların kaynak gösterilmeden kullanımı, 5846 sayılı Fikir ve Sanat Eserleri Kanunundaki hükümlere tabidir.

I
ÖZ
YÜKSEK LİSANS TEZİ
TERMOFİL Bacillus sp. BAKTERİSİNDEN LICHENAZ (β-1,3 VE 1,4 GLUCANASE) ENZİMİ ÜRETİMİ, KARAKTERİZASYONU VE
BİYOTEKNOLOJİK KULLANILABİRLİĞİ
Fatma GÖZÜKARA
ÇUKUROVA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
BİYOTEKNOLOJİ ANABİLİM DALI
Danışman : Prof.Dr. Burhan ARIKAN Yıl : 2009, Sayfa : 91 Jüri : Prof.Dr. Burhan ARIKAN
Prof.Dr. Mustafa CANLI Doç.Dr. Hatice KORMAZ GÜVENMEZ
Bu çalışmada, değişik ortamlardan izole edilen termofil Bacillus sp. suşlarından likenaz enzimi izolasyonu ve karakterizasyonu gerçekleştirilmiştir. Bu amaçla bakterilerin farklı sıcaklık ve pH değerlerinde katı besiyerinde üreme ve enzim üretme yetenekleri araştırılmıştır. İzole edilen enzimin karakterizasyonu gerçekleştirilmiştir.
Bacillus sp. L-12 suşundan üretilen likenazın nativ-PAGE analizinde 100.74 ve 54.4 kDa moleküler ağırlıklarına sahip iki band elde edilmiştir. Enzimin optimum aktivite gösterdiği sıcaklık 100ºC bulunmuştur. Enzim 30-110 ºC aralığında ortalama %66.5 aktivitesini korumuştur. Enzim, optimum aktivitesini pH 6.0’da gösterirken, 24 saat süreyle 55 ºC de pH 3.0-6.0 aralığında ortalama %88 stabilite göstermiştir. L-12 enzimi %7.5 NaCl konsantrasyonunda %67’lik optimum aktivite göstermiştir. Enzim aktivitesi 5 mM EDTA (%97), 5 mM CaCl2 (%95), 3 mM PMSF (%56), %1 v/v triton X-100 (%101), %1 w/v SDS (%38) ve 5mM ZnCl2 ile (%36) korunmuştur.
Bu sonuçlara göre L-12 likenaz enzimi, hipertermofil, termostabil, asidik ve asidostabil, halotolerant, surfaktanlardan etkilenmeyen ve kükürtlü amino asitlerce zengin enzim özelliği göstermektedir.
Enzim optimum aktivitesini asidik pH da gösteren termostabil ve asidik pH stabil özellikte olduğu için, özellikle tavuk yemi hazırlama alanında öncelikle kullanılabilecek özelliktedir.
Anahtar Kelimeler: Bacillus, likenaz, termofil, asidostabil, halotolerant

II
ABSTRACT
M.Sc. THESIS
ISOLATION OF THERMOPILIC Bacillus sp., PRODUCTION, CHARACTERIZATION AND DETERMINATION OF
BIOTECHNOLOGICAL APPLICATION OF LICHENASE (β-1,3 AND 1,4 GLUCANASE)
Fatma GÖZÜKARA
DEPARTMENT OF BIOTECHNOLOGY
INSTITUTE OF NATURAL AND APPLIED SCIENCES UNIVERSITY OF CUKUROVA
Supervisor : Prof.Dr. Burhan ARIKAN
Year : 2009, Page : 91 Jury : Prof.Dr. Burhan ARIKAN
Prof.Dr. Mustafa CANLI Assoc.Prof.Dr. Hatice KORMAZ GÜVENMEZ
In this study, the lichenase enzymes from Bacillus sp. strains isolated from different environments were produced and characterized. For this purpose, different temperatures and pH values of bacteria in solid plate and enzymes to produce reproductive capabilities were investigated. Characterization of the enzyme has been isolated.
Molecular weight of the lichenase from Bacillus sp. LB-12 was estimated as 100.74 and 54.4 kDa on native-PAGE. The temperature of the enzyme shows optimum activity was found 100 ºC. The enzyme was retained it`s activity around %66.5 at the end of 24 hours between 30-110 ºC. The optimum activity was obtained at pH 6.0 the enzyme also presented the enzyme was 88% stable at 55 ºC between pH 3.0-6.0 for 24 hours. The optimum activity of L-12 lichenase has shown 67'lik% in the 7.5% NaCl concentration. The activity of enzyme was protected by 5 mM EDTA (%97), 5 mM CaCl2 (%95), 3 mM PMSF (%56), %1 v/v triton X-100 (%101), %1 w/v SDS (%38) and 5mM ZnCl2 ile (%36).
According to these result, L-12 lichenase is hiperthermophile, thermostable, acidic and asidostable, halotolerant, sulfur-rich amino acids and not affected by surfactants.
For the optimum activity of enzyme is indicating that acidic pH stable and acidic pH thermostable are stable properties, preparation of chicken feed, especially in the first available properties.
Key Words: Bacillus, lichenase, thermostable, acidostable, halotolerant

III
TEŞEKKÜR
Çalışmalarım sırasında ve tez konumun belirlenmesinde bana yardımcı olan
danışman hocam Sayın Prof. Dr. Burhan ARIKAN’a teşekkür ederim.
Çalışmalarım sırasında ve tezimin çalışmalarında bana yardım eden İnci Suna
HÜRTAŞ’a teşekkür ederim.
Çalışmalarım sırasında, iş hayatımda ve manevi hayatta desteğini ve
yardımını esirgemeyen Sayın Uzm. Dr. Hilal ONAÇ’a teşekkür ederim.
Tez aşamasında iyi günde, kötü günde her zaman benim yanımda olan ve
tezimin yazımındaki katkılarından dolayı Murat TAPANYİĞİT ve Mehmet
COŞKUN’a teşekkür ederim.
Her zaman yanımda olan ve benden maddi manevi desteklerini esirgemeyen
değerli aileme özellikle annem Hülya GÖZÜKARA’ya ne kadar teşekkür etsem
azdır.

IV
İÇİNDEKİLER SAYFA
ÖZ ……………………………………………………..……………………..……………..I
ABSTRACT …………………………………………………………………….....………II
TEŞEKKÜR ……………………………………….……………….…………………….III
İÇİNDEKİLER ………………………………….…………………................................IV
ÇİZELGELER DİZİNİ ……………………………………………..….........................VII
ŞEKİLLER DİZİNİ ………...……………………………………………………....…VIII
SİMGE VE KISALTMALAR ………….…………………………….........................….X
1.GİRİŞ ………………………...…..…………..……………………………………......…1
1.1. Proteazlar .…………………………………………………………..………….5
1.2. Ksilanazlar ………………..………………………………….………...........…6
1.3. Amilazlar ………...……………………………………...…..…………………6
1.4. Selülaz (Glukanaz) ………………………….…………………………………7
1.4.1. Glukanaz’ların Uygulama Alanları …………………………………..9
1.4.1.1. Gıda Endüstrisinde Glukanazlar ………………………….10
1.4.1.2. İçecek ve Şarap Endüstrisinde Selülazlar …………….......10
1.4.1.3. Hayvan Yemi Endüstrisinde Glukanazlar ……………..….11
1.4.1.4. Tekstil Endüstrisinde Glukanazlar ………………………..11
1.4.2. β-(1,3)-(1,4)- Glukanaz (Likenaz)’ların Uygulama Alanları …….…11
1.5. Ekstremofilik Enzimler (Ekstremezimler) ………………………………...…12
1.6. Elektroforez Uygulamaları …………………………………………...…...…13
1.6.1. Jel Elektroforezleri ………………………………………………….14
1.6.2. Doğal (Native) Jel Elektroforezi ……………………………………17
2. ÖNCEKİ ÇALIŞMALAR ………………………………………………………..…..18
3. MATERYAL VE METOD …………………………………………………….….…22
3.1. Materyal ……………………………………………………………………....22
3.1.1. Kullanılan Çözeltiler ……………………………………….……….22
3.1.1.1. NaOH Çözeltisi ………………………………………..….22
3.1.1.2. Etanol …………………………………………………..…22

V
3.1.1.3. Kongo Kırmızısı (%0.1) …………………………………..22
3.1.1.4. NaCl Çözeltisi (1M) ………………………………………22
3.1.1.5. Sodyum Fosfat Tamponu (0.1 M) ………………………...23
3.1.1.6. Sitrat Tamponu ……………………………………………23
3.1.1.7. Sodyum-Fosfat Tamponu …………………………………23
3.1.1.8. Glisin-NaOH Tamponu …………………………….......…24
3.1.1.9. DNS (Dinitro Salisilik Asit) ………………………...…....24
3.1.2. Bakteri İzolasyonunda ve Teşhisinde Kullanılan Besiyerleri ………25
3.1.2.1. LB Agar …………………………………………….…….25
3.1.2.2. Likenanlı Besiyeri ………………………………...………25
3.1.2.3. Likenaz Enzimi Üretimi Besiyeri ………………………...26
3.1.3. Elektroforez İşleminde Kullanılan Çözeltiler ………………………26
3.1.3.1. Solüsyon A (Akrilamid Solüsyonu) ………………………26
3.1.3.2. Solüsyon B (4X) ………………………………………….26
3.1.3.3. Solüsyon C (4X) ………………………………………….27
3.1.3.4. Amonyum Persülfat (AMPS) %10 ……………………….27
3.1.3.5. Elektroforez Tamponu ……………………………………27
3.1.3.6. Örnek Yükleme Tamponu (5X) …………………………..27
3.1.3.7. Jel Boyama (Staining) Solüsyonu ………………………..28
3.1.3.8. Jelden Boyayı Geri Alma (Destaining) Solüsyonu ……….28
3.2. Metod …………………………………………………………………………29
3.2.1. Bakteri İzolasyonu ve Teşhisi ………………………………………29
3.2.1.1. Bacillus sp. Suşlarının İzolasyonu ………………………..29
3.2.2. Bakterilerin İdentifikasyonu ………………………………….…….29
3.2.3. Katı Besiyerinde Likenaz Aktivitesinin Saptanması ……….………29
3.2.4. Bakterinin Ürediği ve Enzim Sentezinin Gerçekleştiği pH
Aralığının Saptanması ……......……………………………….........30
3.2.5. Bakterinin Ürediği ve Enzim Sentezinin Gerçekleştiği Sıcaklık ve
pH Aralığının Saptanması ..……………………………………..…30
3.2.6. Enzim Üretimi ve Kısmi Saflaştırma ……………………….………30

VI
3.2.7. Enzimin pH Optimumunun Saptanması …………………………....31
3.2.8. Enzimin Optimum Sıcaklık Aktivitesinin Saptanması …………..…32
3.2.9. Enzimin Sıcaklık Stabilitesinin Saptanması ……………………..…32
3.2.10. Likenaz Enziminin pH Stabilitesinin Saptanması ………………...33
3.2.11. NaCl’ün Enzim Aktiviteleri Üzerine Etkisinin Belirlenmesi ……..33
3.2.12. Enzim Aktivitesine İnhibitör, Şelatör, Deterjan ve Metal
İyonlarının Etkisi ………………………………………….......…..34
3.2.13. Nativ (Doğal) Jel Elektroforezinde Moleküler Ağırlık ve
Zimogram Analizi ….………………………………………..…….34
3.2.13.1. Nativ Jel Sisteminin Hazırlanması ……………………....34
3.2.13.2. Ayırıcı Jel’in Hazırlanması (%10’luk) …………………..34
3.2.13.3. Dengeleme Jel’inin Hazırlanması ……………………….35
3.2.13.4. Enzim Örneklerinin Jele Yüklenmesi ve Yürütülmesi …35
3.2.13.5. Nativ (Doğal) Jelinin Boyanması ve Zimogram
Analizi ….……………………………………………….36
4. BULGULAR VE TARTIŞMA ………….………………………..…………….……37
4.1. Bacillus sp. Suşlarının İzolasyonu …………………………...………………37
4.2. L-12 Suşunun Katı Besiyerinde Üreme ve Enzim Üretme Sonuçları ……..... 38
4.3. Enziminin Optimum pH Aralığına Ait Bulgular ……………………..………52
4.4. L–12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık
Değerine Ait Sonuçlar ….…...………………………………………………..54
4.5. L-12 Likenaz Enziminin pH Stabilitesine Ait Sonuçlar ……...………………57
4.6. L–12 Likenaz Enziminin Termal Stabilite Analizlerine Ait Sonuçlar ……….59
4.7. L-12 Likenaz Enzim Aktivitesi ve Stabilitesi Üzerine NaCl’ün
Etkisine Ait Sonuçlar ……….…………….……………..……………………63
4.8. L-12 Likenaz Enzimi Üzerine İhibitör, Şelatör, Deterjan ve Metal
İyonlarının Etkisine Ait Sonuçlar …………….…………………………........64
4.9. L-12 Likenaz Enziminin Native-PAGE ve Zimogram Analizi …………........66
5. SONUÇ VE ÖNERİLER ….…………………………………………….………...…69
KAYNAKLAR ……...…………………………………………………..…….….........…81
ÖZGEÇMİŞ …….………………......................................................................................91

VII
ÇİZELGELER DİZİNİ SAYFA
Çizelge 4.1. 20°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp.
Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi ……..…....38
Çizelge 4.2. 30°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp.
Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi ..………....40
Çizelge 4.3. 40°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp.
Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi …..……....42
Çizelge 4.4. 50°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp.
Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi …..……....43
Çizelge 4.5. 60°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp.
Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi ………......45
Çizelge 4.6. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği pH
Değerine Ait Bulgular ...…………………….……..…………....52
Çizelge 4.7 L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık
Değerine Ait Bulgular …….…..…...….……….………………..54
Çizelge 4.8. L-12 Likenaz Enzimine Ait pH Stabilite Sonuçları ….………....57
Çizelge 4.9. L-12 Likenaz Enzimine Ait Termal Stabilite Sonuçları ………...60
Çizelge 4.10. Farklı NaCl Konsantrasyonlarının L-12 Likenaz Enzimin
Aktivitesine Etkisi …………………………….………………...62
Çizelge 4.11. L-12 Likenaz Enzimi Üzerine Şelatör, Metal İyonları,
Deterjanlar ve İnhibitörlerin Etkisi ..….…………….…………...64

VIII
ŞEKİLLER DİZİNİ SAYFA
Şekil 4.1.
L-12 suşunun katı besiyerinde üremesi 20°C, pH 5.0, 6.0,
7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm .…....…47
Şekil 4.2. L-12 suşunun katı besiyerinde üremesi 20°C, pH 9.0 ve 10.0 da
gerçekleşen enzim sentezine ait görünüm ...…………….……....47
Şekil 4.3. L-12 suşunun katı besiyerinde üremesi 30°C, pH 5.0, 6.0,
7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm ……......48
Şekil 4.4. L-12 suşunun katı besiyerinde üremesi 30°C, pH 9.0 ve 10.0 da
gerçekleşen enzim sentezine ait görünüm ………………….…...48
Şekil 4.5. L-12 suşunun katı besiyerinde üremesi 40°C, pH 5.0, 6.0, 7.0 ve
8.0 de gerçekleşen enzim sentezine ait görünüm ………..……...49
Şekil 4.6. L-12 suşunun katı besiyerinde üremesi 40°C, pH 9.0 ve 10.0 da
gerçekleşen enzim sentezine ait görünüm ………………………49
Şekil 4.7. L-12 suşunun katı besiyerinde üremesi 50°C, pH 5.0, 6.0, 7.0 ve
8.0 de gerçekleşen enzim sentezine ait görünüm ………….……50
Şekil 4.8. L-12 suşunun katı besiyerinde üremesi 50°C, pH 9.0 ve 10.0 da
gerçekleşen enzim sentezine ait görünüm ………………………50
Şekil 4.9. L-12 suşunun katı besiyerinde üremesi 60°C, pH 5.0, 6.0, 7.0 ve
8.0 de gerçekleşen enzim sentezine ait görünüm ……………….51
Şekil 4.10. L-12 suşunun katı besiyerinde üremesi 60°C, pH 9.0 ve 10.0 da
gerçekleşen enzim sentezine ait görünüm ………………………51
Şekil 4.11. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği pH
Değerine Ait Sonuçlar …..……………………………………....53
Şekil 4.12. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık
Değerine Ait Sonuçlar …………………….………….………....55
Şekil 4.13. L-12 Likenaz Enziminin pH Stabilite Grafiği ..………………...58
Şekil 4.14. L–12 Likenaz Enziminin Termal Stabilite Grafiği ……………..61

IX
Şekil 4.15. L-12 Likenaz Enzim Aktivitesi Üzerine Farklı NaCl
Konsantrasyonlarının Etkisi …..………………………………...63
Şekil 4.16. L12 Likenaz Enzimine Metal İyonu, Şelatör, İnhibitör ve
Deterjanların Etkisi …………….……………………………….66
Şekil 4.17. L-12 Likenaz Enziminin Molekül Büyüklüğü ve Zimogram
Analizi ..………………………………………………….……...68

X
SİMGELER VE KISALTMALAR
AMPS : Amonyum persülfat DNS : Dinitrosalisilikasit EDTA : Etilendiaminotetraasetik asit L : Litre LB : Luria Bertani M : Molar mL : Mililitre mM : Milimolar mg : Miligram µg : Mikrogram µL : Mikrolitre SDS : Sodyum dodesil Sülfat α : Alfa β : Beta rpm : dakikada devir sayısı Tris : 2-Amino-2-Hidroksimetilpropan-1,3-diol TEMED : N,N,N,N-Tetrametil etilendiamin TAE :Tris-Asetik Asit-Edta

1. GİRİŞ Fatma GÖZÜKARA
1
1.GİRİŞ
Biyoteknoloji, çok çeşitli alanlarda gelişme gösteren ve günümüzde moleküler
biyolojik yöntemlerinde yaygın şekilde kullanımıyla birlikte, giderek moleküler
biyoteknoloji şeklinde transformasyona uğrayan ve çağımıza damgasını vuran bir
alandır.
Dünyada, 1980-1983 yılları arasında sadece 300 küçük biyoteknoloji şirketi
çalışma gerçekleştirirken, bu sayı 1985 yılında sadece Amerika Birleşik
Devletlerinde (A.B.D) 400 düzeyine ulaşmıştır. Günümüzde A.B.D de 900, bütün
dünyada ise yaklaşık 1200 biyoteknoloji şirketi çalışmalarını çeşitli alanlarda
sürdürmektedirler. Örneğin sadece farmasötik alanında 2000’li yıllarda kullanılacak
biyoteknolojik ürünlerin toplam değerinin yaklaşık 60 milyar dolar/yıl düzeyinde
olacağı düşünülmektedir. Biyoteknoloji kaynaklı çalışmalar A.B.D. de odaklanmış
olmakla birlikte, günümüzde, Japonya ve Kanada, biyoteknolojiyi (özellikle
moleküler biyoteknolojiyi) stratejik alan kategorisinde değerlendirerek, özel
şirketlerin yanı sıra, hükümetler düzeyinde destekleme ve geliştirme kararı
almışlardır (Glick ve Pasternak, 1994).
Enzim teknolojisinin giderek gelişmesi, ürünlerin kullanım alanlarının
çeşitliliği ve ekonomik değerinin çok yüksek olması nedeniyle, biyoteknolojinin
endüstriyel enzimlerle ilgili alanında yapılan çeşitli araştırmalar, daha da önem
kazanmaktadır. Özellikle birkaç ülke dışında diğer ülkelerin bu konuda tamamen
dışa bağımlı durumdadırlar.
Enzimler, doğal olarak canlılar tarafından sentezlenen protein yapısında ya da
bir kısmı protein olan biyo-moleküllerdir. Enzimler, binlerce yıldır içecek, ekmek
ve peynir yapımı gibi işlemlerde varlığı ve görevi bilinmeden kullanılmıştır. İlk
bulgular eski Mısıra kadar dayanmaktadır. Yakın tarihte ise Doğu ülkelerinde
birçok gıda fermentasyonu için ipliksi mantarlar enzim kaynağı olarak
kullanılmaktadır. Batıda 1896’da gerçek modern mikrobiyal enzim teknolojisi
‘takadiastase’ın ticareti ile başlamıştır. Bu Doğudan Batı toplumuna önemli bir
teknolojik transferdir (Uhlig, 1998).

1. GİRİŞ Fatma GÖZÜKARA
2
Dericilikte, derinin yumuşatılması köpek ya da güvercin dışkıları ile muamele
edilerek yapılırken, bu yüzyıl başlarında Alman kimyacı ‘Otto Röhm’ köpek
dışkılarındaki aktif bileşenlerin proteinleri parçalayan proteaz enzimi olduğunu ve
hayvansal organlardan bu enzimin elde edilebileceğini ve bu işlemlerde köpek
dışkısı yerine kullanılabileceğini ortaya koymuştur. Böylece 1905’den itibaren
domuz ve sığır pankreasları sosyal açıdan ve güvenilir bir enzim kaynağı olarak ön
plana çıkmıştır (Smith, 1996). Bitkisel kaynaklı enzimlerden bira üretiminde
kullanılan malt amilazı içecek endüstrisinde önemli bir yer tutmaktadır (John,
1987).
Tarihsel gelişim açısından bakıldığında enzimlerin çok farklı kaynaklardan
elde edildiği görülmektedir. Bunlar bitkisel, hayvansal ya da endüstriyel anlamda
ihtiyacı karşılayabilen mikrobiyal kaynaklı enzimlerdir. Bugüne kadar yaklaşık
2500 farklı enzim tanımlanmış ve bunların ancak %10’u ticari alanda kullanım için
kendilerine yer bulmuşlardır. Bunlardan ise ancak 25 tanesi nişasta sanayi ile
deterjan katkı maddesi olarak kullanılmış olup, ticari alanda yararlanılan bütün
enzimlerin %80’ini oluşturmaktadırlar (Rao ve ark., 1998). Bitkisel ve hayvansal
enzimlerin endüstriyel ihtiyacı karşılayamaması, bu alandaki ilginin giderek artan
bir şekilde mikrobiyal enzimlere yönelmesini sağlamıştır. Mikroorganizmalar,
biyokimyasal çeşitlilikleri ve genetik manipülasyonlara uygunluğu gibi sebeplerden
dolayı mükemmel bir enzim kaynağı olarak değerlendirilmektedir (Rao ve ark.,
1998). Günümüzde endüstride kullanılan enzimlerin yaklaşık %90’ı
mikroorganizmalardan üretilmektedir (Wolfgang, 2004).
Mikrobiyal enzimlerin dünya genelinde yıllık kullanım değerlerine bakıldığı
zaman, alkaline proteaz %25, diğer proteazlar %21, Amilaz %18, Renin %10,
Trypsin %3, Lipaz %3, diğer karbonhidrat parçalayan enzimler (selülaz ve ksilanaz
gibi) %10, analitik ve farmasötik enzimler %10 şeklinde bir dağılımla
karşılaşılmaktadır (Rao ve ark., 1998).
Endüstriyel alanda kullanılan enzimler bitkisel, hayvansal ve mikroorganizma
kökenli olmakla birlikte ağırlıklı olarak mikroorganizmalardan izole
edilmektedirler. Bunun nedeni, mikroorganizma kaynaklı enzimlerin katalitik
aktivitelerinin yüksek olması, istenmeyen yan ürün oluşturmamaları, daha stabil ve

1. GİRİŞ Fatma GÖZÜKARA
3
daha ucuz olmaları, büyük boyutlarda ve yüksek saflıkta elde edilmesi gibi
avantajlara sahip olmasıdır. Örneğin mikrobiyal enzimler ekstrem sıcaklık ve pH
değerlerinde çok yüksek düzeyde aktivite gösterirler (Wiseman, 1987; Horikoshi,
1999).
Endüstride faydalı olan ve ticari olarak kullanılan enzimlerin, bir ürünün
üretilmesinde bazı avantajlara sahip olması gerekmektedir. Buna göre, bir enzimin
herhangi bir endüstri alanında kullanılabilmesi maliyet bakımından ucuz olması,
çok farklı alanlarda kullanılabilme özelliğinde olması ve en önemlisi de enzimin
alerjik ya da toksik etkiye sahip olmaması, yani güvenilir olmasına bağlıdır
(Wiseman, 1987; Sarıkaya, 1995).
Endüstride kullanılan enzimlerin yaklaşık %75’ini hidrolitik enzimler
oluşturmaktadır. Proteaz grubundaki enzimler kullanımda ilk sırayı alırken,
karbonhidratları parçalayanlar ikinci sırada yer almaktadırlar (Hamilton ve ark.,
1999; Bhat, 2000). Karbonhidrat parçalayan enzimlerden amilazlar, enzim
piyasasında yaklaşık %25’lik bir paya sahiptir (Sindhu ve ark.,1997; Rao ve
ark.,1998). Son zamanlardaki en etkin uygulamaları kağıt hamuru beyazlatılması
olan ksilanazların endüstride kullanılması hızlı bir yükselişe geçmiştir (Wainq ve
Ingworsen, 2003).
Hidrolazlardan selülazlar ise selülozik materyalleri şekerlere dönüştürerek
özellikle biyo-etanol gibi biyolojik temelli yakıt katkı maddelerinin üretiminde
önemli bir yer tutmaya başlamışlardır. Bu nedenle selülaz’ların (glukanaz) enzim
piyasasındaki paylarının ciddi bir şekilde artması beklenmektedir. Sadece bitkisel
atıkların hidrolizinde kullanılması durumunda bile, selülaz enzimlerinin Amerika
Birleşik Devletleri’nde yıllık 400 milyon dolarlık bir ticaret hacmi yaratacağı
öngörülmektedir (Boyce ve Walsh, 2007).
Dünya geneli incelendiğinde endüstriyel enzimlerin ticari pazar payının
yaklaşık 1.6 milyar dolar olduğu tahmin edilmektedir. Bu enzimlerin kullanım
alanlarına göre dağılımına bakıldığında ise, %29’unun gıda endüstrisi, %15’inin
hayvan yemi sektörü ve %56’sının genel amaçlı teknik alanlar şeklinde dağıldığı
görülmektedir (Uhlig, 1998).

1. GİRİŞ Fatma GÖZÜKARA
4
Endüstride yaygın şekilde kullanılan enzimler arasında Bacillus cinsine ait
tür ve alttürleri tarafından sentezlenen ondan fazla enzim vardır. Örneğin ticari
olarak üretilen ve kullanılan termostabil amilaz enzimlerinin üretilmelerinde
Bacillus amyloliquefaciens ve Bacillus licheniformis en çok kullanılan iki Bacillus
türüdür (Horikoshi, 1996; Lin ve ark., 1998; Horikoshi, 1999; Kumar ve Takagi,
1999).
Bacillus cinsi bakteriler toprak, hayvan dışkıları ve bitkisel atıklar üzerinde
yaygın olarak bulunurlar. Bu cinsin bireylerinin çoğu zararsız, izolasyonu ve teşhisi
kolay, hızlı büyüme oranı ile fermentasyon süresi kısadır.
Genel olarak güvenli olmaları, sentezledikleri proteinleri dış ortama salgılama
kapasiteleri gibi birçok nedenden dolayı cazip endüstriyel organizmalardır. Çünkü,
gram negatif bakteriler ürettikleri proteinleri protoplazmalarında yada periplazmik
boşluklarında biriktirirler ve bu da üretilen ürünün izolasyonunu güçleştirerek
suştan birden fazla kez yararlanılmasını engeller. Ayrıca sentezlenen ürünlerin
organizmaya karşı toksik etki oluşturması söz konusudur. İntrasellüler ortamda
sentez ürünlerinin biriktirilmesi çözünmeyen protein oluşumu, yanlış protein
katlanmaları ve etkin olmayan disülfit bağ formasyonu gibi sorunları beraberinde
getirmektedir. Ayrıca gram negatif bakteriler, insanlara toksik olan endotoksin ve
intrasellüler protein üretmeleri nedeniyle, proteinlerin izolasyonu ve
saflaştırılmasında ekstra maliyet oluşturmaktadırlar (Boyce ve Walsh, 2007).
Mantarlardaki aflatoksin gibi toksik yada alerjen bileşik üretimi de göz
önünde bulundurulduğunda, gram pozitif bakteriler, özellikle Bacillus türleri
endüstriyel enzim üretiminde öncelikli olarak tercih edilmektedirler.
Bacillus sp. suşları, proteolitik enzimler ve karbonhidratazların en önemli
kaynağını oluşturmaktadırlar. Örneğin α-amilaz enzimi Bacillus sp. grubunda
oldukça yaygın şekilde bulunur ve başlıca, gıda endüstrisinde nişastanın maltoza
hidroliz edilmesi, maltoz şuruplarının hazırlanması, ekmek ve bira üretiminde
kullanılmaktadır. Diğer taraftan, gıdalarda tatlandırıcı ve kaliteyi arttırıcı olarak
(Anonymous, 1988a), tekstil endüstrisinde haşıl alma işlemlerinde (Tarakçıoğlu,
1979) ve alkol fermantasyonunda yaygın bir kullanım alanı vardır (Bhat, 2000).
Özellikle termofilik alkali amilazlar, pH 9’un üzerinde yüksek düzeyde aktif olup,

1. GİRİŞ Fatma GÖZÜKARA
5
deterjan endüstrisinin en önemli katkı maddesini oluşturmaktadırlar. Bunların
dışında kağıt ve tekstil endüstrisi, çeşitli içeceklerin saflaştırılması ve
berraklaştırılması gibi geniş bir kullanım alanına sahiptirler (Rao ve ark., 1998).
Bacillus türleri çeşitli kompleks substratlara karşı aktivite gösteren çok sayıda
ve çeşitli hidrolitik enzimler üretmekte ve salgılamaktadırlar. Bu nedenle Bacillus
cinsindeki organizmalar, endüstriyel alanda α-amilaz, proteaz, glukanaz, glukoz
izomeraz ve endonükleaz gibi enzimlerin üretiminde yaygın şekilde
kullanılmaktadırlar (Uhlig, 1998).
Bu enzimler arasında endo β-(1,3)-(1,4)- glukanazlar (likenazlar), endosperm
hücrelerinin nişastasına α-amilazların ulaşımını kolaylaştırdığından dolayı içki
mayalama endüstrisinde spesifik uygulamalara sahiptirler (Beckmann ve ark.,
2006). β -1,3-1,4 glukanlar, yüksek yapılı bitkilerin hücre duvarlarının yapısında
bulunan hem β-1,3 hemde β-1,4 bağlı D-glukoz içeren lineer polisakkaritlerdir.
Likenazların, β-glukanları diğer glukanazlara göre daha yüksek düzeyde hidroliz
etmeleri nedeniyle, kümes hayvanları ve domuz yemlerinin hazırlanmasında
giderek ön plana çıkmaktadırlar (Beckmann ve ark., 2006).
1.1. Proteazlar
Proteazlar, doğada bitkisel, hayvansal ve mikrobiyal kalıntıların
dekompozisyonunda önemli rol oynayarak besin döngüsününün sürekliliğini
sağlayarak bitkilerin besinlerini alabilmelerini sağlamaktadır (Aoki, 1995).
Mikrobiyal proteazlar geniş pH aralıklarında aktivite gösterdikleri için asidik, nötral
ve alkali olmak üzere 3 gruba ayrılmışlardır. Proteaz enzimi üretimi birçok bakteri
ve mantar türünde oldukça yaygındır. Bununla birlikte Bacillus cinsinde yer alan
çeşitli suşlar enzim endüstrisinde çok önemli bir yere sahiptirler (Fogarty ve Kelly,
1979; Aunstrup, 1981).

1. GİRİŞ Fatma GÖZÜKARA
6
1.2. Ksilanazlar
Ksilan, yüksek bitkilerin hücre duvarının hemiselülozik kısmının temel
bileşeni olup kullanılırlığı dikkate alındığında doğada en fazla bulunan ikinci
kaynaktır (Christov ve ark.,1996; Gessesse, 1997). Pek çok bakteri ve mantar suşu
ksilanı parçalayabilmek için ksilanaz enzimine gereksinim duyarlar (Gamerith ve
ark., 1992; Gessesse ve Gashe, 1997). Son on yıldır, ksilan ve ksilenaz enzimlerinin
endüstriyel uygulamaları araştırıcıların giderek ilgisini çekmektedir. Enzimin
kullanım alanlarına örnek olarak kağıt hamuru hazırlama sistemleri ve kağıt
endüstrileri, gıda ve hayvan yemi endüstrileri verilebilir (Kulkarni ve ark., 1999).
1.3. Amilazlar
Karbohidratazların en önemli kaynağını Bacillus cinsindeki organizmalar
oluşturmaktadır. Bir karbohidraz olan α-amilaz enzimi, ticari olarak kullanılan ilk
enzimdir (Radley, 1976). Amilazlar, nişasta ve glikojen moleküllerini hidrolize
ederek glikoz, maltoz, maltotrioz ve α-limit dekstrinlerin oluşmasını sağlayan
ekstraselüler enzimlerdir. Bu enzimler hayvanlar ve bitkiler tarafından
sentezlenmesine rağmen, uygun koşullarda kısa sürede elde edilmesinden dolayı
asıl kaynağı mikroorganizmalar oluşturmaktadır. Fungal α-amilazların
termostabiliteleri bakteriyel α-amilazlardan daha düşüktür. Araştırmalar daha çok
bakteriyel özellikle de Bacillus amilazları üzerinde yoğunlaşmıştır (Wiseman,
1987).
Bacillus cinsine ait 8 suştan izole edilen α-amilaz enzimi çeşitli araştırıcılar
tarafından tanımlanmış ve bütün özellikleri karakterize edilmiştir. Bu suşlar B.
subtilis (Coleman ve Elliott, 1962.; Pazur, 1965.; Matsuzaki ve ark., 1974), B
.amyloliquefaciens (Borgia ve Campbell, 1978), B. caldolytcus (Grootegoed ve ark,
.,1973), B.coagulans (Bliesmer ve Hartman., 1973), B. licheniformis (Meer, 1972.,
Saito, 1973), B. macerans (Lane ve Pirt, 1973), B. stearothermophilus (Ogasahara
ve ark., 1970) ve B. subtilis var. amylosacchariticus (Matsuzaki ve ark., 1974)’dur.
Amilolitik enzimler bakteri ve mantarlarda oldukça yaygındır. Eskiden amilaz

1. GİRİŞ Fatma GÖZÜKARA
7
terimi amilaz ve amilopektinin α-1,4-glukozidik bağlarını hidrolize eden endo-
amilaz enzimler için kullanılırdı.
Günümüzde amilazlar 3 grup altında sınıflandırılmaktadırlar. Bunlar ekzo-
amilaz, endo-amilaz ve dallanma göstermeyen amilaz enzimleridir (John, 1987).
Sıradışı bakteriyel amilazlar asidofilik, alkalifilik ve termoasidofilik bakterilerde
bulunmuştur (Boyer ve ark., 1972). Termoasidofilik ve alkalifilik bakterilerden elde
edilen enzimler ekstrem pH ve sıcaklık koşullarında kullanıma uygundur (Kindle,
1983). Çok yüksek sıcaklıklarda (80-90°C) optimal aktiviteye sahip olan α-amilaz
enzimi termofil ortamda üreyen B.licheniformis tarafından üretilmektedir (John,
1987).
1.4. Selülaz (Glukanaz)
Selüloz, doğada en bol bulunan yenilenebilir bir biyopolimer olup yaygın
olarak bitki hücre duvarlarında bulunur. Glikoz birimlerinin β-1,4 glikozidik
bağlarla bağlanmasıyla oluşan polimerlerdir. Nişastadan farklı olarak düz bir zincir
şeklinde olup herhangi bir sarmal yapı göstermez. Selüloz zincirleri hidrojen
bağları ile bağlanarak bir arada tutulurlar. Bazı durumlarda selüloz neredeyse saf
olarak bulunur. Birçok durumda ise selüloz lifleri hemiselüloz ve lignin gibi diğer
yapısal biyopolimerler arasına katılabilmektedir. Selülozun önemli bir özelliği,
diğer polisakkaritlerden farklı olarak kristal yapı oluşturabilmeleridir. Doğada
glikoz birimleri bir zincir halinde sentezlenirken biyosentez bölgesinde
kendiliğinden birleşerek elementer fibril olarak adlandırılan yaklaşık 30 selüloz
zincirinden oluşan birimlere dönüşür. Bunlar da daha geniş üniteler halinde
paketlenerek mikrofibril denen yapıları oluştururlar. Mikrofibriller ise selüloz
fibrillerini oluştururlar. Hidrojen bağları, zincirleri zincir içi ve zincirler arası
bağlarla birbirlerine bağlar ve sert bir yapı oluşmasını sağlar.
Selüloz, β-1,4 bağlarıyla birbirine bağlanmış glikoz moleküllerinden oluşmuş
bir homopolimerdir (Bath, 2000). Selülozun bakteriyel parçalanmasında hem
aerobik hem de anaerobik mikroorganizmalar görev yapmaktadır. Bu proses
sırasında selüloz glikoza kadar parçalanır. Bakterilerin selülolitik enzimleri ekzo-β-

1. GİRİŞ Fatma GÖZÜKARA
8
1,4 ve endo-β-1,4 glukanazlar şeklinde tanımlanmışlardır (Bath, 2000).
Glukanazlar endo ve ekzo glukanazlar şeklinde ya β-1,3 veya β-1,4 bağlarına
etki göstererek glukan molekülünü parçalarken, likenaz enzimi β-1,3 ve β-1,4
bağlarını tek başına parçalayarak etki göstermektedir. Endoglukanazlar selüloz
molekülünde zincirin iç bölgelerinde rastgele hidroliz yaparlar ve değişik
uzunluklarda oligosakkaritler meydana getirirler. Ekzoglukanazlar ise selüloz
molekülünde zincirin indirgenen ve indirgenmeyen ucundan itibaren hidroliz
yaparak glukoz (glukanohidrolaz) yada sellobiozu (Sellobiohidrolaz) ana ürün
olarak açığa çıkarır. Ekzoglukanazlar aynı zamanda mikrokristal selüloz üzerine de
etki gösterirler. β-Glukozidazlar sellodekstrin ve sellobiyozu glukoza parçalarlar.
Genellikle bir selülaz sistemi birden fazla farklı enzimden oluşmaktadır. Üç
ana enzimatik aktivite tipi bulunmaktadır.
1.Endoglukanazlar (endo-β-1,4-glukanazlar, veya β-1,4-D-glukan-4-
glukanohidrolazlar, EC 3.2.1.4).
2.Ekzoglukanazlar (Sellodekstrinazlar, β-1,4-D-glukan glukanohidrolazlar)
(EC 3.2.1.74) ve Sellobiyohidrolazlar (ekzo-β-1,4-glukanazlar, veya β-1,4-D-
glukan sellobiyohidrolazlar, EC 3.2.1.91).
3.Sellobiyazlar (β-glukosidazlar, veya β-D-glukozid glukohidrolazlar, EC
3.2.1.21).
Endo-(1,3-1,4)- β- glukanaz enzimi, arpa ve yulaf gibi tahılların endosperm
hücre duvarlarının büyük kısmını oluşturan karma bağlı (1,3-1,4)-β-glukanları
hidrolize edebilme özelliğindedir. β- glukanaz üretme yeteneği B. subtilis, B.
amyloliquefaciens, B. macerans ve B. licheniformis’te oldukça yaygın olup,
sentezden sorumlu genler diğer organizmalarda klonlanmışlardır (Liming ve
Xueliang, 2004).
Endo-(1,3-1,4)- β- glukanaz enzimi bira üretimi sırasında malt enzimlerinin
daha iyi çalışmasını sağlamak için yoğun şekilde kullanılmaktadır. Termostabl β-
glukanazlar, maltın kurutulması sırasında inaktive olmadıkları için özellikle tercih
edilirler. Maltlanmış arpada bulunan β-1,3-1,4-glukanaza benzer özgüllük gösteren
bakteriyel β-1,3-1,4-glukanaz önemli bir endüstriyel enzimdir ve çoğunlukla ezme

1. GİRİŞ Fatma GÖZÜKARA
9
işlemi esnasında akışkanlığı azaltmak için kullanılır. Hayvan yeminde, özellikle
ızgaralık piliç ve küçük domuzlarda, bakteriyel β-1,3-1,4-glukanazlar içeren
enzimatik preparatlar ilavesi arpa kökenli beslenmenin sindirilebilirliğini
geliştirebilir ve sağlıksal sorunları azaltabilir (Zhang ve ark., 2006).
Gelecekte termofil özellik gösteren extremophile mikroorganizmaların enzim
üretim alanında mikrobiyal fabrikalar şeklinde kullanılacakları ve bu şekilde
adlandırılacakları belirtilmektedir (Niehaus ve ark., 1999).
1.4.1. Glukanaz’ların Uygulama Alanları
Hayvan yemlerine enzim katılması ile ilgili ilk uygulama 1980 yılında
Finlandiya’da yapılmıştır. β–glukanaz enziminin arpadan üretilen tavuk yemlerine
karıştırılması ile birlikte, özellikle tavukçuluk sektöründe enzim kullanımı önemli
boyutlara ulaşmıştır. Bu uygulamayı domuz yemleri izlemiştir. Ruminantlar
uygulamanın nispeten daha az gerçekleştirildiği sektör olmuştur. Hayvan yemi
üretiminin boyutları düşünüldüğü zaman enzimler için çok büyük bir potansiyel
ortaya çıkmaktadır. Tek mideli hayvanların besinlerinde şuan yaklaşık %10’luk bir
uygulama söz konusudur (Wolfgang, 2004).
Özellikle hayvan yemi üretiminde tavukçuluk sektörünün hızlı gelişimine
paralel olarak yem gereksinimi önemli boyutlara ulaşmıştır. Tavuk yemi üretiminde
ana hammadde olarak mısırdan yararlanılmakla birlikte buğday, arpa ve çavdar da
kullanılmaktadır. Son yıllarda biyoteknolojik çalışmalarda çok hızlı ivme kazanan
diğer bir çalışma alanını biyoetanol oluşturmaktadır. Etanol üretiminde hammadde
olarak mısırın daha çok tercih edilmesi mısır fiyatlarının artmasına, dolayısı ile yem
fiyatlarının yükselmesine neden olmuştur.
Mısır yerine alternatif yem hammaddesi olarak arpa ön plana çıkmaktadır.
Tavuklarda arpada bulunan β-glukanı parçalayacak enzim sistemi yeterince
gelişmediği için yem üretiminde enzim kullanımı zorunluluğu ortaya çıkmıştır.
Özellikle glukanaz ve xylanase enzimi bu konuda ön plana çıkmaktadır (Wolfgang,
2004).

1. GİRİŞ Fatma GÖZÜKARA
10
β–glukan β–1,3 ve β–1,4 şeklinde bağlanmış lineer glikoz birimlerinden
oluşur. Likenaz enzimi bu bağların hem β-1,3 hemde β-1,4 bağlarını kopartarak etki
gösterir.
Bakteriler arasında Cellulomonas sp., Bacillus sp. ve Thermoactinomyces sp.
ekstraselüler selülolitik akviteye sahip organizmaları oluşturur. Mantarlar arasında
Trichoderma reseii en önemli selülaz üreticisidir. Arpanın yaklaşık %3-4’ü β-
glukandan oluşmaktadır. β-glukan tavuk ve domuzlar tarafından sindirilemediği
için sindirim sisteminde viskos bir yapı oluşmakta ve dışkının yoğunluğu
artmaktadır (Wolfgang, 2004).
1.4.1.1. Gıda Endüstrisinde Glukanazlar
Tohumlardan yağ ve meyve suyu ekstraksiyonunda, meyve sularının
berraklaştırılmasında, tahılların homojen olarak su çekmesini sağlamak ve yeterince
ıslanmasının artırılmasında, soya sosu gibi fermente soya gıdaların üretiminde
soyanın dış zarının uzaklaştırılmasında, kokonat ve soya fasulyesinden protein
izolasyonunda, mısır ve tatlı patatesten nişasta üretiminde, sindirimini artırmak
amacı ile yosunlarının jelatinizasyonunda, su yosunlarından agar ekstraksiyonunda,
gıda katkısı maddesi olarak kullanılan öğütülmüş lignoselülozik materyalin
parçalanmasında, selülozik atıklardan çözünür şeker, glikoz ve sello-oligosakkarit
üretiminde, bioetanol üretimi için substrat eldesinde, kurutulmuş sebze ve çorba
karışımlarının geri sulandırımının artırılmasında, polisakkarit, protein, enzim ve tat
verici maddelerin açığa çıkışını kolaylaştırmak amacıyla bitki hücre duvarlarının
uzaklaştırılmalarında kullanılmaktadır (Bhat ve Bhat, 1997).
1.4.1.2. İçecek ve Şarap Endüstrisinde Glukanazlar
Rekombinant mayalardan elde edilen β-1,3 ve β-1,4 glukanazlar (likenazlar)
şaplarda aroma artışının sağlanması, biranın filtrasyonunun kolaylaştırılması ve
düşük kalite arpada bulunan β-1,3 ve β -1,4 glukan hidrolizinde kullanılmaktadırlar.

1. GİRİŞ Fatma GÖZÜKARA
11
1.4.1.3. Hayvan Yemi Endüstrisinde Glukanazlar
Ruminant ve monogastrik hayvanlar yemlerinde sindirilebilirliği artırmak
amacı ile, lignoselülozik materyallerin ön işlemden geçirilmesinde, hububatların
kabuklarından arındırılmasında, ruminant ve monogastrik hayvanların selülozda
yararlanmalarını artırmak için silaj yapımında kullanılırlar.
1.4.1.4. Tekstil Endüstrisinde Glukanazlar
Kumaşlarda boyanın fazlasını almada (biostoning), bir çok yıkama sonunda
pamuk kumaşlardan çıkan mikrofibrillerin giderilmesinde, pamuk yada pamuklu
kumaşların yıkanması, renk parlaklığının artırılması ve yumuşaklığının geri
kazandırılmasında kullanılır (Bhat ve Bhat, 1997).
1.4.2. β-(1,3)-(1,4)- Glukanaz (Likenaz)’ların Uygulama Alanları
Bacillus türleri çeşitli kompleks substratlara karşı aktivite gösteren çok sayıda
ve çeşitli hidrolitik enzimler üretmekte ve salgılamaktadırlar. Bu nedenle Bacillus
cinsindeki organizmalar, endüstriyel alanda α-amilaz, proteaz, glukanaz, glukoz
izomeraz ve endonükleaz gibi enzimlerin üretiminde yaygın şekilde
kullanılmaktadırlar (Uhlig, 1998). Bu enzimler arasında endo β-(1,3)-(1,4)-
glukanazlar (likenazlar), endosperm hücrelerinin nişastasına α-amilazların ulaşımını
kolaylaştırdığından dolayı içki mayalama endüstrisinde spesifik uygulamalara
sahiptirler (Beckmann ve ark., 2006). β -1,3-1,4 glukanlar yüksek yapılı bitkilerin
hücre duvarlarının yapısında bulunan hem β-1,3 hemde β-1,4 bağlı D-glukoz
içeren lineer polisakkaritlerdir. Likenazların, β-glukanları diğer glukanazlara göre
daha yüksek düzeyde hidroliz etmeleri nedeniyle, kümes hayvanları ve domuz
yemlerinin hazırlanmasında giderek ön plana çıkmaktadırlar (Beckmann ve ark.,
2006).
Endo-(1,3-1,4)- β- glukanaz enzimi bira üretimi sırasında malt enzimlerinin
daha iyi çalışmasını sağlamak için yoğun şekilde kullanılmaktadır. Termostabl β-

1. GİRİŞ Fatma GÖZÜKARA
12
glukanazlar, maltın kurutulması sırasında inaktive olmadıkları için özellikle tercih
edilirler. Maltlanmış arpada bulunan β-1,3-1,4-glukanaza benzer özgüllük gösteren
bakteriyel β-1,3-1,4-glukanaz önemli bir endüstriyel enzimdir ve çoğunlukla ezme
işlemi esnasında akışkanlığı azaltmak için kullanılır. Hayvan yeminde, özellikle
etlik tavuk ve küçük domuzlarda, bakteriyel β-1,3-1,4-glukanazlar içeren enzimatik
preparatlar ilavesi arpa kökenli beslenmenin sindirilebilirliğini geliştirebilir ve
sağlıksal sorunları azaltabilir (Zhang ve ark., 2006).
1.5. Ekstremofilik Enzimler (Ekstremezimler)
Ekstremofilik enzimler sıcaklık, basınç, pH ve tuzluluk gibi ekstrem şartlarda
çalışabilen ve endüstriyel uygulamalar için büyük öneme sahip enzimlerdir.
Mezofilik enzimler enzim stabilitelerindeki eksikliklerden dolayı endüstriyel
enzimlerden istenen zorlu reaksiyon şartları için, pek uygun değillerdir.
Ekstremofilik organizmlar ise ekstrem ortamlarda bulunabilen organizmalar
olup termofil, asidofil, halofil, alkalofil, psikrofil gibi bir çok farklı sınıflar altında
grublandırılabilirler. Dolayısı ile bu organzimalar, mezofillerin yaşayamlarını
sürdüremiyecekleri koşullarda aktivite gösteren enzimleri üretebilirler.
Mikroorganizmalar optimum büyüme sıcaklıkları dikkate alındığında
psikrofiller (20oC altında), mezofiller (20-55oC) ve termofiller (55oC üzeri) olmak
üzere üç ana gruba ayrılırlar. Bunlara ilaveten Kristjansson ve Stetter (1992)’
termofil grubu daha da genişletilerek 60-80oC arasında üreyenleri ekstermofil,
80oC’nin üzerinde üreyenler için hipertermofil tanımını kullanılmaktadırlar.
Gerek termofilik gerekse hipertermofilik enzimler karakteristik olarak
40oC’nin altında etkin bir aktivite göstermezler (Gomes ve Steiner, 2004). Sıcaklığa
dirençli proteinlerin amino asit kompozisyonu mezofilik organizmaların amino asit
kompozisyonu ile karşılaştırıldığında glisin yerine alanin, lizin yerine arjinin amino
asitinin sıcaklığa dirençli proteinlerde daha fazla yer aldığı görülmüştür. Aynı
zamanda arjinin termofilik proteinlerin yapısında bol bulunurken sıcaklığa en
hassas amino asit olan sistein bu proteinlerin yapısında daha sınırlı düzeyde
bulunmaktadır.

1. GİRİŞ Fatma GÖZÜKARA
13
Deneysel çalışmalar hidrofobik interaksiyonların termofilik proteinlerin
stabilizasyonunda önemli bir rol aldığını göstermiştir. Sıcaklığa dirençli proteinler
hidrofobik interaksiyonlarla dimer oluşturarak termal hidrolize karşı daha dirençli
hale geçmektedir. Bunlardan başka sıcaklığa dirençli proteinler disülfit bağları,
molekül içi aromatik yapılar, zengin hidrojen bağları içermekte ve elektrostatik
mekanizmalarla yüksek sıcaklığı tolere edebilmektedirler (Kumar ve Nussinov,
2001).
Deterjan, gıda, yem, nişasta, tekstil, deri, kağıt hamuru, farmasötik endüstrisi,
termofilik enzimlerin en geniş kullanıldığı alanlardır (Gomez ve Steiner, 2004).
Örneğin nişasta endüstrisi, termostable amilazın en yaygın kullanıldığı alandır.
1.6. Elektroforez Uygulamaları
İyonlaşabilir gruplara sahip amino asitler, peptidler, proteinler, nükleotidler ve
nükleik asitler gibi biyolojik moleküller, ayırıcı bir ortamın bulunduğu elektriksel
alanda, sahip oldukları elektrik yüküne bağlı olarak, anod veya katoda bölgesine
göç ederler. Moleküller aynı yüke sahip olsalar dahi, molekül ağırlık farklılığı
nedeniyle sahip olunan yükün molekül ağırlığına oranı (yük/kütle) farklı olacaktır.
Bu farklılıklar sayesinde solüsyon içindeki iyonlar bir elektriksel alana tabii
tutulduklarında moleküllerin göçüne yönelik farklılıklar ortaya çıkar.
Elektroforez için gereken gereçler güç kaynağı ve elektroforez tankından
oluşur. Güç kaynağı elektrodlar arasında doğru akımın dengeli ve düzgün şekilde
akışını sağlayarak elektriksel alanın istenilen özellikte oluşmasını neden olur.
Elektriksel alanda sürekli olarak, pozitif yüklü moleküller negatif kutba, negatif
yüklü moleküller pozitif kutba doğru göç ederler.
Örnekler elektroforezin düzgün gerçekleşmesi için tampon içinde çözülürerek
hazırlanmalıdır. Ayrıca elektroforzde istenilen başarının elde edilmesi için ayırıcı
ortmın mutlaka elektroforez tamponu ile hazırlanması şarttır. pH değişikliği
elektroforezi yapılan molekülün iyonik yükünü değiştirebileceğinden, tampon
pH’sının stabil tutlması önemlidir.

1. GİRİŞ Fatma GÖZÜKARA
14
Ayırıcı ortam olarak filtre kağıdı, selüloz asetat, agaroz, nişasta, agar veya
poliakrilamid gibi sitemler kullanılabilir. Her sistemin ayırma gücü bir birinden
farklılık gösterir. Elektroforez işlemi ayırıcı ortamın konsantrasyonu, ortam
sıcaklığı, uygulanan akımın türü ve şiddeti, kullanılan tamponun iyonik gücü,
elektroforez süresi, molekül şekli ve ağırlığı gibi parametrelerden etkilenmektedir
(Aygan, 2008).
Elektrodlar arasındaki akımı genel olarak tampon iyonları az bir kısmı örnek
iyonları iletilir. Voltajdaki iletilen toplam yükün artışına neden olur. Sıcaklık artışı
elektroforez sırasında direncin düşmesine neden olur. Isı artışı tomponun
buharlamasına neden olduğunda iyonik gücün değişmesine yol açar.
Elektroforez düzeyi örneğin net elektriksel yükü arttıkça artacaktır. Diğer
taraftan molekül büyüklüğü arttıkça göç oranı düşecektir. Örnekler aynı molekül
büyüklüğüne sahip olsalar bile moleküllerin globüler yada fibröz oluşu moleüllerin
göçünde farklılığa neden olacaktır. Elektroforez işlemi, kullanılan tamponun
bileşimi ve iyonik yükünden faklı şekilde etkilenir. Yaygın olarak kullanılan
tamponlar agaroz için tris-asetat, tris-fosfat ve tris-borat, poliakrilamid için tris-
glisin karışımlarıdır.
Örnekle bağlanma davranışı göstermeyen tamponlar molekülün göç oranını
değiştirken, karbonhidratların analizinde kullanılan borik asit tamponu moleküle
bağlandığı için elektroforezde olumsuzluğa yol açar. Aynı şekilde tamponun iyonik
gücü artarsa, tampondan geçen elektrik akımıda artacaktır. (Wilson ve Goulding,
1986; Aygan, 2008).
1.6.1. Jel Elektroforezleri
Ayırıcı ortam olarak jellerin kullanılmaya başlanması, nükleik asit ve protein
gibi büyük moleküllü maddelerin separasyonunda “düşük voltajlı ince tabaka
elektroforez” sistemlerinin devre dışı kalmasına sebep olmuştur. Suda çözünmez,
hidrofilik ve yarı-katı kolloid yapıda olmaları gibi fiziksel özellikleri tercih
edilmelerinde önemli faktörleri oluşmuştur. Nişasta, agar ve poliakrilamid jel
elektroforez uygulamalarında kullanılan malzemeler olup, kullanmadan hemen

1. GİRİŞ Fatma GÖZÜKARA
15
önce hazırlanırlar.
Nişasta jeller, uygun tampon içerisinde kısmen hidrolize olmuş nişasta
karışımı ısıtılıp soğutularak hazırlanır. Nişastanın amilopektin bileşeni dallanmış
zincirlerinin sarılmaları ile yarı katı jelin oluşumunu sağlar. Nişastanın moleküler
elek özelliğinde olması, fizyolojik aktif proteinler ve kompleks yapısal molekül
karışımlarının analizinde tercih edilmelerine neden olmuştur.
Agar ucuz, toksik olmayan, zararsız bir malzeme olup agaroz ve
agaropektinden oluşmuştur. Agar tampon içerisinde kaynatılarak çözülürken ortam
sıcaklığı 40oC ye düşürüldüğünde katılaşarak jelleşir. Geniş por çapına sahip
olmaları nedeniyle elektroforez esnasında iyonların hareketi çok hızlı
gerçekleşmektedir.
Bu özellik makromoleküllerin separasyonunu için bir avantajdır. Agar,
separasyondan sonra boyama için çok uygun bir materyal olup, özellikle difüzyon
direncinin düşük olması nedeniyle, immünelektroforez uygulamalarında fazlaca
tercih edilmektedir. Saflaştırılmış agaroz jel saf halde izole edilmiş nükleik asit
yapılarının analizi için yaygın bir kullanım alanına sahiptir.
Poliakrilamid jeller oldukça toksik kimyasal bileşiklerle hazırlanır. Akrilamid
monomerleri (CH2=CHCONH2) taze hazırlanmış amonyumpersülfat ve TEMED
(Kimyasal katalizör sistemi) varlığında genellikle N,N-metilenbisakrilamidin
(CH2(NHCOCH=CH2)2) çapraz bağlanmaları ile polimerize olurlar. Moleküler
oksijenin polimerizasyonu inhibe etmesi nedeniyle monomer, tampon ve su
sistemine AMPS ve TEMED in ilavesinden önce degaz işlemi gerçekleştirilir. Bu
amaçla karışım vakum pompası yada vakumlu desikatörde veya ultrasonik su
banyosunda 5 dakika bekletilir.
Jelin dökümünden hemen sonra üst yüzeydeki yüzey gerilimi nedeniyle
meydana gelen eğimli yüzey oluşumunun separasyonu olumsuz etkilemesi ve
atmosferik oksijenin difüzyonunun engellenmesi için jelin üst tabakası distile su
veya suya doyurulmuş bütanol ile kapatılır. Jelin polimerizasyonunda riboflavin ve
TEMED sisteminden yararlanılacaksa (foto-polimerizasyon) reaksiyon ışık altında
gerçekleştirilir (Wilson ve Goulding, 1986; Aygan, 2008).

1. GİRİŞ Fatma GÖZÜKARA
16
Akrilamid jel uygulamaları kesiksiz (Continuous) ve kesikli (discontinuous)
olmak üzere iki farklı şekilde gerçekleştirilebilir. Kesiksiz sistemde tek bir ayırıcı
jel vardır ve tanklarla jelde aynı tampon kullanılır. Kesikli sistemde ise jel, farklı
tamponlarla hazırlanmış iki kısımdan oluşur. Büyük porlu stacking (dengeleme jeli)
ve küçük porlu separating (ayırıcı) jel şeklindedir.
Jelin porlarının büyüklüğü/küçüklüğü akrilamid monomerinin konsantrasyonu
ile değişir. Jeller mevcut akrilamidin toplam yüzdesi ile %3 ila %30 arasında bir
konsantrasyonda hazırlanabilir. Düşük konsantrasyonlarda geniş por yapıları oluşur
ve büyük moleküllerin separasyonu için tecih edilir.
Proteinlerin seperasyonu çoğunlukla %5-15 lik jellerde gerçekleştirilmektedir.
Benzer yüklere sahip olsalar bile farklı şekil ve büyüklükteki moleküllerin
analizinde çok uygun bir elektroforez tekniğidir.
Elektroforetik separasyon sonunda proteinler ya doğrudan jel içerisinde yada
sabit bir yüzeye (örn: nitroselüloz veya naylon membran) aktarıldıktan sonra
saptanır ve analiz edilir. Protein bandlarının görünür hale getirilmesinde en uygun
yöntem boyamadır. Jeldeki proteinler genellikle Coomassie Brillant Blue veya
Gümüş Nitrat ile, filtreye emdirilmiş proteinler ise amido black ve ponceau S gibi
boyalarla boyanır.
Boyama sonunda bir birinden ayrılmış proteinler jel yada membran üzerinde
bantlar şeklinde görünürler. Herhangi bir yöntemle boyanmış protein bantlarının
bulunduğu jel çeşitli çözeltiler içinde birkaç ay veya jel kurutma aletinde
kurutulduktan sonra yıllarca saklanabilir. Ancak görüntünün kalıcı olması açısından
en pratik yol fotoğrafının çekilmesidir (Aygan, 2008).
Proteinlerin elektroforez uygulamaları farklı amaçlara göre denatüre veya
native (Doğal) jel elektroforezi olmak üzere iki şekilde gerçekleştirilir. Denatüre
jeller SDS veya üre gibi denatüre edici ajanların jel içerisine karıştırılması ile
hazırlanırlar. Bu tür uygulama başta molekül ağırlık belirlenmesi olmak üzere,
proteinlerin saflıklarının araştırılması, konsantrasyonlarının tanımlanması ve enzim
aktivitelerinin saptanmasına olanak verir.

1. GİRİŞ Fatma GÖZÜKARA
17
1.6.2. Doğal (Native) Jel Elektroforezi
Doğal jel elektroforezinde kullanılan tamponlar, deterjan ve diğer denatüre
edici ajanları içermediğinden, işlemler doğal şartlarda gerçekleştirilir. Fazla sayıda
protein içeren karışımların bir birinden ayrılmasında, bir protein ya da protein
kompleksinin aktivite veya yapısı ile ilgili elektroforetik çalışmalarda, protein saflık
kontrolü ve proteinin denatüre olup olmadığının saptanmasında kullanılır.
Doğal jel elektroforez uygulamasında, örneklerin molekül ağırlıkları hakkında
kesin bir bilgi edinmek mümkün değildir. Bunun nedeni, molekül büyüklüğünün
yanında molekül şekli ve yükünün de ayrımı etkilemesidir (Aygan, 2008).
Bu çalışmada, glukan molekülünü oluşturan β–1,3 ve β–1,4 glikozid bağları
ile bağlanmış birimlerin yüksek sıcaklıkta tam hidrolizini gerçekleştirecek termofil
ve termostabil Likenaz enzim üreticilerinin izolasyonu, enzim üretiminin
gerçekleştirilmesi ve enzimin karakterizasyonunun yapılarak, biyoteknolojik
kullanılabilirliğinin (Arpa, yulaf ve buğdaydan hayvan yemi üretimi alanında)
araştırılması amaçlanmaktadır.

2. ÖNCEKİ ÇALIŞMALAR Fatma GÖZÜKARA
18
2. ÖNCEKİ ÇALIŞMALAR
Biodin ve Effront (1917), B. subtilis kültürlerinden enzim hazırlanmasıyla
ilgili çalışmalar yapmıştır.
Miller (1951), Karbonhidrataz etkisi gösteren enzimlerin aktivite
analizlerinde indirgen şeker miktarının saptanması için Dinitrosalisilik asit (DNS)
yöntemini kullanmıştır. Bu metod ile 3,5-dinitrosalisalisilik asitin, 3-amino-5-
nitrosalisilik asite indirgendiğini ve aldehit grupların karboksilik gruplara okside
olarak gelişen renk reaksiyonunu oluşturduklarını bildirmiştir.
Morgan ve Priest (1981), 40°C’nin üzerinde aktivitelerini kaybeden
Bacillus suşlarında termostabilite yeteneklerini araştırmışlardır.
Louw ve ark. (1993), Bacillus brevis bakterisinden endo β-(1,3)-(1,4)-
glukanaz (likenaz) genini Escherichia coli’de klonlamışlardır. Üretilen enzimin
optimum aktivite sıcaklığı 65-70°C, optimum aktivite pH’ı 8-10 aralığında
bulunmuştur. Enzim 1 saat süreyle 70°C’de inkübe edildiği zaman orijinal
aktivitesini %85 oranında korumuştur. Likenaz enziminin SDS-PAGE analizi
sonucunda moleküler ağırlığı 29 kDa olarak saptanmıştır.
Akita ve ark (2005), Bacillus halodurans C-125 suşundan izole edilen
enzimin ince tabaka kromatografisi analizinde endo özellik gösterdiğini
bulmuşlardır. Enzim optimum aktivitesini 60°C de pH 6 ve 8 de göstermiştir.
Enzimin orijinal aktivitesini 50 ve 60°C de 2 saatlik inkübasyondan sonra %100
koruduğunu saptamışlardır.
Oyekola ve ark. (2007), izole ettikleri endoglukanaz enziminin pH
optimumunun 6-6.5, sıcaklık optimumunun ise 50°C olduğunu saptamışlardır.
Enzim Cu, Ni ve Zn gibi divalent katyonlarla inhibe olurken, Fe, Mg ve Ca ile
aktive olmuştur.
Endo ve ark. (2001) alkalifilik Bacillus suşundan izole edilen alkaline
endoglucanase enziminin optimum aktivite pH’ının 10.0, sıcaklık optimumunun
ise 55°C olduğunu saptamışlardır. SDS-PAGE ile yapılan analizlerde enzimin
moleküler ağırlığının 50 kDa olduğu bulunmuştur.

2. ÖNCEKİ ÇALIŞMALAR Fatma GÖZÜKARA
19
Hirasawa ve ark. (2006), alkalifilik Bacillus agaradhaerens suşundanizole
edilen endoglucanase enziminin moleküler ağırlığının 38 kDa, optimum aktivite
gösterdiği pH aralığının 7.0-9.4 ve optimum aktivite sıcaklığının 60°C olduğunu
bulmuşlardır.
Zhang ve ark. (2006), β-1,3-1,4-glukanaz enziminin termostabilitesini
yönlendirilmiş mutasyonla artırmışlardır.
Hreggvidsson ve ark. (1996), izole ettikleri termofilik selülaz enziminn
moleküler ağırlığının 49 kDa, optimum aktivite pH’ının 7.0 olduğunu
saptamışlardır. Enzim 100°C de 3.5 saat inkübe edildiğinde orijinal aktivitesini
%50 korurken, 90°C de 16 saat inkübe edildiğinde %80 korumuştur. Enzim
yüksek düzeyde termostabil olarak tanımlanmıştır.
Robson ve Chambliss (1984), Bacillus sp.’den üretilen selülaz enziminde
aktivite analizi yapmışlar ve maksimum selülaz aktivitesin pH 4.8 ve 58°C’de
gerçekleştiğini bulmuşlardır.
Soutschek-Bauer ve Staudenbauer (1987), Clostridium thermocellum’dan
elde edilen genini B.subtilis’e ve B.stearothermophilus’a transfer ederek CMC-az
üretimi gerçekleştirmişlerdir.
Hakamada ve ark (1997), alkalifilik Bacillus sp. KSM-S237’den izole
edilmiş termostabil alkali selülazın optimum aktivitesini pH 8.6-9.0 ve 45°C’de
gösterdiği bildirmişlerdir.
Yernool ve Eveleigh (1998), iki termostabil endoselülaz izolasyonu yapmış
ve moleküler ağırlıkları 29 ve 30kDa olan enzimlerin optimum pH 6.0-6.6 ve
95°C’de aktif olduklarını bulmuştur.
Krishna (1999), katı faz fermentasyon ile B. subtilis’ten selülaz enzimi
üretimini gerçekleştirmişler ve substrat, nem, parça büyüklüğü, ortam pH’sı,
inkübasyon sıcaklığı azot ve karbonun, enzim üretimine etkilerini araştırmışlardır.
Mawadza ve ark (2000), iki Bacillus suşundan üretilen selülazın
karakterizasyonunu gerçekleştirmişlerdir. Her iki enzimin molekül ağırlığı 40 kDa
olup, enzimlerin otimum aktivitelerini pH 5.0-6.5’de ve 65 ve 70°C’de
gösterdiklerini belirlemişlerdir.

2. ÖNCEKİ ÇALIŞMALAR Fatma GÖZÜKARA
20
Hakamada ve ark (2002), Bacillus circulans’dan izole ettikleri alkalin
endoglukanazın pH 8.5’da 55°C’de optimal çalıştığını ve 43 kDa moleküler
ağırlığa sahip olduğunu saptamışlardır.
Singh ve ark (2001), termostabil alkali CMC-az enzimini Bacillus sp. VG1
suşundan izole etmişler ve enzimin optimum aktivite gösterdiği pH değerinin 9.0-
10.0, yarılanma ömrünün ise 100°C’de 12 dakika olduğunu bulmuşlardır.
Coral ve ark (2002), Aspergillus niger’den moleküler ağırlıkları 83 ve
50kDa olan, optimum aktivitesini pH 3.0-9.0 aralığında gösteren CMC-az enzimi
izole etmişlerdir. Enzimin optimum aktivite sıcaklığını 40°C bulmuşlardır.
Kang ve ark (2004), Aspergillus niger KK2 suşundan solid state
fermentasyon tekniği ile pirinç ve buğday kabukları kullanılarak selülaz enzimi
üretimi gerçekleştirmişlerdir.
Saha (2004), Mucor circinelloides’ten izole ettikleri endoglukanazın
optimum aktivitesini pH 4.0-6.0 ve 55°C’de gösterdiğini, enzimin moleküler
ağırlığının 27 kDa olduğunu belirtmiştir.
Huang ve Monk (2004), Caldibacillus cellulovorans’dan izolasyonunu
yaptıkları CMC-az enziminin 85.1 kDa molekül ağırlığa sahip olduğunu ve
maksimum aktivitenin ise 80°C’de gerçekleştiğini bildirmişlerdir.
Singh ve ark (2004), Bacillus sphaericus JS1suşundan alkali selülaz
enzimi izole ederek enzimin üretimi ve karakterizasyonunu gerçekleştirmişlerdir.
Enzimin SDS-PAGE analizinde moleküler ağırlığının 42 kDa olduğu saptanmış ve
enzimin termostabilite, pH stabilitesi ve hidrolitik kapasitesi açısından, deterjan
sanayi için uygun olduğunu belirtmişlerdir.
Kim ve ark (2005), alkalifilik Bacillus sp. suşundan HSH-810’dan alkali
selülaz izolasyonu yapmışlar ve enzimin pH 10.0’da ve 50°C’de optimum aktivite
gösterdiğini belirtmişlerdir.
Zverava ve ark (2006), haloalkalifilik anaerobik bakteriden izole ettikleri
selülaz enziminin moleküler ağırlığının 75 ve 84 kDa olduğunu bildirmişlerdir.
Hirasawa ve ark (2006), Bacillus agaradhaerens suşundan endoglukanaz
enzimi izolasyonu gerçekleştirmişler, farklı sıcaklık ve pH’larda enzim
aktivitesine bakarak, NaCl ile enzim aktivitesinde artış olduğunu belirlemişlerdir.

2. ÖNCEKİ ÇALIŞMALAR Fatma GÖZÜKARA
21
NaCl’ün enzim aktivitesini artırıcı yöndeki etkisinin pH 6.5-7.0 arasında ve
60°C’de, altı kat daha yüksek olduğunu ortaya koymuşlardır.
Kang ve ark (2007), Pyrococcus horikoshii’den, kristalin selüloza karşı
oldukça etkin hipertermofilik selülaz izolasyonu gerçekleştirmişler ve enzimin
disülfit bağına sahip olduğunu belirtmişlerdir.

3. MATERYAL VE METOD Fatma GÖZÜKARA
22
3. MATERYAL VE METOD
3.1. Materyal
3.1.1. Kullanılan Çözeltiler
3.1.1.1. NaOH Çözeltisi
Besiyerlerinin pH’sını ayarlamak ve tampon çözeltilerin hazırlanması amacı
ile 0.2N ve 2N NaOH çözeltisi kullanılmıştır.
3.1.1.2. Etanol
Sıvı besiyerlerinde üretilen enzimlerin çöktürülmesi amacı ile daha önceden
soğutulmuş %96’lık soğuk etanol kullanılmıştır.
3.1.1.3. Kongo Kırmızısı (%0.1)
Katı besiyerindeki enzim aktivitesi ve Nativ Jel uygulamasında jeldeki
likenaz aktivitesinin gösterilmesi için %0.1’lik kongo kırmızısı 100 mL distile suda
çözülerek hazırlanmıştır (Voget ve ark, 2006).
3.1.1.4. NaCl Çözeltisi (1M)
Petri kutusunda likenaz aktivitesini saptamak ve zimogram analizinde jelde
bulunan aktivite bandını görünür hale getirmek amacıyla kullanılmıştır (Voget ve
ark, 2006).
3. MATERYAL VE METOD Fatma GÖZÜKARA
23
3.1.1.5. Sodyum Fosfat Tamponu (0.1 M)
Çöktürme sonucu elde edilen enzimin çözülerek resüspanse edilmesi için
kullanılmıştır.
3.1.1.6. Sitrat Tamponu
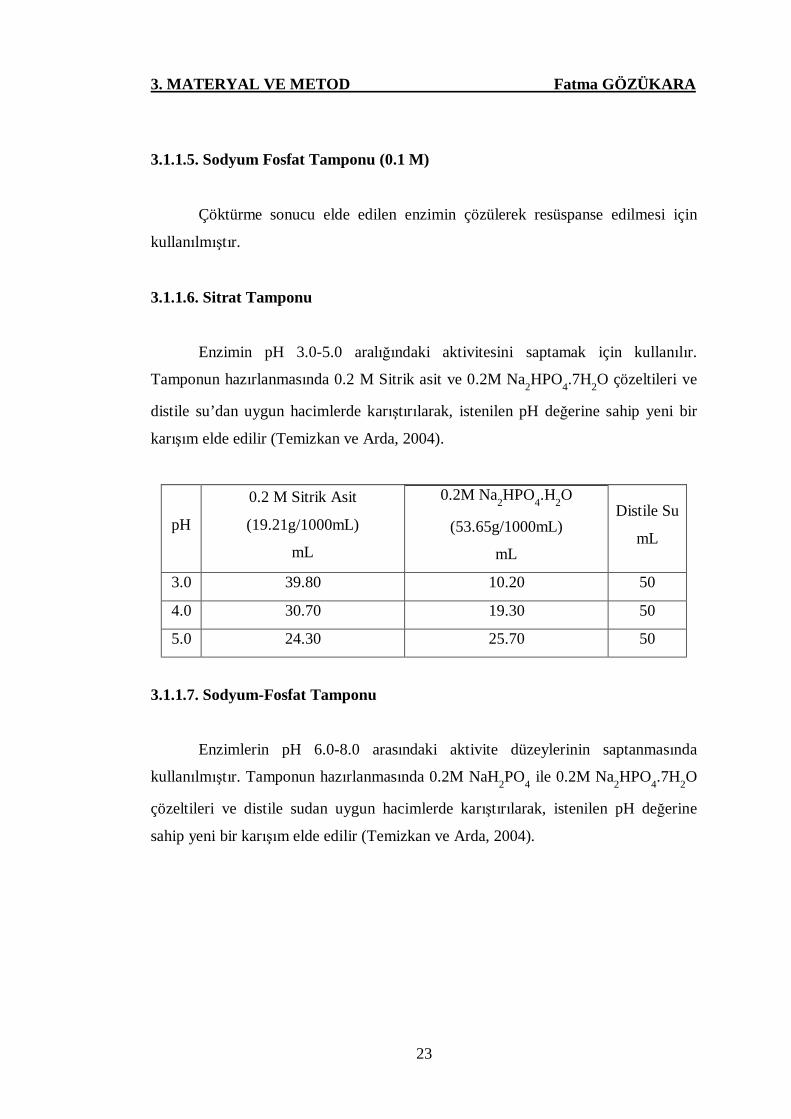
Enzimin pH 3.0-5.0 aralığındaki aktivitesini saptamak için kullanılır.
Tamponun hazırlanmasında 0.2 M Sitrik asit ve 0.2M Na2HPO4.7H2O çözeltileri ve
distile su’dan uygun hacimlerde karıştırılarak, istenilen pH değerine sahip yeni bir
karışım elde edilir (Temizkan ve Arda, 2004).
pH
0.2 M Sitrik Asit
(19.21g/1000mL)
mL
0.2M Na2HPO4.H2O
(53.65g/1000mL)
mL
Distile Su
mL
3.0 39.80 10.20 50
4.0 30.70 19.30 50
5.0 24.30 25.70 50
3.1.1.7. Sodyum-Fosfat Tamponu
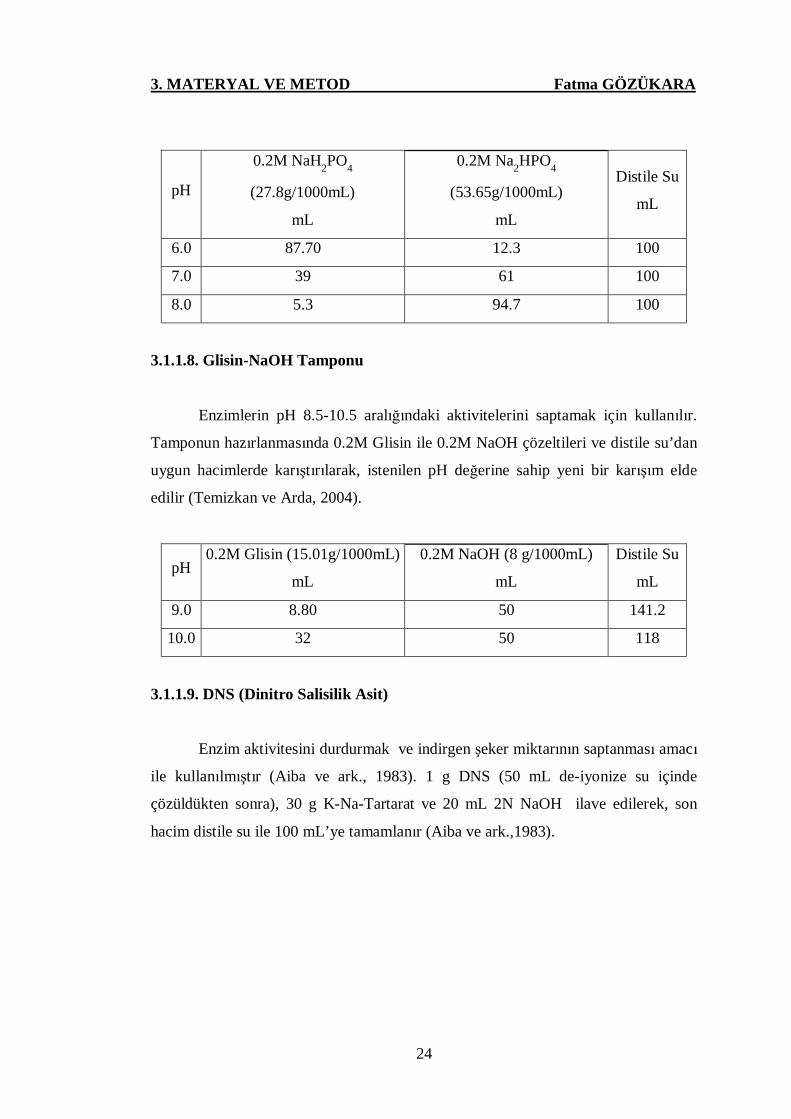
Enzimlerin pH 6.0-8.0 arasındaki aktivite düzeylerinin saptanmasında
kullanılmıştır. Tamponun hazırlanmasında 0.2M NaH2PO4 ile 0.2M Na2HPO4.7H2O
çözeltileri ve distile sudan uygun hacimlerde karıştırılarak, istenilen pH değerine
sahip yeni bir karışım elde edilir (Temizkan ve Arda, 2004).
3. MATERYAL VE METOD Fatma GÖZÜKARA
24
pH
0.2M NaH2PO4
(27.8g/1000mL)
mL
0.2M Na2HPO4
(53.65g/1000mL)
mL
Distile Su
mL
6.0 87.70 12.3 100
7.0 39 61 100
8.0 5.3 94.7 100
3.1.1.8. Glisin-NaOH Tamponu
Enzimlerin pH 8.5-10.5 aralığındaki aktivitelerini saptamak için kullanılır.
Tamponun hazırlanmasında 0.2M Glisin ile 0.2M NaOH çözeltileri ve distile su’dan
uygun hacimlerde karıştırılarak, istenilen pH değerine sahip yeni bir karışım elde
edilir (Temizkan ve Arda, 2004).
pH 0.2M Glisin (15.01g/1000mL)
mL
0.2M NaOH (8 g/1000mL)
mL
Distile Su
mL
9.0 8.80 50 141.2
10.0 32 50 118
3.1.1.9. DNS (Dinitro Salisilik Asit)
Enzim aktivitesini durdurmak ve indirgen şeker miktarının saptanması amacı
ile kullanılmıştır (Aiba ve ark., 1983). 1 g DNS (50 mL de-iyonize su içinde
çözüldükten sonra), 30 g K-Na-Tartarat ve 20 mL 2N NaOH ilave edilerek, son
hacim distile su ile 100 mL’ye tamamlanır (Aiba ve ark.,1983).

3. MATERYAL VE METOD Fatma GÖZÜKARA
25
3.1.2. Bakteri İzolasyonunda ve Teşhisinde Kullanılan Besiyerleri
3.1.2.1. LB Agar
Bakterilerin izolasyonu ve stok kültürlerin saklanması için kullanılmıştır
(Sambrook ve Russell, 2001).
Bileşimi g/L
Tripton 10
Maya Özütü 5
NaCl 10
Agar 15
3.1.2.2. Likenanlı Besiyeri
Likenaz aktivitesinin saptanması amacıyla kullanılmıştır (Kim ve ark, 2005).
Bileşimi g/L
Likenan 10
Pepton 5
Maya Özütü 5
KH2PO4 1
MgSO4.7H2O 0.2
NaCl 10
Agar 15

3. MATERYAL VE METOD Fatma GÖZÜKARA
26
3.1.2.3. Likenaz Enzimi Üretimi Besiyeri (Krishna,1999).
Bileşimi g/L
Na2HPO4.7H2O 1.18
KH2PO4 0.9
NaNO3 1
KCl 0.5
MgSO4.7H2O 0.5
Maya Özütü 0.5
Pepton 0.5
Likenan 10
3.1.3. Elektroforez İşleminde Kullanılan Çözeltiler
3.1.3.1. Solüsyon A (Akrilamid Solüsyonu)
Bileşimi g
Akrilamid 29.2
Bisakrilamid 0.8
Distile su 100 mL
Yukardaki bileşikler bir miktar distile su içerisinde çözüldükten sonra son
hacim distile su ile 100 mL’ye tamamlanır. (Bollag ve ark, 1996).
3.1.3.2. Solüsyon B (4X)
Nativ jel uygulamalarında SDS kullanılmadığı için 75 mL 2M Tris-HCl (pH
8.8) ve son hacim distile su ile 100 mL’ye tamamlanır. (Bollag ve ark, 1996).

3. MATERYAL VE METOD Fatma GÖZÜKARA
27
3.1.3.3. Solüsyon C (4X)
Nativ jel uygulamalarında SDS kullanılmadığı için 50 mL 1M Tris-HCl (pH
6.8) ve son hacim distile su ile 100 mL’ye tamamlanır. (Bollag ve ark, 1996).
3.1.3.4. Amonyum Persülfat (AMPS) %10
0.5g AMPS 5mL distile su içerisinde çözülerek hazırlanır (Bollag ve ark,
1996).
3.1.3.5. Elektroforez Tamponu
Bileşimi g/L
Tris 3
Glisin 14.4
Distile su 1000mL
Nativ Jel uygulandığından SDS kullanılmamıştır (Bollag ve ark, 1996).
3.1.3.6. Örnek Yükleme Tamponu (5X)
Bileşimi mL
1 M Tris-HCl (pH 6.8) 0.6
Gliserol %50 5
β-Merkaptoetanol 0.5
Bromfenol mavisi %1 1
Distile su 0.9
Nativ Jel uygulandığından SDS kullanılmamıştır (Bollag ve ark, 1996).

3. MATERYAL VE METOD Fatma GÖZÜKARA
28
3.1.3.7. Jel Boyama (Staining) Solüsyonu
Bileşimi
Comassie Brillant Blue R-250 1 g
Metanol 450 mL
Glasial Asetik Asit 100 mL
Distile su 450 mL
1g Coomassie Brillant Blue R-250, 450 mL metanol içerisinde çözülüp filtre
kağıdından süzüldükten sonra, karışımın üzerine 100 mL Glasial asetik asit ve 450
mL distile su eklenerek hazırlanır (Bollag ve ark, 1996).
3.1.3.8. Jelden Boyayı Geri Alma (Destaining) Solüsyonu (Bollag ve ark, 1996).
Bileşimi mL
Metanol 100
Glasial Asetik Asit 100
Distile su 800

3. MATERYAL VE METOD Fatma GÖZÜKARA
29
3.2. Metod
3.2.1. Bakteri İzolasyonu ve Teşhisi
3.2.1.1. Bacillus sp. Suşlarının İzolasyonu
Değişik ortamlardan örnekler alınarak 1’er g tartılıp 5 mL steril distile su
içerisinde süspansiyon haline getirilmiştir. Bacillus sp. sporlu bir bakteri olduğundan,
elde edilen süspansiyon 80ºC’de 10 dakika inkübe edilerek ısı şoku uygulanmıştır
(Lennete ve ark., 1985).
Isı şoku uygulamasından sonra tek koloni oluşumunu sağlayabilmek için
örnekten seri sulandırmalar yapılarak, pH sı 7.0 olan LB agar besiyerine yayma
şeklinde ekim yapılmış ve 55ºC’de 1 gece inkübe edilmiştir. İnkübasyon sonunda
besiyerinde tek düşmüş farklı morfolojik görünüme sahip koloniler seçilmiş ve LB
agar besiyerine çizgi şeklinde ekim yapılarak identifikasyon ve Likenaz aktivitesinin
saptanması amacı ile stok kültür şeklinde saklanmışlardır.
3.2.2. Bakterilerin İdentifikasyonu
Seçilen bakteri örneklerini tanımlamak amacı ile indol, hidrojensülfür,
hareket, asit, katalaz, jelatin ve kazeini kullanma gibi biyokimyasal testler ile spor,
yüzeyde zar oluşturma, dipte çökelti oluşturma ve gram boyama testleri yapılmıştır
(Jin ve ark., 1990).
3.2.3. Katı Besiyerinde Likenaz Aktivitesinin Saptanması
Tanımlanmaları yapılan stok Bacillus sp. suşları Likenan içeren pH sı 7.0’ye
ayarlanmış LB agar besiyerine çizgi ekim tekniği kullanılarak aşılanmışlar ve
55ºC’de 24 saat üremeye bırakılmışlardır. İnkübasyon sonunda koloniler %0.1’lik
kongo kırmızısı ile 15 dakika boyandıktan sonra 1M’lık NaCl ile 15 dakika boyanın
geri alınması işlemi uygulanmıştır. Kolonilerin etrafında kırmızı zeminde sarı zon

3. MATERYAL VE METOD Fatma GÖZÜKARA
30
oluşumu likenaz pozitif şeklinde değerlendirilmiştir (Hols ve ark., 1994; Burhan ve
ark., 2003).
3.2.4.Bakterinin Ürediği ve Enzim Sentezinin Gerçekleştiği pH Aralığının
Saptanması
Bu amaçla 5.0, 6.0, 7.0, 8.0, 9.0, 10.0 pH değerlerine sahip Likenan’lı LB
agar besiyerlerine, tanımlanması yapılan ve likenaz pozitif özellik gösteren suşlardan
çizgi şeklinde ekim yapılmıştır. Örnekler bakteri izolasyonunun gerçekleştirildiği
sıcaklık olan 55ºC’de 24 saat üretildikten sonra en iyi üreme ve enzim sentezinin
gerçekleştiği pH değeri ve aralığı koloni çapı ve aktivite zon genişliği ölçülerek (mm
olarak) saptanmıştır (Hols ve ark.,1994; Burhan ve ark. 2003).
3.2.5. Bakterinin Ürediği ve Enzim Sentezinin Gerçekleştiği Sıcaklık ve pH
Aralığının Saptanması
Tanımlamaları yapılan ve ilk analizlerde iyi aktivite gösteren 20 suş 20, 30,
40, 50 ve 60 ºC lerde pH değerleri 5.0, 6.0, 7.0, 8.0, 9.0 ve 10.0 a ayarlanmış
likenanlı LB agar besiyerinde üretilerek üreme ve enzim sentezinin sıcaklık ve pH
değerlerine göre değişimi araştırılmıştır. Örnekler 24 saat süreyle belirtilen
sıcaklıklarda inkübe edildikten sonra gereçekleşen koloni çapları ve aktivite zon
genişlikleri ölçülerek (mm olarak) en iyi üreme ve enzim sentezinin gerçekleştiği
sıcaklık değeri ve aralığı saptanmıştır (Arikan, 2008).
3.2.6. Enzim Üretimi ve Kısmi Saflaştırma
Seçilen bakteri örneklerinden en geniş zon çapı oluşturan L-12 suşu enzim
üretimi için kullanılmıştır. Bu amaçla seçilen suşun bir gecelik taze kültüründen
içerisinde likenan bulunan ve pH sı 7.0 olan sıvı LB besiyerine uygun şekilde
aşılama yapılarak 24 saat süreyle 55ºC’de 250 devir/dakikada çalkalama kültür
şeklinde üretim gerçekleştirilmiştir.

3. MATERYAL VE METOD Fatma GÖZÜKARA
31
Elde edilen karışık kültür +4ºC ve 6000 rpm’de 20 dakika santrifüj edilerek
bakteriler kültürden uzaklaştırılmış ve sıvı faz temiz bir tüpe aktarılmıştır. Örneğin
üzerine orijinal hacmin %70’i oranında %96’lık soğuk etanol eklenmiş ve –33ºC’de
bir gece bekletilerek alkol presipitasyonu yapılmıştır.
Soğuk çöktürme ortamından alınan örnek +4ºC de 20 dakika süreyle 8000
rpm’de santrifüj edilerek çöktürülmüştür. Dipte toplanan enzim çökeltisi 0.1 M’lik
pH’sı 6.8 olan sodyum fosfat tamponunda çözülerek, toplam 250 mL’lik enzim
solüsyonu elde edilmiştir. Elde edilen enzimden likenan bulunan katı besiyerine 50
µL damlatılarak aktivite kontrolü yapılmıştır (Srivastava, 1987; Arikan, 2008).
3.2.7. Enzimin pH Optimumunun Saptanması
Alkol çöktürme tekniği kullanılarak kısmi saflaştırma işlemi ile elde edilen
enzim çözeltisinin optimum aktivite gösterdiği pH değerinin saptanması için Sitrat-
fosfat (pH 3.0-5.0), Na-fosfat (pH 6.0-8.0) ve Glisin-NaOH (pH 8.5-10.5) tamponları
kullanılarak %0.1’lik likenan çözeltileri hazırlanmıştır (Anonymous, 1917).
Aktivite tayini için 0.5 mL enzim çözeltisi ve her pH değerindeki substrat
çözeltisinden 0.5 mL örnek alınarak tüpe konulmuştur. Her pH değeri için üç seri tüp
analiz edilmiştir. Hazırlanan enzim substrat karışımları enzimin üretildiği sıcaklık
olan 55ºC’lik su banyosunda 30 dakika süreyle inkübe edilmiştir.
İnkübasyonun sonunda her bir örnek tüpüne eşit miktarda DNS ayıracı
konularak (1 mL enzim+substrat, 1 mL DNS) 5 dakika süreyle kaynatma işlemi
uygulanmıştır. Tüpler soğutulduktan sonra 540 nm dalga boyu kullanılarak Cecil5500
UV-visible spektrofotometresinde köre karşı (eşit hacimde substrat ve DNS ayıracı
kullanılarak hazırlanmıştır) okuma işlemi yapılmıştır (Arikan, 2008).
En yüksek absorbans değerinin elde edildiği pH değeri 100 kabul edilerek
diğer pH değerleri buna göre oranlanarak relatif enzim aktivitesi saptanmıştır
(Gessesse ve Gashe., 1997; Burhan ve ark., 2003).

3. MATERYAL VE METOD Fatma GÖZÜKARA
32
3.2.8. Enzimin Optimum Sıcaklık Aktivitesinin Saptanması
Kısmi saflaştırma ile elde edilen enzimin optimum aktivite gösterdiği sıcaklık
değerinin saptanması için 30, 40, 50, 60, 70, 80, 90, 100 ve 110ºC’lik sıcaklık
değerleri seçilmiştir. İnkübasyonlardan 30-90ºC aralığındaki denemeler su
banyosunda, 100-110 ºC aralığındaki denemeler ise yağ banyosunda yapılmıştır.
Aktivite tayini için 0.5 mL enzim ve 0.5 mL substrat çözeltisi (optimum
aktivitenin gerçekleştiği pH değerinde hazırlanmış) karıştırılarak seçilen sıcaklık
değerlerinde 30 dakika süreyle inkübasyon gerçekleştirilmiştir.
İnkübasyonun sonunda her bir örnek tüpüne eşit miktarda DNS ayıracı
konularak (1 mL enzim+substrat, 1 mL DNS) 5 dakika süreyle kaynatma işlemi
uygulanmıştır. Tüpler soğutulduktan sonra 540 nm dalga boyu kullanılarak Cecil5500
UV-visible spektrofotometresinde köre karşı (eşit hacimde substrat ve DNS ayıracı
kullanılarak hazırlanmıştır) okuma işlemi yapılmıştır (Arikan, 2008).
En yüksek absorbans değerinin elde edildiği sıcaklık değeri 100 kabul
edilerek diğer sıcaklık değerleri buna göre oranlanarak relatif enzim aktivitesi
saptanmıştır (Gessesse ve Gashe., 1997; Burhan ve ark., 2003).
3.2.9. Enzimin Sıcaklık Stabilitesinin Saptanması
Likenaz enziminin sıcaklık stabilitesinin (termal stabilite) saptanması için 30-
110ºC arasındaki sıcaklık değerlerinde 15 dakikalık ön inkübasyon
gerçekleştirilmiştir (sadece enzim kullanılarak). Ön inkübasyon işleminden sonra 0.5
mL enzim ve 0.5 mL substrat solusyonu karıştırılarak optimum aktivitenin
gerçekleştiği sıcaklık değerinde 30 dakika süreyle inkübasyon gerçekleştirilip,
standart aktivite tayini yapılmıştır (Aiba ve ark., 1983; Srivastava, 1987; Gessesse ve
Gashe, 1997; Mehrotta ve ark., 1999; Burhan ve ark. 2003; Arikan, 2008).
İnkübasyonun sonunda her bir örnek tüpüne eşit miktarda DNS ayıracı
konularak (1 mL enzim+substrat, 1 mL DNS) 5 dakika süreyle kaynatma işlemi
uygulanmıştır. Tüpler soğutulduktan sonra 540 nm dalga boyu kullanılarak Cecil5500
UV-visible spektrofotometresinde köre karşı (eşit hacimde substrat ve DNS ayıracı

3. MATERYAL VE METOD Fatma GÖZÜKARA
33
kullanılarak hazırlanmıştır) okuma işlemi yapılmıştır (Arikan, 2008).
3.2.10. Likenaz Enziminin pH Stabilitesinin Saptanması
Likenaz enziminin pH stabilitesinin saptanması için pH 3.0, 4.0, 5.0, 6.0, 7.0,
8.0, 9.0 ve 10.0 değerine sahip çözeltiler kullanılarak enzim 24 saat süreyle 37ºC ön
inkübasyon işlemine tutulmuştur. Kısmi saflaştırma ile elde edilmiş sıvı enzim
örneğinden (bütün aktiviteler için kullanılan stok enzim) her pH değeri için üçer
örnek olacak şekilde her tüpe 2 mL enzim konularak 10.000 rpm de 5 dakika
çöktürülmüştür. Üst faz atıldıktan sonra tüplere (üçerli) önceden hazırlanmış farklı
pH değerindeki tamponlardan eklenmiş ve enzim sıvılaştırılarak 37ºC de 24 saat ön
inkübasyona bırakılmıştır (Arikan, 2008).
Aktivite analizi için ön inkübasyonu yapılmış enzim ve optimum pH daki
tamponda hazırlanmış substrat çözeltisinden 0.5’er mL alınarak optimum sıcaklığın
gerçekleştiği sıcaklık değerinde 30 dakika inkübasyon gerçekleştirilmiş ve standart
aktivite tayini yapılmıştır.
3.2.11. NaCl’ün Enzim Aktiviteleri Üzerine Etkisinin Belirlenmesi
Sıvı enzim örneğinden 2 mL alınarak eppendorf tüpünde çöktürülmüş, sıvı
faz atılarak tüpe farklı konsantrasyonda NaCl (%3.5, %5.0, %7.5, %10.0, %15.0 ve
%20.0) çözeltilerinden 2 mL eklenmiş ve enzim tekrar resüspanse edilmiştir. Her
konsantrasyon için üç örnek hazırlanmıştır. Sıvılaştırılan enzim 37°C de 15 dakika
inkübasyona bırakılmıştır. Ön inkübasyon aşamasından sonra, 0.5 mL enzim ve 0.5
mL substrat (optimum aktivitenin görüldüğü pH değerinde hazırlanmış) karıştırılarak
optimum aktivitenin gerçekleştiği sıcaklıkta 30 dakika inkübasyon gerçekleştirilip
standart aktivite tayini yapılmıştır (Aygan, 2008; Arikan, 2008).

3. MATERYAL VE METOD Fatma GÖZÜKARA
34
3.2.12. Enzim Aktivitesine İnhibitör, Şelatör, Deterjan ve Metal İyonlarının
Etkisi
Stok enzim örneğinden her bir inhibitör için üç seri analiz yapılacak şekilde
0.5’er mL alınarak 10000 rpm’de çöktürülmüştür. Sıvı faz dökülerek ependorf
tüpüne (her tüp için ayrı ayrı olacak şekilde) 0.5 mL uygun konsantrasyonda
inhibitör madde ilave edilerek enzim süspansiyon haline getirilmiş ve 37ºC de 15
dakika süreyle inkübasyon yapılmıştır. İnhibitör madde olarak 5mM EDTA, 5mM
CaCl2, 3mM PMSF, %1v/v SDS, %1v/v Triton X-100, 5mM Na-Sülfide, 5mM
ZnCl2 ve 8M üre kullanılmıştır.
Ön inkübasyon işleminden sonra enzim örneğinin üzerine 0.5 mL optimum
pH değerinde hazırlanmış substrat karıştırılarak optimum aktivitenin elde edildiği
sıcaklıkta 30 dakika süreyle inkübasyon gerçekleştirilmiştir. İnkübasyon sonunda
standart aktivite tayini yapılmıştır. Başlangıç enzim aktivitesini saptamak için enzim
ve substrat karışımı optimum aktivitenin gerçekleştiği sıcaklıkta 30 dakika inkübe
edilerek başlangıç enzim aktivitesi saptanmıştır. Başlangıç testinden elde edilen
değer 100 olarak değerlendirilip inhibitör, şelatör, deterjan ve metal iyonları ile
yapılan analiz sonunda elde edilen değerler % olarak oranlanıp, relatif olarak kalan
enzim aktivitesi saptanmıştır. (Burhan ve ark., 2003).
3.2.13. Nativ (Doğal) Jel Elektroforezinde Moleküler Ağırlık ve Zimogram
Analizi
Enzimlerin moleküler ağırlıkları ve enzim fraksiyonuna ait aktivitenin
(zimogram) analizi için Nativ jel sistemlerinden yararlanılmıştır (Laemmli, 1970).
3.2.13.1. Nativ Jel Sisteminin Hazırlanması
3.2.13.2. Ayırıcı Jel’in Hazırlanması (%10’luk)
Doğal (nativ) jel elektroforezi için kullanılacak jel bileşenleri (Sol B, Sol C,

3. MATERYAL VE METOD Fatma GÖZÜKARA
35
Elektroforez tamponu, ve örnek yükleme tamponu) SDS’siz olarak hazırlanır.
%10’luk homojen native jel hazırlamak için 6.5 mL Sol A, 5 mL Sol B ve 8.5
mL distile su 50 mL’lik cam bir şişe içerisine ilave edilerek moleküler oksijenin
uzaklaştırılması amacı ile vakum pompasıyla 5 dakika degaz işlemi yapılmıştır.
Karışıma 66 µL %10’luk AMPS çözeltisinden, 13 µL TEMED (N,N,N,N -
tetrametilen-etilendiamin) çözeltisinden ilave edilmiş ve şişe karıştırılmıştır.
Hazırlanan solüsyon bir enjektör yardımı ile cam plaklar arasına dökülmüştür. Jel
yüzeyinin düzgün olması ve atmosferik oksijenin jele difüzyonunu engellemek amacı
ile üst yüzey ince bir su tabakası ile kapatılmıştır. Jelin polimerize olabilmesi için,
oda sıcaklığında 30-60 dakika bekletilmiştir (Bollag ve ark.,1996; Coral ve ark.,
2002; Burhan ve ark., 2003).
3.2.13.3. Dengeleme Jel’inin Hazırlanması
1 mL Sol A, 1.5 mL Sol C ve 3.5 mL distile su 20 mL’lik cam bir şişe
içerisine konmuş ve vakum pompası ile 5 dakika degaz işlemi yapılmıştır. Karışımın
üzerine 20 µL AMPS ve 5 µL TEMED çözeltisinden ilave edilerek şişe birkaç kez
karıştırılmıştır. Hazırlanan karışım, ayırıcı jelin üzerine döküldükten sonra örnek
gözlerinin oluşturulması için üst taraftan tarak yerleştirilmiştir. Atmosferik oksijen
difüzyonunu önlemek amacıyla üst yüzey su ile kaplanmış ve oda sıcaklığında 30-60
dakika süreyle polimerizasyon gerçekleştirilmiştir. Polimerizasyon işlemi
tamamlandıktan sonra tarak jelden çıkarılmış ve oluşan gözlere örneklerin daha iyi
yerleşmesi ve gözlerin bozulmaması için 100 µL elektroforez tamponu
doldurulmuştur (Bollag ve ark.,1996; Coral ve ark., 2002; Burhan ve ark., 2003).
3.2.13.4. Enzim Örneklerinin Jele Yüklenmesi ve Yürütülmesi
Polimerize olmuş jeldeki tarak, oluşan örnek yükleme gözleri yırtılmadan
çıkarılır ve örnek yükleme tamponu ile karıştırılmış protein örnekleri ceplere 30 µL
miktarda doldurulmuştur. Elektroforez işlemi sonunda moleküler ağırlıkları doğru
saptamak için gözlerden birine içeriği ve moleküler ağırlığı bilinen protein standart’ı

3. MATERYAL VE METOD Fatma GÖZÜKARA
36
(68.000 Da ağırlığa sahip sığır serum albumininden 5 µL konmuştur) konur. Örnek
yükleme işlemi tamamlandıktan sonra elektroforez uygulaması başlatılır. Örnekte
bulunan izleme boyası ayırma jeli içine girinceye kadar yaklaşık 30 mA, daha sonra
20 mA akım uygulanır. İzleme boyası jelin diğer ucuna 2 cm uzaklığa ulaştığı zaman
elektroforez işlemi sonlandırılır.
3.2.13.5. Nativ (Doğal) Jelinin Boyanması ve Zimogram Analizleri
Elektroforez işlemi tamamlandıktan sonra marker proteinin yürütüldüğü
bölüm jelde kesilerek çıkartılır ve boya çözeltisine konularak 12 saat boyama
gerçekleştirilmiştir. Boyama işleminden sonra marker proteinin bulunduğu jel
destaining solüsyonuna konularak boyanın geri alınması işlemi yapılmıştır.
Analiz edilecek örneğin bulunduğu diğer jel bölümü içerisinde %3 likenan
bulunan pH’sı 6.0 olan (optimum enzim aktivitesinin gözlendiği pH değeri) 0.2 M
Na-fosfat tamponu içerisine alınmıştır. Jel 55 ºC ye konularak 50 dev/dk. da
çalkalamalı karıştırıcı kullanılarak 30 dakika süreyle inkübasyon gerçekleştirilmiştir.
İnkübasyon işleminden sonra jel %0.1’lik Kongo Kırmızısı (w/v) içerisinde
15 dakika süreyle boyanmıştır. Aktivite zonlarının görülebilmesi için fazla boyanın
giderilmesi amacıyla jel 1M NaCl çözeltisinde 15 dakika inkübe edilmiştir. Kırmızı
zeminde sarı zon bölgeleri enzimin bulunduğu bölgeler şeklinde değerlendirilmiştir.
Enzim bandlarının moleküler ağırlığını saptamak amacıyla marker proteinin
bulunduğu jel bölgesi ile enzimin bulunduğu jel bölgesi birleştirilerek
görüntülenmiştir (Aygan, 2008).

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
37
4. BULGULAR VE TARTIŞMA
4.1. Bacillus sp. Suşlarının İzolasyonu
Değişik ortamlardan örnekler alınarak 1’er gr tartılıp 5 ml steril distile su ile
karıştırıldıktan sonra süspansiyon haline getirilmiştir. Bacillus sp. sporlu bir bakteri
olduğundan, elde edilen süspansiyon 80ºC’de 10 dakika inkübe edilerek ısı şoku
uygulanmıştır (Lennete ve ark., 1985).
Isı şoku uygulamasından sonra tek koloni oluşumunu sağlayabilmek için
örnekten seri sulandırmalar yapılarak, LB agar besiyerine yayma şeklinde ekim
yapılmış ve 55ºC’lik etüve konularak 1 gece süreyle inkübasyon gerçekleştirilmiştir.
İnkübasyon sonunda besiyerinde tek düşmüş farklı morfolojik görünüme sahip 20
koloni seçilmiş ve LB agar besiyerine çizgi şeklinde ekim yapılarak identifikasyon
ve Likenaz aktivitesinin saptanması amacı ile stok kültür şeklinde saklanmışlardır.
Enzim üretimi ve karakterizasyonu için kullanılmak üzere seçilen suşlar,
geniş ve dalgalı kenarlı koloni morfolojisine sahiptirler. Suşlar gram pozitif, aerob,
spor oluşturan, çubuk şekilli ve hareketli bakterilerdir.
İzolasyonu ve biyokimyasal testlere göre tanımlanmaları tamamlanan 20
Bacillus sp. suşu farklı sıcaklık ve pH değerlerindeki üreme ve enzim sentezleme
yeteneklerinin saptanması için likenan içeren katı besiyerlerine çizgi yöntemine göre
ekilerek analiz edilmişlerdir. Elde edilen sonuçlar Çizelge 4.1. , 4.2. , 4.3. , 4.4. ve
4.5. de gösterilmiştir.
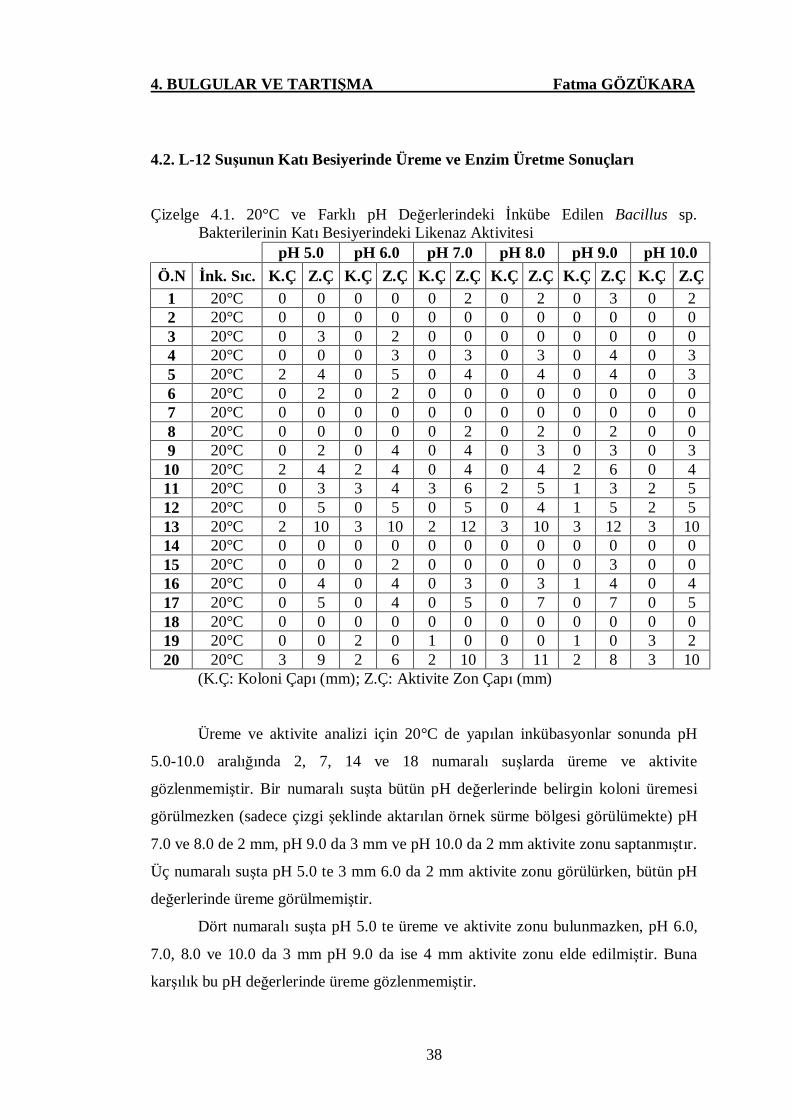
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
38
4.2. L-12 Suşunun Katı Besiyerinde Üreme ve Enzim Üretme Sonuçları
Çizelge 4.1. 20°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp. Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi
pH 5.0 pH 6.0 pH 7.0 pH 8.0 pH 9.0 pH 10.0 Ö.N İnk. Sıc. K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç
1 20°C 0 0 0 0 0 2 0 2 0 3 0 2 2 20°C 0 0 0 0 0 0 0 0 0 0 0 0 3 20°C 0 3 0 2 0 0 0 0 0 0 0 0 4 20°C 0 0 0 3 0 3 0 3 0 4 0 3 5 20°C 2 4 0 5 0 4 0 4 0 4 0 3 6 20°C 0 2 0 2 0 0 0 0 0 0 0 0 7 20°C 0 0 0 0 0 0 0 0 0 0 0 0 8 20°C 0 0 0 0 0 2 0 2 0 2 0 0 9 20°C 0 2 0 4 0 4 0 3 0 3 0 3
10 20°C 2 4 2 4 0 4 0 4 2 6 0 4 11 20°C 0 3 3 4 3 6 2 5 1 3 2 5 12 20°C 0 5 0 5 0 5 0 4 1 5 2 5 13 20°C 2 10 3 10 2 12 3 10 3 12 3 10 14 20°C 0 0 0 0 0 0 0 0 0 0 0 0 15 20°C 0 0 0 2 0 0 0 0 0 3 0 0 16 20°C 0 4 0 4 0 3 0 3 1 4 0 4 17 20°C 0 5 0 4 0 5 0 7 0 7 0 5 18 20°C 0 0 0 0 0 0 0 0 0 0 0 0 19 20°C 0 0 2 0 1 0 0 0 1 0 3 2 20 20°C 3 9 2 6 2 10 3 11 2 8 3 10
(K.Ç: Koloni Çapı (mm); Z.Ç: Aktivite Zon Çapı (mm)
Üreme ve aktivite analizi için 20°C de yapılan inkübasyonlar sonunda pH
5.0-10.0 aralığında 2, 7, 14 ve 18 numaralı suşlarda üreme ve aktivite
gözlenmemiştir. Bir numaralı suşta bütün pH değerlerinde belirgin koloni üremesi
görülmezken (sadece çizgi şeklinde aktarılan örnek sürme bölgesi görülümekte) pH
7.0 ve 8.0 de 2 mm, pH 9.0 da 3 mm ve pH 10.0 da 2 mm aktivite zonu saptanmıştır.
Üç numaralı suşta pH 5.0 te 3 mm 6.0 da 2 mm aktivite zonu görülürken, bütün pH
değerlerinde üreme görülmemiştir.
Dört numaralı suşta pH 5.0 te üreme ve aktivite zonu bulunmazken, pH 6.0,
7.0, 8.0 ve 10.0 da 3 mm pH 9.0 da ise 4 mm aktivite zonu elde edilmiştir. Buna
karşılık bu pH değerlerinde üreme gözlenmemiştir.

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
39
Beş numaralı suş pH 5.0 de 2 mm koloni çapı oluşturarak üremiş fakat diğer
pH değerlerinde üreme gerçekleşmemiştir. Bununla birlikte bütün pH değerlerinde
aktivite zonu (pH 5.0 te 4 mm, 6.0 da 5 mm, 7.0, 8.0 ve 9.0 da 4 mm ve 10.0 da 3
mm) ölçülmüştür. Altı numaralı suşta pH 5.0 ve 6.0 da üreme gözlenmezken 2 mm
aktivite zonu elde edilmiştir. Diğer pH değerlerinde ise üreme ve aktivite zonu
saptanmamıştır.
Sekiz numaralı suşta bütün pH değerlerinde belirgin üreme saptanmamıştır.
Bununla birlikte pH 7.0-9.0 da 2 mm aktivite zonu saptanmıştır. Dokuz numaralı suş
bütün pH değerlerinde üreme göstermezken pH 5.0 de 2, 6.0 ve 7.0 de 4, 8.0, 9.0 ve
10.0 da 3 mm aktivite zonu ölçülmüştür. On numaralı suşta pH 5.0 ve 6.0 da 2 mm
bakteri 4 mm aktivite zonu gözlenmiştir. pH 7.0, 8.0 ve 10.0 da üreme gözlenmediği
halde 4 er mm aktivite zonu saptanmıştır. pH 9.0 da ise 2 mm bakteri, 6 mm aktivite
zonu elde edilmiştir. On bir numaralı suşta pH 5.0 te üreme görülmeden 3 mm çaplı
aktivite zonu oluşmuştur. Diğer bütün pH değerlerinde üreme ve aktivite zonu
meydana gelirken en yüksek aktivite 6 mm ile pH 6.0 da saptanmıştır. On iki
numaralı suşta ortalama 5 mm aktivite zonu saptanırken, pH 5.0, 6.0, 7.0 ve 8.0 de
üreme görülmezken diğer pH değerlerinde 1-2 mm çaplı üreme gerçekleşmiştir.
On üç numaralı suş bu sıcaklık değerinde en yüksek aktivite zonlarının elde
edildiği örnek olmuştur. Ortalama koloni çapı 2 mm olarak saptanırken, ortalama
aktivite zon çapı 10.6 mm en geniş zon ise pH 7.0 ve 9.0 da 12 mm olarak
gözlenmiştir. On beş numaralı suşta bütün pH değerlerinde belirgin üreme
görülmezken pH 6.0 da 2, 9.0 da 3 mm aktivite zonu saptanmıştır.
On altı numaralı suşta pH 9.0 da 1 mm koloni çapı oluşumu söz konusu iken
diğer pH larda üreme olmamıştır. Bununla birlikte bütün pH değerlerinde ortalama 4
mm aktivite zonu saptanmıştır. On yedi numaralı suşta hiçbir pH değerinde üreme
gözlenmezken, pH 8.0 ve 9.0 da 7 mm, diğer pH’larda ise ortalama 5 mm’lik aktivite
zonu elde edilmiştir. On dokuz numaralı suşta pH 5.0 ve 8.0 de hem üreme hemde
aktivite gözlenmemiştir. Buna karşılık pH 10.0 da 3 mm koloni çapı ve 2 mm
aktivite zonu saptanmıştır. Diğer pH değerlerinde ise üreme gerçekleşmemiş fakat
ortalama 1 mm aktivite zonu elde edilmiştir. Yirmi Numaralı suşta bütün pH
değerlerinde üreme gerçekleşmiş ve ortalama 2 mm’lik koloniler üremiştir. Bütün pH
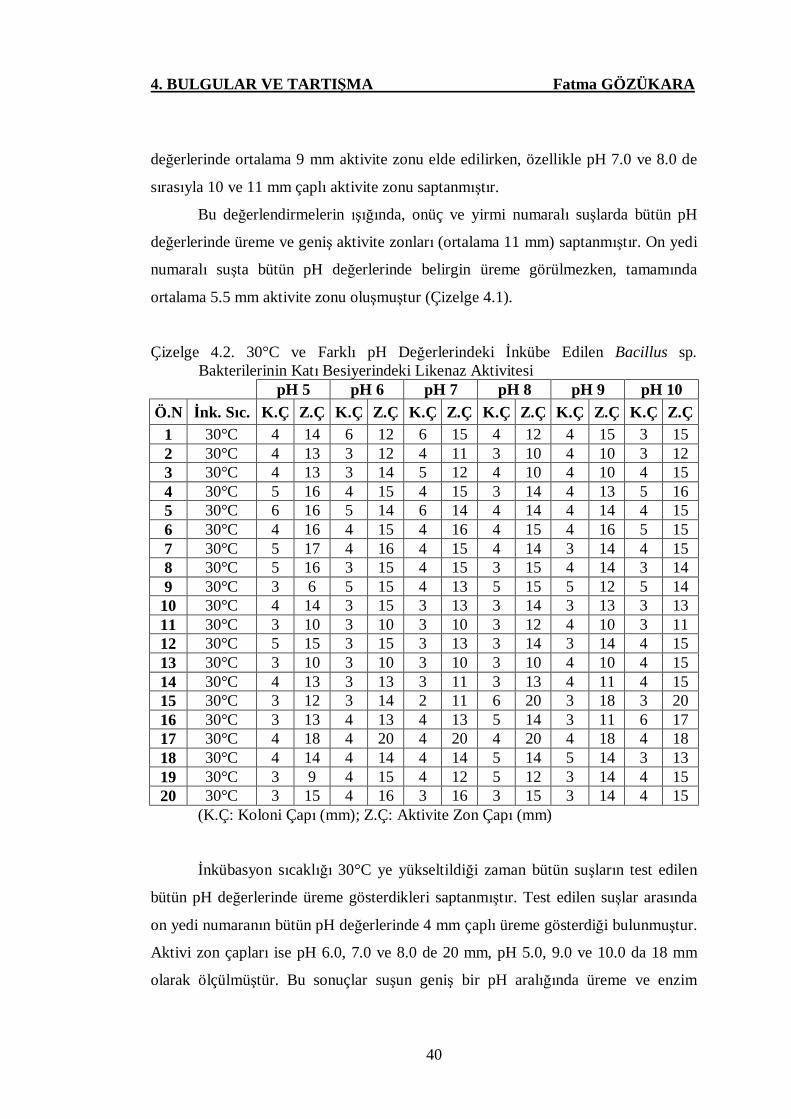
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
40
değerlerinde ortalama 9 mm aktivite zonu elde edilirken, özellikle pH 7.0 ve 8.0 de
sırasıyla 10 ve 11 mm çaplı aktivite zonu saptanmıştır.
Bu değerlendirmelerin ışığında, onüç ve yirmi numaralı suşlarda bütün pH
değerlerinde üreme ve geniş aktivite zonları (ortalama 11 mm) saptanmıştır. On yedi
numaralı suşta bütün pH değerlerinde belirgin üreme görülmezken, tamamında
ortalama 5.5 mm aktivite zonu oluşmuştur (Çizelge 4.1).
Çizelge 4.2. 30°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp. Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi
pH 5 pH 6 pH 7 pH 8 pH 9 pH 10 Ö.N İnk. Sıc. K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç
1 30°C 4 14 6 12 6 15 4 12 4 15 3 15 2 30°C 4 13 3 12 4 11 3 10 4 10 3 12 3 30°C 4 13 3 14 5 12 4 10 4 10 4 15 4 30°C 5 16 4 15 4 15 3 14 4 13 5 16 5 30°C 6 16 5 14 6 14 4 14 4 14 4 15 6 30°C 4 16 4 15 4 16 4 15 4 16 5 15 7 30°C 5 17 4 16 4 15 4 14 3 14 4 15 8 30°C 5 16 3 15 4 15 3 15 4 14 3 14 9 30°C 3 6 5 15 4 13 5 15 5 12 5 14
10 30°C 4 14 3 15 3 13 3 14 3 13 3 13 11 30°C 3 10 3 10 3 10 3 12 4 10 3 11 12 30°C 5 15 3 15 3 13 3 14 3 14 4 15 13 30°C 3 10 3 10 3 10 3 10 4 10 4 15 14 30°C 4 13 3 13 3 11 3 13 4 11 4 15 15 30°C 3 12 3 14 2 11 6 20 3 18 3 20 16 30°C 3 13 4 13 4 13 5 14 3 11 6 17 17 30°C 4 18 4 20 4 20 4 20 4 18 4 18 18 30°C 4 14 4 14 4 14 5 14 5 14 3 13 19 30°C 3 9 4 15 4 12 5 12 3 14 4 15 20 30°C 3 15 4 16 3 16 3 15 3 14 4 15
(K.Ç: Koloni Çapı (mm); Z.Ç: Aktivite Zon Çapı (mm)
İnkübasyon sıcaklığı 30°C ye yükseltildiği zaman bütün suşların test edilen
bütün pH değerlerinde üreme gösterdikleri saptanmıştır. Test edilen suşlar arasında
on yedi numaranın bütün pH değerlerinde 4 mm çaplı üreme gösterdiği bulunmuştur.
Aktivi zon çapları ise pH 6.0, 7.0 ve 8.0 de 20 mm, pH 5.0, 9.0 ve 10.0 da 18 mm
olarak ölçülmüştür. Bu sonuçlar suşun geniş bir pH aralığında üreme ve enzim

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
41
sentezleme yeteneğinde olduğunu göstermektedir. Suşlar arasından en iyi üreme pH
5 te beş numaralı şuşta 6 mm, pH 6 da bir numaralı suşta 6 mm, pH 7.0 de bir ve beş
numaralı suşlarda 6 mm, pH 8.0 de 15 numaralı suşta 6 mm, pH 9.0 da dokuz ve
onsekiz numaralı suşlarda 5 mm ve pH 10.0 da 16 numaralı suşta 6 mm olarak
gerçekleşmiştir. Aktivite zon çaplarının genişliği dikkate alındığında ise ph 5.0, 6.0
ve 7.0 de onyedi numaralı suş sırasıyla 18, 20 ve 20 mm ile en geniş zonu
oluşturmuştur. pH 8.0 de on beş ve on yedi numaralı suşlar 20 mm, pH 9.0 da on beş
ve on yedi numaralı suşlar 18 mm, pH 10.0 da ise on beş numaralı suş 20 mm, on
yedi numaralı suş 18 mm aktivite zonu oluşturmuşlardır. Bu sonuçlara göre bütün
suşların geniş bir pH aralığında (pH 5.0-10.0) üreme ve enzim sentezleme yeteneğine
sahip oldukları görülmektedir. Bununla birlikte özellikle on yedi numaralı suşun
bütün pH değerlerinde yüksek bir aktivite sahip olduğu saptanmıştır. On beş
numaralı suş ise daha çok pH 8.0-10.0 aralığında (alkali) geniş aktivite zonu
oluşturmuştur (Çizelge 4.2).
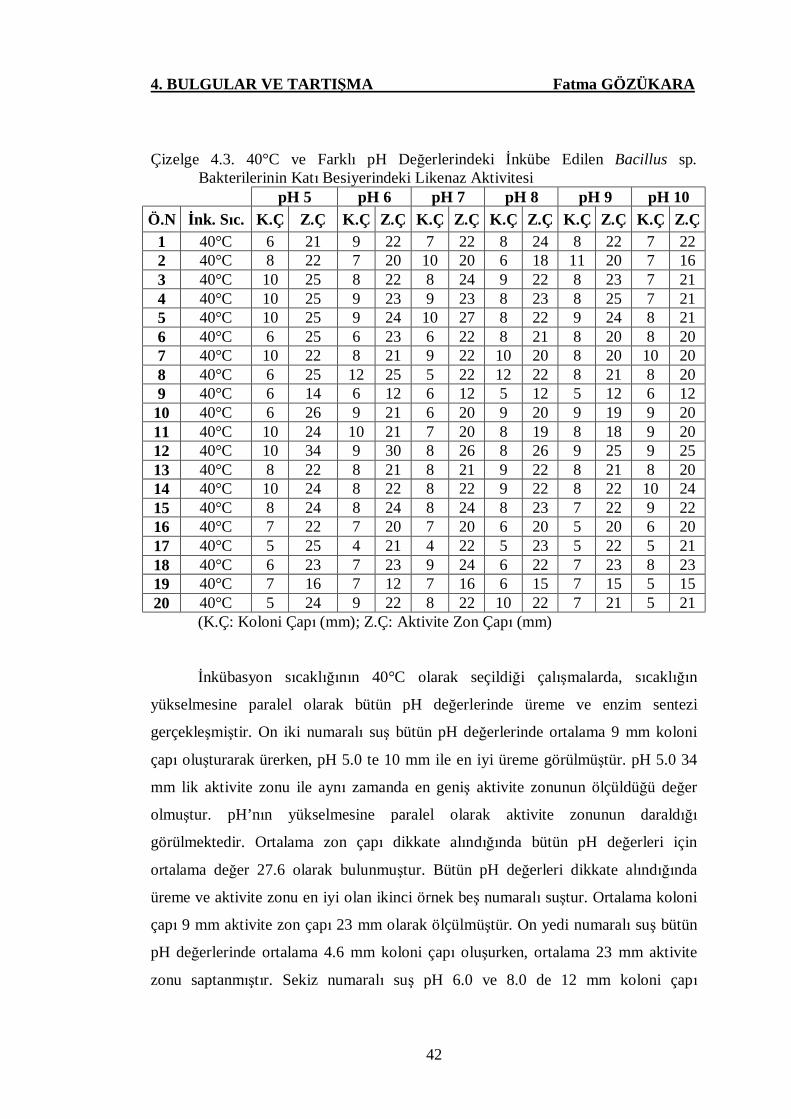
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
42
Çizelge 4.3. 40°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp. Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi
pH 5 pH 6 pH 7 pH 8 pH 9 pH 10 Ö.N İnk. Sıc. K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç
1 40°C 6 21 9 22 7 22 8 24 8 22 7 22 2 40°C 8 22 7 20 10 20 6 18 11 20 7 16 3 40°C 10 25 8 22 8 24 9 22 8 23 7 21 4 40°C 10 25 9 23 9 23 8 23 8 25 7 21 5 40°C 10 25 9 24 10 27 8 22 9 24 8 21 6 40°C 6 25 6 23 6 22 8 21 8 20 8 20 7 40°C 10 22 8 21 9 22 10 20 8 20 10 20 8 40°C 6 25 12 25 5 22 12 22 8 21 8 20 9 40°C 6 14 6 12 6 12 5 12 5 12 6 12
10 40°C 6 26 9 21 6 20 9 20 9 19 9 20 11 40°C 10 24 10 21 7 20 8 19 8 18 9 20 12 40°C 10 34 9 30 8 26 8 26 9 25 9 25 13 40°C 8 22 8 21 8 21 9 22 8 21 8 20 14 40°C 10 24 8 22 8 22 9 22 8 22 10 24 15 40°C 8 24 8 24 8 24 8 23 7 22 9 22 16 40°C 7 22 7 20 7 20 6 20 5 20 6 20 17 40°C 5 25 4 21 4 22 5 23 5 22 5 21 18 40°C 6 23 7 23 9 24 6 22 7 23 8 23 19 40°C 7 16 7 12 7 16 6 15 7 15 5 15 20 40°C 5 24 9 22 8 22 10 22 7 21 5 21
(K.Ç: Koloni Çapı (mm); Z.Ç: Aktivite Zon Çapı (mm)
İnkübasyon sıcaklığının 40°C olarak seçildiği çalışmalarda, sıcaklığın
yükselmesine paralel olarak bütün pH değerlerinde üreme ve enzim sentezi
gerçekleşmiştir. On iki numaralı suş bütün pH değerlerinde ortalama 9 mm koloni
çapı oluşturarak ürerken, pH 5.0 te 10 mm ile en iyi üreme görülmüştür. pH 5.0 34
mm lik aktivite zonu ile aynı zamanda en geniş aktivite zonunun ölçüldüğü değer
olmuştur. pH’nın yükselmesine paralel olarak aktivite zonunun daraldığı
görülmektedir. Ortalama zon çapı dikkate alındığında bütün pH değerleri için
ortalama değer 27.6 olarak bulunmuştur. Bütün pH değerleri dikkate alındığında
üreme ve aktivite zonu en iyi olan ikinci örnek beş numaralı suştur. Ortalama koloni
çapı 9 mm aktivite zon çapı 23 mm olarak ölçülmüştür. On yedi numaralı suş bütün
pH değerlerinde ortalama 4.6 mm koloni çapı oluşurken, ortalama 23 mm aktivite
zonu saptanmıştır. Sekiz numaralı suş pH 6.0 ve 8.0 de 12 mm koloni çapı

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
43
oluşturarak üreme gösterirken, aktivite zonu asidik pH değerlerinde daha geniş
ölçülmüştür. Genel olarak değerlendirildiğinde bütün suşların pH 5.0 ve 6.0 da hem
daha iyi üredikleri hemde daha geniş aktivite zonu oluşturdukları bulunmuştur.
Özellikle 3, 4, 5, 7, 11, 12 ve 14 numaralı suşlar diğerlerine göre (ortalama 10 mm
koloni çapı) daha iyi üreme göstermişlerdir. Aynı suşların aktivite zon çaplarıda
diğer suşlardan belirgin şekilde geniştir (pH 5.0 te 25.8 mm, pH 6.0 da 23.3mm)
(Çizelge 4.3).
Çizelge 4.4. 50°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp. Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi pH 5 pH 6 pH 7 pH 8 pH 9 pH 10
Ö.N İnk. Sıc. K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç 1 50°C 7 20 6 19 5 20 6 20 7 20 5 18 2 50°C 8 17 15 19 8 19 8 18 6 18 6 17 3 50°C 8 19 8 21 9 22 9 22 7 22 10 21 4 50°C 9 21 8 21 9 24 11 23 13 25 9 21 5 50°C 8 21 8 23 10 22 9 25 9 23 9 21 6 50°C 13 20 16 28 10 21 11 21 14 21 16 21 7 50°C 17 20 11 20 10 18 13 20 14 20 15 20 8 50°C 15 23 7 24 11 22 12 22 18 27 9 22 9 50°C 8 14 6 14 7 12 5 12 6 11 7 11 10 50°C 9 18 11 22 9 20 10 21 8 21 8 19 11 50°C 8 19 9 19 14 21 11 21 8 20 10 20 12 50°C 12 27 16 29 23 36 20 35 19 35 21 31 13 50°C 10 21 9 19 10 21 11 24 8 20 9 19 14 50°C 10 22 11 21 17 27 11 24 23 27 10 21 15 50°C 10 24 10 21 11 27 11 21 6 17 6 16 16 50°C 7 20 11 22 9 22 8 21 8 22 8 21 17 50°C 5 21 6 21 7 17 7 20 6 16 6 16 18 50°C 10 22 7 22 17 24 7 23 16 22 12 21 19 50°C 6 11 6 12 8 13 6 11 5 12 4 - 20 50°C 11 21 10 21 10 22 9 25 7 22 6 22
(K.Ç: Koloni Çapı (mm); Z.Ç: Aktivite Zon Çapı (mm)
İnkübasyon sıcaklığı 50°C ye yükseltildiği zaman bütün pH değerlerinde
bütün suşların daha iyi üredikleri görülmektedir. On iki numaralı suşun bütün pH
değerlerinde ortalama 18.5 mm koloni çapı oluşturarak ürediği saptanmıştır. En iyi
üreme ise 23 mm ile pH 7.0 de gerçekleşmiştir. En geniş aktivite zonu 36 mm ile pH

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
44
7.0 de gözlenirken, pH 8.0 ve 9.0 da 35 er mm’lik aktivite zonu ölçülmüştür. On iki
numaralı suşun bütün pH değerlerinde ortalama 32.6 mm aktivite zonuna sahip
olduğu saptanmıştır. On iki numaralı suşta pH 7.0 den itibaren hem koloni çapı
hemde aktivite zon genişliği artmıştır. Zon koloni oranlaması yapıldığında aktivite
zonunun koloni çapına göre 1.6 kat olduğu görülmektedir. On yedi numaralı suşta ise
üreme ve zon oluşumu pH 5.0 ve 6.0 da daha yüksek (ortalama koloni çapı 5.5 mm,
zon çapı 21 mm) iken pH arttıkça üremenin daha iyi olduğu buna kariışık aktivite
zonunun daraldığı görülmektedir. On yedi numaralı suşta zon koloni oranlaması
yapıldığı zaman zon çapının koloni çapından ortalama 4 kat geniş olduğu
görülmektedir. Ondokuz numaralı suşta pH 10 da 4 mm koloni çapı oluşurken,
aktivite zonu saptanmamıştır. Bu sonuca göre ondokuz numaralı suşun enzim
sentezleme yeteneğinin pH 9.0 da sona erdiği görülmektedir (Çizelge 4.4).
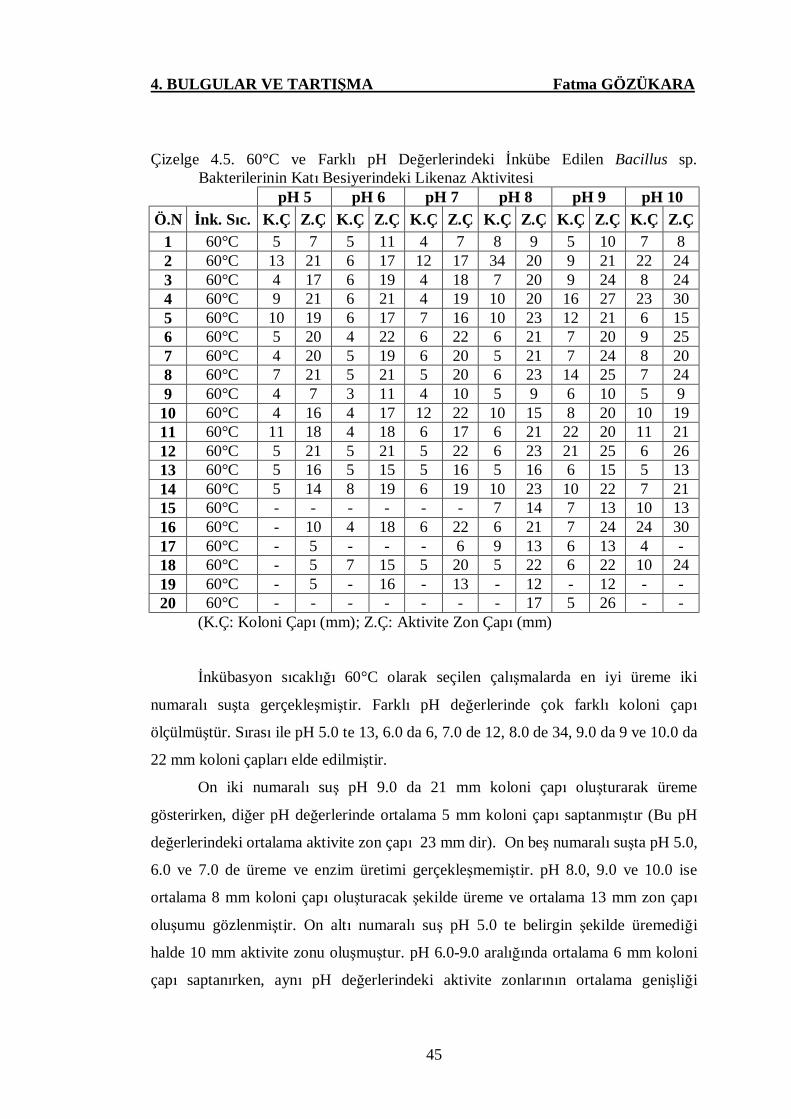
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
45
Çizelge 4.5. 60°C ve Farklı pH Değerlerindeki İnkübe Edilen Bacillus sp. Bakterilerinin Katı Besiyerindeki Likenaz Aktivitesi pH 5 pH 6 pH 7 pH 8 pH 9 pH 10
Ö.N İnk. Sıc. K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç K.Ç Z.Ç 1 60°C 5 7 5 11 4 7 8 9 5 10 7 8 2 60°C 13 21 6 17 12 17 34 20 9 21 22 24 3 60°C 4 17 6 19 4 18 7 20 9 24 8 24 4 60°C 9 21 6 21 4 19 10 20 16 27 23 30 5 60°C 10 19 6 17 7 16 10 23 12 21 6 15 6 60°C 5 20 4 22 6 22 6 21 7 20 9 25 7 60°C 4 20 5 19 6 20 5 21 7 24 8 20 8 60°C 7 21 5 21 5 20 6 23 14 25 7 24 9 60°C 4 7 3 11 4 10 5 9 6 10 5 9
10 60°C 4 16 4 17 12 22 10 15 8 20 10 19 11 60°C 11 18 4 18 6 17 6 21 22 20 11 21 12 60°C 5 21 5 21 5 22 6 23 21 25 6 26 13 60°C 5 16 5 15 5 16 5 16 6 15 5 13 14 60°C 5 14 8 19 6 19 10 23 10 22 7 21 15 60°C - - - - - - 7 14 7 13 10 13 16 60°C - 10 4 18 6 22 6 21 7 24 24 30 17 60°C - 5 - - - 6 9 13 6 13 4 - 18 60°C - 5 7 15 5 20 5 22 6 22 10 24 19 60°C - 5 - 16 - 13 - 12 - 12 - - 20 60°C - - - - - - - 17 5 26 - -
(K.Ç: Koloni Çapı (mm); Z.Ç: Aktivite Zon Çapı (mm)
İnkübasyon sıcaklığı 60°C olarak seçilen çalışmalarda en iyi üreme iki
numaralı suşta gerçekleşmiştir. Farklı pH değerlerinde çok farklı koloni çapı
ölçülmüştür. Sırası ile pH 5.0 te 13, 6.0 da 6, 7.0 de 12, 8.0 de 34, 9.0 da 9 ve 10.0 da
22 mm koloni çapları elde edilmiştir.
On iki numaralı suş pH 9.0 da 21 mm koloni çapı oluşturarak üreme
gösterirken, diğer pH değerlerinde ortalama 5 mm koloni çapı saptanmıştır (Bu pH
değerlerindeki ortalama aktivite zon çapı 23 mm dir). On beş numaralı suşta pH 5.0,
6.0 ve 7.0 de üreme ve enzim üretimi gerçekleşmemiştir. pH 8.0, 9.0 ve 10.0 ise
ortalama 8 mm koloni çapı oluşturacak şekilde üreme ve ortalama 13 mm zon çapı
oluşumu gözlenmiştir. On altı numaralı suş pH 5.0 te belirgin şekilde üremediği
halde 10 mm aktivite zonu oluşmuştur. pH 6.0-9.0 aralığında ortalama 6 mm koloni
çapı saptanırken, aynı pH değerlerindeki aktivite zonlarının ortalama genişliği

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
46
ortalama 21 mm olmuştur. pH 10.0 da ise koloni çapı 24 mm, aktivite zon genişliği
30 mm olmuştur. Yirmi numaralı suş ile yapılan çalışmalarda pH 5.0-7.0 aralığında
üreme ve enzim sentezi saptanmamıştır. pH 8.0 de üreme gerçekleşmediği halde
(belirgin) 17 mm genişliğinde aktivite zonu oluşmuştur. pH 9.0 da 5 mm çapında
koloni oluşumu ve 26 mm genişliğide aktivite zonu gözlenmiştir. pH 10.0 da ise
üreme ve enzim sentezi gerçekleşmemiştir (Çizelge 4.6).
Bu sıcaklık değerindeki üreme ve aktivite zon çapının ortalama genişlikleri
dikkate alındığında on iki numaralı suşta aktivite zon genişliğinin koloni çapının
ortalama üç katı olduğu saptanmıştır. Bu özelliği, bütün pH değerlerinde birbirine
yakın aktivite zon genişliği oluşturması ve bütün sıcaklık değerlerinde üreme ve
yüksek enzim sentezi gerçekleştirmesi nedeni ile on iki numaralı suş, çalışmanın
amaçları doğrultusunda enzim üreticisi olarak seçilmiştir.
Literatür verilerinde Bacillus sp. sularının katı besiyerinde farklı pH ve
sıcaklık değerlerinde üreme ve aktivite zonu ile ilgili değişik çalışmalar vardır.
Lin ve ark (1998), Bacillus sp. TS-23 suşunun enzim sentezinin 42-60°C
sıcaklıkta ve optimum 55°C’de meydana geldiğini belirtmişlerdir. Hakamada ve ark
(2002), Bacillus circulans’dan izole ettikleri alkalin endoglukanazın pH 8.5’da
55°C’de optimal aktivite gösterdiğini saptamışlardır. L-12 suşu ile yapılan
çalışmalarda optimum enzim sentezinin 50°C de gerçekleştiğinin bulunması
Hakamada ve arkadaşlarının bulguları ile tam bir uyum göstermektedir. Yine, Bajbai
ve Bajbai (1987)’de yapmış oldukları çalışmalarda Bacillus licheniformis TCRDC-
B13 suşu için enzim üretimi ve üreme sıcaklığının 25-50°C de ve maksimum enzim
üretiminin ise 35°C’de gerçekleştiğini bulmuşlardır.
Katı besiyerinde farklı sıcaklık ve pH değerlerinde L-12 suşu ve enzim
aktivitesine ait görüntüler şekil 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 ve 15 de
görülmektedir.

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
47
Şekil 4.1. L-12 suşunun katı besiyerinde üremesi 20°C, pH 5.0, 6.0, 7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm.
Şekil 4.2. L-12 suşunun katı besiyerinde üremesi 20°C, pH 9.0 ve 10.0 da gerçekleşen enzim sentezine ait görünüm.

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
48
Şekil 4.3. L-12 suşunun katı besiyerinde üremesi 30°C, pH 5.0, 6.0, 7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm.
Şekil 4.4. L-12 suşunun katı besiyerinde üremesi 20°C, pH 9.0 ve 10.0 da gerçekleşen enzim sentezine ait görünüm.
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
49
Şekil 4.5. L-12 suşunun katı besiyerinde üremesi 40°C, pH 5.0, 6.0, 7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm.
Şekil 4.6. L-12 suşunun katı besiyerinde üremesi 40°C, pH 9.0 ve 10.0 da gerçekleşen enzim sentezine ait görünüm.

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
50
Şekil 4.7. L-12 suşunun katı besiyerinde üremesi 50°C, pH 5.0, 6.0, 7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm.
Şekil 4.8. L-12 suşunun katı besiyerinde üremesi 50°C, pH 9.0 ve 10.0 da gerçekleşen enzim sentezine ait görünüm.
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
51
Şekil 4.9. L-12 suşunun katı besiyerinde üremesi 60°C, pH 5.0, 6.0, 7.0 ve 8.0 de gerçekleşen enzim sentezine ait görünüm.
Şekil 4.10. L-12 suşunun katı besiyerinde üremesi 60°C, pH 9.0 ve 10.0 da gerçekleşen enzim sentezine ait görünüm.
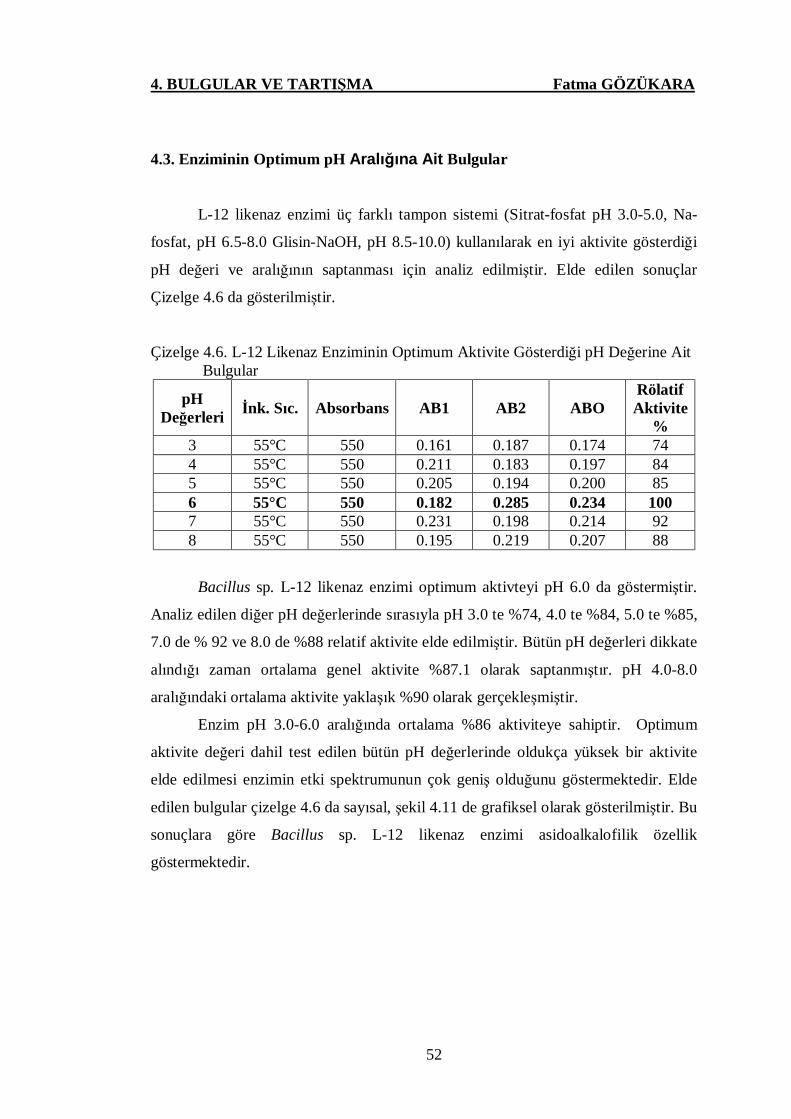
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
52
4.3. Enziminin Optimum pH Aralığına Ait Bulgular
L-12 likenaz enzimi üç farklı tampon sistemi (Sitrat-fosfat pH 3.0-5.0, Na-
fosfat, pH 6.5-8.0 Glisin-NaOH, pH 8.5-10.0) kullanılarak en iyi aktivite gösterdiği
pH değeri ve aralığının saptanması için analiz edilmiştir. Elde edilen sonuçlar
Çizelge 4.6 da gösterilmiştir.
Çizelge 4.6. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği pH Değerine Ait Bulgular
pH Değerleri İnk. Sıc. Absorbans AB1 AB2 ABO
Rölatif Aktivite
% 3 55°C 550 0.161 0.187 0.174 74 4 55°C 550 0.211 0.183 0.197 84 5 55°C 550 0.205 0.194 0.200 85 6 55°C 550 0.182 0.285 0.234 100 7 55°C 550 0.231 0.198 0.214 92 8 55°C 550 0.195 0.219 0.207 88
Bacillus sp. L-12 likenaz enzimi optimum aktivteyi pH 6.0 da göstermiştir.
Analiz edilen diğer pH değerlerinde sırasıyla pH 3.0 te %74, 4.0 te %84, 5.0 te %85,
7.0 de % 92 ve 8.0 de %88 relatif aktivite elde edilmiştir. Bütün pH değerleri dikkate
alındığı zaman ortalama genel aktivite %87.1 olarak saptanmıştır. pH 4.0-8.0
aralığındaki ortalama aktivite yaklaşık %90 olarak gerçekleşmiştir.
Enzim pH 3.0-6.0 aralığında ortalama %86 aktiviteye sahiptir. Optimum
aktivite değeri dahil test edilen bütün pH değerlerinde oldukça yüksek bir aktivite
elde edilmesi enzimin etki spektrumunun çok geniş olduğunu göstermektedir. Elde
edilen bulgular çizelge 4.6 da sayısal, şekil 4.11 de grafiksel olarak gösterilmiştir. Bu
sonuçlara göre Bacillus sp. L-12 likenaz enzimi asidoalkalofilik özellik
göstermektedir.
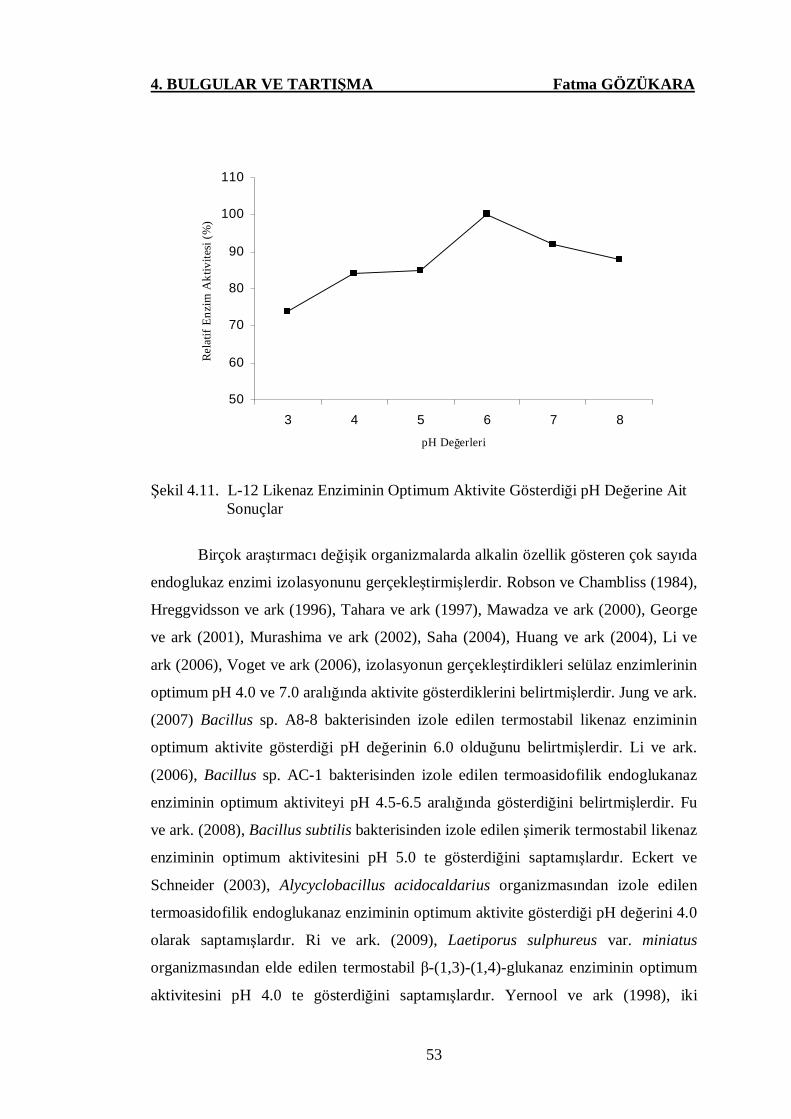
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
53
50
60
70
80
90
100
110
3 4 5 6 7 8
pH Değerleri
Rela
tif E
nzim
Akt
ivite
si (%
)
Şekil 4.11. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği pH Değerine Ait
Sonuçlar
Birçok araştırmacı değişik organizmalarda alkalin özellik gösteren çok sayıda
endoglukaz enzimi izolasyonunu gerçekleştirmişlerdir. Robson ve Chambliss (1984),
Hreggvidsson ve ark (1996), Tahara ve ark (1997), Mawadza ve ark (2000), George
ve ark (2001), Murashima ve ark (2002), Saha (2004), Huang ve ark (2004), Li ve
ark (2006), Voget ve ark (2006), izolasyonun gerçekleştirdikleri selülaz enzimlerinin
optimum pH 4.0 ve 7.0 aralığında aktivite gösterdiklerini belirtmişlerdir. Jung ve ark.
(2007) Bacillus sp. A8-8 bakterisinden izole edilen termostabil likenaz enziminin
optimum aktivite gösterdiği pH değerinin 6.0 olduğunu belirtmişlerdir. Li ve ark.
(2006), Bacillus sp. AC-1 bakterisinden izole edilen termoasidofilik endoglukanaz
enziminin optimum aktiviteyi pH 4.5-6.5 aralığında gösterdiğini belirtmişlerdir. Fu
ve ark. (2008), Bacillus subtilis bakterisinden izole edilen şimerik termostabil likenaz
enziminin optimum aktivitesini pH 5.0 te gösterdiğini saptamışlardır. Eckert ve
Schneider (2003), Alycyclobacillus acidocaldarius organizmasından izole edilen
termoasidofilik endoglukanaz enziminin optimum aktivite gösterdiği pH değerini 4.0
olarak saptamışlardır. Ri ve ark. (2009), Laetiporus sulphureus var. miniatus
organizmasından elde edilen termostabil β-(1,3)-(1,4)-glukanaz enziminin optimum
aktivitesini pH 4.0 te gösterdiğini saptamışlardır. Yernool ve ark (1998), iki
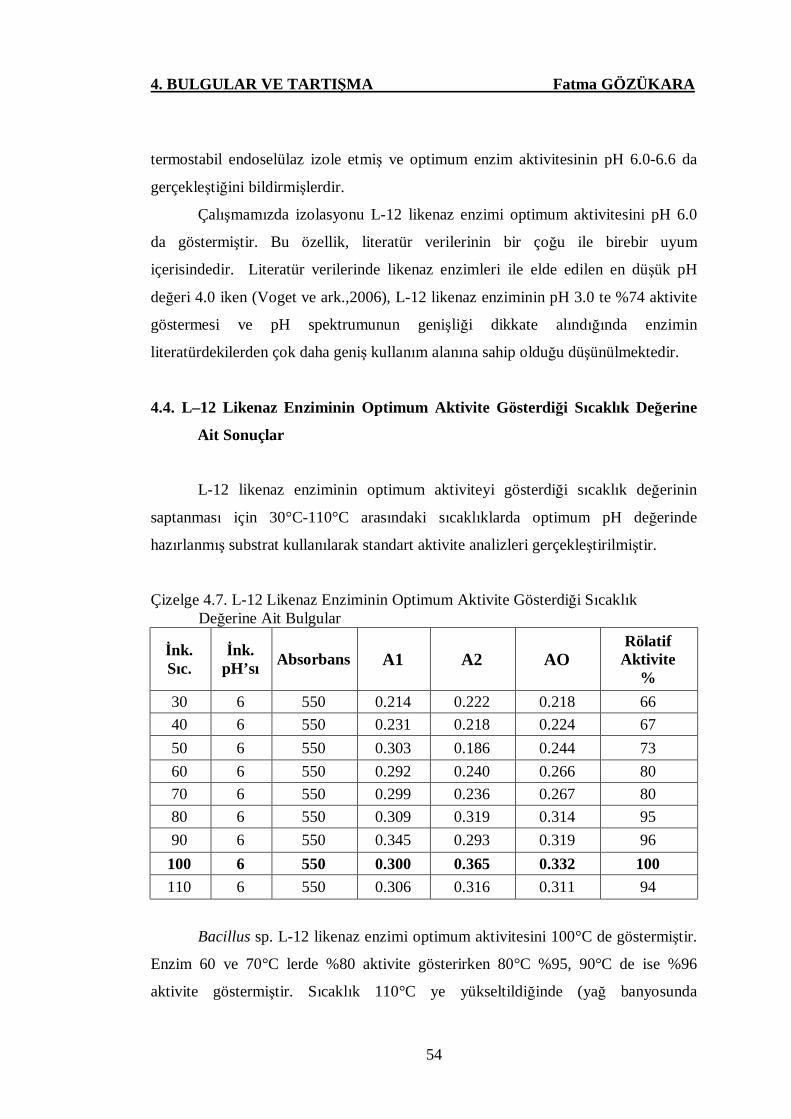
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
54
termostabil endoselülaz izole etmiş ve optimum enzim aktivitesinin pH 6.0-6.6 da
gerçekleştiğini bildirmişlerdir.
Çalışmamızda izolasyonu L-12 likenaz enzimi optimum aktivitesini pH 6.0
da göstermiştir. Bu özellik, literatür verilerinin bir çoğu ile birebir uyum
içerisindedir. Literatür verilerinde likenaz enzimleri ile elde edilen en düşük pH
değeri 4.0 iken (Voget ve ark.,2006), L-12 likenaz enziminin pH 3.0 te %74 aktivite
göstermesi ve pH spektrumunun genişliği dikkate alındığında enzimin
literatürdekilerden çok daha geniş kullanım alanına sahip olduğu düşünülmektedir.
4.4. L–12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık Değerine
Ait Sonuçlar
L-12 likenaz enziminin optimum aktiviteyi gösterdiği sıcaklık değerinin
saptanması için 30°C-110°C arasındaki sıcaklıklarda optimum pH değerinde
hazırlanmış substrat kullanılarak standart aktivite analizleri gerçekleştirilmiştir.
Çizelge 4.7. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık Değerine Ait Bulgular
İnk. Sıc.
İnk. pH’sı Absorbans A1 A2 AO
Rölatif Aktivite
% 30 6 550 0.214 0.222 0.218 66 40 6 550 0.231 0.218 0.224 67 50 6 550 0.303 0.186 0.244 73 60 6 550 0.292 0.240 0.266 80 70 6 550 0.299 0.236 0.267 80 80 6 550 0.309 0.319 0.314 95 90 6 550 0.345 0.293 0.319 96
100 6 550 0.300 0.365 0.332 100 110 6 550 0.306 0.316 0.311 94
Bacillus sp. L-12 likenaz enzimi optimum aktivitesini 100°C de göstermiştir.
Enzim 60 ve 70°C lerde %80 aktivite gösterirken 80°C %95, 90°C de ise %96
aktivite göstermiştir. Sıcaklık 110°C ye yükseltildiğinde (yağ banyosunda
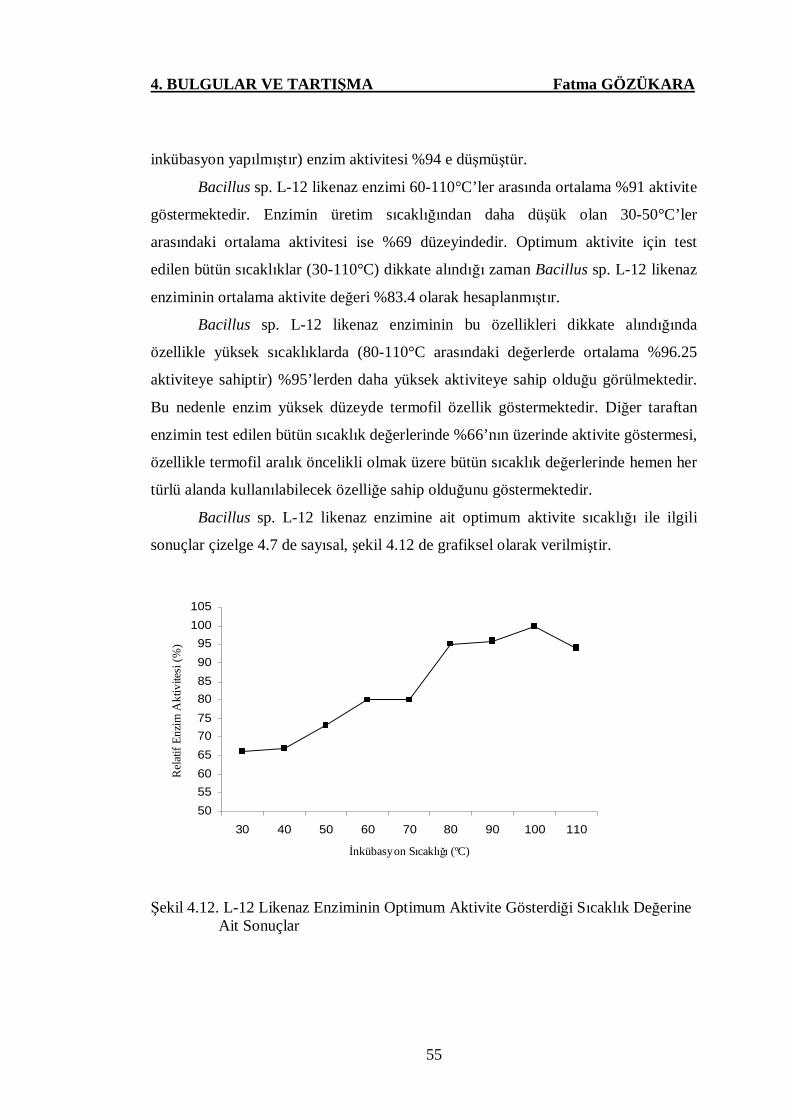
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
55
inkübasyon yapılmıştır) enzim aktivitesi %94 e düşmüştür.
Bacillus sp. L-12 likenaz enzimi 60-110°C’ler arasında ortalama %91 aktivite
göstermektedir. Enzimin üretim sıcaklığından daha düşük olan 30-50°C’ler
arasındaki ortalama aktivitesi ise %69 düzeyindedir. Optimum aktivite için test
edilen bütün sıcaklıklar (30-110°C) dikkate alındığı zaman Bacillus sp. L-12 likenaz
enziminin ortalama aktivite değeri %83.4 olarak hesaplanmıştır.
Bacillus sp. L-12 likenaz enziminin bu özellikleri dikkate alındığında
özellikle yüksek sıcaklıklarda (80-110°C arasındaki değerlerde ortalama %96.25
aktiviteye sahiptir) %95’lerden daha yüksek aktiviteye sahip olduğu görülmektedir.
Bu nedenle enzim yüksek düzeyde termofil özellik göstermektedir. Diğer taraftan
enzimin test edilen bütün sıcaklık değerlerinde %66’nın üzerinde aktivite göstermesi,
özellikle termofil aralık öncelikli olmak üzere bütün sıcaklık değerlerinde hemen her
türlü alanda kullanılabilecek özelliğe sahip olduğunu göstermektedir.
Bacillus sp. L-12 likenaz enzimine ait optimum aktivite sıcaklığı ile ilgili
sonuçlar çizelge 4.7 de sayısal, şekil 4.12 de grafiksel olarak verilmiştir.
50556065707580859095
100105
30 40 50 60 70 80 90 100 110
İnkübasyon Sıcaklığı (ºC)
Rel
atif
Enzi
m A
ktiv
itesi
(%)
Şekil 4.12. L-12 Likenaz Enziminin Optimum Aktivite Gösterdiği Sıcaklık Değerine
Ait Sonuçlar

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
56
Literatür verilerinde optimum aktivite sıcaklığı farklı olan çok sayıda
endoglukanaz enzimi yer almaktadır. Bunlar içerisinde asidik özellikte olan
endoglukanaz enzimlerinin optimum aktivite gösterdiği sıcaklık değerleri farklılık
göstermektedir. Mawadza ve ark, (2000), izole edilen endoglukanaz enziminin
optimum aktivitesini 65-70°C, Li ve ark. (2006), elde edilen endoglukanaz enziminin
70°C Huang ve Monk (2004) izole ettikleri endoglukanaz enziminin optimum relatif
aktivitesini 80°C de gösterdiğini saptamışlardır. Bischoff ve ark, (2006), termofilik
Bacillus licheniformis B-41361’den izole edilen endoglukanaz enziminin optimum
sıcaklığını 65ºC olarak belirtmişlerdir. Ri ve ark. (2009), Laetiporus sulphureus var.
miniatus suşundan elde edilen β-(1,3)-(1,4)-glukanaz enziminin optimum aktivitesini
75°C de gösterdiğini bulmuşlardır.
Yernool ve ark (1998), izolasyonunu gerçekleştirdikleri iki termostabil
endoglukanaz enziminin optimum aktivitesini 95°C de gösterdiğini saptamışlardır.
Zverlov ve ark. (1997) Thermotoga neopolitana suşundan izole edilen yüksek
düzeyde termostabil 1,3-beta endoglukanaz enzimlerinden 73 kDa büyüklükteki
fragmentin optimum aktivitesini 95°C, 52 kDa büyüklükteki fragmentin 85°C de
göstediğini saptamışlardır. Çalışmamızda Termofil Bacillus sp. suşundan izole
ettiğimiz L-12 likenaz enzimi optimum aktivitesini 100ºC de göstermektedir.
Literatür verilerinden daha yüksek optimum aktivite sıcaklığına sahip olan enzim,
hiper termofilik özelliktedir.
4.5. L-12 Likenaz Enziminin pH Stabilitesine Ait Sonuçlar
Bacillus sp. L-12 likenaz enzimi pH stabilitesinin saptanması için pH 3.0-
10.0 aralığında 24 saat 37ºC de ön inkübasyona tabi tutulmuştur. Sıvılaştırma
tamponu içerisindeki enzimle başlangıç aktivite değerinin saptanması için analiz
yapılmış ve bu değer 100 şeklinde alınarak, ön inkübasyon sonucu elde edilen
değerler oranlanmışlardır. Analizler sonunda elde edilen bulgular Çizelge 4.8 de
gösterilmiştir.
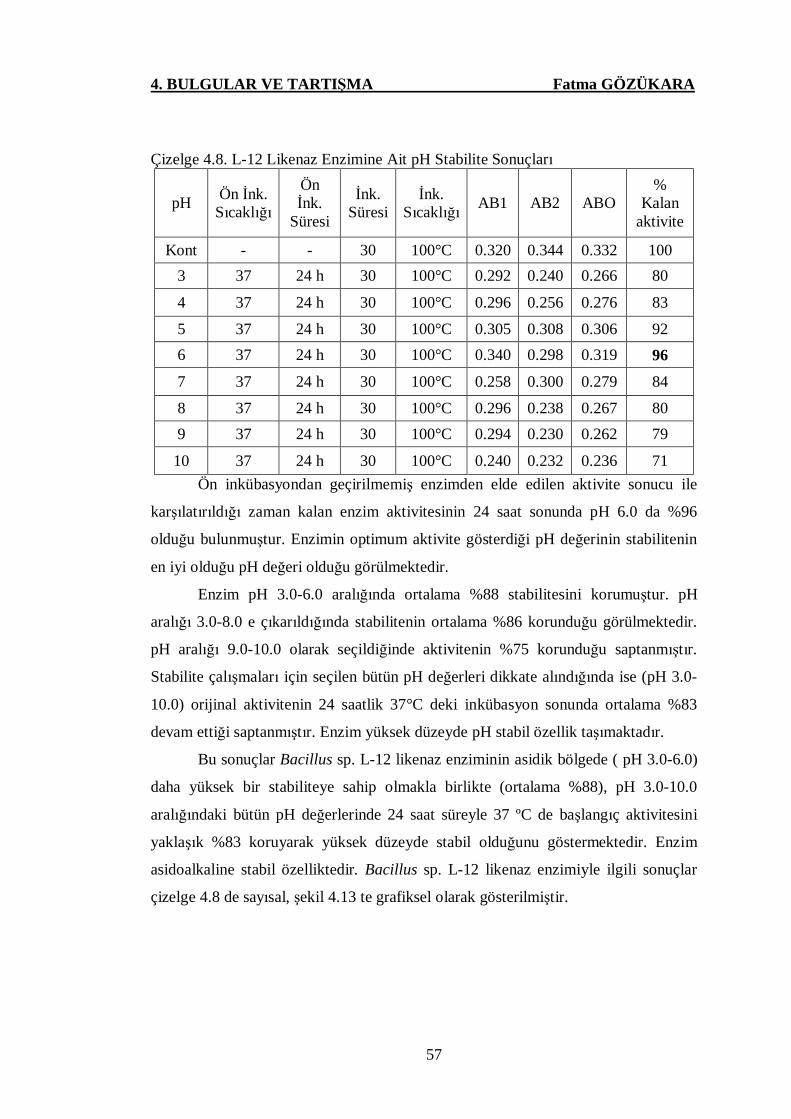
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
57
Çizelge 4.8. L-12 Likenaz Enzimine Ait pH Stabilite Sonuçları
pH Ön İnk. Sıcaklığı
Ön İnk.
Süresi
İnk. Süresi
İnk. Sıcaklığı AB1 AB2 ABO
% Kalan
aktivite
Kont - - 30 100°C 0.320 0.344 0.332 100 3 37 24 h 30 100°C 0.292 0.240 0.266 80
4 37 24 h 30 100°C 0.296 0.256 0.276 83
5 37 24 h 30 100°C 0.305 0.308 0.306 92 6 37 24 h 30 100°C 0.340 0.298 0.319 96 7 37 24 h 30 100°C 0.258 0.300 0.279 84
8 37 24 h 30 100°C 0.296 0.238 0.267 80 9 37 24 h 30 100°C 0.294 0.230 0.262 79
10 37 24 h 30 100°C 0.240 0.232 0.236 71 Ön inkübasyondan geçirilmemiş enzimden elde edilen aktivite sonucu ile
karşılatırıldığı zaman kalan enzim aktivitesinin 24 saat sonunda pH 6.0 da %96
olduğu bulunmuştur. Enzimin optimum aktivite gösterdiği pH değerinin stabilitenin
en iyi olduğu pH değeri olduğu görülmektedir.
Enzim pH 3.0-6.0 aralığında ortalama %88 stabilitesini korumuştur. pH
aralığı 3.0-8.0 e çıkarıldığında stabilitenin ortalama %86 korunduğu görülmektedir.
pH aralığı 9.0-10.0 olarak seçildiğinde aktivitenin %75 korunduğu saptanmıştır.
Stabilite çalışmaları için seçilen bütün pH değerleri dikkate alındığında ise (pH 3.0-
10.0) orijinal aktivitenin 24 saatlik 37°C deki inkübasyon sonunda ortalama %83
devam ettiği saptanmıştır. Enzim yüksek düzeyde pH stabil özellik taşımaktadır.
Bu sonuçlar Bacillus sp. L-12 likenaz enziminin asidik bölgede ( pH 3.0-6.0)
daha yüksek bir stabiliteye sahip olmakla birlikte (ortalama %88), pH 3.0-10.0
aralığındaki bütün pH değerlerinde 24 saat süreyle 37 ºC de başlangıç aktivitesini
yaklaşık %83 koruyarak yüksek düzeyde stabil olduğunu göstermektedir. Enzim
asidoalkaline stabil özelliktedir. Bacillus sp. L-12 likenaz enzimiyle ilgili sonuçlar
çizelge 4.8 de sayısal, şekil 4.13 te grafiksel olarak gösterilmiştir.
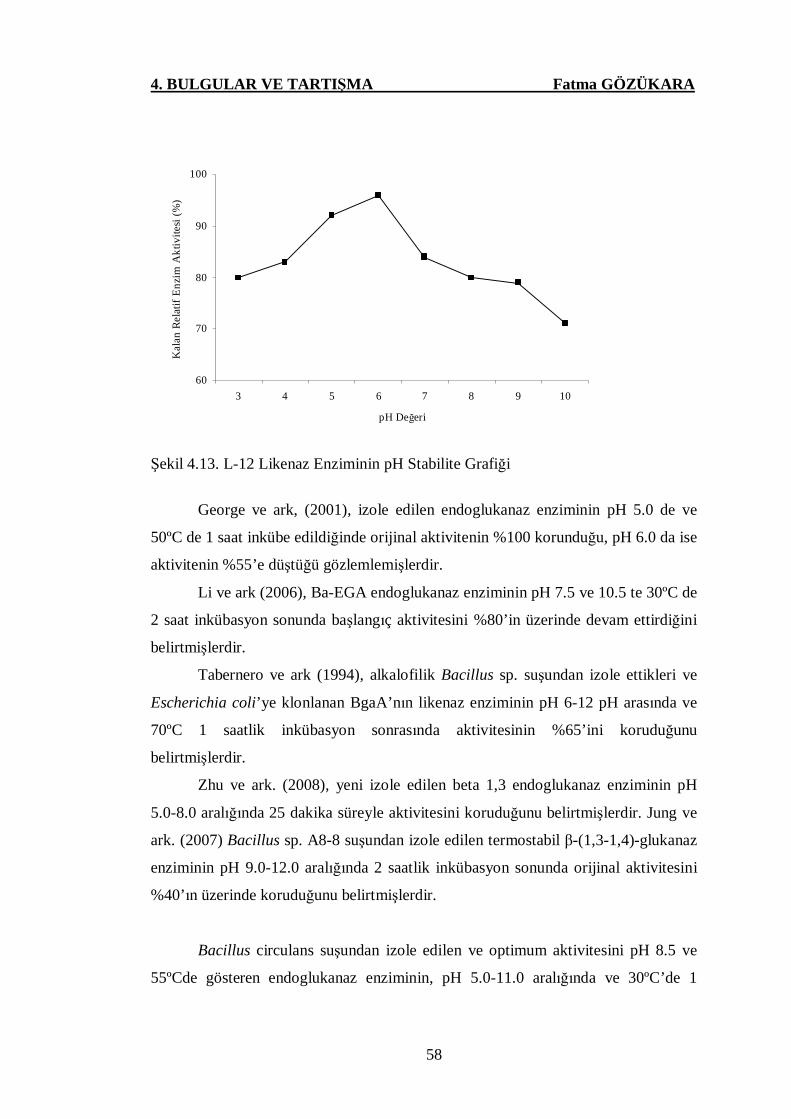
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
58
60
70
80
90
100
3 4 5 6 7 8 9 10
pH Değeri
Kal
an R
elat
if En
zim
Akt
ivite
si (%
)
Şekil 4.13. L-12 Likenaz Enziminin pH Stabilite Grafiği
George ve ark, (2001), izole edilen endoglukanaz enziminin pH 5.0 de ve
50ºC de 1 saat inkübe edildiğinde orijinal aktivitenin %100 korunduğu, pH 6.0 da ise
aktivitenin %55’e düştüğü gözlemlemişlerdir.
Li ve ark (2006), Ba-EGA endoglukanaz enziminin pH 7.5 ve 10.5 te 30ºC de
2 saat inkübasyon sonunda başlangıç aktivitesini %80’in üzerinde devam ettirdiğini
belirtmişlerdir.
Tabernero ve ark (1994), alkalofilik Bacillus sp. suşundan izole ettikleri ve
Escherichia coli’ye klonlanan BgaA’nın likenaz enziminin pH 6-12 pH arasında ve
70ºC 1 saatlik inkübasyon sonrasında aktivitesinin %65’ini koruduğunu
belirtmişlerdir.
Zhu ve ark. (2008), yeni izole edilen beta 1,3 endoglukanaz enziminin pH
5.0-8.0 aralığında 25 dakika süreyle aktivitesini koruduğunu belirtmişlerdir. Jung ve
ark. (2007) Bacillus sp. A8-8 suşundan izole edilen termostabil β-(1,3-1,4)-glukanaz
enziminin pH 9.0-12.0 aralığında 2 saatlik inkübasyon sonunda orijinal aktivitesini
%40’ın üzerinde koruduğunu belirtmişlerdir.
Bacillus circulans suşundan izole edilen ve optimum aktivitesini pH 8.5 ve
55ºCde gösteren endoglukanaz enziminin, pH 5.0-11.0 aralığında ve 30ºC’de 1

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
59
saatlik inkübasyondan sonra orijinal aktivitesini %100’e yakın koruduğu
bulunmuştur (Hakamada ve ark, 2002). Bacillus sp. HSH-810 suşundan izole edilen
alkalin endoglukanaz enziminin pH 7.0–9.0 aralığında ve 50ºC 10 dakika inkübe
edildiğinde aktivitenin yaklaşık %10 kaybolduğu saptanmıştır (Kim ve ark, 2005).
L–12 likenaz enzimin 24 saat süreyle 37ºC de ön inkübasyondan geçirilip
aktivite analizi yapıldığı zaman pH 3.0–10.0 aralığında orjinal aktivitesini ortalama
%83.1 koruduğu bulunmuştur. Ön inkübasyon süresinin uzunluğu ve stabilite
sonuçlarının yüksekliği L-12 likenaz enziminin literatür bilgilerinden çok daha
yüksek ve geniş pH aralığında stabiliteye sahip olduğunu göstermektedir. L-12
likenaz enzimi asido-alkali pH stabil özelliktedir.
4.6. L–12 Likenaz Enziminin Termal Stabilite Analizlerine Ait Sonuçlar
L–12 likenaz enziminin termal stabilitesini saptamak için 30–110ºC
aralığında 15 dakikalık ön inkübasyon uygulamaları gerçekleştirilmiştir. Enzimin 30-
60 ºC aralığındaki sıcaklık değerlerinde orijinal aktivitesini ortalama %92.5, 30-70
ºC aralığında ortalama %90.6, 70-90 ºC aralığında ortalama %79, ve 100-110 ºC
aralığında ortalama %66.5 koruduğu belirlenmiştir.
Ön inkübasyon sıcaklık aralığı 80-110 ºC seçildiğinde enzimin ortalama %72
kalan aktiviteye sahip olduğu bulunmuştur. Ön inkübasyon işleminden sonra ölçülen
kalan aktivite değerleri bütün sıcaklıklar dikkate alındığında ortalama %82.2 olarak
saptanmıştır.
Elde edilen sonuçlar sayısal olarak çizelge 4.9, grafiksel olarak şekil 4.12 de
gösterilmiştir. Bu bulgular likenaz enziminin termostabil özellikte olduğunu
göstermektedir.
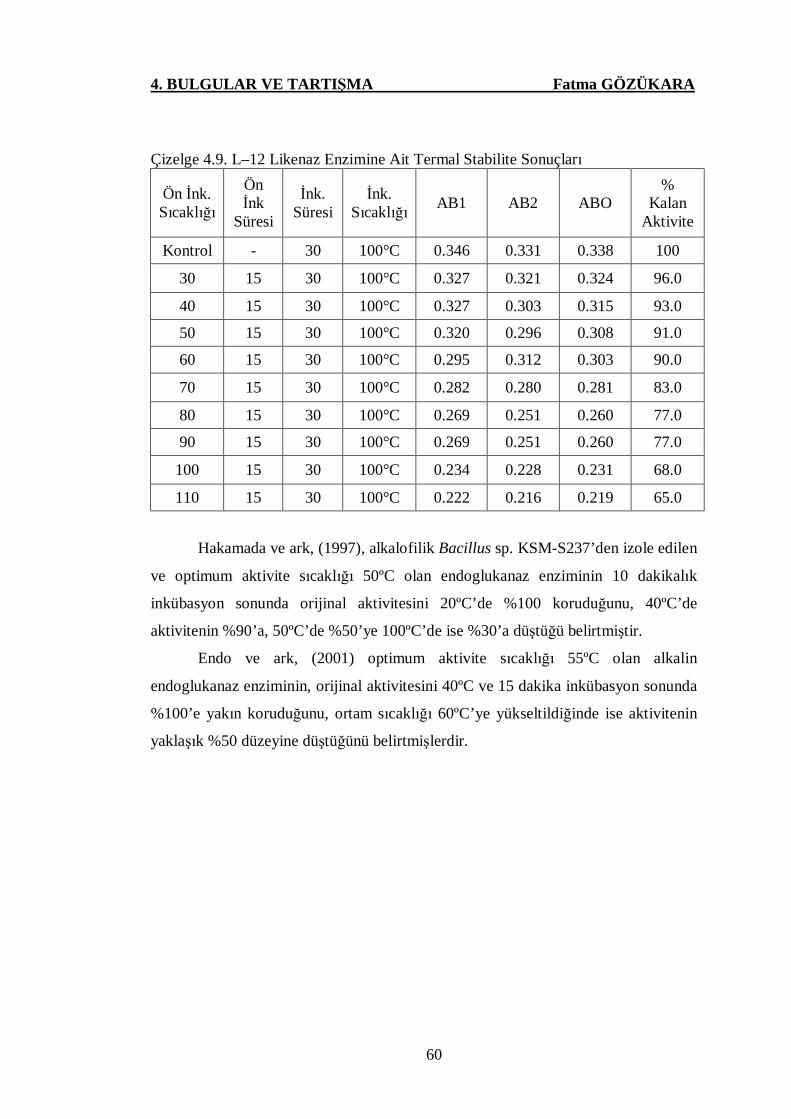
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
60
Çizelge 4.9. L–12 Likenaz Enzimine Ait Termal Stabilite Sonuçları
Ön İnk. Sıcaklığı
Ön İnk
Süresi
İnk. Süresi
İnk. Sıcaklığı AB1 AB2 ABO
% Kalan
Aktivite
Kontrol - 30 100°C 0.346 0.331 0.338 100
30 15 30 100°C 0.327 0.321 0.324 96.0
40 15 30 100°C 0.327 0.303 0.315 93.0
50 15 30 100°C 0.320 0.296 0.308 91.0
60 15 30 100°C 0.295 0.312 0.303 90.0
70 15 30 100°C 0.282 0.280 0.281 83.0
80 15 30 100°C 0.269 0.251 0.260 77.0
90 15 30 100°C 0.269 0.251 0.260 77.0
100 15 30 100°C 0.234 0.228 0.231 68.0
110 15 30 100°C 0.222 0.216 0.219 65.0
Hakamada ve ark, (1997), alkalofilik Bacillus sp. KSM-S237’den izole edilen
ve optimum aktivite sıcaklığı 50ºC olan endoglukanaz enziminin 10 dakikalık
inkübasyon sonunda orijinal aktivitesini 20ºC’de %100 koruduğunu, 40ºC’de
aktivitenin %90’a, 50ºC’de %50’ye 100ºC’de ise %30’a düştüğü belirtmiştir.
Endo ve ark, (2001) optimum aktivite sıcaklığı 55ºC olan alkalin
endoglukanaz enziminin, orijinal aktivitesini 40ºC ve 15 dakika inkübasyon sonunda
%100’e yakın koruduğunu, ortam sıcaklığı 60ºC’ye yükseltildiğinde ise aktivitenin
yaklaşık %50 düzeyine düştüğünü belirtmişlerdir.
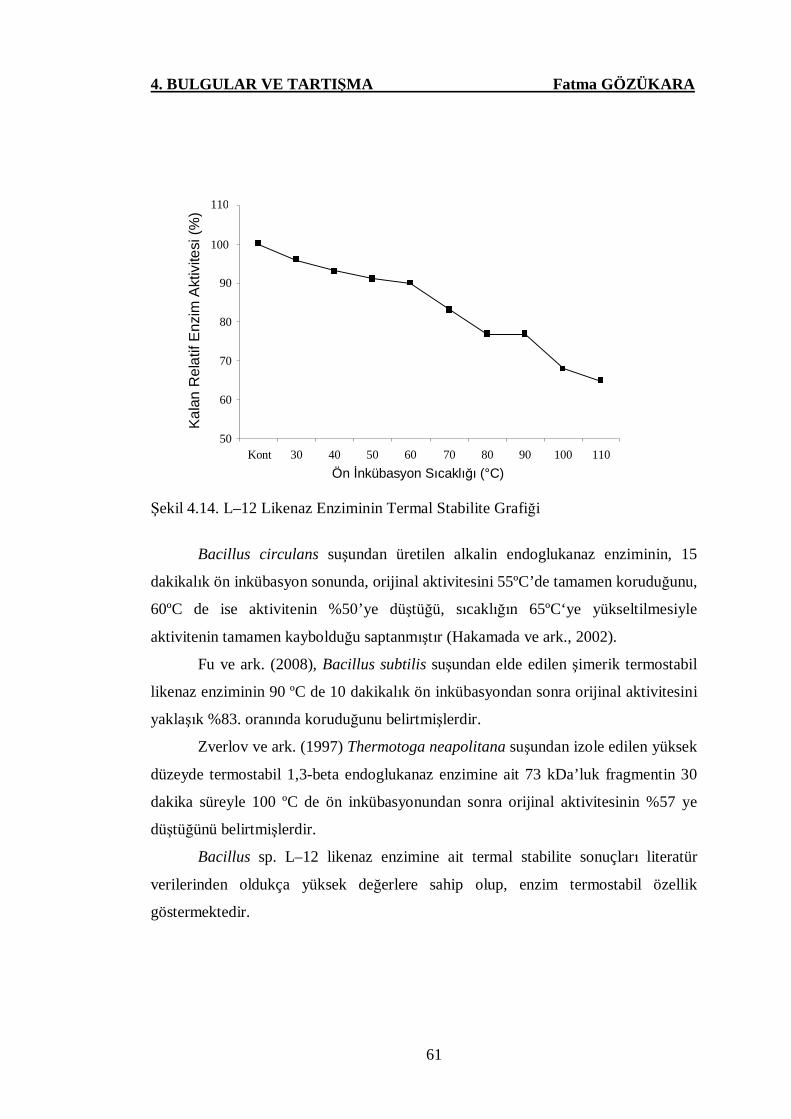
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
61
Şekil 4.14. L–12 Likenaz Enziminin Termal Stabilite Grafiği
Bacillus circulans suşundan üretilen alkalin endoglukanaz enziminin, 15
dakikalık ön inkübasyon sonunda, orijinal aktivitesini 55ºC’de tamamen koruduğunu,
60ºC de ise aktivitenin %50’ye düştüğü, sıcaklığın 65ºC‘ye yükseltilmesiyle
aktivitenin tamamen kaybolduğu saptanmıştır (Hakamada ve ark., 2002).
Fu ve ark. (2008), Bacillus subtilis suşundan elde edilen şimerik termostabil
likenaz enziminin 90 ºC de 10 dakikalık ön inkübasyondan sonra orijinal aktivitesini
yaklaşık %83. oranında koruduğunu belirtmişlerdir.
Zverlov ve ark. (1997) Thermotoga neapolitana suşundan izole edilen yüksek
düzeyde termostabil 1,3-beta endoglukanaz enzimine ait 73 kDa’luk fragmentin 30
dakika süreyle 100 ºC de ön inkübasyonundan sonra orijinal aktivitesinin %57 ye
düştüğünü belirtmişlerdir.
Bacillus sp. L–12 likenaz enzimine ait termal stabilite sonuçları literatür
verilerinden oldukça yüksek değerlere sahip olup, enzim termostabil özellik
göstermektedir.
50
60
70
80
90
100
110
Kont 30 40 50 60 70 80 90 100 110Ön İnkübasyon Sıcaklığı (°C)
Kal
an R
elat
if E
nzim
Akt
ivite
si (%
)
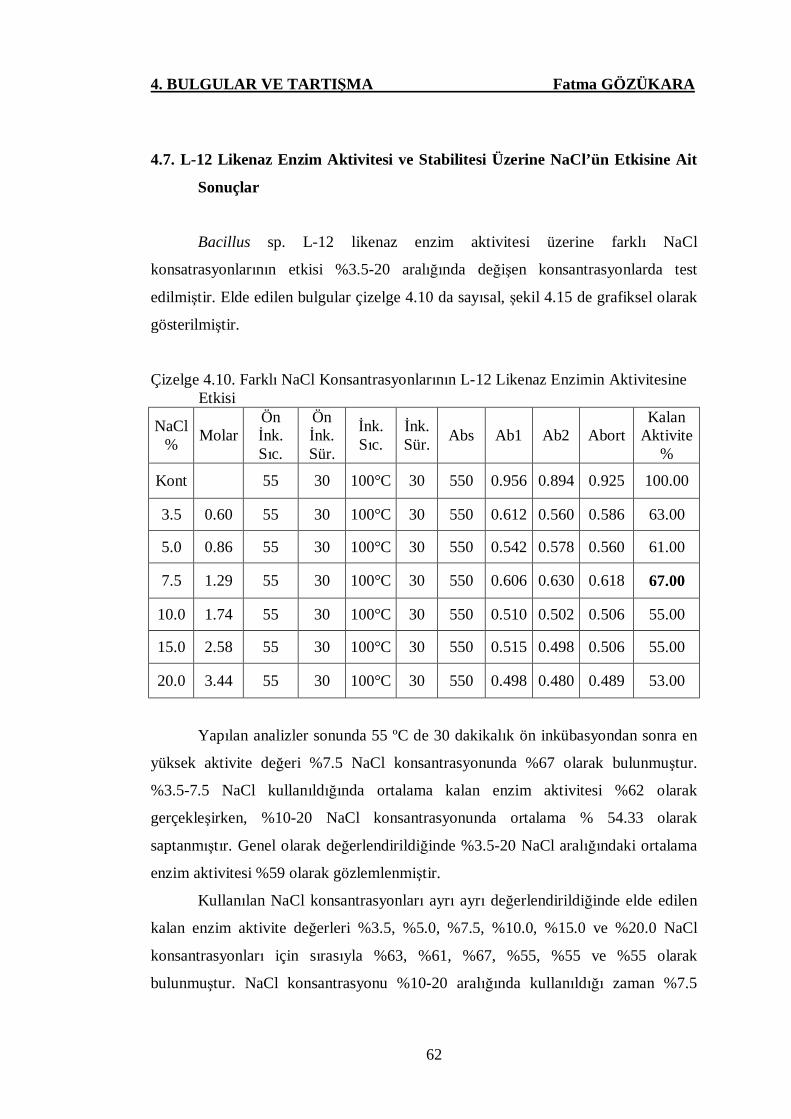
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
62
4.7. L-12 Likenaz Enzim Aktivitesi ve Stabilitesi Üzerine NaCl’ün Etkisine Ait
Sonuçlar
Bacillus sp. L-12 likenaz enzim aktivitesi üzerine farklı NaCl
konsatrasyonlarının etkisi %3.5-20 aralığında değişen konsantrasyonlarda test
edilmiştir. Elde edilen bulgular çizelge 4.10 da sayısal, şekil 4.15 de grafiksel olarak
gösterilmiştir.
Çizelge 4.10. Farklı NaCl Konsantrasyonlarının L-12 Likenaz Enzimin Aktivitesine Etkisi
NaCl % Molar
Ön İnk. Sıc.
Ön İnk. Sür.
İnk. Sıc.
İnk. Sür. Abs Ab1 Ab2 Abort
Kalan Aktivite
%
Kont 55 30 100°C 30 550 0.956 0.894 0.925 100.00
3.5 0.60 55 30 100°C 30 550 0.612 0.560 0.586 63.00
5.0 0.86 55 30 100°C 30 550 0.542 0.578 0.560 61.00
7.5 1.29 55 30 100°C 30 550 0.606 0.630 0.618 67.00
10.0 1.74 55 30 100°C 30 550 0.510 0.502 0.506 55.00
15.0 2.58 55 30 100°C 30 550 0.515 0.498 0.506 55.00
20.0 3.44 55 30 100°C 30 550 0.498 0.480 0.489 53.00
Yapılan analizler sonunda 55 ºC de 30 dakikalık ön inkübasyondan sonra en
yüksek aktivite değeri %7.5 NaCl konsantrasyonunda %67 olarak bulunmuştur.
%3.5-7.5 NaCl kullanıldığında ortalama kalan enzim aktivitesi %62 olarak
gerçekleşirken, %10-20 NaCl konsantrasyonunda ortalama % 54.33 olarak
saptanmıştır. Genel olarak değerlendirildiğinde %3.5-20 NaCl aralığındaki ortalama
enzim aktivitesi %59 olarak gözlemlenmiştir.
Kullanılan NaCl konsantrasyonları ayrı ayrı değerlendirildiğinde elde edilen
kalan enzim aktivite değerleri %3.5, %5.0, %7.5, %10.0, %15.0 ve %20.0 NaCl
konsantrasyonları için sırasıyla %63, %61, %67, %55, %55 ve %55 olarak
bulunmuştur. NaCl konsantrasyonu %10-20 aralığında kullanıldığı zaman %7.5
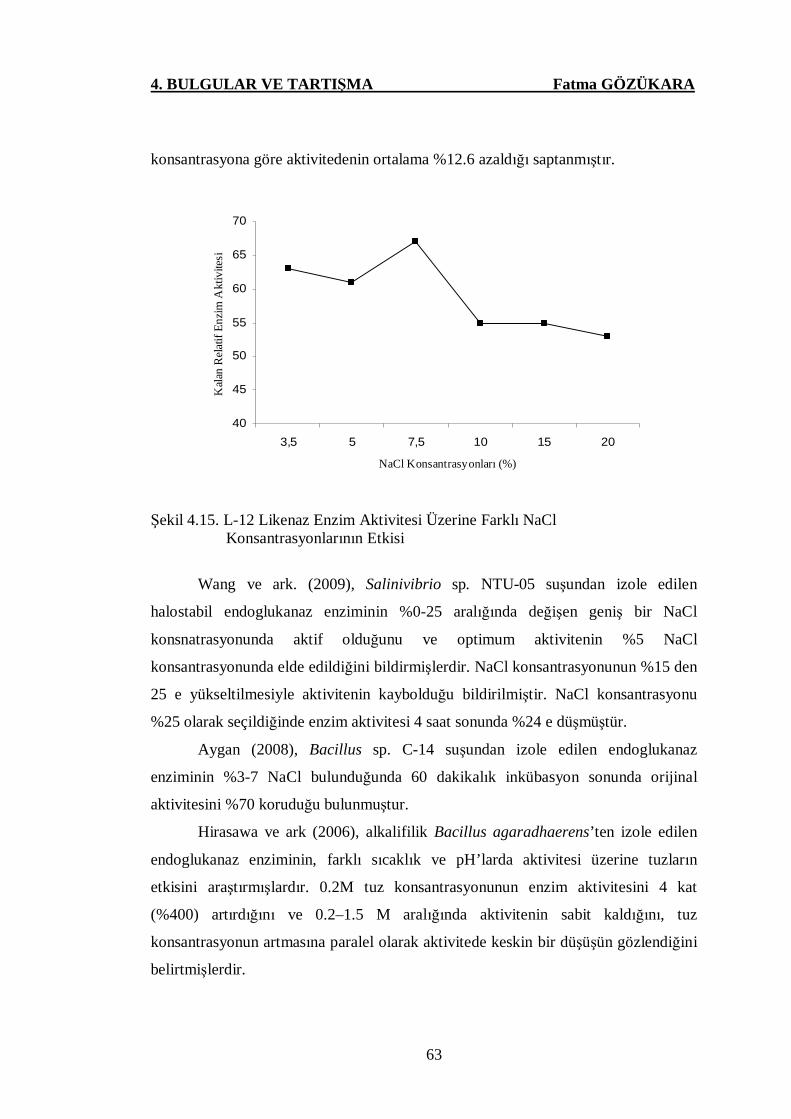
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
63
konsantrasyona göre aktivitedenin ortalama %12.6 azaldığı saptanmıştır.
40
45
50
55
60
65
70
3,5 5 7,5 10 15 20
NaCl Konsantrasyonları (%)
Kal
an R
elat
if En
zim
Akt
ivite
si
Şekil 4.15. L-12 Likenaz Enzim Aktivitesi Üzerine Farklı NaCl
Konsantrasyonlarının Etkisi
Wang ve ark. (2009), Salinivibrio sp. NTU-05 suşundan izole edilen
halostabil endoglukanaz enziminin %0-25 aralığında değişen geniş bir NaCl
konsnatrasyonunda aktif olduğunu ve optimum aktivitenin %5 NaCl
konsantrasyonunda elde edildiğini bildirmişlerdir. NaCl konsantrasyonunun %15 den
25 e yükseltilmesiyle aktivitenin kaybolduğu bildirilmiştir. NaCl konsantrasyonu
%25 olarak seçildiğinde enzim aktivitesi 4 saat sonunda %24 e düşmüştür.
Aygan (2008), Bacillus sp. C-14 suşundan izole edilen endoglukanaz
enziminin %3-7 NaCl bulunduğunda 60 dakikalık inkübasyon sonunda orijinal
aktivitesini %70 koruduğu bulunmuştur.
Hirasawa ve ark (2006), alkalifilik Bacillus agaradhaerens’ten izole edilen
endoglukanaz enziminin, farklı sıcaklık ve pH’larda aktivitesi üzerine tuzların
etkisini araştırmışlardır. 0.2M tuz konsantrasyonunun enzim aktivitesini 4 kat
(%400) artırdığını ve 0.2–1.5 M aralığında aktivitenin sabit kaldığını, tuz
konsantrasyonun artmasına paralel olarak aktivitede keskin bir düşüşün gözlendiğini
belirtmişlerdir.
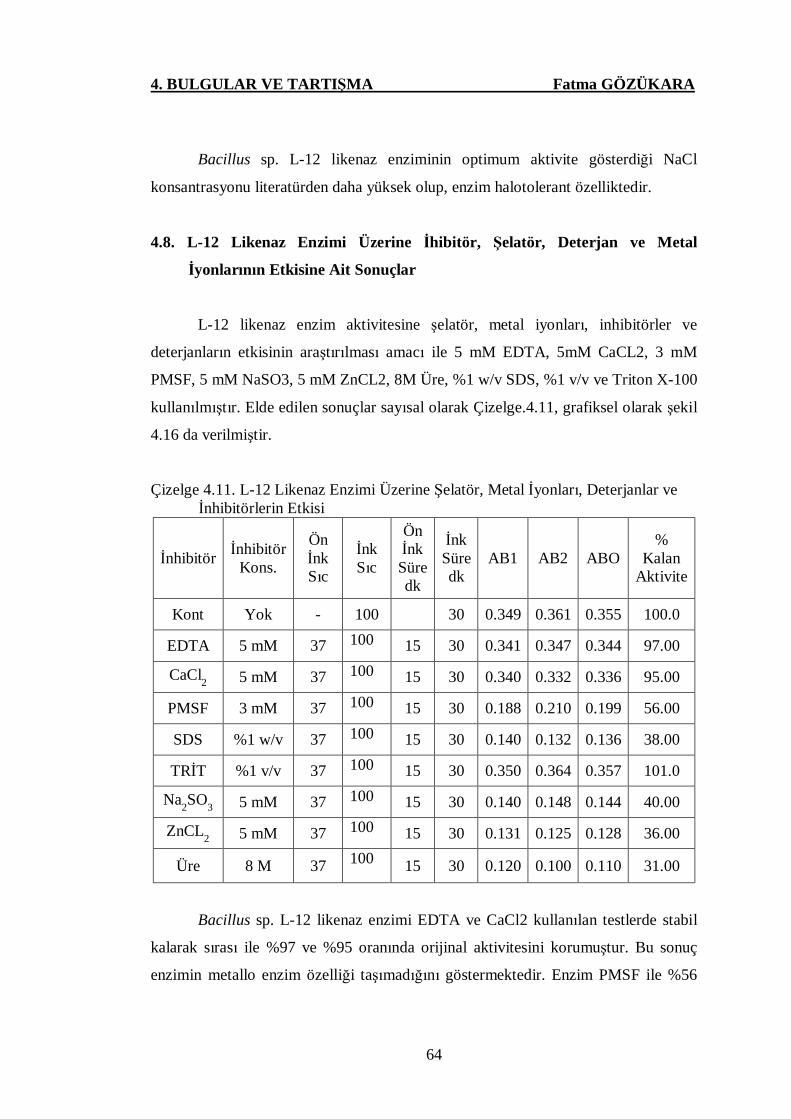
4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
64
Bacillus sp. L-12 likenaz enziminin optimum aktivite gösterdiği NaCl
konsantrasyonu literatürden daha yüksek olup, enzim halotolerant özelliktedir.
4.8. L-12 Likenaz Enzimi Üzerine İhibitör, Şelatör, Deterjan ve Metal
İyonlarının Etkisine Ait Sonuçlar
L-12 likenaz enzim aktivitesine şelatör, metal iyonları, inhibitörler ve
deterjanların etkisinin araştırılması amacı ile 5 mM EDTA, 5mM CaCL2, 3 mM
PMSF, 5 mM NaSO3, 5 mM ZnCL2, 8M Üre, %1 w/v SDS, %1 v/v ve Triton X-100
kullanılmıştır. Elde edilen sonuçlar sayısal olarak Çizelge.4.11, grafiksel olarak şekil
4.16 da verilmiştir.
Çizelge 4.11. L-12 Likenaz Enzimi Üzerine Şelatör, Metal İyonları, Deterjanlar ve İnhibitörlerin Etkisi
İnhibitör İnhibitör Kons.
Ön İnk Sıc
İnk Sıc
Ön İnk Süre dk
İnk Süre dk
AB1 AB2 ABO %
Kalan Aktivite
Kont Yok - 100 30 0.349 0.361 0.355 100.0
EDTA 5 mM 37 100 15 30 0.341 0.347 0.344 97.00
CaCl2 5 mM 37 100 15 30 0.340 0.332 0.336 95.00
PMSF 3 mM 37 100 15 30 0.188 0.210 0.199 56.00
SDS %1 w/v 37 100 15 30 0.140 0.132 0.136 38.00
TRİT %1 v/v 37 100 15 30 0.350 0.364 0.357 101.0
Na2SO3 5 mM 37 100 15 30 0.140 0.148 0.144 40.00
ZnCL2 5 mM 37 100 15 30 0.131 0.125 0.128 36.00
Üre 8 M 37 100 15 30 0.120 0.100 0.110 31.00
Bacillus sp. L-12 likenaz enzimi EDTA ve CaCl2 kullanılan testlerde stabil
kalarak sırası ile %97 ve %95 oranında orijinal aktivitesini korumuştur. Bu sonuç
enzimin metallo enzim özelliği taşımadığını göstermektedir. Enzim PMSF ile %56

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
65
oranında inhibe olmuştur. L-12 likenaz enzimi SDS ile %62 oranında aktivitesini
kaybederken non iyonik deterjan olan triton-X 100 ile %101 oranında aktivite
göstermiştir. Bacillus sp. L-12 likenaz enzimi Na2SO3, ZnCL2 Üre kullanılan
çalışmalarda orijinal aktivitesini sırası ile % 60, %64 ve %69 oranında kaybetmiştir.
Literatür bulgularında EDTA ile gerçekleştirilen aktivite çalışmalarında kalan
enzim aktivitesinin %90-95 aralığında gerçekleştiği belirtilmiştir (Singh ve ark,
2001; Huang ve Monk, 2004; Li ve ark, 2006).
Huang ve Monk, (2004), Termofilik aerobik Caldibacillus cellulovorans
bakterisinden izole edilen CMCase enzimine ait orijinal aktivitesinin Zn ile önemli
derecede inhibe edildiğini belirtmişlerdir. L-12 likenaz enziminden elde edilen sonuç
literatürle uyum içindedir. Jung ve ark. (2007), Bacillus sp. A8-8 bakterisinden izole
edilen termostabil β-1,3-1,4 glukanaz enziminin 5 mM EDTA ile aktivitesini
koruduğunu ve etkilenmediğini, buna karşılık Ca ile aktivitenin arttığını
belirtmişlerdir. Aynı araştırma ekibi birçok β-1,3-1,4 glukanaz enziminin Ca ve Co
iyonları ile aktivitelerinin arttığını buna karşılık EDTA, Zn ve Fe ve Mn ile
aktivitenin inhibe edildiğini belirtmişlerdir.
Ng Son ve ark. (2009), termofilik Geobacillus sp. 70PC53 bakterisinden izole
edilen ve geniş bir sıcaklık aralığında stabilite gösteren CelA endoglukanaz
enziminin konsantrasyona bağlı olarak Zn ve Cu iyonları ile güçlü şekilde inhibe
edildiğini, buna karşılık Ca ile azda olsa sitümüle edildiğini belirtmişlerdir. Aynı
araştırma grubu CelA endoglukanaz enzim aktivitesinin Na, Mg ve EDTA ile
etkilenmediğini belirtmişlerdir. Ng Son ve ark. (2009), CelA endoglukanaz enzim
aktivitesinin 2-merkaptoethanol (β-mercaptoethanol) ile dramatik şekilde arttığını
belirtmişlerdir.
Li ve ark. (2006) Bacillus sp. AC.1 bakterisinden izole edilen termoasidofilik
endoglukanaz enziminin %0.5 SDS ile %66 oranında inhibe olurken, %0.25 triton X-
100 ile %127 aktive edildiğini belirtmişlerdir. Aynı enzimin 5 mM EDTA ile orijinal
aktivitesini %95 koruduğu belirlenmiştir. İzole edilen Bacillus sp. L-12 likenaz
enziminin Li ve ark (2006) nın bulguları ile çok büyük benzerlik gösterdiği
görülmektedir.
Akita ve ark. (2005) Bacillus halodurans C-125 bakterisinden izole edilen

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
66
beta glukanaz enziminin 1 mM Zn ile inhibi olduğunu buna karşılık 10 mM CaCl2ve
EDTA’dan etkilenmediğini bildirmişlerdir.
Bacillus sp. L-12 likenaz enziminden elde edilen sonuçlar literatürde
belirtilen termostable endoglukanaz enzimlerine ait sonuçlarla çok büyük uyum
göstermektedir.
Şekil 4.16. L-12 Likenaz Enzimine Metal İyonu, Şelatör, İnhibitör ve Deterjanların
Etkisi
4.9. L-12 Likenaz Enziminin Native-PAGE ve Zimogram Analizi
Analiz edilen L-12 likenaz enzimi %10’luk Native-PAGE’de (%1’lik likenan
içeren) yürütüldükten sonra jel iki parçaya ayrılarak, marker bölgesi Comassie
Brillant Blue R-250 ile boyanırken, diğer parça renatürasyon işlemlerinden sonra
aktivitenin saptanması için kongo kırmızısı ile boyanarak analiz edilmiştir. Elde
edilen sonuçlar Şekil 4.17 de gösterilmiştir.
0
20
40
60
80
100
120
Kont EDTA CaCl2 PMSF SDS Trit-X100 Na-Sülfit ZnCl
2 Üre
İnhibitörler
Kal
an R
ölat
if E
nzim
Akt
ivite
si (%
)

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
67
Marker Enzim Şekil 4.17. L-12 Likenaz Enziminin Molekül Büyüklüğü ve Zimogram Analizi
(Marker olarak Sığır serum albumin kullanılmıştır, 68.000 Da)
Likenaz enzimi zymogram analizi sonucunda moleküler ağırlıkları 54.4 ve
100.74 kDa olan bir birinden bağımsız iki aktivite bandı elde edilmiştir.
Literatürde endoglukanaz enzimlerinin büyük çoğunluğunda moleküler
ağırlıkların 35 kDa 85 kDa arasında değiştiği belirtilmiştir (Han ve ark.,1995;
Hakamada ve ark., 1997; Mawadza ve ark.,2000; Hakamada ve ark., 2002; Singh ve
ark., 2001; Endo ve ark.,2001; Kim ve ark., 2005; Voget ve ark., 2006; Zvereva ve
ark, 2006; Zhang ve ark, 2006; Aygan, 2008).
Li ve ark., (2006), Bacillus sp. AC.1 bakterisinden izole edilen termo-
asidofilik endoglukanaz (Ba-EGA) enziminin SDS-PAGE analizi sonucunda 67 kDa
moleküler ağırlıkta tek bir bandan meydana geldiğini saptanmıştır.
Eckert ve Schneider (2003) Alycyclobacillus acidocaldarius bakterisinden
izole edilen termoasidofilik CelB endoglukanaz enziminin PAGE gel analizi
sonucunda moleküler ağırlıkları 30-100 kDa arasında değişen 5 farklı alt birimden
oluştuğunu bulmuşlardır.
Ri ve ark. (2009) Laetiporus sulphureus var. miniatus organizmasından izole
edilen termostabil β-1,3-1,4 glukanaz enziminin SDS-PAGE elektroforez analizi
sonucunda 52 kDa ağırlığa sahip tek bir bandan meydana geldiğini bulmuşlardır.
100.74
68 kDa
54.4 kDa

4. BULGULAR VE TARTIŞMA Fatma GÖZÜKARA
68
Zverlov ve ark. (1997), Thermotoga naepolitana organizmasından izole
edilen yüksek düzeyde termostabil endo-1,3- β-glukanaz enziminin 52 ve 73 kDa
ağırlığa sahip bağımsız iki alt birimden oluştuğunu belirtmişlerdir.
Huang ve Monk (2004), Termofilik aerobik Caldibacillus cellulovorans
bakterisinden izole edilen CMCase enziminin SDS-PAGE analizi sonunda moleküler
ağırlığı 85.1 kDa olan tek bir bandan meydana geldiğini belirtmişlerdir.
Bacillus sp. L-12 likenaz enziminin native-PAGE jelinde yapılan analiz
sonunda iki farklı badın saptanması ve moleküler ağırlıklar, termoasidofilik
endoglukanaz enzimlerinin özellikleriyle uygunluk göstermektedir.

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
69
5. SONUÇLAR VE ÖNERİLER
Enzimler, doğal olarak canlılar tarafından sentezlenen protein yapısında ya da
bir kısmı protein olan biyo-moleküllerdir. Enzimler, binlerce yıldır içecek, ekmek ve
peynir yapımı gibi işlemlerde varlığı ve görevi bilinmeden kullanılmıştır. İlk bulgular
eski Mısıra kadar dayanmaktadır. Yakın tarihte ise Doğu ülkelerinde birçok gıda
fermantasyonu için ipliksi mantarlar enzim kaynağı olarak kullanılmaktadır. Batıda
1896’da gerçek modern mikrobiyal enzim teknolojisi ‘takadiastase’ın ticareti ile
başlamıştır. Bu Doğudan Batı toplumuna önemli bir teknolojik transferdir (Uhlig,
1998).
Enzim teknolojisinin giderek gelişmesi, ürünlerin kullanım alanlarının
çeşitliliği ve ekonomik değerinin çok yüksek olması nedeniyle, biyoteknolojinin
endüstriyel enzimlerle ilgili alanında yapılan çeşitli araştırmalar, daha da önem
kazanmaktadır. Özellikle birkaç ülke dışında diğer ülkelerin bu konuda tamamen dışa
bağımlı durumdadırlar.
Dünyada, 1980-1983 yılları arasında sadece 300 küçük biyoteknoloji şirketi
çalışma gerçekleştirirken, bu sayı 1985 yılında sadece Amerika Birleşik
Devletlerinde (A.B.D) 400 düzeyine ulaşmıştır. Günümüzde A.B.D de 900, bütün
dünyada ise yaklaşık 1200 biyoteknoloji şirketi çalışmalarını çeşitli alanlarda
sürdürmektedirler. Örneğin sadece farmasötik alanında 2000’li yıllarda kullanılacak
biyoteknolojik ürünlerin toplam değerinin yaklaşık 60 milyar dolar/yıl düzeyinde
olacağı düşünülmektedir.
Biyoteknoloji kaynaklı çalışmalar A.B.D. de odaklanmış olmakla birlikte,
günümüzde, Japonya ve Kanada, biyoteknolojiyi (özellikle moleküler
biyoteknolojiyi) stratejik alan kategorisinde değerlendirerek, özel şirketlerin yanı
sıra, hükümetler düzeyinde destekleme ve geliştirme kararı almışlardır (Glick ve
Pasternak, 1994).
Tarihsel gelişim açısından bakıldığında enzimlerin çok farklı kaynaklardan
elde edildiği görülmektedir. Bunlar bitkisel, hayvansal ya da endüstriyel anlamda
ihtiyacı karşılayabilen mikrobiyal kaynaklı enzimlerdir. Bugüne kadar yaklaşık 2500
farklı enzim tanımlanmış ve bunların ancak %10’u ticari alanda kullanım için

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
70
kendilerine yer bulmuşlardır. Bunlardan ise ancak 25 tanesi nişasta sanayi ile
deterjan katkı maddesi olarak kullanılmış olup, ticari alanda yararlanılan bütün
enzimlerin %80 ini oluşturmaktadırlar (Rao ve ark. 1998). Bitkisel ve hayvansal
enzimlerin endüstriyel ihtiyacı karşılayamaması, bu alandaki ilginin giderek artan bir
şekilde mikrobiyal enzimlere yönelmesini sağlamıştır. Mikroorganizmalar,
biyokimyasal çeşitlilikleri ve genetik manipülasyonlara uygunluğu gibi sebeplerden
dolayı mükemmel bir enzim kaynağı olarak değerlendirilmektedir (Rao ve ark,
1998).
Günümüzde endüstride kullanılan enzimlerin yaklaşık %90’ı
mikroorganizmalardan üretilmektedir (Wolfgang, 2004). Bunun nedeni,
mikroorganizma kaynaklı enzimlerin katalitik aktivitelerinin yüksek olması,
istenmeyen yan ürün oluşturmamaları, daha stabil ve daha ucuz olmaları, büyük
boyutlarda ve yüksek saflıkta elde edilmesi gibi avantajlara sahip olmasıdır. Örneğin
mikrobiyal enzimler ekstrem sıcaklık ve pH değerlerinde çok yüksek düzeyde
aktivite gösterirler (Wiseman, 1987; Horikoshi, 1999).
Bir enzimin bir endüstri alanında kullanılabilmesi maliyet bakımından ucuz
olması, çok farklı alanlarda kullanılabilme özelliğinde olması ve en önemlisi de
enzimin alerjik ya da toksik etkiye sahip olmaması, yani güvenilir olmasına bağlıdır
(Wiseman, 1987; Sarıkaya, 1995).
Mikrobiyal enzimlerin dünya genelinde yıllık kullanım değerlerine bakıldığı
zaman, alkaline proteaz %25, diğer proteazlar %21, Amilaz %18, Renin %10,
Trypsin %3, Lipaz %3, diğer karbonhidrat parçalayan enzimler (selülaz ve xylanaz
gibi) %10, analitik ve farmasötik enzimler %10 şeklinde bir dağılımla
karşılaşılmaktadır (Rao ve ark., 1998).
Hidrolazlardan selülazlar ise selülozik materyalleri şekerlere dönüştürerek
özellikle biyo-etanol gibi biyolojik temelli yakıt katkı maddelerinin üretiminde
önemli bir yer tutmaya başlamışlardır. Bu nedenle selülaz’ların (glukanaz) enzim
piyasasındaki paylarının ciddi bir şekilde artması beklenmektedir. Sadece bitkisel
atıkların hidrolizinde kullanılması durumunda bile, selülaz enzimlerinin Amerika
Birleşik Devletleri’nde yıllık 400 milyon dolarlık bir ticaret hacmi yaratacağı
öngörülmektedir (Boyce ve Walsh, 2007).

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
71
Dünya geneli incelendiğinde endüstriyel enzimlerin ticari pazar payının
yaklaşık 1.6 milyar dolar olduğu tahmin edilmektedir. Bu enzimlerin kullanım
alanlarına göre dağılımına bakıldığında ise, %29’unun gıda endüstrisi, %15’inin
hayvan yemi sektörü ve %56’sının genel amaçlı teknik alanlarda şeklinde dağıldığı
görülmektedir (Uhlig, 1998).
Endüstride yaygın şekilde kullanılan enzimler arasında Bacillus cinsine ait
tür ve alttürleri tarafından sentezlenen ondan fazla enzim vardır. Örneğin ticari
olarak üretilen ve kullanılan termostabil amilaz enzimlerinin üretilmelerinde Bacillus
amyloliquefaciens ve Bacillus licheniformis en çok kullanılan iki Bacillus türüdür
(Horikoshi, 1996; Lin ve ark., 1998; Horikoshi, 1999; Kumar ve Takagi, 1999).
Bacillus cinsi bakteriler toprak, hayvan dışkıları ve bitkisel atıklar üzerinde
yaygın olarak bulunurlar. Bu cinsin bireylerinin çoğu zararsız, izolasyonu ve teşhisi
kolay, hızlı büyüme oranı ile fermentasyon süresi kısadır. Genel olarak güvenli
olmaları, sentezledikleri proteinleri dış ortama salgılama kapasiteleri gibi birçok
nedenden dolayı endüstriyel organizmalardır.
Bacillus türleri çeşitli kompleks substratlara karşı aktivite gösteren çok sayıda
ve çeşitli hidrolitik enzimler üretmekte ve salgılamaktadırlar. Bu nedenle Bacillus
cinsindeki organizmalar, endüstriyel alanda α-amilaz, proteaz, glukanaz, glukoz
izomeraz ve endonükleaz gibi enzimlerin üretiminde yaygın şekilde
kullanılmaktadırlar (Uhlig, 1998). Bu enzimler arasında endo β-(1,3)-(1,4)-
glukanazlar (likenazlar), endosperm hücrelerinin nişastasına α-amilazların ulaşımını
kolaylaştırdığından dolayı içki mayalama endüstrisinde spesifik uygulamalara
sahiptirler (Beckmann ve ark., 2006).
Çalışmalar sırasında tanımlanmaları yapılarak morfolojik, boyanma davranışı
ve biyokimyasal testler sonucunda Bacillus sp. şeklinde adlandırılan toplam 68
bakteri suşu öncelikli olarak üreme ve enzim sentezleme özelliklerinin saptanması
amacı ile 20-60°C ortam sıcaklığı ve 5.0-10.0 aralığındaki pH değerlerinde analiz
edilmişlerdir. Analizler içerisinde likenan bulunan katı besiyerinde
gerçekleştirilmiştir.
Çalışmamızda izole edilen suşlar arasından 20 adet Bacillus sp. suşu 20, 30,
40, 50 ve 60°C sıcaklık ve pH 5.0, 6.0, 7.0, 8.0, 9.0 ve 10.0 da analiz edilerek katı

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
72
besiyerindeki üreme ve enzim sentezleme davranışları ayrıntılı olarak analiz
edilmiştir (Şekil 4.1., Şekil 4.2., Şekil 4.3., Şekil 4.4., Şekil 4.5., Şekil 4.6., Şekil
4.7., Şekil 4.8., Şekil 4.9., Şekil 4.10.).
Sıcaklık ve pH değerleri farklılaştıkça suşların üreme ve enzim sentezleme
davranışlarında da farklılıklar saptanmıştır.
İnkübasyon sıcaklığı 20°C seçildiğinde 2, 7, 14 ve 18 numaralı suşların bütün
pH değerlerinde üremedikleri saptanmıştır. Bir numaralı suş bütün pH değerlerinde
belirgin şekilde üremediği halde pH 7.0, 8.0, 9.0 ve 10.0 da ortalama 2 mm
genişliğinde aktivite zonu oluşturmuştur. Üç numaralı suş bütün pH değerlerinde
belirgin üreme göstermemiş fakat pH 5.0 ve 6.0 da ortalama 3 mm genişliğinde
aktivite zonu saptanmıştır. On üç numaralı suş bütün pH değerlerinde ortalama 2.6
mm çaplı koloni oluşturarak ürerken, ortalama 10.6 mm genişliğinde aktivite zonu
elde edilmiştir. On iki numaralı suş pH 5.0, 6.0, 7.0 ve 8.0 de üremediği halde bu pH
değerlerinde ortalama 5 mm genişliğinde aktivite zonu saptanmıştır. Beş numaralı
suş sadece pH 5.0 te üremiş fakat bütün pH değerlerinde (ortalama 4 mm) enzim
aktivitesi saptanmıştır. Bu inkübasyon sıcaklığında on üç numaralı suş bütün pH
değerlerinde üremiş ve ortalama 10.6 mm genişliğinde aktivite zonu saptanmıştır
(Çizelge 4.1.).
İnkübasyon sıcaklığı 30°C ye yükseltildiğinde bütün suşların analiz edilen
bütün pH değerlerinde üreme ve enzim sentezi gerçekleştirdiği saptanmıştır.
İnkübasyon sıcaklığının yükseltilmesine paralel olarak koloni çapı ve aktivite zon
genişliğinde belirgin artış görülmüştür. Bu inkübasyon sıcaklığında en geniş aktivite
zonu (bütün pH değerleri için ortalama 20 mm) 17 numaralı suşta elde edilmiştir. On
beş numaralı suşta pH 5.0, 6.0 ve 7.0 de koloni çapı ortalama 2.5 mm, aktivite zon
genişliği 12 mm olarak ölçülürken, pH 8.0, 9.0 ve 10.0 da koloni çapı ortalama 4 mm
zon genişliği 20 mm bulunmuştur. On iki numaralı suşta ortalama koloni çapı 3.5,
aktivite zon genişliği 14.3 mm olarak saptanmıştır (Çizelge 4.2.).
Ortam sıcaklığı 40°C ye yükseltildiğinde özellikle 5 ve 12 numaralı suşların
koloni ve aktivite zon çapı genişlikleri diğer örneklerden daha yüksek değerlere
ulaşmıştır. Beş numaralı suşta ortalama koloni çapı 9 mm, aktivite zon genişliği 23.8
mm olmuştur.

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
73
On iki numaralı suşta ise koloni çapı 8.8 mm, aktivite zon genişliği 27.6 mm
ölçülmüştür. Özellikle pH 5.0 ve 6.0 da oralama 32 mm aktivite zon genişliği
saptanırken, pH değerleri yükseldikçe aktivite zon genişliği giderek düşmüş ve
ortalama 25.5 mm olarak gerçekleşmiştir (Çizelge 4.3.).
Ortam sıcaklığı 50°C ye yükseltildiğinde koloni çapı ve aktivite zon
genişliğinin diğer suşlardan daha yüksek değerlere ulaştığı saptanmıştır. Altı
numaralı suşta ortalama koloni çapı 13.3 mm, aktivite zon genişliği ise 22 mm olarak
bulunmuştur. Bu suşta en geniş zon 28 mm ile pH 6.0 da elde edilmiştir.
On iki numaralı suşta ortalama koloni çapı 18.5 mm aktivite zon genişliği
32.16 mm’ye ulaşmıştır. On iki numaralı suşta en geniş zon 36 mm ile pH 7.0 de elde
edilirken, sırası ile pH 8.0 de 35, 9.0 da 35 ve 10.0 da 31 mm olarak bulunmuştur
(Şekil 4.7., Şekil 4.8.). Bu sıcaklık değerinde özellikle pH 5.0 te 7 numaralı suşun 17,
sekiz numaralı suşun ise 15 mm koloni çapı ile en iyi üreme gerçekleşmiştir
(Çizelge 4.4.).
Ortam sıcaklığı 60°C ye yükseltildiğinde 15 numaralı suşun pH 5.0, 6.0 ve
7.0 de ürediği halde enzim sentezinin gerçekleşmediği saptanmıştır. On yedi
numaralı suşta pH 5.0 te üreme olmadığı halde 5 mm’lik aktivite zonu elde
edilmiştir. Buna karşılık aynı suş pH 10.0 da ürediği halde enzim sentezi
gerçekleşmemiştir. On dokuz numaralı suş bütün pH değerlerinde üreme
göstermemiş fakat pH 5.0 te 5 mm, 6.0 da 16 mm, 8.0 ve 9.0 da 12 mm aktivite zonu
elde edilmiştir. pH 10.0 da ise üreme ve enzim sentezi gerçekleşmemiştir.
Yirmi numaralı suşta sadece pH 9.0 da üreme gerçekleşmiş (5 mm koloni
çapı) ve 26 mm aktivite zon genişliği saptanmıştır. pH 8.0 de ise üreme olmadığı
halde 17 mm aktivite zonu görülmüştür. Diğer pH değerlerinde üreme ve enzim
sentezi meydana gelmemiştir.
Bu inkübasyon sıcaklığında iki numaralı suş bütün pH değerlerinde ortalama
16 mm koloni çapı oluşturacak şekilde üremiştir. Özellikle pH 8.0 de 34 mm’lik
koloni çapı saptanmıştır. Bu suşun aktivite zon çapı ise ortalama 20 mm olarak
bulunmuştur.
On iki numaralı suş, bütün pH değerlerinde ortalama 8 mm koloni çapı
oluşturacak şekilde üreme göstermiştir. pH 9.0 da 21 mm’lik koloni çapı elde

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
74
edilirken, pH 8.0 ve 10.0 da 6 mm çaplı üremenin gerçekleşmesi bu suş için pH 9.0
un ekstrem bir değer olduğunu düşündürmektedir. Bununla birlikte aktivite zon
genişliğinin 25 mm olması, bu pH’ın enzim sentezi açısından ekstrem olmadığını
göstermektedir (örneğin pH 5.0 te 5 mm koloni, 21 mm, pH 10.0 da 6 mm koloni 26
mm aktivite zonu ölçülmüştür). On iki numaralı suşun bütün pH değerlerinde yüksek
bir aktivite zonuna sahip olduğu ve değerlerin birbirine yakın olduğu bulunmuştur
(pH 5.0 te 21 mm, pH 10.0 da 26 mm aktivite zonu). Bütün pH değerleri için
ortalama 23 mm aktivite zonu elde edilmiştir (Çizelge 4.5.).
Aktivite zonunun koloni çapına oranlaması (ortalama zon genişliği/ortalama
koloni çapı) yapıldığında on iki numaralı suşun aktivite zon genişliği koloni çapının
yaklaşık 3 katı değerindedir.
L-12 likenaz enziminin optimum aktivite gösterdiği pH değeri 6.0 olarak
ölçülmüştür (Şekil 4.11.). Enzim pH 4.0-8.0 aralığında ortalama % 90 relatif
aktiviteye sahiptir. Bütün pH değerleri dikkate alındığında ortalama relatif aktivite
%87 düzeyinde gerçekleşmiştir. Likenaz enzimi bu özellikleri ile çok geniş bir pH
aralığında aktive gösterme özelliğine sahiptir. Bununla birlikte optimum pH
değerinin 6.0 olması likenaz enziminin asidik, aynı zamanda alkali bölgede yüksek
aktiviteye sahip olduğunu göstermektedir (Çizelge 4.6.).
Jung ve ark. (2007) Bacillus sp. A8-8 bakterisinden izole edilen termostabil
likenaz enziminin optimum aktivite gösterdiği pH değerinin 6.0 olduğunu
belirtmişlerdir. Huang ve Monk (2004), termofilik aerobik bakteri Caldibacillus
cellulovarans izole edilen CMCase enziminin optimum aktivite gösterdiği pH
aralığının 6.5-7.0 olduğunu belirtmişlerdir. Zhu ve ark. (2008), Stichopus japonicus
organizmasından izole edilen beta-1,3 glukanaz enziminin optimum aktivitesini pH
5.5 te gösterdiğini belirtmişlerdir. Zverlov ve ark (1997), yüksek düzeyde termostabil
1,3-beta endoglukanaz enziminin optimum relatif aktiviteyi pH 6.2 de gösterdiğini
bulmuşlardır. Li ve ark. (2006), Bacillus sp. AC-1 bakterisinden izole edilen
termoasidofilik endoglukanaz enziminin optimum aktiviteyi pH 4.5-6.5 aralığında
gösterdiğini belirtmişlerdir. Fu ve ark. (2008), Bacillus subtilis bakterisinden izole
edilen şimerik termostabil likenaz enziminin optimum aktivitesini pH 5.0 te
gösterdiğini saptamışlardır. Bacillus sp. L-12 likenaz enzimi ile elde edilen sonuçlar

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
75
literatür verileri ile birebir uyum içinde olup, enzim asidofiliktir.
Ön inkübasyondan geçirilmemiş enzimden elde edilen aktivite sonucu ile
karşılaştırıldığı zaman kalan enzim aktivitesinin 24 saat sonunda pH 6.0 da %96
olduğu bulunmuştur. Enzimin optimum aktivite gösterdiği pH değerinin stabilitenin
en iyi olduğu pH değeri olduğu görülmektedir.
Enzim pH 3.0-6.0 aralığında ortalama %88 stabilitesini korumuştur. pH
aralığı 3.0-8.0 e çıkarıldığında stabilitenin ortalama %86 korunduğu görülmektedir.
pH aralığı 9.0-10.0 olarak seçildiğinde aktivitenin %75 korunduğu saptanmıştır.
Stabilite çalışmaları için seçilen bütün pH değerleri dikkate alındığında ise (pH 3.0-
10.0) orijinal aktivitenin 24 saatlik 37°C deki inkübasyon sonunda ortalama %83
devam ettiği saptanmıştır. Enzim yüksek düzeyde pH stabil özellik taşımaktadır
(Çizelge 4.8.).
Li ve ark (2006), Ba-EGA endoglukanaz enziminin pH 7.5 ve 10.5 te 30ºC de
2 saat inkübasyon sonunda başlangıç aktivitesini %80’in üzerinde devam ettirdiğini
belirtmişlerdir.
Zhu ve ark. (2008), yeni izole edilen beta 1,3 endoglukanaz enziminin pH
5.0-8.0 aralığında 25 dakika süreyle aktivitesini koruduğunu belirtmişlerdir. Jung ve
ark. (2007) Bacillus sp. A8-8 suşundan izole edilen termostabil β-(1,3-1,4)-glukanaz
enziminin pH 9.0-12.0 aralığında 2 saatlik inkübasyon sonunda orijinal aktivitesini
%40’ın üzerinde koruduğunu belirtmişlerdir.
George ve ark, (2001) İzole edilen endoglukanaz enziminin optimum
aktivitesinin pH 6.0-8.0 aralığında gösterdiğini saptanmışlardır. Enzim pH 5.0 de ve
50ºC’de 1 saat inkübe edildiğinde orijinal aktivitenin %100 korunduğu, pH 6.0 da ise
aktivitenin %55’e düştüğü gözlemişlerdir. Likenaz enzimi asidofilik ve asidostabil
özelliktedir.
Bacillus sp. L-12 likenaz enzimi optimum aktivitesini 100°C de göstermiştir
(Şekil 4.12.). Enzim 60 ve 70°C lerde %80 aktivite gösterirken 80°C %95, 90°C de
ise %96 aktivite göstermiştir. Sıcaklık 110°C ye yükseltildiğinde (yağ banyosunda
inkübasyon yapılmıştır) enzim aktivitesi %94 gibi oldukça yüksek bir değerde
ölçülmüştür (Çizelge 4.7.).
Bacillus sp. L-12 likenaz enziminin bu özellikleri dikkate alındığında

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
76
özellikle yüksek sıcaklıklarda (80-110°C arasındaki değerlerde ortalama %96.25
aktiviteye sahiptir) %95’lerden daha yüksek aktiviteye sahip olduğu görülmektedir.
Bu nedenle enzim yüksek düzeyde termofil özellik göstermektedir.
Yernool ve ark (1998), izolasyonunu gerçekleştirdikleri iki termostabil
endoglukanaz enziminin optimum aktivitesini 95°C de gösterdiğini saptamışlardır.
Zverlov ve ark. (1997) Thermotoga neopolitana suşundan izole edilen yüksek
düzeyde termostabil 1,3-beta endoglukanaz enzimlerinden 73 kDa büyüklükteki
fragmentin optimum aktivitesini 95°C, 52 kDa büyüklükteki fragmentin 85°C de
göstediğini saptamışlardır. Çalışmamızda Termofil Bacillus sp. suşundan izole
ettiğimiz L-12 likenaz enzimi optimum aktivitesini 100ºC de göstermektedir (Şekil
4.12.). Literatür verilerinden daha yüksek optimum aktivite sıcaklığına sahip olan
enzim, hiper termofilik özelliktedir. L-12 likenaz enzimi hipertermofil özelliktedir.
Enzimin 30-60 ºC aralığındaki sıcaklık değerlerinde orijinal aktivitesini
ortalama %92.5, 30-70 ºC aralığında ortalama %90.6, 70-90 ºC aralığında ortalama
%79, ve 100-110 ºC aralığında ortalama %66.5 koruduğu belirlenmiştir (Çizelge
4.9.).
Fu ve ark. (2008), Bacillus subtilis suşundan elde edilen şimerik termostabil
likenaz enziminin 90 ºC de 10 dakikalık ön inkübasyondan sonra orijinal aktivitesini
yaklaşık %83. oranında koruduğunu belirtmişlerdir.
Zverlov ve ark. (1997) Thermotoga neapolitana suşundan izole edilen yüksek
düzeyde termostabil 1,3-beta endoglukanaz enzimine ait 73 kDa’luk fragmentin 30
dakika süreyle 100 ºC de ön inkübasyonundan sonra orijinal aktivitesinin %57 ye
düştüğünü belirtmişlerdir.
Hakamada ve ark, (1997), alkalofilik Bacillus sp. KSM-S237’den izole edilen
ve optimum aktivite sıcaklığı 50ºC olan endoglukanaz enziminin 10 dakikalık
inkübasyon sonunda orijinal aktivitesini 20ºC’de %100 koruduğunu, 40ºC’de
aktivitenin %90’a, 50ºC’de %50’ye 100ºC’de ise %30’a düştüğü belirtmiştir.
Bacillus sp. L–12 likenaz enzimine ait termal stabilite sonuçları literatür
verilerinden oldukça yüksek değerlere sahip olup, enzim yüksek düzeyde termostabil
özellik göstermektedir.
Bacillus sp. L-12 likenaz enzimi değişik NaCl konsantrasyonlarında analiz

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
77
edilmiş ve en yüksek aktivite değeri %7.5 NaCl konsantrasyonunda %67 olarak
bulunmuştur. %3.5-7.5 NaCl kullanıldığında ortalama kalan enzim aktivitesi %62
olarak gerçekleşirken, %10-20 NaCl konsantrasyonunda ortalama % 54.33 olarak
saptanmıştır (Çizelge 4.10.).
Simankova ve ark. (1993), Hallocella cellulosilytica bakterisinden izole
edilen enzimin mikrokristalin selülozu %5-20 arasındaki NaCl konsantrasyonlarında
parçaladığını, fakat optimum aktivitenin %10 NaCl konsantrasyonunda
gerçekleştiğini belirtmişlerdir.
L-12 likenaz enziminin %3.5-20.0 NaCl konsantrasyonlarında ortalama %59
aktivite göstermesi, enzimin halotolerant olduğununun kanıtıdır.
Bacillus sp. L-12 likenaz enzimi EDTA ve CaCl2 kullanılan testlerde stabil
kalarak sırası ile %97 ve %95 oranında orijinal aktivitesini korumuştur.
L-12 likenaz enzimi SDS ile %62 oranında aktivitesini kaybederken non
iyonik deterjan olan triton-X 100 ile %101 oranında aktivite göstermiştir. Bacillus sp.
L-12 likenaz enzimi Na2SO3, ZnCL2 Üre kullanılan çalışmalarda orijinal aktivitesini
sırası ile % 60, %64 ve %69 oranında kaybetmiştir (Çizelge 4.11.).
Joung ve ark. (2007), Bacillus sp. A8-8 bakterisinden izole edilen termostabil
β-1,3-1,4 glukanaz enziminin 5 mM EDTA ile aktivitesini koruduğunu ve
etkilenmediğini, buna karşılık Ca ile aktivitenin arttığını belirtmişlerdir.
Ng Son ve ark. (2009), termofilik Geobacillus sp. 70PC53 bakterisinden izole
edilen ve geniş bir sıcaklık aralığında stabilite gösteren CelA endoglukanaz
enziminin konsantrasyona bağlı olarak Zn+2 ve Cu+2 iyonları ile güçlü şekilde inhibe
edildiğini, buna karşılık Ca+2 ile azda olsa sitümüle edildiğini belirtmişlerdir. Aynı
araştırma grubu CelA endoglukanaz enzim aktivitesinin Na+, Mg+2 ve EDTA ile
etkilenmediğini belirtmişlerdir.
Li ve ark. (2006) Bacillus sp. AC.1 bakterisinden izole edilen termoasidofilik
endoglukanaz enziminin %0.5 SDS ile %66 oranında inhibe olurken, %0.25 triton X-
100 ile %127 aktive edildiğini belirtmişlerdir. Aynı enzimin 5 mM EDTA ile orijinal
aktivitesini %95 koruduğu belirlenmiştir.
Akita ve ark. (2005) Bacillus halodurans C-125 bakterisinden izole edilen
beta glukanaz enziminin 1 mM Zn+2 ile inhibi olduğunu buna karşılık 10 mM

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
78
CaCl2ve EDTA’dan etkilenmediğini bildirmişlerdir.
Bacillus sp. L-12 likenaz enzimi metalloenzim özelliği göstermeyen, PMSF
ile inhibe olan (%44), ZnCl2ile önemli düzeyde inhibe olan (%64) ve noniyonik
surfaktan olan Triton X-100 den etkilenmeyen ve azda olsa aktivitesi uyarılan
(%101) enzim özelliğindedir. Çinko iyonları ve üre ile önemli derecede inhibe
olması (ZnCl2 ile %64, Üre ile %69) enzimin hiper termofil, termostabil ve
halotolerant karakterde olduğunu göstermektedir (Lin ve ark., 1998).
Bu özellikleri ile Li ve ark. (2006), nın Bacillus sp. AC-1 termoasidofilik
endoglukanaz enzimi ile tam bir uyum göstermektedir.
Likenaz enziminin zymogram analizi sonucunda birbirinden bağımsız
moleküler ağırlıkları 100740 Da ve 54400 Da olan iki aktivite bandı elde edilmiştir
(Şekil 4.17.).
Literatürde endoglukanaz enzimlerinin büyük çoğunluğunda moleküler
ağırlıkların 35 kDa 85 kDa arasında değiştiği belirtilmektedir (Han ve ark,1995;
Hakamada ve ark., 1997; Mawadza ve ark.,2000; Hakamada ve ark., 2002; Singh ve
ark., 2001; Endo ve ark.,2001; Kim ve ark., 2005;Voget ve ark., 2006; Zvereva ve
ark, 2006; Zhang ve ark, 2007; Aygan, 2008).
Zverlov ve ark. (1997), Thermotoga naepolitana organizmasından izole
edilen yüksek düzeyde termostabil endo-1,3- β-glukanaz enziminin 52 ve 73 kDa
ağırlığa sahip bağımsız iki alt birimden oluştuğunu belirtmişlerdir.
Huang ve Monk (2004), termofilik Caldibacillus cellulovorans bakterisinden
izole edilen endoglukanaz enziminin 85.1 kda moleküler ağırlığa sahip olduğunu,
optimum aktivitesini pH6.5-7.0 de gösterdiğini ve enzimin Zn+2 iyonu ile inhibe
olduğunu belirtmişlerdir.
Geçen son 20 yıl içerisinde selülaz kullanımı özellikle tekstil, temizlik, kağıt,
içecek ve şarap endüstrisinde önemli düzeyde artış göstermiştir. Ayrıca hayvan
yemlerinin üretimi, tekstil ve temizlik endüstrisi, kağıt hamuru hazırlama işletmeleri
ile tarımsal uygulamalar endoglukanazların kullanım alanları arasındadır.
Günümüzde selülaz (enduglukanaz) enziminin dünya enzim ticaretindeki yeri
yaklaşık %20 düzeyindedir. Selülazların ana uygulama alanları gıda, hayvan yemi
üretimi, tekstil, biyo-yakıt, kimya, kağıt ve kağıt hamuru endüstrisi, atıkların

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
79
giderimi, tıbbi ve farmasötik endüstrisi, protoplast üretimi, genetik mühendisliği ve
kirlilik giderimidir (Bhat ve Bhat, 1997).
İzolasyonunu gerçekleştirdiğimiz Bacillus sp. L-12 likenaz enziminin
optimum aktivitesinin pH 6.0 da gerçekleşmesi, pH stabilitesinin yüksekliği,
optimum aktivite sıcaklığının 100°C ve termostabil olması, orta düzeyde halofilik
özellik göstermesi ve triton X-100 gibi surfaktanlara dirençli olması, enzimin tekstil
uygulamaları, kağıt endüstrisi, temizlik alanı ve özellikle hayvan yemi hazırlanması
gibi bir çok biyoteknolojik alanda uygulanabilir özellikte olduğunu göstermektedir.
Bacillus türleri çeşitli kompleks substratlara karşı aktivite gösteren çok sayıda
ve çeşitli hidrolitik enzimler üretmekte ve salgılamaktadırlar. Bu nedenle Bacillus
cinsindeki organizmalar, endüstriyel alanda α-amilaz, proteaz, glukanaz, glukoz
izomeraz ve endonükleaz gibi enzimlerin üretiminde yaygın şekilde
kullanılmaktadırlar (Uhlig, 1998). Bu enzimler arasında endo β-(1,3)-(1,4)-
glukanazlar (likenazlar), endosperm hücrelerinin nişastasına α-amilazların ulaşımını
kolaylaştırdığından dolayı içki mayalama endüstrisinde spesifik uygulamalara
sahiptirler (Beckmann ve ark., 2006). β -1,3-1,4 glukanlar yüksek yapılı bitkilerin
hücre duvarlarının yapısında bulunan hem β-1,3 hemde β-1,4 bağlı D-glukoz içeren
lineer polisakkaritlerdir. Likenazların, β-glukanları diğer glukanazlara göre daha
yüksek düzeyde hidroliz etmeleri nedeniyle, kümes hayvanları ve domuz yemlerinin
hazırlanmasında giderek ön plana çıkmaktadırlar (Beckmann ve ark., 2006).
Endo-(1,3-1,4)- β- glukanaz enzimi bira üretimi sırasında malt enzimlerinin
daha iyi çalışmasını sağlamak için yoğun şekilde kullanılmaktadır. Termostabl β-
glukanazlar, maltın kurutulması sırasında inaktive olmadıkları için özellikle tercih
edilirler. Maltlanmış arpada bulunan β-1,3-1,4-glukanaza benzer özgüllük gösteren
bakteriyel β-1,3-1,4-glukanaz önemli bir endüstriyel enzimdir ve çoğunlukla ezme
işlemi esnasında akışkanlığı azaltmak için kullanılır. Hayvan yeminde, özellikle etlik
tavuk ve küçük domuzlarda, bakteriyel β-1,3-1,4-glukanazlar içeren enzimatik
preparatlar ilavesi arpa kökenli beslenmenin sindirilebilirliğini geliştirebilir ve
sağlıksal sorunları azaltabilir (Zhang ve ark., 2006).

5. SONUÇLAR VE ÖNERİLER Fatma GÖZÜKARA
80
Sonuç olarak;
1. Bacillus sp. L-12 likenaz enzimi asidik pH koşullarında (pH 3.0-6.0)
yüksek düzeyde aktivite göstermesi, geniş bir pH aralığında 24 saat süreyle yüksek
düzeyde stabil kalması, optimum enzimatik aktivitesini 100°C de gösterip, aynı
zamanda termostabil karakterde olması nedeniyle hayvan yemi hazırlama amacı ile
kullanılabilecek ideal bir enzimdir. Özellikle tavuk yemlerinin hazırlanması sırasında
peletleme işleminin 80°C de gerçekleştirilmesi ve beslenme sırasında yemin barsağa
ulaşıncaya kadar (kursak-mide yolunda pH 2-4 arasında değişmektedir) ortalama 4
saat beklemesi dikkate alınacak olursa, enzimin 24 saat süreyle pH 3.0-6.0
aralığındaki aktivite ve stabilitesi (ortalama %88) özel bir önem kazanmaktadır. Bu
özellikleri nedeniyle tavuk yem endüstrisi için ideal bir enzimdir.
2. Selülozik materyali parçalayan enzimler ya β-1,3 yada β-1,4 glikozid
bağlarını koparma ve molekülü hidroliz etme yeteneğindedirler. Bu nedenle hidroliz
ürünlerinin verimi düşük olmakta ve selülozik materyalden yararlanma oranı
azalmaktadır. İzolasyonunu gerçekleştirdiğimiz likenaz enzimi β-1,3-1,4 glikozid
bağlarını aynı derecede parlama yeteneğinde olduğu için selülozik materyalden elde
edilen verim üst düzeylere ulaşmaktadır. Bu özellik selüloz parçalanma ürünlerinin
kullanımının hedeflendiği, yem endüstrisi, içecek endüstrisi ve biyoetanol üretimi
alanları için, L-12 likenaz enziminin önemini artırmaktadır.
3. Likenaz enzimi arpadan malt üretimi sırasında amilaz enziminin
endosperme ulaşmasını kolaylaştırmak amacıyla yaygın şekilde kullanılmaktadır.
Malt oluşumu sırasında kurutma işlemi yüksek sıcaklıkta yapıldığı için izolasyonunu
gerçekleştirdiğimiz likenaz enziminden termofil özelliği nedeniyle bu alanda çok
daha yüksek verim alınmasında yararlanılması mümkündür.

81
KAYNAKLAR
AIBA, S., KILAL, K., and IMANAKA, T., 1983. Cloning and expression of
thermostable α-amilase gene from Bacillus stearotermophilus, Bacillus
subtilis. Appl. And Environm. Microbiol. 46 (5):1059-1065
AKITA, M., KAYATAMA, K., HATADA, Y., ITO, S., and HORIKOSHI, K. 2005.
A novel β-glucanase gene from Bacillus halodurans C-125. FEMS
Microbiology Letters. 248:9-15.
ANONYMOUS, 1917. Buffer solutions. Handbook of Biochemistry and Molecular
Biology, s. 370-382.
ANONYMOUS., 1988a., Handbook of amylases and related enzymes. Edited by the
amylase research society of japan. Pergamon press, Oxford.
ARIKAN, B. 2008. Highly Thermostable, Thermophilic, Alkaline, SDS and Chelator
Resistant Amylase from a Thermophilic Bacillus sp. Isolate A3-15.
Bioresource Technology. 98 (8):3071-3078.
AOKI, K., MIYAMOTO, K., MURAKAMI, S., and SHINKE, R., 1995. Anaerobic
Synthesis of Extracellular Proteases by The Soil Bacterium Bacillus sp. AM-
23: Putrification and Characterization of The Enzymes. Soil Biol. Biochem.
27(11):1377-1382.
AUNSTRUP, K., 1981. Industrial Aspects of Biochemistry (B. Spencer editör),
pp:23-29. FEES, Dublin.
AYGAN, A., 2008. Haloalkalofil Bacillus sp. İzolasyonu, Amilaz, Selülaz ve
Ksilanaz Enzimlerinin Üretimi, Karakterizasyonu ve Biyoteknolojik
Uygulamalarda Kullanılabilirliği. Ç.Ü. FBE. Doktora Tezi.
BAJPAI, P. and BAJPAI, K.P., 1987. High temperature alkaline a-amylase from
Bacillus licheniformis TCRDC-B13. Biotechnology and Bioengineering,
33:72-78.
BECKMANN, L., SIMON, O., and VAHJEN, W. 2006. Isolation and identification
of mixed linked β-glucan egrading bacteria in the intestine of broiler chickens
and partial characteriation of respective 1,3-1,4-glucanase activities. J Basic
Microbiol. 46 (3): 175-185.

82
BHAT, M.K., 2000. Cellulase and related enzymes in biotechnology. Biotechnology
Advences. 18 (5): 355-458.
BIODIN, A. and EFFRONT,J.,1917. Process manufacturing diastases and toxins by
oxidizing ferments. U.S.Patent 1, 227,525.
BISCHOFF, K.M., ROONEY, A.P., and LI, X.L., 2006. Purification and
characterization of a family 5 endoglucanase from a moderately thermophilic
strain of Bacillus licheniformis. Biotechnol Lett. 28:1761-1765
BHAT, M.K., and BHAT, S., 1997. Cellulose degrading enzymes and their potential
industrial applications. Biotechnology Advances, 15: 583-620.
BLEISMER, B.O., and HARTMAN, P.A., 1973. Diferential heat stabilities of
Bacillus subtilis amylases. J.Bacteriol. 113:526-528.
BOLLAG,D.M., ROZYCKI, M.D., EDELSTEIN, S.J., 1996. Protein Methods (2nd
Edt). Viley-Liss Press, USA. 414s.
BORGIA, P.T. and CAMPBELL, L.L., 1978. α-Amylases from Five Strains of
Bacillus amyloliquefaciens: Evidence for Identical Primary Structures.
J.Bacteriol. 134(2):389-393.
BOYCE, A. and WALSH, G. 2007. Production, purification and application-relevant
characterisation of an endo-1,2 (4)-β-glucanase from Rhizomucor miegei.
Appl Microbiol Biotechnol. 76:835-841.
BOYER, F.W., INGLE, M.B., and MERCER, G.D. 1972. Starch. 31:66-71
BURHAN, A., ÜNALDI, N., CORAL, G., COLAK, O., AYGAN, A., and
GÜLNAZ, O. 2003. Enzymatic properties of a novel thermostable,
thermophilic, alkaline and chelator resistant amylase from an alkaliphilic
Bacillus sp. İsolate ANT-6. Process Biochemistry. 38:1397-1403.
COLEMAN, G., and ELLIOTT, W.H., 1962. Studies on α-Amylase formation by
Bacillus subtilis, Biochcm. J. 83:256-263.
CORAL, G., ARIKAN, B., ÜNALDI, M.N., and GÜVENMEZ KORKMAZ, H.
2002. Some properties of thermostable xylanase from an Aspergillus niger
strain. Ann. Microbiol. 52:299-306.

83
CHRISTOV, L.P., AKHTAR, M., and PRIOR, B.A. 1996. Impact of Xylanase and
Fungal Pretreatment on Alkali Solubility and Brightness of Dissolving Pulp.
Biobleaching of Sulphite Pulp. 50: 579-582.
ECKERT, K., and SCHNEIDER, E. 2003. A thermoacidophilic endoglucanase
(CelB) from Alicyclobacillus acidocaldarius displays high sequence
similarity to arabinofuranosidases belonging to family 51 of glycoside
hydrolases. Eur. J. Biochem. 270:3593-3602 (2003).
ENDO, K., HAKAMADA, Y., TAKIZAWA, S., KUBOTA, H., SUMITOMO, N.,
KOBAYASHI, T., and ITO, S. 2001. A novel alkaline endoglucanase from
an alkaliphilic Bacillus isolate: Enzymatic properties, and nucleotide and
deduced amino acid sequences. Appl Microbiol Biotechnology. 57:109-116.
FOGARTY, W.M., and KELLY, C.T., 1979. Developments in microbial
extracellular enzymes. In: Wiseman A, editor. Topics in enzyme and
fermentaion biotechnology. 3:45-108.
FU, L.L., XU, Z.R., SHUAI J.B., HU, C.X., DAI, W., and LI, W.F., 2008. High-
Level Secretion of a Chimeric Thermostable Lichenase from Bacillus subtilis
by Screening of Site- Mutated Signal Peptides with Structural Alterations.
Curr Microbiol 56:287-292.
GAMERITH, G., GROICHER, R., ZEILINGER, S., HERZOG, P., KUBICEK, C.P.,
1992. Cellulase-Poor Xylanases Produced by Trichoderma reesei RUTC-30
on Hemicellulase Substrates. Appl Microbiol Biotechnol. 38: 315-322.
GEORGE, S.P., AHMAD, A., RAO,M.B., 2001. Studies on carboxymethyl cellulase
produced by an alkalothermophilic actinomycete. Bioresource Tech., 77:171-
175.
GESSESSE, A., and GASHE. B:A., 1997. Production of Alkaline Xylanase by an
Alkaliphilic Bacillus sp. Isolated From an Alkaline Soda Lake. Journal of
Applied Microbiology., 83: 402-406.
GLICK, B.R., and PASTERNAK, S.S., 1994. Molecular biotechnology: Principles
and Applications of Recombinant DNA. Cold Spring Harbor Laboratory,
New York.

84
GOMES, J. and STEINER,W., 2004. The Biocatalytic Potential of Extremophiles
and Extremozymes. Food Technology and Biotechnology, 42(4): 223-235.
GROOTEGOED, J.A., and LAUVERS, and A.M., 1973. Separation and partial
purification of extracellular amylase and protease from Bacillus caldolyticus
Arch. Mikrobiol 90:223-232.
HAN, S.J., Y.J. YOO, and H.S. KANG. 1995. Characterization of a bifunctional
cellulase and its structural gene. J.Biol. Chem. 270:26012-26019.
HAKAMADA, Y., ENDO, K., TAKIZAWA, S., KOBAYASHI, T., SHIRAI, T.,
YAMANE, T., and ITO, S., 2002. Enzymatic properties, crystallization and
deduced aminoacid sequence of an alkaline endoglucanase from Bacillus
circulans. Biochimica et Biophysica Acta,1570: 174-180.
HAKAMADA, Y., KOIKE, K., YOSHIMATSU, T., MORI, H., KOBAYASHI, T.,
and ITO,S., 1997. Thermostable alkaline cellulase from an alkaliphilic
isolate, Bacillus sp. KSM-S237. Extremophiles, 1:151-156.
HAMILTON, LM., KELLY, C.T., and FOGARTY, W.M. 1999. Purification and
properties of the raw starch degrading α-amylase of Bacillus sp. IMD434.
Biotechnol. Lett. 2:111-115.
HIRASAWA, K., UCHIMURA, K., KASHIWA, M., GRANT, W.D., ITO, S.,
KOBAYASHI, T., and HORIKOSHI, K. 2006. Salt-activated endoglucanase
of a strain of alkaliphilic Bacillus agaradhaerens. Antonie van Leeuwenhoek.
89:211-219.
HOLS, P., FERAIN, T., GARMYN, D., BERNARD, N., and DELCOUR, J., 1994.
Use of expression secretion signals and vector free stable chromosomal
integration in engineering of Lactobacillus plantarum for α-amilaseand
Levanase expression. Appl. And Environm. Microbiol. 60 (5): 1401-1403.
HORIKOSHI, K., 1996. Alkaliphiles-from an industrial point of view. FEMS
Microbiology Reviews, 18:259-270.
HORIKOSHI, K., 1999. Alkaliphiles: Some applications of their products for
biotechnolgy. Microbiology and Molecular Biology Reviews. 63 (4): 735-
750.

85
HREGGVIDSSON, G.O., KAISTE, E., HOLST, O., EGGERTSSON, G.,
PALDSDOTTIR, A., and KRISTJANSSON, J. 1996. An extremely
thermostable cellulase from the thermophilic eubacterium Rhodothermus
marinus. App. Environmental Microbiology, 62 (8):3047-3049.
HUANG, X.P., and MONK, C., 2004. Purification and characterization of a cellulase
(CMCase) from a newly isolated thermophilic aerobic bacterium
Caldibacillus cellulovarans gen. nov., sp. nov. World J. Microbiol.
Biotechnol., 20: 85-92.
JIN, F., CHENG, X., SHI, Y., and ZHANG, C., 1990. Isolation of New
Thermophilic Aerobic Bacteria Which Produce Thermostable α-Amylase. J.
Gen. Appl. Microbiol. 36:415-424.
JOHN, F.K., 1987. Enzyme technology. Biotechnology, Vol 7A. NewYork. P. 37-62
JUNG, Y.J., YOO, J.S., and LEE, Y.S. 2007. Purification and characterization of
thermostable β-1,3-1,4 glucanase from Bacillus sp. A8-8. Biotechnol.
Bioprocess Eng. 12:265-270.
KANG, H-J., UEGAKI, K., FUKADA, H., and ISHIKAWA, K., 2007. Improvement
of the enzymatic activity of the hyperthermophilic cellulase from Pyrococcus
horikoshii. Extremophiles,11, 251-256.
KANG, S.W., PARK, Y.S., LEE, J.S., HONG, S.I., and KIM, S.W., 2004.
Production of cellulases and hemicellulases by Aspergillus niger KK2 from
lignocellulosic biomass. Bioresource Tech.,91: 153-156.
KIM, J-Y., HUR, S-H., HONG, J-H., 2005. Purification and characterization of an
alkaline cellulase from a newly isolated alkalophilic Bacillus sp. HSH-810.
Biotechnol. Lett., 27:313-316.
KINDLE, K.L., 1983. Characterization and production of thermostable α-amilase.
Appl. Biochem. Biotechnol. 8:153-158.
KRISHNA, C., 1999. Production of bacterial cellulases by solid state bioprocessing
of banana waste. Bioresorce Tech., 69:231-239.
KRISTJANSSON, J.K., and STETTER, K.O., 1992. Thermophilic Bacteria.
(J.K.Kristjansson, editör) Thermophilic Bacteria. CRC Pres, London,1-18s.

86
KUMAR, H.D. and NUSSINOV, R., 2001. How Do Thermophilic Proteins Deal
with Heat? A review. Cellular and Molecular Life Sciences, 58: 1216-1233.
KULKARNI, N., SHENDYE, A., and RAO, M., 1999. Moleculer and
Biotechnological Aspects of Xylanases. FEMS Microbiology Reviews 23:
411-456.
KUMAR, G.C., and TAKAGI, H., 1999. Microbial alkaline proteases from a
bioindustrial viewpoint. Biotechnology Advances. 17:561-594.
LAEMMLI, U.K., 1970., Cleauage of Structural Proteins During the Assembly of the
Head of the Bacteriophage T4. Nature, 227: 680-685.
LANE, A.G. and PIRT, S.J., 1973. Production of Cyclodextrin Glycoyltransferase by
Batch and Chemostat Cultures of Bacillus macerans in Chemically Defined
Mediom. J. Appi. C hem. Biotech., 23:309-321.
LENNETE, E.H., BALOWS, A., HAUSLER, J.W. Jr., and SHADOMY, J.H., 1985.
Manuel of clinical Microbiology. USA, Vol 4, p.1149.
LI, Y-H., DING,M., WANG, J., XU, G-J., and ZHAO, F., 2006. A novel
thermoacidophilic endoglucanas, Ba-EGA, from a new cellulosedegrading
bacterium, Bacillus sp. AC-1. Appl. Microbiol.Biotechnol., 70:430-436.
LIMING, X. AND XUELIANG, S. 2004. High-yield celllase production by
Trichoderma reesei ZU-02 on corn cob residue. Bioresource Technology.
91:259-262.
LIN, L.L., CHYAU, C.C., and HSU, W.H., 1998. Production and properties of a raw
strach degrading amylase from the thermophilic and alkaliphilic Bacillus sp.
Biotechnol. Appl. Biochem. 28:61-68
LOUW, M.E., REID, S.J. and WATSON, T.G. 1993. Characterization, cloning and
sequencing of a thermostable endo β-(1,3-1,4) glucanase-encoding gene from
an alkalophilic Bacillus brevis. Appl. Microbiol. Biotechnol. 38, 507–513.
MATSUZAKI, H., YAMANE, K., YAMAGUCHI, K, NAGATA, K., and MAROU,
B., 1974. Hybrid Alpha-Amylases Produced by Transformants of Bacillus
subtilis. I Purification and Characterization of Extracellular ct-Amylases
Produced by the Parental Strains and Transformants: Biochim. Biophy. Acta.,
365, 235-247.

87
MAWADZA, C., HATTI-KAUL, R., ZVAUYA, R., and MATTIASON, B., 2000.
Purification and characterization of cellulases produced by Bacillus Strain. J.
Biotechnol., 83: 177-187.
MEER, J.L., 1972. The Regulation Of Alpha-Amylase Production in Bacillus
licheniformis. Antonievon Leuvenhoek: J. Microb. Serol., 38:570-585.
MEHROTRA ,S., PANDEY, P.K., GAUR, R., and DARMWAL, N.S., 1999. The
prufication of alkaline protease by a Bacillus sp. isolated. Bioresource
Technology. 67:201-203.
MILLER, G.L., 1951. Use of Dinitrosalicylic Acid Reagent for Determination of
Reducing Sugar. Analytical Chemistry,426-428.
MORGAN, F.J. and PRIEST, F.G., 1981. Characterization of a thermostable α-
amylase from Bacillus licheniformis NCIB 6346. J.Appl. Bacteriol., 50:107-
114.
MURASHIMA, K., NISHIMURA, T., NAKAMURA, Y., KOGA, J., MORIYA, T.,
SUMIDA, N., YAGUCHI, T., and KONO,T., 2002. Purification and
characterization of new endo-1,4-β-D-glucanases from Rhizopus oryzae.
Enzyme. Microbial Technol., 30: 319-326
NG, L.S., LI, C.W., YEH, Y.F., and CHEN, P.T., 2009. A novel endoglucanase from
the thermophilic bacterium Geobacillus sp. 70PC53 with high activity and
stability over a broad range of temperatures. Extremophiles, 13:425-435.
NIEHAUS, F., BERTOLDO, C., KAHLER, M., and ANTRANIKIAN, G., 1999.
Extremophiles as a source of novel enzymes for industrial application. App
Microbiol Biotechnol, 51:711-729
OGASAHARA, K., IMANISHI, A., and ISEMURA, T., 1970. Studies on
Thermophilic α- Amylase from Bacillus stearothermophilus. I. Some General
and Physiochemical Properties of Thermophilic α- Amylase. J. Biochem.,
Vol.67, No:1 65-75.
OYEKOLA, O.O., NGESI, N., and WHITELEY, C.G. 2007. Isolation, purification
and characterisation of an endogluanase and β-glucosidase from an anaerobic
sulphidogenic bioreactor. Enzyme and Microbial Technology. 40:637-644.

88
PAZUR, H.H., 1965. Starch Chemistry and Technology, p: 133 Academic Press,
New York.
RADLEY, J.A., 1976. Production of Microbial Amylolytic Enzymes: Starch
Production Technology (L.A. Underkofler Editör). Chapter 16., Applied
Science Publisher Ld., England, p: 295-309.
RAO, B.M., TANKSALE, M.A., GHATHE, S.M., and DESHPANDE, V.V., 1998.
Molecular and biotechnological aspects of microbial proteases. Microbiology
and Molecular Biology Reviews. 62 (3):597-635.
ROBSON, L.M. and CHAMBLISS, G.H., 1984. Characterization of the cellulolytic
activity of a Bacillus isolate. Appl. Environ.Microbiol.,47:1039-1046.
RI, M.H., KIM, Y.S., JOO, A.R., LEE, J.K., KIM, Y.S., and OH, D.K., 2009.
Purification and Characterization of a Thermostable β-1,3-1,4-Glucanase
from Laetiporus sulphureus var. miniatus. J. Microbiol. Biotechnol. (2009),
19(2), 000–000 doi: 10.4014/jmb.0812.674
SAHA, B.C. 2004. Endoglucanase from a newly isolated strain of Mucor
circinelloides. Process Biochemistry 39(12): 1871-1876
SAITO, N., 1973. Thermophilic extracellular α-amilase from Bacillus licheniformis.
Arc. Biochem. Biophy. 155:290-298.
SAMBROOK, J. and RUSSELL, D.W.,2001. Molecular Cloning: A Laboratory
Manual, 3rd ed., Cold Spring Harbor Laboratory Press, New York, USA.
SARIKAYA, E., 1995. α-amilaz üreten bazı Bacillus suşlarının gelişme
parametreleri, enzim özellik ve üretim koşullarının optimizasyonu. Ankara
Üniversitesi Fen Bilimleri Enstitüsü Doktora Tezi.
SIMANKOVA, M.V.,CHERNYCH, N.A., OSIPOV, G.A. and ZAVARZIN, G.A.,
1993. Hellocella cellulolytica gen. nov., sp. nov., a new obligately anaerobic,
halophilic, cellulolytic bacterium. Syst. Appl. Microbiol. 16. 385-389.
SINDHU, G.S., SHAMA, P., CHAKRABARTI, T., and GUPTA, J.K. 1997. Strain
improvement for the production of a themostable α-amylase. Enzyme
Microbil. Technol. 24:584-59.

89
SINGH, J., BATRA, N., and SOBTI, R.C., 2001. A highly thermostable, alkaline
CMCase produced by a newly isolated Bacillus sp. VG1. World J. Microbiol.
Biotechnol., 17: 761-765.
SMITH, J.E.,1996. Biotechnology. Chambridge University Pres, Chambridge, 233s.
SOUTSCHEK-BAUER, E. and STAUDENBAUER L., 1987. Synthesis and
secretion of a heat stable carboxymethylcellulase from Clostridium
thermocellum in Bacillus subtilis and Bacillus stearothermophilus. Mol.Gen.
Genet., 208: 537-541.
SRIVASTAVA, R.A.K., 1987. Purification and Chemical Characterization of
Thermostable Amylases Produced by Bacillus stearothermophilus.
Biochemistry Laboratory, Botany Department, University of Gorakhpur,
Gorakhpur 273001, India.
TABERNERO, C., COLL, P.M., FERNANDEZ-ABALOS, J.M., PEREZ, P., and
SANTAMARIA, R.I., 1994. Cloning and DNA squencing of bgaA, a gene
encoding an endo-β-1,3-1,4-glucanase, from an alkalophilic Bacillus strain
(N137). Appl. Environ. Micribiol. 60,1213-1220.
TARAKÇIOĞLU, Y., 1979. An amylase producing maltotriose from Bacillus
subtilis. Agric. Biol. Chem. 49 (4):1091-1097.
TAHARA, N., YANA, H., YOSHINAGA, F., 1997. Two types of Cellulase Activity
Produced by a cellulose-producing Acetobacter Strain. Journal of
Fermentation and Bioengineering, 83: 389-392.
TEMİZKAN, G. ve ARDA, N., 2004. Moleküler Biyolojide Kullanılan Yöntemler.
Nobel Tıp Kitabevi, İst.345s.
UHLIG, H. 1998. Industrial enzymes and their applications. JOHN WILEY&SONS,
INC. NEW YORK.
VOGET, S., STEELE, H.L., and STREIT, W.R., 2006. Characterization of
metagenome-derived halotolerant cellulase. J. Biotechnol 126:26-36.
WAINQ, M., and INGWORSEN, K. 2003. Roduction of β-xylnase and β-xylosidase
by the extremelyhalophilic arceon Halorhabdus utahensis. Extremophiles,
7:87-93.

90
WANG, C.H., HSİEH, Y.R., NG, C.C., CHAN, H., LIN H.T., TZENG W.S., and
SHYU, Y.T., 2009. Purification and characterization of a novel halostable
cellulase from Salinivibrio sp. strain NTU-05. Enzyme and Microbial Techn.,
44:373-379
WILSON, K. And GOULDING, K.H., 1986. A Biologist’s Guide to Principles and
Techniques of Practical Biochemistry. Edward Arnold, London.396s.
WISEMAN, A., 1987. Handbook of enzyme biotechnology. Second Edition. Chapter
3. The application of enzymes in industry, p.274-373.
WOLFGANG, A. 2004. Enzymes in industry: Production ad applications. WILEY-
VCH Verlag GmbH&Co. KgaA, Weinheim.
YERNOOL, J-D.B.D., and EVELEİGH, D.E., 1998. Purification, characterization
and molecular analysis of thermostable cellulase CelA and CelB from
Thermotoga neapolitana. Appl.Environ.Microbiol,64:4774-4781.
ZHANG, X.Y., RUAN, H., MU, L., HE, G., TANG, X.J., and CHEN, O.H. 2006.
Enhancement of the thermostability of β-1,3-1,4-glucanase by directed
evolution. J Zhejiang Univ SCIENCE A .7(11):1948-1955.
ZHU, B.W., ZHAO, J.G., YANG, J.F., MIKIRO, T., ZHANG, Z.S., and ZHOU,
D.Y., 2008. Purification and partial characterization of a novel β-1,3-
glucanase from the gut of sea cucumber Stichopus japonicus. Process
Biochem (2008) 43:1102-1106
ZVERAVA, E.A., FEDOROVA,T.V., KEVBRIN, V.V., ZHILINA, T.N. and
RABINOVICH, M.L. , 2006. Cellulase activity of a haloalkaliphilic
anaerobic bacterium, strain Z-7026. Extremophiles, 10: 53-60.
ZVERLOV, V. V., VOLKOV, I. Y., VELIKODVORSKAYA, T. V. and
SCHWARZ, W. H. (1997) Highly thermostable endo-1,3-β-glucanase
(laminarinase) LamA from Thermotoga neapolitana : nucleotide sequence of
the gene and characterization of the recombinant gene product. Microbiology
143, 1701–1708.

91
ÖZGEÇMİŞ
1983 yılında Adana’da doğdum. İlkokul, ortaokul ve lise öğrenimimi
Adana’da tamamladıktan sonra 2001 yılında Çukurova Üniversitesi Fen-Edebiyat
Fakültesi Biyoloji bölümünü kazandım.
2005 yılında mezun olup aynı yıl Çukurova Üniversitesi Fen Bilimleri
Enstitüsü Biyoteknoloji Anabilim Dalında Yüksek Lisansa başladım.
2006 yılında Acıbadem Hastanesi LabMed Adana’da mikrobiyoloji
laboratuvar teknisyeni olarak işe başladım. 2009 yılında Kan Bankası sorumlu
teknisyenliğine atandım ve halen çalışmaktayım.