Thermal Stress and the Heat Shock Response in Microbes
13
1 | Page THERMAL STRESS AND THE HEAT SHOCK RESPONSE
-
Upload
amna-jalil -
Category
Education
-
view
249 -
download
0
Transcript of Thermal Stress and the Heat Shock Response in Microbes

1 | P a g e
THERMAL STRESS AND THE HEAT
SHOCK RESPONSE

2 | P a g e
Contents Types of Microbes on the basis of Temperature
Thermophile
Psychrophile
Mesophile
Thermal Stress
Heat Shock Response
Description
Types of Heat Shock Responses
Heat Shock Response against Cytoplasmic Thermal
Stress
σᴴ Regulation
Heat Shock Response against Periplasmic Thermal
Stress
σᴱ Regulation
Cpx systems

3 | P a g e
THERMAL STRESS AND THE HEAT SHOCK RESPONSE
As with pH, microbes exhibit a wide range of temperatures at which they can grow.
Types of Microbes on the basis of Temperature
There are three types of microbes on the basis of temperature.
1. Thermophiles
2. Psychrophiles
3. Mesophiles
Thermophile
A thermophile is an organism — a type of extremophile — that thrives at
relatively high temperatures, between 45 and 122 °C (113 and 252 °F).
Many thermophiles are archaea. Thermophilic eubacteria are suggested to have
been among the earliest bacteria.
Thermophiles are found in various geothermally heated regions of the Earth, such
as hot springs like those in Yellowstone National Park and deep sea hydrothermal
vents, as well as decaying plant matter, such as peat bogs and compost.
As a prerequisite for their survival, thermophiles contain enzymes that can
function at high temperatures. Some of these enzymes are used in molecular
biology (for example, heat-stable DNA polymerases for PCR), and in washing
agents.

4 | P a g e
"Thermophile" is derived from the Greek: θερμότητα (thermotita),
meaning heat, and Greek: υίλια (philia), love
A colony of thermophiles in the outflow of Mickey Hot Springs, Oregon, the water temperature is approximately 60°C.
Psychrophile
Psychrophiles or cryophiles are extremophilic organisms that are capable of
growth and reproduction in cold temperatures, ranging from −15°C to +10°C.
Temperatures as low as −15°C are found in pockets of very salty water (brine)
surrounded by sea ice.
They can be contrasted with thermophiles, which thrive at unusually hot
temperatures. The environments they inhabit are ubiquitous on Earth, as a large
fraction of our planetary surface experiences temperatures lower than 15°C.
They are present in alpine and arctic soils, high-latitude and deep ocean waters,
polar ice, glaciers, and snowfields.

5 | P a g e
Examples are Arthrobacter sp., Psychrobacter sp. and members
of the genera Halomonas, Pseudomonas, Hyphomonas and
Sphingomonas.
Cold-loving extremophile bacteria
Mesophile
A mesophile is an organism that grows best in moderate temperature, neither too
hot nor too cold, typically between 20 and 45 °C (68 and 113 °F). The term is
mainly applied to microorganisms.
The habitats of these organisms include especially cheese, yogurt, and mesophile
organisms are often included in the process of beer and wine making.
Organisms that prefer cold environments are termed psychrophilic, those
preferring warmer temperatures are termed thermophilic and those thriving in
extremely hot environments are hyperthermophilic. All bacteria have their own
optimum environmental surroundings and temperatures in which they thrive the
most.
Examples include E.coli and Streptococcus.
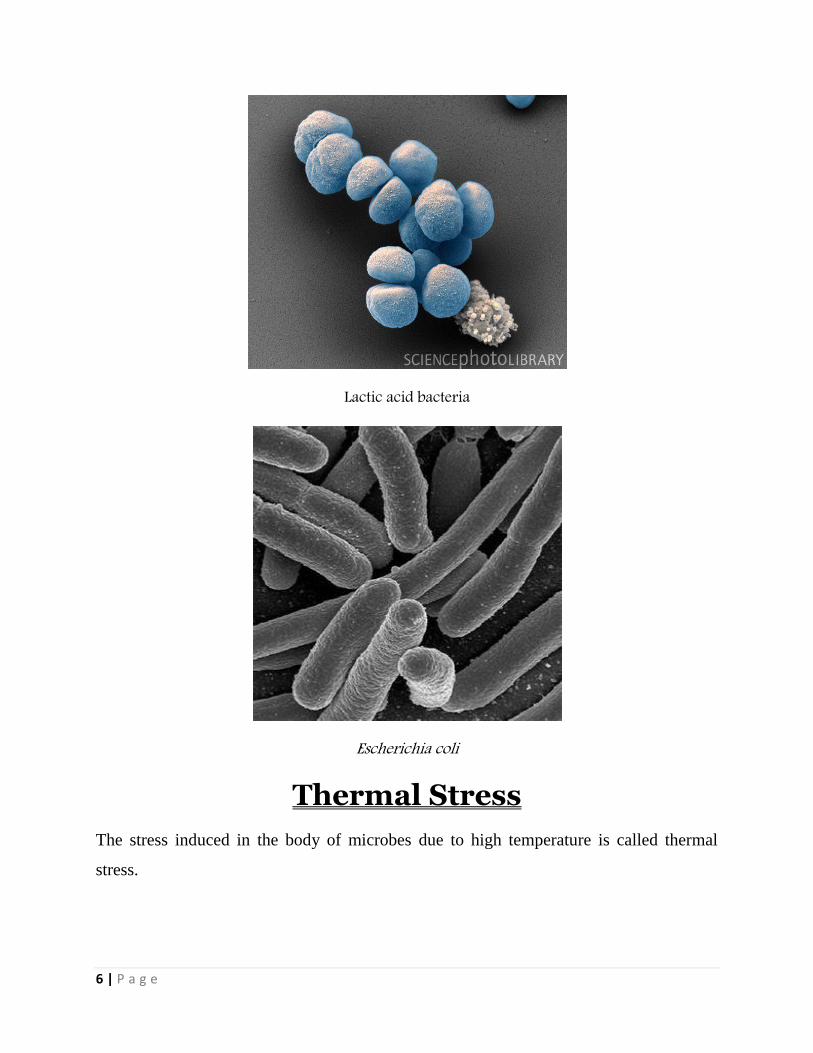
6 | P a g e
Lactic acid bacteria
Escherichia coli
Thermal Stress
The stress induced in the body of microbes due to high temperature is called thermal
stress.

7 | P a g e
Heat Shock Response
The heat shock represents a ubiquitous protective and homeostatic cellular response to
cope with heat-induced damage in proteins.
Description
Upon a shift from 30◦ to 42 ◦C, E. coli and other bacteria transiently increase the
rate of synthesis of a set of proteins called heat shock proteins (HSPs).
Many of these HSPs are required for cell growth or survival at more elevated
temperatures (thermotolerance).
Among the induced proteins are DnaJ and DnaK, the RNA polymerase σ70
subunit (rpoD), GroES, GroEL, Lon protease, and LysU.
There are nearly 50 heat shock–inducible proteins identified in E. coli.
Types of Heat Shock Responses
There are two types of heat shock responses.
1. Heat shock response against cytoplasmic thermal stress
2. Heat shock response against periplasmic thermal stress
The hallmark ofthe Gram-negative cell is the existence of two membrane-bound
subcellular compartments:
Cytoplasm
Periplasm
Conditions in each of these compartments differ markedly.
The cytoplasm is energy rich, reducing (low redox potential), and osmotically
stable.

8 | P a g e
Whereas the periplasm lacks ATP, is oxidizing, and is in contact withthe
external milieu.
Since optimal cell growth requires that the cell senses and responds to changes in these
disparate sub cellular compartments, it is not surprising that the stress responses in E. coli
and S. typhimurium are compartmentalized into cytoplasmic and extracytoplasmic
responses.
1. Heat Shock Response against
Cytoplasmic Thermal Stress
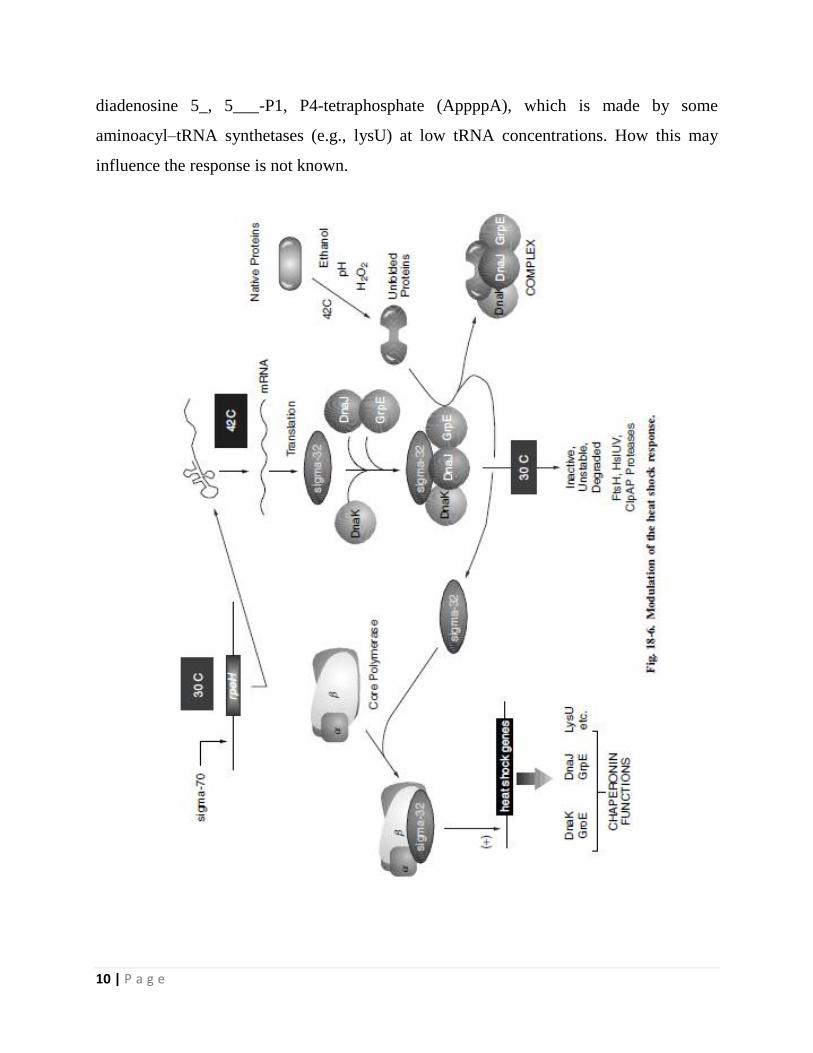
The σᴴ regulon provides protection against cytoplasmic thermal stress.
The E. coli rpoH locus (formerly called htpR) encodes a 32 kDa σ factor,
alternatively called σᴴ or σ³², which redirects promoter specificity of RNA
polymerase.
The σᴴ protein regulates the expression of 34 heat shock genes.
The simple explanation for how heat shock increases expression of the σᴴ regulon
is that heat shock first causes an elevation in σᴴ levels, which in turn increases
expression of the σᴴ target genes.
Although the principle is simple, the controls governing σᴴ production are
complex.
First, a temperature upshifts from 30 ◦ to 42 ◦C results in the increased translation
of rpoH message.
Cis-acting mRNA sites within the 5’ region of rpoH message form temperature-
sensitive secondary structures that sequester the ribosome-binding site. At higher
temperatures, these secondary structures melt, thereby enabling more efficient
translation of the rpoH message.
In addition to the increased translation of rpoH message, the σᴴ protein itself
becomes more stable, at least transiently.

9 | P a g e
The mechanism regulating proteolysis centers on whether σᴴ associates with RNA
polymerase. During growth at 30 ◦C, σᴴ can be degraded by several proteases
including FtsH, HslVU, and ClpAP.
However, if σᴴ is bound to RNAP, σᴴ is protected from degradation.
The cell uses the DnaK-DnaJ-GrpE chaperone team to interact with σᴴ at low
temperature, sequestering σᴴ from RNA polymerase.
Failure to bind RNAP facilitates degradation of the σ factor.
Upon heat shock, there is an increase in the number of other unfolded or denatured
proteins that can bind to DnaK or DnaJ. This reduces the level of free DnaK/DnaJ
molecules available to bind σᴴ, allowing σᴴ tobind RNAP, which protects σᴴ from
degradation.
As the cell reaches the adaptation phase following heat shock, the levels of DnaK
and DnaJ rise (both are induced by σᴴ) and can again bind σᴴ, redirecting it toward
degradation. Nevertheless, even though σᴴ degradation resumes, translation of
rpoH remains high at the elevated temperature and σᴴ continues to accumulate,
although at a slower rate.
In addition to translational and proteolytic controls, production of σᴴ is regulated at the
transcriptional level via a feedback mechanism. There are four promoters driving rpoH
expression, three of which are dependent on σ70, the housekeeping σ factor.
The gene encoding σ70, rpoD, is also a heat shock gene induced by σᴴ. So increased
production of σᴴ increases σ70, which increases transcription of rpoH. The fourth rpoH
promoter is recognized by another σ factor, σᴱ, encoded by rpoE.
The heat shock response is also triggered by a variety of environmental agents such as
ethanol, UV irradiation, and agents that inhibit DNA gyrase. Induction by all of these
stimuli occurs through σᴴ. The only explanation that appears reasonable is the
accumulation of denatured or incomplete peptides. There is a potential alarmone that has
been implicated in signaling expression of this global network. The molecule is
10 | P a g e
diadenosine 5_, 5___-P1, P4-tetraphosphate (AppppA), which is made by some
aminoacyl–tRNA synthetases (e.g., lysU) at low tRNA concentrations. How this may
influence the response is not known.

11 | P a g e
2. Heat Shock Response against
Periplasmic Thermal Stress
The extra cytoplasmic response pathways involve two partially overlapping signal
transduction cascades:
I. σᴱ regulation
II. Cpx systems
These pathways are induced following the accumulation of misfolded proteins in the
periplasmas a result of stresses such as high temperature, pH extremes, or carbon/energy
starvation.
I. σᴱ Regulation
σᴱis a member of the extracytoplasmic function (ECF) subfamily of σ factors.In E. coli,
σᴱ is responsible for the transcription of up to genes, including:
rpoH (σᴴ)
degP (htrA) encoding a periplasmic protease for the degradation of
misfolded proteins
fkpA encoding a periplasmic peptidyl prolyl isomerase
rpoE rseABC operon
The gene encoding σᴱ, rpoE, is the first member of an operon followed by the genes rseA,
B, and C (rse, meaning regulators of σE).

12 | P a g e
RseA is a transmembrane protein whose cytoplasmic C-terminal domain interacts
with σᴱ, acting as an anti-σfactor.
The periplasmic face of RseA binds to the periplasmic RseB.
Extracellular stress in some way signals increased proteolysis of RseA by the
periplasmic protease DegS, thus relieving the anti-σ effect of RseA on σE.
It has been proposed that Rse Band perhaps other periplasmic proteins involved in
protein folding protect RseA from degradation by binding to the RseA periplasmic
domain, capping the target site of DegS.
Stress-induced misfolding of periplasmic proteins would titrate the RseA cap
proteins off of RseA, rendering the anti-σ factor vulnerable to attack by DegS.
The result would be increased activity of σᴱ leading to increased levels of σᴱ
protein and RseA anti-σ (since they form an operon).
The increased amount of σᴱ will drive further expression of genes whose products
handle the periplasmic damage while the of RseA will enable the cell to down
regulate the system once the capping proteins are again free to bind and protect
RseA from DegS degradation.
II. Cpx systems
A second system dedicated to protecting the periplasmic perimeter of the cellis the
CpxRA two-component system with:
CpxA playing the role of membrane-localized sensor histidine kinase
CpxR as the cytoplasmicresponse regulator
CpxA responds to envelope stress by autophosphorylation followed by
phosphotransfer to CpxR. CpxR∼P activates expression of:
dsbA (disulfideoxidoreductase)
ppiA and ppiD encoding peptidyl-prolyl-isomerases

13 | P a g e
In E. coli ,in conjunction with σE, degP (htrA)
In addition, CpxR∼P activates transcription of cpxP encoding a small protein that
negatively regulates the CpxAR regulon, probablyby binding to a periplasmic
domain of CpxA.
The ability to autoactivate and then repress (via CpxP) enables a temporary
amplification of the Cpx response that maybe important to rescue cells from transitory
stresses.
PrpA and PrpB are type Iserine/threonine phosphatases that also participate in the ECF
pathway at least inpart by affecting the phosphorylation level of CpxR.
B. subtilis has four classes of heat shock genes. One is particularly interesting because
of the mechanism used to control the genes. Class I heat shock genes include the major
chaperones DnaK-DnaJ-GrpE and GroEL-GroES. Their transcription requires the
housekeeping σ factor, σA (σ70) and is negatively controlled by a repressor called HrcA.
HrcA binds to a class I geneoperator, a well-conserved 9 bp inverted repeat with a 9 bp
spacer, called CIRCE.
TheCIRCE/HrcA regulon is normally repressed by the HrcA repressor but can be heat
induced by inactivating the repressor. The molecular switch involves the chaperone
GroE. Unlike σ³² in E. coli, the GroE chaperones bind to and facilitate folding of HrcA
and thereby modulate repressor function. Titration of GroE by stress-induced misfolded
proteins results in lower HrcA repressor activity.