
This includes
26
www.pnas.org/cgi/doi/10.1073/pnas. 115 Supplementary Information for Conserved fungal effector suppresses PAMP-triggered immunity by targeting plant immune kinases Hiroki Irieda, Yoshihiro Inoue, Masashi Mori, Kohji Yamada, Yuu Oshikawa, Hiromasa Saitoh, Aiko Uemura, Ryohei Terauchi, Saeko Kitakura, Ayumi Kosaka, Suthitar Singkaravanit-Ogawa, and Yoshitaka Takano Email: [email protected] This PDF file includes: Supplementary text Figs. S1 to S9 Tables S1 to S3 References for SI reference citations 1807297
Transcript of This includes

www.pnas.org/cgi/doi/10.1073/pnas. 115
SupplementaryInformationfor
ConservedfungaleffectorsuppressesPAMP-triggeredimmunityby
targetingplantimmunekinases
HirokiIrieda,YoshihiroInoue,MasashiMori,KohjiYamada,YuuOshikawa,Hiromasa
Saitoh,AikoUemura,RyoheiTerauchi,SaekoKitakura,AyumiKosaka,Suthitar
Singkaravanit-Ogawa,andYoshitakaTakano
Email: [email protected]
ThisPDFfileincludes:
Supplementarytext
Figs.S1toS9
TablesS1toS3
ReferencesforSIreferencecitations
1807297

1
Supplemental information
SI Materials and Methods
Plasmid construction Escherichia coli strain DH5α (1) was used as the host for DNA manipulation. All primers
used in this study are listed in Table S3.
All plasmids used for transient expression in N. benthamiana were derived from
pBICP35, a vector for protein expression under the control of the cauliflower mosaic
virus 35S promoter (2). To generate pBICP35-MoNIS1-mCherry, BamHI-BamHI
fragment of MoNIS1 was amplified from pBICP35-MoNIS1-HA by PCR, and the
amplified fragement was introduced into the corresponding site of pBICP35-mCherry (3).
To generate point mutation series of NIS1∆C30-HA, fragments of
CoNIS1∆C30-S123A-HA, CoNIS1∆C30-Y125A-HA, CoNIS1∆C30-V127A-HA and
CoNIS1∆C30-G132A-HA (all having BamHI and EcoRI sites at their 5′ and 3′ ends,
respectively) were amplified from C. orbiculare cDNA by PCR, digested with BamHI
and EcoRI, and introduced into the corresponding sites in pBICP35. To generate
C-terminally GFP-fused BAK1, BKK1, NbSERK3 and OsBAK1, a fragment of eGFP
was amplified by PCR, digested with EcoRI and KpnI, and introduced into the
corresponding sites of pBICP35, resulting in pBICP35-eGFP-C. Next, fragments of
BAK1, BKK1, NbSERK3 or OsBAK1 amplified from A. thaliana cDNA were digested
with BamHI and EcoRI, and introduced into the corresponding sites of pBICP35-eGFP-C.
For construction of BIK1-GFP, a PCR-amplified fragment of BIK1 was digested with
BamHI and PstI, and introduced into pHYPT-eGFP (3), resulting in
pHYPT-AtBIK1-GFP. A fragment of BIK1-eGFP was then amplified from
pHYPT-AtBIK1-GFP, digested with BamHI and KpnI, and introduced into the
corresponding sites in pBICP35. To generate pBICP35-3xFLAG-RBOHD, a fragment of
3xFLAG-RBOHD was amplified from A. thaliana cDNA by PCR, digested with BamHI
and EcoRI, and introduced into the corresponding sites of pBICP35. To generate
pBICP35-CtNIS1-HA, CtNIS1-HA was amplified from C. tofieldiae cDNA, provided by

2
Dr. K. Hiruma (NAIST), by PCR and introduced into pBICP35 digested with BamHI
using In-Fusion HD Cloning Kit (Takara Bio).
For yeast two-hybrid assay, to generate bait constructs, we amplified fragments of the
cytoplasmic regions of BAK1 and BKK1 from A. thaliana cDNA using PCR, digested
them with EcoRI and BamHI, and introduced them into the corresponding sites of
pGBKT7 (Clontech). To generate prey constructs, we amplified fragments of CoNIS1
and MoNIS1 lacking the signal peptide-encoding sequence from cDNAs of C. orbiculare
and M. oryzae by PCR, digested them with EcoRI and BamHI, and introduced them into
the corresponding sites of pGADT7 (Clontech).
The recombinant CoNIS1 lacking the signal peptide region was expressed in tobacco
BY-2 cells using the tomato mosaic virus (ToMV)-based inducible expression system (4,
5). The ToMV-based expression vector used for expressing this CoNIS1 was constructed
using a modified pBICER8-C0.3-HuIFN-γ-SRz (5). We replaced the IFN gene with
synthesized DNAs encoding CoNIS1 having an Arabidopsis basic chitinase signal
(KTNLFLFLIFSLLLSLSSAEF) sequence and 6xHis-tag sequence
(HHHHHHHHDDDDK) at its N terminus for secretion and purification purposes (the
CoNIS1 protein secreted from BY-2 lacks the basic chitinase signal sequence). The
synthesized NIS1 sequence is below:
5’-ATGAAAACTAACCTCTTCTTATTCCTTATCTTCTCCTTGCTCTTATCTTTGT
CTTCCGCTGAGTTCCACCACCATCACCACCACCATCATGATGATGATGATAA
GAGAATTTCAGGAATTGCTCTTCCACAAACAGTTAAAGCTGGAGATAATAT
TAACGCTATCGTTGTTACTGAGGGATACATCCAATCAGTTCAAGATATCGCT
ATTGCTTTTGGAGTTGCTCCAGCTGCTTCTGCTTATCCTGGTACTCTTTCAAC
ACTTTTGGGATCTTTTTACTTGGGTCCTGAACAATCAAACGTTCAAAACGAT
ATCACTGAACCAATTACAATTCCTGAGTCTTTGGTTCCAGGAGAATATGTTA
TCGCTGCTTCATTGTTTTCTTTGTACGGTGCTTCTTCATCTCCTACTGTTTCTA
ACTACGATGTTACAGTTAACGTTGTTGACGAGACTTCCACTACATACGTTAG
GTCTCAGTTTTATGTTGGTAATAGTAATTCCACAGTTTGCTTAGGAGGATAC
ACAAGAAAGATATAA-3’

3
For MoNIS1 gene disruption, an 8.7 kb fragment containing the MoNIS1 gene was
amplified and cloned into pCR-XL-TOPO to generate pCR-XL-MoNIS1 using the
TOPO XL PCR Cloning Kit (Invitrogen). To construct the MoNIS1 gene disruption
vector, the 3.1 kb upstream region of the MoNIS1 gene was amplified from
pCR-XL-MoNIS1 with the primers KMNS2 and AMNL2, and the 3.8 kb downstream
region was amplified with the primers EMNU2 and XMNAS2. The amplified products
were digested with KpnI and ApaI (for the upstream region) or with EcoRI and XbaI (for
the downstream region) and then inserted into the 5′ site (for the upstream region) or the
3′ site (for the downstream region) of the hygromycin resistance gene HPH in pCB1636
(6). For complementation assay of an MoNIS1 mutant with MoNIS1 or CoNIS1, a
fragment containing MoNIS1 or CoNIS1 was amplified, digested with NotI and BamHI,
or NotI and EcoRI, respectively, and ligated into the corresponding sites of pCB1531,
which carries the bialaphos resistance (bar) gene (6), generating pCB1531-MoNIS1 or
pBA-CoNIS1proORFter.
Plant lines and growth
Seeds of Arabidopsis thaliana lines Col-0, pen2-1 (7), bak1-5 (8), bik1-1
(SALK_005291) (9), and bik1-1 pbl1-1 (SALK_005291, SAIL_1236_D07) (10) were
sown on soil or rockwool (Grodan, Roermond, The Netherlands) and grown at 25˚C with
16 h of illumination per day. In the case of rockwool, nutrient medium was applied. N.
benthamiana was grown on soil in a controlled environment chamber at 25˚C with 16 h of
illumination per day. Rice (Oryza sativa cv Moukoto) (11), provided by Dr. A. Abe
(Iwate Biotechnology Research Center), and barley (Hordeum vulgare cv Nigrate) (12),
provided by Dr. Y. Tosa (Kobe University), were grown on soil at 28˚C and on
vermiculite at 23˚C, respectively, with 16 h of illumination per day.
Colocalization assay
BAK1-GFP and MoNIS1-mCherry proteins were expressed in N. benthamiana leaves by
Agrobacterium infiltration. The fluorescence of GFP and mCherry was visualized with
Olympus FluoView FV1200 confocal microscope with a Nikon 40x PlanApo objective at
2 dpi.

4
Yeast two-hybrid assays
Yeast two-hybrid assays were performed using the Matchmaker Gold Yeast Two-Hybrid
System (Clontech, 630489) according to the manufacturer’s handbook. pGBKT7-based
bait plasmids and pGADT7-based prey plasmids were transformed into the yeast strains
Y2H Gold and Y187, respectively (Clontech). Bait and prey strains were mixed in
2xYPDA medium (Clontech) and incubated at 30˚C for 20 h. Yeast-diploid colonies were
obtained after mating and plated on selective medium (DDO, SD/-Leu/-Trp).
Protein-protein interaction was analyzed by growing yeast on synthetic minimal medium
with 40 µg/mL X-α-Gal and 125 ng/mL Aureobasidin A (QDO/X/A,
SD/-Ade/-His/-Leu/-Trp/X-α-Gal/Aba). The results were recorded 4 days after plating.
ROS measurement ROS assays were performed by a previously described method (13) with some
modification. Three-millimeter leaf discs were cut from N. benthamiana transiently
expressing the indicated constructs and were incubated overnight in sterile water under
dark conditions. The leaf discs were then immersed in 50 µL distilled water, to which was
added 50 µL of a solution containing 400 µM luminol (Sigma-Aldrich, A8511), 20
µg/mL peroxidase (Sigma-Aldrich, P6782), and either 1 µM flg22 (Invitrogen) or 2
mg/mL chitin (Yaizu Suisan Kagaku Industry, NA-COS-Y). They were then assayed for
ROS production. Luminescence was measured using a Luminoskan Ascent 2.1 (Thermo
Fisher Scientific, Yokohama, Japan).
In vitro kinase activity assays A construct for the expression of NIS1 lacking its signal peptide was introduced into
transgenic BY-2 cells expressing the β-estrogen-activatable transcription factor XVE.
Each transgenic BY2 suspension-cultured cell line was maintained at 26˚C, and 150 µL
of the saturated suspension culture was transferred to 30 mL of fresh Murashige and
Skoog’s liquid medium each week. Recombinant NIS1 expression was induced by
adding 17β-estradiol (final concentration, 10 µM) to 300 mL culture, 2 days after
subculture. The culture medium of the BY-2 cells expressing NIS1 was ultracentrifuged
at 100,000 x g for 30 min and the supernatant was then purified with a Ni2+ affinity

5
column. The eluted fraction was dialyzed in 0.1 M sodium phosphate buffer (pH 7.0), 50
mM NaCl and 1 mM CaCl2. The in vitro kinase assay followed a previously described
method (14) with some modifications. MBP-fused kinases BAK1 and BIK1 were
expressed in E. coli strain BL21 (DE3) (15) carrying plasmid pMal MBP-BAK1 CD and
pMal MBP-BIK1, respectively (14), and purified with amylose resin (E8021, New
England Biolabs). His-tagged proteins (100 or 500 ng) and 100 ng of MBP-fused kinases
were incubated in kinase reaction buffer (25 mM Tris-HCl, pH 7.5, 5 mM MgCl2, 1.5 mM
MnCl2, 0.5 mM CaCl2, 10 µM ATP, 5 mM DTT) for 1 h at room temperature. [γ-32P]ATP
(370 kBq) was then added and incubated for 30 min at 30˚C. The reactions were stopped
by the addition of SDS sample buffer. Proteins were separated by SDS-PAGE.
Radioactivity in the dried gel was detected by BAS2000 (Fujifilm). As controls, proteins
were separated in independent gels and visualized with CBB.

Fig. S1. Amino acid sequence alignment of selected NIS1 homologs in fungi.Amino acid sequences were aligned using Clustal Omega (16) and alignment cleaning was performed with trimAl (17). Columns with percentage of physico-chemically equivalent residues > 50% is shown with frames. Within each framed columns, physico-chemically equivalent residues are shown in bold. Alignment gaps are indicated by dots. Orange hooked arrows indicate the deletion sites of CoNIS1 truncation mutants (CoNIS1∆C30 and CoNIS1∆C60), and blue and red boxes indicate point-mutated residues of CoNIS1∆C30. For full species and gene names, see Table S1 and S2.

Fig. S1. Continued.
ΔC60 ΔC30
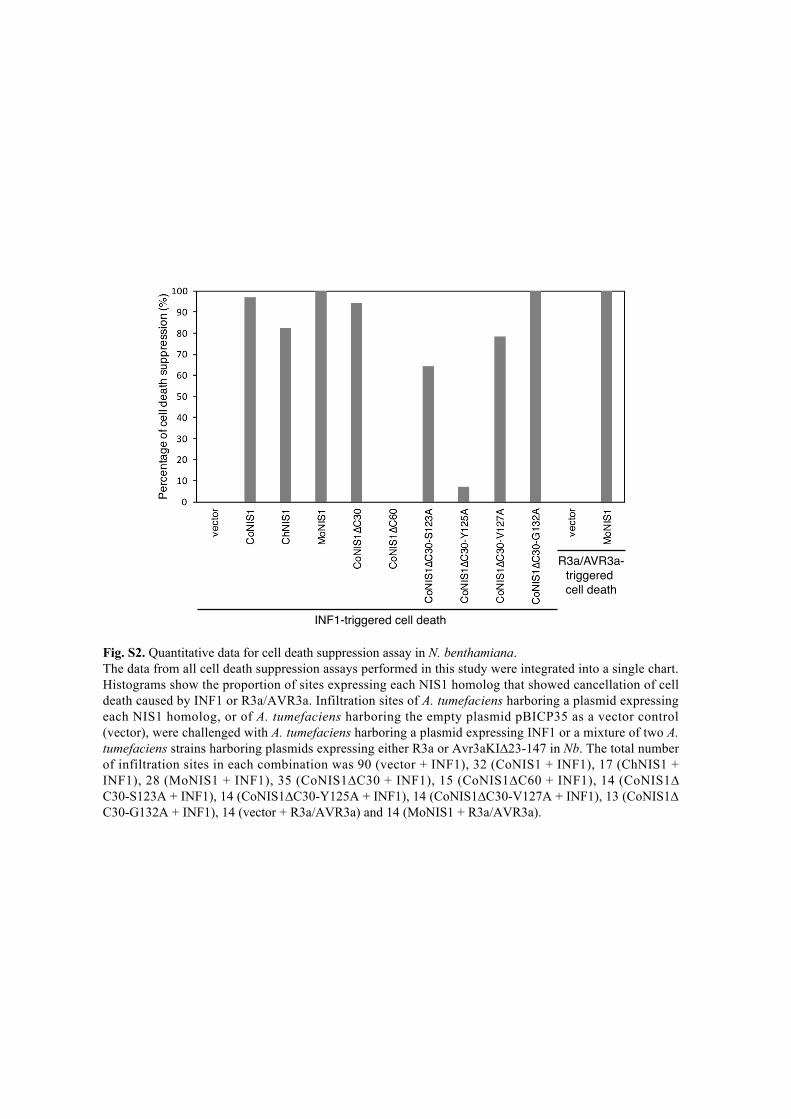
Fig. S2. Quantitative data for cell death suppression assay in N. benthamiana. The data from all cell death suppression assays performed in this study were integrated into a single chart. Histograms show the proportion of sites expressing each NIS1 homolog that showed cancellation of cell death caused by INF1 or R3a/AVR3a. Infiltration sites of A. tumefaciens harboring a plasmid expressing each NIS1 homolog, or of A. tumefaciens harboring the empty plasmid pBICP35 as a vector control (vector), were challenged with A. tumefaciens harboring a plasmid expressing INF1 or a mixture of two A. tumefaciens strains harboring plasmids expressing either R3a or Avr3aKI∆23-147 in Nb. The total number of infiltration sites in each combination was 90 (vector + INF1), 32 (CoNIS1 + INF1), 17 (ChNIS1 + INF1), 28 (MoNIS1 + INF1), 35 (CoNIS1∆C30 + INF1), 15 (CoNIS1∆C60 + INF1), 14 (CoNIS1∆C30-S123A + INF1), 14 (CoNIS1∆C30-Y125A + INF1), 14 (CoNIS1∆C30-V127A + INF1), 13 (CoNIS1∆C30-G132A + INF1), 14 (vector + R3a/AVR3a) and 14 (MoNIS1 + R3a/AVR3a).
INF1-triggered cell death
R3a/AVR3a-triggered cell death
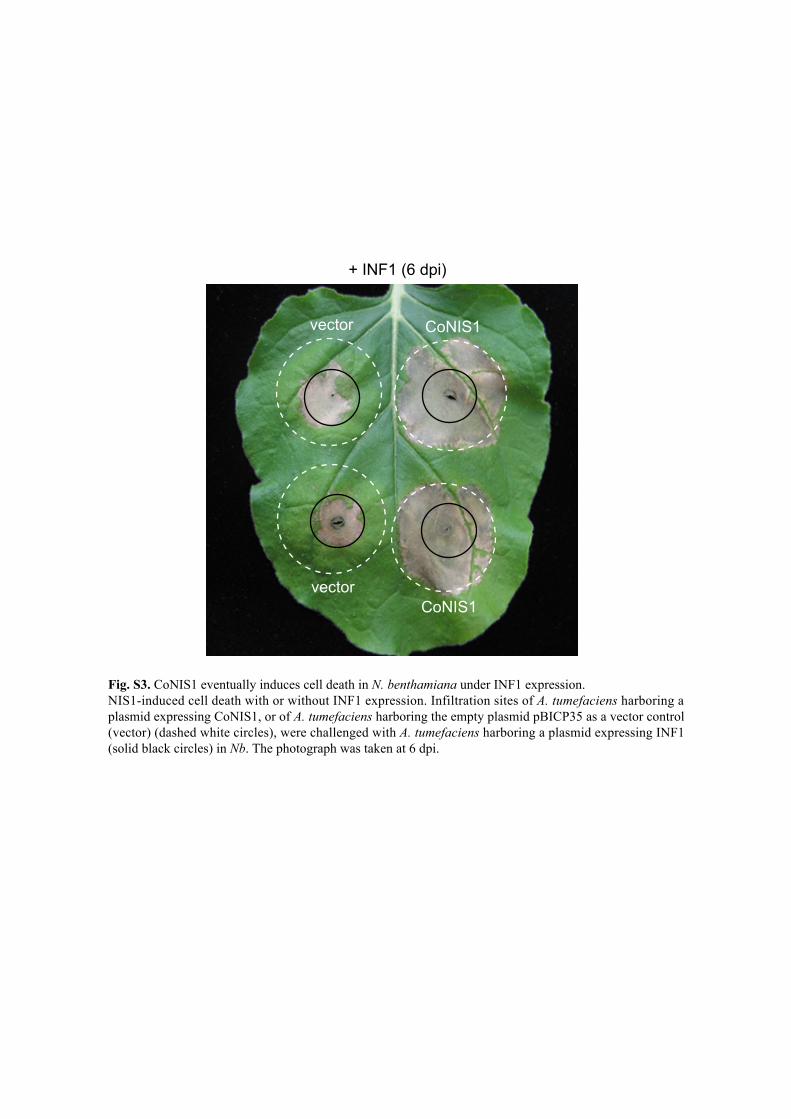
Fig. S3. CoNIS1 eventually induces cell death in N. benthamiana under INF1 expression. NIS1-induced cell death with or without INF1 expression. Infiltration sites of A. tumefaciens harboring a plasmid expressing CoNIS1, or of A. tumefaciens harboring the empty plasmid pBICP35 as a vector control (vector) (dashed white circles), were challenged with A. tumefaciens harboring a plasmid expressing INF1 (solid black circles) in Nb. The photograph was taken at 6 dpi.
+ INF1 (6 dpi)
vector
vector
CoNIS1
CoNIS1
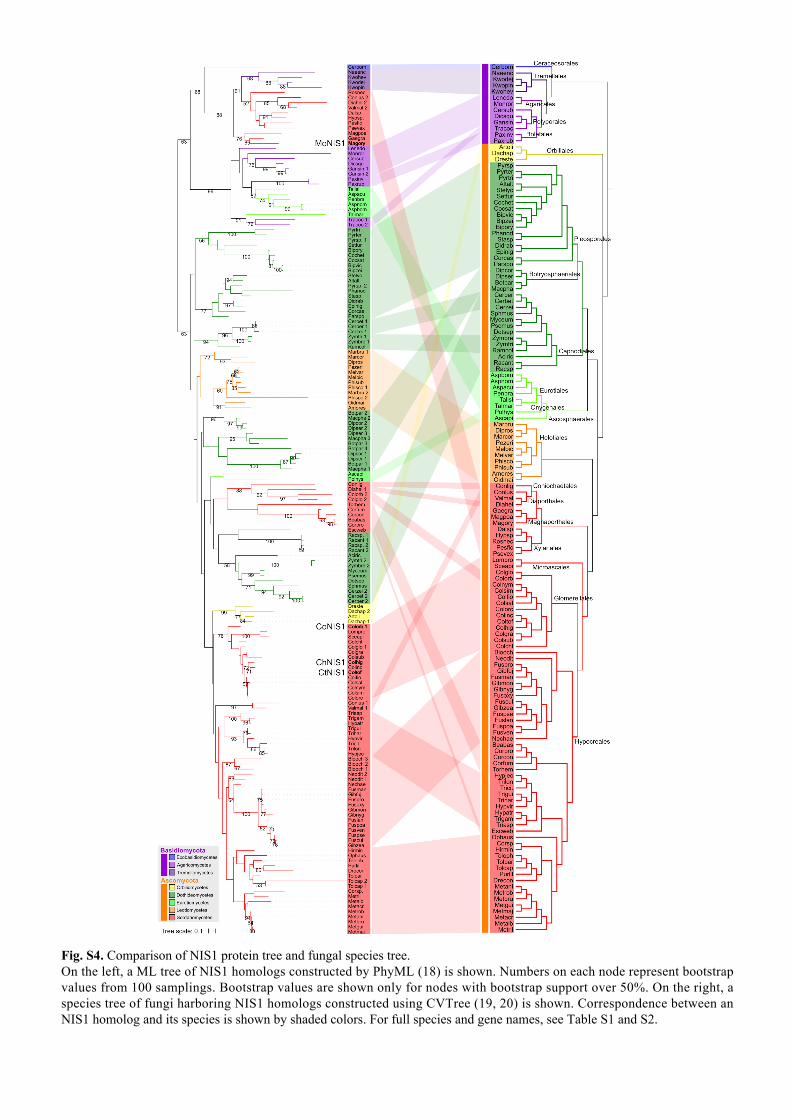
Fig. S4. Comparison of NIS1 protein tree and fungal species tree. On the left, a ML tree of NIS1 homologs constructed by PhyML (18) is shown. Numbers on each node represent bootstrap values from 100 samplings. Bootstrap values are shown only for nodes with bootstrap support over 50%. On the right, a species tree of fungi harboring NIS1 homologs constructed using CVTree (19, 20) is shown. Correspondence between an NIS1 homolog and its species is shown by shaded colors. For full species and gene names, see Table S1 and S2.

Fig. S5. MoNIS1 fused with mCherry suppresses INF1-induced cell death in Nb. Infiltration sites of A. tumefaciens harboring a plasmid expressing MoNIS1-mCherry, or of A. tumefaciens harboring a plasmid expressing mCherry (dashed white circles), were challenged with A. tumefaciens harboring a plasmid expressing INF1 (solid black circles) in Nb. The photograph was taken at 3 dpi.
mCherry MoNIS1-mCherry
+INF1 (3 dpi)
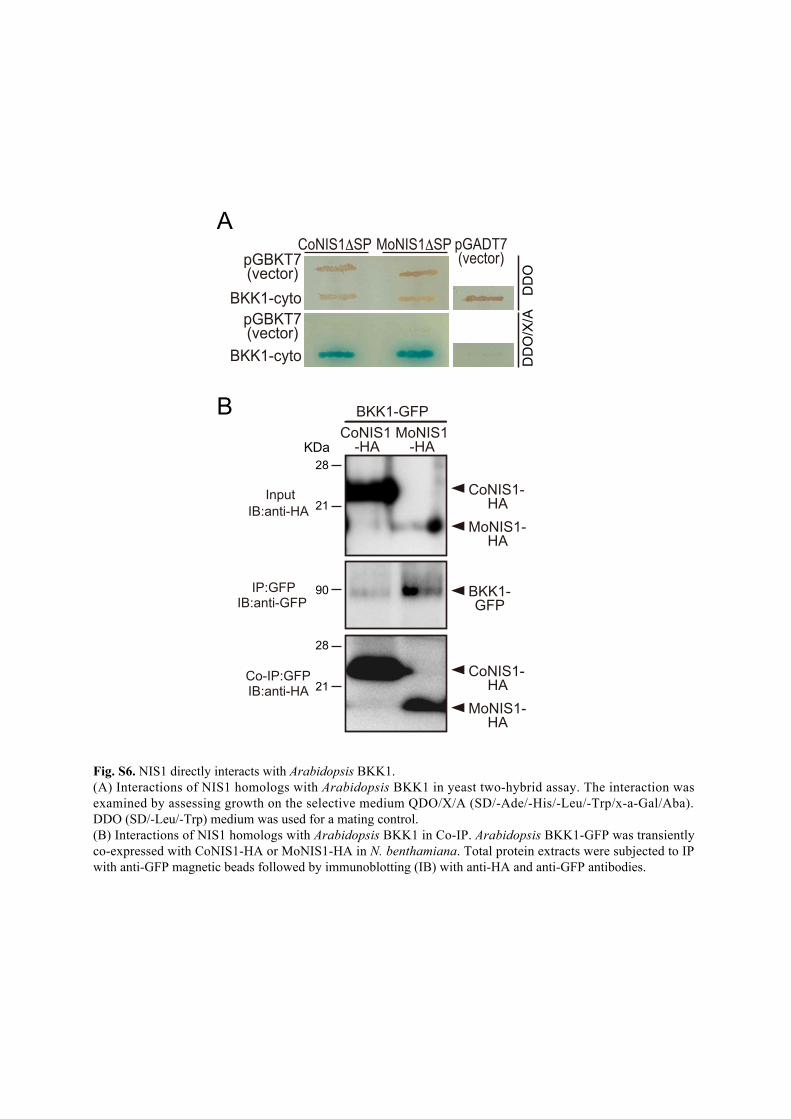
BKK1-cyto
pGBKT7(vector)
BKK1-cyto
pGBKT7(vector)
DD
OD
DO
/X/A
pGADT7(vector)
CoNIS1ΔSP MoNIS1ΔSP
InputIB:anti-HA
IP:GFPIB:anti-GFP
Co-IP:GFPIB:anti-HA
BKK1-GFPCoNIS1
-HAMoNIS1
-HA
A
B
MoNIS1-HA
BKK1-GFP
MoNIS1-HA
CoNIS1-HA
CoNIS1-HA
Fig. S6. NIS1 directly interacts with Arabidopsis BKK1. (A) Interactions of NIS1 homologs with Arabidopsis BKK1 in yeast two-hybrid assay. The interaction was examined by assessing growth on the selective medium QDO/X/A (SD/-Ade/-His/-Leu/-Trp/x-a-Gal/Aba). DDO (SD/-Leu/-Trp) medium was used for a mating control. (B) Interactions of NIS1 homologs with Arabidopsis BKK1 in Co-IP. Arabidopsis BKK1-GFP was transiently co-expressed with CoNIS1-HA or MoNIS1-HA in N. benthamiana. Total protein extracts were subjected to IP with anti-GFP magnetic beads followed by immunoblotting (IB) with anti-HA and anti-GFP antibodies.
KDa28
21
90
28
21
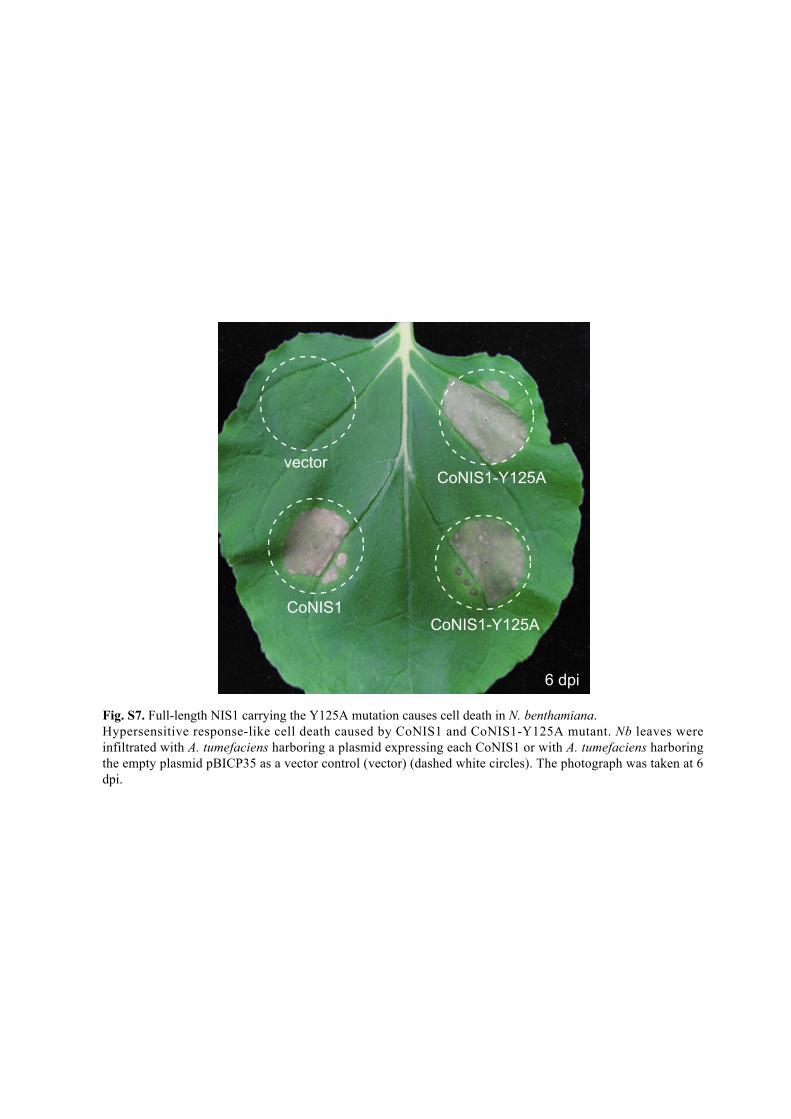
Fig. S7. Full-length NIS1 carrying the Y125A mutation causes cell death in N. benthamiana. Hypersensitive response-like cell death caused by CoNIS1 and CoNIS1-Y125A mutant. Nb leaves were infiltrated with A. tumefaciens harboring a plasmid expressing each CoNIS1 or with A. tumefaciens harboring the empty plasmid pBICP35 as a vector control (vector) (dashed white circles). The photograph was taken at 6 dpi.
vector
CoNIS1
CoNIS1-Y125A
CoNIS1-Y125A
6 dpi
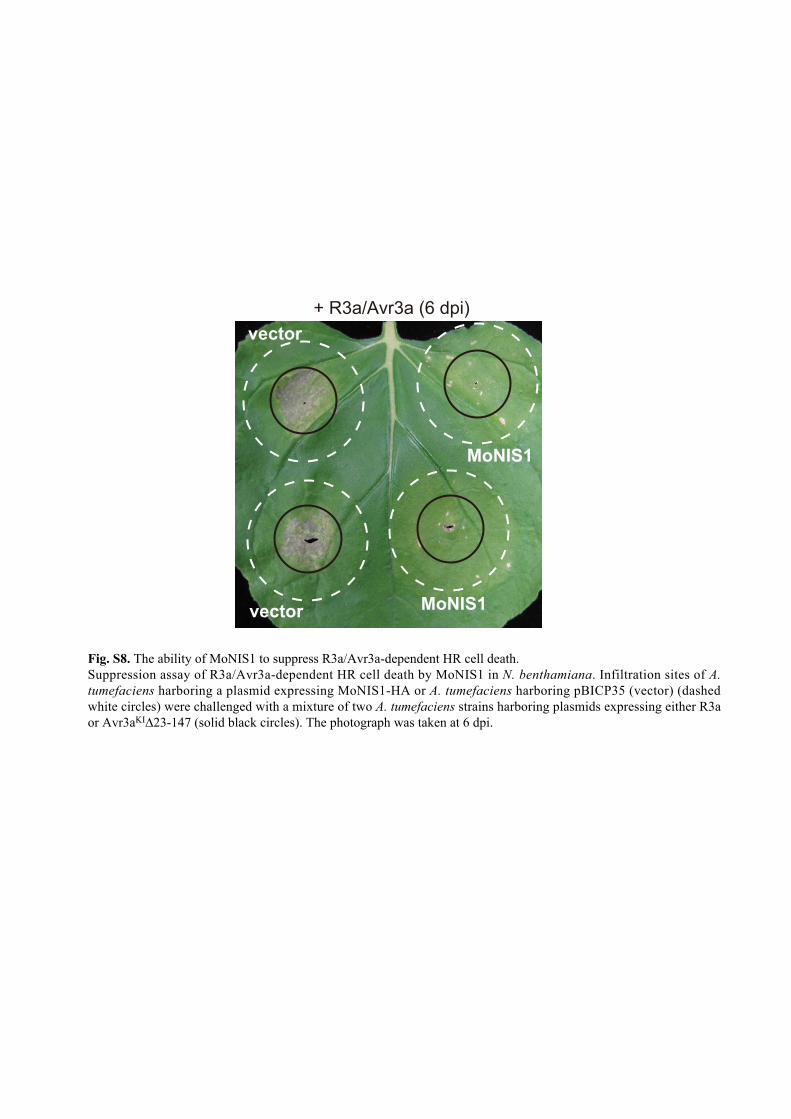
vector MoNIS1
MoNIS1
vector+ R3a/Avr3a (6 dpi)
Fig. S8. The ability of MoNIS1 to suppress R3a/Avr3a-dependent HR cell death. Suppression assay of R3a/Avr3a-dependent HR cell death by MoNIS1 in N. benthamiana. Infiltration sites of A. tumefaciens harboring a plasmid expressing MoNIS1-HA or A. tumefaciens harboring pBICP35 (vector) (dashed white circles) were challenged with a mixture of two A. tumefaciens strains harboring plasmids expressing either R3a or Avr3aKI∆23-147 (solid black circles). The photograph was taken at 6 dpi.
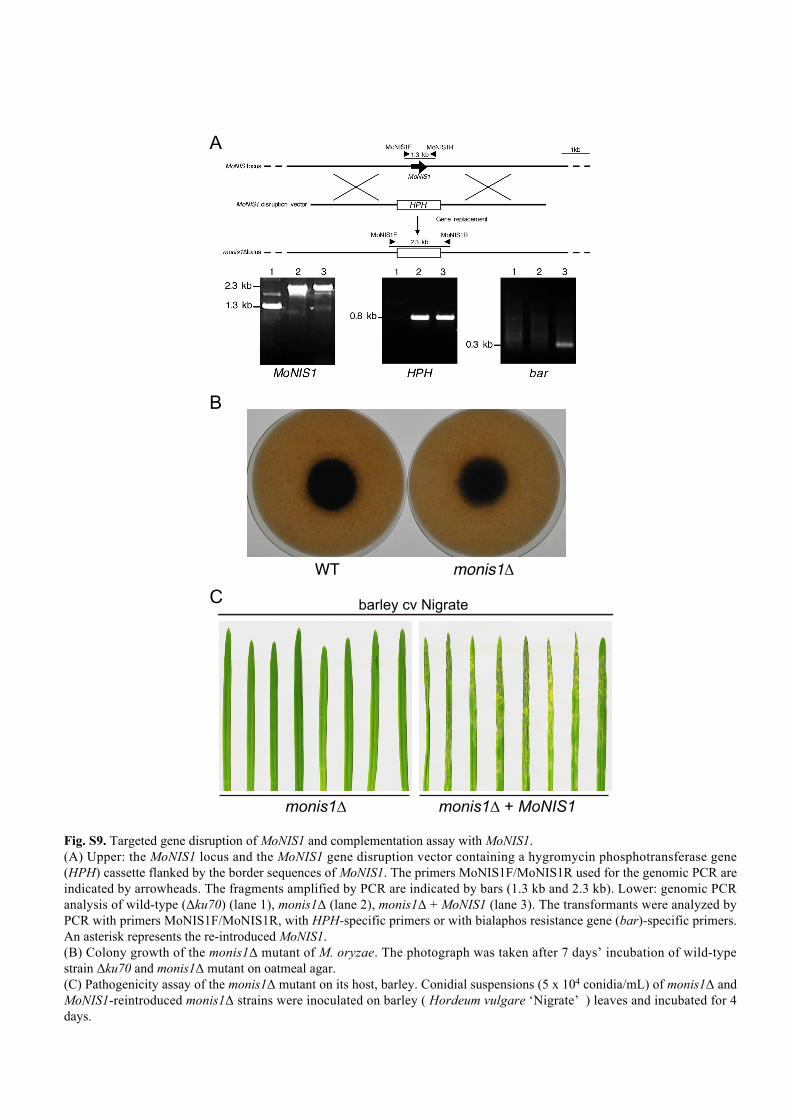
WT monis1Δ
monis1∆ monis1∆ + MoNIS1
A
B
C barley cv Nigrate
Fig. S9. Targeted gene disruption of MoNIS1 and complementation assay with MoNIS1. (A) Upper: the MoNIS1 locus and the MoNIS1 gene disruption vector containing a hygromycin phosphotransferase gene (HPH) cassette flanked by the border sequences of MoNIS1. The primers MoNIS1F/MoNIS1R used for the genomic PCR are indicated by arrowheads. The fragments amplified by PCR are indicated by bars (1.3 kb and 2.3 kb). Lower: genomic PCR analysis of wild-type (Δku70) (lane 1), monis1Δ (lane 2), monis1Δ + MoNIS1 (lane 3). The transformants were analyzed by PCR with primers MoNIS1F/MoNIS1R, with HPH-specific primers or with bialaphos resistance gene (bar)-specific primers. An asterisk represents the re-introduced MoNIS1. (B) Colony growth of the monis1Δ mutant of M. oryzae. The photograph was taken after 7 days’ incubation of wild-type strain Δku70 and monis1Δ mutant on oatmeal agar.(C) Pathogenicity assay of the monis1Δ mutant on its host, barley. Conidial suspensions (5 x 104 conidia/mL) of monis1Δ and MoNIS1-reintroduced monis1Δ strains were inoculated on barley ( Hordeum vulgare ‘Nigrate’ ) leaves and incubated for 4 days.
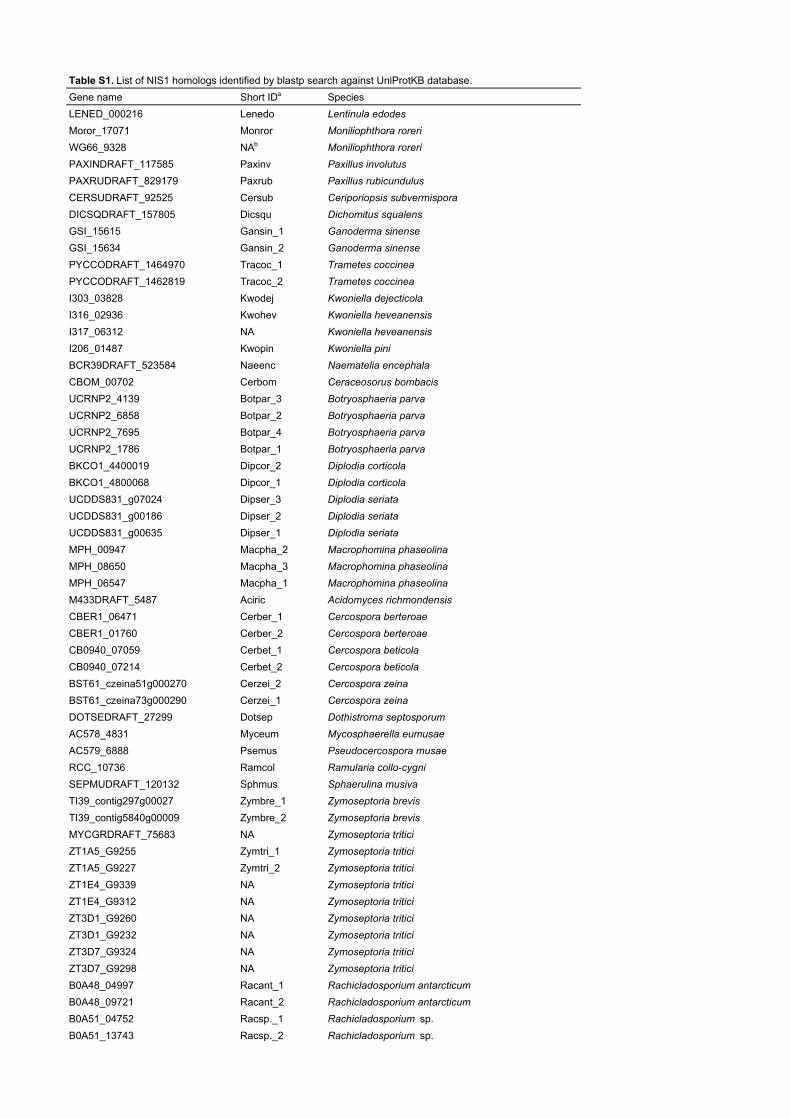
Table S1. List of NIS1 homologs identified by blastp search against UniProtKB database.Gene name Short IDa SpeciesLENED_000216 Lenedo Lentinula edodesMoror_17071 Monror Moniliophthora roreriWG66_9328 NAb Moniliophthora roreriPAXINDRAFT_117585 Paxinv Paxillus involutusPAXRUDRAFT_829179 Paxrub Paxillus rubicundulusCERSUDRAFT_92525 Cersub Ceriporiopsis subvermisporaDICSQDRAFT_157805 Dicsqu Dichomitus squalensGSI_15615 Gansin_1 Ganoderma sinenseGSI_15634 Gansin_2 Ganoderma sinensePYCCODRAFT_1464970 Tracoc_1 Trametes coccineaPYCCODRAFT_1462819 Tracoc_2 Trametes coccineaI303_03828 Kwodej Kwoniella dejecticolaI316_02936 Kwohev Kwoniella heveanensisI317_06312 NA Kwoniella heveanensisI206_01487 Kwopin Kwoniella piniBCR39DRAFT_523584 Naeenc Naematelia encephalaCBOM_00702 Cerbom Ceraceosorus bombacisUCRNP2_4139 Botpar_3 Botryosphaeria parvaUCRNP2_6858 Botpar_2 Botryosphaeria parvaUCRNP2_7695 Botpar_4 Botryosphaeria parvaUCRNP2_1786 Botpar_1 Botryosphaeria parvaBKCO1_4400019 Dipcor_2 Diplodia corticolaBKCO1_4800068 Dipcor_1 Diplodia corticolaUCDDS831_g07024 Dipser_3 Diplodia seriataUCDDS831_g00186 Dipser_2 Diplodia seriataUCDDS831_g00635 Dipser_1 Diplodia seriataMPH_00947 Macpha_2 Macrophomina phaseolinaMPH_08650 Macpha_3 Macrophomina phaseolinaMPH_06547 Macpha_1 Macrophomina phaseolinaM433DRAFT_5487 Aciric Acidomyces richmondensisCBER1_06471 Cerber_1 Cercospora berteroaeCBER1_01760 Cerber_2 Cercospora berteroaeCB0940_07059 Cerbet_1 Cercospora beticolaCB0940_07214 Cerbet_2 Cercospora beticolaBST61_czeina51g000270 Cerzei_2 Cercospora zeinaBST61_czeina73g000290 Cerzei_1 Cercospora zeinaDOTSEDRAFT_27299 Dotsep Dothistroma septosporumAC578_4831 Myceum Mycosphaerella eumusaeAC579_6888 Psemus Pseudocercospora musaeRCC_10736 Ramcol Ramularia collo-cygniSEPMUDRAFT_120132 Sphmus Sphaerulina musivaTI39_contig297g00027 Zymbre_1 Zymoseptoria brevisTI39_contig5840g00009 Zymbre_2 Zymoseptoria brevisMYCGRDRAFT_75683 NA Zymoseptoria triticiZT1A5_G9255 Zymtri_1 Zymoseptoria triticiZT1A5_G9227 Zymtri_2 Zymoseptoria triticiZT1E4_G9339 NA Zymoseptoria triticiZT1E4_G9312 NA Zymoseptoria triticiZT3D1_G9260 NA Zymoseptoria triticiZT3D1_G9232 NA Zymoseptoria triticiZT3D7_G9324 NA Zymoseptoria triticiZT3D7_G9298 NA Zymoseptoria triticiB0A48_04997 Racant_1 Rachicladosporium antarcticumB0A48_09721 Racant_2 Rachicladosporium antarcticumB0A51_04752 Racsp._1 Rachicladosporium sp.B0A51_13743 Racsp._2 Rachicladosporium sp.

BS50DRAFT_575309 Corcas Corynespora cassiicolaCC84DRAFT_1009733 Parspo Paraphaeosphaeria sporulosaIQ06DRAFT_290651 Stasp. Stagonospora sp.ST47_g1078 Didrab Didymella rabieiB5807_02474 Epinig Epicoccum nigrumSNOG_00069 Phanod Phaeosphaeria nodorumCC77DRAFT_1020930 Altalt Alternaria alternataCOCMIDRAFT_79993 Bipory Bipolaris oryzaeCOCVIDRAFT_11212 Bipvic Bipolaris victoriaeCOCCADRAFT_35713 Bipzei Bipolaris zeicolaCOCC4DRAFT_29978 NA Cochliobolus heterostrophusCOCHEDRAFT_1022529 Cochet Cochliobolus heterostrophusCOCSADRAFT_41606 Cocsat Cochliobolus sativusPTT_19513 Pyrter Pyrenophora teres f. teresPTRG_10762 Pyrtri Pyrenophora tritici-repentisSETTUDRAFT_162785 Settur Setosphaeria turcicaTW65_08014 Stelyc Stemphylium lycopersiciIQ07DRAFT_206793 Pyrsp._2 Pyrenochaeta sp.IQ07DRAFT_591166 Pyrsp._1 Pyrenochaeta sp.ASPACDRAFT_1891584 Aspacu Aspergillus aculeatusABOM_001603 Aspbom Aspergillus bombycisANOM_002819 Aspnom Aspergillus nomiusPMG11_09350 Penbra Penicillium brasilianumPISL3812_08033 Talisl Talaromyces islandicusPMAA_006330 Talmar Talaromyces marneffeiGQ26_0270540 NA Talaromyces marneffeiAAP_01282 Ascapi Ascosphaera apisAJ80_02549 Polhys Polytolypa hystricisBUE80_DR003311 Dipros Diplocarpon rosaeMBM_08313 Marbru_1 Marssonina brunnea f. sp. multigermtubiMBM_08390 Marbru_2 Marssonina brunnea f. sp. multigermtubiB2J93_8986 Marcor Marssonina coronariaeK444DRAFT_622704 Melbic Meliniomyces bicolorL207DRAFT_518835 Melvar Meliniomyces variabilisNA56DRAFT_21508 Pezeri Pezoloma ericaeLY89DRAFT_540464 Phisco_1 Phialocephala scopiformisLY89DRAFT_764533 Phisco_2 Phialocephala scopiformisPAC_01149 Phisub Phialocephala subalpinaM430DRAFT_76990 Amores Amorphotheca resinaeOIDMADRAFT_182184 Oidmai Oidiodendron maiusCCHL11_04114 Colchl Colletotrichum chlorophytiCFIO01_00805 Colfio Colletotrichum fioriniaeCGLO_15227 NA Colletotrichum gloeosporioidesCGLO_16733 NA Colletotrichum gloeosporioidesCGGC5_8593 Colglo_1 Colletotrichum gloeosporioidesCGGC5_2528 Colglo_2 Colletotrichum gloeosporioidesGLRG_05338 Colgra Colletotrichum graminicolaCH063_03697 Colhig Colletotrichum higginsianumCI238_06349 Colinc Colletotrichum incanumCSPAE12_00032 NA Colletotrichum incanumCNYM01_04434 Colnym Colletotrichum nymphaeaeCob_01067 Colorb_1 Colletotrichum orbiculareCob_00420 Colorb_2 Colletotrichum orbiculareCORC01_02258 Colorc Colletotrichum orchidophilumCSAL01_06451 Colsal Colletotrichum salicisCSIM01_08905 Colsim Colletotrichum simmondsiiCSUB01_11462 Colsub Colletotrichum sublineolaCT0861_07819 Coltof Colletotrichum tofieldiae

BN869_000006902_1 Biooch_1 Bionectria ochroleucaBN869_000006859_1 Biooch_2 Bionectria ochroleucaBN869_000007630_1 Biooch_3 Bionectria ochroleucaMAC_04101 Metacr Metarhizium acridumMAM_04654 Metalb Metarhizium albumMAN_05946 Metani Metarhizium anisopliaeH634G_05057 NA Metarhizium anisopliaeMBR_05417 Metbru Metarhizium brunneumMGU_05477 Metgui Metarhizium guizhouenseMAJ_06383 Metmaj Metarhizium majusNOR_06771 Metril Metarhizium rileyiMAA_08938 Metrob Metarhizium robertsiiX797_009566 NA Metarhizium robertsiiVHEMI02772 Torhem Torrubiella hemipterigenaBBA_00254 Beabas Beauveria bassianaBBAD15_g2256 NA Beauveria bassianaBB8028_0002g14990 NA Beauveria bassianaBM221_008189 NA Beauveria bassianaBBO_03959 Corbro Cordyceps brongniartiiLEL_01213 Corcon Cordyceps confragosaISF_08379 Corfum Cordyceps fumosoroseaCDD83_4783 Corsp. Cordyceps sp.ESCO_004832 Escweb Escovopsis weberiTRIATDRAFT_53493 Hypatr Hypocrea atroviridisM419DRAFT_130653 NA Hypocrea jecorinaTRIREDRAFT_61526 Hypjec Hypocrea jecorina TRIVIDRAFT_205388 Hypvir Hypocrea virensM441DRAFT_199594 Triasp Trichoderma asperellumBBK36DRAFT_1119116 Tricit Trichoderma citrinovirideTGAMA5MH_10387 Trigam Trichoderma gamsiiTGAM01_v210334 NA Trichoderma gamsiiA0O28_0068280 Trigui Trichoderma guizhouenseM431DRAFT_6545 Trihar Trichoderma harzianumCI102_11310 NA Trichoderma harzianumTHARTR1_09184 NA Trichoderma harzianumTHAR02_07916 NA Trichoderma harzianumM440DRAFT_1402529 Trilon Trichoderma longibrachiatumA9Z42_0038330 NA Trichoderma parareeseiFMAN_07996 Fusman Fusarium mangiferaeFPRO_08436 Fuspro Fusarium proliferatumFFUJ_07852 Gibfuj Gibberella fujikuroi FFC1_03428 NA Gibberella fujikuroi FVEG_09142 Gibmon Gibberella moniliformisFNYG_11156 Gibnyg Gibberella nygamaiFOXB_09017 NA Fusarium oxysporumFOC1_g10008116 NA Fusarium oxysporum f. sp. cubenseFOC4_g10009208 NA Fusarium oxysporum f. sp. cubenseFOIG_10872 NA Fusarium oxysporum f. sp. cubenseFOPG_11950 NA Fusarium oxysporum f. sp. conglutinansFOXG_10488 Fusoxy Fusarium oxysporum f. sp. lycopersiciFOMG_07263 NA Fusarium oxysporum f. sp. melonisFOVG_06740 NA Fusarium oxysporum f. sp. pisiAU210_008318 NA Fusarium oxysporum f. sp. radicis-cucumerinumFOQG_06093 NA Fusarium oxysporum f. sp. raphaniFOTG_09084 NA Fusarium oxysporum f. sp. vasinfectumFOZG_09971 NA Fusarium oxysporumFOYG_08673 NA Fusarium oxysporumFRV6_01449 NA Fusarium oxysporum

FCULG_00011427 Fuscul Fusarium culmorumFLAG1_03029 Fuslan Fusarium langsethiaeFPOA_02551 Fuspoa Fusarium poaeFPSE_08678 Fuspse Fusarium pseudograminearumFVRRES_02981 Fusven Fusarium venenatumFG02560.1 Gibzea Gibberella zeaeFGRA07_08229 NA Gibberella zeaeNECHADRAFT_100964 Nechae Nectria haematococcaAK830_g11716 Neodit_1 Neonectria ditissimaAK830_g6664 Neodit_2 Neonectria ditissimaDCS_02975 Drecon Drechmeria coniosporaHIM_09673 Hirmin Hirsutella minnesotensisCDD81_1416 Ophaus Ophiocordyceps australisCDD82_1745 NA Ophiocordyceps australisPCL_01709 Purlil Purpureocillium lilacinumVFPBJ_02636 NA Purpureocillium lilacinumTCAP_03967 Tolcap_1 Tolypocladium capitatumTCAP_02490 Tolcap_2 Tolypocladium capitatumTOPH_03782 Toloph Tolypocladium ophioglossoidesTPAR_06182 Tolpar Tolypocladium paradoxumjhhlp_007773 Lompro Lomentospora prolificansSAPIO_CDS1471 Sceapi Scedosporium apiospermumCONLIGDRAFT_665122 Conlig Coniochaeta ligniariaBD289DRAFT_344446 Conlus_1 Coniella lustricolaBD289DRAFT_419655 Conlus_2 Coniella lustricolaDHEL01_v204442 Diahel_2 Diaporthe helianthiDHEL01_v205905 Diahel_1 Diaporthe helianthiVM1G_07228 Valmal_1 Valsa mali VM1G_03758 Valmal_2 Valsa mali VP1G_08017 NA Valsa mali var. pyriVP1G_01130 NA Valsa mali var. pyriGGTG_07450 Gaegra Gaeumannomyces graminis var. triticiMGG_02347 Magory Magnaporthe oryzae OOW_P131scaffold01609g1 NA Magnaporthe oryzae OOU_Y34scaffold00129g8 NA Magnaporthe oryzae MAPG_06957 Magpoa Magnaporthiopsis poaeK445DRAFT_321672 Dalsp. Daldinia sp.M426DRAFT_325989 Hypsp. Hypoxylon sp.M434DRAFT_42892 NA Hypoxylon sp.K449DRAFT_319022 NA Hypoxylon sp.PFICI_11520 Pesfic Pestalotiopsis ficiBCR38DRAFT_410336 Psevex Pseudomassariella vexataSAMD00023353_5000550 Rosnec Rosellinia necatrixAOL_s00109g49 Artoli Arthrobotrys oligosporaH072_712 Dachap_1 Dactylellina haptotylaH072_5146 Dachap_2 Dactylellina haptotylaDRE_07509 Dreste Drechslerella stenobrochaa Genes with short ID were used for sequence and phylogenetic analysisb NA, not assigned
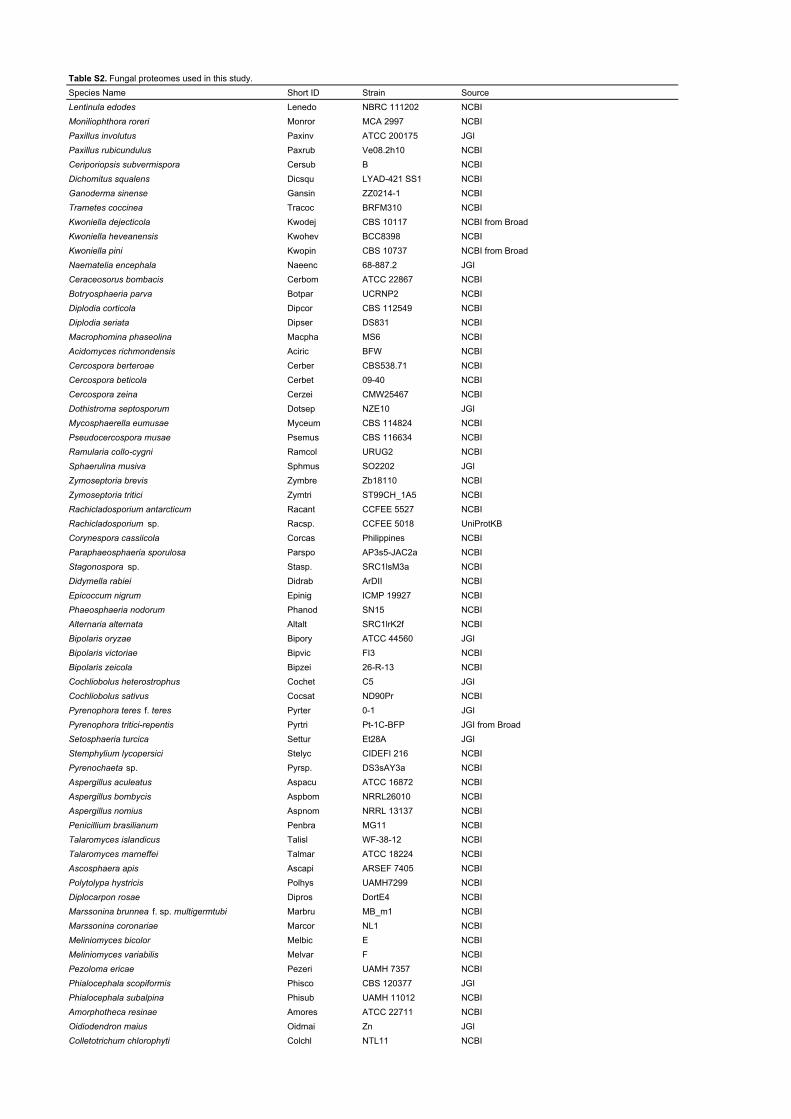
Table S2. Fungal proteomes used in this study.Species Name Short ID Strain Source Lentinula edodes Lenedo NBRC 111202 NCBIMoniliophthora roreri Monror MCA 2997 NCBIPaxillus involutus Paxinv ATCC 200175 JGIPaxillus rubicundulus Paxrub Ve08.2h10 NCBICeriporiopsis subvermispora Cersub B NCBIDichomitus squalens Dicsqu LYAD-421 SS1 NCBIGanoderma sinense Gansin ZZ0214-1 NCBITrametes coccinea Tracoc BRFM310 NCBIKwoniella dejecticola Kwodej CBS 10117 NCBI from BroadKwoniella heveanensis Kwohev BCC8398 NCBIKwoniella pini Kwopin CBS 10737 NCBI from BroadNaematelia encephala Naeenc 68-887.2 JGICeraceosorus bombacis Cerbom ATCC 22867 NCBIBotryosphaeria parva Botpar UCRNP2 NCBIDiplodia corticola Dipcor CBS 112549 NCBIDiplodia seriata Dipser DS831 NCBIMacrophomina phaseolina Macpha MS6 NCBIAcidomyces richmondensis Aciric BFW NCBICercospora berteroae Cerber CBS538.71 NCBICercospora beticola Cerbet 09-40 NCBICercospora zeina Cerzei CMW25467 NCBIDothistroma septosporum Dotsep NZE10 JGIMycosphaerella eumusae Myceum CBS 114824 NCBIPseudocercospora musae Psemus CBS 116634 NCBIRamularia collo-cygni Ramcol URUG2 NCBISphaerulina musiva Sphmus SO2202 JGIZymoseptoria brevis Zymbre Zb18110 NCBIZymoseptoria tritici Zymtri ST99CH_1A5 NCBIRachicladosporium antarcticum Racant CCFEE 5527 NCBIRachicladosporium sp. Racsp. CCFEE 5018 UniProtKBCorynespora cassiicola Corcas Philippines NCBIParaphaeosphaeria sporulosa Parspo AP3s5-JAC2a NCBIStagonospora sp. Stasp. SRC1lsM3a NCBIDidymella rabiei Didrab ArDII NCBIEpicoccum nigrum Epinig ICMP 19927 NCBIPhaeosphaeria nodorum Phanod SN15 NCBIAlternaria alternata Altalt SRC1lrK2f NCBIBipolaris oryzae Bipory ATCC 44560 JGIBipolaris victoriae Bipvic FI3 NCBIBipolaris zeicola Bipzei 26-R-13 NCBICochliobolus heterostrophus Cochet C5 JGICochliobolus sativus Cocsat ND90Pr NCBIPyrenophora teres f. teres Pyrter 0-1 JGIPyrenophora tritici-repentis Pyrtri Pt-1C-BFP JGI from BroadSetosphaeria turcica Settur Et28A JGIStemphylium lycopersici Stelyc CIDEFI 216 NCBIPyrenochaeta sp. Pyrsp. DS3sAY3a NCBIAspergillus aculeatus Aspacu ATCC 16872 NCBIAspergillus bombycis Aspbom NRRL26010 NCBIAspergillus nomius Aspnom NRRL 13137 NCBIPenicillium brasilianum Penbra MG11 NCBITalaromyces islandicus Talisl WF-38-12 NCBITalaromyces marneffei Talmar ATCC 18224 NCBIAscosphaera apis Ascapi ARSEF 7405 NCBIPolytolypa hystricis Polhys UAMH7299 NCBIDiplocarpon rosae Dipros DortE4 NCBIMarssonina brunnea f. sp. multigermtubi Marbru MB_m1 NCBIMarssonina coronariae Marcor NL1 NCBIMeliniomyces bicolor Melbic E NCBIMeliniomyces variabilis Melvar F NCBIPezoloma ericae Pezeri UAMH 7357 NCBIPhialocephala scopiformis Phisco CBS 120377 JGIPhialocephala subalpina Phisub UAMH 11012 NCBIAmorphotheca resinae Amores ATCC 22711 NCBIOidiodendron maius Oidmai Zn JGIColletotrichum chlorophyti Colchl NTL11 NCBI
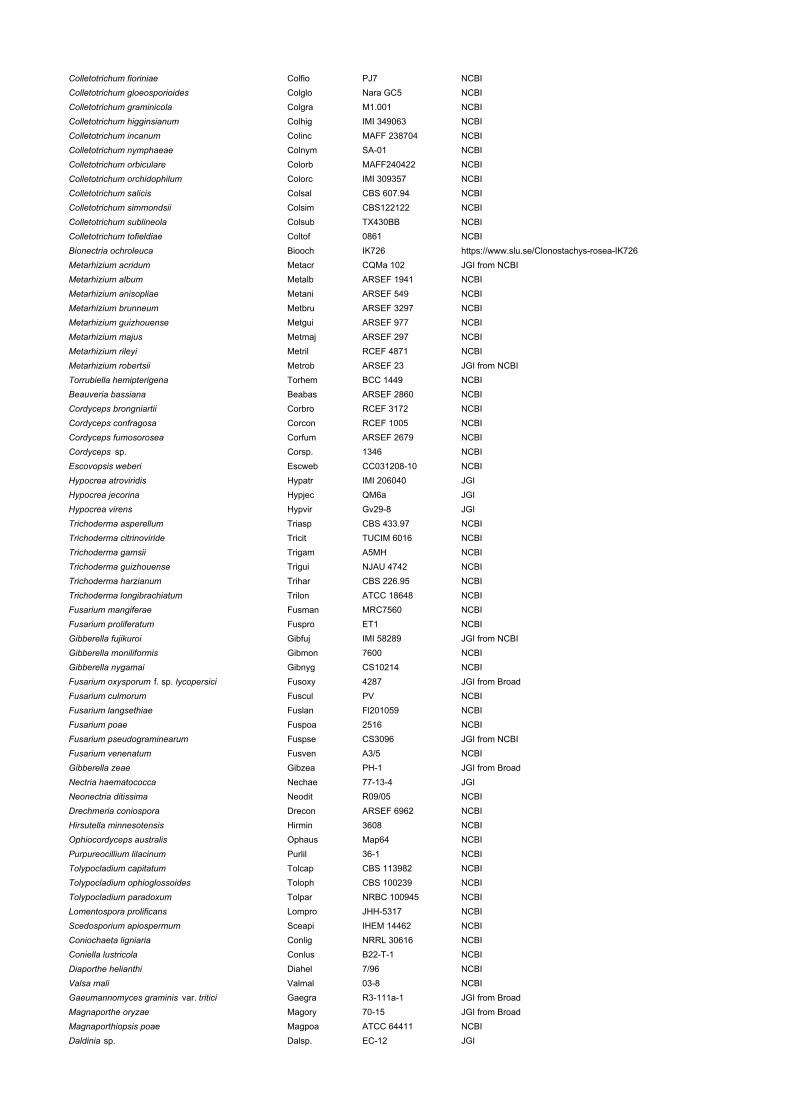
Colletotrichum fioriniae Colfio PJ7 NCBIColletotrichum gloeosporioides Colglo Nara GC5 NCBIColletotrichum graminicola Colgra M1.001 NCBIColletotrichum higginsianum Colhig IMI 349063 NCBIColletotrichum incanum Colinc MAFF 238704 NCBIColletotrichum nymphaeae Colnym SA-01 NCBIColletotrichum orbiculare Colorb MAFF240422 NCBIColletotrichum orchidophilum Colorc IMI 309357 NCBIColletotrichum salicis Colsal CBS 607.94 NCBIColletotrichum simmondsii Colsim CBS122122 NCBIColletotrichum sublineola Colsub TX430BB NCBIColletotrichum tofieldiae Coltof 0861 NCBIBionectria ochroleuca Biooch IK726 https://www.slu.se/Clonostachys-rosea-IK726Metarhizium acridum Metacr CQMa 102 JGI from NCBIMetarhizium album Metalb ARSEF 1941 NCBIMetarhizium anisopliae Metani ARSEF 549 NCBIMetarhizium brunneum Metbru ARSEF 3297 NCBIMetarhizium guizhouense Metgui ARSEF 977 NCBIMetarhizium majus Metmaj ARSEF 297 NCBIMetarhizium rileyi Metril RCEF 4871 NCBIMetarhizium robertsii Metrob ARSEF 23 JGI from NCBITorrubiella hemipterigena Torhem BCC 1449 NCBIBeauveria bassiana Beabas ARSEF 2860 NCBICordyceps brongniartii Corbro RCEF 3172 NCBICordyceps confragosa Corcon RCEF 1005 NCBICordyceps fumosorosea Corfum ARSEF 2679 NCBICordyceps sp. Corsp. 1346 NCBIEscovopsis weberi Escweb CC031208-10 NCBIHypocrea atroviridis Hypatr IMI 206040 JGIHypocrea jecorina Hypjec QM6a JGIHypocrea virens Hypvir Gv29-8 JGITrichoderma asperellum Triasp CBS 433.97 NCBITrichoderma citrinoviride Tricit TUCIM 6016 NCBITrichoderma gamsii Trigam A5MH NCBITrichoderma guizhouense Trigui NJAU 4742 NCBITrichoderma harzianum Trihar CBS 226.95 NCBITrichoderma longibrachiatum Trilon ATCC 18648 NCBIFusarium mangiferae Fusman MRC7560 NCBIFusarium proliferatum Fuspro ET1 NCBIGibberella fujikuroi Gibfuj IMI 58289 JGI from NCBIGibberella moniliformis Gibmon 7600 NCBIGibberella nygamai Gibnyg CS10214 NCBIFusarium oxysporum f. sp. lycopersici Fusoxy 4287 JGI from BroadFusarium culmorum Fuscul PV NCBIFusarium langsethiae Fuslan Fl201059 NCBIFusarium poae Fuspoa 2516 NCBIFusarium pseudograminearum Fuspse CS3096 JGI from NCBIFusarium venenatum Fusven A3/5 NCBIGibberella zeae Gibzea PH-1 JGI from BroadNectria haematococca Nechae 77-13-4 JGINeonectria ditissima Neodit R09/05 NCBIDrechmeria coniospora Drecon ARSEF 6962 NCBIHirsutella minnesotensis Hirmin 3608 NCBIOphiocordyceps australis Ophaus Map64 NCBIPurpureocillium lilacinum Purlil 36-1 NCBITolypocladium capitatum Tolcap CBS 113982 NCBITolypocladium ophioglossoides Toloph CBS 100239 NCBITolypocladium paradoxum Tolpar NRBC 100945 NCBILomentospora prolificans Lompro JHH-5317 NCBIScedosporium apiospermum Sceapi IHEM 14462 NCBIConiochaeta ligniaria Conlig NRRL 30616 NCBIConiella lustricola Conlus B22-T-1 NCBIDiaporthe helianthi Diahel 7/96 NCBIValsa mali Valmal 03-8 NCBIGaeumannomyces graminis var. tritici Gaegra R3-111a-1 JGI from BroadMagnaporthe oryzae Magory 70-15 JGI from BroadMagnaporthiopsis poae Magpoa ATCC 64411 NCBIDaldinia sp. Dalsp. EC-12 JGI

Hypoxylon sp. Hypsp. CI-4A JGIPestalotiopsis fici Pesfic W106-1 NCBIPseudomassariella vexata Psevex CBS 129021 NCBIRosellinia necatrix Rosnec W97 NCBIArthrobotrys oligospora Artoli ATCC 24927 JGI from NCBIDactylellina haptotyla Dachap CBS 200.50 JGI from NCBIDrechslerella stenobrocha Dreste 248 NCBI

Table S3. Primers used in this study.Name Sequence destination35SpNIS1fwd 5΄-CGGGATCCTTAAGGAGATATAACAATGCAGTTCCTCACCTCCCTC-3΄ pBICP35-CoNIS1ΔC30-(S123A, Y125A, V127A, G132A)-HACoNIS1dC30-S123A-HA-EcoRI-r(c) 5΄-CGAATTCTCAAGCGTAATCTGGAACATCGTATGGGTAGCCGACGTTGACGGTGACGTTGTAGTTGGCGACGGTGGGAC-3΄pBICP35-CoNIS1ΔC30-S123A-HACoNIS1dC30-Y125A-HA-EcoRI-r(c) 5΄-CGAATTCTCAAGCGTAATCTGGAACATCGTATGGGTAGCCGACGTTGACGGTGACGTTGGCGTTGGAGACGG-3΄ pBICP35-CoNIS1ΔC30-Y125A-HACoNIS1dC30-V127A-HA-EcoRI-r(c) 5΄-CGAATTCTCAAGCGTAATCTGGAACATCGTATGGGTAGCCGACGTTGACGGTGGCGTTGTAGTTGG-3΄ pBICP35-CoNIS1ΔC30-V127A-HACoNIS1dC30-G132A-HA-EcoRI-r(c) 5΄-CGAATTCTCAAGCGTAATCTGGAACATCGTATGGGTAGGCGACGTTGACGGTGACGT-3΄ pBICP35-CoNIS1ΔC30-G132A-HAeGFP-EcoRI-f 5'-CGGAATTCATGGTGAGCAAGGGCGAGGAG-3' pBICP35-eGFP-CeGFP-KpnI-r(c) 5'-GGGGTACCTTACTTGTACAGCTCGTCCATGC-3' pBICP35-eGFP-C, pBICP35-BIK1-GFPAtBAK1-BamHI-f 5'-CGGGATCCAAGGAGATATAACAATGGAACGAAGATTAATGATCCCTTGC-3' pBICP35-BAK1-GFPAtBAK1-EcoRI-r(c) 5'-GGAATTCTCTTGGACCCGAGGGGTATTC-3' pBICP35-BAK1-GFPAtBKK1-BamHI-f 5'-CGGGATCCAAGGAGATATAACAATGACAAGTTCAAAAATGGAACAAAGATCAC-3' pBICP35-BKK1-GFPAtBKK1-EcoRI-r(c) 5'-GGAATTCTCTTGGACCCGAGGGGTAATCG-3' pBICP35-BKK1-GFPNbSERK3-BamHI-f 5'-CGGGATCCAAGGAGATATAACAATGGATCAGTCGGTCTTGTTG-3' pBICP35-NbSERK3-GFPNbSERK3-EcoRI-r(c) 5'-GGAATTCTCTTGGCCCCGACAATTCATC-3' pBICP35-NbSERK3-GFPOsBAK1-BamHI-f 5'-CGGGATCCAAGGAGATATAACAATGGCGGCGCATCGGTGG-3' pBICP35-OsBAK1-GFPOsBAK1-EcoRI-r(c) 5'-GGAATTCCCTCGGCCCTGATAGCTCAAC-3' pBICP35-OsBAK1-GFPAtBIK1-BamHI-f 5'-CGGGATCCATGGGTTCTTGCTTCAGTTCTC-3' pBICP35-BIK1-GFP, pBICP35-BIK1-FLAGAtBIK1-PstI-r(c) 5'-CATTGGTTCTGCAGCACAAGGTGCCTGCCAAAAGG-3' pBICP35-BIK1-GFPeGFP-PstI-f 5'-CATTGGTTCTGCAGATGGTGAGCAAGGGCGAGGAG-3' pBICP35-BIK1-GFP
FLAG3-AtRBOHD-BamHI-f5'-CGGGATCCAAGGAGATATAACAATGGACTACAAAGACCATGACGGTGATTATAAAGATCATGACATCGACTACAAGGACGACGATGACAAGAAAATGAGACGAGGC-3'
pBICP35-3xFLAG-RBOHD
AtRBOHD-EcoRI-r(c) 5'-GGAATTCCTAGAAGTTCTCTTTGTGGAAGTC-3' pBICP35-3xFLAG-RBOHDAtBAK1-Cyto-EcoRI-f 5'-GGAATTCATGCTAGCTTGGTGGCGAAGG-3' pGBKT7-BAK1-CytoAtBAK1-BamHI-r(c) 5'-CGGGATCCTTATCTTGGACCCGAGGGGTATTCG-3' pGBKT7-BAK1-CytoAtBKK1-Cyto-EcoRI-f 5'-GGAATTCATGTTTGCTTGGTGGCTCAG-3' pGBKT7-BKK1-CytoAtBKK1-BamHI-r(c) 5'-CGGGATCCTTATCTTGGACCCGAGGGGTAATCG-3' pGBKT7-BKK1-CytoCtofNIS1-F_InFu_pBICBamHI 5'-AGGCCTACGGGGATCCAAGGAGATATAACAATGCAGTTCCGATCTTCCCTC-3' pBICP35-CtofNIS1-HACtofNIS1-HA-R_InFu_pBICBamHI 5'-GGAATTCCGGGGATCCCTAAGCGTAATCTGGAACATCGTATGGGTATCGCTGGCTGCCGACGTATGT-3' pBICP35-CtofNIS1-HAMoNIS1-BamHI-f 5'-CGGGATCCAAGGAGATATAACAATGCAGCTCCTCGCCACGACCCTCAC-3' pBICP35-MoNIS1-mCherryMoNIS1-BamHI-NS-r 5'-CGGGATCCACCACCACCACCACCCTCGCAGACCTGAACTGTAGTA-3' pBICP35-MoNIS1-mCherryCoNIS1dSP-EcoR I-f 5'-CGGAATTCATGCGCATCTCCGGCATCGC-3' pGADT7-NIS1ΔSPCoNIS1-BamHI-r(c) 5'-CGGGATCCTCAGATTTTCCTCGTGTACCCGC-3' pGADT7-NIS1ΔSPMoNIS1dSP-EcoR I-f 5'-GGAATTCATGTACATCAACAGCATGAGCGCGCC-3' pGADT7-MoNIS1ΔSPMoNIS1-BamHI-r(c) 5'-CGGGATCCTTACTCGCAGACCTGAACTGTAG-3' pGADT7-MoNIS1ΔSPPro+CoNIS1-Not I-f 5'-ATAAGAATGCGGCCGCATAGAATGAGTTTCGTTATTG-3' pBA-CoNIS1proORFterCoNIS1ter-EcoRI-r(c) 5'-GGAATTCCCAGCTACGGGCATCTCATTAATC-3' pBA-CoNIS1proORFterMoNIS1-S1 5'-TTCACCCTTGCCGTTTGTTTGCCTTCTC-3' pCR-XL-MoNIS1MoNIS1-AS1 5'-GCTAATCAAGAAACCGCTGGGCACAAGG-3' pCR-XL-MoNIS1KMNS2 5'-GGGGTACCCCATCTTCACTCTGCTCGGCC-3' MoNIS1 gene-disruption vectorAMNL2 5'-GAAGGGCCCGGTGGGTGGATGCGATTTTTATTTTTAG-3' MoNIS1 gene-disruption vectorEMNU2 5'-GGAATTCTGGAGCTTTAATGTTTCATATTTG-3' MoNIS1 gene-disruption vectorXMNAS2 5'- CTCTAGAAGTCTGCTAGGTCTACC-3' MoNIS1 gene-disruption vectorNMNpU-1 5'-ATAAGAATGCGGCCGCCGCAGTTGTCTCGGCATGGATC-3' pCB1531-MoNIS1MoNIS1-L2 5'- CCTTATTGTCCCTTTTTAGCAG-3' pCB1531-MoNIS1

1
Supplemental References
1. Grant SG, Jessee J, Bloom FR, Hanahan D (1990) Differential plasmid rescue from
transgenic mouse DNAs into Escherichia coli methylation-restriction mutants. Proc
Natl Acad Sci USA 87: 4645-4649.
2. Mori M, Mise K, Kobayashi K, Okuno T, Furusawa I (1991) Infectivity of plasmids
containing brome mosaic virus cDNA linked to the cauliflower mosaic virus 35S
RNA promoter. J Gen Virol 72: 243-246.
3. Irieda H, Maeda H, Akiyama K, Hagiwara A, Saitoh H, Uemura A, Terauchi R,
Takano Y (2014) Colletotrichum orbiculare secretes virulence effectors to a
biotrophic interface at the primary hyphal neck via exocytosis coupled with
SEC22-mediated traffic. Plant Cell 26: 2265-2281.
4. Dohi K, Nishikiori M, Tamai A, Ishikawa M, Meshi T, Mori M (2006) Inducible
virus-mediated expression of a foreign protein in suspension-cultured plant cells.
Arch Virol 15: 1075-1084.
5. Dohi K, Tamai A, Mori M (2008) Insertion in the coding region of the movement
protein improves stability of the plasmid encoding a tomato mosaic virus-based
expression vector. Arch Virol 153: 1667-1675.
6. Sweigard J, Chumley F, Carroll A, Farrall L, Valent B (1997) A series of vectors for
fungal transformation. Fungal Genet Newsl 44: 52-55.
7. Lipka V, Dittgen J, Bednarek P, Bhat R, Wiermer M, Stein M, Landtag J, Brandt W,
Rosahl S, Scheel D, Llorente F, Molina A, Parker J, Somerville S, Schulze-Lefert P
(2005) Pre- and postinvasion defenses both contribute to nonhost resistance in
Arabidopsis. Science 310: 1180-1183.
8. Schwessinger B, Roux M, Kadota Y, Ntoukakis V, Sklenar J, Jones A, Zipfel C (2011)
Phosphorylation-dependent differential regulation of plant growth, cell death, and
innate immunity by the regulatory receptor-like kinase BAK1. PLoS Genet 7:
e1002046.
9. Veronese P, Nakagami H, Bluhm B, Abuqamar S, Chen X, Salmeron J, Dietrich RA,
Hirt H, Mengiste T (2006) The membrane-anchored BOTRYTIS-INDUCED
KINASE1 plays distinct roles in Arabidopsis resistance to necrotrophic and
biotrophic pathogens. Plant Cell 18: 257-273.

2
10. Zhang J, Li W, Xiang T, Liu Z, Laluk K, Ding X, Zou Y, Gao M, Zhang X, Chen S,
Mengiste T, Zhang Y, Zhou JM (2010) Receptor-like cytoplasmic kinases integrate
signaling from multiple plant immune receptors and are targeted by a Pseudomonas
syringae effector. Cell Host Microbe 7: 290-301.
11. Yoshida K, Saitoh H, Fujisawa S, Kanzaki H, Matsumura H, Yoshida K, Tosa Y,
Chuma I, Takano Y, Win J, Kamoun S, Terauchi R (2009) Association genetics
reveals three novel avirulence genes from the rice blast fungal pathogen
Magnaporthe oryzae. Plant Cell 21: 1573-1591.
12. Hyon G-S, Nga NT, Chuma I, Inoue Y, Asano H, Murata N, Kusaba M, Tosa Y
(2012) Characterization of interactions between barley and various host-specific
subgroups of Magnaporthe oryzae and M. grisea. J Gen Plant Pathol 78: 237-246.
13. Gómez-Gómez L, Felix G, Boller T (1999) A single locus determines sensitivity to
bacterial flagellin in Arabidopsis thaliana. Plant J 18: 277-284.
14. Yamada K, Saijo Y, Nakagami H, Takano Y (2016) Regulation of sugar transporter
activity for antibacterial defense in Arabidopsis. Science 354: 1427-1430.
15. Studier FW, Moffatt BA (1986) Use of bacteriophage T7 RNA polymerase to direct
selective high-level expression of cloned genes. J Mol Biol 189: 113-130.
16. Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H,
Remmert M, Soding J, Thompson JD, Higgins DG (2011) Fast, scalable generation
of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst.
Biol. 7:539.
17. Capella-Gutierrez S, Silla-Martinez JM, Gabaldon T (2009) trimAl: a tool for
automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics
25:1972-1973.
18. Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010) New
algorithms and methods to estimate maximum-likelihood phylogenies: assessing the
performance of PhyML 3.0. Syst Biol 59: 307-321.
19. Qi J, Wang B, Hao BI (2004) Whole proteome prokaryote phylogeny without
sequence alignment: a K-string composition approach. J Mol Evol 58: 1-11.
20. Zuo G, Hao B (2015) CVTree3 web server for whole-genome-based and
alignment-free prokaryotic phylogeny and taxonomy. Genomics Proteomics
Bioinformatics 13: 321-331.


![PC-to-Mac Transfer Kitcache-1].pdf · Common file types: .mpeg, .mov, .avi Desktop Files – This option includes any file formats located in this folder on your PC . Common file](https://static.fdocument.org/doc/165x107/5f4e305cb8f54b47894e5020/pc-to-mac-transfer-kitcache-1pdf-common-file-types-mpeg-mov-avi-desktop.jpg)















